Sjögren's syndrome (SS) is a rare chronic and
systemic autoimmune disorder, primarily characterized by
lymphocytic infiltrations and autoimmune exocrinopathy and
epithelitis as well as a female predominance (9:1 female-to-male
predisposition ratio primarily in perimenopausal women) (1). The characteristic lymphocytic
infiltration consists of activated T and B lymphocytes that affect
exocrine glands and autoantibody production. SS occurs in 0.5-1% of
the population, and the spectrum of the manifestations of the
disorder may extend from organ-specific to systemic ones, including
xerostomia, dry eyes, arthralgias/arthritis, rash,
keratoconjunctivitis sicca, primary liver cirrhosis, lymphoma and
lung involvement (2,3). The disorder is also characterized
by hypergammaglobulinemia and autoantibody production, mainly
against the ribonucleoproteins SS-A/Ro and SS-B/La characteristic
of SS and identified in the serum of the patients (4). The increased levels of various
cytokines such as interferon (IFN)-γ, interleukin (IL)-6 and IL-10
found in patients with SS have suggested their notable role in the
pathogenesis of the disease (5,6).
SS has a complex pathogenesis, involving interacting genetic,
epigenetic, hormonal and environmental factors. Furthermore,
additional activators of the disease include specific viruses, such
as Epstein-Barr virus, human T-lymphotropic virus, retroviruses and
Coxsackie virus (7-9). No effective treatment for SS has
been developed so far. Current medical treatments focus on the
alleviation of the symptoms of the disease as well as the decrease
of inflammatory events (10).
Endometriosis is an enigmatic, multifactorial
disorder, representing one of the most common gynecological
diseases affecting 3-10% of women in their reproductive years, and
it can be a debilitating disease leading to poor quality of life
(11). Despite the frequent
occurrence, its etiology and pathogenesis are poorly understood and
the exact cause is unknown (12-14). Multiple genetic and epigenetic
factors, in combination with interacting environmental factors,
including pollution agents and toxins, lead to the development of
this condition (15). It is
characterized by ectopic localization of endometrial cells and,
therefore, the occurrence of endometrial tissue outside the uterine
cavity on other organs (16,17). Endometriosis can appear as
ovarian endometriotic cysts, peritoneal lesions, fibrosis and
deeply infiltrative endometriosis (15). It is associated with chronic
pelvic pain, dysmenorrhea, irregular menstrual bleeding, deep
dyspareunia and urinary tract symptoms, while ~30% of patients with
endometriosis also suffer from infertility (18-20). Considering that all endometriosis
cases cannot be explained by a uniform theory, apart from the
favorable and most accepted theory of Sampson (21) based on retrograde menstruation
hypothesis, more pathways and cellular processes have been
considered, including angiogenesis, chronic inflammation, increased
oxidative stress and endothelial dysfunction (22,23). Clinical presentation varies
widely, ranging from asymptomatic to severe, and no diagnostic
biomarkers have been approved for routine clinical diagnosis of
endometriosis (24). Notably,
the type and severity of symptoms depend on the extent of the
disease and the location of the involved organ(s).
Advances in the past years have shown that female
patients with endometriosis are at higher risk of developing
chronic diseases as systemic comorbidities, such as cancer
(25), cardiovascular diseases
(26,27), asthma (25), hypothyroidism (25) and psychiatric disorders (28). Most importantly, epidemiological
studies demonstrated that patients with endometriosis were
associated with an increased risk of developing a number of
autoimmune diseases compared with unaffected controls due to
notable changes in immune-related parameters. The autoimmune
diseases included in this category include rheumatoid arthritis
(RA), multiple sclerosis (MS), systemic lupus erythematosus (SLE),
ulcerative colitis, Crohn's disease, coeliac disease, ankylosing
spondylitis and autoimmune thyroid disorder (29-33). However, it is unclear whether
autoimmune diseases represent a risk factor of endometriosis, or
these two types of diseases share similar pathogenetic
mediators.
Previous and recent epidemiological studies have
suggested that endometriosis can increase the susceptibility to SS
in these females compared with unaffected controls (29,34-36). Although the pathogenesis of SS
has not been elucidated, there has been strong evidence pointing
out the important role of genetics in the development of this
disease. Moreover, current research has demonstrated substantial
deregulation of the immune system of female patients with
endometriosis and epidemiological studies have presented evidence
for a link between endometriosis and an increased risk of
developing SS (29,34-36). The etiology of this co-occurrence
remains poorly defined. It is possible that the immunological
alterations and chronic inflammation characterizing endometriosis
may lead to SS. These similarities between molecular and cellular
pathways of endometriosis and SS may implicate a partially shared
genetic background. Thus, in the current review, an overview of the
shared genetic factors known thus far that are associated with an
increased susceptibility for both disorders are presented, while
the review did not focus on the plausible clinical basis and
relevant aspects regarding the co-occurrence of both diseases.
Endometriosis is a highly complex disease with
numerous genetic, epigenetic and environmental factors interacting
with each other, thus contributing to its pathogenesis (15). The identification and functional
analysis of the numerous genetic factors involved in the
development of endometriosis have notably contributed to the better
understanding of the biological processes and molecular mechanisms
leading to the disease, as presented in detail in recent studies by
our research group (27,30,32). In brief, the strong genetic
predisposition of the disease was documented firstly in monozygotic
twin-based and family studies (37,38), followed by linkage analysis,
various candidate and gene association, genome wide association
studies (GWAS), meta-analyses and next-generation sequencing
studies (39-53). As a consequence, a number of
disease-associated gene polymorphisms have been detected, which are
involved in estrogen-induced cell growth (WNT4 rs7521902),
vascular function (KDR rs17773813), cell adhesion
(VEZT rs10859871, KAZN rs10928050), growth and
migration (FN1 rs1250241), matrix remodeling and
angiogenesis (VEGF rs699947, LAMA5 rs2427284), cell
cycle regulation (FAS rs1341643), transcription (ID4
rs7739264, MLLT10 rs1802669), differentiation (GDAP1
rs554964149), proliferation (MUC4 rs882605), oncogenesis
(TP53 rs1042522, CHD5 rs9434741), inflammation
(COX-2 rs20417), sex steroid hormone activity and metabolism
(GREB1 rs13394619, FSHB rs74485684, SYNE1
rs71575922, CCDC170 rs1971256, ESR1 rs2206949),
immunity (STAT4 rs7574865, IL-1A rs6542095,
IL-10 rs1800871, IL-16 rs4072111) and oxidative
stress (HIF-1α rs11549465) (41,42,46,47,52,53). Furthermore, accumulating evidence
suggests that epigenetic aberrations seem to play an important role
in the pathophysiology of endometriosis (54) and the development of some
specific complications such as pain and infertility (11,55). The levels of DNA methylation,
histone modifications and microRNA (miR/miRNA) expression reflect
the main epigenetic information at the cellular level (56,57). These epigenetic changes may have
potential applications in disease diagnosis, prognosis and
therapeutic interventions (58).
The importance of epigenetic modifications regarding numerous
biological processes has been so far demonstrated according to
findings from aberrant methylation studies as well as
epigenome-wide association studies (EWAS) (59,60).
SS is a complex autoimmune disease with a number of
well-established susceptibility loci. Worldwide studies focused on
the detection of SS-associated alleles of human leukocyte antigen
(HLA) genes, which encode cell surface antigen presenting proteins.
Before the era of GWAS, associations between HLA genes and SS were
detected concerning HLA-Dw3, HLA-B8, HLA-DRw3,
HLA-DR3, DRw52 and HLA-B8 20 (61), while more recent studies focused
on HLA identified DRw53 (62), DR2 (63), HLA-DRB1*01:01,
HLA-B*35:01 (64),
HLA-DQA1*050 (65),
HLA-DRB1*0301 (66),
HLA-Cw7, HLA-DR3 and HLA-DR11 (67). Other genetic studies of SS had
been focused on familial aggregation (68). The first GWAS performed focusing
on SS (69) detected six non-HLA
loci, namely IFN regulatory factor 5 (IRF5)-TNPO3,
signal transducer and activator of transcription-4
(STAT4), IL12A, FAM167A-BLK, CXCR5 and
TNIP1, while previously reported HLA associations (61-67) were confirmed (69). Other GWAS established new
associations with GTF2IRD1-GTF2I and tumour necrosis factor
(TNF)AIP3 (70) as well
as IRF5 (71). Other
studies showed association of SS with TNF-α and IL-10
(72), OAS1 (73) and IKZF1 (74). In the most recent GWAS conducted
by Khatri et al (75) on
patients of European ancestry, 10 novel genome-wide marked
SS-associated loci were identified, including CD247, NAB1,
PTTG1-MIR146A, PRDM1-ATG5, TNFAIP3, X Kell blood group complex
subunit-related family member 6 (XKR6), MAPT-CRHR1,
RPTOR-CHMP6-BAIAP6, tyrosine kinase 2 (TYK2) and
SYNGR1, while a subsequent meta-analysis based on
ImmunoChip-derived data revealed three additional SS-loci,
CD247, PRDM1-ATG5 and TNFAIP3 (75). It is worth noting that the SS
susceptibility loci identified by Khatri et al (75) are implicated in alterations in
immune cell function, inflammatory signaling, and cell stress,
survival and proliferation. Regarding the epigenetic studies in SS,
it was reported that in type I IFN-regulated genes, which were
upregulated in the blood and salivary glands (SGs) of patients with
SS (76,77), DNA hypomethylation was observed
(77). Previous studies assessed
global DNA methylation levels and specific CpG sites in various
candidate genes, such as LTA as well as type I IFN-induced
genes, including STAT1, IFI44L, MX1, IFI44L, PARP9
and IFITM1 (68).
Furthermore, numerous large-scale EWAS managed to identify genetic
regions exhibiting a differential DNA methylation pattern between
patients with SS and healthy controls, including IRF5
promoter regions, TNFSF7 promoter, FOXP3,
KRT19 and DNMT1 genes as well as LINE-1 (68,69).
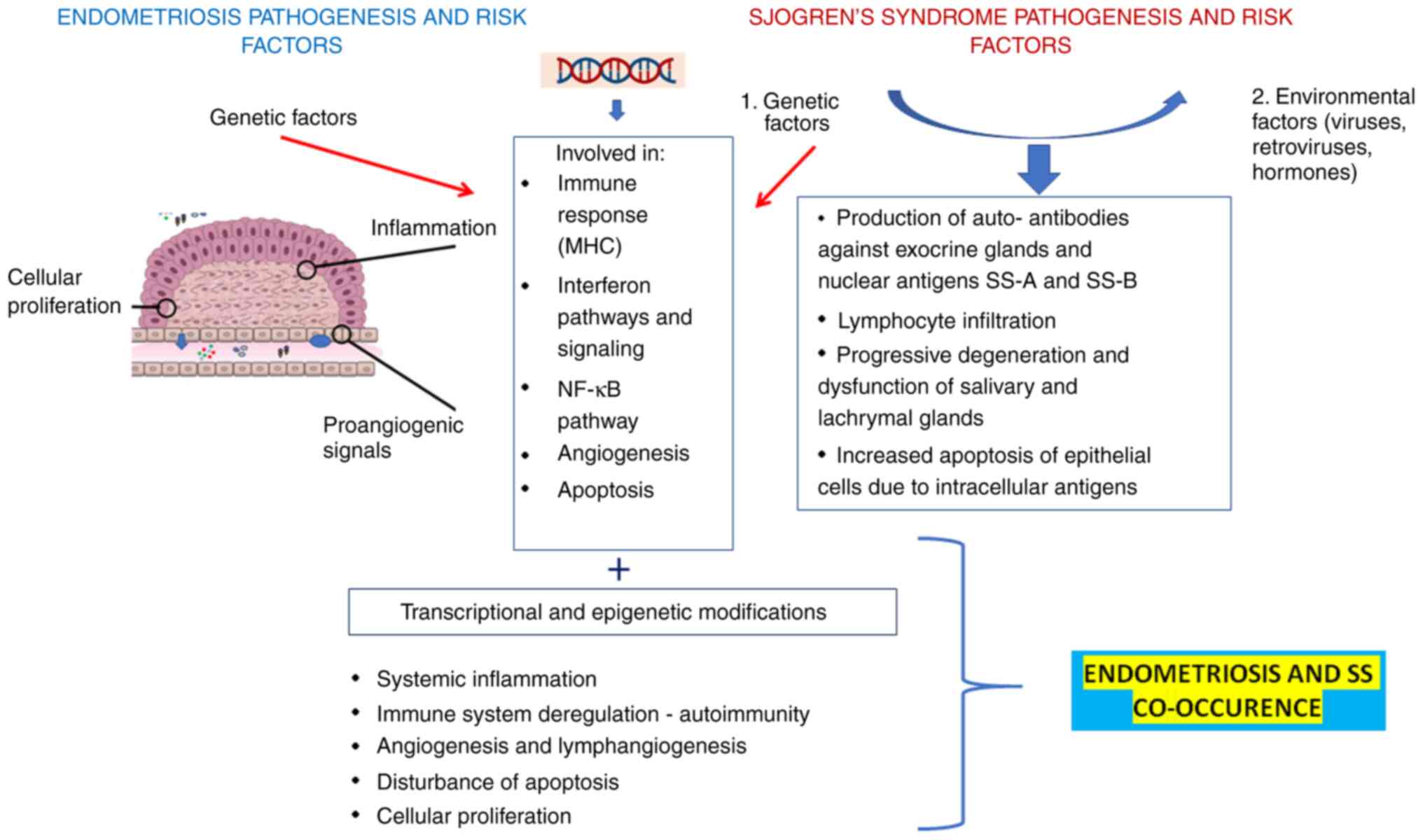
Immune dysregulation leading to chronic inflammatory
response in the ectopic endometrium aggravates a number of
abnormalities in the cell-mediated as well as the humoral immune
systems of patients (30).
