Introduction
Lung cancer (LC) remains the leading cause of
cancer-related mortality worldwide (1). Although standard therapeutic
modalities, including surgery, radiotherapy, chemotherapy, targeted
therapy and immunotherapy, have improved, their clinical efficacy
is often compromised by drug resistance, tumor microenvironment
remodeling and immune evasion (2). Recent breakthroughs in
understanding programmed cell death (PCD) pathways have resulted in
the identification of novel therapeutic options. In particular, the
close crosstalk between autophagy and ferroptosis has emerged as a
critical regulatory axis with notable implications for LC treatment
(3).
Autophagy is an evolutionarily conserved homeostatic
process that exerts a context-dependent role in tumorigenesis.
While it can promote tumor cell survival under metabolic stress by
recycling cellular components (4), it may also trigger
tumor-suppressive effects through autophagic cell death (5). Moreover, autophagy actively shapes
the tumor immune microenvironment. For example, the microRNA
(miR)-127-3p/MAPK/AKT/mechanistic target of rapamycin (mTOR)/p70S6K
axis has been shown to simultaneously stimulate tumor cell
autophagy and polarize macrophages toward an M1 phenotype (6).
Ferroptosis, by contrast, is a distinct form of PCD
driven by iron-dependent lipid peroxidation, leading to membrane
damage and cell death (7). It is
tightly regulated by molecules including glutathione peroxidase 4
(GPX4), iron metabolism proteins such as ferritin heavy chain 1
(FTH1), and the cystine/glutamate antiporter system Xc−
(7,8). Preclinical studies have
consistently demonstrated that triggering ferroptosis effectively
suppresses LC growth, underscoring its therapeutic potential
(9,10). Lung adenocarcinoma (LUAD)
exhibits particular susceptibility to ferroptosis modulation and
acquired resistance to this process represents a recognized barrier
to treatment efficacy (11,12).
The interaction between autophagy and ferroptosis is
complex and biologically consequential. Autophagic degradation of
substrates such as ferritin and GPX4 can elevate intracellular iron
and lipid peroxides, thereby eliciting ferroptosis and stimulating
antitumor immunity in non-small cell LC (NSCLC) (13). This crosstalk is especially
pertinent in LC given the characteristic dysregulation of iron
metabolism, oxidative stress responses and autophagic flux in this
disease. Notably, O-GlcNAcylation has been shown to co-regulate
ferritinophagy and mitophagy to fine-tune ferroptosis sensitivity
in LC cells (14).
Despite growing research interest, critical
knowledge gaps persist. First, existing reviews lack a systematic
synthesis of LC-specific crosstalk patterns and subtype
heterogeneity. Second, the roles of emerging post-translational
modifications in mediating this interaction are not fully
integrated into current models. Third, the contribution of this
axis to therapy resistance and its potential as a therapeutic
target requires a more comprehensive analysis. Finally, the
translational relevance, including novel biomarkers and therapeutic
agents, has not been adequately addressed.
Therefore, an improved mechanistic understanding of
the autophagy-ferroptosis interplay is essential for elucidating
the pathogenesis of LC and developing innovative treatment
strategies. The present review aims to address these gaps by
providing an updated, LC-focused summary of this crosstalk,
spanning molecular mechanisms to therapeutic translation, thereby
offering a solid foundation for future research and clinical
development.
Fundamental insights into autophagy and
ferroptosis
Molecular mechanisms and biological
functions of autophagy
Autophagy is a highly organized catabolic process
essential for cellular homeostasis. Its initiation is triggered by
stressors such as nutrient deprivation or oxidative stress, which
activate the unc-51 like kinase 1 (ULK1) complex. ULK1 directly
interacts with lactate dehydrogenase (LDH)A, phosphorylates
serine-196 and elevates intracellular lactate levels. This promotes
lactylation of vacuolar protein sorting 34 (VPS34), which in turn
amplifies autophagic flux and endolysosomal trafficking (15). VPS34, a class III PI3K, produces
phosphatidylinositol-3-phosphate to recruit autophagy-related (ATG)
proteins and initiate autophagosome nucleation (16). This cascade is not merely a
housekeeping pathway but a dynamic stress response. For example, in
KRASG12C-driven LC cells, KRASG12C inhibition
induces ULK1/2-mediated autophagy as a critical survival mechanism,
highlighting its potential as a pharmacological target (17). However, this dependency is likely
not universal and a systematic understanding of how different
oncogenic drivers rewire autophagic networks remains to be
elucidated.
Autophagosome elongation and maturation rely on two
ubiquitin-like conjugation systems. The ATG12-ATG5-ATG16L1 complex
drives membrane expansion (18),
whereas microtubule-associated protein 1 light chain 3 (LC3) is
conjugated to phosphatidylethanolamine, converting LC3-I to
membrane-bound LC3-II, a hallmark of autophagosomes (19). Mature autophagosomes fuse with
lysosomes to form autolysosomes, where engulfed cargo is degraded
and recycled.
Autophagy serves pivotal roles in postembryonic
development, cell differentiation and modulation of the
inflammatory response, and its dysregulation is implicated in
various diseases (20,21). In LC, autophagy exerts
context-dependent effects. For example, ailanthone blocks autophagy
by upregulating growth arrest specific (GAS)5 via inhibition of
up-frameshift protein 1-mediated nonsense-mediated mRNA decay,
thereby inhibiting NSCLC proliferation (22). Conversely, regulator of G protein
signaling 20 facilitates NSCLC proliferation by activating
autophagy through suppressing the PKA-Hippo pathway (23). Furthermore, the long non-coding
(lnc)RNA KCTD21-AS1, modified by METTL14-mediated
N6-methyladenosine methylation, acts as a competing endogenous RNA
for miR-519d-5p to regulate CD47 and TIPRL expression, thereby
modulating macrophage phagocytosis and autophagy in LC cells
(24). In small cell LC (SCLC),
elevated AMBRA1 promotes CDK6 degradation via autophagy and confers
a favorable prognosis in patients (25). These studies collectively argue
against targeting autophagy as a monotherapy; instead, they argue
for a precision medicine approach based on tumor genotype and
autophagic cargo specificity.
Definition, characteristics and
mechanisms of ferroptosis
Ferroptosis is a distinct form of PCD driven by
iron-dependent lipid peroxidation. Its core mechanisms involve
dysregulated iron metabolism, excessive lipid peroxidation and
compromised antioxidant defense (26).
