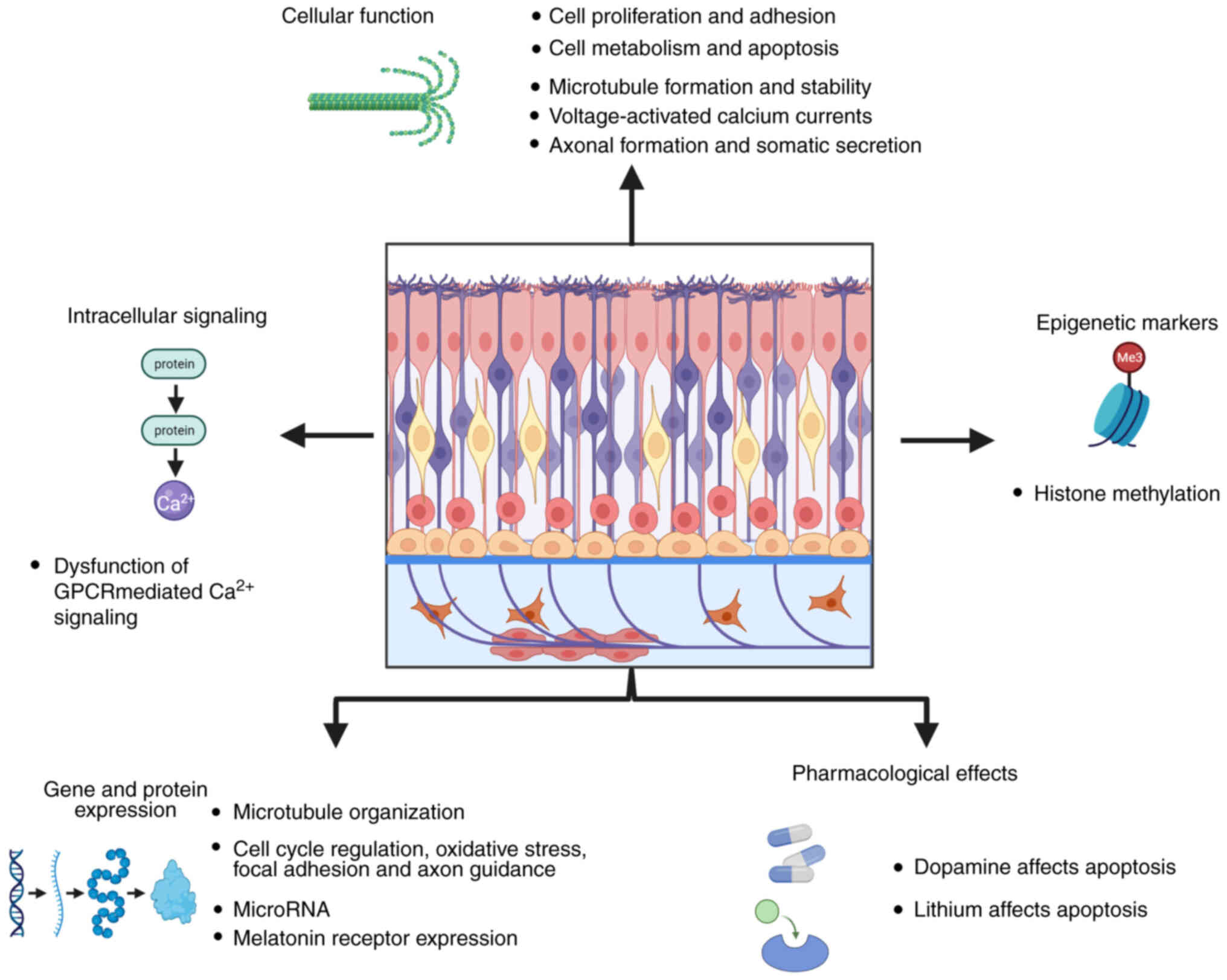
Schizophrenia (SZ) is a multifactorial disease with
an unspecified origin. Although genetic, environmental,
psychosocial and other factors might be involved in its etiology,
the precise pathophysiological mechanisms of its development remain
unclear. SZ diagnosis is primarily based on symptoms classified as
positive (hallucinations, delirium, disorganized speech,
psychomotor disturbances), negative (affective flattening, alogia,
avolition, asociality and anhedonia) or cognitive (memory and
executive function deficits) (1,2).
SZ symptoms, specifically cognitive symptoms, are
associated with the molecular structure of dopaminergic and
serotonergic topology and brain networks (3).
A key objective in the study of neuropsychiatric
disorders is to elucidate the pathophysiological processes
occurring in the brain to improve understanding of the disease and
diagnostic and therapeutic options available. The delicate nature
of the brain complicates study, and although postmortem studies
have yielded insight, there is need for suitable models to overcome
ethical and methodological limitations to obtain brain samples. In
recent years, in vitro models have emerged, such as the
culture of induced pluripotent stem cells (iPSCs) (4) and induced neuronal (iN) cells
(5), which allow reprogramming
of cells into neural and glial cell lines (6). An alternative is the use of human
olfactory neuroepithelial (hONE) cells: Primary neurons and glial
cells can be taken via epithelial cells in the nasal cavity of
living patients with a minimally invasive technique. The
heterogeneous samples include stem cells with multipotent and
regenerative capacities that can be differentiated into neuronal
and glial cells for use in vitro and ex vivo
(6,7). Neuropsychiatric disorders including
SZ (8-12), Alzheimer's disease (13,14) and other mood and anxiety
disorders (15) are associated
with anosmia, and it has been shown that the olfactory epithelial
cells of patients with these illnesses have cellular and molecular
alterations, such as amyloid-β and paired helical filament-tau
aggregates, alterations to the cell cycle and phosphatidylinositol
signaling pathways, membrane phospholipid alterations, dysregulated
neurodevelopmental pathways, dysregulated mitochondrial function,
oxidative stress (16-25). Since the hONE cells of the
olfactory bulb are connected to the olfactory cortex,
neurobiological alterations in the limbic regions may be reflected
in the hONE cells, suggesting these may serve as an appropriate
model for the study of neuropsychiatric disorders.
In patients with SZ or SZ-like animal models,
dysfunctions have been observed in intracellular mechanisms
activated by key hormones, modulators and transmitters such as
dopamine, glutamate, serotonin, acetylcholine (ACh), ATP,
melatonin, endocannabinoids and oxytocin (26-28). These modulators exert action by
binding to G protein-coupled receptors (GPCRs) and triggering
complex downstream intracellular signaling cascades. In the
physiology of the central nervous system (CNS), the GPCR family of
receptors is involved in key cellular functions such as
proliferation, differentiation, migration and neurotransmission
both in undifferentiated and mature neurodevelopmental stages
(29-31). Genomic and proteomic studies have
demonstrated the association of SZ with alterations in expression
of GPCRs and enzymes activated by them, such as phospholipase Cb
(32-34). In addition, drugs (such as
aripiprazole, azepine, chlorpromazine) used in the treatment of
this psychiatric disorder target GPCRs. To the best of our
knowledge, however, there are few studies of the functionality of
these receptors and the actions of these drugs at the cellular
level (6,35). One possibility to study these is
the use of cells cultivated from patients.
The present study conducted a literature review on
PubMed and Google Scholar, selecting articles associated with GPCRs
and their connection to SZ, as well as GPCRs in stem cells and
their relevance to SZ. The following search strategies were used:
Schizophrenia AND olfactory epithelial cells AND GPCR; GPCR AND
schizophrenia and schizophrenia AND stem cells.
To comply with the bioethical and anatomical
restrictions around directly obtaining CNS tissue from patients
with mental disorders or neurodegenerative diseases, several
experimental approaches have been developed to study human neurons
and neuroglial physiological processes at the cellular level
(36-39). Cell models have been
characterized, such as olfactory epithelial SCs, iP cells and
monocytes induced to resemble neurons (6,21). In particular, SCs of the
olfactory epithelium express different types of GPCR and may be a
suitable model to study the function of these receptors at the
cellular level and their alteration in SZ; alterations in
neurodevelopment, stress response and gene/protein expression
regulatory pathways have been found in patients with SZ through the
use of cells in culture obtained from olfactory epithelium
(40). Most of the currently
validated cellular models take advantage of the specific
characteristics of SCs, such as their self-renewal capacity and
their differentiation potency (41,42). These characteristics are also
useful to establish cryopreserved biobanks of neural SCs at
different stages of development. These cells are multipotent and
have been differentiated into neurons (43) and neuroglia (44), making the study of GPCRs at
different stages of development in different cell types
possible.
Studies have observed disease-associated
pathological traits in both neural SCs and their differentiated
progeny, such as alterations in microtubule organization (45), making these models suitable to
investigate cellular and subcellular mechanisms underlying the
pathophysiology of psychiatric disorder. Human olfactory neural
stem cells obtained by the nasal cavity exfoliation procedure
described by Benitez-King et al (37) have revealed cellular and
subcellular alterations in patients with SZ, bipolar disorder and
Alzheimer's disease (46) and in
cannabis users (Fig. 1)
(47,48). Specifically regarding GPCRs and
their signaling, one study reported abnormal 3'-5'-cyclic adenosine
monophosphate (cAMP) accumulation in patient-derived hONE cells
(49). Another study reported
melatonin MT1 and MT2 receptors and their
involvement in the modulation of axonogenesis, associated with
increased levels of phosphorylated (p)GSK3β (Fig. 1) (27); axonogenesis is impaired and
melatonin receptor and pGSK3β levels are lower in cells derived
from patients with SZ compared with those from healthy subjects
(27). In olfactory cells of
patients with SZ, trimethylation of histone H3 lysine and H3 lysine
27 alters expression of genes related to glutamate decarboxylase 1
and other pathways associated with SZ (50). Neural epithelial SCs from living
patients obtained via non-invasive exfoliation allows observation
of the pathophysiological mechanisms and structural and molecular
changes in SZ (7,51,52). Moreover, this model presents an
opportunity to obtain cells from a single patient at different
stages of disease, including naive stages and during treatment.
Numerous in vitro models for the study of SZ have been
developed and standardized using other human biospecimens such as
postmortem brains and genetically engineered cells due to their
accessibility and reliability (37,40,53,54).
The initial sample to develop iPSC and iN stem cells
can be easily collected since, usually, peripheral cells are used.
Meanwhile, the collection of hONE cells has moderate ease with a
minimally invasive technique that a qualified professional should
perform (6,55). hONE cells are ready for use ~4
weeks after collection, while iPSC require a longer waiting time
(6). Additionally, costs to
obtain hONE cells are lower than that for iPSCs and iN cells. hONE
cells are neural tissue and do not require genomic reprogramming.
Both hONE cells and iPSC have moderate or high proliferative
capacity while iN cells do not possess this capacity (5,56). As iPSC and iN cells are induced
models, it is difficult to determine the degree of phenotypical
similarity with brain cells, while in hONE cells neurobiological
properties are preserved (6).
hONE cells are cultured from living patients, which allows the
comparison of cells obtained at different stages of the illness and
treatment.
hONE cells are a relatively new model to study GPCR
expression and function. hONE cultures have multipotent SC features
and express functional purinergic P2 receptors (both ionotropic P2X
and metabotropic P2Y receptors) (57). The activation of the purinergic
pathway in these cells elicits transient increase in the
intracellular calcium (Ca2+) concentration, mainly by
the participation of the P2Y receptors; the calcium increase
induces exocytotic processes in these cells (57).
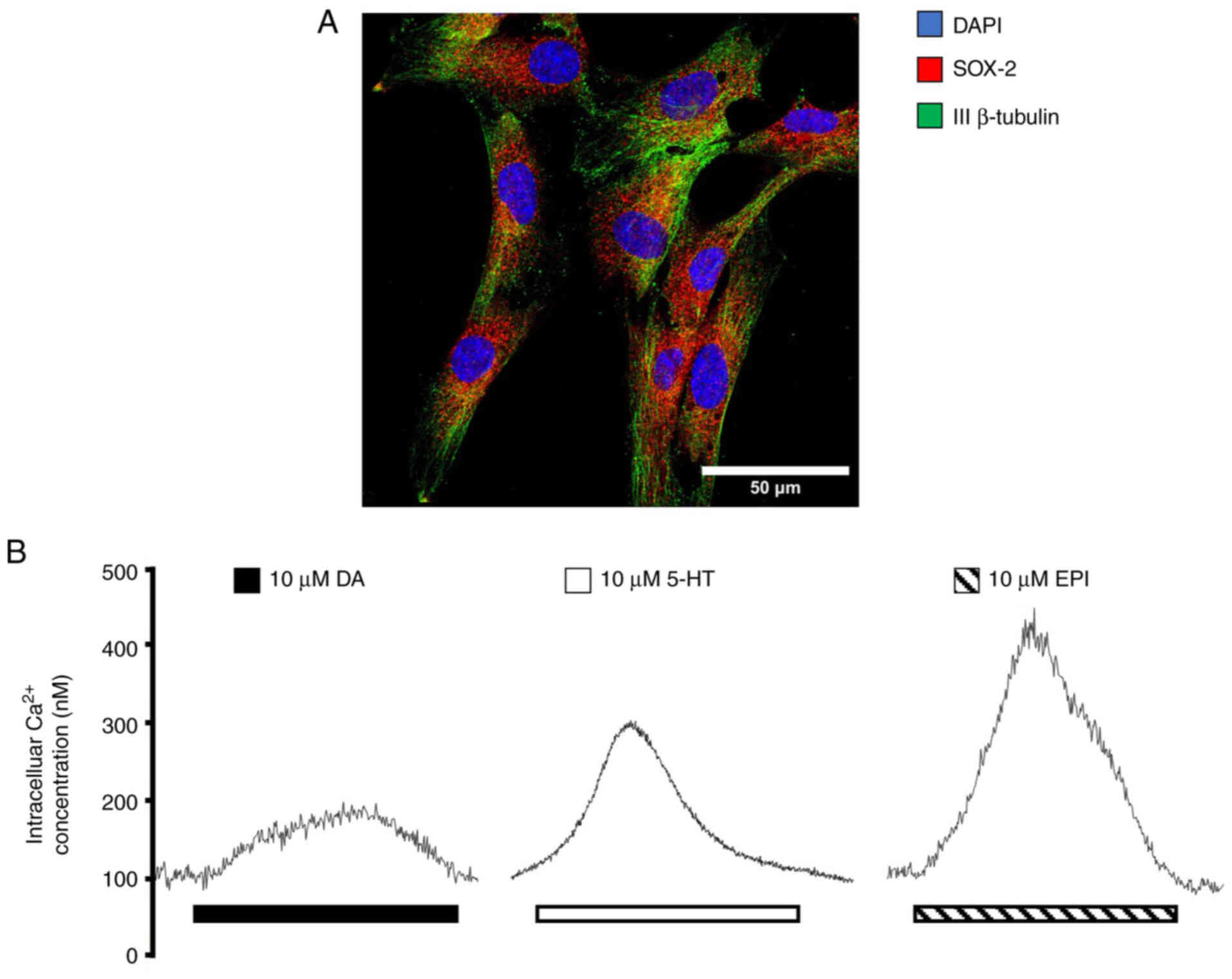
Moreover, other functional GPCRs are expressed in
human olfactory neural SCs, such as dopaminergic, serotoninergic
and adrenergic receptors (ARs). These cells express markers of
multipotency (Fig. 2A) and
elicit an increase in intracellular Ca2+ concentrations
in response to ligand binding (Fig.
2B). These characteristics contribute to a viable, minimally
invasive model for neuronal culture sample from live patients with
SZ to study the GPCR signaling pathways involved in this
pathology.