Furthermore, other studies suggested the association of
endometriosis with both antibody self-reactivity and chronic local
inflammation (11,23,27,30), caused by a variety of
inflammatory factors such as cytokines, macrophages and
prostaglandins, thus characterizing this disorder as an autoimmune
one (74). In this framework,
extensive research has been conducted to understand the influence
of autoimmunity in endometriosis, aiming to gain better insight of
the pathogenetic mechanisms leading to this condition (30). The identification of antibodies
against endometrial antigens and the subsequent detection of
anti-nuclear and anti-phospholipid antibodies in the blood and the
peritoneal fluid (PF) of patients in combination with elevated
levels of inflammatory cytokines, such as IL-6, IL-8 and TNF-α,
supported pre-existing data indicating a critical role of
autoimmunity in endometriosis (78). This role was further strengthened
by the loss of self-tolerance, leading to immune-mediated tissue
destruction and multi-organ involvement, which represent
immunological alterations also occurring in endometriosis (77). Furthermore, natural killer (NK)
cells were found to be decreased in local NK-mediated cytotoxicity
in the peripheral blood and PF of female patients with
endometriosis (79), with this
decrease being more pronounced in patients with endometriosis at
stages II and IV as demonstrated by a notable reduction in NK
activity (79). It has been
suggested that inhibitory factors developing during the
pathogenesis of endometriosis may suppress NK cell function
(79). Abnormalities regarding
the function and concentration of B- and T-lymphocytes as well as
the total number of macrophages have also been observed in patients
(77,80). Dysregulation of the immune system
may prevent the ability to eliminate the endometrium of the pelvic
cavity, while macrophages and NK cells (78) may be unable to destroy cells in
ectopic sites (77). The
hormonal alterations observed in endometriosis, such as higher
levels of estradiol and progesterone (15,17), have been related to the
inflammatory imbalance that is characteristic of this disorder,
given that inflammation affects hormonal regulation (81). The strong association of
endometriosis with inflammation has also been indicated by the
elevated levels of the inflammatory cytokines IL-6, IL-10 and TNF-α
(82) observed in the PF and
peripheral blood of patients (83) (Fig
1).
As it was aforementioned, SS is a systemic,
inflammatory, autoimmune disease characterized by exocrine
dysfunction due to immunologically-mediated mechanisms (84). It follows the classical,
multistep model of human autoimmune diseases. However, the
immunological mechanisms that mediate the self-directed destruction
of SG tissue are still not well understood. A periductal
mononuclear cell infiltration has been found, leading to the
collection of distinct cellular aggregates in SGs and lachrymal
glands, and the subsequent chronic inflammation with signs in
various organs including lungs, liver and kidneys (85). Moreover, it has been reported
that the observed chronic inflammation is caused by an imbalance of
cytokine production locally in the glands and systemically in the
blood (86). The dysregulation
of cytokines refers to alterations of both local and systemic
expression of pro and anti-inflammatory cytokines released by
infiltrating cells in inflamed tissues (87). A notable research finding refers
to functional differences between the glandular and the peripheral
blood lymphocytes, thus suggesting the existence of a distinct
microenvironment in the gland compared with that in the peripheral
blood, while the putative disruption of the T helper (Th) 1 and 2
cell balance cannot be underestimated (88). Notably, the aforementioned
studies have also suggested a differential role for the action of
Th1 and Th2 cytokines in SS (85,86,88); The Th1 cytokines IL-10, IL-6 and
TGF-β are critical for the induction of SS, but Th2 cytokines, such
as IL-4 and IL-5, may be related to the disease progression
(89). It is noteworthy that
IL-6 functions as a key player regarding B-cell differentiation and
the production of autoantibodies. IL-6 is found at higher levels in
patients with SS compared with healthy controls, thus contributing
to the characteristic inflammatory state of the disorder (90). Moreover, it was recently reported
that the proinflammatory cytokine IL-17, produced mainly by Th17
cells, is upregulated in SS as shown in numerous studies, thus
being involved in the development of SS and, in particular,
associated with the degree of inflammation and clinical
manifestations (91-93). TNF-α and TNF-α-receptors also
appear to be dysregulated in peripheral blood and secreting cells
from SGs from patients with SS (94,95). Mackay and Tangye (96) have emphasized the role of the B
cell activating factor (BAFF), which is a protein member of the TNF
family produced by monocytes, T and dendritic cells (DCs). This
protein is upregulated in SS and is associated with autoantibody
production and B-cell tolerance (97). Numerous pathways that are
strongly associated with IFN signaling were found to be upregulated
in patients with SS, especially in SGs and plasmacytoid DCs
(98). TGF-β, a cytokine crucial
in Th17 polarization (93), is
linked to the growth of regulatory T cells which are upregulated in
inflamed SGs of patients with SS (98). It is noteworthy that accumulating
evidence document that a broad spectrum of impaired immune
functions results in the pathogenesis of SS, including the
cytotoxic cellular cytokines IL-1β, IL-12p40, IL-15 and TNF-α, the
humoral cytokine IL-6, and the factors and death receptors TNF-RI
and TNF-RII (99,100).
Although the etiology of endometriosis is highly
complex and far from being fully elucidated, there is compelling
evidence attributing a critical role in angiogenesis and
lymphangiogenesis, which are involved in both the invasion to the
extracellular matrix and ectopic implantation of endometrial
tissue, and the development of endometriotic lesions (101). Angiogenesis confers to the
maintenance of the endometriotic lesions by supplying them with
functional blood vessels and, as a consequence, leading to the
formation of a dense vascularization (102). The angiogenic properties of the
endometrium and its strong potential to attract blood vessels from
the surrounding tissue is well established (103). Moreover, various potent
angiogenic factors such as TGF-α, TGF-β, basic fibroblast growth
factor and angiopoietin (32)
have been found to be altered in endometriosis, thus suggesting the
involvement of angiogenesis in the ectopic implantation of the
endometrial cells (104). A
number of genetic polymorphisms of key components of the
angiogenesis mechanism, including placental growth factor
rs2268614, hypoxia inducible factor-1α (HIF-1a) rs11549465
and vascular endothelial growth factor (VEGF) receptor 1
(VEGFR1) rs9582036, were found to be associated with the
development of endometriosis. Thus, it has been demonstrated that
all these single nucleotide polymorphisms (SNPs) contribute to the
variability in the plasma levels of the encoded proteins (105,106). In the same framework, data have
shown that higher expression levels of VEGF and angiopoietin-1 led
to dysregulated angiogenic activity of the utopic endometrium in
patients with endometriosis (101,107). It is known that VEGF activates
the migration of various inflammatory cells, including monocytes
and lymphocytes, into the extracellular matrix (108). It is noteworthy that another
study showed that predominantly in adolescent and young adult
female patients with endometriosis, increased levels and marked
activation of circulating proteins related to angiogenesis and cell
migration were observed (109).
The further identification of new factors associated with
angiogenesis may be important for developing novel therapeutic
approaches for endometriosis.
Marked discoveries regarding the molecular
mechanisms leading to SS have highlighted the role of angiogenesis
(110), a fundamental process
in growth and development (111). Previous experiments have shown
that the metalloproteinase TNF-α-converting enzyme, which
participates in proangiogenic pathways leading to the formation of
vessels (112,113), is involved in
VEGF/VEGFR2-mediated angiogenesis in SS (114). Furthermore, it has been shown
that infiltrating T-cells, in combination with human SG epithelial
cells (SGECs), produce increased amounts of proangiogenic factors
through the activation of the VEGF-A/VEGFR-2 system (115). Various findings have shown that
a proangiogenic protein, the metalloproteinase TNF-α-converting
enzyme (TACE) is overproduced in SS, thus emphasizing the role of
the VEGF-A/TACE/VEGFR2/NF-κB axis dysfunction in the pathogenesis
of SS (114). In the same
framework, neuropilin, which represents a transmembrane co-receptor
of the VEGF protein family members, was reported to promote
angiogenesis in SS by activating NF-κB (116). Moreover, quantification and
characterization of circulating angiogenic T (Tang) cells in minor
SGs (MSGs) and in the peripheral blood of patients with SS showed
that this type of cells may participate in endothelial dysfunction
and glandular angiogenesis observed in SS (117). Importantly, Tang cells
infiltrate MSGs and are directly associated with disease activity
in patients with SS (117).
Previous findings have suggested an association
between endometriosis and a higher risk of SS in various
populations, including cases from the USA, Denmark and Taiwan
(29,34-36), an observation that posed a
reasonable question concerning the putative existence of shared
genetic factors that are involved in the co-occurrence of these
diseases. Notably, preexisting data that are presented in detail
below, have demonstrated that various autoimmunity- and
inflammation-associated genes play a crucial role in the
development of both conditions and, therefore, it seems intriguing
to further explore plausible shared mechanisms underlying
endometriosis and SS. Endometriosis may be a risk factor for, or
share a common cause with SS.
Thus, the results of the literature search carried
out as part of the present review showed that the IRF5
rs10488631 (118,119), STAT4 rs7574865 (71,120-122), protein tyrosine phosphatase
non-receptor type 2 (PTPN22) rs2476601 (30,123), TYK2 rs2304256 (75,124), TNF-α rs1800629 (125,126), HIF-1a rs11549465
(64,127), XKR6 rs11250098 SNPs
(75,128) as well as the
HLA-associated alleles DQB1*0301 (65,129,130) and DRB1*01:01 (64,131) are associated with both diseases
(Table I).
STAT4 is a key transcription factor expressed in
activated peripheral blood monocytes, macrophages and DCs in humans
(132), and it is involved in
numerous processes, including transduction of IL-12, IL-23 and type
1 IFN-mediated signals into Th1 and Th17 differentiation, IFN-γ
production and monocyte activation (133). STAT4 is encoded by the
STAT4 gene, which is located at 2q32.2-q32.3, consists of 24
exons and spans a region of 120 kb (121). STAT4 is vital to signaling
pathways in the immune response and autoimmune diseases; the
requirement for a STAT4-dependent cytokine regulation has been well
documented (134). The
STAT4 rs7574865 G/T SNP is a well-known SLE- and
RA-associated polymorphism (134) that was found to be associated
with an increased risk of SS (133) as well as endometriosis
(120). A second STAT4
SNP, rs7582694 (C/G), was reported to be associated with both SS
and endometriosis (121,122).
More specifically, the 'T' allele of the STAT4 rs7574865 SNP
was more common in patients with SS (133). This SNP has been demonstrated
to be weakly associated with the mRNA levels of several IFN-induced
genes when peripheral blood mononuclear cells (PBMCs) from patients
with SS were analyzed (122).
The SNP rs7574865 was shown to be associated with increased
sensitivity to IFN-α signaling in patients with SLE (135). Although the functional role of
rs7574865 SNP in SS is still unclear, it has been assumed that the
rate of transcription is altered due to the resulting nucleotide
change, leading an alteration of the binding of histones in this
genomic area (133). Notably, a
previous bioinformatics analysis showed that this SNP did not
disrupt any activator or transcription factor binding site
(136). Moreover, experiments
performed by the same research group showed a distinct impairment
in STAT4 production as well as in STAT4 phosphorylation in the
presence of the 'T' allele (136). Regarding endometriosis, the
frequency of the TT genotype of the rs7574865 SNP was increased in
female patients with minimal or mild endometriosis compared with
that in controls (120).
Furthermore, it has been suggested that the rs7574865 SNP may
affect either gene expression or mRNA splicing, thus playing an
important role in the regulation of Th17 pathways, due to its
involvement in the induction of Th1 and Th17 cytokine responses and
IFN signaling (120). A
relationship between Th1 response pattern and deep infiltrating
endometriosis has also been suggested (137).
miRNAs are naturally-occurring, small (~22
nucleotides in length), non-coding, post-transcriptional regulatory
molecules, expressed in a tissue-specific and cell-type-specific
manner, targeting numerous mRNAs (196,197). miRNAs represent an evolutionary
system that regulates numerous biological processes such as
embryonic development, cell cycle, migration and proliferation,
differentiation, immune responses as well as apoptosis (198,199), while also being implicated in
the development of various autoimmune diseases including SLE, RA,
MS, AS, SS, experimental autoimmune encephalomyelitis, type 1
diabetes mellitus, inflammatory bowel disease, psoriasis, primary
biliary cirrhosis and idiopathic thrombocytopenic purpura (199-201). To date, various studies have
demonstrated an aberrant expression of miRNAs in affected tissues
or blood serum samples of patients with endometriosis (56,57,202). Moreover, the critical role of
miRNAs has been documented in SS with regards to the
post-transcriptional mRNA expression on PBMCs and SGs (203). Thus, several studies have
investigated the participation and contribution of miRNAs in
endometriosis as well as SS etiopathogenesis (204,205). Notably, various miRNAs have
been reported to have a similar expression profile in both
diseases, including miR16 (203,206-210), miR18a-5p (203,211,212), miR19b-3p (208,213-215), miR26a-5p (208,209,213), miR30c-5p (59,208,209), miR122-3p (214,216), miR142 (217,218), miR146a-5p (217,219,220), miR155-5p (218,221), miR181a (222-225), miR200b (220,226), miR223-3p (203,213) and miR378a-3p (213,227) (Table II).
The present review is a first attempt at searching
the literature and analyzing the genetic and epigenetic factors
that are involved in the co-occurrence of endometriosis and SS,
aiming to shed a light in some shared mechanisms involved in both
conditions while also pointing to the delineation of the relevant
biochemical and molecular pathways. In the present review, the
clinical basis underlying the co-occurrence of both diseases was
not investigated. As presented already in the current review, the
influence of autoimmunity, inflammation, tissue remodeling and
angiogenesis in the underlying biochemical, cellular and
pathophysiological mechanisms leading to the reported association
between endometriosis and SS has been documented. Although it was
beyond the scope of the present review to discuss all these
processes in detail, some aspects regarding their role in the
development of SS and endometriosis were clearly shown in previous
sections. The role of autoimmunity in endometriosis has been
hypothesized and/or established considering that a series of
anti-nuclear, anti-phospholipid and anti-endometrial antibodies are
present in this condition, in combination with elevated levels of
inflammatory cytokines and various immune cell-mediated
abnormalities (74,78). Aiming to unravel the
aforementioned mechanisms, different explanations can be given,
including the putative role of chronic inflammation and immune
dysregulation appearing in endometriosis when it co-occurs with SS.