Cellular iron uptake is primarily mediated by the
transferrin receptor. Intracellularly, iron is stored in ferritin
to maintain homeostasis; dysregulation increases the labile iron
pool, which catalyzes the Fenton reaction to generate hydroxyl
radicals that initiate lipid peroxidation (27). This process is central to
ferroptosis: Polyunsaturated fatty acids within cellular membranes
are peroxidized to form lipid hydroperoxides (LOOH). Under normal
conditions, the glutathione (GSH)-GPX4 axis reduces LOOH to
nontoxic alcohols, preserving membrane integrity (28). Depletion of GSH or inactivation
of GPX4 leads to LOOH accumulation, resulting in the buildup of
toxic peroxidation products, consequent membrane damage, and cell
death (29). Other antioxidant
systems, including superoxide dismutase and catalase, provide
additional protection against oxidative stress. However, the
clinical translatability of these in vitro findings remain
equivocal owing to the absence of corroborative human tissue or
cohort data.
In NSCLC, ferroptosis is regulated by diverse
molecular pathways. For example, exosomal ROR1-AS1 from
cancer-associated fibroblasts suppresses ferroptosis by stabilizing
solute carrier family 7 member 11 (SLC7A11) mRNA via insulin-like
growth factor 2 mRNA binding protein 1, thereby enhancing cystine
uptake and GSH synthesis (30).
Similarly, lysosome associated protein transmembrane 4B (LAPTM4B)
blocks ferroptosis by suppressing NEDD4L/ZRANB1-mediated
ubiquitination and degradation of SLC7A11 (31). Conversely, other mechanisms
promote ferroptosis. Combined suppression of STAT3 and HOXC10
induces ferroptosis, effectively suppressing bone metastasis in
KRAS-mutant LC (32). Oncogenic
RIT1 mutations not only activate canonical RAS/MAPK and PI3K/AKT
pathways but also modulate nuclear factor, erythroid 2 like 2
(NRF2) target expression, markedly increasing cellular
susceptibility to ferroptosis inducers (33). Notably, a number of
ferroptosis-regulating pathways intersect with autophagy networks,
forming a complex interactive landscape.
Critical perspectives and unresolved
questions
While the core frameworks of autophagy and
ferroptosis are established, their functional heterogeneity in LC
presents notable dichotomies. The determinants of this duality,
such as upstream signaling pathways or driver mutations, remain
unclear. In ferroptosis, GPX4, typically a suppressor,
paradoxically sensitizes GPX4-overexpressing NSCLC cells to RAS
selective lethal 3 (RSL3)-induced ferroptosis (34). Similarly, SLC7A11 can be
upregulated by LAPTM4B to inhibit ferroptosis or downregulated by
circPOLA2 to promote it (35),
yet the contextual switches governing its function are unknown.
These contradictions highlight critical questions:
How do LC-specific adaptations of autophagy and ferroptosis differ
from those in normal lung epithelium? Can these differences be
therapeutically exploited? Future research should employ
subtype-specific models and single-cell multi-omics to map
molecular signatures, clarify how driver mutations shape pathway
heterogeneity and validate these insights for targeted therapy
development.
Regulatory mechanisms of autophagy and
ferroptosis crosstalk
Negative regulation
Autophagy can act as a context-dependent negative
regulator of ferroptosis, a pathway frequently dysregulated in LC
that fosters tumor cell survival and therapy resistance. By
degrading lipid peroxidation-derived peroxisomes, misfolded
proteins and damaged organelles, autophagy helps maintain redox
homeostasis and suppresses ferroptosis (36,37).
This regulatory interplay involves crosstalk with
other cell death pathways. Autophagy can modulate the expression
and activity of apoptosis-related proteins, thereby indirectly
altering cellular susceptibility to ferroptosis (38). Furthermore, mechanical tension
has been shown to govern iron metabolism via nuclear receptor
coactivator 4 (NCOA4)-FTH1 phase-separation-mediated autophagy,
influencing ferroptosis sensitivity (39).
mTOR serves as a central nutrient and energy sensor
within this network (40). Under
nutrient-replete conditions, activated mTOR suppresses autophagy
and strengthens antioxidant defenses, thereby restraining
ferroptosis. Conversely, stress conditions inhibit mTOR, triggering
autophagy and metabolic reprogramming that sensitize cells to
ferroptosis. In LC, Plin2 drives NSCLC proliferation by inhibiting
autophagy via AKT/mTOR activation (41), whereas circFAM190B propels tumor
progression by suppressing autophagy through the SFN/mTOR/ULK1 axis
(42). The compound
(+)-anthrabenzoxocinone inhibits PI3K/AKT/mTOR signaling, inducing
cell cycle arrest, apoptosis and autophagy, while elevating
reactive oxygen species (ROS) in NSCLC cells (43). However, the specific downstream
effectors of PI3K/AKT/mTOR that mediate autophagy-ferroptosis
crosstalk, along with potential subtype-specific differences,
require further definition.
Mitophagy, the selective autophagic clearance of
mitochondria, serves a key inhibitory role against ferroptosis
(44). During mitophagy,
mitochondria undergo characteristic morphological changes,
including swelling, cristae rupture and vacuolization, which are
distinct from the cristae reduction and fragmentation observed in
ferroptosis (45). Tumor cells
deficient in mitophagy exhibit heightened susceptibility to
ferroptosis inducers due to the loss of this protective mechanism
(46). Molecules such as
fibroblast growth factor 21 (47), BCL2 interacting protein
3/NIP3-like protein X (48) and
acteoside (via the NRF2-mitophagy axis) (49) mitigate ferroptosis by restoring
mitochondrial function or removing damaged mitochondria,
underscoring the protective role of mitophagy against
ferroptosis.
Positive regulation
Under specific conditions, autophagy can promote
ferroptosis, a switch often associated with malignant progression
in LC. Overactivation of ferritinophagy drives rapid ferritin
degradation, releasing free iron that catalyzes lipid peroxidation
and induces ferroptosis (50,51). Additional promoting mechanisms
include Par-4-stimulated, NCOA4-mediated ferritinophagy (52) and autophagic degradation of GSH
reductase, which reduces GSH synthesis and antioxidant capacity,
further promoting ferroptosis (53).
In LC, depletion of ubiquitin-specific peptidase
(USP)13 facilitates the transition from autophagy to ferroptosis in
KRAS-mutant LUAD cells via the NRF2-p62-kelch like ECH associated
protein 1 (KEAP1) axis (54).
Rapamycin, a classical autophagy inducer, can exacerbate
mitochondrial damage and provoke ferroptosis when combined with
multi-walled carbon nanotubes (55). Beclin-1 also induces
autophagy-triggered ferroptosis by upregulating ATG5 (56).