SZ clinical onset usually happens in early
adulthood. It occurs in ~1% of the human population and in the US
it is estimated to decrease lifespan by 28.5 years (84). Patients with SZ present brain
structural alterations as well as dysfunction in several
neurotransmission systems (dopaminergic, glutamatergic, GABAergic,
ACh and serotonergic signaling), in addition to inflammation and
oxidative stress. Patients also present loss of cerebral gray
matter and abnormal distribution of neurons in the prefrontal
cortex (PFC) (85-87). Patients with SZ present
structural alterations in heavily myelinated brain tracts that
comprise mostly white matter, which suggests that impaired brain
connectivity and an overall dysfunction of the axo-myelin unit is a
key mechanism underlying the pathophysiology of SZ (88). SZ has a complex genetic
background and development depends on environmental factors
(89,90). GPCRs play a key role in the
development, progression and treatment of SZ (Table I).
The biological functions of the catecholaminergic
neurotransmitter dopamine in the brain and periphery are mediated
by dopamine receptors D1-5. These functions include
regulation of sleep, feeding, synaptic regulation, attention,
cognitive function, hormonal regulation, affection, reward systems,
voluntary movement, vision and smell (91).
Variation in dopamine levels and the symptoms of SZ
are dependent on the associated brain region; increased release in
the striatum is associated with positive symptoms (hallucinations
and delusions) where the binding of the D2 receptor
predicts the response to treatment with antipsychotics. However,
the occupation of D2 receptors in the ventral region of
the striatum is associated with negative symptoms such as
passivity, apathy and social withdrawal (98). These conclusions are supported by
genetic research showing a clear association between the dopamine
receptor D2 gene and SZ (86,99). Although the majority of the
currently authorized antipsychotic drugs block D2-type
dopamine receptors, clinical symptomology is not completely treated
in most patients. However, they have effects on other receptors in
the brain, such as dopamine, serotonin, histamine, norepinephrine
and ACh receptors, resulting in other abnormality, such as the risk
of extrapyramidal side effects (100). D2 receptors are
involved in postsynaptic activation and autoreceptor-mediated
inhibition of dopamine release in the striatum and the
D1 receptor modulates actions of dopamine in the
corticostriatal circuitry; alterations in dopamine D1
receptors and key molecules in their signaling pathways have been
found in the PFC of patients with SZ (101). Other studies have visualized
expression in limbic and cortical areas of D3 and
D4 dopamine receptors (102,103). Moreover, clozapine, a second
generation antipsychotic drug, has a higher affinity for the
D4 receptor, which supports its participation in the
pathophysiology of SZ (102).
On the other hand, the distribution and low cerebral abundance of
D3 receptors, as well as their close homology with the
D2 receptor, indicate they may serve as pharmacological
targets, especially since their implementation could avoid the
adverse motor effects produced by the inhibition of the
D2 receptor (104).
The positive symptoms of SZ are exacerbated by
selective and indirect AR agonists (ephedrine, clonidine and
desipramine), while they are decreased by antagonists (yohimbine,
propranolol and oxypertine) (85,108). Additionally, α-ARs are linked
to cognitive deficit in SZ (109) and PFC impairment via PKC
activation (85,110,111). In neocortical pyramidal cells,
adrenergic arousal controls coupling between apical and somatic
integration regions by the regulation of
hyperpolarization-activated currents (Ih) and altering
apical amplification (AA) (112). Higher levels of cAMP lead to
excessive Ih, therefore increasing AA. Patients with SZ
exhibit translocation in the disrupted in schizophrenia 1 (DISC1)
gene and DISC1-regulated phosphodiesterase 4 (PDE4) activity; in
the presence of high concentration of cAMP, this increases
hydrolysis; however, but this process is altered in these patients
(113,114). This area is key for spatial
working memory (WM), in which α2A receptors serve a key
role by inhibiting the cAMP/PKA pathway, thus reducing the
persistent firing by increasing the open state of hyperpolarization
and cyclic nucleotide-gated channels (115,116). The effects of adrenergic
signaling are subtype-specific and could be influenced by
noradrenaline concentration and receptor affinity. The effect is
mediated through the persistent firing of the α2A
receptors, and the use of an exogenous general β agonist does not
alter the outcome. This phenomenon may be related to the
upregulation of cAMP (117). In
another study, the use of a β1 antagonist improved WM
and the activation of β2 enhanced this effect,
illustrating the complex modulation by adrenergic receptors
(117-119).
Certain single nucleotide polymorphisms (SNPs) have
been associated with SZ, including two SNPs in the promoter region
of the α1A receptor gene (120), as well as
methylenetetrahydrofolate reductase (MTHFR) (121,122). A detection system has been
proposed to measure levels of 5-MTHF in patients with MTHFR SNPs
(123).
ACh is a crucial neurotransmitter that participates
both in the CNS and PNS. There are two types of receptors activated
by ACh, nicotinic ionotropic and muscarinic metabotropic receptors
(mAChRs) (124). There are five
types of muscarinic receptors that can be classified as those
coupled to Gq/G11 (M1, -3 and -5)
and those coupled to Gi/o (M2 and -4)
(124-126). The M1 receptor is
the most prevalent receptor in the CNS, located in postsynaptic
neurons and some peripheral tissues (126). Meanwhile, in the presynaptic
neurons, the M2 and M4 receptors are
expressed, while in the postsynaptic neurons, the M3,
M4 and M5 receptors are expressed, with the
M3 typically being the less abundant (126). Lower levels of M1
and M4 expression have been detected in the cortex
(127,128), hippocampus (129) and striatum (130).
Genetic alterations in the muscarinic signaling
pathway have been associated with SZ, including SNPs in the gene
for the muscarinic acetylcholine receptor M1 (CHRM1) (131), as well as changes in
methylation of the promoter of this gene, caused by the increase in
microRNA (miRNA or miR) that regulates this gene (miR-107)
(132). SNPs for CHRM4
(126,133) and CHRM5 (126,134) have also been linked with an
increased risk of SZ.
The use of animal models has demonstrated the
participation of the mAChRs in the pathogenesis of SZ. In
M1 knock-out (KO) mice, impaired WM and long-term
potentiation are observed (126,135). In a double KO mice model for
M1 and M4, impaired prepulse inhibition (PPI)
is observed (126,136). M4 KO mice models
have been reported to present impaired PPI, abnormal social
behavior, locomotor activity, sensorimotor gating, abnormal
antipsychotic function, dopaminergic hyperexcitability and altered
striatal dopamine release regulation (126,137-142). It has been observed that
M5 KO mice present changes in PPI and reduced striatal
dopamine release (126,142-144).
Alterations in other participants of this signaling
pathway affect SZ. Acetylcholinesterase inhibitors, the enzyme that
hydrolyses ACh, decrease visual hallucinations (85,145,146). Additionally, choline
acetyltransferase (ChAt), the enzyme that synthesizes Ach, has
decreased activity in the nucleus accumbens and pontine tegmentum
of patients with SZ, which is associated with cognitive
performance. An SNP for ChAt is associated with SZ (147).
Glutamate is the primary excitatory
neurotransmitter in the CNS responsible for modulation of synaptic
transmission and neuronal excitability. This modulation is mediated
by the activity of ionotropic and metabotropic glutamate receptors
(mGluRs) (85,148,149). There are eight subtypes of
mGluRs encoded by the glutamate metabotropic receptor 1 (GRM1)-8
genes and these receptors are be classified into three groups:
Group I includes receptors coupled to a Gq/11 protein (mGluR1 and
mGluR5) and group II (mGluR2 and mGluR3) and III (mGluR4, -6, -7
and -8) are coupled to Gi/Go protein (85,148,149). All receptor subtypes are
expressed in neurons and glial cells, except mGluR6, which is
primarily expressed in the retina (85,150).
Alterations in mGluR1 are associated with SZ.
Patients with SZ may have deleterious GRM1 non-synonymous SNPs
(85,151); in postmortem studies, patients
with SZ have higher levels of mGluR1α in the PFC (85,152). The role of mGluR1 has been
studied through KO mice. These animals have decreased hippocampal
long-term potentiation leading to a deficit in associative learning
(148,153,154) and activity-dependent synaptic
plasticity (154). mGluR1
deficiency causes long-term depression in the cerebellum and motor
learning impairment (148,155) and a decrease in PPI (148,156). Use of mGluR1 negative
allosteric modulators is effective in the treatment of positive SZ
symptom models (85,148,157).
mGluR5 may be involved in SZ as this receptor
potentiates the NMDAR in brain regions of interest in SZ (158). In mGluR5KO mice, there is a
deficit in PPI (148,159). Furthermore, a KO model of
miR-50103p induces dendritic structural defects, glutamatergic
transmission enhancement and sociability, memory and sensorimotor
gating deficits, which are attenuated when restoring miR-50103p
expression. These effects were attributed to the upregulation of
mGluR5 since this miRNA negatively regulates the expression of the
receptor. When using a negative allosteric modulator of mGluR5,
similar effects were observed (160). In animal models of positive and
negative symptoms, a positive allosteric modulator of mGluR5
effectively improves all types of SZ symptom (85,148,157). Furthermore, mGluR5-selective
negative allosteric modulators in adult rats causes social
interaction deficits, impaired WM, reduced instrumental learning,
decreased overall response in 5-choice serial reaction time task
(5-CSRT) and increased NMDAR antagonist side effects (158,161-165). Postsynaptic mGluR2/3 activation
can augment NMDAR currents via Src kinase in pyramidal cells of the
hippocampal CA1 (166) and in
the PFC via PKC activity (167)
and soluble N-ethylmaleimide-sensitive factor attachment protein
receptor proteins (157,168).
Although the group II receptors have not been as
extensively studied, they may serve as therapeutic targets. In
animal models of SZ, the activation of mGluR2/3 decreases the
psychomotor activity and neurochemical effects produced by
psychostimulants (85,169). Agonists of mGluR2/3 decrease
extracellular dopamine efflux in the substantia nigra, nucleus
accumbens and dorsal striatum (157,170-173). The activation of mGluR2/3
functions as an autoregulator to decrease glutamate release, makes
it a target for the development of agonists for treatment of SZ
(157,174). Additionally, in preclinical
trials, mGluR2/3 agonists (LY354740 and LY379268) decrease NMDAR
antagonist-induced hyperlocomotion (175-178) and behavioral stereotypes
(175,179) and behavioral and
electrophysiological effects and head twitches induced by
(+/-)1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane (DOI) in mice
(180) and improve SZ-like
symptoms induced by prenatal stress and postnatal isolation
(157,181,182). In negative symptom models, the
agonists improve deficits in social interaction (183-185) and mobility attenuated by
dizocilpine in the swimming test (157,178). For cognitive symptoms, mGluR2/3
agonists decrease deficits in discrete-trial delayed alternation
task (175) and errors in the
5-CSRT (157,186). However, it has also been shown
that agonists can impair cognitive symptoms. Impaired cognition by
inhibiting hippocampal synaptic transmission (187) and exacerbated deficits in the
5-CSRT have been observed (157,188).
The least explored receptors are those in group
III. All receptors in this group have been studied in KO mice
models (180,184-197). The administration of a group
III agon ist (1S,3R,4S)-1-aminocyclo-pentane-1,3,4-tricarboxylic
acid (ACTP-1) decreases hyperlocomotion induced by MK-801 and
amphetamines and improves head twitches induced by DOI in mice
(157,193). The mGluR4 is expressed
throughout the brain but is most densely expressed in the
cerebellum; KO mice can present impairments in cerebellar synaptic
plasticity and motor learning of complicated tasks and altered
spatial memory performance. These receptors are key in regulation
of GABAergic absence seizures in the thalamocortical region
(148,194-196). In positive symptom animal
models, the administration of mGluR4 agonist (LSP1-2111 and
LSP4-2022) improves psychosis symptoms (hyperlocomotion and head
twitches) (157,197,198). mGluR4 agonists also improve
deficits in social interaction and novel object recognition
(157,198). mGluR6, which is primarily
expressed in the retina, presents delayed response when retinal
bipolar cells are stimulated with light in mGluR6 KO mice (148,199,200). There have been reports of
photoreceptor and bipolar and retinal ganglion cell (RGC)
dysfunction in SZ (201,202).
RGC signaling deficit is associated with SZ, particularly in
patients that experience visual hallucinations (202).
The mGluR7 receptor is widely expressed but has a
lower affinity to glutamate than other receptors and downregulates
overstimulation by glutamate (148), as indicated by the epileptic
phenotype observed in mGluR7 KO mice (148,203). mGluR7 KO mice exhibit worse
short-term neural plasticity in the hippocampus (85,204,205), memory and learning deficits
(204,206-209) and an altered fear (209) and anxiety response (20,85,148,204,210). In preclinical studies, mGluR7
negative allosteric modulators
6-(4-methoxyphenyl)-5-methyl-3-pyridin-4-ylisoxazolo[4,5-c]
pyridin-4(5H)-one and
(+)-6-(2,4-dimethylphenyl)-2-ethyl-6,7-dihydrobenzo[d]oxazol-4(5H)-one
improves symptoms caused by MK-801 and DOI-induced head twitches
(85,211-213), while an mGuR7 agonist (AMN082)
produces the opposite effects (157,197).
mGluR8 is less expressed than mGluR4 and -7; it is
primarily expressed presynaptically and widely throughout the brain
(148,157). This receptor serves as an
autoreceptor in the lateral prefrontal path of the dentate gyrus,
therefore gating glutamatergic transmission into the hippocampus
(157,214), which is why mGluR8 KO mice
exhibit deficiency in hippocampal-mediated learning (157,215). Unlike the other group III
receptors, mGluR8 agonist [(S)-3,4-DCP] does not affect NMDAR or
amphetamine hyperactivity, suggesting that it might be an
ineffective target for SZ treatment (157,216).
P2Y metabotropic purinoceptors are a family of
proteins divided into eight subtypes (P2Y1, 2, 4, 6 and 11-14) that
can be activated by several nucleotides such as ATP, ADP, UTP, UDP
and UDP-glucose (217).
Activation of these receptors induces biological effects due to the
subsequent activation of different effectors, including MAPK,
ρ-associated protein kinase, phospholipase A2, nitric oxide and the
transactivation of growth receptors (218). Several signaling pathways
activated by ATP and other nucleotides via P2Y, participate in
regulation of CNS development. Stimulation of the P2Y1
receptor promotes adult neurogenesis (219,220). The P2Y receptors have been
suggested to be involved in SZ. P2Y1 receptor agonist
(MRS2365) to the PFC in rats impairs WM and other behavioral
responses that may be involved in conditions that increase ATP
concentration, such as SZ (221). Perisomatic interneurons, which
modulate γ oscillations, express P2Y1Rs (222). These cells have been implicated
in SZ and cognitive deficit (222) and γ oscillations and PPI
alterations have been reported in SZ animal models (223,224).
Wnts transduce signaling cascades to regulate SC
differentiation in various types of tissues such as skin, muscle,
colon and bone marrow; in addition, they promote cell proliferation
and differentiation to regulate maintenance of the adult
hippocampus and neuronal progenitors of the subventricular zone
(230,231). A distinctive aspect of Wnt
signaling is its ability to favor tissue growth while inducing cell
proliferation, serving as a directional growth factor and
preventing the formation of amorphous structures, an essential
feature during tissue development and homeostasis in adults
(232,233).