The development of ectopic endometrial cells and lesions may
provoke an increased immune response, which may be combined with
pathological causes leading to SS. Furthermore, the hypothesis that
endometriosis should be considered an autoimmune disease has been
strengthened by the beneficial effects of danazol and GnRH agonists
as part of endometriosis treatment, likely due to their
immunomodulatory action (259)
as well as the increased number of peritoneal macrophages, high T
and B lymphocyte counts (260)
and the increased levels of circulating anti-endometrial antibodies
(261). However, the putative
mechanisms leading to endometriosis in female patients with SS
remain largely uncertain, given that studies from different
countries examining this relationship have not yet reached a
consensus. Together, the possible role of SS in the etiology of
endometriosis or the occurrence of SS as a secondary response to
endometriosis needs further exploration from a genetic and
biological aspect (36).
It is known that understanding the genetic basis of
complex diseases may provide a unique window into human disease
pathogenesis, which will facilitate the development of improved
diagnostic and therapeutic strategies and enable personalized
medicine. However, apart from the substantial contribution of
various GWAS to the identification of a number of SNPs associated
with an increased risk of endometriosis or SS development, only a
small number of these associations have been analyzed in depth from
a functional aspect, thus minimizing the potential of these SNPs to
be considered as putative therapeutic targets. Therefore, despite
the efforts to analyze the biochemical pathways leading to
endometriosis, it remains an emerging public health problem of
reproductive-age women and the pathogenesis remains elusive. It has
been reported that the prevalence rate of endometriosis in women
with chronic pelvic pain is >33%, and in patients with SS the
prevalence rate is 6.3% (262).
In the same study (262), it
was reported that patients with endometriosis were more likely to
also have SS compared with controls (262). The risk of endometriosis has
been strongly linked to ethnicity but the main differences between
population groups have not been well defined (263). A nine-fold increase in the risk
of developing endometriosis among females from the East Asian
population was found compared with European or American female
populations (263). It is
noteworthy that various studies are currently trying to delineate
how disease risk variation is linked to ethnicity, and to identify
minor differences in SNP variation and differences in autoimmune
disease risk variants reported across different continental
populations (263). The
worldwide range of prevalence of SS is 0.05-4.8% with an overall
9:1 female-to-male ratio, which appears highest in Asian females
(36). However, a limitation of
this type of studies is the lack of information on the patient's
socioeconomic status, personal health behaviors or toxic habits,
with all of them representing confounding factors of the
association between endometriosis and SS (36).
Taking into account that endometriosis has been
associated with various autoimmune diseases, this condition may be
considered a risk factor for SS, which requires specific counseling
and medical management. Immune and inflammatory dysfunctions are
considered challenging therapeutic targets for endometriosis and
SS. The ultimate task of this type of study is the development of
either novel therapeutic alternative by using the in depth
understanding of the role of the associated shared genetic factors
or the use of some miRNAs, a class of agents considered possible
immunomodulators, considering that miRNAs exhibiting a deregulation
in both endometriosis and SS have been identified. Thus,
translation of recent discoveries based on miRNAs may allow the
development of novel therapeutics for endometriosis and SS in the
near future, considering that it has been shown that this type of
drugs provide specificity and reduced toxicity compared with other
therapeutic agents (264,265). The human miRNAome has been
recently analyzed to define a saliva-based diagnostic miRNA
signature for patients with endometriosis, based on their
expression profile, and the results of this study may contribute to
the early diagnosis of this condition (266).
The treatment of complex diseases has undergone
substantial change over the last few years and novel, promising
therapies have been developed. As presented in the current review,
two SNPs of the STAT4 gene, a member of the JAK/STAT pathway
playing a pivotal role in IFN signaling related to immunological
processes promoting chronic inflammation, are associated with both
endometriosis and SS. As a consequence, a promising therapeutic
option may be JAK inhibitors that interrupt the transduction of the
aforementioned JAK/STAT pathway (267,268). Accumulated results suggest that
the JAK inhibitor tofacitinib, a signal transducer associated with
inflammation, can be used as an anti-inflammatory agent in patients
with SS (269). The JAK/STAT
pathway, especially STAT3 phosphorylation, is upregulated in the
utopic endometrium of female patients with endometriosis (270), and it has been reported that
inhibition of JAK/STAT signaling using tofacitinib may be a viable
method for the treatment of endometriosis as well (271). One of the genes that are
activated by STAT3 is HIF1α, which is associated with both
diseases (64,127).
Clinicians should always keep in mind that patients
with endometriosis may also have additional autoimmune diseases,
including SS, while the possible co-occurrence of endometriosis in
patients with SS should not be underestimated. Thus, female
patients with endometriosis, apart from their reference for
follow-up with the gynecologist have to be alerted if they have any
symptoms characterizing SS, such as xerophthalmia, xerostomia,
fatigue, vaginal dryness, myalgia or arthralgia, and report it
immediately to the rheumatologist. It has been reported that female
patients with endometriosis have a higher risk of developing SS
within the first 5 years (36).
Thus, a suitable medication must be provided to these female
patients by clinicians. In this framework, Bardi et al
(281) presented the new
concept of 'holism' for endometriosis, which leads physicians to
evaluate this disorder in a complex and global way, considering its
increased risk to co-occur with various autoimmune diseases. This
combined, global approach is expected to result in beneficial
patient management, taking into account the heterogeneous character
of these diseases. In conclusion, information that can be derived
by analyzing the intersection between autoimmunity, inflammation
and angiogenesis, and the identified shared genetic factors may be
of high value in understanding the underlying biochemical and
cellular mechanisms of the association between endometriosis and
SS, thus contributing to the development of novel therapeutic
alternatives for both disorders.
Not applicable.
MIZ, BCT and GNG designed the current study and
drafted the manuscript. GNG, TBN, BCT, GFG and MIZ searched the
literature. MIZ, GNG and DAS analyzed and organized the data. DAS,
GFG and TBN critically revised the manuscript. All authors have
read and approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
MIZ, BCT, GFG, TBN and GNG (all authors other than
DAS) declare that they have no competing interests. DAS is the
Editor-in-Chief for the journal, but had no personal involvement in
the reviewing process, or any influence in terms of adjudicating on
the final decision, for this article.
Not applicable.
No funding was received.
|
1
|
Mariette X and Criswell LA: Primary
Sjögren's syndrome. N Engl J Med. 378:931–939. 2018.
|
|
2
|
Baimpa E, Dahabreh IJ, Voulgarelis M and
Moutsopoulos HM: Hematologic manifestations and predictors of
lymphoma development in primary Sjögren syndrome: Clinical and
pathophysiologic aspects. Medicine (Baltimore). 88:284–293.
2009.
|
|
3
|
Voulgarelis M, Dafni UG, Isenberg DA and
Moutsopoulos HM: Malignant lymphoma in primary Sjögren's syndrome:
A multicenter, retrospective, clinical study by the European
concerted action on Sjögren's syndrome. Arthritis Rheum.
42:1765–1772. 1999.
|
|
4
|
Nordmark G, Alm GV and Rönnblom L:
Mechanisms of disease: Primary Sjögren's syndrome and the type I
interferon system. Nat Clin Pract Rheumatol. 2:262–269. 2006.
|
|
5
|
Llorente L, Richaud-Patin Y, Fior R,
Alcocer-Varela J, Wijdenes J, Fourrier BM, Galanaud P and Emilie D:
In vivo production of interleukin-10 by non-T cells in rheumatoid
arthritis, Sjögren's syndrome, and systemic lupus erythematosus. A
potential mechanism of B lymphocyte hyperactivity and autoimmunity.
Arthritis Rheum. 37:1647–1655. 1994.
|
|
6
|
Halse A, Tengnér P, Wahren-Herlenius M,
Haga H and Jonsson R: Increased frequency of cells secreting
interleukin-6 and interleukin-10 in peripheral blood of patients
with primary Sjögren's syndrome. Scand J Immunol. 49:533–538.
1999.
|
|
7
|
Fox RI, Pearson G and Vaughan JH:
Detection of Epstein-Barr virus-associated antigens and DNA in
salivary gland biopsies from patients with Sjogren's syndrome. J
Immunol. 137:3162–3168. 1986.
|
|
8
|
Igoe A and Scofield RH: Autoimmunity and
infection in Sjögren's syndrome. Curr Opin Rheumatol. 25:480–487.
2013.
|
|
9
|
Sipsas NV, Gamaletsou MN and Moutsopoulos
HM: Is Sjögren's syndrome a retroviral disease? Arthritis Res Ther.
13:2122011.
|
|
10
|
Bowman SJ: Primary Sjögren's syndrome.
Lupus. 27(Suppl 1): S32–S35. 2018.
|
|
11
|
Bulun SE, Yilmaz BD, Sison C, Miyazaki K,
Bernardi L, Liu S, Kohlmeier A, Yin P, Milad M and Wei J:
Endometriosis. Endocr Rev. 40:1048–1079. 2019.
|
|
12
|
Simpson JL and Bischoff FZ: Heritability
and molecular genetic studies of endometriosis. Ann N Y Acad Sci.
955:239–251. 2002.
|
|
13
|
Stefansson H, Geirsson RT,
Steinthorsdottir V, Jonsson H, Manolescu A, Kong A, Ingadottir G,
Gulcher J and Stefansson K: Genetic factors contribute to the risk
of developing endometriosis. Hum Reprod. 17:555–559. 2002.
|
|
14
|
Montgomery GW, Nyholt DR, Zhao ZZ, Treloar
SA, Painter JN, Missmer SA, Kennedy SH and Zondervan KT: The search
for genes contributing to endometriosis risk. Hum Reprod Update.
14:447–457. 2008.
|
|
15
|
Symons LK, Miller JE, Kay VR, Marks RM,
Liblik K, Koti M and Tayade C: The immunopathophysiology of
endometriosis. Trends Mol Med. 24:748–762. 2018.
|
|
16
|
Berkley KJ, Rapkin AJ and Papka RE: The
pains of endometriosis. Science. 308:1587–1589. 2005.
|
|
17
|
Parente Barbosa C, Bentes De Souza AM,
Bianco B and Christofolini DM: The effect of hormones on
endometriosis development. Minerva Ginecol. 63:375–386. 2011.
|
|
18
|
Simpson JL, Elias S, Malinak LR and
Buttram VC Jr: Heritable aspects of endometriosis. I. Genetic
studies. Am J Obstet Gynecol. 137:327–331. 1980.
|
|
19
|
De Ziegler D, Borghese B and Chapron C:
Endometriosis and infertility: Pathophysiology and management.
Lancet. 376:730–738. 2010.
|
|
20
|
Maddern J, Grundy L, Castro J and Brierley
SM: Pain in endometriosis. Front cell neurosci. 14:5908232020.
|
|
21
|
Sampson JA: Peritoneal endometriosis due
to menstrual dissemination of endometrial tissue into the
peritoneal cavity. Am J Obstet Gynecol. 14:422–469. 1927.
|
|
22
|
Vercellini P, Frontino G, Pietropaolo G,
Gattei U, Daguati R and Crosignani PG: Deep endometriosis:
Definition, pathogenesis, and clinical management. J Am Assoc
Gynecol Laparosc. 11:153–161. 2004.
|
|
23
|
Yovich JL, Rowlands PK, Lingham S,
Sillender M and Srinivasan S: Pathogenesis of endometriosis: Look
no further than John Sampson. Reprod Biomed Online. 40:7–11.
2020.
|
|
24
|
Parasar P, Ozcan P and Terry KL:
Endometriosis: Epidemiology, diagnosis and clinical management.
Curr Obstet Gynecol Rep. 6:34–41. 2017.
|
|
25
|
Kvaskoff M, Mu F, Terry KL, Harris HR,
Poole EM, Farland L and Missmer SA: Endometriosis: A high-risk
population for major chronic diseases? Hum Reprod Update.
21:500–516. 2015.
|
|
26
|
Rafi U, Ahmad S, Bokhari SS, Iqbal MA, Zia
A, Khan MA and Roohi N: Association of inflammatory
markers/cytokines with cardiovascular risk manifestation in
patients with endometriosis. Mediators Inflamm.
2021:34255602021.
|
|
27
|
Vazgiourakis VM, Zervou MI, Papageorgiou
L, Chaniotis D, Spandidos DA, Vlachakis D, Eliopoulos E and
Goulielmos GN: Association of endometriosis with cardiovascular
disease: Genetic aspects (review). Int J Mol Med. 51:292023.
|
|
28
|
Surrey ES, Soliman AM, Johnson SJ, Davis
M, Castelli-Haley J and Snabes MC: Risk of developing comorbidities
among women with endometriosis: A retrospective matched cohort
study. J Womens Health (Larchmt). 27:1114–1123. 2018.
|
|
29
|
Shigesi N, Kvaskoff M, Kirtley S, Feng Q,
Fang H, Knight JC, Missmer SA, Rahmioglu N, Zondervan KT and Becker
CM: The association between endometriosis and autoimmune diseases:
A systematic review and meta-analysis. Hum Reprod Update.
25:486–503. 2019.
|
|
30
|
Zervou MI, Vlachakis D, Papageorgiou L,
Eliopoulos E and Goulielmos GN: Increased risk of rheumatoid
arthritis in patients with endometriosis: Genetic aspects.
