1. Introduction
Atherosclerosis, a clinically important
pathophysiological process characterized by lipid deposition in the
vessel wall, fuels the onset and progression of diverse
cardiovascular diseases responsible for the overwhelming majority
of deaths worldwide (1,2) and accounting for ~17.8 million
deaths annually (3).
Atherosclerosis is a slow and progressive process that involves
multifocal structural alterations in the vessel walls of large- and
medium-sized arteries, followed by secondary atherosclerotic plaque
formation (4). Several
pathophysiological processes, including endothelial dysfunction,
angiogenesis, inflammatory responses, lipid metabolism, aberrant
cellular proliferation and apoptosis, mechanistically provide the
foundation for this disease as key determinants of atherosclerosis
and ultimately the development of thrombotic plaque complications,
such as myocardial infarction (MI), stroke and sudden cardiac death
(1,5). Despite significant advances in the
clinical management of atherosclerosis and related risk factors,
such as hypertension, hyperlipidemia and diabetes,
atherosclerosis-associated acute ischemic events remain the primary
cause of death worldwide (6), and
thus, new cellular and molecular mechanisms are urgently needed to
develop novel therapeutic strategies for intervention.
Long non-coding RNAs (lncRNAs) are RNA transcripts
of >200 bp in length with little or no protein-coding potential
(7). Based on their genomic
locations relative to adjacent protein-coding genes, lncRNAs can be
classified as sense, antisense, bidirectional, intronic or
intergenic (8). On the grounds of
their functions, they can also be classified as signaling, decoy,
guide or scaffold lncRNAs (9). In
addition, lncRNAs are localized in the nucleus or cytoplasm and
exhibit tissue- or cell-specific expression, thus endowing them
extraordinary potential and multifarious functionality, and they
can serve as biomarkers for diagnostics and treatments (10). Mechanistically, lncRNAs perform
either in cis or trans mode to modulate the expression of target
genes through a series of molecular mechanisms, such as acting as a
scaffold for the recruitment of chromatin modifiers or
transcription factors or as decoys for protein sequestration and as
microRNA (miRNA) sponges to activate or repress genes (11,12). In addition, lncRNAs are also
involved in the regulation of mRNA splicing, translation and
turnover (13-15). Taken together, they are a unique
class of functional molecules that orchestrate diverse cellular
processes via multiple mechanisms involving, but not limited to,
direct mRNA, DNA or protein binding (9,16).
Several recent studies have shown that lncRNAs can affect the
initiation and progression of cardiovascular disease. Hence, it is
crucial to elucidate such functional roles for lncRNAs.
Among the several thousand lncRNAs, homeobox (HOX)
cluster-embedded lncRNAs (HOX-lncRNAs) have key roles in
controlling protein-coding genes under normal and pathogenic
conditions (17). Burgeoning
findings indicate that aberrantly expressed HOX-lncRNAs influence
cancer hallmarks, such as proliferation, migration and invasion,
and may also serve as potential regulators of cardiovascular
disease. Thus, the underlying molecular mechanisms could provide
theoretical evidence for the development of targeted therapies
against cardiovascular diseases. More recently, a plethora of data
has shown that HOX-lncRNAs are pivotal modulators of numerous
pathophysiological processes in cardiovascular biology. As an
example, the oncogenic lncRNA is HOX transcript antisense RNA
(HOTAIR) alleviates cardiac injury in chronic heart failure (HF) by
sponging miR-30a-5p to regulate lysine demethylase 3A (18). In addition, HOXA transcript at the
distal tip (HOTTIP) knockdown blocks acute MI (AMI) progression,
and further mechanistic studies have shown that HOTTIP exerts its
effects through miR-92a-2 sponging (19). Moreover, its expression is
significantly upregulated in sepsis-induced cardiac dysfunction and
is closely associated with the development of cardiac dysfunction
(20). In a
streptozotocin-induced type 1 diabetes mouse model, HOTAIR
overexpression ameliorated cardiac function and alleviated
oxidative stress and the inflammatory response, thus providing a
possible direction for targeting diabetic cardiomyopathy (21). In summary, these investigations
provide a foundation for understanding the role of HOX-lncRNAs in
cardiovascular disease. A follow-up delineation of how HOX-lncRNAs
regulate cardiovascular biology will advance the current
understanding of cardiovascular diseases, including
atherosclerosis. Furthermore, molecular insight into
atherosclerosis involving HOX-lncRNAs is a prerequisite for
developing ground-breaking therapies for early prevention and
intervention.
The main aim of the present review was to outline
the contributions of HOX-lncRNAs to atherosclerosis and other
crucial cardiovascular processes that directly result in the
development and progression of cardiovascular events. Collectively,
these investigations have provided a landscape in which HOX-lncRNAs
can mediate various pathophysiological processes, including lipid
metabolism, inflammatory response, angiogenesis, cellular
proliferation and apoptosis, cardiac hypertrophy and fibrosis,
myocardial ischemia/reperfusion (I/R) injury, cardiomyocyte
apoptosis, essential hypertension and pulmonary hypertension, which
are closely linked to the pathogenesis of cardiovascular disease.
Disentangling the biological mechanism ascribed to HOX-lncRNAs is
expected to yield compelling insight for a deeper understanding of
cardiovascular dysfunction.
2. Brief primer on HOX-lncRNAs and their
association with atherosclerosis
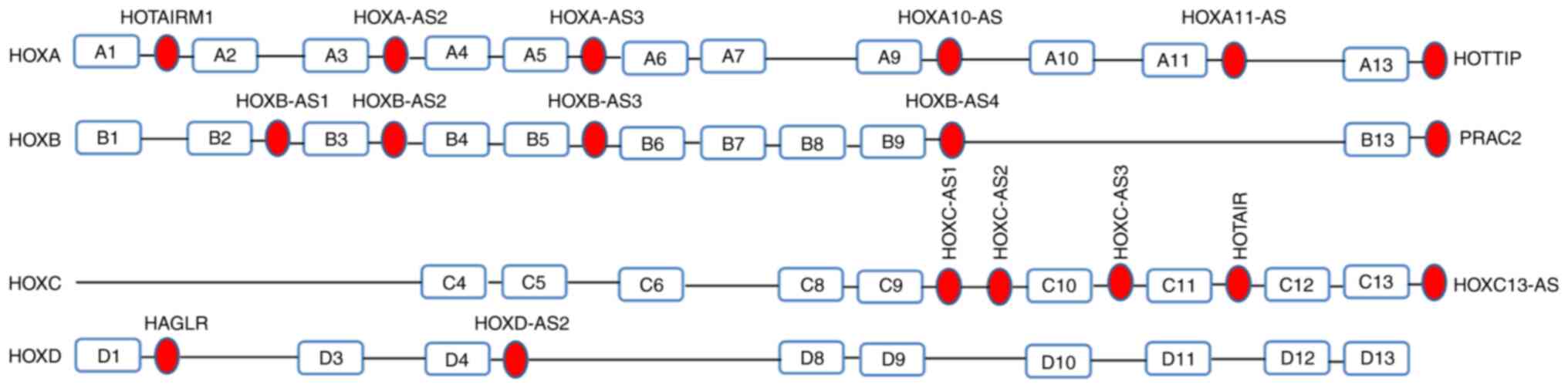
HOX genes are homeobox-containing
protein-coding genes arranged into four HOX clusters at different
chromosomal locations that encode numerous highly conserved lncRNAs
(17,22). In humans, 18 antisense RNA genes,
within the four HOX gene clusters, are deposited in the
National Center for Biotechnology Information (NCBI) GeneBank
database (https://www.ncbi.nlm.nih.gov/). The HOXA gene
contains six HOXA antisense RNA genes, five HOXB and
HOXC regions, and two HOXD regions (Fig. 1). HOX-lncRNAs have multifaceted
roles in mediating the expression of HOX and non-HOX genes
(23). Similar to diverse
lncRNAs, HOX-lncRNAs govern multiple biological functions,
including cell proliferation, apoptosis and the cell cycle, under
normal or abnormal pathophysiological conditions (17,24). They exert unique effects owing to
their tissue-specific expression in different cellular contexts.