Cytochrome c oxidase subunit 7A1 (COX7A1)
exhibits a distinct dual role: It enhances mitochondrial metabolism
to sensitize NSCLC cells to cysteine deprivation-induced
ferroptosis, while concurrently inhibiting autophagy to block
mitochondrial remodeling, two seemingly opposing effects. Rapamycin
can reverse COX7A1-mediated autophagy blockade, synergistically
enhancing ferroptosis under cysteine deprivation (57). The molecular switch controlling
the opposing functions of COX7A1remains unknown, limiting its
therapeutic exploitation. The core mechanisms of autophagy,
ferroptosis and their crosstalk are summarized in Fig. 1.
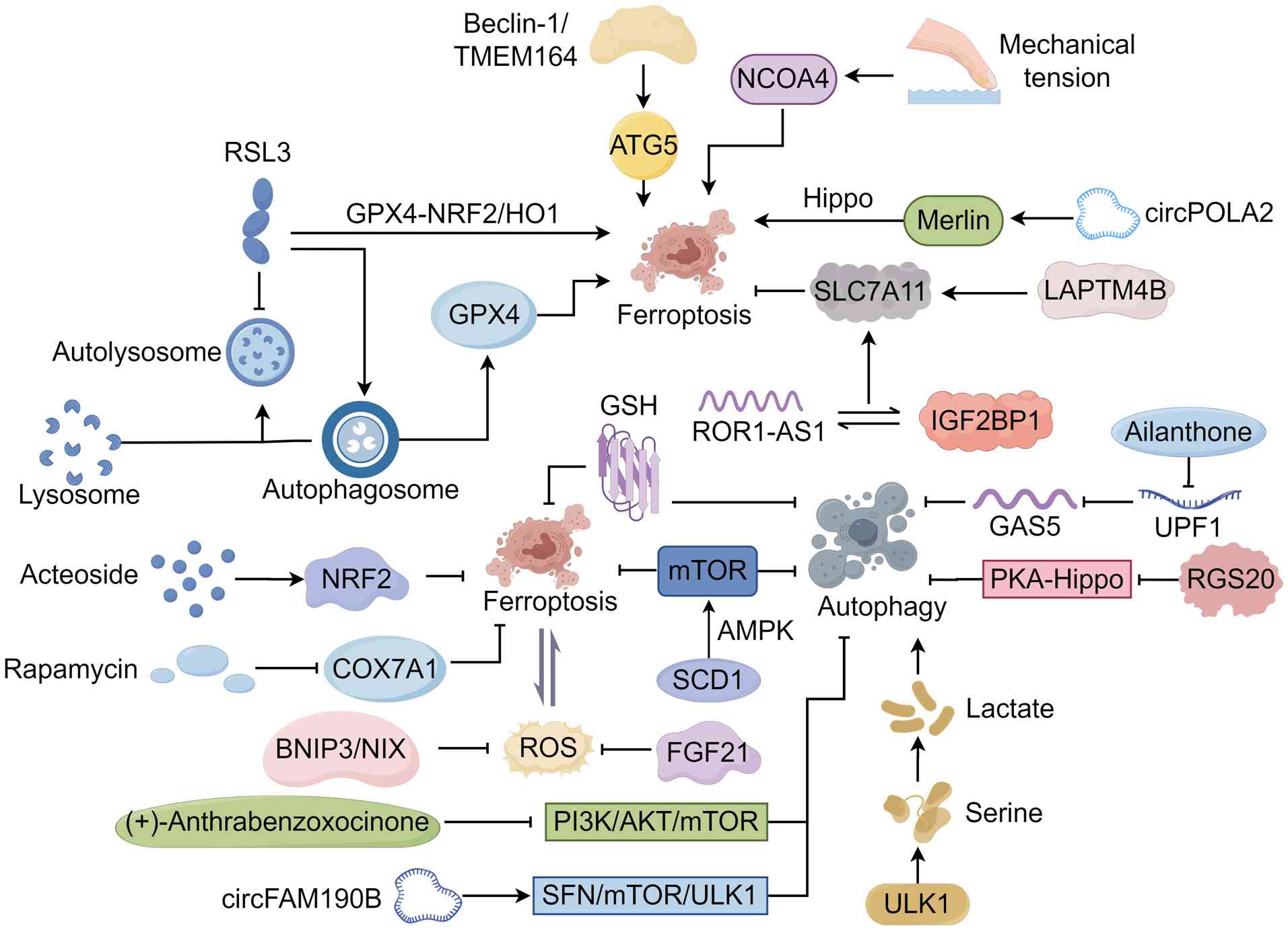 | Figure 1Mechanisms of autophagy and
ferroptosis, and the regulatory mechanisms between them. The figure
was created by Figdraw (www.figdraw.com). RSL3, RAS selective lethal 3; GPX4,
glutathione peroxidase 4; TMEM164, transmembrane protein 164; ATG5,
autophagy related 5; NCOA4, nuclear receptor coactivator 4;
LAPTM4B, lysosome associated protein transmembrane 4B; SLC7A11,
solute carrier family 7 member 11; IGF2BP1, insulin-like growth
factor 2 mRNA binding protein 1; GSH, glutathione; UPF1,
up-frameshift protein 1; GAS5, growth arrest specific 5; RGS20,
regulator of G protein signaling 20; SCD1, stearoyl-CoA desaturase
1; FGF21, fibroblast growth factor 21; ROS, reactive oxygen
species; BNIP3, BCL2 interacting protein 3; NIX, NIP3-like protein
X; COX7A1, cytochrome c oxidase subunit 7A1; NRF2, nuclear
factor, erythroid 2 like 2; ULK1, Unc-51 like kinase 1; mTOR,
mechanistic target of rapamycin. |
Feedback regulation of autophagy by
ferroptosis
Ferroptosis, through its associated oxidative stress
and iron dysregulation, can reciprocally modulate autophagy,
forming a dynamic bidirectional network crucial for cell fate
decisions in the tumor microenvironment.
Under ferroptotic stress, JNK activation leads to
phosphorylation of Beclin-1 at Ser90/93, strengthening its
interaction with the VPS34 complex and stimulating autophagosome
formation (58,59). This constitutes an adaptive
response to ferroptosis-induced damage.
Iron overload enhances p62 phosphorylation, which
strengthens its binding to KEAP1 and leads to NRF2 release and
nuclear translocation. NRF2 then transcriptionally activates ATG
genes, helping to mitigate ROS and maintain homeostasis (60-62). A critical unresolved question is
how NRF2 balances autophagy activation and ferroptosis mitigation
in the LC microenvironment, as this balance may determine tumor
cell fate and therapeutic response.