Rheumatology (Oxford). 61:4252–4262. 2022.
|
|
31
|
Yin Z, Low HY, Chen BS, Huang KS, Zhang Y,
Wang YH, Ye Z and Wei JCC: Risk of ankylosing spondylitis in
patients with endometriosis: A population-based retrospective
cohort study. Front Immunol. 13:8779422022.
|
|
32
|
Zervou MI, Papageorgiou L, Vlachakis D,
Spandidos DA, Eliopoulos E and Goulielmos GN: Genetic factors
involved in the co-occurrence of endometriosis with ankylosing
spondylitis (review). Mol Med Rep. 27:962023.
|
|
33
|
Zizolfi B, Foreste V, Bonavita S, Rubino
V, Ruggiero G, Brescia Morra V, Lanzillo R, Carotenuto A, Boscia F,
Taglialatela M and Guida M: Epidemiological and immune profile
analysis of Italian subjects with endometriosis and multiple
sclerosis. J Clin Med. 12:20432023.
|
|
34
|
Nielsen NM, Jørgensen KT, Pedersen BV,
Rostgaard K and Frisch M: The co-occurrence of endometriosis with
multiple sclerosis, systemic lupus erythematosus and Sjogren
syndrome. Hum Reprod. 26:1555–1559. 2011.
|
|
35
|
Ma L, Li Z, Li W, Ai J and Chen X:
MicroRNA-142-3p suppresses endometriosis by regulating
KLF9-mediated autophagy in vitro and in vivo. RNA Biol.
16:1733–1748. 2019.
|
|
36
|
Chao YH, Liu CH, Pan YA, Yen FS, Chiou JY
and Wei JCC: Association between endometriosis and subsequent risk
of Sjögren's syndrome: A nationwide population-based cohort study.
Front Immunol. 13:8459442022.
|
|
37
|
Treloar SA, O'Connor DT, O'Connor VM and
Martin NG: Genetic influences on endometriosis in an Australian
twin sample. simplesueT@qimr.edu.au. Fertil
Steril. 71:701–710. 1999.
|
|
38
|
Saha R, Pettersson HJ, Svedberg P,
Olovsson M, Bergqvist A, Marions L, Tornvall P and Kuja-Halkola R:
Heritability of endometriosis. Fertil Steril. 104:947–952.
2015.
|
|
39
|
Treloar SA, Wicks J, Nyholt DR, Montgomery
GW, Bahlo M, Smith V, Dawson G, Mackay IJ, Weeks DE, Bennett ST, et
al: Genomewide linkage study in 1,176 affected sister pair families
identifies a significant susceptibility locus for endometriosis on
chromosome 10q26. Am J Hum Genet. 77:365–376. 2005.
|
|
40
|
Falconer H, D'Hooghe T and Fried G:
Endometriosis and genetic polymorphisms. Obstet Gynecol Surv.
62:616–628. 2007.
|
|
41
|
Nyholt DR, Low SK, Anderson CA, Painter
JN, Uno S, Morris AP, MacGregor S, Gordon SD, Henders AK, Martin
NG, et al: Genome-wide association meta-analysis identifies new
endometriosis risk loci. Nat Genet. 44:1355–1359. 2012.
|
|
42
|
Falconer H, Sundqvist J, Xu H, Vodolazkaia
A, Fassbender A, Kyama C, Bokor A and D'Hooghe TM: Analysis of
common variations in tumor-suppressor genes on chr1p36 among
Caucasian women with endometriosis. Gynecol Oncol. 127:398–402.
2012.
|
|
43
|
Kobayashi H, Imanaka S, Nakamura H and
Tsuji A: Understanding the role of epigenomic, genomic and genetic
alterations in the development of endometriosis (Review). Mol Med
Rep. 9:1483–1505. 2014.
|
|
44
|
Rahmioglu N, Nyholt DR, Morris AP, Missmer
SA, Montgomery GW and Zondervan KT: Genetic variants underlying
risk of endometriosis: Insights from meta-analysis of eight
genome-wide association and replication datasets. Hum Reprod
Update. 20:702–716. 2014.
|
|
45
|
Sapkota Y, Fassbender A, Bowdler L, Fung
JN, Peterse D, O D, Montgomery GW, Nyholt DR and D'Hooghe TM:
Independent replication and meta-analysis for endometriosis risk
loci. Twin Res Hum Genet. 18:518–525. 2015.
|
|
46
|
Sapkota Y, Steinthorsdottir V, Morris AP,
Fassbender A, Rahmioglu N, De Vivo I, Buring JE, Zhang F, Edwards
TL, Jones S, et al: Meta-analysis identifies five novel loci
associated with endometriosis highlighting key genes involved in
hormone metabolism. Nat Commun. 8:155392017.
|
|
47
|
Zondervan KT, Becker CM, Koga K, Missmer
SA, Taylor RN and Viganò P: Endometriosis. Nat Rev Dis Primers.
4:92018.
|
|
48
|
Matalliotakis M, Zervou MI, Matalliotaki
C, Rahmioglu N, Koumantakis G, Kalogiannidis I, Prapas I, Zondervan
K, Spandidos DA, Matalliotakis I and Goulielmos GN: The role of
gene polymorphisms in endometriosis. Mol Med Rep. 16:5881–5886.
2017.
|
|
49
|
Matalliotakis M, Zervou MI, Eliopoulos E,
Matalliotaki C, Rahmioglu N, Kalogiannidis I, Zondervan K,
Spandidos DA, Matalliotakis I and Goulielmos GN: The role of IL-16
gene polymorphisms in endometriosis. Int J Mol Med. 41:1469–1476.
2018.
|
|
50
|
Uimari O, Rahmioglu N, Nyholt DR, Vincent
K, Missmer SA, Becker C, Morris AP, Montgomery GW and Zondervan KT:
Genome-wide genetic analyses highlight mitogen-activated protein
kinase (MAPK) signaling in the pathogenesis of endometriosis. Hum
Reprod. 32:780–793. 2017.
|
|
51
|
Albertsen HM, Matalliotaki C,
Matalliotakis M, Zervou MI, Matalliotakis I, Spandidos DA, Chettier
R, Ward K and Goulielmos GN: Whole exome sequencing identifies
hemizygous deletions in the UGT2B28 and USP17L2 genes in a
three-generation family with endometriosis. Mol Med Rep.
19:1716–1720. 2019.
|
|
52
|
Vassilopoulou L, Matalliotakis M, Zervou
MI, Matalliotaki C, Krithinakis K, Matalliotakis I, Spandidos DA
and Goulielmos GN: Defining the genetic profile of endometriosis.
Exp Ther Med. 17:3267–3281. 2019.
|
|
53
|
Rahmioglu N, Mortlock S, Ghiasi M, Møller
PL, Stefansdottir L, Galarneau G, Turman C, Danning R, Law MH,
Sapkota Y, et al: The genetic basis of endometriosis and
comorbidity with other pain and inflammatory conditions. Nat Genet.
55:423–436. 2023.
|
|
54
|
Goulielmos GN, Matalliotakis M,
Matalliotaki C, Eliopoulos E, Matalliotakis I and Zervou MI:
Endometriosis research in the-omics era. Gene. 741:1445452020.
|
|
55
|
Zelenko Z, Aghajanova L, Irwin JC and
Giudice LC: Nuclear receptor, coregulator signaling, and chromatin
remodeling pathways suggest involvement of the epigenome in the
steroid hormone response of endometrium and abnormalities in
endometriosis. Reprod Sci. 19:152–162. 2012.
|
|
56
|
Tan M, Luo H, Lee S, Jin F, Yang JS,
Montellier E, Buchou T, Cheng Z, Rousseaux S, Rajagopal N, et al:
Identification of 67 histone marks and histone lysine crotonylation
as a new type of histone modification. Cell. 146:1016–1028.
2011.
|
|
57
|
Koukoura O, Sifakis S and Spandidos DA:
DNA methylation in endometriosis (Review). Mol Med Rep.
13:939–2948. 2016.
|
|
58
|
Hsiao KY, Wu MH and Tsai SJ: Epigenetic
regulation of the pathological process in endometriosis. Reprod Med
Biol. 16:314–319. 2017.
|
|
59
|
Kim YJ, Yeon Y, Lee WJ, Shin YU, Cho H,
Sung YK, Kim DR, Lim HW and Kang MH: Comparison of MicroRNA
expression in tears of normal subjects and Sjögren syndrome
patients. Investig Ophthalmol Vis Sci. 60:4889–4895. 2019.
|
|
60
|
Horvath S: DNA methylation age of human
tissues and cell types. Genome Biol. 14:R1152013.
|
|
61
|
Teos LY and Alevizos I: Genetics of
Sjögren's syndrome. Clin Immunol. 182:41–47. 2017.
|
|
62
|
Moriuchi J, Ichikawa Y, Takaya M, Shimizu
H, Uchiyama M, Sato K, Tsuji K and Arimori S: Association between
HLA and Sjögren's syndrome in Japanese patients. Arthritis Rheum.
29:1518–1521. 1986.
|
|
63
|
Wang J, Jiang M and Qiu C: Study on the
relationship between primary Sjögren syndrome and HLA-DRbeta gene.
Zhonghua Nei Ke Za Zhi. 36:398–401. 1997.In Chinese.
|
|
64
|
Hernández-Molina G, Vargas-Alarcón G,
Rodríguez-Pérez JM, Martínez-Rodríguez N, Lima G and
Sánchez-Guerrero J: High-resolution HLA analysis of primary and
secondary Sjögren's syndrome: A common immunogenetic background in
Mexican patients. Rheumatol Int. 35:643–649. 2015.
|
|
65
|
Roitberg-Tambur A, Friedmann A, Safirman
C, Markitziu A, Ben-Chetrit E, Rubinow A, Moutsopoulos HM,
Stavropoulos E, Skopouli FN, Margalit H, et al: Molecular analysis
of HLA class II genes in primary Sjögren's syndrome. A study of
Israeli Jewish and Greek non-Jewish patients. Hum Immunol.
36:235–242. 1993.
|
|
66
|
Manoussakis MN, Georgopoulou C, Zintzaras
E, Spyropoulou M, Stavropoulou A, Skopouli FN and Moutsopoulos HM:
Sjögren's syndrome associated with systemic lupus erythematosus:
Clinical and laboratory profiles and comparison with primary
Sjögren's syndrome. Arthritis Rheum. 50:882–891. 2004.
|
|
67
|
García Portales R, Belmonte Lope MA, Camps
García MT, Ocón Sánchez P, Alonso Ortiz A, Guil García M and de
Ramón Garrido E: Immunogenetics of the Sjogren's syndrome in
southern Spain. An Med Interna. 11:56–61. 1994.In Spanish.
|
|
68
|
Imgenberg-Kreuz J, Sandling JK and
Nordmark G: Epigenetic alterations in primary Sjögren's syndrome-an
overview. Clin Immunol. 196:12–20. 2018.
|
|
69
|
Lessard CJ, Li H, Adrianto I, Ice JA,
Rasmussen A, Grundahl KM, Kelly JA, Dozmorov MG, Miceli-Richard C,
Bowman S, et al: Variants at multiple loci implicated in both
innate and adaptive immune responses are associated with Sjögren's
syndrome. Nat Genet. 45:1284–1292. 2013.
|
|
70
|
Li Y, Zhang K, Chen H, Sun F, Xu J, Wu Z,
Li P, Zhang L, Du Y, Luan H, et al: A genome-wide association study
in Han Chinese identifies a susceptibility locus for primary
Sjögren's syndrome at 7q11.23. Nat Genet. 45:1361–1365. 2013.
|
|
71
|
Taylor KE, Wong Q, Levine DM, McHugh C,
Laurie C, Doheny K, Lam MY, Baer AN, Challacombe S, Lanfranchi H,
et al: Genome-wide association analysis reveals genetic
heterogeneity of Sjögren's syndrome according to ancestry.
Arthritis Rheumatol. 69:1294–1305. 2017.
|
|
72
|
Bolstad AI and Jonsson R: Genetic aspects
of Sjögren's syndrome. Arthritis Res. 4:353–359. 2002.
|
|
73
|
Li H, Reksten TR, Ice JA, Kelly JA,
Adrianto I, Rasmussen A, Wang S, He B, Grundahl KM, Glenn SB, et
al: Identification of a Sjögren's syndrome susceptibility locus at
OAS1 that influences isoform switching, protein expression, and
responsiveness to type I interferons. PLoS Genet.
13:e10068202017.
|
|
74
|
Eisenberg VH, Zolti M and Soriano D: Is
there an association between autoimmunity and endometriosis?
Autoimmun Rev. 11:806–814. 2012.
|
|
75
|
Khatri B, Tessneer KL, Rasmussen A,
Aghakhanian F, Reksten TR, Adler A, Alevizos I, Anaya JM, Aqrawi
LA, Baecklund E, et al: Genome-wide association study identifies
Sjögren's risk loci with functional implications in immune and
glandular cells. Nat Commun. 13:42872022.
|
|
76
|
Gottenberg JE, Cagnard N, Lucchesi C,
Letourneur F, Mistou S, Lazure T, Jacques S, Ba N, Ittah M,
Lepajolec C, et al: Activation of IFN pathways and plasmacytoid
dendritic cell recruitment in target organs of primary Sjögren's
syndrome. Proc Natl Acad Sci USA. 103:2770–2775. 2006.
|
|
77
|
Wildenberg ME, van Helden-Meeuwsen CG, van
de Merwe JP, Drexhage HA and Versnel MA: Systemic increase in type
I interferon activity in Sjögren's syndrome: A putative role for
plasmacytoid dendritic cells. Eur J Immunol. 38:2024–2033.
2008.
|
|
78
|
Matarese G, De Placido G, Nikas Y and
Alviggi C: Pathogenesis of endometriosis: Natural immunity
dysfunction or autoimmune disease? Trends Mol Med. 9:223–228.
2003.
|
|
79
|
Wilson TJ, Hertzog PJ, Angus D, Munnery L,
Wood EC and Kola I: Decreased natural killer cell activity in
endometriosis patients: Relationship to disease pathogenesis.