Neuroinflammation and immune dysfunction could be
involved in the pathogenesis of SZ, supported by the higher
incidence of autoimmune disease in patients with SZ. The
inflammatory process is mediated by Wnt/β-catenin dysregulation,
with the primary effector being NF-κB, stimulating production of
inflammatory markers, including various cytokines, and favoring
oxidative stress. Many of these processes promote psychotic
symptoms. SZ is associated with a decrease in Wnt/β-catenin pathway
activity, leading to an upregulation of PPARγ and downregulation of
PPARα (234-236). The increase of PPARγ increases
oxidative stress and inflammation (234). The Wnt/β-catenin pathway is
involved in the pathogenesis of numerous neuropsychiatric
disorders. There have been reports of myelin and oligodendrocyte
dysfunction in SZ (88,237), indicating that the
Wnt/β-catenin pathway could be altered in this illness. The levels
of β-catenin are decreased in the hippocampal region of patients
with SZ and downstream alterations in this pathway have been also
observed (238).
Genome-wide SNP analysis has identified multiple
SNPs associated with SZ, including the FZD1 gene at chromosome
7q21.13 (239), as well as FZD3
gene on the chromosome 8p21 (240-242). FZD3 SNPS are also implicated in
methamphetamine psychosis (243). There is aberrant Wnt gene
expression at multiple levels of the signaling pathway. Microarray
analysis demonstrates that patients with SZ exhibit dysregulated
mRNA expression of genes that attenuate β-catenin signaling and
favor non-canonical signaling, while transcription factor nuclear
factor of activated T cells 3, which is activated downstream by the
non-canonical pathway, is upregulated (244).
S1P is produced in all cell types during the
catabolic degradation of membrane glycosphingolipids and
sphingomyelin, which results in sphingosine that is phosphorylated
by sphingosine kinase (SphK) to S1P, a bioactive signaling molecule
that serves as a ligand for GPCRs of the Gi/o, G12/13, and Gq types
(263). Various hormones,
cytokines and growth factors can activate the SphK/S1P signaling
pathway, modulating cell proliferation, migration and survival. The
SphK/S1P pathway has been associated with stem/progenitor cells and
tissue self-renewal in the vascular, immune, muscular and nervous
systems (264-267).
In the pathogenesis of SZ, there are alterations in
myelin, white matter integrity and metabolism of lipids. Recent
targeted mass spectrometry-based analysis found that postmortem
samples of the corpus callosum of patients with SZ have lower
levels of S1P (268).
Furthermore, one study divided patients with SZ into those that
present an upregulation of S1PR1 and those that have levels
comparable to controls (269).
This may be used as a biomarker since S1PR1 can be detected through
positron emission tomography (269).
NPY is a 36-amino acid peptide produced by
GABAergic interneurons that is widely expressed in the CNS and PNS
during development and adulthood. The Y receptors are a family of
proteins divided into five subtypes (Y1, Y2,
Y4, Y5, and Y6) that are activated
by the NPY family of hormones, which consists of three native
peptide ligands (NPY, pancreatic polypeptide and peptide YY). All
NPY receptors are involved in the Gi signaling cascade; upon
activation, the α subunit decreases cAMP production and the b/g
subunit activates various kinase cascades. This ligand-receptor
interaction can lead to decreased Ca2+ channel activity
and increased G-protein-coupled inward rectifying potassium
currents (270,271).
NPY serves an important role in the regulation of
learning, memory, feeding and endocrine secretion (272). NPY is found in the olfactory
neuroepithelium, where it stimulates proliferation of olfactory SCs
(273). Additionally, NPY
regulates the response of olfactory receptors, apoptosis and cell
regeneration (274) and
protects sensory neurons from death due to excessive GluR
activation by decreasing Ca2+ entry into the presynaptic
nerve terminal via PKA- and p38K-associated signaling (275).
NPY participates in adult neurogenesis in the
hippocampal dentate gyrus, caudal subventricular zone and
subcallosal zone (276). In
vivo, by fusing NPY vectors with a brain transport peptide
(apolipoprotein B), proliferation of neural precursor cells in the
subgranular zone of the hippocampus increases substantially without
neuronal differentiation (277). Furthermore, NPY promotes the
proliferation of olfactory and hippocampal SCs (272,273,278).
Chemokines are a family of small cytokines (CXC, CC
or β-chemokines, C, and CX3C), that regulate chemotaxis,
hematopoiesis, angiogenesis, survival, proliferation, migration and
degranulation of leukocytes by coupling with their respective GPCRs
(283). Chemokine receptors are
divided into four subtypes according to their activating chemokine
ligands (284). Chemokines are
key regulators of SCs in specific tissues (268,285) and can mediate migration of
multipotent SCs (286). CXCR4
modulates growth factor signaling and is expressed in vitro
in adult human and murine NSCs and cells from the embryonic murine
subventricular zone (287).
In addition to chemotactic functions, it has been
observed that chemokines participate in neuromodulation,
neurotransmission and neurogenesis, exert a pleiotropic effect and
exacerbate inflammation, which is why their dysregulation is
associated with neurobiological processes associated with mental
illnesses such as SZ (284,288). A systematic review demonstrated
an association between chemokines and neuroinflammation and the
pathogenesis of SZ, highlighting that there is a genetic
association of SZ with polymorphisms of chemokine receptor genes,
blood levels of CXCL8/IL-8, CCL2/(monocyte chemoattractant protein
1, chemokine (C-C motif) ligands 4 (CCL4)/macrophage inflammatory
protein 1β (MIP-1β), and CCL11/eotaxin-1 are increased and
chemokine expression and their receptors are changed in brain
regions and peripheral immune cells of patients with SZ and animal
models have revealed molecular mechanisms associated with
deregulation of the CX3CL1-CX3CR1 and CXCL12-CXCR4 axes,
demonstrating that deregulation of chemokine expression may
contribute to the neurobiological processes that cause SZ (284).
Management of patients with SZ consists of
pharmacotherapy and/or psychotherapy and its principal goal is to
improve quality of life and limiting side effects of treatment to
maintain adherence to the treatment. The primary pharmacological
therapy used in SZ is based on total or partial antagonists of the
dopamine D2 receptor, however, few patients fully
recover or exhibit reversed negative symptoms (Table II). Moreover, the cognitive
impairments of SZ are usually resistant to current antipsychotic
treatment (289).
GPCRs play an important role in the treatment of SZ
because they transmit the extracellular signal into cells by
activating the signaling cascade coupled to G proteins. Advances in
pharmacology have made it possible to identify drugs that can
modify the interaction of GPCRs related to dopaminergic and
serotonergic activity in the treatment and management of SZ
(290,291). Understanding the role of GPCRs
in the signal transduction of SZ is fundamental for the discovery
of pharmacological targets. The basis of pharmacological treatment
for SZ requires a complete understanding of GPCR-mediated
signaling, transducers and associated second messengers. Structural
plasticity of GPCR proteins underlying physiological regulation
with pharmacological implications in clinical use has been
summarized previously (292).
Considering SZ pathophysiology and ineffective
antipsychotic therapy with severe side effects and poor adherence
to the therapeutic regimen that diminishes quality of life and
undermines the beneficial effects of the drugs, novel treatments
directed at the whole symptomatology as well as specific symptoms
are needed. There are numerous clinical studies of GPCR targets,
including those directed at general, positive, negative and
cognitive symptoms (30,293,294).
The present review demonstrated that GPCR
alterations can be associated with the pathophysiology of
psychiatric disorders and neurodegenerative diseases, such as SZ.
GPCRs are a therapeutic target of antipsychotics used in the
treatment of SZ. To the best of our knowledge, however,
experimental evidence regarding the functionality of these
receptors in patients is scarce. Knowledge of GPCR signaling in
human multipotent SCs and their progeny differentiated in neurons
or neuroglia could widen the study of the pathophysiology of SZ and
other diseases such as diabetes, myocardial infarction, stroke,
Parkinson's disease, Alzheimer's disease and multiple
sclerosis.
Some of the limitations of hONE as a model of study
in SZ include lack of information about GPCRs functionality in hONE
cells; also, since these cells are undifferentiated, they may have
a distinct expression of channels and receptors than their
differentiated progeny, and the results obtained in the
undifferentiated cells should be corroborated in conventional SZ
models based on differentiated dopaminergic and serotoninergic
neurons.
Models such as patient-derived iPSCs,
transdifferentiated neurons, olfactory sensory neurons and cerebral
organoids can provide understanding of SZ and facilitate the
development of treatment. Particularly, the culture and
cryopreservation of olfactory SCs have been characterized and used
to identify several dysfunctional processes at a cellular level;
this has been proposed as a model to understand the pathophysiology
of neuropsychiatric disorders and detect biomarkers for diagnosis.
This model could be useful to study the functionality of GPCR in
SZ. GPCRs and their associated signaling pathways are possible
therapeutic targets for SZ, although further research using
experimental and bioinformatic tools is needed.
Not applicable.
ZASF, BSRM, EFS, BS and HSC conceived the study.
HS, LMM, MVT, EC, AAG, GOLR, RA and JA edited the manuscript. All
authors wrote the manuscript. All authors have read and approved
the final manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The authors would also like to thank Ms Maria del
Pilar Romo and Ms Rosalba Linares for administrative and technical
support (National Autonomous University of Mexico, Mexico City
04510).
The present study was supported by Consejo Nacional de
Humanidades Ciencia y Tecnología (grant no. CF/2019/137725) and
Programa Presupuestario F003 (grant no. 287115).
|
1
|
Tandon R, Gaebel W, Barch DM, Bustillo J,
Gur RE, Heckers S, Malaspina D, Owen MJ, Schultz S, Tsuang M, et
al: Definition and description of schizophrenia in the DSM-5.
Schizophr Res. 150:3–10. 2013.
|
|
2
|
Gaebel W and Zielasek J: Schizophrenia in
2020: Trends in diagnosis and therapy. Psychiatry Clin Neurosci.
69:661–673. 2015.
|
|
3
|
Chen J, Müller VI, Dukart J, Hoffstaedter
F, Baker JT, Holmes AJ, Vatansever D, Nickl-Jockschat T, Liu X,
Derntl B, et al: Intrinsic Connectivity Patterns of Task-defined
brain networks allow individual prediction of cognitive symptom
dimension of schizophrenia and are linked to molecular
architecture. Biol Psychiatry. 89:308–319. 2021.
|
|
4
|
Marchetto MC, Brennand KJ, Boyer LF and
Gage FH: Induced pluripotent stem cells (iPSCs) and neurological
disease modeling: Progress and promises. Hum Mol Genet.
20:R109–R115. 2011.
|
|
5
|
Yang N, Ng YH, Pang ZP, Südhof TC and
Wernig M: Induced neuronal cells: How to make and define a neuron.
Cell Stem Cell. 9:517–525. 2011.
|
|
6
|
Borgmann-Winter K, Willard SL, Sinclair D,
Mirza N, Turetsky B, Berretta S and Hahn CG: Translational
potential of olfactory mucosa for the study of neuropsychiatric
illness. Transl Psychiatry. 5:e5272015.
|
|
7
|
Benítez-King G, Valdés-Tovar M, Trueta C,
Galván-Arrieta T, Argueta J, Alarcón S, Lora-Castellanos A and
Solís-Chagoyán H: The microtubular cytoskeleton of olfactory
neurons derived from patients with schizophrenia or with bipolar
disorder: Implications for biomarker characterization, neuronal
physiology and pharmacological screening. Mol Cell Neurosci.
73:84–95. 2016.
|
|
8
|
Moberg PJ, Kamath V, Marchetto DM, Calkins
ME, Doty RL, Hahn CG, Borgmann-Winter KE, Kohler CG, Gur RE and
Turetsky BI: Meta-analysis of olfactory function in schizophrenia,
first-degree family members, and youths at-risk for psychosis.
Schizophr Bull. 40:50–59. 2014.
|
|
9
|
Kamath V, Turetsky BI, Calkins ME, Bilker
WB, Frishberg N, Borgmann-Winter K, Kohler CG, Conroy CG, Gur RE
and Moberg PJ: The effect of odor valence on olfactory performance
in schizophrenia patients, unaffected relatives and at-risk youth.
J Psychiatr Res. 47:1636–1641. 2013.
|
|
10
|
Malaspina D, Goetz R, Keller A, Messinger
JW, Bruder G, Goetz D, Opler M, Harlap S, Harkavy-Friedman J and
Antonius D: Olfactory processing, sex effects and heterogeneity in
schizophrenia. Schizophr Res. 135:144–151. 2012.
|
|
11
|
Turetsky BI, Hahn C-G, Arnold SE and
Moberg PJ: Olfactory receptor neuron dysfunction in schizophrenia.
Neuropsychopharmacology. 34:767–774. 2009.
|
|
12
|
Rupp CI, Fleischhacker WW, Kemmler G,
Oberbauer H, Scholtz AW, Wanko C and Hinterhuber H: Various
bilateral olfactory deficits in male patients with schizophrenia.
Schizophr Bull. 31:155–165. 2005.
|
|
13
|
Alvarado-Martínez R, Salgado-Puga K and
Peña-Ortega F: Amyloid beta inhibits olfactory bulb activity and
the ability to smell. PLoS One. 8:e757452013.
|
|
14
|
Conti MZ, Vicini-Chilovi B, Riva M,
Zanetti M, Liberini P, Padovani A and Rozzini L: Odor
identification deficit predicts clinical conversion from mild
cognitive impairment to dementia due to Alzheimer's disease. Arch
Clin Neuropsychol. 28:391–399. 2013.
|
|
15
|
Burón E and Bulbena A: Olfaction in
affective and anxiety disorders: A review of the literature.
Psychopathology. 46:63–74. 2013.
|
|
16
|
Arnold SE, Lee EB, Moberg PJ, Stutzbach L,
Kazi H, Han LY and Trojanowski JQ: Olfactory epithelium amyloid-β
and paired helical filament-tau pathology in Alzheimer disease. Ann
Neurol. 67:462–469. 2010.
|
|
17
|
Arnold SE, Smutzer GS, Trojanowski JQ and
Moberg PJ: Cellular and molecular neuropathology of the olfactory
epithelium and central olfactory pathways in Alzheimer's disease
and schizophrenia. Ann N Y Acad Sci. 855:762–775. 1998.
|
|
18
|
Arnold SE, Han L-Y, Moberg PJ, Turetsky
BI, Gur RE, Trojanowski JQ and Hahn CG: Dysregulation of olfactory
receptor neuron lineage in schizophrenia. Arch Gen Psychiatry.
58:829–835. 2001.
|
|
19
|
McCurdy RD, Féron F, Perry C, Chant DC,
McLean D, Matigian N, Hayward NK, McGrath JJ and Mackay-Sim A: Cell
cycle alterations in biopsied olfactory neuroepithelium in
schizophrenia and bipolar I disorder using cell culture and gene
expression analyses. Schizophr Res. 82:163–173. 2006.
|
|
20
|
Féron F, Perry C, Hirning MH, McGrath J
and Mackay-Sim A: Altered adhesion, proliferation and death in
neural cultures from adults with schizophrenia. Schizophr Res.
40:211–218. 2006.
|
|
21
|
Matigian N, Abrahamsen G, Sutharsan R,
Cook AL, Vitale AM, Nouwens A, Bellette B, An J, Anderson M,
Beckhouse AG, et al: Disease-specific, neurosphere-derived cells as
models for brain disorders. Dis Models Mech. 3:785–798. 2010.
|
|
22
|
Fan Y, Abrahamsen G, McGrath JJ and
Mackay-Sim A: Altered cell cycle dynamics in schizophrenia. Biol
Psychiatry. 71:129–135. 2012.
|
|
23
|
Fan Y, Abrahamsen G, Mills R, Calderón CC,
Tee JY, Leyton L, Murrell W, Cooper-White J, McGrath JJ and
Mackay-Sim A: Focal adhesion dynamics are altered in schizophrenia.