Ferroptosis-associated oxidative stress also
influences post-translational modifications of ATG proteins,
fine-tuning their activity and localization (63-65). Ubiquitin ligase subunit FBXO9
blocks V-ATPase assembly to impede LC metastasis (66). Additionally, ferroptosis-derived
ROS can activate the AMPK/ULK1 pathway to initiate autophagy
(67), although the precise
molecular cascade in LC cells is not fully elucidated. In summary,
the crosstalk between autophagy and ferroptosis is governed by a
complex, context-dependent network encompassing three core modes:
i) Negative regulation, primarily through mTOR signaling and
mitophagy, to inhibit ferroptosis; ii) positive regulation, driven
by ferritinophagy overactivation, degradation of antioxidant
proteins and regulators such as USP13, to promote ferroptosis; iii)
feedback regulation, involving the JNK/Beclin-1, NRF2/p62 and
ROS/AMPK pathways, creating a bidirectional adaptive loop. A
defining feature is the dual role of autophagy, which can either
suppress or induce ferroptosis depending on the genetic background,
LC subtype and microenvironmental cues. Critical unresolved issues
include identifying the molecular switches that govern the
transition of autophagy from an anti- to a pro-ferroptotic state,
elucidating subtype-specific differences in crosstalk mechanisms,
and clarifying the molecular basis underlying proteins such as
COX7A1 to exhibit opposing functions. Addressing these questions is
essential for deciphering LC cell survival strategies and
developing targeted therapies that exploit this dynamic
interplay.
Dual role of autophagy-ferroptosis crosstalk
in LC
The interplay between autophagy and ferroptosis
exerts a marked yet context-dependent influence on LC progression,
functioning as a precise biological rheostat that can be co-opted
to either promote tumor progression or trigger tumor
suppression.
Tumor-promoting effects
The nutrient-deprived and hypoxic tumor
microenvironment of LC frequently drives the exploitation of
autophagy as a cytoprotective mechanism that counteracts
ferroptosis. This mechanism maintains redox homeostasis, enabling
cancer cell proliferation under hostile microenvironmental stress.
Genetic or pharmacological inhibition of core autophagy machinery
undoes this protective mechanism, markedly sensitizing LC cells to
ferroptosis inducers. For example, in cells expressing an active
Rab37 mutant, knockdown of ATG5/7 not only reduces autophagic flux
but also diminishes tissue inhibitor of metalloproteinase 1
secretion, leading to suppressed lung nodule formation and
metastasis (68). This evidence
underscores the effects of autophagy as a critical adaptive
response, and demonstrates that its disruption unveils a latent
vulnerability to ferroptotic death.
The regulatory interplay between autophagy and
ferroptosis notably influences both the survival and metabolic
processes of LC cells, particularly through metabolic remodeling. A
key adaptation is the enhancement of fatty acid oxidation; a
process associated with aggressive proliferation and poor patient
prognosis (69,70). Dysregulated fatty acid metabolism
fuels proliferation and confers targeted-therapy resistance in
liver tumors; whether an analogous axis operates in LC, and whether
it is further modulated by autophagy and ferroptosis, remains to be
investigated (71). Furthermore,
lipid metabolism enzymes directly interface with this axis. For
example, stearoyl-CoA desaturase 1 suppresses autophagy by tuning
lipid peroxidation and the mTOR pathway, thereby promoting NSCLC
proliferation and metastasis (72). This suggests that metabolic
rewiring via lipid pathways is a non-canonical mechanism by which
the autophagy-ferroptosis balance is tilted toward tumor
promotion.
Beyond cell-intrinsic effects, this crosstalk
actively sculpts an immunosuppressive microenvironment. Autophagy
can induce the secretion of immunosuppressive cytokines and
regulate the expression of immune checkpoints such as PD-L1
(73). It also facilitates
crosstalk with tumor-associated macrophages, particularly the M2
phenotype, to further dampen antitumor immunity. Consequently,
strategic inhibition of autophagy has been shown to restore antigen
presentation and synergize with immunotherapy in preclinical NSCLC
models (74). These findings
suggest that high baseline autophagic activity, particularly when
coupled with elevated checkpoint expression, could serve as a
biomarker for identifying patients who may benefit from combining
autophagy inhibitors with immunotherapy, a hypothesis awaiting
robust clinical validation.
Tumor-suppressive effects
Inducing ferroptotic cell death
Paradoxically, under specific pharmacological or
genetic perturbations, autophagy can function as a tumor suppressor
by actively promoting ferroptosis. Agents such as rapamycin or
curcumin enhance autophagic flux, leading to the degradation of key
anti-ferroptotic proteins such as GPX4; this depletes cellular
antioxidant defenses, resulting in uncontrolled lipid peroxidation
and execution of ferroptosis (57,75). Similarly, non-pharmacological
interventions such as plasma-activated medium therapy can induce LC
cell death via ferroptosis pathways (76). However, the complexity is
highlighted by paradoxical regulators; for example, the autophagy
receptor hippocalcin-like 1 (HPCAL1) promotes ferroptosis yet
paradoxically facilitates NSCLC growth in vivo, suggesting
that its net oncogenic role may involve pleiotropic functions
beyond regulating this cell death pathway (77).
Enhancing antitumor immunity
The tumor-suppressive function extends to
potentiation of antitumor immunity. Autophagy amplifies antigen
processing and presentation, enhancing T-cell activation and
antitumor immunity (78,79). Furthermore, ferroptosis itself is
an immunogenic form of cell death; its induction can release
damage-associated molecular patterns and tumor antigens, thereby
remodeling the tumor immune landscape (80). This synergy is exemplified by
transmembrane protein 164 (TMEM164), which promotes
autophagy-dependent ferroptosis in NSCLC. TMEM164 upregulation
synergizes powerfully with anti-PD-1 therapy in vivo,
demonstrating how engaging this axis can overcome immunotherapy
resistance (13). Complementary
strategies, such as vaccination with autophagosome-enriched
dendritic cells, have also shown efficacy in reducing metastatic
burden (81), further supporting
the immunostimulatory potential of modulating this crosstalk.
In summary, the autophagy-ferroptosis axis exhibits
dual roles in LC biology. It can be hijacked to promote
tumorigenesis via stress adaptation, metabolic reprogramming and
immune evasion. Conversely, it can be therapeutically engaged to
suppress tumors by inducing ferroptotic death and stimulating
antitumor immunity.