Fertil Steril. 62:1086–1088. 1994.
|
|
80
|
Burns KA, Thomas SY, Hamilton KJ, Young
SL, Cook DN and Korach KS: Early endometriosis in females is
directed by immune-mediated estrogen receptor a and IL-6
cross-talk. Endocrinology. 159:103–118. 2018.
|
|
81
|
Graziottin A, Skaper SD and Fusco M: Mast
cells in chronic inflammation, pelvic pain and depression in women.
Gynecol Endocrinol. 30:472–477. 2014.
|
|
82
|
Rana N, Braun DP, House R, Gebel H, Rotman
C and Dmowski WP: Basal and stimulated secretion of cytokines by
peritoneal macrophages in women with endometriosis. Fertil Steril.
65:925–930. 1996.
|
|
83
|
Bedaiwy MA and Falcone T: Peritoneal fluid
environment in endometriosis. Clinicopathological implications.
Minerva Ginecol. 55:333–345. 2003.
|
|
84
|
Nikolov NP and Illei GG: Pathogenesis of
Sjogren's syndrome. Curr Opin Rheumatol. 21:465–470. 2009.
|
|
85
|
Voulgarelis M and Tzioufas AG:
Pathogenetic mechanisms in the initiation and perpetuation of
Sjögren's syndrome. Nat Rev Rheumatol. 6:529–537. 2010.
|
|
86
|
Manukyan G, Ghazaryan K, Ktsoyan Z,
Khachatryan Z, Kelly D, Tatyan M, Agababova M and Aminov R:
Comparative analysis of cytokine profiles in autoinflammatory and
autoimmune conditions. Cytokine. 50:146–151. 2010.
|
|
87
|
Fox PC and Speight PM: Current concepts of
autoimmune exocrinopathy: Immunologic mechanisms in the salivary
pathology of Sjögren's syndrome. Crit Rev Oral Biol Med. 7:144–158.
1996.
|
|
88
|
Merrill JE, Kono DH, Clayton J, Ando DG,
Hinton DR and Hofman FM: Inflammatory leukocytes and cytokines in
the peptide-induced disease of experimental allergic
encephalomyelitis in SJL and B10.PL mice. Proc Natl Acad Sci USA.
89:574–578. 1992.
|
|
89
|
Ohyama Y, Nakamura S, Matsuzaki G,
Shinohara M, Hiroki A, Fujimura T, Yamada A, Itoh K and Nomoto K:
Cytokine messenger RNA expression in the labial salivary glands of
patients with Sjögren's syndrome. Arthritis Rheum. 39:1376–1384.
1996.
|
|
90
|
Sjögren E, Leanderson P, Kristenson M and
Ernerudh J: Interleukin-6 levels in relation to psychosocial
factors: Studies on serum, saliva, and in vitro production by blood
mononuclear cells. Brain Behav Immun. 20:270–278. 2006.
|
|
91
|
Sakai A, Sugawara Y, Kuroishi T, Sasano T
and Sugawara S: Identification of IL-18 and Th17 cells in salivary
glands of patients with Sjögren's syndrome, and amplification of
IL-17-mediated secretion of inflammatory cytokines from salivary
gland cells by IL-18. J Immunol. 181:2898–2906. 2008.
|
|
92
|
Nguyen CQ, Hu MH, Li Y, Stewart C and Peck
AB: Salivary gland tissue expression of interleukin-23 and
interleukin-17 in Sjögren's syndrome: Findings in humans and mice.
Arthritis Rheum. 58:734–743. 2008.
|
|
93
|
Katsifis GE, Rekka S, Moutsopoulos NM,
Pillemer S and Wahl SM: Systemic and local interleukin-17 and
linked cytokines associated with Sjögren's syndrome
immunopathogenesis. Am J Pathol. 175:1167–1177. 2009.
|
|
94
|
Oxholm P, Daniels TE and Bendtzen K:
Cytokine expression in labial salivary glands from patients with
primary Sjögren's syndrome. Autoimmunity. 12:185–191. 1992.
|
|
95
|
Koski H, Janin A, Humphreys-Beher MG,
Sorsa T, Malmström M and Konttinen YT: Tumor necrosis factor-alpha
and receptors for it in labial salivary glands in Sjögren's
syndrome. Clin Exp Rheumatol. 19:131–137. 2001.
|
|
96
|
Mackay F and Tangye SG: The role of the
BAFF/APRIL system in B cell homeostasis and lymphoid cancers. Curr
Opin Pharmacol. 4:347–354. 2004.
|
|
97
|
Carrillo-Ballesteros FJ, Palafox-Sánchez
CA, Franco-Topete RA, Muñoz-Valle JF, Orozco-Barocio G,
Martínez-Bonilla GE, Gómez-López CE, Marín-Rosales M,
López-Villalobos EF, Luquin S, et al: Expression of BAFF and BAFF
receptors in primary Sjögren's syndrome patients with ectopic
germinal center-like structures. Clin Exp Med. 20:615–626.
2020.
|
|
98
|
Mavragani CP: Mechanisms and new
strategies for primary Sjögren's syndrome. Annu Rev Med.
68:331–343. 2017.
|
|
99
|
Hagiwara E, Pando J, Ishigatsubo Y and
Klinman DM: Altered frequency of type 1 cytokine secreting cells in
the peripheral blood of patients with primary Sjögren's syndrome. J
Rheumatol. 25:89–93. 1998.
|
|
100
|
Garcíc-Carrasco M, Font J, Filella X,
Cervera R, Ramos-Casals M, Sisó A, Aymamí A, Ballesta AM and
Ingelmo M: Circulating levels of Th1/Th2 cytokines in patients with
primary Sjögren's syndrome: Correlation with clinical and
immunological features. Clin Exp Rheumatol. 19:411–415. 2001.
|
|
101
|
McLaren J: Vascular endothelial growth
factor and endometriotic angiogenesis. Hum Reprod Update. 6:45–55.
2000.
|
|
102
|
Malhotra N, Karmakar D, Tripathi V, Luthra
K and Kumar S: Correlation of angiogenic cytokines-leptin and IL-8
in stage, type and presentation of endometriosis. Gynecol
Endocrinol. 28:224–227. 2012.
|
|
103
|
Maas JW, Le Noble FA, Dunselman GA, De
Goeij AF, Struyker Boudier HA and Evers Jl: The chick embryo
chorioallantoic membrane as a model to investigate the angiogenic
properties of human endometrium. Gynecol Obstet Invest. 48:108–112.
1999.
|
|
104
|
Griffioen AW and Molema G: Angiogenesis:
Potentials for pharmacologic intervention in the treatment of
cancer, cardiovascular diseases, and chronic inflammation.
Pharmacol Rev. 52:237–268. 2000.
|
|
105
|
Barleon B, Reusch P, Totzke F, Herzog C,
Keck C, Martiny-Baron G and Marmé D: Soluble VEGFR-1 secreted by
endothelial cells and monocytes is present in human serum and
plasma from healthy donors. Angiogenesis. 4:143–154. 2001.
|
|
106
|
Carmeliet P, Moons L, Luttun A, Vincenti
V, Compernolle V, De Mol M, Wu Y, Bono F, Devy L, Beck H, et al:
Synergism between vascular endothelial growth factor and placental
growth factor contributes to angiogenesis and plasma extravasation
in pathological conditions. Nat Med. 7:575–583. 2001.
|
|
107
|
Donnez J, Smoes P, Gillerot S,
Casanas-Roux F and Nisolle M: Vascular endothelial growth factor
(VEGF) in endometriosis. Hum Reprod. 13:1686–1690. 1998.
|
|
108
|
Melder RJ, Koenig GC, Witwer BP,
Safabakhsh N, Munn LL and Jain R: During angiogenesis, vascular
endothelial growth factor and basic fibroblast growth factor
regulate natural killer cell adhesion to tumor endothelium. Nat
Med. 2:992–997. 1996.
|
|
109
|
Kirchhoff D, Kaulfuss S, Fuhrmann U,
Maurer M and Zollner TM: Mast cells in endometriosis: Guilty or
innocent bystanders? Expert Opin Ther Targets. 16:237–241.
2012.
|
|
110
|
Sisto M, Ribatti D and Lisi S: Molecular
mechanisms linking inflammation to autoimmunity in Sjögren's
syndrome: Identification of new targets. Int J Mol Sci.
23:132292022.
|
|
111
|
Ribatti D and Crivellato E: 'Sprouting
angiogenesis', a reappraisal. Dev Biol. 372:157–165. 2012.
|
|
112
|
Rundhaug JE: Matrix metalloproteinases and
angiogenesis. J Cell Mol Med. 9:267–285. 2005.
|
|
113
|
Swendeman S, Mendelson K, Weskamp G,
Horiuchi K, Deutsch U, Scherle P, Hooper A, Rafii S and Blobel CP:
VEGF-A stimulates ADAM17-dependent shedding of VEGFR2 and crosstalk
between VEGFR2 and ERK signaling. Circ Res. 103:916–918. 2008.
|
|
114
|
Sisto M, Lisi S, Lofrumento DD, D'Amore M,
Frassanito MA and Ribatti D: Sjögren's syndrome pathological
neovascularization is regulated by VEGF-A-stimulated TACE-dependent
crosstalk between VEGFR2 and NF-κB. Genes Immun. 13:411–420.
2012.
|
|
115
|
Gerli R, Vaudo G, Bocci EB, Schillaci G,
Alunno A, Luccioli F, Hijazi R, Mannarino E and Shoenfeld Y:
Functional impairment of the arterial wall in primary Sjögren's
syndrome: Combined action of immunologic and inflammatory factors.
Arthritis Care Res (Hoboken). 62:712–718. 2010.
|
|
116
|
Sisto M, Lisi S, Lofrumento DD, D'Amore M
and Ribatti D: Neuropilin-1 is upregulated in Sjögren's syndrome
and contributes to pathological neovascularization. Histochem Cell
Biol. 137:669–677. 2012.
|
|
117
|
Alunno A, Ibba-Manneschi L, Bistoni O,
Cipriani S, Topini F, Gerli R and Manetti M: Angiogenic T cells in
primary Sjögren's syndrome: A double-edged sword? Clin Exp
Rheumatol. 37(Suppl 118): S36–S41. 2019.
|
|
118
|
Bianco B, André GM, Vilarino FL, Peluso C,
Mafra FA, Christofolini DM and Barbosa CP: The possible role of
genetic variants in autoimmune-related genes in the development of
endometriosis. Hum Immunol. 73:306–315. 2012.
|
|
119
|
Nordmark G, Kristjansdottir G, Theander E,
Eriksson P, Brun JG, Wang C, Padyukov L, Truedsson L, Alm G,
Eloranta ML, et al: Additive effects of the major risk alleles of
IRF5 and STAT4 in primary Sjögren's syndrome. Genes Immun.
10:68–76. 2009.
|
|
120
|
Bianco B, Fernandes RFM, Trevisan CM,
Christofolini DM, Sanz-Lomana CM, de Bernabe JV and Barbosa CP:
Influence of STAT4 gene polymorphisms in the pathogenesis of
endometriosis. Ann Hum Genet. 83:249–255. 2019.
|
|
121
|
Zamani MR, Salmaninejad A, Akbari Asbagh
F, Masoud A and Rezaei N: STAT4 single nucleotide gene
polymorphisms and susceptibility to endometriosis-related
infertility. Eur J Obstet Gynecol Reprod Biol. 203:20–24. 2016.
|
|
122
|
Gestermann N, Mekinian A, Comets E,
Loiseau P, Puechal X, Hachulla E, Gottenberg JE, Mariette X and
Miceli-Richard C: STAT4 is a confirmed genetic risk factor for
Sjögren's syndrome and could be involved in type 1 interferon
pathway signaling. Genes Immun. 11:432–438. 2010.
|
|
123
|
Gomes FMCS, Bianco B, Teles JS,
Christofolini DM, de Souza AMB, Guedes AD and Barbosa CP: PTPN22
C1858T polymorphism in women with endometriosis. Am J Reprod
Immunol. 63:227–232. 2010.
|
|
124
|
Veena KV, Manolla ML, Deenadayal M,
Shivaji S and Bhanoori M: The rs2304256, a non-synonymous
polymorphism in tyrosine kinase 2 gene is associated with the risk
of endometriosis. Austin J Obstet Gynecol. 10:12162023.
|
|
125
|
Mier-Cabrera J, Cruz-Orozco O, de la
Jara-Díaz J, Galicia-Castillo O, Buenrostro-Jáuregui M,
Parra-Carriedo A and Hernández-Guerrero C: Polymorphisms of
TNF-alpha (-308), IL-1beta (+3954) and IL1-Ra (VNTR) are associated
to severe stage of endometriosis in Mexican women: A case control
study. BMC Womens Health. 22:3562022.
|
|
126
|
Qin B, Wang J, Liang Y, Yang Z and Zhong
R: The association between TNF-α, IL-10 gene polymorphisms and
primary Sjögren's syndrome: A meta-analysis and systemic review.
PLoS One. 8:e634012013.
|
|
127
|
Vodolazkaia A, Yesilyurt BT, Kyama CM,
Bokor A, Schols D, Huskens D, Meuleman C, Peeraer K, Tomassetti C,
Bossuyt X, et al: Vascular endothelial growth factor pathway in
endometriosis: Genetic variants and plasma biomarkers. Fertil
Steril. 105:988–996. 2016.
|
|
128
|
Adewuyi EO, Mehta D; International
Endogene Consortium (IEC); 23andMe Research Team; Nyholt DR:
Genetic overlap analysis of endometriosis and asthma identifies
shared loci implicating sex hormones and thyroid signalling
pathways. Hum Reprod. 37:366–383. 2022.
|
|
129
|
Ishii K, Takakuwa K, Kashima K, Tamura M
and Tanaka K: Associations between patients with endometriosis and
HLA class II; the analysis of HLA-DQB1 and HLA-DPB1 genotypes. Hum
Reprod. 18:985–989. 2003.
|
|
130
|
Cruz-Tapias P, Rojas-Villarraga A,
Maier-Moore S and Anaya JM: HLA and Sjögren's syndrome
susceptibility. A meta-analysis of worldwide studies. Autoimmun
Rev. 11:281–287. 2012.
|
|
131
|
Kitawaki J, Obayashi H, Kado N, Ishihara
H, Koshiba H, Maruya E, Saji H, Ohta M, Hasegawa G, Nakamura N, et
al: Association of HLA class I and class II alleles with
susceptibility to endometriosis. Hum Immunol. 63:1033–1038.