Atherosclerosis occurs mainly through stepwise and multistep
biological processes involving angiogenesis, lipid metabolism,
inflammatory responses, cellular proliferation and apoptosis
(5). Various studies have
indicated that HOX-lncRNAs participate in these processes through
various mechanisms. Therefore, it is important to determine their
roles in cardiovascular biology and atherosclerosis. One-third of
these antisense RNAs are associated with atherosclerosis
development. In the following section, the role of HOX-lncRNAs in
atherosclerosis was reviewed, including lipid metabolism,
inflammatory responses, angiogenesis, and cellular proliferation
and apoptosis, principally focusing on their aberrant expression,
cellular biological functions and mechanisms of action in
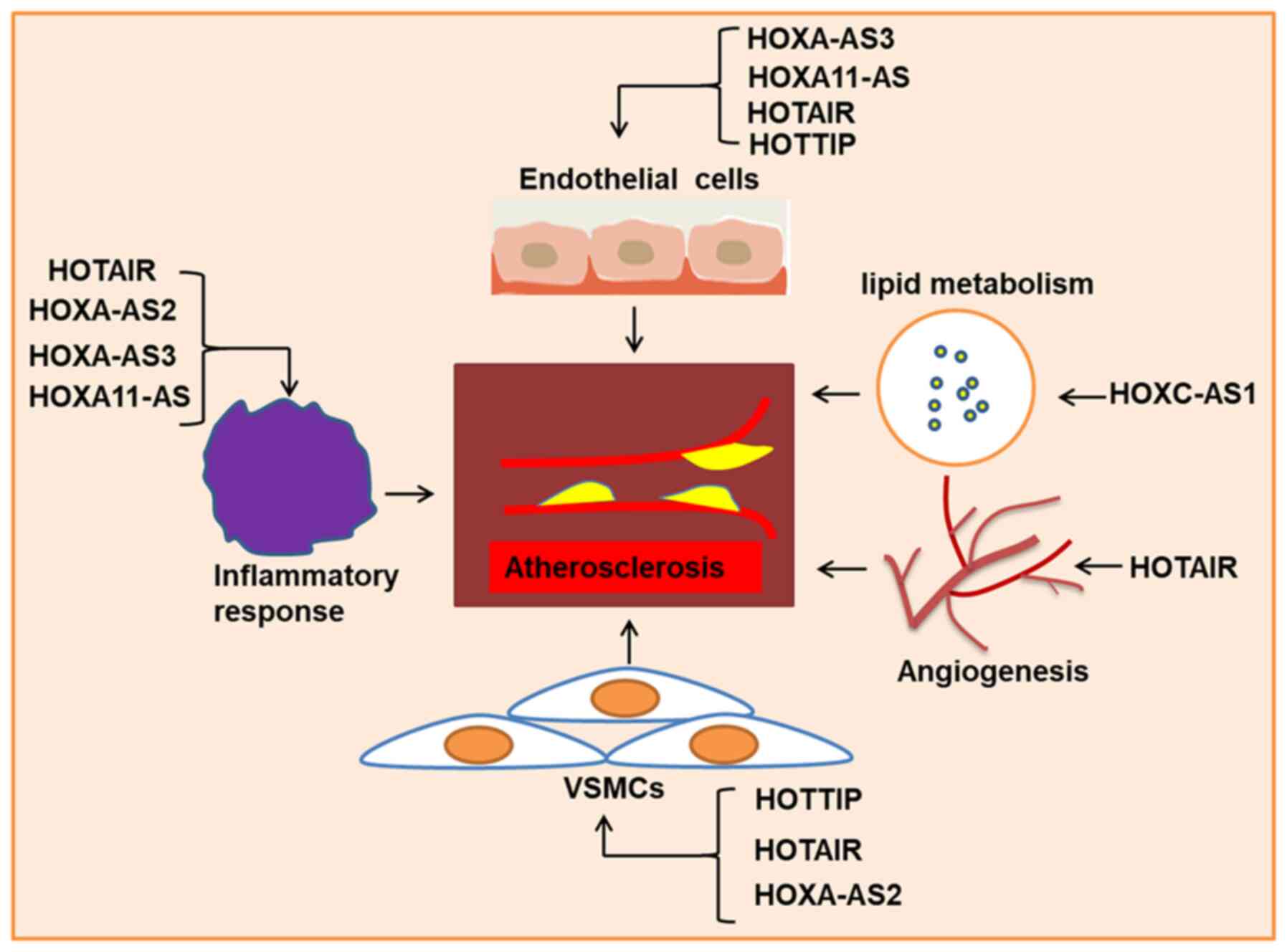
atherosclerosis (Fig. 2 and
Table I).
 | Table IInvolvement of HOX-lncRNAs in diverse
pathophysiological processes in atherosclerosis. |
Table I
Involvement of HOX-lncRNAs in diverse
pathophysiological processes in atherosclerosis.
| Functional
classification | HOX-lncRNA | Model | Biological
functions | Target or
pathway | Mode of action | (Refs.) |
|---|
| Lipid
metabolism | HOXC-AS1 | THP-1
macrophages | ↓ Ox-LDL-induced
cholesterol accumulation | HOXC6 | N.D. | (28) |
| Inflammatory
response | HOTAIR | Raw264.7 cells | ↓ Inflammatory
response | FXR1/NF-κB
pathway | N.D. | (31) |
| Macrophages | ↑ Inflammatory
response | miR-330-5p | ceRNA | (32) |
| Macrophages | ↓ Inflammatory
response | NF-κB pathway | N.D. | (40) |
| H9C2 cells | ↑ Inflammatory
response | miR-138/NF-κB
pathway | ceRNA | (33) |
| H9C2 cells | ↓ H/R-induced
inflammatory | NF-κB pathway | N.D. | (36) |
| HOXA-AS2 | HAEC/HUECs | ↓ Inflammatory
response | NF-κB pathway | N.D. | (37) |
| HOXA-AS3 | HUVECs | ↑ Endothelium
inflammation | NF-κB pathway | N.D. | (38) |
| HOXA11-AS | VSMCs/VECs | ↑ Endothelium
inflammation | PI3K/AKT
pathway | N.D. | (39) |
| Angiogenesis | HOTAIR | ECs | ↓ Proliferation,
migration, and tube formation | VEGFA | N.D. | (43) |
| hBMVECs | ↓ Migratory and
tube formation | EZH2 | N.D. | (45) |
| EC proliferation or
apoptosis | HOXA-AS3 | HUVECs/mouse | ↑
Atherosclerosis | miR-455-5p/p27
Kip1 | ceRNA | (49) |
| HOXA11-AS | HUVECs/mouse | ↑
Atherosclerosis |
miR-515-5p/ROCK1 | ceRNA | (51) |
| HOTAIR | ECs | ↑ Proliferation; ↓
Apoptosis | PI3K/AKT-IRF1
pathway | N.D. | (52) |
| HOTTIP | ECs | ↑ Proliferation and
migration | Wnt/β-catenin
pathway | N.D. | (53) |
| VSMCs proliferation
or apoptosis | HOTTIP | HAVSMCs | ↑ Proliferation and
migration |
miR-490-3p/HMGB1/PI3K-AKT | ceRNA | (54) |
| HOTAIR | VSMCs | ↓ Proliferation; ↑
Apoptosis |
miRNA-130b-3p/PPARα | ceRNA | (55) |
| VSMCs | ↑ Viability and
migration | miR-148b-3p | ceRNA | (56) |
| HOXA-AS2 | VSMCs | ↑ Proliferation; ↓
Apoptosis | miRNA-877-3p | ceRNA | (57) |
| VSMCs | ↑
Proliferation |
miR-520d-3p/IGF2BP3 | ceRNA | (58) |
HOX-lncRNA-mediated lipid metabolism may
drive atherosclerosis
Lipid metabolism disorders are recognized as key
risk factors for atherosclerosis. Macrophages and vascular smooth
muscle cells (VSMCs) engulf the accumulated lipoproteins
[particularly oxidized low-density lipoprotein (ox-LDL)] in the
damaged artery wall and finally transform into foam cells (25). At the same time, the accumulation
of ox-LDL may result in the formation of lipid streaks and even
lipid plaques, thus accelerating the development of
atherosclerosis. Several studies have indicated that HOX-lncRNAs
participate in lipid metabolism. Huang et al (26) reported that lncRNA HOXC cluster
antisense RNA 1 (HOXC-AS1) expression is downregulated in carotid
atherosclerosis. Subsequent research has shown that HOXC-AS1
overexpression partly blocks the cholesterol accumulation induced
by ox-LDL in THP-1 macrophages (26), which may provide insight into the
regulation of cholesterol homeostasis and suggests that HOXC-AS1 is
a promising therapeutic target that governs atherosclerosis.
Another study indicated that HOXB-AS3 regulates lipid metabolism in
endometrial cancer (27). Gariani
and Jornayvaz (28) revealed that
HOTAIR knockdown decreases total cholesterol and triglyceride
levels. Overall, these data preliminarily show that HOX-lncRNAs
have a causal role in lipid metabolism.
HOX-lncRNAs as key players in the
inflammatory response
The inflammatory response is well-known to have a
predominant role in driving the pathogenesis of atherosclerosis
(1). Several mechanisms
underlying the inflammatory activation of endothelial cells (ECs),
foam cell formation and foam cell secretion of inflammatory
cytokines contribute to the development of atherosclerosis
(29). The persistent activation
of the inflammatory response leads to increased EC permeability and
higher rates of lipid entry, thus fueling the development of
atherosclerosis. Overwhelming findings have displayed the causal
effect of HOX-lncRNAs in the inflammatory response. For instance,
one of the most studied HOX cluster-embedded lncRNAs is HOTAIR,
which resides between HOXC11 and HOXC12 in the HOXC
cluster on human chromosome 12q13.13 (30). HOTAIR overexpression attenuates
the expression of pro-inflammatory cytokines (TNF-α and IL-1β) and
potentiates the expression of anti-inflammatory cytokines (IL-4 and
IL-10) in Raw264.7 cells exposed to ox-LDL (31). Likewise, another study also
reported that HOTAIR facilitates oxidative stress and the
inflammatory response by sponging miR-330-5p in human macrophages
in the presence of ox-LDL (32).