Persisting unresolved paradoxes underscore the need
for a more nuanced understanding. First, the same process can
either enhance or subvert antitumor immunity depending on the
cellular and cytokine context. Second, some molecular regulators
exhibit seemingly contradictory pro-tumor functions despite their
ability to induce a tumor-suppressive form of death such as
ferroptosis. Therefore, key questions for future research include:
i) What are the subtype-specific regulatory networks that dictate
the pro- vs. antitumor switch? ii) What are the definitive
molecular determinants and microenvironmental cues that establish
this context-dependence? iii) What is the mechanistic basis of
paradoxical regulators such as HPCAL1? Prioritizing these questions
is essential to move beyond descriptive phenomenology and towards
the rational design of therapeutic strategies that can reliably
steer this dynamic crosstalk toward tumor eradication.
Therapeutic strategies targeting the
autophagy-ferroptosis axis in LC
Repurposing traditional drugs
Traditional pharmacological agents have demonstrated
renewed potential in LC by modulating the autophagy-ferroptosis
axis through diverse and sometimes unexpected mechanisms.
Autophagy inhibitors: Chloroquine (CQ)
and hydroxychloroquine (HCQ)
Antimalarial agents CQ and HCQ are repurposed for LC
by blocking autophagosome-lysosome fusion, thereby blocking
autophagic flux (82). Their
therapeutic efficacy in LC is markedly subtype-dependent, a
critical factor for clinical translation. In NSCLC, HCQ shows
promise as both a chemosensitizer and immunomodulator, potentially
by reducing lysosomal drug sequestration and boosting
CD8+ T-cell responses (83).
Conversely, in a previous study on extensive-stage
SCLC, the addition of HCQ to platinum-based chemotherapy failed to
improve overall survival and even increased toxicity (84), underscoring that HCQ is not a
universal agent and thus necessitates careful patient
stratification. Nanodelivery strategies, such as encapsulation in
human ferritin nanocages, can potentiate the antitumor activity of
HCQ, particularly in cancer with elevated transferrin receptor 1
expression (85). Furthermore,
in LKB1/KRAS co-mutant NSCLC, combining HCQ with the MEK inhibitor
trametinib synergizes to induce ferroptosis. Paradoxically,
autophagy induction under different conditions can confer
trametinib resistance by suppressing ferroptosis (86). This bidirectional effect
highlights the necessity for context-specific modulation of the
autophagy-ferroptosis axis.
Ferroptosis inducers and pathway
modulators
Classic ferroptosis inducers such as erastin can
surmount radioresistance in NSCLC by triggering ferroptosis
(87). The WEE1 G2
checkpoint kinase inhibitor AZD1775 also blocks cystine uptake and
synergizes with SLC7A11 inhibitors to amplify ferroptotic cell
death (88). These findings
validate the therapeutic targeting of the system Xc−
pathway, although challenges related to tumor selectivity and
potential systemic toxicity remain.
Other agents function at the intersection of
multiple cell death pathways. The quinoline derivative DFIQ blocks
autophagic flux, leading to mitochondrial dysfunction and thereby
rendering cells more susceptible to ferroptosis (89). Rabeprazole produces analogous
effects and concurrently upregulates markers of pyroptosis, hinting
at an uncharacterized tripartite crosstalk among autophagy,
ferroptosis and pyroptosis (90). In NSCLC, carbon monoxide (CO)
directly triggers ferroptosis through the ROS/GSK3β/GPX4 axis,
resulting in the accumulation of LOOH (91). These findings establish the
ROS/GSK3β/GPX4 axis as a key driver of CO-induced ferroptosis, but
how this pathway intersects with other cell-death pathways remains
to be clarified.
Metabolic interventions
Reprogramming cellular metabolism presents another
option to adjust ferroptosis sensitivity. Metformin, known
primarily for stimulating autophagy and apoptosis via the
EGFR/AKT/AMPK/mTOR pathway in SCLC (92), may indirectly influence
ferroptosis given the central role of the AMPK/mTOR hub in this
crosstalk, although direct evidence is required.
Targeting lipid metabolism is a promising strategy
for reversing therapy resistance. The co-administration of
ferroptosis inducers and diacylglycerol acyltransferase inhibitors
suppresses tumor growth in LC models resistant to multiple
chemotherapeutic agents (93).
Similarly, modulating homocysteine metabolism through hydrogen
sulfide increases ferroptosis susceptibility in NSCLC, although the
precise mechanistic link requires further definition (94). These findings collectively
validate 'metabolic remodeling' as a viable approach to overcome
resistance. A key future direction involves defining optimal drug
ratios and identifying predictive biomarkers to guide patient
selection for such metabolically targeted combination
therapies.
Novel pharmacological agents
Beyond repurposing traditional drugs, the
development of novel agents specifically designed to target the
autophagy-ferroptosis axis represents a novel option in LC therapy.
These efforts focus on distinct regulatory nodes within this
interactive network.
Targeting autophagy-related pathways
Research into natural compounds has identified
modulators of autophagic signaling as potential antitumor agents in
LC. These agents converge on key regulatory nodes: The AMPK/mTOR
axis and the Sestrin-2/LKB1/AMPK cascade to provoke autophagy and
cell death (95-97). Others, such as corynoxine, target
upstream phosphatases such as protein phosphatase 2A to dually
inhibit AKT signaling (98),
whereas Gboxin and its analog Y9 trigger lysosomal dysfunction and
autophagic impairment (99).
However, a critical gap across these studies is the general lack of
investigation into whether and how these autophagy-modulating
agents affect ferroptosis, which limits the assessment of their
full relevance to the core crosstalk theme. Furthermore, the tumor
selectivity and potential off-target toxicity of agents such as
Gboxin on normal lung epithelium remain unclear, posing challenges
for clinical translation.
Targeting ferroptosis and shared
regulatory nodes
Drug development has also progressed towards core
components of the ferroptosis pathway and its key intersections
with autophagy. Targeting the upstream autophagy initiator ULK1/2
presents a strategic node. The selective ULK1/2 inhibitor,
DCC-3116, synergizes with the KRASG12C inhibitor
sotorasib to suppress KRAS-driven LC, whereas this combination has
been assessed only in early preclinical stages and lacks predictive
biomarkers for patient stratification (17). By contrast, hesperetin inhibits
ferritinophagy and mitigates ferroptosis by modulating the
PI3K/AKT/mTOR/ULK1 pathway (100), further underscoring the
context-dependent role of ULK1.