Biol Psychiatry. 74:418–426. 2013.
|
|
24
|
Hahn CG, Gomez G, Restrepo D, Friedman E,
Josiassen R, Pribitkin EA, Lowry LD, Gallop RJ and Rawson NE:
Aberrant intracellular calcium signaling in olfactory neurons from
patients with bipolar disorder. Am J Psychiatry. 162:616–618.
2005.
|
|
25
|
Pantazopoulos H, Boyer-Boiteau A, Holbrook
EH, Jang W, Hahn CG, Arnold SE and Berretta S: Proteoglycan
abnormalities in olfactory epithelium tissue from subjects
diagnosed with schizophrenia. Schizophr Res. 150:366–372. 2013.
|
|
26
|
Yang A and Tsai SJ: New targets for
schizophrenia treatment beyond the dopamine hypothesis. Int J Mol
Sci. 18:16892017.
|
|
27
|
Galván-Arrieta T, Trueta C, Cercós MG,
Valdés-Tovar M, Alarcón S, Oikawa J, Zamudio-Meza H and
Benítez-King G: The role of melatonin in the neurodevelopmental
etiology of schizophrenia: A study in human olfactory neuronal
precursors. J Pineal Res. 63:2017. View Article : Google Scholar
|
|
28
|
Ferretjans R, de Souza RP, Panizzutti B,
Ferrari P, Mantovani L, de Campos-Carli SM, Santos RR, Guimarães
FC, Teixeira AL, Gama CS and Salgado JV: Cannabinoid receptor gene
polymorphisms and cognitive performance in patients with
schizophrenia and controls. Braz J Psychiatry. 44:26–34. 2022.
|
|
29
|
Borroto-Escuela DO, Cuesta-Marti C,
Lopez-Salas A, Chruścicka-Smaga B, Crespo-Ramírez M, Tesoro-Cruz E,
Palacios-Lagunas DA, Perez de la Mora M, Schellekens H and Fuxe K:
The oxytocin receptor represents a key hub in the GPCR
heteroreceptor network: Potential relevance for brain and behavior.
Front Mol Neurosci. 15:10553442022.
|
|
30
|
Rahman MM, Islam MR, Mim SA, Sultana N,
Chellappan DK, Dua K, Kamal MA, Sharma R and Emran TB: Insights
into the promising prospect of G protein and GPCR-mediated
signaling in neuropathophysiology and its therapeutic regulation.
Oxid Med Cell Longev. 2022:84256402022.
|
|
31
|
Komatsu H, Fukuchi M and Habata Y:
Potential utility of biased GPCR signaling for treatment of
psychiatric disorders. Int J Mol Sci. 20:201906292019.
|
|
32
|
Udawela M, Scarr E, Hannan AJ, Thomas EA
and Dean B: Phospholipase C beta 1 expression in the dorsolateral
prefrontal cortex from patients with schizophrenia at different
stages of illness. Aust N Z J Psychiatry. 45:140–147. 2011.
|
|
33
|
Udawela M, Scarr E, Boer S, Um JY, Hannan
AJ, McOmish C, Felder CC, Thomas EA and Dean B: Isoform specific
differences in phospholipase C beta 1 expression in the prefrontal
cortex in schizophrenia and suicide. NPJ Schizophr. 3:192017.
|
|
34
|
Vasco VRL, Cardinale G and Polonia P:
Deletion of PLCB1 gene in schizophrenia-affected patients. J Cell
Mol Med. 16:844–851. 2012.
|
|
35
|
Deng C, Pan B, Engel M and Huang XF:
Neuregulin-1 signalling and antipsychotic treatment: Potential
therapeutic targets in a schizophrenia candidate signalling
pathway. Psychopharmacology (Berl). 226:201–215. 2013.
|
|
36
|
Féron F, Perry C, Girard SD and Mackay-Sim
A: Isolation of adult stem cells from the human olfactory mucosa.
Methods Mol Biol. 1059:107–114. 2013.
|
|
37
|
Benitez-King G, Riquelme A, Ortiz-Lopez L,
Berlanga C, Rodriguez-Verdugo MS, Romo F, Calixto E, Solís-Chagoyán
H, Jímenez M, Montaño LM, et al: A non-invasive method to isolate
the neuronal linage from the nasal epithelium from schizophrenic
and bipolar diseases. J Neurosci Methods. 201:35–45. 2011.
|
|
38
|
Bellon A, Wegener A, Lescallette AR,
Valente M, Yang SK, Gardette R, Matricon J, Mouaffak F, Watts P,
Vimeux L, et al: Transdifferentiation of human circulating
monocytes into neuronal-like cells in 20 days and without
reprograming. Front Mol Neurosci. 11:3232018.
|
|
39
|
Stoddard-Bennett T and Reijo Pera R:
Treatment of Parkinson's disease through personalized medicine and
induced pluripotent stem cells. Cells. 8:262019.
|
|
40
|
Lavoie J, Sawa A and Ishizuka K:
Application of olfactory tissue and its neural progenitors to
schizophrenia and psychiatric research. Curr Opin Psychiatry.
30:176–183. 2017.
|
|
41
|
Ellis P, Fagan BM, Magness ST, Hutton S,
Taranova O, Hayashi S, McMahon A, Rao M and Pevny L: SOX2, a
persistent marker for multipotential neural stem cells derived from
embryonic stem cells, the embryo or the adult. Dev Neurosci.
26:148–165. 2004.
|
|
42
|
Caprnda M, Kubatka P, Gazdikova K,
Gasparova I, Valentova V, Stollarova N, La Rocca G, Kobyliak N,
Dragasek J, Mozos I, et al: Immunomodulatory effects of stem cells:
Therapeutic option for neurodegenerative disorders. Biomed
Pharmacother. 91:60–69. 2017.
|
|
43
|
Zhang X, Klueber KM, Guo Z, Lu C and
Roisen FJ: Adult human olfactory neural progenitors cultured in
defined medium. Exp Neurol. 186:112–123. 2004.
|
|
44
|
Zhang X, Cai J, Klueber KM, Guo Z, Lu C,
Qiu M and Roisen FJ: Induction of oligodendrocytes from adult human
olfactory epithelial-derived progenitors by transcription factors.
Stem Cells. 23:442–453. 2005.
|
|
45
|
Solís-Chagoyán H, Calixto E, Figueroa A,
Montaño LM, Berlanga C, Rodríguez-Verdugo MS, Romo F, Jiménez M,
Gurrola CZ, Riquelme A and Benítez-King G: Microtubule organization
and L-type voltage-activated calcium current in olfactory neuronal
cells obtained from patients with schizophrenia and bipolar
disorder. Schizophr Res. 143:384–389. 2013.
|
|
46
|
Riquelme A, Valdés-Tovar M, Ugalde O,
Maya-Ampudia V, Fernández M, Mendoza-Durán L, Rodríguez-Cárdenas L
and Benítez-King G: Potential use of exfoliated and cultured
olfactory neuronal precursors for in vivo Alzheimer's disease
diagnosis: A pilot study. Cell Mol Neurobiol. 40:87–98. 2020.
|
|
47
|
Barrera-Conde M, Ausin K, Lachén-Montes M,
Fernández-Irigoyen J, Galindo L, Cuenca-Royo A, Fernández-Avilés C,
Pérez V, de la Torre R, Santamaría E and Robledo P: Cannabis use
induces distinctive proteomic alterations in olfactory
neuroepithelial cells of schizophrenia patients. J Pers Med.
11:1602021.
|
|
48
|
Delgado-Sequera A, Hidalgo-Figueroa M,
Barrera-Conde M, Duran-Ruiz MC, Castro C, Fernández-Avilés C, de la
Torre R, Sánchez-Gomar I, Pérez V, Geribaldi-Doldán N, et al:
Olfactory neuroepithelium cells from cannabis users display
alterations to the cytoskeleton and to markers of adhesion,
proliferation and apoptosis. Mol Neurobiol. 58:1695–1710. 2021.
|
|
49
|
Muñoz-Estrada J, Benítez-King G, Berlanga
C and Meza I: Altered subcellular distribution of the 75-kDa DISC1
isoform, cAMP accumulation, and decreased neuronal migration in
schizophrenia and bipolar disorder: Implications for
neurodevelopment. CNS Neurosci Ther. 21:446–453. 2015.
|
|
50
|
Kano S, Colantuoni C, Han F, Zhou Z, Yuan
Q, Wilson A, Takayanagi Y, Lee Y, Rapoport J, Eaton W, et al:
Genome-wide profiling of multiple histone methylations in olfactory
cells: Further implications for cellular susceptibility to
oxidative stress in schizophrenia. Mol Psychiatry. 18:740–742.
2013.
|
|
51
|
Borgmann-Winter KE, Rawson NE, Wang HY,
Wang H, Macdonald ML, Ozdener MH, Yee KK, Gomez G, Xu J, Bryant B,
et al: Human olfactory epithelial cells generated in vitro express
diverse neuronal characteristics. Neuroscience. 158:642–653.
2009.
|
|
52
|
Mackay-Sim A: Concise review:
Patient-derived olfactory stem cells: New models for brain
diseases. Stem Cells. 30:2361–2365. 2012.
|
|
53
|
Rabadan MA, De La Cruz ED, Rao SB, Chen Y,
Gong C, Crabtree G, Xu B, Markx S, Gogos JA, Yuste R and Tomer R:
An in vitro model of neuronal ensembles. Nat Commun.
13:33402022.
|
|
54
|
Hoffmann A, Ziller M and Spengler D:
Progress in iPSC-based modeling of psychiatric disorders. Int J Mol
Sci. 20:48962019.
|
|
55
|
Kolagar TA, Farzaneh M, Nikkar N and
Khoshnam SE: Human pluripotent stem cells in neurodegenerative
diseases: Potentials, advances and limitations. Curr Stem Cell Res
Ther. 15:102–110. 2020.
|
|
56
|
Nicholson MW, Ting CY, Chan DZH, Cheng YC,
Lee YC, Hsu CC, Huang CY and Hsieh PCH: Utility of iPSC-derived
cells for disease modeling, drug development, and cell therapy.
Cells. 11:18532022.
|
|
57
|
Solis-Chagoyan H, Flores-Soto E,
Valdes-Tovar M, Cercos MG, Calixto E, Montano LM, Barajas-López C,
Sommer B, Aquino-Gálvez A, Trueta C and Benítez-King GA: Purinergic
signaling pathway in human olfactory neuronal precursor cells. Stem
Cells Int. 2019:27287862019.
|
|
58
|
Berridge MJ, Bootman MD and Roderick HL:
Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev
Mol Cell Biol. 4:517–529. 2003.
|
|
59
|
Berridge MJ: Calcium signalling and
psychiatric disease: Bipolar disorder and schizophrenia. Cell and
Tissue Res. 357:477–492. 2014.
|
|
60
|
Berridge MJ: Dysregulation of neural
calcium signaling in Alzheimer disease, bipolar disorder and
schizophrenia. Prion. 7:2–13. 2013.
|
|
61
|
Schwartz RD, Wagner JP, Yu X and Martin D:
Bidirectional modulation of GABA-gated chloride channels by
divalent cations: Inhibition by Ca2+ and enhancement by Mg2+. J
Neurochemistry. 62:916–922. 1994.
|
|
62
|
Olney JW, Newcomer JW and Farber NB: NMDA
receptor hypofunction model of schizophrenia. J Psychiatric Res.
33:523–533. 1999.
|
|
63
|
Olney JW and Farber NB: Glutamate receptor
dysfunction and schizophrenia. Arch Gen Psychiatry. 52:998–1007.
1995.
|
|
64
|
Sharp FR, Butman M, Koistinaho J, Aardalen
K, Nakki R, Massa SM, Swanson RA and Sagar SM: Phencyclidine
induction of the hsp 70 stress gene in injured pyramidal neurons is
mediated via multiple receptors and voltage gated calcium channels.
Neuroscience. 62:1079–1092. 1994.
|
|
65
|
Novak G, Seeman P and Tallerico T:
Schizophrenia: Elevated mRNA for calcium-calmodulin-dependent
protein kinase IIbeta in frontal cortex. Brain Res Mol Brain Res.
82:95–100. 2000.
|
|
66
|
Benfenati F, Valtorta F, Rubenstein JL,
Gorelick FS, Greengard P and Czernik AJ: Synaptic
vesicle-associated Ca2+/calmodulindependent protein kinase II is a
binding protein for synapsin I. Nature. 359:417–420. 1992.
|
|
67
|
Greengard P, Benfenati F and Valtorta F:
Synapsin I, an actin-binding protein regulating synaptic vesicle
traffic in the nerve terminal. Adv Second Messenger Phosphoprotein
Res. 29:31–45. 1994.
|
|
68
|
Kantor L, Hewlett GHK and Gnegy ME:
Enhanced amphetamine- and K+-mediated dopamine release in rat
striatum after repeated amphetamine: Differential requirements for
Ca2+- and Calmodulin-dependent phosphorylation and synaptic
vesicles. J Neurosci. 19:3801–3808. 1999.
|
|
69
|
Popov N and Matthies H: Influence of
dopamine receptor agonists and antagonists on calmodulin
translocation in different brain regions. Eur J Pharmacol.
172:205–210. 1989.
|
|
70
|
Selemon LD and Goldman-Rakic PS: The
reduced neuropil hypothesis: A circuit based model of
schizophrenia. Biol Psychiatry. 45:17–25. 1999.
|
|
71
|
Broadbelt K, Byne W and Jones LB: Evidence
for a decrease in basilar dendrites of pyramidal cells in
schizophrenic medial prefrontal cortex. Schizophr Res. 58:75–81.
2002.
|
|
72
|
Mattson MP: Calcium as sculptor and
destroyer of neural circuitry. Exp Gerontol. 27:29–49. 1992.
|
|
73
|
Bird MM and Owen A: The effect of calcium
ionophore A23187 on neurites from embryonic mouse spinal cord
explants in culture. J Eectron Microscopy. 49:379–386. 2000.
|
|
74
|
Lidow MS: Calcium signaling dysfunction in
schizophrenia: A unifying approach. Brain Res Brain Res Rev.
43:70–84. 2003.
|
|
75
|
Benes FM, McSparren J, Bird ED,
SanGiovanni JP and Vincent SL: Deficits in small interneurons in
prefrontal and cingulate cortices of schizophrenic and
schizoaffective patients. Arch Gen Psychiatry. 48:996–1001.
1991.
|
|
76
|
Benes FM, Davidson J and Bird ED:
Quantitative cytoarchitectural studies of the cerebral cortex of
schizophrenics. Arch Gen Psychiatry. 43:31–35. 1986.
|
|
77
|
Benes FM, Kwok EW, Vincent SL and
Todtenkopf MS: A reduction of nonpyramidal cells in sector CA2 of
schizophrenics and manic depressives. Biol Psychiatry. 44:88–97.
1998.
|
|
78
|
Falkai P and Bogerts B: Cell loss in the
hippocampus of schizophrenics. Eur Arch Psychiatry Neurol Sci.