In addition, Lu et al (33) found that HOTAIR expression was
upregulated in an AMI group compared to that in its counterparts.
Functionally, HOTAIR overexpression boosts the secretion of
inflammatory cytokines, such as IL-6 and TNF-α, in H9C2 cells in
the presence of a hypoxia stimulus (33). Nuclear factor-κB (NF-κB), as a
major transcription factor involved in inflammatory responses, was
first discovered in 1986 (34).
NF-κB activators and NF-κB-mediated genes have been determined to
participate directly or indirectly in the pathogenesis of
atherosclerosis (35). HOTAIR
modulates the inflammatory response and oxidative stress in H9C2
cells by affecting the NF-κB pathway, thus protecting
cardiomyocytes (36). As a
similar regulatory mechanism, it was also reported that lncRNA HOXA
cluster antisense RNA 2 (HOXA-AS2)-mediated endothelium protection
is partly attributed to inhibition of the NF-κB pathway (37). Another study showed that HOXA-AS3
can interact with NF-κB and positively regulate its activity by
modulating the NF-κB inhibitor protein IκBα (38). HOXA11-AS knockdown markedly
inhibits the expression of inflammation-related genes induced by
TNF-α in VSMCs and by platelet-derived growth factor in vascular
endothelial cells (VECs) (39).
In addition, HOTAIR expression is upregulated in macrophages upon
exposure to lipopolysaccharide (LPS). Furthermore, HOTAIR depletion
diminishes NF-κB-mediated inflammatory gene and cytokine
expression. Further mechanistic studies have indicated that HOTAIR
knockdown leads to reduced expression of NF-κB target genes via a
decrease in the recruitment of NF-κB and associated cofactors at
the target gene promoters (40).
The inflammatory response normally requires high energy and, hence,
is closely related to glucose metabolism. One study showed that
HOTAIR has key roles in the expression of glucose transporter
isoform 1, which controls glucose uptake by macrophages (41). Ultimately, in LPS-induced H9C2
cells, HOTAIR promotes the inflammatory response and apoptosis by
enhancing programmed cell death 4 stability. In vivo
experiments verified that HOTAIR knockdown alleviates cardiac
injury and the secretion of inflammatory factors caused by sepsis
(42). These studies suggest that
HOTAIR has a significant role in mediating inflammatory responses.
In conclusion, HOX-lncRNAs have important roles in inflammatory
responses through several mechanisms. Ongoing research and the
increasing awareness of HOX-lncRNAs will help to elucidate the
important role of HOX-lncRNAs in inflammation and
inflammation-mediated atherosclerosis.
HOX-lncRNA regulation:
Angiogenesis-driven atherosclerosis
EC dysfunction is a primary contributor, driven by
pathological angiogenesis within the arterial wall, to the
pathogenesis of atherosclerosis (43). Angiogenesis markedly affects
plaque growth and the instability of atherosclerotic lesions.
Important research has shown that HOTAIR is involved in
angiogenesis, and its overexpression resulted in a substantial
reduction in EC proliferation, migration and tube formation.
Mechanistically, HOTAIR transcriptionally inhibits vascular
endothelial growth factor (VEGF) via direct binding to its
promoter (44). VEGF is a crucial
factor associated with angiogenesis (45). Furthermore, a recent study
demonstrated that HOTAIR knockdown enhances the migratory and tube
formation abilities of oxygen-glucose
deprivation/reperfusion-induced human brain microvascular ECs
(46). In summary, these data
demonstrate a HOTAIR-mediated anti-angiogenic effect, thus blocking
the progression of atherosclerosis. However, the role of
HOX-lncRNAs in angiogenesis remains unclear and further in-depth
studies on this topic will improve the current understanding of
angiogenesis relevant to atherosclerosis.
Modulation of HOX-lncRNAs in cellular
proliferation and apoptosis
VSMCs and VECs are the main cell types involved in
atherosclerosis, and their abnormal proliferation and apoptosis
have a key role in the development and progression of
atherosclerosis (47). Given that
the development of atherosclerosis comprises the orchestrated
interplay between ECs and SMCs, the following chapters outline the
contribution of HOX-lncRNAs to cellular proliferation and
apoptosis.
ECs
Dysfunction of the endothelial lining of blood
vessels is a critical event in atherosclerosis initiation (48). As such, abnormal EC proliferation
and apoptosis contribute to this disease (49). Mounting evidence indicates that
HOX-lncRNAs are closely related to the proliferation and apoptosis
of VSMCs and VECs, suggesting an important role for HOX-lncRNAs in
the process of atherosclerosis. HOXA-AS3 expression is upregulated
in human umbilical vein ECs (HUVECs) upon ox-LDL stimulation, and
its depletion significantly suppresses the progression of
atherosclerosis. Mechanistically, it acts as a competing endogenous
(ceRNA) for miR-455-5p to decrease the protein level of
p27Kip1 (50), a cell
cycle regulator first identified as a cyclin-dependent kinase
antagonist (51). Similarly, the
expression of HOXA11-AS is increased in both atherosclerotic mouse
aortic tissue and ox-LDL-stimulated HUVECs. HOXA11-AS knockdown
markedly blunts the ox-LDL-induced inhibitory effect on cell
proliferation and diminished apoptosis in HUVECs, suggesting that
HOXA11-AS sponges miR-515-5p to stimulate the expression of
rho-associated coiled-coil containing protein kinase 1 (ROCK1), a
direct target of miR-515-5p, thus contributing to atherosclerotic
injury by directly regulating the miR-515-5p/ROCK1 axis (52). In summary, these observations
indicate that HOX-lncRNAs affect the proliferation and apoptosis of
VECs through miRNA sponging mechanisms to modulate their target
genes. Beyond functioning as ceRNA machinery, HOX-lncRNAs can also
modulate the proliferation and migration of ECs by regulating
related signaling pathways. An increasing number of studies have
shown that HOTAIR has a significant role in cardiovascular biology
and diseases. Specifically, its expression is much lower in ECs
from atherosclerotic plaques. Functional assays further indicated
that HOTAIR facilitates proliferation and migration and suppresses
apoptosis in ECs. Mechanistically, HOTAIR, activated by thymic
stromal lymphopoietin, regulates the proliferation and migration of
ECs via the PI3K/AKT-interferon regulatory factor 1 pathway
(53). Finally, HOTTIP expression
was reported to be higher in coronary artery disease tissues than
in normal tissues, and its depletion was observed to block EC
proliferation and migration. Further mechanistic studies indicated
that HOTTIP may govern EC proliferation and migration via
Wnt/β-catenin pathway activation (54). In addition, HOTAIR facilitates
pulmonary VEC apoptosis via the DNMT1-mediated hypermethylation of
the Bcl-2 promoter in chronic obstructive pulmonary disease
(55). Overall, the increasing
evidence presented here underscores the important contribution of
HOX-lncRNAs to EC functions, particularly in human EC models.
VSMCs
The proliferation and migration of VSMCs are key
events in the progression of atherosclerotic lesions and
restenosis. Several studies have investigated the potential
functions of HOX-lncRNAs in VSMC proliferation and migration.
HOTTIP is a well-characterized HOX-lncRNA that is induced in human
aortic VSMCs (HAVSMCs) in response to ox-LDL and in the sera of
patients with atherosclerosis. Functionally, HOTTIP knockdown
suppresses the ox-LDL-induced proliferation and migration of
HAVSMCs. Mechanistically, HOTTIP depletion blocks cell
proliferation and migration through the regulation of the
miR-490-3p/HMGB1 and PI3K-AKT pathways in ox-LDL-induced HAVSMCs
(56). Given the recent interest
in the interplay between proliferative pathways and atherosclerosis
development, HOTAIR may be a prospective target for atherosclerosis
interventions. Xue et al (57) revealed that HOTAIR overexpression
facilitates viability and suppresses apoptosis in VSMCs.
Mechanistically, HOTAIR serves as a ceRNA for miR-130b-3p to
diminish the expression of peroxisome proliferators-activated
receptors α (57). In addition,
HOTAIR binds to and negatively regulates miR-148b-3p, leading to
increased VSMC viability and migration (58). The lncRNA HOXA-AS2, located
between and antisense to human HOXA3 and HOXA4, has
been reported to potentiate the proliferative and migratory
abilities and decrease the apoptosis of VSMCs by absorbing
miR-877-3p (59). Recently,
HOXA-AS2 was found to be highly expressed in thoracic aortic
aneurysm (TAA) tissues. HOXA-AS2 upregulates the expression of NHS
like 3 by targeting miR-520d-3p/IGF2BP3 to drive VSMC growth in TAA
(60). Therefore, HOX lncRNAs are
important regulators of VSMC proliferation and migration. However,
further investigations are necessary to determine the significance
of HOX-lncRNAs in regulating VSMC proliferation.