2002.
|
|
132
|
Frucht DM, Aringer M, Galon J, Danning C,
Brown M, Fan S, Centola M, Wu CY, Yamada N, El Gabalawy H and
O'Shea JJ: Stat4 is expressed in activated peripheral blood
monocytes, dendritic cells, and macrophages at sites of
Th1-mediated inflammation. J Immunol. 164:4659–4664. 2000.
|
|
133
|
Korman BD, Kastner DL, Gregersen PK and
Remmers EF: STAT4: Genetics, mechanisms, and implications for
autoimmunity. Curr Allergy Asthma Rep. 8:398–403. 2008.
|
|
134
|
Remmers EF, Plenge RM, Lee AT, Graham RR,
Hom G, Behrens TW, de Bakker PI, Le JM, Lee HS, Batliwalla F, et
al: STAT4 and the risk of rheumatoid arthritis and systemic lupus
erythematosus. N Engl J Med. 357:977–986. 2007.
|
|
135
|
Kariuki SN, Kirou KA, MacDermott EJ,
Barillas-Arias L, Crow MK and Niewold TB: Cutting edge: Autoimmune
disease risk variant of STAT4 confers increased sensitivity to
IFN-alpha in lupus patients in vivo. J Immunol. 182:34–38.
2009.
|
|
136
|
Kofteridis D, Krasoudaki E, Kavousanaki M,
Zervou MI, Panierakis C, Boumpas DT and Goulielmos GN: STAT4 is not
associated with type 2 diabetes in the genetically homogeneous
population of Crete. Genet Test Mol Biomarkers. 13:281–284.
2009.
|
|
137
|
Podgaec S, Dias Junior JA, Chapron C,
Oliveira RM, Baracat EC and Abrão MS: Th1 and Th2 ummune responses
related to pelvic endometriosis. Rev Assoc Med Bras (1992).
56:92–98. 2010.
|
|
138
|
Ryzhakov G, Eames HL and Udalova IA:
Activation and function of interferon regulatory factor 5. J
Interferon Cytokine Res. 35:71–78. 2015.
|
|
139
|
Taniguchi T, Ogasawara K, Takaoka A and
Tanaka N: IRF family of transcription factors as regulators of host
defense. Annu Rev Immunol. 19:623–655. 2001.
|
|
140
|
Chang Foreman HC, Van Scoy S, Cheng TF and
Reich NC: Activation of interferon regulatory factor 5 by site
specific phosphorylation. PLoS One. 7:e330982012.
|
|
141
|
Krausgruber T, Blazek K, Smallie T,
Alzabin S, Lockstone H, Sahgal N, Hussell T, Feldmann M and Udalova
IA: IRF5 promotes inflammatory macrophage polarization and TH1-TH17
responses. Nat Immunol. 12:231–238. 2011.
|
|
142
|
Bianco B, De Camargo CR, Christofolini DM
and Barbosa CP: Involvement of interferon regulatory factor 5
(IRF5) gene polymorphisms and haplotype in endometriosis-related
infertility. J Endometr Pelvic Pain Disord. 9:188–192. 2017.
|
|
143
|
Miceli-Richard C, Comets E, Loiseau P,
Puechal X, Hachulla E and Mariette X: Association of an IRF5 gene
functional polymorphism with Sjögren's syndrome. Arthritis Rheum.
56:3989–3994. 2007.
|
|
144
|
Niewold TB, Kelly JA, Kariuki SN, Franek
BS, Kumar AA, Kaufman KM, Thomas K, Walker D, Kamp S, Frost JM, et
al: IRF5 haplotypes demonstrate diverse serological associations
which predict serum interferon alpha activity and explain the
majority of the genetic association with systemic lupus
erythematosus. Ann Rheum Dis. 71:463–468. 2012.
|
|
145
|
Wang K, Li M and Hakonarson H: Analysing
biological pathways in genome-wide association studies. Nat Rev
Genet. 11:843–854. 2010.
|
|
146
|
Burbelo PD, Ambatipudi K and Alevizos I:
Genome-wide association studies in Sjögren's syndrome: What do the
genes tell us about disease pathogenesis? Autoimmun Rev.
13:756–761. 2014.
|
|
147
|
Zervou MI, Dorschner JM, Ghodke-Puranik Y,
Boumpas DT, Niewold TB and Goulielmos GN: Association of IRF5
polymorphisms with increased risk for systemic lupus erythematosus
in population of Crete, a southern-eastern European Greek island.
Gene. 610:9–14. 2017.
|
|
148
|
Sankararaman S, Mallick S, Dannemann M,
Prüfer K, Kelso J, Pääbo S, Patterson N and Reich D: The genomic
landscape of Neanderthal ancestry in present-day humans. Nature.
507:354–357. 2014.
|
|
149
|
Wu J, Katrekar A, Honigberg LA, Smith AM,
Conn MT, Tang J, Jeffery D, Mortara K, Sampang J, Williams SR, et
al: Identification of substrates of human protein-tyrosine
phosphatase PTPN22. J Biol Chem. 281:11002–11010. 2006.
|
|
150
|
Cohen S, Dadi H, Shaoul E, Sharfe N and
Roifman CM: Cloning and characterization of a lymphoid-specific,
inducible human protein tyrosine phosphatase, Lyp. Blood.
93:2013–2024. 1999.
|
|
151
|
Gomez LM, Anaya JM, Gonzalez CI,
Pineda-Tamayo R, Otero W, Arango A and Martín J: PTPN22 C1858T
polymorphism in Colombian patients with autoimmune diseases. Genes
Immun. 6:628–631. 2005.
|
|
152
|
Cloutier JF and Veillette A: Cooperative
inhibition of T-cell antigen receptor signaling by a complex
between a kinase and a phosphatase. J Exp Med. 189:111–121.
1999.
|
|
153
|
Lee YH, Rho YH, Choi SJ, Ji JD, Song GG,
Nath SK and Harley JB: The PTPN22 C1858T functional polymorphism
and autoimmune diseases-a meta-analysis. Rheumatology (Oxford).
46:49–56. 2007.
|
|
154
|
Bottini N, Musumeci L, Alonso A, Rahmouni
S, Nika K, Rostamkhani M, MacMurray J, Meloni GF, Lucarelli P,
Pellecchia M, et al: A functional variant of lymphoid tyrosine
phosphatase is associated with type I diabetes. Nat Genet.
36:337–338. 2004.
|
|
155
|
Dmowski WP, Gebel HM and Braun DP: The
role of cell-mediated immunity in pathogenesis of endometriosis.
Acta Obstet Gynecol Scand Suppl. 159:7–14. 1994.
|
|
156
|
Eyre S, Bowes J, Diogo D, Lee A, Barton A,
Martin P, Zhernakova A, Stahl E, Viatte S, McAllister K, et al:
High-density genetic mapping identifies new susceptibility loci for
rheumatoid arthritis. Nat Genet. 44:1336–1340. 2012.
|
|
157
|
Diogo D, Bastarache L, Liao KP, Graham RR,
Fulton RS, Greenberg JD, Eyre S, Bowes J, Cui J, Lee A, et al: TYK2
protein-coding variants protect against rheumatoid arthritis and
autoimmunity, with no evidence of major pleiotropic effects on
non-autoimmune complex traits. PLoS One. 10:e01222712015.
|
|
158
|
Kyogoku C, Morinobu A, Nishimura K,
Sugiyama D, Hashimoto H, Tokano Y, Mimori T, Terao C, Matsuda F,
Kuno T and Kumagai S: Lack of association between tyrosine kinase 2
(TYK2) gene polymorphisms and susceptibility to SLE in a Japanese
population. Mod Rheumatol. 19:401–406. 2009.
|
|
159
|
Tokumasa N, Suto A, Kagami S, Furuta S,
Hirose K, Watanabe N, Saito Y, Shimoda K, Iwamoto I and Nakajima H:
Expression of Tyk2 in dendritic cells is required for IL-12, IL-23,
and IFN-gamma production and the induction of Th1 cell
differentiation. Blood. 110:553–560. 2007.
|
|
160
|
Li Z, Rotival M, Patin E, Michel F and
Pellegrini S: Two common disease-associated TYK2 variants impact
exon splicing and TYK2 dosage. PLoS One. 15:e02252892020.
|
|
161
|
Chegini N, Roberts M and Ripps B:
Differential expression of interleukins (IL)-13 and IL-15 in
ectopic and eutopic endometrium of women with endometriosis and
normal fertile women. Am J Reprod Immunol. 49:75–83. 2003.
|
|
162
|
Wang F, Wang H, Jin D and Zhang Y: Serum
miR-17, IL-4, and IL-6 levels for diagnosis of endometriosis.
Medicine (Baltimore). 97:e108532018.
|
|
163
|
Kelly-Welch AE, Hanson EM, Boothby MR and
Keegan AD: Interleukin-4 and interleukin-13 signaling connections
maps. Science. 300:1527–1528. 2003.
|
|
164
|
Roberts M, Luo X and Chegini N:
Differential regulation of interleukins IL-13 and IL-15 by ovarian
steroids, TNF-alpha and TGF-beta in human endometrial epithelial
and stromal cells. Mol Hum Reprod. 11:751–760. 2005.
|
|
165
|
Karjalainen A, Shoebridge S, Krunic M,
Simonović N, Tebb G, Macho-Maschler S, Strobl B and Müller M: TYK2
in tumor immunosurveillance. Cancers (Basel). 12:1502020.
|
|
166
|
Goronzy JJ, Zettl A and Weyand CM: T cell
receptor repertoire in rheumatoid arthritis. Int Rev Immunol.
17:339–363. 1998.
|
|
167
|
Morling N, Andersen V, Fugger L, Georgsen
J, Halberg P, Oxholm P, Odum N and Svejgaard A: Immunogenetics of
rheumatoid arthritis and primary Sjögren's syndrome: DNA
polymorphism of HLA class II genes. Dis Markers. 9:289–296.
1991.
|
|
168
|
Marsh SG and Bodmer JG: HLA class II
nucleotide sequences, 1991. Hum Immunol. 31:207–227. 1991.
|
|
169
|
Cowan EP, Pierce ML, McFarland HF and
McFarlin DE: HLA-DR and -DQ allelic sequences in multiple sclerosis
patients are identical to those found in the general population.
Hum Immunol. 32:203–210. 1991.
|
|
170
|
Reveille JD, Fischbach M, Mcnearney T,
Friedman AW, Aguilar MB, Lisse J, Fritzler MJ, Ahn C and Arnett FC;
GENISOS Study Group: Systemic sclerosis in 3 US ethnic groups: A
comparison of clinical, sociodemographic, serologic, and
immunogenetic determinants. Semin Arthritis Rheum. 30:332–346.
2001.
|
|
171
|
Hodak E, Lapidoth M, Kohn K, David D,
Brautbar B, Kfir K, Narinski N, Safirman S, Maron M and Klein K:
Mycosis fungoides: HLA class II associations among Ashkenazi and
non-Ashkenazi Jewish patients. Br J Dermatol. 145:974–980.
2001.
|
|
172
|
Semino C, Semino A, Pietra G, Mingari MC,
Barocci S, Venturini PL, Ragni N and Melioli G: Role of major
histocompatibility complex class I expression and natural
killer-like T cells in the genetic control of endometriosis. Fertil
Steril. 64:909–916. 1995.
|
|
173
|
Fox RI, Kang HI, Ando D, Abrams J and Pisa
E: Cytokine mRNA expression in salivary gland biopsies of Sjögren's
syndrome. J Immunol. 152:5532–5539. 1994.
|
|
174
|
Abutorabi R, Baradaran A, Sadat Mostafavi
F, Zarrin Y and Mardanian F: Evaluation of tumor necrosis factor
alpha polymorphism frequencies in endometriosis. Int J Fertil
Steril. 9:329–337. 2015.
|
|
175
|
Harada T, Iwabe T and Terakawa N: Role of
cytokines in endometriosis. Fertil Steril. 76:1–10. 2001.
|
|
176
|
Brennan FM and Feldmann M: Cytokines in
autoimmunity. Curr Opin Immunol. 8:872–877. 1996.
|
|
177
|
Gupta V, Gupta A, Jafar T, Gupta V,
Agrawal S, Srivastava N, Kumar S, Singh AK, Natu SM, Agarwal CG and
Agarwal GG: Association of TNF-α promoter gene G-308A polymorphism
with metabolic syndrome, insulin resistance, serum TNF-α and leptin
levels in Indian adult women. Cytokine. 57:32–36. 2012.
|
|
178
|
Bischoff F and Simpson JL: Genetic basis
of endometriosis. Ann N Y Acad Sci. 1034:284–299. 2004.
|
|
179
|
Cramer DW and Missmer SA: The epidemiology
of endometriosis. Ann NY Acad Sci. 955:11–22. 34–36. 396–406.
2002.
|
|
180
|
Kyama CM, Overbergh L, Debrock S, Valckx
D, Vander Perre S, Meuleman C, Mihalyi A, Mwenda JM, Mathieu C and
D'Hooghe TM: Increased peritoneal and endometrial gene expression
of biologically relevant cytokines and growth factors during the
menstrual phase in women with endometriosis. Fertil Steril.
85:1667–1675. 2006.
|
|
181
|
Bertorello R, Cordone MP, Contini P, Rossi
P, Indiveri F, Puppo F and Cordon G: Increased levels of
interleukin-10 in saliva of Sjögren's syndrome patients.