236:154–161. 1986.
|
|
79
|
Jeste DV and Lohr JB: Hippocampal
pathologic findings in schizophrenia. A morphometric study. Arch
Gen Psychiatry. 46:1019–1024. 1989.
|
|
80
|
Popken GJ, Bunney WE, Potkin SG and Jones
EG: Subnucleus-specific loss of neurons in medial thalamus of
schizophrenics. Proc Natl Acad Sci USA. 97:9276–9280. 2000.
|
|
81
|
Akbarian S, Kim JJ, Potkin SG, Hetrick WP,
Bunney WE Jr and Jones EG: Maldistribution of interstitial neurons
in prefrontal white matter of the brains of schizophrenic patients.
Arch Gen Psychiatry. 53:425–436. 1996.
|
|
82
|
Hirai K, Yoshioka H, Kihara M, Hasegawa K,
Sakamoto T, Sawada T and Fushiki S: Inhibiting neuronal migration
by blocking NMDA receptors in the embryonic rat cerebral cortex: A
tissue culture study. Brain Res Dev Brain Res. 114:63–67. 1999.
|
|
83
|
Soria JM and Valdeolmillos M:
Receptor-activated calcium signals in tangentially migrating
cortical cells. Cerebral Cortex. 12:831–839. 2002.
|
|
84
|
Velligan DI and Rao S: The epidemiology
and global burden of schizophrenia. J Clin Psychiatry.
84:MS21078COM52023.
|
|
85
|
Boczek T, Mackiewicz J, Sobolczyk M,
Wawrzyniak J, Lisek M, Ferenc B, Guo F and Zylinska L: The role of
G Protein-coupled receptors (GPCRs) and calcium signaling in
schizophrenia. Focus on GPCRs activated by neurotransmitters and
chemokines. Cells. 10:12282021.
|
|
86
|
Ermakov EA, Dmitrieva EM, Parshukova DA,
Kazantseva DV, Vasilieva AR and Smirnova LP: Oxidative
Stress-related mechanisms in schizophrenia pathogenesis and new
treatment perspectives. Oxid Med Cell Longev. 2021:88817702021.
|
|
87
|
Özdemir H, Eker M, Zengin B, Yılmaz DA,
İşman Haznedaroğlu D, Çınar C, Kitiş Ö, Akay A and Gönül AS: Gray
matter changes in patients with deficit schizophrenia and
non-deficit schizophrenia. Turk Psikiyatri Derg. 23:237–246.
2012.
|
|
88
|
Valdés-Tovar M, Rodríguez-Ramírez AM,
Rodríguez-Cárdenas L, Sotelo-Ramírez CE, Camarena B,
Sanabrais-Jiménez MA, Solís-Chagoyán H, Argueta J and
López-Riquelme GO: Insights into myelin dysfunction in
schizophrenia and bipolar disorder. World J Psychiatry. 12:264–285.
2022.
|
|
89
|
Jimerson DC, Post RM, Carman JS, van
Kammen DP, Wood JH, Goodwin FK and Bunney WE Jr: CSF calcium:
Clinical correlates in affective illness and schizophrenia. Biol
Psychiatry. 14:37–51. 1979.
|
|
90
|
Lewis DA and Moghaddam B: Cognitive
dysfunction in schizophrenia: Convergence of gamma-aminobutyric
acid and glutamate alterations. Arch Neurol. 63:1372–1376.
2006.
|
|
91
|
Beaulieu JM, Espinoza S and Gainetdinov
RR: Dopamine receptors-IUPHAR Review 13. Br J Pharmacol. 172:1–23.
2015.
|
|
92
|
Li YC, Kellendonk C, Simpson EH, Kandel ER
and Gao WJ: D2 receptor overexpression in the striatum leads to a
deficit in inhibitory transmission and dopamine sensitivity in
mouse prefrontal cortex. Proc Natl Acad Sci USA. 108:12107–12112.
2011.
|
|
93
|
Takahashi H, Kato M, Takano H, Arakawa R,
Okumura M, Otsuka T, Kodaka F, Hayashi M, Okubo Y, Ito H and Suhara
T: Differential contributions of prefrontal and hippocampal
dopamine D(1) and D(2) receptors in human cognitive functions. J
Neurosci. 28:12032–12038. 2008.
|
|
94
|
Beaulieu JM and Gainetdinov RR: The
physiology, signaling, and pharmacology of dopamine receptors.
Pharmacol Rev. 63:182–217. 2011.
|
|
95
|
Speranza L, di Porzio U, Viggiano D, de
Donato A and Volpicelli F: Dopamine: The Neuromodulator of
Long-term synaptic plasticity, reward and movement control. Cells.
10:7352021.
|
|
96
|
Conio B, Martino M, Magioncalda P,
Escelsior A, Inglese M, Amore M and Northoff G: Opposite effects of
dopamine and serotonin on resting-state networks: Review and
implications for psychiatric disorders. Mol Psychiatry. 25:82–93.
2020.
|
|
97
|
Seeman P: Targeting the dopamine D2
receptor in schizophrenia. Expert Opin Ther Targets. 10:515–531.
2006.
|
|
98
|
Simpson EH, Gallo EF, Balsam PD, Javitch
JA and Kellendonk C: How changes in dopamine D2 receptor levels
alter striatal circuit function and motivation. Mol Psychiatry.
27:436–444. 2022.
|
|
99
|
Schizophrenia Working Group of the
Psychiatric Genomics Consortium: Biological insights from 108
schizophrenia-associated genetic loci. Nature. 511:421–427.
2014.
|
|
100
|
Howes O, Mccutcheon R and Stone J:
Glutamate and dopamine in schizophrenia: An update for the 21st
century. J Psychopharmacol. 29:97–115. 2015.
|
|
101
|
Goldman-Rakic P, Castner S, Svensson T,
Siever L and Williams G: Targeting the dopamine D1 receptor in
schizophrenia: Insights for cognitive dysfunction.
Psychopharmacology (Berl). 174:3–16. 2004.
|
|
102
|
Jardemark K, Wadenberg ML, Grillner P and
Svensson TH: Dopamine D3 and D4 receptor antagonists in the
treatment of schizophrenia. Curr Opin Investig Drugs. 3:101–105.
2002.
|
|
103
|
Gross G, Wicke K and Drescher KU: Dopamine
D3 receptor antagonism-still a therapeutic option for
the treatment of schizophrenia. Naunyn Schmiedebergs Arch
Pharmacol. 386:155–166. 2013.
|
|
104
|
Maramai S, Gemma S, Brogi S, Campiani G,
Butini S, Stark H and Brindisi M: Dopamine D3 receptor antagonists
as potential therapeutics for the treatment of neurological
diseases. Front Neurosci. 10:4512016.
|
|
105
|
Motiejunaite J, Amar L and Vidal-Petiot E:
Adrenergic receptors and cardiovascular effects of catecholamines.
Ann Endocrinol (Paris). 82:193–197. 2021.
|
|
106
|
Perez DM and Doze VA: Cardiac and
neuroprotection regulated by α(1)-adrenergic receptor subtypes. J
Recept Signal Transduct Res. 31:98–110. 2011.
|
|
107
|
Jensen BC, Swigart PM, De Marco T, Hoopes
C and Simpson PC: {alpha}1-Adrenergic receptor subtypes in
nonfailing and failing human myocardium. Circ Heart Fail.
2:654–663. 2009.
|
|
108
|
Yamamoto K and Hornykiewicz O: Proposal
for a noradrenaline hypothesis of schizophrenia. Prog
Neuropsychopharmacol Biol Psychiatry. 28:913–922. 2004.
|
|
109
|
Arnsten AT: Adrenergic targets for the
treatment of cognitive deficits in schizophrenia.
Psychopharmacology (Berl). 174:25–31. 2004.
|
|
110
|
Atzori M, Cuevas-Olguin R, Esquivel-Rendon
E, Garcia-Oscos F, Salgado-Delgado RC, Saderi N, Miranda-Morales M,
Treviño M, Pineda JC and Salgado H: Locus ceruleus norepinephrine
release: A central regulator of CNS Spatio-temporal activation?
Front Synaptic Neurosci. 8:252016.
|
|
111
|
Birnbaum SG, Yuan PX, Wang M,
Vijayraghavan S, Bloom AK, Davis DJ, Gobeske KT, Sweatt JD, Manji
HK and Arnsten AF: Protein kinase C overactivity impairs prefrontal
cortical regulation of working memory. Science. 306:882–884.
2004.
|
|
112
|
Phillips WA, Larkum ME, Harley CW and
Silverstein SM: The effects of arousal on apical amplification and
conscious state. Neurosci Conscious. 2016:niw0152016.
|
|
113
|
Millar JK, Pickard BS, Mackie S, James R,
Christie S, Buchanan SR, Malloy MP, Chubb JE, Huston E, Baillie GS,
et al: DISC1 and PDE4B are interacting genetic factors in
schizophrenia that regulate cAMP signaling. Science. 310:1187–1191.
2005.
|
|
114
|
Millar JK, Mackie S, Clapcote SJ, Murdoch
H, Pickard BS, Christie S, Muir WJ, Blackwood DH, Roder JC, Houslay
MD and Porteous DJ: Disrupted in schizophrenia 1 and
phosphodiesterase 4B: towards an understanding of psychiatric
illness. J Physiol. 584:401–405. 2007.
|
|
115
|
Wang M, Ramos BP, Paspalas CD, Shu Y,
Simen A, Duque A, Vijayraghavan S, Brennan A, Dudley A, Nou E, et
al: α2A-adrenoceptors strengthen working memory networks by
inhibiting cAMP-HCN channel signaling in prefrontal cortex. Cell.
129:397–410. 2007.
|
|
116
|
Wang M, Gamo NJ, Yang Y, Jin LE, Wang XJ,
Laubach M, Mazer JA, Lee D and Arnsten AF: Neuronal basis of
age-related working memory decline. Nature. 476:210–213. 2011.
|
|
117
|
Valero-Aracama MJ, Reboreda A, Arboit A,
Sauvage M and Yoshida M: Noradrenergic suppression of persistent
firing in hippocampal CA1 pyramidal cells through cAMP-PKA pathway.
eNeuro. 8:ENEURO.0440–20.2020. 2021.
|
|
118
|
Ramos BP, Colgan L, Nou E, Ovadia S,
Wilson SR and Arnsten AF: The beta-1 adrenergic antagonist,
betaxolol, improves working memory performance in rats and monkeys.
Biol Psychiatry. 58:894–900. 2005.
|
|
119
|
Ramos BP, Colgan LA, Nou E and Arnsten
AFT: β2 adrenergic agonist, clenbuterol, enhances working memory
performance in aging animals. Neurobiol Aging. 29:1060–1069.
2008.
|
|
120
|
Clark DA, Arranz MJ, Mata I,
Lopéz-Ilundain J, Pérez-Nievas F and Kerwin RW: Polymorphisms in
the promoter region of the alpha1A-adrenoceptor gene are associated
with schizophrenia/schizoaffective disorder in a Spanish isolate
population. Biol Psychiatry. 58:435–439. 2005.
|
|
121
|
Lochman J, Plesník J, Janout V, Povová J,
Míšek I, Dvořáková D and Šerý O: Interactive effect of MTHFR and
ADRA2A gene polymorphisms on pathogenesis of schizophrenia. Neuro
Endocrinol Lett. 34:792–797. 2013.
|
|
122
|
Vares M, Saetre P, Deng H, Cai G, Liu X,
Hansen T, Rasmussen HB, Werge T, Melle I, Djurovic S, et al:
Association between methylenetetrahydrofolate reductase (MTHFR)
C677T polymorphism and age of onset in schizophrenia. Am J Med
Genet B Neuropsychiatr Genet. 153B:610–618. 2010.
|
|
123
|
Lu ML, Ku WC, Syifa N, Hu SC, Chou CT, Wu
YH, Kuo PH, Chen CH, Chen WJ and Wu TH: Developing a sensitive
platform to measure 5-methyltetrahydrofolate in subjects with MTHFR
and PON1 gene polymorphisms. Nutrients. 14:33202022.
|
|
124
|
Dean B and Scarr E: Muscarinic M1 and M4
receptors: Hypothesis driven drug development for schizophrenia.
Psychiatry Res. 288:1129892020.
|
|
125
|
Dean B, Bakker G, Ueda HR, Tobin AB, Brown
A and Kanaan RAA: A growing understanding of the role of muscarinic
receptors in the molecular pathology and treatment of
schizophrenia. Front Cell Neurosci. 17:11243332023.
|
|
126
|
Teal LB, Gould RW, Felts AS and Jones CK:
Selective allosteric modulation of muscarinic acetylcholine
receptors for the treatment of schizophrenia and substance use
disorders. Adv Pharmacol. 86:153–196. 2019.
|
|
127
|
Crook JM, Tomaskovic-Crook E, Copolov DL
and Dean B: Low muscarinic receptor binding in prefrontal cortex
from subjects with schizophrenia: A study of Brodmann's Areas 8, 9,
10, and 46 and the effects of neuroleptic drug treatment. Am J
Psychiatry. 158:918–925. 2001.
|
|
128
|
Zavitsanou K, Katsifis A, Mattner F and
Huang XF: Investigation of M1/M4 muscarinic receptors in the
anterior cingulate cortex in schizophrenia, bipolar disorder, and
major depression disorder. Neuropsychopharmacology. 29:619–625.
2004.
|
|
129
|
Crook JM, Tomaskovic-Crook E, Copolov DL
and Dean B: Decreased muscarinic receptor binding in subjects with
schizophrenia: A study of the human hippocampal formation. Biol
Psychiatry. 48:381–388. 2000.
|
|
130
|
Dean B, Crook JM, Opeskin K, Hill C, Keks
N and Copolov DL: The density of muscarinic M1 receptors is
decreased in the caudate-putamen of subjects with schizophrenia.
Mol Psychiatry. 1:54–58. 1996.
|
|
131
|
Liao DL, Hong CJ, Chen HM, Chen YE, Lee
SM, Chang CY, Chen H and Tsai SJ: Association of muscarinic m1
receptor genetic polymorphisms with psychiatric symptoms and
cognitive function in schizophrenic patients. Neuropsychobiology.
48:72–76. 2003.
|
|
132
|
Scarr E, Craig JM, Cairns MJ, Seo MS,
Galati JC, Beveridge NJ, Gibbons A, Juzva S, Weinrich B,
Parkinson-Bates M, et al: Decreased cortical muscarinic M1
receptors in schizophrenia are associated with changes in gene
promoter methylation, mRNA and gene targeting microRNA. Transl
Psychiatry. 3:e2302013.
|
|
133
|
Scarr E, Um JY, Cowie TF and Dean B:
Cholinergic muscarinic M4 receptor gene polymorphisms: A potential
risk factor and pharmacogenomic marker for schizophrenia. Schizophr
Res. 146:279–284. 2013.
|
|
134
|
De Luca V, Wang H, Squassina A, Wong GW,
Yeomans J and Kennedy JL: Linkage of M5 muscarinic and
alpha7-nicotinic receptor genes on 15q13 to schizophrenia.
Neuropsychobiology. 50:124–127. 2004.
|
|
135
|
Anagnostaras SG, Murphy GG, Hamilton SE,
Mitchell SL, Rahnama NP, Nathanson NM and Silva AJ: Selective
cognitive dysfunction in acetylcholine M1 muscarinic receptor
mutant mice. Nat Neurosci. 6:51–58. 2003.
|
|
136
|
Thomsen M, Wess J, Fulton BS, Fink-Jensen
A and Caine SB: Modulation of prepulse inhibition through both M1
and M4 muscarinic receptors in mice. Psychopharmacology.