3. HOX-lncRNAs and other aspects of
cardiovascular biology
Beyond the discussion of relevant events concerning
HOX-lncRNAs described previously herein, HOX-lncRNAs have been
implicated in other aspects of cardiovascular biology associated
with diverse cardiovascular diseases. Given that the role of
HOX-lncRNAs in cardiovascular diseases has been increasingly
confirmed in recent years, in addition to discussing the function
of HOX-lncRNAs in atherosclerosis, the following section reviews
the relevance of HOX-lncRNAs to other cardiovascular processes,
such as cardiac hypertrophy, fibrosis, myocardial I/R injury,
cardiomyocyte apoptosis, essential hypertension and pulmonary
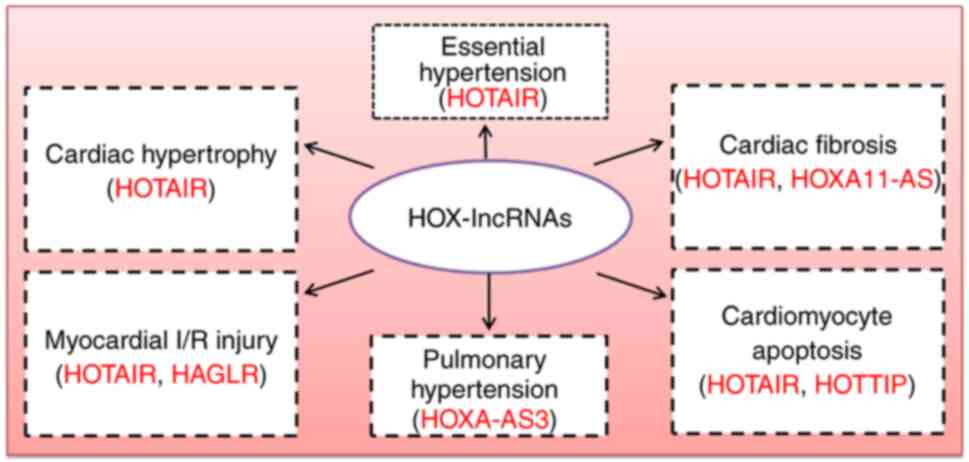
hypertension (Fig. 3 and Table II). These studies are expected to
reveal a more profound molecular landscape of HOX-lncRNAs involved
in cardiovascular dysfunction.
| Table IIHOX-lncRNAs in other cardiovascular
biology. |
Table II
HOX-lncRNAs in other cardiovascular
biology.
| Functional
classification | HOX-lncRNA | Model | Biological
functions | Target or
pathway | Mode of action | (Refs.) |
|---|
| Cardiac
hypertrophy | HOTAIR | Cardiomyocytes | ↓ Cardiac
hypertrophy | miR-19/PTEN | ceRNA | (59) |
| Cardiac
fibrosis | HOTAIR | Fibroblasts | ↑ Myocardial
fibrosis | Wnt signaling
pathway | N.D. | (60) |
|
Fibroblasts/mice | ↑ Myocardial
fibrosis | Wnt signaling
pathway | N.D. | (61) |
| Fibroblasts | ↑ Myocardial
fibrosis | miR-124 | ceRNA | (62) |
| Myocardial I/R
injury | HOTAIR | H9c2
cells/mice | ↑ Myocardial I/R
injury | miR-126 | ceRNA | (64) |
| HAGLR | mice | ↑ Myocardial I/R
injury |
miR-133a-3p/MAPK1 | ceRNA | (70) |
| Cardiomyocyte
apoptosis | HOTAIR | H9c2 | ↓ Apoptosis | miR130a3p/MDM4 | ceRNA | (72) |
| H9c2 | ↑ Cell viability; ↓
Apoptosis | miR-206/FN1 | ceRNA | (73) |
| HOTTIP | Cardiomyocytes | ↓ Apoptosis after
HOTTIP knockdown |
miR-92a-2/c-Met | ceRNA | (19) |
| Pulmonary
hypertension | HOXA-AS3 | HPASMCs | ↑ Growth and
migration; ↓ Apoptosis | miR-675-3p | ceRNA | (83) |
HOX-lncRNA involvement in cardiac
hypertrophy and fibrosis
Cardiac hypertrophy is an initial adaptive response
to various stresses, including pressure or volume overload, which
lowers the increase in the wall tension and aids in maintaining
cardiac output. This adaptive process is beneficial and can
ameliorate cardiac functions; however, persistent exposure of the
heart to increased workload leads to impaired blood flow, resulting
in relative hypoxia and the subsequent loss of cardiomyocytes,
ultimately causing HF. Cardiac fibrosis is closely associated with
numerous heart diseases, such as chronic HF, MI, malignant
arrhythmia and sudden cardiac death, and is a key pathological
feature of cardiac remodeling.
Lai et al (61) reported that HOTAIR expression is
downregulated in heart tissues from transverse aortic
constriction-operated mice in vivo and in cultures treated
with Ang-II in vitro. Its overexpression reduces the cell
surface area and expression of the hypertrophic markers atrial
natriuretic peptide, brain natriuretic peptide and β-myosin heavy
chain in response to Ang-II. Regarding its molecular mechanisms in
cardiac hypertrophy, HOTAIR may act as a ceRNA for miR-19, thereby
modulating its target PTEN and playing an important role in
inhibiting cardiac hypertrophy progression (61). Therefore, HOTAIR may have a key
role in the regulation of cardiac hypertrophy. Ang-II upregulates
HOTAIR expression in cardiac fibroblasts (CFs), which promotes cell
proliferation and inhibits apoptosis. The mechanism by which HOTAIR
regulates myocardial fibrosis may be related to activation of the
Wnt signaling pathway by targeting uRI1 prefoldin like chaperone
(62). Similarly, Tan et
al (63) found that HOTAIR
silencing significantly diminishes Ang-II-induced proliferation,
migration and fibrosis in fibroblasts. Moreover, HOTAIR knockdown
was found to markedly inhibit fibrosis in the heart tissues of
atrial fibrillation (AF)-model mice via the regulation of Wnt
signaling. Recently, Jiang et al (64) showed that HOTAIR functions as a
ceRNA of miR-124 to promote myocardial fibrosis. Therefore, HOTAIR
may have a role in cardiac fibrosis. In addition, another study
showed that HOXA11-AS overexpression in CFs enhances the expression
of transforming growth factor β1, a classic and powerful fibrogenic
pathway mediator pertinent to cardiac fibrosis, thus contributing
to cardiac fibrosis progression (65). Collectively, HOX-lncRNAs, such as
HOTAIR and HOXA11-AS, appear to have prominent roles in regulating
cardiac hypertrophy and fibrosis.
Regulation of myocardial I/R injury by
HOX-lncRNAs
I/R injury is a major cause of the necrotic,
apoptotic and autophagic death of cardiomyocytes. In recent years,
studies have shown that HOX-lncRNAs have a protective effect
against myocardial I/R injury. For instance, Sun and Hu (66) found that HOTAIR expression is
upregulated in mice after I/R and in
H2O2-induced H9c2 cells. Overexpression of
HOTAIR markedly suppressed viability and increased lactate
dehydrogenase release and caspase-3 activity in H9c2 cells treated
with H2O2, and further mechanistic studies
implied that HOTAIR aggravates myocardial I/R injury by sponging
miR-126 to upregulate serine and arginine rich splicing factor 1
expression (66). Furthermore,
propofol (PPF) has certain protective effects against myocardial
I/R injury (67). Chen et
al (68) showed that PPF
pretreatment markedly upregulated HOTAIR expression and that HOTAIR
knockdown partially reversed the protective effects of PPF on
myocardial I/R injury. Likewise, HOTAIR, as a ceRNA, affects
myocardial I/R injury via a HOTAIR/miRNA-1/connexin-43 axis
(69). In conclusion, these data
suggest that HOTAIR is of paramount importance for myocardial I/R
injury. In addition to HOTAIR, expression of HOXD antisense
growth-associated lncRNA (HAGLR), also known as HOXD-AS1 and Mdgt,
whose gene is located in the intergenic region between HOXD1
and HOXD3, has been reported to be significantly increased
upon I/R in vivo or hypoxia-reoxygenation in vitro.
HAGLR knockdown attenuated myocardial I/R injury, and
mechanistically, HAGLR increases myocardial I/R injury by
inhibiting miR-133a-3p, thereby promoting MAPK1 expression
(70). Taken together, HOTAIR and
HAGLR may be therapeutic targets for the treatment of myocardial
I/R injury and much work is needed to fully understand how
HOX-lncRNAs coordinate myocardial I/R injury.