Correlation with disease activity. Clin Exp Med. 4:148–151.
2004.
|
|
182
|
Jaakkola P, Mole DR, Tian YM, Wilson MI,
Gielbert J, Gaskell SJ, von Kriegsheim A, Hebestreit HF, Mukherji
M, Schofield CJ, et al: Targeting of HIF-alpha to the von
Hippel-Lindau ubiquitylation complex by O2-regulated prolyl
hydroxylation. Science. 292:468–472. 2001.
|
|
183
|
Palazon A, Goldrath AW, Nizet V and Ohnson
RS: HIF transcription factors, inflammation, and immunity.
Immunity. 41:518–528. 2014.
|
|
184
|
Shan B, Gerez J, Haedo M, Fuertes M,
Theodoropoulou M, Buchfelder M, Losa M, Stalla GK, Arzt E and
Renner U: RSUME is implicated in HIF-1-induced VEGF-A production in
pituitary tumour cells. Endocr Relat Cancer. 19:13–27. 2012.
|
|
185
|
Hua S and Dias TH: Hypoxia-inducible
factor (HIF) as a target for novel therapies in rheumatoid
arthritis. Front Pharmacol. 7:1842016.
|
|
186
|
Yang ZC and Liu Y: Hypoxia-inducible
factor-1α and autoimmune lupus, arthritis. Inflammation.
39:1268–1273. 2016.
|
|
187
|
Clifford SC, Astuti D, Hooper L, Maxwell
PH, Ratcliffe PJ and Maher ER: The pVHL-associated SCF ubiquitin
ligase complex: Molecular genetic analysis of elongin B and C, Rbx1
and HIF-1alpha in renal cell carcinoma. Oncogene. 20:5067–5074.
2001.
|
|
188
|
Dang EV, Barbi J, Yang HY, Jinasena D, Yu
H, Zheng Y, Bordman Z, Fu J, Kim Y, Yen HR, et al: Control of
T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell.
146:772–784. 2011.
|
|
189
|
Hu F, Liu H, Xu L, Li Y, Liu X, Shi L, Su
Y, Qiu X, Zhang X, Yang Y, et al: Hypoxia-inducible factor-1α
perpetuates synovial fibroblast interactions with T cells and B
cells in rheumatoid arthritis. Eur J Immunol. 46:742–751. 2016.
|
|
190
|
Nezos A, Gravani F, Tassidou A,
Kapsogeorgou EK, Voulgarelis M, Koutsilieris M, Crow MK and
Mavragani C: Type I and II interferon signatures in Sjogren's
syndrome pathogenesis: Contributions in distinct clinical
phenotypes and Sjogren's related lymphomagenesis. J Autoimmun.
63:47–58. 2015.
|
|
191
|
Tanimoto K, Yoshiga K, Eguchi H, Kaneyasu
M, Ukon K, Kumazaki T, Oue N, Yasui W, Imai K, Nakachi K, et al:
Hypoxia-inducible factor-1alpha polymorphisms associated with
enhanced transactivation capacity, implying clinical significance.
Carcinogenesis. 24:1779–1783. 2003.
|
|
192
|
Collec E, Colin Y, Carbonnet F, Hattab C,
Bertrand O, Cartron JP and Kim CL: Structure and expression of the
mouse homologue of the XK gene. Immunogenetics. 50:16–21. 1999.
|
|
193
|
Joo YB, Lim J, Tsao BP, Nath SK, Kim K and
Bae SC: Genetic variants in systemic lupus erythematosus
susceptibility loci, XKR6 and GLT1D1 are associated with
childhood-onset SLE in a Korean cohort. Sci Rep. 8:99622018.
|
|
194
|
Cordero RY, Cordero JB, Stiemke AB, Datta
LW, Buyske S, Kugathasan S, McGovern DPB, Brant SR and Simpson CL:
Trans-ancestry, Bayesian meta-analysis discovers 20 novel risk loci
for inflammatory bowel disease in an African American, East Asian
and European cohort. Hum Mol Genet. 32:873–882. 2023.
|
|
195
|
Kottyan LC, Davis BP, Sherrill JD, Liu K,
Rochman M, Kaufman K, Weirauch MT, Vaughn S, Lazaro S, Rupert AM,
et al: Genome-wide association analysis of eosinophilic esophagitis
provides insight into the tissue specificity of this allergic
disease. Nat Genet. 46:895–900. 2014.
|
|
196
|
Saj A and Lai EC: Control of microRNA
biogenesis and transcription by cell signaling pathways. Curr Opin
Genet Dev. 21:504–510. 2011.
|
|
197
|
Marí-Alexandre J, Sánchez-Izquierdo D,
Gilabert-Estellés J, Barceló-Molina M, Braza-Boïls A and Sandoval
J: miRNAs regulation and its role as biomarkers in endometriosis.
Int J Mol Sci. 17:932016.
|
|
198
|
Dai R and Ahmed SA: MicroRNA, a new
paradigm for understanding immunoregulation, inflammation, and
autoimmune diseases. Transl Res. 157:163–179. 2011.
|
|
199
|
Garo LP and Murugaiyan G: Contribution of
microRNAs to autoimmune diseases. Cell Mol Life Sci. 73:2041–2051.
2016.
|
|
200
|
Qu Z, Li W and Fu B: MicroRNAs in
autoimmune diseases. Biomed Res Int. 2014:5278952014.
|
|
201
|
Yang Y, Peng L, Ma W, Yi F, Zhang Z, Chen
H, Guo Y, Wang L, Zhao LD, Zheng W, et al: Autoantigen-targeting
microRNAs in Sjögren's syndrome. Clin Rheumatol. 35:911–917.
2016.
|
|
202
|
Monnaka VU, Hernandes C, Heller D and
Podgaec S: Overview of miRNAs for the non-invasive diagnosis of
endometriosis: Evidence, challenges and strategies. A systematic
review. Einstein (Sao Paulo). 19:eRW57042021.In English,
Portuguese.
|
|
203
|
De Benedittis G, Ciccacci C, Latini A,
Novelli L, Novelli G and Borgiani P: Emerging role of microRNAs and
long non-coding RNAs in Sjögren's syndrome. Genes (Basel).
12:9032021.
|
|
204
|
Ghafouri-Fard S, Shoorei H and Taheri M:
Role of non-coding RNAs in the pathogenesis of endometriosis. Front
Oncol. 10:13702020.
|
|
205
|
Reale M, D'Angelo C, Costantini E, Laus M,
Moretti A and Croce A: MicroRNA in Sjögren's syndrome: Their
potential roles in pathogenesis and diagnosis. J Immunol Res.
2018:75101742018.
|
|
206
|
Suryawanshi S, Vlad AM, Lin HM,
Mantia-Smaldone G, Laskey R, Lee M, Lin Y, Donnellan N, Klein-Patel
M, Lee T, et al: Plasma microRNAs as novel biomarkers for
endometriosis and endometriosis-associated ovarian cancer. Clin
Cancer Res. 19:1213–1224. 2013.
|
|
207
|
Cha S, Mona M, Lee KE, Kim DH and Han K:
MicroRNAs in autoimmune Sjögren's syndrome. Genomics Inform.
16:e192018.
|
|
208
|
Abdel-Rasheed M, Nour Eldeen G, Mahmoud M,
ElHefnawi M, Abu-Shahba N, Reda M, Elsetohy K, Nabil M, Elnoury A,
Taha T and Azmy O: MicroRNA expression analysis in endometriotic
serum treated mesenchymal stem cells. EXCLI J. 16:852–867.
2017.
|
|
209
|
Wang Y, Zhang G, Zhang L, Zhao M and Huang
H: Decreased microRNA-181a and -16 expression levels in the labial
salivary glands of Sjögren syndrome patients. Exp Ther Med.
15:426–432. 2018.
|
|
210
|
Alevizos I and Illei GG: MicroRNAs in
Sjögren's syndrome as a prototypic autoimmune disease. Autoimmun
Rev. 9:618–621. 2010.
|
|
211
|
Cosar E, Mamillapalli R, Ersoy GS, Cho S,
Seifer B and Taylor HS: Serum microRNAs as diagnostic markers of
endometriosis: A comprehensive array-based analysis. Fertil Steril.
106:402–409. 2016.
|
|
212
|
Yan T, Shen J, Chen J, Zhao M, Guo H and
Wang Y: Differential expression of miR-17-92 cluster among varying
histological stages of minor salivary gland in patients with
primary Sjögren's syndrome. Clin Exp Rheumatol. 37(Suppl 118):
S49–S54. 2019.
|
|
213
|
Wang-Renault SF, Boudaoud S, Nocturne G,
Roche E, Sigrist N, Daviaud C, Bugge Tinggaard A, Renault V,
Deleuze JF, Mariette X and Tost J: Deregulation of microRNA
expression in purified T and B lymphocytes from patients with
primary Sjögren's syndrome. Ann Rheum Dis. 77:133–140. 2018.
|
|
214
|
Qi H, Liang G, Yu J, Wang X, Liang Y, He
X, Feng T and Zhang J: Genome-wide profiling of miRNA expression
patterns in tubal endometriosis. Reproduction. 157:525–534.
2019.
|
|
215
|
Chen X, Jiang Y and Pan D: miR-30c may
serve a role in endometriosis by targeting plasminogen activator
inhibitor-1. Exp Ther Med. 14:4846–4852. 2017.
|
|
216
|
Maged AM, Deeb WS, El Amir A, Zaki SS, El
Sawah H, Al Mohamady M, Metwally AA and Katta MA: Diagnostic
accuracy of serum miR-122 and miR-199a in women with endometriosis.
Int J Gynecol Obstetr. 141:14–19. 2018.
|
|
217
|
Yang P, Wu Z, Ma C, Pan N, Wang Y and Yan
L: Endometrial miR-543 is downregulated during the implantation
window in women with endometriosis-related infertility. Reprod Sci.
26:900–908. 2019.
|
|
218
|
Cortes-Troncoso J, Jang SI, Perez P,
Hidalgo J, Ikeuchi T, Greenwell-Wild T, Warner BM, Moutsopoulos NM
and Alevizos I: T cell exosome-derived miR-142-3p impairs glandular
cell function in Sjögren's syndrome. JCI Insight.
5:e1334972020.
|
|
219
|
Shi H, Zheng LY, Zhang P and Yu CQ:
miR-146a and miR-155 expression in PBMCs from patients with
Sjögren's syndrome. J Oral Pathol Med. 43:792–797. 2014.
|
|
220
|
Ibáñez-Cabellos JS, Pallardó FV,
García-Giménez JL and Seco-Cervera M: Oxidative stress and
epigenetics: miRNA involvement in rare autoimmune diseases.
Antioxidants (Basel). 12:8002023.
|
|
221
|
Nisenblat V, Sharkey DJ, Wang Z, Evans SF,
Healey M, Ohlsson Teague EMC, Print CG, Robertson SA and Hull ML:
Plasma miRNAs display limited potential as diagnostic tools for
endometriosis. J Clin Endocrinol Metab. 104:1999–2022. 2019.
|
|
222
|
Azmy OM and Elgarf WT: MiRNA-130a, a
potential endometriosis inducing factor. Med Res J. 11:40–47.
2012.
|
|
223
|
Williams AEG, Choi K, Chan AL, Lee YJ,
Reeves WH, Bubb MR, Stewart CM and Cha S: Sjögren's
syndrome-associated microRNAs in CD14(+) monocytes unveils targeted
TGFβ signaling. Arthritis Res Ther. 18:952016.
|
|
224
|
Le Dantec C, Varin MM, Brooks WH, Pers JO,
Youinou P and Renaudineau Y: Epigenetics and Sjögren's syndrome.
Curr Pharm Biotechnol. 13:2046–2053. 2012.
|
|
225
|
Peng L, Ma W, Yi F, Yang YJ, Lin W, Chen
H, Zhang X, Zhang LH, Zhang F and Du Q: MicroRNA profiling in
Chinese patients with primary Sjögren syndrome reveals elevated
miRNA-181a in peripheral blood mononuclear cells. J Rheumatol.
41:2208–2213. 2014.
|
|
226
|
Saare M, Rekker K, Laisk-Podar T, Sõritsa
D, Roost AM, Simm J, Velthut-Meikas A, Samuel K, Metsalu T, Karro
H, et al: High-throughput sequencing approach uncovers the miRNome
of peritoneal endometriotic lesions and adjacent healthy tissues.
PLoS One. 9:e1126302014.
|
|
227
|
Pateisky P, Pils D, Szabo L, Kuessel L,
Husslein H, Schmitz A, Wenzl R and Yotova I: hsa-miRNA-154-5p
expression in plasma of endometriosis patients is a potential
diagnostic marker for the disease. Reprod Biomed Online.
37:449–466. 2018.
|
|
228
|
Yang RQ, Teng H, Xu XH, Liu SY, Wang YH,
Guo FJ and Liu XJ: Microarray analysis of microRNA deregulation and
angiogenesis-related proteins in endometriosis. Genet Mol Res.
15:150278262016.
|
|
229
|
Jia SZ, Yang Y, Lang J, Sun P and Leng J:
Plasma miR-17-5p, miR-20a and miR-22 are down-regulated in women
with endometriosis. Hum Reprod. 28:322–330. 2013.
|
|
230
|
Taguchi A, Yanagisawa K, Tanaka M, Cao K,
Matsuyama Y, Goto H and Takahashi T: Identification of
hypoxia-inducible factor-1 alpha as a novel target for miR-17-92
microRNA cluster. Cancer Res. 68:5540–5545. 2008.
|
|
231
|
Lei Z, Li B, Yang Z, Fang H, Zhang GM,
Feng ZH and Huang B: Regulation of HIF-1alpha and VEGF by miR-20b
tunes tumor cells to adapt to the alteration of oxygen
concentration. PLoS One. 4:e76292009.
|
|
232
|
Gutierrez LS and Gutierrez J:
Thrombospondin 1 in metabolic diseases. Front Endocrinol
(Lausanne). 12:6385362021.
|
|
233
|
Mogilyansky E and Rigoutsos I: The
miR-17/92 cluster: A comprehensive update on its genomics,
genetics, functions and increasingly important and numerous roles
in health and disease. Cell Death Differ. 20:1603–1614. 2013.
|
|
234
|
Baulina NM, Kulakova OG and Favorova OO:
MicroRNAs: The role in autoimmune inflammation. Acta Naturae.