208:401–416. 2010.
|
|
137
|
Felder CC, Porter AC, Skillman TL, Zhang
L, Bymaster FP, Nathanson NM, Hamilton SE, Gomeza J, Wess J and
McKinzie DL: Elucidating the role of muscarinic receptors in
psychosis. Life Sci. 68:2605–2613. 2001.
|
|
138
|
Koshimizu H, Leiter LM and Miyakawa T: M4
muscarinic receptor knockout mice display abnormal social behavior
and decreased prepulse inhibition. Mol Brain. 5:102012.
|
|
139
|
Dencker D, Wörtwein G, Weikop P, Jeon J,
Thomsen M, Sager TN, Mørk A, Woldbye DP, Wess J and Fink-Jensen A:
Involvement of a subpopulation of neuronal M4 muscarinic
acetylcholine receptors in the antipsychotic-like effects of the
M1/M4 preferring muscarinic receptor agonist xanomeline. J
Neurosci. 31:5905–5908. 2011.
|
|
140
|
Woolley ML, Carter HJ, Gartlon JE, Watson
JM and Dawson LA: Attenuation of amphetamine-induced activity by
the non-selective muscarinic receptor agonist, xanomeline, is
absent in muscarinic M4 receptor knockout mice and attenuated in
muscarinic M1 receptor knockout mice. Eur J Pharmacol. 603:147–149.
2009.
|
|
141
|
Tzavara ET, Bymaster FP, Davis RJ, Wade
MR, Perry KW, Wess J, McKinzie DL, Felder C and Nomikos GG: M4
muscarinic receptors regulate the dynamics of cholinergic and
dopaminergic neurotransmission: Relevance to the pathophysiology
and treatment of related CNS pathologies. FASEB J. 18:1410–1412.
2004.
|
|
142
|
Zhang W, Yamada M, Gomeza J, Basile AS and
Wess J: Multiple muscarinic acetylcholine receptor subtypes
modulate striatal dopamine release, as studied with M1-M5
muscarinic receptor knock-out mice. J Neurosci. 22:6347–6352.
2002.
|
|
143
|
Thomsen M, Wörtwein G, Fink-Jensen A,
Woldbye DP, Wess J and Caine SB: Decreased prepulse inhibition and
increased sensitivity to muscarinic, but not dopaminergic drugs in
M5 muscarinic acetylcholine receptor knockout mice.
Psychopharmacology. 192:97–110. 2007.
|
|
144
|
Wang H, Ng K, Hayes D, Gao X, Forster G,
Blaha C and Yeomans J: Decreased Amphetamine-induced locomotion and
improved latent inhibition in mice mutant for the M5 muscarinic
receptor gene found in the human 15q schizophrenia region.
Neuropsychopharmacology. 29:2126–2139. 2004.
|
|
145
|
Abad NH, Doulatabad NS, Mohammadi A and
Srazi HR: Treatment of visual hallucinations in schizophrenia by
acetylcholinesterase inhibitors: A case report. Iran J Osychiatry.
6:161–163. 2011.
|
|
146
|
Patel SS, Attard A, Jacobsen P and
Shergill S: Acetylcholinesterase Inhibitors (AChEI's) for the
treatment of visual hallucinations in schizophrenia: A case report.
BMC Psychiatry. 10:682010.
|
|
147
|
Mancama D, Mata I, Kerwin RW and Arranz
MJ: Choline acetyltransferase variants and their influence in
schizophrenia and olanzapine response. Am J Med Genet B
Neuropsychiatr Genet. 144B:849–853. 2007.
|
|
148
|
Niswender CM and Conn PJ: Metabotropic
glutamate receptors: Physiology, pharmacology, and disease. Ann Rev
Pharmacol Toxicol. 50:295–322. 2010.
|
|
149
|
Kim JH, Marton J, Ametamey SM and Cumming
P: A review of molecular imaging of glutamate receptors. Molecules.
25:47492020.
|
|
150
|
Crupi R, Impellizzeri D and Cuzzocrea S:
Role of metabotropic glutamate receptors in neurological disorders.
Front Mol Neurosci. 12:202019.
|
|
151
|
Ayoub MA, Angelicheva D, Vile D, Chandler
D, Morar B, Cavanaugh JA, Visscher PM, Jablensky A, Pfleger KD and
Kalaydjieva L: Deleterious GRM1 mutations in schizophrenia. PLoS
One. 7:e328492012.
|
|
152
|
Volk DW, Eggan SM and Lewis DA:
Alterations in metabotropic glutamate receptor 1α and regulator of
G protein signaling 4 in the prefrontal cortex in schizophrenia. Am
J Psychiatry. 167:1489–1498. 2010.
|
|
153
|
Aiba A, Chen C, Herrup K, Rosenmund C,
Stevens CF and Tonegawa S: Reduced hippocampal long-term
potentiation and context-specific deficit in associative learning
in mGluR1 mutant mice. Cell. 79:365–375. 1994.
|
|
154
|
Gil-Sanz C, Delgado-García JM, Fairén A
and Gruart A: Involvement of the mGluR1 receptor in hippocampal
synaptic plasticity and associative learning in behaving mice.
Cerebral Cortex. 18:1653–1663. 2008.
|
|
155
|
Aiba A, Kano M, Chen C, Stanton ME, Fox
GD, Herrup K, Zwingman TA and Tonegawa S: Deficient cerebellar
long-term depression and impaired motor learning in mGluR1 mutant
mice. Cell. 79:377–388. 1994.
|
|
156
|
Brody SA, Conquet F and Geyer MA:
Disruption of prepulse inhibition in mice lacking mGluR1. Eur J
Neurosci. 18:3361–3366. 2003.
|
|
157
|
Maksymetz J, Moran SP and Conn PJ:
Targeting metabotropic glutamate receptors for novel treatments of
schizophrenia. Mol Brain. 10:152017.
|
|
158
|
Matosin N and Newell KA: Metabotropic
glutamate receptor 5 in the pathology and treatment of
schizophrenia. Neurosci Biobehav Rev. 37:256–268. 2013.
|
|
159
|
Brody SA, Dulawa SC, Conquet F and Geyer
MA: Assessment of a prepulse inhibition deficit in a mutant mouse
lacking mGlu5 receptors. Mol Psychiatry. 9:35–41. 2004.
|
|
160
|
Liang W, Hou Y, Huang W, Wang Y, Jiang T,
Huang X, Wang Z, Wu F, Zheng J, Zhang J, et al: Loss of
schizophrenia-related miR-501-3p in mice impairs sociability and
memory by enhancing mGluR5-mediated glutamatergic transmission. Sci
Adv. 8:eabn73572022.
|
|
161
|
Campbell UC, Lalwani K, Hernandez L,
Kinney GG, Conn PJ and Bristow LJ: The mGluR5 antagonist
2-methyl-6-(phenylethynyl)-pyridine (MPEP) potentiates PCP-induced
cognitive deficits in rats. Psychopharmacology. 175:310–318.
2004.
|
|
162
|
Henry SA, Lehmann-Masten V, Gasparini F,
Geyer MA and Markou A: The mGluR5 antagonist MPEP, but not the
mGluR2/3 agonist LY314582, augments PCP effects on prepulse
inhibition and locomotor activity. Neuropharmacology. 43:1199–1209.
2002.
|
|
163
|
Homayoun H, Stefani MR, Adams BW, Tamagan
GD and Moghaddam B: Functional interaction between NMDA and mGlu5
receptors: Effects on working memory, instrumental learning, motor
behaviors, and dopamine release. Neuropsychopharmacology.
29:1259–1269. 2004.
|
|
164
|
Zou D, Huang J, Wu X and Li L:
Metabotropic glutamate subtype 5 receptors modulate
fear-conditioning induced enhancement of prepulse inhibition in
rats. Neuropharmacology. 52:476–486. 2007.
|
|
165
|
Semenova S and Markou A: The effects of
the mGluR5 antagonist MPEP and the mGluR2/3 antagonist LY341495 on
rats' performance in the 5-choice serial reaction time task.
Neuropharmacology. 52:863–872. 2007.
|
|
166
|
Trepanier C, Lei G, Xie YF and Macdonald
JF: Group II metabotropic glutamate receptors modify
N-methyl-D-aspartate receptors via Src kinase. Sci Rep.
3:9262013.
|
|
167
|
Tyszkiewicz JP, Gu Z, Wang X, Cai X and
Yan Z: Group II metabotropic glutamate receptors enhance NMDA
receptor currents via a protein kinase C-dependent mechanism in
pyramidal neurones of rat prefrontal cortex. J Physiol.
554:765–777. 2004.
|
|
168
|
Cheng J, Liu W, Duffney LJ and Yan Z:
SNARE proteins are essential in the potentiation of NMDA receptors
by group II metabotropic glutamate receptors. J Physiol.
591:3935–3947. 2013.
|
|
169
|
Uslaner JM, Smith SM, Huszar SL,
Pachmerhiwala R, Hinchliffe RM, Vardigan JD and Hutson PH: Combined
administration of an mGlu2/3 receptor agonist and a 5-HT 2A
receptor antagonist markedly attenuate the psychomotoractivating
and neurochemical effects of psychostimulants. Psychopharmacology.
206:641–651. 2009.
|
|
170
|
Campusano JM, Abarca J, Forray MI, Gysling
K and Bustos G: Modulation of dendritic release of dopamine by
metabotropic glutamate receptors in rat substantia nigra. Biochem
Pharmacol. 63:1343–1352. 2002.
|
|
171
|
Chaki S, Yoshikawa R and Okuyama S: Group
II metabotropic glutamate receptor-mediated regulation of dopamine
release from slices of rat nucleus accumbens. Neurosci Lett.
404:182–186. 2006.
|
|
172
|
Hu G, Duffy P, Swanson C, Ghasemzadeh MB
and Kalivas PW: The regulation of dopamine transmission by
metabotropic glutamate receptors. J Pharmacol Exp Ther.
289:412–416. 1999.
|
|
173
|
Johnson KA, Mateo Y and Lovinger DM:
Metabotropic glutamate receptor 2 inhibits thalamically-driven
glutamate and dopamine release in the dorsal striatum.
Neuropharmacology. 117:114–123. 2017.
|
|
174
|
Schoepp DD, Jane DE and Monn JA:
Pharmacological agents acting at subtypes of metabotropic glutamate
receptors. Neuropharmacology. 38:1431–1476. 1999.
|
|
175
|
Moghaddam B and Adams BW: Reversal of
phencyclidine effects by a group II metabotropic glutamate receptor
agonist in rats. Science. 281:1349–1352. 1998.
|
|
176
|
Cartmell J, Monn JA and Schoepp DD: The
metabotropic glutamate 2/3 receptor agonists LY354740 and LY379268
selectively attenuate phencyclidine versus d-amphetamine motor
behaviors in rats. J Pharmacol Exp Ther. 291:161–170. 1999.
|
|
177
|
Galici R, Echemendia NG, Rodriguez AL and
Conn PJ: A selective allosteric potentiator of metabotropic
glutamate (mGlu) 2 receptors has effects similar to an orthosteric
mGlu2/3 receptor agonist in mouse models predictive of
antipsychotic activity. J Pharmacol Exp Ther. 315:1181–1187.
2005.
|
|
178
|
Kawaura K, Karasawa J and Hikichi H:
Stimulation of the metabotropic glutamate (mGlu) 2 receptor
attenuates the MK-801-induced increase in the immobility time in
the forced swimming test in rats. Pharmacol Rep. 68:80–84.
2016.
|
|
179
|
Homayoun H, Jackson ME and Moghaddam B:
Activation of metabotropic glutamate 2/3 receptors reverses the
effects of NMDA receptor hypofunction on prefrontal cortex unit
activity in awake rats. J Neurophysiol. 93:1989–2001. 2005.
|
|
180
|
Kłodzinska A, Bijak M, Tokarski K and Pilc
A: Group II mGlu receptor agonists inhibit behavioural and
electrophysiological effects of DOI in mice. Pharmacol Biochem
Behav. 73:327–332. 2002.
|
|
181
|
Matrisciano F, Tueting P, Maccari S,
Nicoletti F and Guidotti A: Pharmacological activation of group-II
metabotropic glutamate receptors corrects a schizophrenia-like
phenotype induced by prenatal stress in mice.
Neuropsychopharmacology. 37:929–938. 2012.
|
|
182
|
Jones CA, Brown AM, Auer DP and Fone KCF:
The mGluR2/3 agonist LY379268 reverses post-weaning social
isolation-induced recognition memory deficits in the rat.
Psychopharmacology. 214:269–283. 2011.
|
|
183
|
Harich S, Gross G and Bespalov A:
Stimulation of the metabotropic glutamate 2/3 receptor attenuates
social novelty discrimination deficits induced by neonatal
phencyclidine treatment. Psychopharmacology. 192:511–519. 2007.
|
|
184
|
Hikichi H, Kaku A, Karasawa JI and Chaki
S: Stimulation of metabotropic glutamate (mGlu) 2 receptor and
blockade of mGlu1 receptor improve social memory impairment
elicited by MK-801 in rats. J Pharmacol Sci. 122:10–16. 2013.
|
|
185
|
Wierońska JM, Acher FC, Sławińska A, Gruca
P, Łasoń-Tyburkiewicz M, Papp M and Pilc A: The antipsychotic-like
effects of the mGlu group III orthosteric agonist, LSP1-2111,
involves 5-HT1A signalling. Psychopharmacology. 227:711–725.
2013.
|
|
186
|
Greco B, Invernizzi RW and Carli M:
Phencyclidine-induced impairment in attention and response control
depends on the background genotype of mice: Reversal by the mGLU2/3
receptor agonist LY379268. Psychopharmacology. 179:68–76. 2005.
|
|
187
|
Higgins GA, Ballard TM, Kew JN, Richards
JG, Kemp JA, Adam G, Woltering T, Nakanishi S and Mutel V:
Pharmacological manipulation of mGlu2 receptors influences
cognitive performance in the rodent. Neuropharmacology. 46:907–917.
2004.
|
|
188
|
Amitai N and Markou A: Effects of
metabotropic glutamate receptor 2/3 agonism and antagonism on
schizophrenia-like cognitive deficits induced by phencyclidine in
rats. Eur J Pharmacol. 639:67–80. 2010.
|
|
189
|
González-Maeso J, Ang RL, Yuen T, Chan P,
Weisstaub NV, López-Giménez JF, Zhou M, Okawa Y, Callado LF,
Milligan G, et al: Identification of a serotonin/glutamate receptor
complex implicated in psychosis. Nature. 452:93–97. 2008.
|
|
190
|
Fribourg M, Moreno JL, Holloway T, Provasi
D, Baki L, Mahajan R, Park G, Adney SK, Hatcher C, Eltit JM, et al:
Decoding the signaling of a GPCR heteromeric complex reveals a
unifying mechanism of action of antipsychotic drugs. Cell.
147:1011–1023. 2011.
|
|
191
|
Moreno JL, Miranda-Azpiazu P, García-Bea
A, Younkin J, Cui M, Kozlenkov A, Ben-Ezra A, Voloudakis G, Fakira
AK, Baki L, et al: Allosteric signaling through an mGlu2 and 5-HT2A
heteromeric receptor complex and its potential contribution to
schizophrenia. Sci Signal. 9:ra52016.
|
|
192
|
Kurita M, Holloway T, García-Bea A,
Kozlenkov A, Friedman AK, Moreno JL, Heshmati M, Golden SA, Kennedy
PJ, Takahashi N, et al: HDAC2 regulates atypical antipsychotic
responses through the modulation of mGlu2 promoter activity. Nat
Neurosci. 15:1245–1254. 2012.
|
|
193
|
Pałucha-Poniewiera A, Kłodzińska A,
Stachowicz K, Tokarski K, Hess G, Schann S, Frauli M, Neuville P
and Pilc A: Peripheral administration of group III mGlu receptor
agonist ACPT-I exerts potential antipsychotic effects in rodents.