Involvement of HOX-lncRNAs in
cardiomyocyte apoptosis
Cardiomyocyte apoptosis, or programmed cell death,
is a pivotal pathological manifestation of I/R injury and the
primary cause of cardiac dysfunction (71). Therefore, an in-depth
understanding of the mechanism underlying cardiomyocyte apoptosis
is key to preventing myocardial injury and treating heart disease.
Recently, HOX-lncRNAs were reported to be involved in cardiomyocyte
apoptosis. For instance, HOTAIR, which is significantly
downregulated in the ischemic myocardium of rats, inhibits H9c2
cell apoptosis induced by H2O2 in
vitro. Mechanistically, HOTAIR binds to miR-130a-3p and acts as
a sponge to block its functions in I/R injury (72). Similar results were obtained when
determining the role of HOTAIR in AMI. Its expression was found to
be substantially downregulated in the serum of patients with AMI
and in mice. Furthermore, HOTAIR overexpression was found to
promote H9c2 cell viability and inhibit apoptosis under hypoxic
conditions in vitro. The mechanism was attributed to miR-206
binding fibronectin 1 (73). In
addition, HOTTIP levels were determined to be significantly
upregulated in the ischemic myocardium of mice with MI and in
hypoxia-induced cardiomyocytes. Furthermore, HOTTIP functions as a
ceRNA of miR-92a-2 to enhance c-Met expression and is involved in
the modulation of cardiomyocyte growth and apoptosis, indicating
that it is a potential therapeutic target for AMI (19). Ongoing investigations are needed
to further determine their relevant roles.
HOX-lncRNAs and essential
hypertension
Essential hypertension is a common and frequently
occurring disease and a risk factor for various cardiovascular and
cerebrovascular diseases. In recent years, the search for ncRNAs
has attracted the attention of many researchers. According to
reports, increasing studies have shown that lncRNAs have important
roles in the occurrence and development of essential hypertension
(74-78). As a special type of lncRNA, the
roles of HOX-lncRNAs in essential hypertension have rarely been
studied. However, studies have shown that HOTAIR expression is
significantly downregulated in stroke patients with hypertension
compared to that in stroke patients without hypertension (79), suggesting that it may be closely
related to the occurrence and development of hypertension and that
it could be used as a biomarker for the diagnosis and treatment of
this condition.
HOX-lncRNAs and pulmonary
hypertension
Pulmonary arterial hypertension (PAH) is a vascular
remodeling disease characterized by vasoconstriction and
progressive obliteration of the distal pulmonary arteries, leading
to elevated pulmonary pressure and eventually right ventricular HF
(80,81). Pulmonary vascular remodeling is
characterized by the excessive proliferation of pulmonary arterial
smooth muscle cells (PASMCs) and the dysfunction of pulmonary
arterial ECs (82). Therefore, it
is important to explore the cellular mechanisms underlying the
effects of HOX-lncRNAs in pulmonary VSMCs and in the pathogenesis
of pulmonary hypertension. HOXA-AS3 is a novel lncRNA that is
transcribed from the HOXA cluster. A previous study showed
increased HOXA-AS3 levels in human (H)PASMCs exposed to hypoxia.
Furthermore, its knockdown decreased growth and migration and
induced apoptosis in HPASMCs. The mechanism associated with the
effects of HOXA-AS3 was also examined; it was found to upregulate
phosphodiesterase 5A expression by sponging miR-675-3p (83). Another study showed that HOXA-AS3
knockdown represses PASMC proliferation in vitro (84), thus providing a potential novel
strategy for the treatment of PAH.
4. HOX-lncRNAs as potential diagnostic
biomarkers for cardiovascular disease
Given the significant recent studies demonstrating
the role of HOX-lncRNAs in cardiovascular diseases, outlined
previously herein, there may be opportunities to treat these
diseases by targeting these molecules. The cell and tissue
specificity of HOX-lncRNAs makes them potential diagnostic markers
for cardiovascular diseases. Mounting evidence suggests that
HOX-lncRNAs are differentially expressed in various cardiovascular
diseases. For instance, HOTAIRM1, HOXA-AS2 and HOXB-AS2 expression
was found to be significantly downregulated in epicardial adipose
tissue associated with AF using RNA sequencing (85). Likewise, HAGLR and HOTAIRM1 have
robust diagnostic value for AF, which may facilitate the discovery
of novel diagnostic biomarkers or therapeutic targets (86). Studies have indicated that HOTAIR
can be used as a biomarker for the diagnosis of congenital heart
disease (87) and AMI (88). Furthermore, another study
indicated that the expression of HOTAIR is significantly
downregulated with chronic HF (89). Finally, HOXA11-AS expression is
downregulated in the plasma of patients with in-stent restenosis
(ISR) compared to that in patients without ISR, and it was found to
be an independent protective factor for ISR (90). In summary, the large amounts of
data presented provide evidence for the use of HOX-lncRNAs as
diagnostic markers of cardiovascular diseases, and future in-depth
studies of these markers are expected to contribute to their use as
potential therapeutic targets. However, more studies regarding
HOX-lncRNAs are essential to affirm their feasibility as diagnostic
biomarkers.
5. Concluding remarks
For decades, investigations into cardiovascular
biology have primarily highlighted the involvement of
protein-coding genes. Recently, a new class of molecules, termed
lncRNAs, has been shown to have pivotal regulatory roles in
coordinating multiple cellular functions, coupled with a high
degree of tissue and cell specificity, making them promising target
candidates for clinical practice. LncRNAs have also emerged as
potential modulators of atherosclerosis and other cardiovascular
events. Recently, a unique class of functional molecules,
HOX-lncRNAs, has emerged. Of note, the modulation of HOX-lncRNAs
has an impact on diverse aspects cardiovascular biology to a
certain extent. The present review focused on the contributions of
HOX-lncRNAs to atherosclerosis and the key biological processes
relevant to cardiovascular problems, including cardiac hypertrophy,
fibrosis, myocardial I/R injury, cardiomyocyte apoptosis, pulmonary
hypertension and essential hypertension.
First, the research progress associated with
HOX-lncRNAs in atherosclerosis was discussed, including abnormal
lipid metabolism, inflammatory responses, angiogenesis and the
proliferation and apoptosis of vascular cells. In view of this
research, HOX-lncRNAs have broad prospects in atherosclerosis,
indicating that they may serve as biomarkers or targets for the
diagnosis and treatment of this disease. The present review then
continued to explore the effects of HOX-lncRNAs on cardiomyocyte
hypertrophy, cardiomyocyte apoptosis, myocardial fibrosis,
myocardial I/R injury, essential hypertension and pulmonary
hypertension, indicating that HOX-lncRNAs have an important role in
these cardiovascular events. This report also provides a
theoretical foundation for future research. In terms of the
mechanism of action, it was indicated that HOX-lncRNAs can regulate
the expression of downstream target genes as ceRNAs, via
RNA-binding proteins and through epigenetic mechanisms.
Transcription factors can bind to the promoter region of
HOX-lncRNAs, thereby activating or inhibiting their transcription.
In the present review, HOX-lncRNAs were mainly considered to be
involved in the regulation of gene expression through ceRNA
mechanisms. Other related upstream and downstream regulatory
mechanisms have not yet been reported.
A preponderance of research suggests that
HOX-lncRNAs are vital mediators with multifaceted roles in the
pathogenesis of atherosclerosis and other cardiovascular diseases.
This report provides insight into the regulation and function of
HOX-lncRNAs in atherosclerosis and other cardiovascular diseases.
However, given that the investigation of HOX-lncRNAs still seems to
be preliminary, with deeper insight into their functions and
mechanisms needed, work in this field appears to have merely
scratched the surface. Consequently, a further in-depth
understanding of how HOX-lncRNAs govern atherosclerosis could be
very informative for the development of novel diagnostic and
therapeutic strategies. The increasingly substantial data on the
role of HOX-lncRNAs in the regulation of cardiovascular dysfunction
will help to understand atherosclerosis or atherosclerotic
cardiovascular disease.
6. Future outlook and challenge
To date, several HOX-lncRNAs have been identified as
having a role in atherosclerosis, potentially providing a promising
direction for numerous novel research areas pertaining to the
cardiovascular system. However, to unequivocally understand the
roles of HOX-lncRNAs in atherosclerosis, numerous important
pressing questions remain.
Based on the detailed overview of the role of
HOX-lncRNAs in atherosclerosis and cardiovascular biology, it is
not difficult to see that the knowledge of the regulatory effects
of HOX-lncRNAs on atherosclerosis is still not complete, with the
focus of research in this area being principally concentrated on
in vitro culture experiments and less known concerning such
roles in vivo. Moreover, the mechanisms underlying the
effects of HOX-lncRNAs are mainly limited to their role as ceRNAs,
and other mechanisms, such as RNA-RNA, RNA-DNA and RNA-protein
interactions have not yet been characterized. The upstream
regulatory circuits of HOX-lncRNAs also remain elusive and the
reasons for their abnormal expression in atherosclerosis remain
largely unexplored. In addition, in terms of HOX-lncRNAs, HOTAIR
has been most extensively studied in the cardiovascular field;
however, other HOX-lncRNAs have not been studied in detail.