Inhibition of the antioxidant transcription factor
NRF2 is another validated strategy to enhance ferroptosis
sensitivity. The NRF2 inhibitor, ML385, synergizes with silymarin
(101), and NRF2 knockdown
facilitates S-3'-hydroxy-7',2',4'-trimethoxyisoflavone-mediated
ferritinophagy (102),
revealing a reciprocal regulatory loop between NRF2 and autophagy.
Other agents include sanggenol L, which acts through the
miR-26a-1-3p/MDM2/p53/SLC7A11 axis (103), and curcumin, which activates
autophagy to subsequently trigger ferroptosis (75). Polyoxometalates exhibit
synergistic anti-NSCLC activity with ferroptosis and apoptosis
inducers (104), although the
specific nodes of crosstalk involved remain to be defined.
Optimizing ferroptosis inducers
To address the pharmacokinetic and selectivity
limitations of classic ferroptosis inducers, advanced formulation
strategies are being explored. A folic acid-modified liposomal
nanoformulation co-delivering erastin and the metallothionein 1D
pseudogene markedly augments bioavailability and cytotoxicity in LC
(105). Concurrently,
understanding resistance mechanisms is crucial. Upregulation of the
SLC7A11/HOXB9 axis can mitigate cold atmospheric plasma-induced
ferroptosis while fueling tumor progression (106). Conversely, inducing ferroptosis
sensitizes LC cells to IFN-γ secreted by ROR1-targeted chimeric
antigen receptor (CAR)-T cells, revealing a potent synergy with
immunotherapy (107). A
recurring limitation in both therapeutic development and resistance
studies is the failure to consider the potential contributory role
of autophagy, leaving the integrated network perspective
incomplete.
Gene therapy
Gene editing technologies
The advent of precision gene-editing tools, notably
CRISPR-Cas9, has enabled the direct targeting of specific nodes
within the autophagy-ferroptosis regulatory network. This approach
allows for the functional characterization of key genes and the
exploration of their therapeutic potential. For example,
CRISPR-Cas9-mediated knockout of the anti-apoptotic gene Bcl-2
triggers ferroptosis in LUAD cells by disrupting the
Bax/voltage-dependent anion channel axis (108). Similarly, targeting the central
ferroptosis inhibitor SLC7A11, the expression of which is
upregulated via the MISP/MST/yes-associated protein signaling
pathway, represents a rational strategy to sensitize cells to
ferroptosis (109). Beyond
ferroptosis-specific genes, editing regulators of autophagy can
also influence this crosstalk. Ablation of DEAD-box helicase 24
facilitates autophagy through NF-κB-mediated transcriptional
upregulation of Beclin-1 (110). While these studies provide
compelling proof-of-concept, a notable translational gap exists.
Most findings are derived from in vitro models, underscoring
the urgent need for robust in vivo validation in relevant
preclinical models. Furthermore, future research should prioritize
exploring synergistic combinations of such genetic interventions
with pharmacological modulators of autophagy or ferroptosis to
achieve enhanced therapeutic efficacy.
Gene delivery vectors
Effective gene therapy requires advanced delivery
vectors. While viral vectors offer high efficiency, their
immunogenicity and limited cargo capacity pose notable drawbacks.
By contrast, non-viral vectors, including liposomes and polymeric
nanoparticles, provide improved biocompatibility and lower
immunogenicity, although often at the expense of reduced
transfection efficiency (111).
To overcome these limitations, multifunctional
nanomaterial-based platforms have emerged as promising
alternatives. These systems can simultaneously act as gene carriers
and therapeutic agents. For example, Cu2O@Au nanozymes
deplete intracellular GSH and downregulate SLC7A11, thereby
provoking ferroptosis (112).
Iron-platinum alloy nanoparticles have been shown to reverse
tyrosine kinase inhibitor (TKI) resistance by activating
ferroptosis (113). Other
engineered nanocarriers are designed either to co-activate both
ferroptosis and autophagy (114) or to synergistically combine
chemotherapeutic agents such as paclitaxel with ferroptosis
induction (115).
Despite their promising preclinical performance, the
clinical translation of nanocarriers faces notable hurdles. Key
challenges include unresolved long-term biosafety profiles, the
absence of standardized large-scale manufacturing protocols, and
the insufficient investigation into their potential effects on
autophagy and ferroptosis in normal tissues. Addressing these
issues is therefore paramount to realizing the full potential of
nanomaterial-mediated gene therapy in LC.
Combination treatment strategies
Beyond monotherapies, the strategic modulation of
autophagy and ferroptosis offers a powerful approach to augment the
efficacy of established cancer treatments and overcome resistance
to them. The following sections detail how this axis interacts with
and can be leveraged to enhance chemotherapy, radiotherapy,
targeted therapy and immunotherapy.
With chemotherapy
Chemotherapy efficacy in LC is frequently
constrained by drug resistance, which can be alleviated by tuning
the autophagy-ferroptosis interplay. The role of autophagy is
particularly context-dependent. It can serve as a cytoprotective
mechanism, promoting resistance, as seen in SCLC where upregulation
of lncRNA LYPLAL1-DT enhances autophagy and contributes to
multidrug resistance (116).
Furthermore, suppressing such protective autophagy can be
beneficial, exemplified in NSCLC where silencing staphylococcal
nuclease and tudor domain containing 1 sensitizes cells to various
agents by downregulating PDCD4 (117). By contrast, in a mouse model of
NSCLC, inactivation of the essential autophagy gene ATG5 has been
shown to accelerate the early stages of oncogenesis (118). This divergence underscores the
need for subtype-specific targeting of autophagy.
Concurrently, inducing ferroptosis has proven
effective in surmounting chemoresistance. Suppression of SPTBN2 or
enhancement of HAR1A-mediated MYC degradation increases cisplatin
sensitivity by inhibiting adaptive autophagy or directly triggering
ferroptosis (119,120). Furthermore, targeting
regulators within the crosstalk itself, such as knocking down
deltex E3 ubiquitin ligase 2 to stabilize NCOA4 and promote
ferritinophagy, can mitigate cisplatin resistance in NSCLC
(121). Fig. 2 depicts the mechanisms underlying
the regulation of autophagy and ferroptosis crosstalk in the
context of LC and LC pharmacotherapy. These findings collectively
validate that precise intervention in this axis may markedly
augment chemotherapeutic efficacy, though the successful clinical
translation of such combinations require careful patient
stratification based on molecular and histological subtypes.