Neuropharmacology. 55:517–524. 2008.
|
|
194
|
Pekhletski R, Gerlai R, Overstreet LS,
Huang X-P, Agopyan N, Slater NT, Abramow-Newerly W, Roder JC and
Hampson DR: Impaired cerebellar synaptic plasticity and motor
performance in mice lacking the mGluR4 subtype of metabotropic
glutamate receptor. J Neurosci. 16:6364–6373. 1996.
|
|
195
|
Gerlai R, Roder JC and Hampson DR: Altered
spatial learning and memory in mice lacking the mGluR4 subtype of
metabotropic glutamate receptor. Behav Neurosci. 112:525–532.
1998.
|
|
196
|
Snead OC, Banerjee PK, Burnham M and
Hampson D: Modulation of absence seizures by the GABA(A) receptor:
A critical role for metabotropic glutamate receptor 4 (mGluR4). J
Neurosci. 20:6218–6224. 2000.
|
|
197
|
Wierońska JM, Stachowicz K, Acher F, Lech
T and Pilc A: Opposing efficacy of group III mGlu receptor
activators, LSP1-2111 and AMN082, in animal models of positive
symptoms of schizophrenia. Psychopharmacology. 220:481–494.
2012.
|
|
198
|
Woźniak M, Acher F, Marciniak M,
Lasoń-Tyburkiewicz M, Gruca P, Papp M, Pilc A and Wierońska JM:
Involvement of GABAB receptor signaling in Antipsychotic-like
action of the novel orthosteric agonist of the mGlu4 receptor,
LSP4-2022. Curr Neuropharmacol. 14:413–426. 2016.
|
|
199
|
Masu M, Iwakabe H, Tagawa Y, Miyoshi T,
Yamashita M, Fukuda Y, Sasaki H, Hiroi K, Nakamura Y, Shigemoto R,
et al: Specific deficit of the ON response in visual transmission
by targeted disruption of the mGluR6 gene. Cell. 80:757–765.
1995.
|
|
200
|
Sugihara H, Inoue T, Nakanishi S and
Fukuda Y: A late ON response remains in visual response of the
mGluR6-deficient mouse. Neuroscience Lett. 233:137–140. 1997.
|
|
201
|
Hosak L, Sery O, Sadykov E and Studnicka
J: Retinal abnormatilites as a diagnostic or prognostic marker of
schizophrenia. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub.
162:159–164. 2018.
|
|
202
|
Bernardin F, Schwitzer T, Angioi-Duprez K,
Giersch A, Jansen C, Schwan R and Laprevote V: Retinal ganglion
cells dysfunctions in schizophrenia patients with or without visual
hallucinations. Schizophr Res. 219:47–55. 2020.
|
|
203
|
Sansig G, Bushell TJ, Clarke VRJ, Rozov A,
Burnashev N, Portet C, Gasparini F, Schmutz M, Klebs K, Shigemoto
R, et al: Increased seizure susceptibility in mice lacking
metabotropic glutamate receptor 7. J Neurosci. 21:8734–8745.
2001.
|
|
204
|
O'Connor RM, Finger BC, Flor PJ and Cryan
JF: Metabotropic glutamate receptor 7: At the interface of
cognition and emotion. Eur J Pharmacol. 639:123–131. 2010.
|
|
205
|
Bushell TJ, Sansig G, Collett VJ, Van Der
Putten H and Collingridge GL: Altered Short-term synaptic
plasticity in mice lacking the metabotropic glutamate receptor
mGlu7. ScientificWorldJournal. 2:730–737. 2002.
|
|
206
|
Hölscher C, Schmid S, Pilz PK, Sansig G,
van der Putten H and Plappert CF: Lack of the metabotropic
glutamate receptor subtype 7 selectively impairs short-term working
memory but not long-term memory. Behav Brain Res. 154:473–481.
2004.
|
|
207
|
Hölscher C, Schmid S, Pilz PKD, Sansig G,
Van Der Putten H and Plappert CF: Lack of the metabotropic
glutamate receptor subtype 7 selectively modulates Theta rhythm and
working memory. Learning Memory. 12:450–455. 2005.
|
|
208
|
Goddyn H, Callaerts-Vegh Z, Stroobants S,
Dirikx T, Vansteenwegen D, Hermans D, van der Putten H and D'Hooge
R: Deficits in acquisition and extinction of conditioned responses
in mGluR7 knockout mice. Neurobiol Learn Mem. 90:103–111. 2008.
|
|
209
|
Callaerts-Vegh Z, Beckers T, Ball SM,
Baeyens F, Callaerts PF, Cryan FJ, Molnar E and D'Hooge R:
Concomitant deficits in working memory and fear extinction are
functionally dissociated from reduced anxiety in metabotropic
glutamate receptor 7-deficient mice. J Neurosci. 26:6573–6582.
2006.
|
|
210
|
Masugi M, Yokoi M, Shigemoto R, Muguruma
K, Watanabe Y, Sansig G, van der Putten H and Nakanishi S:
Metabotropic glutamate receptor subtype 7 ablation causes deficit
in fear response and conditioned taste aversion. J Neurosci.
19:955–963. 1999.
|
|
211
|
Kalinichev M, Rouillier M, Girard F,
Royer-Urios I, Bournique B, Finn T, Charvin D, Campo B, Le Poul E,
Mutel V, et al: ADX71743, a potent and selective negative
allosteric modulator of metabotropic glutamate receptor 7: In vitro
and In vivo characterization. J Pharmacol Exp Ther. 344:624–636.
2013.
|
|
212
|
Suzuki G, Tsukamoto N, Fushiki H,
Kawagishi A, Nakamura M, Kurihara H, Mitsuya M, Ohkubo M and Ohta
H: In vitro pharmacological characterization of novel
isoxazolopyridone derivatives as allosteric metabotropic glutamate
receptor 7 antagonists. J Pharmacol Exp Ther. 323:147–156.
2007.
|
|
213
|
Cieślik P, Woźniak M, Kaczorowska K,
Brański P, Burnat G, Chocyk A, Bobula B, Gruca P, Litwa E,
Pałucha-Poniewiera A, et al: Negative allosteric modulators of
mGlu7 receptor as putative antipsychotic drugs. Front Mol Neurosci.
11:3162018.
|
|
214
|
Zhai J, Tian MT, Wang Y, Yu JL, Köster A,
Baez M and Nisenbaum ES: Modulation of lateral perforant path
excitatory responses by metabotropic glutamate 8 (mGlu8) receptors.
Neuropharmacology. 43:223–230. 2002.
|
|
215
|
Gerlai R, Adams B, Fitch T, Chaney S and
Baez M: Performance deficits of mGluR8 knockout mice in learning
tasks: The effects of null mutation and the background genotype.
Neuropharmacology. 43:235–249. 2002.
|
|
216
|
Robbins MJ, Starr KR, Honey A, Soffin EM,
Rourke C, Jones GA, Kelly FM, Strum J, Melarange RA, Harris AJ, et
al: Evaluation of the mGlu8 receptor as a putative therapeutic
target in schizophrenia. Brain Res. 1152:215–227. 2007.
|
|
217
|
Gicquel T, Le Daré B, Boichot E and
Lagente V: Purinergic receptors: New targets for the treatment of
gout and fibrosis. Fundam Clin Pharmacol. 31:136–146. 2017.
|
|
218
|
Magni G and Ceruti S: P2Y purinergic
receptors: New targets for analgesic and antimigraine drugs.
Biochem Pharmacol. 85:466–477. 2013.
|
|
219
|
Weissman TA, Riquelme PA, Ivic L, Flint AC
and Kriegstein AR: Calcium waves propagate through radial glial
cells and modulate proliferation in the developing neocortex.
Neuron. 43:647–661. 2004.
|
|
220
|
Ribeiro DE, Glaser T, Oliveira-Giacomelli
Á and Ulrich H: Purinergic receptors in neurogenic processes. Brain
Res Bull. 151:3–11. 2019.
|
|
221
|
Koch H, Bespalov A, Drescher K, Franke H
and Krügel U: Impaired cognition after stimulation of P2Y1
receptors in the rat medial prefrontal cortex.
Neuropsychopharmacology. 40:305–314. 2015.
|
|
222
|
Huang L, Otrokocsi L and Sperlágh B: Role
of P2 receptors in normal brain development and in
neurodevelopmental psychiatric disorders. Brain Res Bull.
151:55–64. 2019.
|
|
223
|
Leung LS and Ma J: Medial septum modulates
hippocampal gamma activity and prepulse inhibition in an
N-methyl-d-aspartate receptor antagonist model of schizophrenia.
Schizophr Res. 198:36–44. 2018.
|
|
224
|
Schroeder A, Hudson M, Du X, Wu YWC,
Nakamura J, van den Buuse M, Jones NC and Hill RA: Estradiol and
raloxifene modulate hippocampal gamma oscillations during a spatial
memory task. Psychoneuroendocrinology. 78:85–92. 2017.
|
|
225
|
Lindberg D, Shan D, Ayers-Ringler J,
Oliveros A, Benitez J, Prieto M, McCullumsmith R and Choi DS:
Purinergic signaling and energy homeostasis in psychiatric
disorders. Curr Mol Med. 15:275–295. 2015.
|
|
226
|
Cheffer A, Castillo ARG, Corrêa-Velloso J,
Gonçalves MCB, Naaldijk Y, Nascimento IC, Burnstock G and Ulrich H:
Purinergic system in psychiatric diseases. Mol Psychiatry.
23:94–106. 2018.
|
|
227
|
Lara DR, Dall'Igna OP, Ghisolfi ES and
Brunstein MG: Involvement of adenosine in the neurobiology of
schizophrenia and its therapeutic implications. Prog
Neuropsychopharmacol Biol Psychiatry. 30:617–629. 2006.
|
|
228
|
Matos M, Shen HY, Augusto E, Wang Y, Wei
CJ, Wang YT, Agostinho P, Boison D, Cunha RA and Chen JF: Deletion
of adenosine A2A receptors from astrocytes disrupts glutamate
homeostasis leading to psychomotor and cognitive impairment:
Relevance to schizophrenia. Biol Psychiatry. 78:763–774. 2015.
|
|
229
|
Marques TR, Natesan S, Rabiner EA, Searle
GE, Gunn R, Howes OD and Kapur S: Adenosine A2A receptor in
schizophrenia: An In vivo brain PET imaging study.
Psychopharmacology. 239:3439–3445. 2022.
|
|
230
|
Wexler EM, Paucer A, Kornblum HI, Palmer
TD and Geschwind DH: Endogenous Wnt signaling maintains neural
progenitor cell potency. Stem Cells. 27:1130–1141. 2009.
|
|
231
|
Niehrs C and Acebron SP: Mitotic and
mitogenic Wnt signalling. EMBO J. 31:2705–2713. 2012.
|
|
232
|
Huang YL and Niehrs C: Polarized wnt
signaling regulates ectodermal cell fate in xenopus. Dev Cell.
29:250–257. 2014.
|
|
233
|
Loh Kyle M, van Amerongen R and Nusse R:
Generating cellular diversity and spatial form: Wnt signaling and
the evolution of multicellular animals. Dev Cell. 38:643–655.
2016.
|
|
234
|
Vallée A: Neuroinflammation in
schizophrenia: The key role of the WNT/β-catenin pathway. Int J Mol
Sci. 23:28102022.
|
|
235
|
Anand AA, Khan M, V M and Kar D: The
molecular basis of Wnt/β-catenin signaling pathways in
neurodegenerative diseases. Int J Cell Biol. 2023:92960922023.
|
|
236
|
Al-Harthi L: Wnt/β-catenin and its diverse
physiological cell signaling pathways in neurodegenerative and
neuropsychiatric disorders. J Neuroimmune Pharmacol. 7:725–730.
2012.
|
|
237
|
Schmitt A, Simons M, Cantuti-Castelvetri L
and Falkai P: A new role for oligodendrocytes and myelination in
schizophrenia and affective disorders? Eur Arch Psychiatry Clin
Neurosci. 269:371–372. 2019.
|
|
238
|
Cotter D, Kerwin R, al-Sarraji S, Brion
JP, Chadwich A, Lovestone S, Anderton B and Everall I:
Abnormalities of Wnt signalling in schizophrenia-evidence for
neurodevelopmental abnormality. Neuroreport. 9:1379–1383. 1998.
|
|
239
|
Liu X, Low SK, Atkins JR, Wu JQ, Reay WR,
Cairns HM, Green MJ, Schall U, Jablensky A, Mowry B, et al: Wnt
receptor gene FZD1 was associated with schizophrenia in genome-wide
SNP analysis of the Australian Schizophrenia Research Bank cohort.
Aust N Z J Psychiatry. 54:902–908. 2020.
|
|
240
|
Katsu T, Ujike H, Nakano T, Tanaka Y,
Nomura A, Nakata K, Takaki M, Sakai A, Uchida N, Imamura T and
Kuroda S: The human frizzled-3 (FZD3) gene on chromosome 8p21, a
receptor gene for Wnt ligands, is associated with the
susceptibility to schizophrenia. Neurosci Lett. 353:53–56.
2003.
|
|
241
|
Zhang Y, Yu X, Yuan Y, Ling Y, Ruan Y, Si
T, Lu T, Wu S, Gong X, Zhu Z, et al: Positive association of the
human frizzled 3 (FZD3) gene haplotype with schizophrenia in
Chinese Han population. Am J Med Genet B Neuropsychiatr Genet.
129B:16–19. 2004.
|
|
242
|
Yang J, Si T, Ling Y, Ruan Y, Han Y, Wang
X, Zhang H, Kong Q, Li X, Liu C, et al: Association study of the
human FZD3 locus with schizophrenia. Biol Psychiatry. 54:1298–1301.
2003.
|
|
243
|
Kishimoto M, Ujike H, Okahisa Y, Kotaka T,
Takaki M, Kodama M, Inada T, Yamada M, Uchimura N, Iwata N, et al:
The Frizzled 3 gene is associated with methamphetamine psychosis in
the Japanese population. Behav Brain Funct. 4:372008.
|
|
244
|
Hoseth EZ, Krull F, Dieset I, Mørch RH,
Hope S, Gardsjord ES, Steen NE, Melle I, Brattbakk HR, Steen VM, et
al: Exploring the Wnt signaling pathway in schizophrenia and
bipolar disorder. Transl Psychiatry. 8:552018.
|
|
245
|
Castillo PE, Younts TJ, Chávez AE and
Hashimotodani Y: Endocannabinoid signaling and synaptic function.
Neuron. 76:70–81. 2012.
|
|
246
|
Pertwee RG, Howlett AC, Abood ME,
Alexander SP, Di Marzo V, Elphick MR, Greasley PJ, Hansen HS, Kunos
G, Mackie K, et al: International union of basic and clinical
pharmacology. LXXIX. Cannabinoid receptors and their ligands:
Beyond CB1 and CB2. Pharmacol Rev.
62:588–631. 2010.
|
|
247
|
Downer EJ: Cannabinoids and innate
immunity: Taking a toll on neuroinflammation.
ScientificWorldJournal. 11:855–865. 2011.
|
|
248
|
Galindo L, Moreno E, López-Armenta F,
Guinart D, Cuenca-Royo A, Izquierdo-Serra M, Xicota L, Fernandez C,
Menoyo E, Fernández-Fernández JM, et al: Cannabis users show
enhanced expression of CB1-5HT2A receptor
heteromers in olfactory neuroepithelium cells. Mol Neurobiol.