Consequently, HOTAIR is likely to be a biomarker for the diagnosis
or treatment of cardiovascular diseases, but further research is
needed to investigate this function. Meanwhile, abundant
investigations will be launched on HOX-lncRNAs, and their functions
in cardiovascular diseases have been rigorously assessed using the
most robust approaches available, and these HOX-lncRNAs may have
unique and critical roles in the cardiovascular field.
The expression levels of multiple lncRNAs are
relatively low, which leads to uncertainties regarding the
reliability and reproducibility of large-scale lncRNA
investigations. Therefore, a novel platform for HOX-lncRNA capture
and quantification is urgently required to enhance the efficiency
of the detection of HOX-lncRNA expression.
The roles of more than half of the HOX-lncRNAs in
atherosclerosis, i.e., HOXA10-AS, HOXB-AS1, HOXB-AS3, HOXB-AS4,
PRAC2, HOXC-AS2, HOXC-AS3, HOXC13-AS and HOXD-AS2, are currently
unclear. Therefore, the further investigation of HOX-lncRNAs in
atherosclerosis is a prospective area for future research.
Eventually, and perhaps the most key and unaddressed
issue in this discussion, is how to translate these findings
regarding HOX-lncRNAs from seemingly far-reaching fundamental
insight into daily clinical practice. One challenge remains in
terms of the conservation of HOX-lncRNAs across species; hence, the
functional significance of substantially defined HOX-lncRNAs in
atherosclerosis is frequently unclear. A wealth of clinical trials
is also necessary to address off-target effects and determine the
safety and efficacy of targeting HOX-lncRNAs. Integrated approaches
based on multiomics and multilevel data are prerequisites for
transforming molecular findings into clinical practice to treat
cardiovascular diseases, including atherosclerosis. Addressing
these pressing questions promises to yield pivotal insight into not
only atherosclerosis, but also the increasingly sophisticated
cardiovascular events with which it is associated. The insight on
this horizon will advance our understanding of diverse HOX-lncRNAs,
thereby enabling the information gained from the study of one
HOX-lncRNA to be more reliably leveraged to further understand
numerous other related molecules, ultimately providing a firm grasp
of the number of the thousands of HOX-lncRNA genes found in the
cell that are functional.
Availability of data and materials
Not applicable.
Authors' contributions
YZ wrote the manuscript. QW provided the research
direction. All authors have read and approved the final manuscript.
Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
AF
|
atrial fibrillation
|
|
AMI
|
acute myocardial infarction
|
|
ceRNA
|
competing endogenous RNA
|
|
CFs
|
cardiac fibroblasts
|
|
EC
|
endothelial cell
|
|
HAVSMCs
|
human aortic VSMCs
|
|
HF
|
heart failure
|
|
HOX
|
homeobox
|
|
HOXC-AS1
|
lncRNA HOXC cluster antisense RNA
1
|
|
HOXA-AS3
|
lncRNA HOXA cluster antisense RNA
3
|
|
HPASMCs
|
human pulmonary artery smooth muscle
cells
|
|
HUVECs
|
human umbilical vein ECs
|
|
I/R
|
ischemia/reperfusion
|
|
ISR
|
in-stent restenosis
|
|
lncRNA
|
long non-coding RNA
|
|
LPS
|
lipopolysaccharide
|
|
MI
|
myocardial infarction
|
|
NF-κB
|
nuclear factor-κB
|
|
PAH
|
pulmonary arterial hypertension
|
|
PASMCs
|
pulmonary arterial smooth muscle
cells
|
|
PPF
|
propofol
|
|
TAA
|
thoracic aortic aneurysm
|
|
VEGF
|
vascular endothelial growth factor
|
|
VSMCs
|
vascular smooth muscle cells
|
Acknowledgments
Not applicable.
Funding
This study was financially supported by the National Natural
Science Foundation of China (grant no. 82260084), the Guizhou
Provincial Science and Technology Project (grant no.
QKHJC-ZK[2022]YB268) and the Science and Technology fund project of
Health and Family Planning Commission of Guizhou province (grant
no. gzwkj2023-132).
References
|
1
|
Libby P, Buring JE, Badimon L, Hansson GK,
Deanfield J, Bittencourt MS, Tokgözoğlu L and Lewis EF:
Atherosclerosis. Nat Rev Dis Primers. 5:562019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barquera S, Pedroza-Tobías A, Medina C,
Hernández-Barrera L, Bibbins-Domingo K, Lozano R and Moran AE:
Global overview of the epidemiology of atherosclerotic
cardiovascular disease. Arch Med Res. 46:328–338. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
GBD 2017 Causes of Death Collaborators:
Global, regional, and national age-sex-specific mortality for 282
causes of death in 195 countries and territories, 1980-2017: A
systematic analysis for the Global Burden of Disease Study 2017.
Lancet. 392:1736–1788. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Libby P, Ridker PM and Maseri A:
Inflammation and atherosclerosis. Circulation. 105:1135–1143. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Camaré C, Pucelle M, Nègre-Salvayre A and
Salvayre R: Angiogenesis in the atherosclerotic plaque. Redox Biol.
12:18–34. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nowbar AN, Gitto M, Howard JP, Francis DP
and Al-Lamee R: Mortality From Ischemic Heart Disease: Analysis of
Data From the World Health Organization and Coronary Artery Disease
Risk Factors From NCD Risk Factor Collaboration. Circ Cardiovasc
Qual Outcomes. 12:e0053752019. View Article : Google Scholar :
|
|
7
|
Klattenhoff CA, Scheuermann JC, Surface
LE, Bradley RK, Fields PA, Steinhauser ML, Ding H, Butty VL, Torrey
L, Haas S, et al: Braveheart, a long noncoding RNA required for
cardiovascular lineage commitment. Cell. 152:570–583. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pang KC, Frith MC and Mattick JS: Rapid
evolution of noncoding RNAs: Lack of conservation does not mean
lack of function. Trends Genet. 22:1–5. 2006. View Article : Google Scholar
|
|
9
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Haemmig S, Simion V, Yang D, Deng Y and
Feinberg MW: Long Noncoding RNAs in cardiovascular pathology,
diagnosis, and therapy. Curr Opin Cardiol. 32:7762017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ulitsky I and Bartel DP: lincRNAs:
Genomics, evolution, and mechanisms. Cell. 154:26–46. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gong C and Maquat LE: lncRNAs
transactivate STAU1-mediated mRNA decay by duplexing with 3' UTRs
via Alu elements. Nature. 470:284–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoon JH, Abdelmohsen K, Srikantan S, Yang
X, Martindale JL, De S, Huarte M, Zhan M, Becker KG and Gorospe M:
LincRNA-p21 suppresses target mRNA translation. Mol Cell.
47:648–655. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tripathi V, Ellis JD, Shen Z, Song DY, Pan
Q, Watt AT, Freier SM, Bennett CF, Sharma A, Bubulya PA, et al: The
nuclear-retained noncoding RNA MALAT1 regulates alternative
splicing by modulating SR splicing factor phosphorylation. Mol
Cell. 39:925–938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chowdhary A, Satagopam V and Schneider R:
Long non-coding RNAs: mechanisms, experimental, and computational
approaches in identification, characterization, and their biomarker
potential in cancer. Front. Genet. 12:6496192021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Botti G, De Chiara A, Di Bonito M, Cerrone
M, Malzone MG, Collina F and Cantile M: Noncoding RNAs within the
HOX gene network in tumor pathogenesis and progression. J Cell
Physiol. 234:395–413. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang X, Gao Y, Wu H, Mao Y and Qi Y:
LncRNA HOX transcript antisense RNA mitigates cardiac function
injury in chronic heart failure via regulating microRNA-30a-5p to
target KDM3A. J Cell Mol Med. 26:1473–1485. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang B, Ma L and Wang J: LncRNA HOTTIP
Knockdown Attenuates Acute Myocardial Infarction via Regulating
miR-92a-2/c-Met Axis. Cardiovasc Toxicol. 22:352–364. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fan H, Shao H and Gao X: Long Non-Coding
RNA HOTTIP is elevated in patients with sepsis and promotes cardiac
dysfunction. Immunol Invest. 51:2086–2096. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao L, Wang X, Guo S, Xiao L, Liang C,
Wang Z, Li Y, Liu Y, Yao R, Liu Y and Zhang Y: LncRNA HOTAIR
functions as a competing endogenous RNA to upregulate SIRT1 by
sponging miR-34a in diabetic cardiomyopathy. J Cell Physiol.
234:4944–4958. 2019. View Article : Google Scholar
|
|
22
|
Wang Y, Dang Y, Liu J and Ouyang X: The
function of homeobox genes and lncRNAs in cancer. Oncol. Lett.
12:1635–1641. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
De Kumar B and Krumlauf R: HOXs and
lincRNAs: Two sides of the same coin. Sci Adv. 2:e15014022016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dasen JS: Long noncoding RNAs in
development: Solidifying the Lncs to Hox gene regulation. Cell Rep.