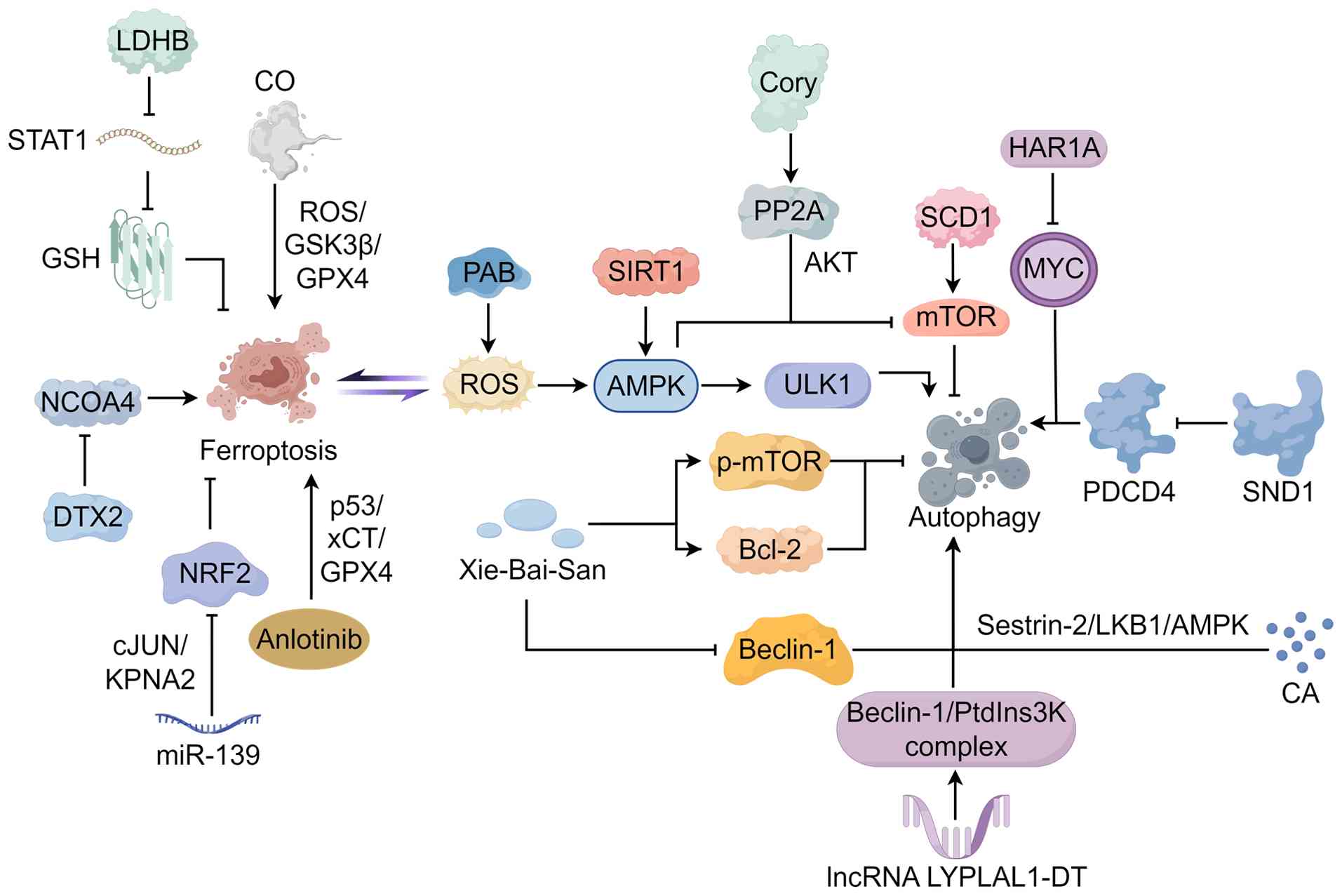 | Figure 2Mechanisms of the crosstalk
regulation between autophagy and ferroptosis in lung cancer and
lung cancer pharmacotherapy. The figure was created by Figdraw
(www.figdraw.com). LDHB, lactate dehydrogenase B;
GSH, glutathione; CO, carbon monoxide; PAB, pseudolaric acid B;
ROS, reactive oxygen species; Cory, corynoxine; PP2A, protein
phosphatase 2A; ULK1, unc-51 like kinase 1; SCD1, stearoyl-CoA
desaturase 1; SND1, staphylococcal nuclease and tudor domain
containing 1; PDCD4, programmed cell death 4; CA, carnosic acid;
lncRNA, long non-coding RNA; p-mTOR, phosphorylated mechanistic
target of rapamycin; miR-139, microRNA-139; NRF2, nuclear factor,
erythroid 2 like 2; DTX2, deltex E3 ubiquitin ligase 2; NCOA4,
nuclear receptor coactivator 4. |
With radiotherapy
Modulating the autophagy-ferroptosis axis also
represents a viable strategy to counteract radioresistance, as it
functions bidirectionally to either sensitize cells or promote
resistance.
Several mechanisms have been identified to augment
radiosensitivity through ferroptosis induction. For example,
miR-139 increases the radiosensitivity of NSCLC by directly
targeting cJUN and KPNA2, disrupting the NRF2 pathway and
amplifying radiation-induced lipid peroxidation (122). Similarly, targeting NRF2 to
upregulate phosphorylase kinase catalytic subunit γ2 promotes
ferritinophagy and mitochondrial stress-dependent ferroptosis,
increasing radiosensitivity (123). MAF bZIP transcription factor F
enhances sensitivity to both cisplatin and ionizing radiation by
regulating SLC7A11, CDK6 and CDKN2C to induce ferroptosis and
arrest the cell cycle (124).
Conversely, certain pathways activated by
radiotherapy can promote resistance via pro-survival autophagy.
Upregulation of long-chain acyl-CoA synthase (ACSL)6 enhances
post-radiotherapy invasiveness in LC by augmenting
FLI1-collagen-mediated autophagic flux (125). Lactoferrin establishes a
positive feedback loop involving AMPK/SP2/NEAT1/miR-214-5p to
foster autophagy and contribute to radioresistance in lung squamous
cell carcinoma (126). These
opposing effects highlight how the net impact of the crosstalk on
radiosensitivity is determined by specific molecular contexts,
presenting both a challenge and an opportunity for personalized
radiosensitization strategies.
With targeted therapy
Targeted therapy resistance, a major clinical
challenge, is amenable to intervention via the
autophagy-ferroptosis axis, a key determinant of cell fate under
stress, through two primary strategies. i) Inducing ferroptosis to
reverse targeted therapy resistance.
Ferroptosis induction has demonstrated considerable
potential to circumvent resistance to targeted therapies,
particularly in EGFR-mutant and KRAS-driven LC.