8:21–33. 2016.
|
|
235
|
Xu X, Li Z, Liu J, Yu S and Wei Z:
MicroRNA expression profiling in endometriosis-associated
infertility and its relationship with endometrial receptivity
evaluated by ultrasound. J Xray Sci Technol. 25:523–532. 2017.
|
|
236
|
Infante-Menéndez J, López-Pastor AR,
González-López P, Gómez-Hernández A and Escribano O: The Interplay
between oxidative stress and miRNAs in obesity-associated hepatic
and vascular complications. Antioxidants (Basel). 9:6072020.
|
|
237
|
Qi H, Zhang J, Shang Y, Yuan S and Meng C:
Argon inhibits reactive oxygen species oxidative stress via the
miR-21-mediated PDCD4/PTEN pathway to prevent myocardial
ischemia/reperfusion injury. Bioengineered. 12:5529–5539. 2021.
|
|
238
|
Zhang B, Xu ZW, Wang KH, Lu TC and Du Y:
Complex regulatory network of microRNAs, transcription factors,
gene alterations in adrenocortical cancer. Asian Pacific J Cancer
Prev. 14:2265–2268. 2013.
|
|
239
|
Alsaleh G, François A, Philippe L, Gong
YZ, Bahram S, Cetin S, Pfeffer S, Gottenberg JE, Wachsmann D,
Georgei P and Sibilia J: MiR-30a-3p negatively regulates BAFF
synthesis in systemic sclerosis and rheumatoid arthritis
fibroblasts. PLoS One. 9:e1112662014.
|
|
240
|
Wang WT, Zhao YN, Han BW, Hong SJ and Chen
YQ: Circulating microRNAs identified in a genome-wide serum
microRNA expression analysis as noninvasive biomarkers for
endometriosis. J Clin Endocrinol Metab. 98:281–289. 2013.
|
|
241
|
Nahid MA, Pauley KM, Satoh M and Chan EK:
miR-146a is critical for endotox-ininduced tolerance: Implication
in innate immunity. J Biol Chem. 284:34590–34599. 2009.
|
|
242
|
Nahid MA, Satoh M and Chan EK: Mechanistic
role of microRNA-146a in endotoxin-induced differential
cross-regulation of TLR signaling. J Immunol. 186:1723–1734.
2010.
|
|
243
|
Lu LF, Boldin MP, Chaudhry A, Lin LL,
Taganov KD, Hanada T, Yoshimura A, Baltimore D and Rudensky AY:
Function of miR-146a in controlling Treg cell-mediated regulation
of Th1 responses. Cell. 142:914–929. 2010.
|
|
244
|
Curtale G, Citarella F, Carissimi C,
Goldoni M, Carucci N, Fulci V, Franceschini D, Meloni F, Barnaba V
and Macino G: An emerging player in the adaptive immune response:
microRNA-146a is a modulator of IL-2 expression and
activation-induced cell death in T lymphocytes. Blood. 115:265–273.
2010.
|
|
245
|
Tsitsiou E and Lindsay MA: microRNAs and
the immune response. Curr Opin Pharmacol. 9:514–520. 2009.
|
|
246
|
Zilahi E, Tarr T, Papp G, Griger Z, Sipka
S and Zeher M: Increased microRNA-146a/b, TRAF6 gene and decreased
IRAK1 gene expressions in the peripheral mononuclear cells of
patients with Sjögren's syndrome. Immunol Lett. 141:165–168.
2012.
|
|
247
|
Wang X, Xin S, Wang Y, Ju D, Wu Q, Qiu Y,
Niu X, Liu W, Li J and Ji P: MicroRNA-146a-5p enhances T helper 17
cell differentiation via decreasing a disintegrin and
metalloprotease 17 level in primary sjögren's syndrome.
Bioengineered. 2:310–324. 2021.
|
|
248
|
Gottfried E, Kreutz M and Mackensen A:
Tumor metabolism as modulator of immune response and tumor
progression. Semin Cancer Biol. 22:335–341. 2012.
|
|
249
|
Appel S, Le Hellard S, Bruland O, Brun JG,
Omdal R, Kristjansdottir G, Theander E, Nordmark G, Kvarnström M,
Eriksson P, et al: Potential association of muscarinic receptor 3
gene variants with primary Sjogren's syndrome. Ann Rheum Dis.
70:1327–1329. 2011.
|
|
250
|
Whisnant AW, Bogerd HP, Flores O, Ho P,
Powers JG, Sharova N, Stevenson M, Chen CH and Cullen BR: In-depth
analysis of the interaction of HIV-1 with cellular microRNA
biogenesis and effector mechanisms. mBio. 4:e0001932013.
|
|
251
|
Eggers JC, Martino V, Reinbold R, Schäfer
SD, Kiesel L, Starzinski-Powitz A, Schüring AN, Kemper B, Greve B
and Götte M: microRNA miR-200b affects proliferation, invasiveness
and stemness of endometriotic cells by targeting ZEB1, ZEB2 and
KLF4. Reprod Biomed Online. 32:434–445. 2016.
|
|
252
|
Matsuzaki S and Darcha C: Epithelial to
mesenchymal transition-like and mesenchymal to epithelial
transition-like processes might be involved in the pathogenesis of
pelvic endometriosis. Hum Reprod. 27:712–721. 2012.
|
|
253
|
Proestling K, Birner P, Gamperl S, Nirtl
N, Marton E, Yerlikaya G, Wenzl R, Streubel B and Husslein H:
Enhanced epithelial to mesenchymal transition (EMT) and upregulated
MYC in ectopic lesions contribute independently to endometriosis.
Reprod Biol Endocrinol. 13:752015.
|
|
254
|
Kapsogeorgou EK, Gourzi VC, Manoussakis
MN, Moutsopoulos HM and Tzioufas AG: Cellular microRNAs (miRNAs)
and Sjögren's syndrome: Candidate regulators of autoimmune response
and autoantigen expression. J Autoimmun. 37:129–135. 2011.
|
|
255
|
Jones Buie JN, Goodwin AJ, Cook JA,
Halushka PV and Fan H: The role of miRNAs in cardiovascular disease
risk factors. Atherosclerosis. 254:271–281. 2016.
|
|
256
|
Zeng Y, Zhang X, Kang K, Chen J, Wu Z,
Huang J, Lu W, Chen Y, Zhang J, Wang Z, et al: MicroRNA-223
attenuates hypoxia-induced vascular remodeling by targeting
RhoB/MLC2 in pulmonary arterial smooth muscle cells. Sci Rep.
6:249002016.
|
|
257
|
Fulci V, Scappucci G, Sebastiani GD,
Giannitti C, Franceschini D, Meloni F, Colombo T, Citarella F,
Barnaba V, Minisola G, et al: miR-223 is overexpressed in
T-lymphocytes of patients affected by rheumatoid arthritis. Hum
Immunol. 71:206–211. 2010.
|
|
258
|
Pan Q, Ma J and Guo K: miR-223 enhances
the neuroprotection of estradiol against oxidative stress injury by
inhibiting the FOXO3/TXNIP axis. Neurochem Res. 47:1865–1877.
2022.
|
|
259
|
Seli E and Arici A: Endometriosis:
Interaction of immune and endocrine systems. Semin Reprod Med.
21:135–144. 2003.
|
|
260
|
Badawy SZ, Cuenca V, Stitzel A and Tice D:
Immune rosettes of T and B lymphocytes in infertile women with
endometriosis. J Reprod Med. 32:194–197. 1987.
|
|
261
|
Nothnick WB: Treating endometriosis as an
autoimmune disease. Fertil Steril. 76:223–231. 2001.
|
|
262
|
Ma K: Endometriosis increases the risk for
Sjögren syndrome: A nationwide prospective cohort study. Oral Surg
Oral Med Oral Pathol Oral Radiol. 129:e192–e193. 2020.
|
|
263
|
Papageorgiou L, Andreou A, Zervou M,
Vlachakis D, Goulielmos GN and Eliopoulos E: A global population
genomic analysis shows novel insights into the genetic
characteristics of endometriosis. World Acad Sci J. 5:122023.
|
|
264
|
Shomron N and Levy C: MicroRNA-biogenesis
and pre-mRNA splicing crosstalk. J Biomed Biotechnol.
2009:5946782009.
|
|
265
|
Gregory RI, Yan KP, Amuthan G, Chendrimada
T, Doratotaj B, Cooch N and Shiekhattar R: The Microprocessor
complex mediates the genesis of microRNAs. Nature. 432:235–240.
2004.
|
|
266
|
Bendifallah S, Suisse S, Puchar A, Delbos
L, Poilblanc M, Descamps P, Golfier F, Jornea L, Bouteiller D,
Touboul C, et al: Salivary MicroRNA signature for diagnosis of
endometriosis. J Clin Med. 11:6122022.
|
|
267
|
Sutton S, Clutterbuck A, Harris P, Gent T,
Freeman S, Foster N, Barrett-Jolley R and Mobasheri A: The
contribution of the synovium, synovial derived inflammatory
cytokines and neuropeptides to the pathogenesis of osteoarthritis.
Vet J. 179:10–24. 2009.
|
|
268
|
Gaffen SL, Jain R, Garg AV and Cua DJ: The
IL-23-IL-17 immune axis: From mechanisms to therapeutic testing.
Nat Rev Immunol. 14:585–600. 2014.
|
|
269
|
Barrera MJ, Aguilera S, Castro I, Matus S,
Carvajal P, Molina C, González S, Jara D, Hermoso M and González
MJ: Tofacitinib counteracts IL-6 overexpression induced by
deficient autophagy: Implications in Sjögren's syndrome.
Rheumatology (Oxford). 60:1951–1962. 2021.
|
|
270
|
Kim BG, Yoo JY, Kim TH, Shin JH,
Langenheim JF, Ferguson SD, Fazleabas AT, Young SL, Lessey BA and
Jeong JW: Aberrant activation of signal transducer and activator of
transcription-3 (STAT3) signaling in endometriosis. Hum Reprod.
30:1069–1078. 2015.
|
|
271
|
Kotlyar AM, Mamillapalli R, Flores VA and
Taylor HS: Tofacitinib alters STAT3 signaling and leads to
endometriosis lesion regression. Mol Hum Reprod.
27:gaab0162021.
|
|
272
|
Iwabe T, Harada T, Tsudo T, Nagano Y,
Yoshida S, Tanikawa M and Terakawa N: Tumor necrosis factor-alpha
promotes proliferation of endometriotic stromal cells by inducing
interleukin-8 gene and protein expression. J Clin Endocrinol Metab.
85:824–829. 2000.
|
|
273
|
Xue YH, You LT, Ting HF, Chen YW, Sheng
ZY, Xie YD, Wang YH, Chiou JY and Wei JC: Increased risk of
rheumatoid arthritis among patients with endometriosis: A
nationwide population-based cohort study. Rheumatology (Oxford).
60:3326–3333. 2021.
|
|
274
|
Mohler KM, Torrance DS, Smith CA, Goodwin
RG, Stremler KE, Fung VP, Madani H and Widmer MB: Soluble tumor
necrosis factor (TNF) receptors are effective therapeutic agents in
lethal endotoxemia and function simultaneously as both TNF carriers
and TNF antagonists. J Immunol. 151:1548–1561. 1993.
|
|
275
|
Braun DP, Ding J and Dmowski WP:
Peritoneal fluid-mediated enhancement of eutopic and ectopic
endometrial cell proliferation is dependent on tumor necrosis
factor-alpha in women with endometriosis. Fertil Steril.
78:727–732. 2002.
|
|
276
|
Zandbelt MM, de Wilde P, van Damme P,
Hoyng CB, van de Putte L and van den Hoogen F: Etanercept in the
treatment of patients with primary Sjögren's syndrome: a pilot
study. J Rheumatol. 31:96–101. 2004.
|
|
277
|
Sankar V, Brennan MT, Kok MR, Leakan RA,
Smith JA, Manny J, Baum BJ and Pillemer SR: Etanercept in Sjögren's
syndrome: A twelve-week randomized, double-blind,
placebo-controlled pilot clinical trial. Arthritis Rheum.
50:2240–2245. 2004.
|
|
278
|
Moutsopoulos NM, Katsifis GE, Angelov N,
Leakan RA, Sankar V, Pillemer S and Wahl SM: Lack of efficacy of
etanercept in Sjögren syndrome correlates with failed suppression
of tumour necrosis factor alpha and systemic immune activation. Ann
Rheum Dis. 67:1437–1443. 2008.
|
|
279
|
Mariette X, Ravaud P, Steinfeld S, Baron
G, Goetz J, Hachulla E, Combe B, Puéchal X, Pennec Y, Sauvezie B,
et al: Inefficacy of infliximab in primary Sjögren's syndrome:
Results of the randomized, controlled trial of remicade in primary
Sjögren's syndrome (TRIPSS). Arthritis Rheum. 50:1270–1276.
2004.
|
|
280
|
Falconer H, Mwenda JM, Chai DC, Wagner C,
Song XY, Mihalyi A, Simsa P, Kyama C, Cornillie FJ, Bergqvist A, et
al: Treatment with anti-TNF monoclonal antibody (c5N) reduces the
extent of induced endometriosis in the baboon. Hum Reprod.
21:1856–1862. 2006.
|
|
281
|
Bardi M, Cornolti S, Iliakis A, Scorpiniti
A and Vitali M: Endometriosis therapy: Not only hormones and
surgery-the importance of a holistic approach. Am J Biomed Sci Res.
18:0025102023.
|