5:1–2. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Maguire EM, Pearce SWA and Xiao Q: Foam
cell formation: A new target for fighting atherosclerosis and
cardiovascular disease. Vascul Pharmacol. 112:54–71. 2019.
View Article : Google Scholar
|
|
26
|
Huang C, Hu YW, Zhao JJ, Ma X, Zhang Y,
Guo FX, Kang CM, Lu JB, Xiu JC, Sha YH, et al: Long Noncoding RNA
HOXC-AS1 Suppresses Ox-LDL-Induced Cholesterol Accumulation Through
Promoting HOXC6 Expression in THP-1 Macrophages. DNA Cell Biol.
35:722–729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou Q, Kong D, Li W, Shi Z, Liu Y, Sun R,
Ma X, Qiu C, Liu Z, Hou Y and Jiang J: LncRNA HOXB-AS3 binding to
PTBP1 protein regulates lipid metabolism by targeting SREBP1 in
endometrioid carcinoma. Life Sci. 320:1215122023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gariani K and Jornayvaz FR:
Pathophysiology of NASH in endocrine diseases. Endocr Connect.
10:R52–R65. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Geovanini GR and Libby P: Atherosclerosis
and inflammation: Overview and updates. Clin Sci (Lond).
132:1243–1252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Woo CJ and Kingston RE: HOTAIR lifts
noncoding RNAs to new levels. Cell. 129:1257–1259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pang JL, Wang JW, Hu PY, Jiang JS and Yu
C: HOTAIR alleviates ox-LDL-induced inflammatory response in
Raw264.7 cells via inhibiting NF-κB pathway. Eur Rev Med Pharmacol
Sci. 22:6991–6998. 2018.PubMed/NCBI
|
|
32
|
Liu J, Huang GQ and Ke ZP: Silence of long
intergenic noncoding RNA HOTAIR ameliorates oxidative stress and
inflammation response in ox-LDL-treated human macrophages by
upregulating miR-330-5p. J Cell Physiol. 234:5134–5142. 2019.
View Article : Google Scholar
|
|
33
|
Lu W, Zhu L, Ruan ZB, Wang MX, Ren Y and
Li W: HOTAIR promotes inflammatory response after acute myocardium
infarction by upregulating RAGE. Eur Rev Med Pharmacol Sci.
22:7423–7430. 2018.PubMed/NCBI
|
|
34
|
Sen R and Baltimore D: Inducibility of
kappa immunoglobulin enhancer-binding protein Nf-kappa B by a
posttranslational mechanism. Cell. 47:921–928. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pamukcu B, Lip GY and Shantsila E: The
nuclear factor-kappa B pathway in atherosclerosis: A potential
therapeutic target for atherothrombotic vascular disease. Thromb
Res. 128:117–123. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang G, Wang Q and Xu W: Effects of Long
Noncoding RNA HOTAIR Targeting miR-138 on Inflammatory Response and
Oxidative Stress in Rat Cardiomyocytes Induced by Hypoxia and
Reoxygenation. Dis Markers. 2021:42732742021. View Article : Google Scholar :
|
|
37
|
Zhu X, Liu Y, Yu J, Du J, Guo R, Feng Y,
Zhong G, Jiang Y and Lin J: LncRNA HOXA-AS2 represses endothelium
inflammation by regulating the activity of NF-κB signaling.
Atherosclerosis. 281:38–46. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu X, Chen D, Liu Y, Yu J, Qiao L, Lin S,
Chen D, Zhong G, Lu X, Wang Y, et al: Long Noncoding RNA HOXA-AS3
Integrates NF-κB Signaling To Regulate Endothelium Inflammation.
Mol Cell Biol. 39:e00139–19. 2019. View Article : Google Scholar :
|
|
39
|
Jin QS, Huang LJ, Zhao TT, Yao XY, Lin LY,
Teng YQ, Kim SH, Nam MS, Zhang LY and Jin YJ: HOXA11-AS regulates
diabetic arteriosclerosis-related inflammation via PI3K/AKT
pathway. Eur Rev Med Pharmacol Sci. 22:6912–6921. 2018.PubMed/NCBI
|
|
40
|
Obaid M, Udden SMN, Deb P, Shihabeddin N,
Zaki MH and Mandal SS: LncRNA HOTAIR regulates
lipopolysaccharide-induced cytokine expression and inflammatory
response in macrophages. Sci Rep. 8:156702018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Obaid M, Udden SMN, Alluri P and Mandal
SS: LncRNA HOTAIR regulates glucose transporter Glut1 expression
and glucose uptake in macrophages during inflammation. Sci Rep.
11:2322021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ni SY, Xu WT, Liao GY, Wang YL and Li J:
LncRNA HOTAIR Promotes LPS-Induced inflammation and apoptosis of
cardiomyocytes via Lin28-Mediated PDCD4 Stability. Inflammation.
44:1452–1463. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xiao W, Jia Z, Zhang Q, Wei C, Wang H and
Wu Y: Inflammation and oxidative stress, rather than hypoxia, are
predominant factors promoting angiogenesis in the initial phases of
atherosclerosis. Mol Med Rep. 12:3315–3322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu K, Liu F, Wu W, Chen Y, Wu H and Zhang
W: Long non-coding RNA HOX transcript antisense RNA (HOTAIR)
suppresses the angiogenesis of human placentation by inhibiting
vascular endothelial growth factor A expression. Reprod Fertil Dev.
31:377–385. 2019. View Article : Google Scholar
|
|
45
|
Di Stefano R, Felice F and Balbarini A:
Angiogenesis as risk factor for plaque vulnerability. Curr Pharm
Des. 15:1095–1106. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang Y, Mao J, Li X, Wang B and Zhou X:
lncRNA HOTAIR mediates OGD/R-induced cell injury and angiogenesis
in a EZH2-dependent manner. Exp Ther Med. 23:992022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Davignon J and Ganz P: Role of endothelial
dysfunction in atherosclerosis. Circulation. 109(23 Suppl 1):
III27–III32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sun GB, Qin M, Ye JX, Pan RL, Meng XB,
Wang M, Luo Y, Li ZY, Wang HW and Sun XB: Inhibitory effects of
myricitrin on oxidative stress-induced endothelial damage and early
atherosclerosis in ApoE-/-mice. Toxicol Appl Pharmacol.
271:114–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chi K, Zhang J, Sun H, Liu Y, Li Y, Yuan T
and Zhang F: Knockdown of lncRNA HOXA-AS3 suppresses the
progression of atherosclerosis via Sponging miR-455-5p. Drug Des
Devel Ther. 14:3651–3662. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Choi BK, Fujiwara K, Dayaram T, Darlington
Y, Dickerson J, Goodell MA and Donehower LA: WIP1 dephosphorylation
of p27Kip1 serine 140 destabilizes p27Kip1
and reverses anti-proliferative effects of ATM phosphorylation.
Cell Cycle. 19:479–491. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gao F, Wang XC, Luo ZD, Hu GQ, Ma MQ,
Liang Y, Xu BL and Lin XH: LncRNA HOXA11-AS promotes vascular
endothelial cell injury in atherosclerosis by regulating the
miR-515-5p/ROCK1 axis. ESC Heart Fail. 9:2259–2271. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Peng Y, Meng K, Jiang L, Zhong Y, Yang Y,
Lan Y, Zeng Q and Cheng L: Thymic stromal lymphopoietin-induced
HOTAIR activation promotes endothelial cell proliferation and
migration in atherosclerosis. Biosci Rep. 37:BSR201703512017.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liao B, Chen R, Lin F, Mai A, Chen J, Li
H, Xu Z and Dong S: Long noncoding RNA HOTTIP promotes endothelial
cell proliferation and migration via activation of the
Wnt/β-catenin pathway. J Cell Biochem. 119:2797–2805. 2018.
View Article : Google Scholar
|
|
55
|
Dai Z, Liu X, Zeng H and Chen Y: Long
noncoding RNA HOTAIR facilitates pulmonary vascular endothelial
cell apoptosis via DNMT1 mediated hypermethylation of Bcl-2
promoter in COPD. Respir Res. 23:3562022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Guo X, Liu Y, Zheng X, Han Y and Cheng J:
HOTTIP knockdown inhibits cell proliferation and migration via
regulating miR-490-3p/HMGB1 axis and PI3K-AKT signaling pathway in
ox-LDL-induced VSMCs. Life Sci. 248:1174452020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xue H, Wang B and Xue YS: LncRNA HOTAIR
regulates the proliferation and apoptosis of vascular smooth muscle
cells through targeting miRNA-130b-3p/PPARα axis. Eur Rev Med
Pharmacol Sci. 23:10989–10995. 2019.PubMed/NCBI
|
|
58
|
Shen D, Chen Q, Li J, Wang S, Song H and
Wang F: Clinical Value of Long Non-Coding RNA HOTAIR in carotid
artery stenosis and its role in vascular smooth muscle cell
proliferation. Crit Rev Eukaryot Gene Expr. 33:15–23. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fan TT, Liu YX, Wang XC, Xu BL, Chen ZC,
Lu HA and Zhang M: LncRNA HOXA-AS2 accelerates the proliferation
and migration and inhibits the apoptosis of vascular smooth muscle
cells by absorbing miRNA-877-3p. Eur Rev Med Pharmacol Sci.