55:6347–6361. 2018.
|
|
249
|
Compagnucci C, Di Siena S, Bustamante MB,
Di Giacomo D, Di Tommaso M, Maccarrone M, Grimaldi P and Sette C:
Type-1 (CB1) cannabinoid receptor promotes neuronal differentiation
and maturation of neural stem cells. PLoS One. 8:e542712013.
|
|
250
|
Goncalves MB, Suetterlin P, Yip P,
Molina-Holgado F, Walker DJ, Oudin MJ, Zentar MP, Pollard S,
Yáñez-Muñoz RJ, Williams G, et al: A diacylglycerol lipase-CB2
cannabinoid pathway regulates adult subventricular zone
neurogenesis in an age-dependent manner. Mol Cell Neurosci.
38:526–536. 2008.
|
|
251
|
Molina-Holgado F, Rubio-Araiz A,
García-Ovejero D, Williams RJ, Moore JD, Arévalo-Martín A,
Gómez-Torres O and Molina-Holgado E: CB2 cannabinoid receptors
promote mouse neural stem cell proliferation. Eur J Neurosci.
25:629–634. 2007.
|
|
252
|
Palazuelos J, Aguado T, Egia A, Mechoulam
R, Guzmán M and Galve-Roperh I: Non-psychoactive CB2 cannabinoid
agonists stimulate neural progenitor proliferation. FASEB J.
20:2405–2407. 2006.
|
|
253
|
Palazuelos J, Ortega Z, Díaz-Alonso J,
Guzmán M and Galve-Roperh I: CB2 cannabinoid receptors promote
neural progenitor cell proliferation via mTORC1 signaling. J Biol
Chem. 287:1198–1209. 2012.
|
|
254
|
Müller-Vahl KR and Emrich HM: Cannabis and
schizophrenia: Towards a cannabinoid hypothesis of schizophrenia.
Expert Rev Neurother. 8:1037–1048. 2008.
|
|
255
|
Navarro D, Gasparyan A, Navarrete F,
Torregrosa AB, Rubio G, Marín-Mayor M, Acosta GB, Garcia-Gutiérrez
MS and Manzanares J: Molecular alterations of the endocannabinoid
system in psychiatric disorders. Int J Mol Sci. 23:47642022.
|
|
256
|
Tao R, Li C, Jaffe AE, Shin JH,
Deep-Soboslay A, Yamin R, Weinberger DR, Hyde TM and Kleinman JE:
Cannabinoid receptor CNR1 expression and DNA methylation in human
prefrontal cortex, hippocampus and caudate in brain development and
schizophrenia. Transl Psychiatry. 10:1582020.
|
|
257
|
Volk DW, Eggan SM, Horti AG, Wong DF and
Lewis DA: Reciprocal alterations in cortical cannabinoid receptor 1
binding relative to protein immunoreactivity and transcript levels
in schizophrenia. Schizophr Res. 159:124–129. 2014.
|
|
258
|
Wong DF, Kuwabara H, Horti AG, Raymont V,
Brasic J, Guevara M, Ye W, Dannals RF, Ravert HT, Nandi A, et al:
Quantification of cerebral cannabinoid receptors subtype 1 (CB1) in
healthy subjects and schizophrenia by the novel PET radioligand
[11C]OMAR. Neuroimage. 52:1505–1513. 2010.
|
|
259
|
García-Gutiérrez MS, García-Bueno B, Zoppi
S, Leza JC and Manzanares J: Chronic blockade of cannabinoid CB2
receptors induces anxiolytic-like actions associated with
alterations in GABA(A) receptors. Br J Pharmacol. 165:951–964.
2012.
|
|
260
|
Hu B, Doods H, Treede RD and Ceci A:
Depression-like behaviour in rats with mononeuropathy is reduced by
the CB2-selective agonist GW405833. Pain. 143:206–212. 2009.
|
|
261
|
Suárez J, Llorente R, Romero-Zerbo SY,
Mateos B, Bermúdez-Silva FJ, de Fonseca FR and Viveros MP: Early
maternal deprivation induces gender-dependent changes on the
expression of hippocampal CB(1) and CB(2) cannabinoid receptors of
neonatal rats. Hippocampus. 19:623–632. 2009.
|
|
262
|
Hamdani N, Tabeze JP, Ramoz N, Ades J,
Hamon M, Sarfati Y, Boni C and Gorwood P: The CNR1 gene as a
pharmacogenetic factor for antipsychotics rather than a
susceptibility gene for schizophrenia. Eur Neuropsychopharmacol.
18:34–40. 2008.
|
|
263
|
Pan Y, Gao F, Zhao S, Han J and Chen F:
Role of the SphK-S1P-S1PRs pathway in invasion of the nervous
system by SARS-CoV-2 infection. Clin Exp Pharmacol Physiol.
48:637–650. 2021.
|
|
264
|
Calise S, Blescia S, Cencetti F,
Bernacchioni C, Donati C and Bruni P: Sphingosine 1-phosphate
stimulates proliferation and migration of satellite cells: Role of
S1P receptors. Biochim Biophys Acta. 1823:439–450. 2012.
|
|
265
|
Shen H, Zhou E, Wei X, Fu Z, Niu C, Li Y,
Pan B, Mathew AV, Wang X, Pennathur S, et al: High density
lipoprotein promotes proliferation of adipose-derived stem cells
via S1P1 receptor and Akt, ERK1/2 signal pathways. Stem Cell Res
Ther. 6:952015.
|
|
266
|
Kimura T, Boehmler AM, Seitz G, Kuçi S,
Wiesner T, Brinkmann V, Kanz L and Möhle R: The sphingosine
1-phosphate receptor agonist FTY720 supports CXCR4-dependent
migration and bone marrow homing of human CD34+ progenitor cells.
Blood. 103:4478–4486. 2004.
|
|
267
|
Anderson G and Maes M: Reconceptualizing
adult neurogenesis: Role for sphingosine-1-phosphate and fibroblast
growth factor-1 in co-ordinating astrocyte-neuronal precursor
interactions. CNS Neurol Disord Drug Targets. 13:126–136. 2014.
|
|
268
|
Esaki K, Balan S, Iwayama Y,
Shimamoto-Mitsuyama C, Hirabayashi Y, Dean B and Yoshikawa T:
Evidence for altered metabolism of sphingosine-1-phosphate in the
corpus callosum of patients with schizophrenia. Schizophr Bull.
46:1172–1181. 2020.
|
|
269
|
Chand GB, Jiang H, Miller JP, Rhodes CH,
Tu Z and Wong DF: Differential Sphingosine-1-phosphate Receptor-1
protein expression in the dorsolateral prefrontal cortex between
schizophrenia type 1 and type 2. Front Psychiatry.
13:8279812022.
|
|
270
|
Brothers SP and Wahlestedt C: Therapeutic
potential of neuropeptide Y (NPY) receptor ligands. EMBO Mol Med.
2:429–439. 2010.
|
|
271
|
Lindner D, Stichel J and Beck-Sickinger
AG: Molecular recognition of the NPY hormone family by their
receptors. Nutrition. 24:907–917. 2008.
|
|
272
|
Howell OW, Doyle K, Goodman JH, Scharfman
HE, Herzog H, Pringle A, Beck-Sickinger AG and Gray WP:
Neuropeptide Y stimulates neuronal precursor proliferation in the
post-natal and adult dentate gyrus. J Neurochem. 93:560–570.
2005.
|
|
273
|
Hansel DE, Eipper BA and Ronnett GV:
Neuropeptide Y functions as a neuroproliferative factor. Nature.
410:940–944. 2001.
|
|
274
|
Montani G, Tonelli S, Elsaesser R, Paysan
J and Tirindelli R: Neuropeptide Y in the olfactory microvillar
cells. Eur J Neurosci. 24:20–24. 2006.
|
|
275
|
Santos-Carvalho A, Elvas F, Alvaro AR,
Ambrósio AF and Cavadas C: Neuropeptide Y receptors activation
protects rat retinal neural cells against necrotic and apoptotic
cell death induced by glutamate. Cell Death Dis. 4:e6362013.
|
|
276
|
Thiriet N, Agasse F, Nicoleau C, Guégan C,
Vallette F, Cadet JL, Jaber M, Malva JO and Coronas V: NPY promotes
chemokinesis and neurogenesis in the rat subventricular zone. J
Neurochem. 116:1018–1027. 2011.
|
|
277
|
Spencer B, Potkar R, Metcalf J, Thrin I,
Adame A, Rockenstein E and Masliah E: Systemic central nervous
system (CNS)-targeted delivery of neuropeptide Y (NPY) reduces
neurodegeneration and increases neural precursor cell proliferation
in a mouse model of alzheimer disease. J Biol Chem. 291:1905–1920.
2016.
|
|
278
|
Decressac M, Prestoz L, Veran J, Cantereau
A, Jaber M and Gaillard A: Neuropeptide Y stimulates proliferation,
migration and differentiation of neural precursors from the
subventricular zone in adult mice. Neurobiol Dis. 34:441–449.
2009.
|
|
279
|
Mellios N, Huang HS, Baker SP, Galdzicka
M, Ginns E and Akbarian S: Molecular determinants of dysregulated
GABAergic gene expression in the prefrontal cortex of subjects with
schizophrenia. Biol Psychiatry. 65:1006–1014. 2009.
|
|
280
|
Morris HM, Stopczynski RE and Lewis DA:
NPY mRNA expression in the prefrontal cortex: Selective reduction
in the superficial white matter of subjects with schizoaffective
disorder. Schizophr Res. 115:261–269. 2009.
|
|
281
|
Stadlbauer U, Langhans W and Meyer U:
Administration of the Y2 receptor agonist PYY3-36 in mice induces
multiple behavioral changes relevant to schizophrenia.
Neuropsychopharmacology. 38:2446–2455. 2013.
|
|
282
|
Stahl SM: Beyond the dopamine hypothesis
of schizophrenia to three neural networks of psychosis: Dopamine,
serotonin, and glutamate. CNS Spectrums. 23:187–191. 2018.
|
|
283
|
Miller MC and Mayo KH: Chemokines from a
structural perspective. Int J Mol Sci. 18:20882017.
|
|
284
|
Ermakov EA, Mednova IA, Boiko AS, Buneva
VN and Ivanova SA: Chemokine dysregulation and neuroinflammation in
schizophrenia: A systematic review. Int J Mol Sci. 24:22152023.
|
|
285
|
Sugiyama T, Kohara H, Noda M and Nagasawa
T: Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4
chemokine signaling in bone marrow stromal cell niches. Immunity.
25:977–988. 2006.
|
|
286
|
Ishida Y, Kimura A, Kuninaka Y, Inui M,
Matsushima K, Mukaida N and Kondo T: Pivotal role of the CCL5/CCR5
interaction for recruitment of endothelial progenitor cells in
mouse wound healing. J Clin Invest. 122:711–721. 2012.
|
|
287
|
Jiang Z, Li Y, Ji X, Tang Y, Yu H, Ding L,
Yu M, Cui Q, Zhang M, Ma Y and Li M: Protein profiling identified
key chemokines that regulate the maintenance of human pluripotent
stem cells. Sci Rep. 7:145102017.
|
|
288
|
Bajetto A, Bonavia R, Barbero S, Florio T
and Schettini G: Chemokines and their receptors in the central
nervous system. Front Neuroendocrinol. 22:147–184. 2001.
|
|
289
|
Tiihonen J, Koskuvi M, Lähteenvuo M,
Trontti K, Ojansuu I, Vaurio O, Cannon TD, Lönnqvist J, Therman S,
Suvisaari J, et al: Molecular signaling pathways underlying
schizophrenia. Schizophr Res. 232:33–41. 2021.
|
|
290
|
Simon IA, Bjørn-Yoshimoto WE, Harpsøe K,
Iliadis S, Svensson B, Jensen AA and Gloriam DE: Ligand selectivity
hotspots in serotonin GPCRs. Trends Pharmacol Sci. 44:978–990.
2023.
|
|
291
|
Littlepage-Saunders M, Hochstein MJ, Chang
DS and Johnson KA: G protein-coupled receptor modulation of
striatal dopamine transmission: Implications for psychoactive drug
effects. Br J Pharmacol. May 31–2023. View Article : Google Scholar : Epub ahead of
print.
|
|
292
|
Valencia M, Medina R, Calixto E and
Rodríguez N: Cerebral, psychosocial, family functioning and
disability of persons with schizophrenia. Neuropsychiat Dis Treat.
18:2069–2082. 2022.
|
|
293
|
Krogmann A, Peters L, Von Hardenberg L,
Bödeker K, Nöhles VB and Correll CU: Keeping up with the
therapeutic advances in schizophrenia: A review of novel and
emerging pharmacological entities. CNS Spectrums. 24:38–69.
2019.
|
|
294
|
Borroto-Escuela DO, Carlsson J, Ambrogini
P, Narváez M, Wydra K, Tarakanov AO, Li X, Millón C, Ferraro L,
Cuppini R, et al: Understanding the role of GPCR heteroreceptor
complexes in modulating the brain networks in health and disease.
Front Cell Neurosci. 11:372017.
|
|
295
|
Yamamoto K and Hornykiewicz O: Proposal
for a noradrenaline hypothesis of schizophrenia. Prog
Neuropsychopharmacol Biol Psychiatry. 28:913–922. 2004.
|
|
296
|
Newell KA, Zavitsanou K, Jew SK and Huang
XF: Alterations of muscarinic and GABA receptor binding in the
posterior cingulate cortex in schizophrenia. Prog
Neuropsychopharmacol Biol Psychiatry. 31:225–233. 2007.
|
|
297
|
Erskine D, Taylor JP, Bakker G, Brown AJH,
Tasker T and Nathan PJ: Cholinergic muscarinic M(1) and M(4)
receptors as therapeutic targets for cognitive, behavioural, and
psychological symptoms in psychiatric and neurological disorders.
Drug Discov Today. 24:2307–2314. 2019.
|
|
298
|
Krystal JH, Abi-Saab W, Perry E, D'Souza
DC, Liu N, Gueorguieva R, McDougall L, Hunsberger T, Belger A,
Levine L and Breier A: Preliminary evidence of attenuation of the
disruptive effects of the NMDA glutamate receptor antagonist,
ketamine, on working memory by pretreatment with the group II
metabotropic glutamate receptor agonist, LY354740, in healthy human
subjects. Psychopharmacology (Berl). 179:303–309. 2005.
|
|
299
|
Yasuno F, Suhara T, Ichimiya T, Takano A,
Ando T and Okubo Y: Decreased 5-HT1A receptor binding in amygdala
of schizophrenia. Biol Psychiatry. 55:439–444. 2004.
|
|
300
|
Rasmussen H, Frokjaer VG, Hilker RW,
Madsen J, Anhøj S, Oranje B, Pinborg LH, Glenthøj B and Knudsen GM:
Low frontal serotonin 2A receptor binding is a state marker for
schizophrenia? Eur Neuropsychopharmacol. 26:1248–1250. 2016.
|
|
301
|
Nikiforuk A: Serotonergic and cholinergic
strategies as potential targets for the treatment of schizophrenia.
Curr Pharm Des. 22:2093–2116. 2016.
|
|
302
|
Fatemi SH, Folsom TD and Thuras PD:
Deficits in GABA(B) receptor system in schizophrenia and mood
disorders: A postmortem study. Schizophr Res. 128:37–43. 2011.
|
|
303
|
Correll CU: Current treatment options and
emerging agents for schizophrenia. J Clin Psychiatry.
81:MS19053BR3C2020.
|