24:362–368. 2020.PubMed/NCBI
|
|
60
|
Ou M, Chu Y, Zhang Q, Zhao H and Song Q:
HOXA cluster antisense RNA 2 elevates KIAA1522 expression through
microRNA-520d-3p and insulin like growth factor 2 mRNA binding
protein 3 to promote the growth of vascular smooth muscle cells in
thoracic aortic aneurysm. ESC Heart Fail. 9:2955–2966. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lai Y, He S, Ma L, Lin H, Ren B, Ma J, Zhu
X and Zhuang S: HOTAIR functions as a competing endogenous RNA to
regulate PTEN expression by inhibiting miR-19 in cardiac
hypertrophy. Mol Cell Biochem. 432:179–187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Pan SC, Cui HH and Qiu CG: HOTAIR promotes
myocardial fibrosis through regulating URI1 expression via Wnt
pathway. Eur Rev Med Pharmacol Sci. 22:6983–6990. 2018.PubMed/NCBI
|
|
63
|
Tan W, Wang K, Yang X, Wang K, Wang N and
Jiang TB: LncRNA HOTAIR promotes myocardial fibrosis in atrial
fibrillation through binding with PTBP1 to increase the stability
of Wnt5a. Int J Cardiol. 369:21–28. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jiang J, Xu ST and Ren K: LncRNA HOTAIR
promotes myocardial fibrosis by suppressing miR-124. Int J Cardiol.
374:942023. View Article : Google Scholar
|
|
65
|
Wang J, Liu X, Zhuang Q, Pan R, Zou L, Cen
Z and Tang L: Long noncoding RNA homeobox A11 antisense promotes
transforming growth factor β1induced fibrogenesis in cardiac
fibroblasts. Mol Med Rep. 19:2817–2824. 2019.PubMed/NCBI
|
|
66
|
Sun Y and Hu ZQ: LncRNA HOTAIR aggravates
myocardial ischemia-reperfusion injury by sponging microRNA-126 to
upregulate SRSF1. Eur Rev Med Pharmacol Sci. 24:9046–9054.
2020.PubMed/NCBI
|
|
67
|
Wang B, Wu Q, Liao J, Zhang S, Liu H, Yang
C, Dong Q, Zhao N, Huang Z, Guo K and Du Y: Propofol induces
cardioprotection against ischemia-reperfusion injury via
suppression of transient receptor potential vanilloid 4 channel.
Front Pharmacol. 10:11502019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen J, Li X, Zhao F and Hu Y:
HOTAIR/miR-17-5p axis is involved in the propofol-mediated
cardioprotection against ischemia/reperfusion injury. Clin Interv
Aging. 16:621–632. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu Z, Liu Y, Zhu Y and Gong J:
HOTAIR/miRNA-1/Cx43: A potential mechanism for treating myocardial
ischemia-reperfusion injury. Int J Cardiol. 308:112020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang Z, Luo W, Zhong P, Feng Y and Wang H:
lncRNA HAGLR modulates myocardial ischemia-reperfusion injury in
mice through regulating miR-133a-3p/MAPK1 axis. Open Med (Wars).
17:1299–1307. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Chouchani ET, Pell VR, James AM, Work LM,
Saeb-Parsy K, Frezza C, Krieg T and Murphy MP: A Unifying mechanism
for mitochondrial superoxide production during ischemia-reper
fusion injury. Cell Metab. 23:2542632016. View Article : Google Scholar
|
|
72
|
Fang J, Zheng W, Hu P and Wu J:
Investigating the effect of lncRNA HOTAIR on apoptosis induced by
myocardial ischemia-reperfusion injury. Mol Med Rep. 23:1692021.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yao J, Ma R, Wang C and Zhao G:
LncRNA-HOTAIR inhibits H9c2 apoptosis after acute myocardial
infarction via miR-206/FN1 axis. Biochem Genet. 60:1781–1792. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhao X, Wang C, Liu M, Meng F and Liu K:
LncRNA FENDRR servers as a possible marker of essential
hypertension and regulates human umbilical vein endothelial cells
dysfunction via miR-423-5p/Nox4 Axis. Int J Gen Med. 15:2529–2540.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Li R, Yu X, Chen Y, Xiao M, Zuo M, Xie Y,
Yang Z and Kuang D: Association of lncRNA PVT1 gene polymorphisms
with the risk of essential hypertension in Chinese Population.
Biomed Res Int. 2022:99769092022.PubMed/NCBI
|
|
76
|
Peng W, Cao H, Liu K, Guo C, Sun Y, Qi H,
Liu Z, Xie Y, Liu X, Li B and Zhang L: Identification of
lncRNA-NR_104160 as a biomarker and construction of a
lncRNA-related ceRNA network for essential hypertension. Am J
Transl Res. 12:6060–6075. 2020.PubMed/NCBI
|
|
77
|
Yin L, Yao J, Deng G, Wang X, Cai W and
Shen J: Identification of candidate lncRNAs and circRNAs regulating
WNT3/β-catenin signaling in essential hypertension. Aging (Albany
NY). 12:8261–8288. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Fang G, Qi J, Huang L and Zhao X: LncRNA
MRAK048635_P1 is critical for vascular smooth muscle cell function
and phenotypic switching in essential hypertension. Biosci Rep.
39:BSR201822292019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Ali MA, Shaker OG, Khalifa AA, Ezzat EM,
Elghobary HA, Abdel Mawla TS, Elkhateeb AF, Elebiary AMA and Elamir
AM: LncRNAs NEAT1, HOTAIR, and GAS5 expression in hypertensive and
non-hypertensive associated cerebrovascular stroke patients, and
its link to clinical characteristics and severity score of the
disease. Noncoding RNA Res. 8:96–108. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Lau EMT, Giannoulatou E, Celermajer DS and
Humbert M: Epidemiology and treatment of pulmonary arterial
hypertension. Nat Rev Cardiol. 14:603–614. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhang ZQ, Zhu SK, Wang M, Wang XA, Tong
XH, Wan JQ and Ding JW: New progress in diagnosis and treatment of
pulmonary arterial hypertension. J Cardiothorac Surg. 17:2162022.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Rabinovitch M: Pathobiology of pulmonary
hypertension. Annu Rev Pathol. 2:369–399. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Li ZK, Gao LF, Zhu XA and Xiang DK: LncRNA
HOXA-AS3 Promotes the Progression of Pulmonary Arterial
Hypertension through Mediation of miR-675-3p/PDE5A Axis. Biochem
Genet. 59:1158–1172. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang H, Liu Y, Yan L, Wang S, Zhang M, Ma
C, Zheng X, Chen H and Zhu D: Long noncoding RNA Hoxaas3
contributes to hypoxia-induced pulmonary artery smooth muscle cell
proliferation. Cardiovasc Res. 115:647–657. 2019. View Article : Google Scholar
|
|
85
|
Shi X, Shao X, Liu B, Lv M, Pandey P, Guo
C, Zhang R and Zhang Y: Genome-wide screening of functional long
noncoding RNAs in the epicardial adipose tissues of atrial
fibrillation. Biochim Biophys Acta Mol Basis Dis. 1866:1657572020.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Qian C, Li H, Chang D, Wei B and Wang Y:
Identification of functional lncRNAs in atrial fibrillation by
integrative analysis of the lncRNA-mRNA network based on competing
endogenous RNAs hypothesis. J Cell Physiol. 234:11620–11630. 2019.
View Article : Google Scholar
|
|
87
|
Jiang Y, Mo H, Luo J, Zhao S, Liang S,
Zhang M and Yuan J: HOTAIR is a potential novel biomarker in
patients with congenital heart diseases. Biomed Res Int.
2018:28506572018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Gao L, Liu Y, Guo S, Yao R, Wu L, Xiao L,
Wang Z, Liu Y and Zhang Y: Circulating Long Noncoding RNA HOTAIR is
an Essential Mediator of Acute Myocardial Infarction. Cell Physiol
Biochem. 44:1497–1508. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Dai W, Chao X, Li S, Zhou S, Zhong G and
Jiang Z: Long Noncoding RNA HOTAIR Functions as a Competitive
Endogenous RNA to Regulate Connexin43 Remodeling in Atrial
Fibrillation by Sponging MicroRNA-613. Cardiovasc Ther.
2020:59253422020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Jin Z, Shen H, Cha W, Xia H and Liu L:
Predictive value of using plasma long non-coding RNAs ANRIL and
HOXA11-AS for in-stent restenosis. Exp Ther Med. 23:1152022.
View Article : Google Scholar : PubMed/NCBI
|