In EGFR-mutant NSCLC, multiple approaches have been
shown to restore sensitivity by activating ferroptosis. For
example, Tenovin-3 triggers mitochondrial pathway-dependent
ferroptosis in EGFR 19del cells (127). Downregulation of
apoptosis-associated tyrosine kinase promotes iron accumulation via
enhanced endosome recycling, sensitizing EGFR-TKI-resistant cells
to ferroptosis inducers such as RSL3 (128). Beyond classic TKIs, the
multi-targeted agent anlotinib exerts potent antitumor effects in
NSCLC by specifically inducing ferroptosis via the p53/xCT/GPX4
axis (129). Conversely,
upregulation of the anti-apoptotic protein MCL1 has been linked to
acquired resistance against sorafenib-induced ferroptosis,
revealing a potential adaptive mechanism (130).
In KRAS-driven LC, targeting metabolic dependencies
provides another strategic entry point. Inhibition of LDHB
upregulates STAT1, suppresses SLC7A11 expression and disrupts GSH
metabolism, thereby surmounting intrinsic ferroptosis resistance in
this subtype (131).
Collectively, these studies establish ferroptosis
induction as a viable strategy to overcome targeted therapy
resistance. They reveal actionable targets spanning signaling,
metabolic, and vesicular recycling pathways, offering a robust
foundation for designing novel drug combinations.
ii) Modulating autophagy to overcome drug
resistance. Targeting autophagy represents another critical
strategy to overcome resistance to targeted therapies. Its role is
context-dependent, functioning as either a pro-survival mechanism
that confers resistance or a process that can be co-opted to
enhance drug sensitivity. Autophagy can be co-opted by cancer cells
to drive resistance to EGFR-TKIs. For example, PD-L1 upregulation
activates pro-survival autophagy via the MAPK pathway, contributing
to gefitinib resistance and NSCLC progression (132). Similarly, autophagy mediated by
the STAT3/FOXM1/ATG7 axis has been identified as a novel mechanism
underlying icotinib resistance (133). Pharmacological inhibition of
such pro-survival autophagy can restore drug sensitivity. The
traditional formulation Xie-Bai-San, for example, suppresses
gefitinib-induced autophagic flux by modulating the mTOR/Beclin-1
pathway, thereby blocking this escape route (134). Beyond direct suppression,
strategic combination therapies can redirect cellular responses.
The concurrent inhibition of MEK (trametinib) and glutamine
metabolism (V-9302) enhances antitumor efficacy by orchestrating
the FOXO3a/FOXM1 axis and autophagy, leading to pyroptosis and cell
cycle arrest (135).
A similar duality is observed in ALK-rearranged
NSCLC. While ALK inhibition itself can trigger an LC3B-independent
macroautophagic flux that promotes the survival of
EML4-ALK+ cells (136), strategically inducing autophagy
in a controlled manner can be beneficial. Activating SIRT1
upregulates AMPK, inhibits mTOR phosphorylation and induces
autophagy, thereby sensitizing EML4-ALK mutant cells (L1196M,
G1202R) to crizotinib (137).
These findings underscore the complex, dual role of
autophagy in targeted therapy resistance. The central challenge
lies in precisely determining whether to inhibit or activate
autophagic pathways, a decision that depends on the specific
molecular context, including the driver mutation and the resistance
mechanism at play.
Combination with immunotherapy
Patient selection for combination strategies is
critical. In NSCLC, 18F-RGD uptake detected by PET is negatively
associated with tumoral PD-L1 expression (138), a finding that may inform
patient selection for combined immunotherapy and
autophagy-ferroptosis modulation. The integration of immune
checkpoint inhibitors (ICIs) with modulators of autophagy and
ferroptosis has emerged as a promising strategy to overcome
resistance to immunotherapy in LC. This approach seeks to enhance
tumor immunogenicity, reactivate antitumor immunity and improve
durable clinical responses. i) Modulating autophagy to improve
immunotherapeutic efficacy. Autophagy exerts a dual influence on
tumor-immune interactions, which can be strategically modulated to
overcome ICI resistance. On one hand, induction of autophagy can
promote the degradation of immunosuppressive proteins. For example,
the PPARγ-LC3 interaction facilitates autophagic degradation of
PD-L1 in lysosomes, thereby attenuating immune evasion in NSCLC
(139). Similarly, the small
molecule USP24-i-101 facilitates autophagy-dependent PD-L1
degradation and may help mitigate acquired resistance to ICIs
(140). On the other hand,
tumor cells can exploit autophagy as a protective mechanism. Under
glutamine-depleted conditions in the tumor microenvironment,
autophagy mediates degradation of the IFN-γ receptor, impairing
IFN-γ signaling and contributing to resistance against
anti-PD-1/PD-L1 therapy (141).
This duality underscores the importance of context-specific
modulation and highlights the current lack of reliable biomarkers
to guide patient selection for autophagy-targeting
combinations.
ii) Inducing ferroptosis to synergize with
immunotherapy. Ferroptosis inducers can potentiate immunotherapy by
triggering immunogenic cell death, releasing tumor antigen and
activating antitumor immune responses. For example, combining
ROR1-targeted CAR-T cells with ferroptosis inducers enhances lipid
peroxidation and augments antitumor efficacy in a mouse model of
NSCLC (107). The natural
compound fascaplysin induces both apoptosis and ferroptosis,
thereby sensitizing NSCLC to anti-PD-1 therapy (142). Moreover, the protein LRRC1B has
been shown to modulate ICI efficacy by regulating ferroptosis
sensitivity in NSCLC cells (143). However, excessive or systemic
induction of ferroptosis may cause normal tissue toxicity and
impair immune cell function, emphasizing the need for precise
spatiotemporal control in therapeutic regimens.
The toxicities of ferroptosis inducers manifest as
systemic iron overload and lipid peroxidation-mediated multi-organ
injury, requiring rigorous longitudinal monitoring and dynamic
management (11). Beyond classic
bone-marrow suppression and transaminase elevation, four
pulmonary-specific and systemic hazards demand particular vigilance
in patients with LC: Radiation-like pneumonitis/fibrosis,
iron-catalyzed oxidative stress, CD8+ T-cell functional
exhaustion and QT-interval prolongation (144).
iii) Coordinated targeting of autophagy and
ferroptosis. Simultaneous modulation of autophagy and ferroptosis
may yield synergistic benefits and overcome compensatory resistance
mechanisms. For example, combining radiotherapy with PD-L1 blockade
and autophagy inhibition activates the cGAS-STING pathway and
promotes T-cell-mediated antitumor immunity (145). Activation of the cGAS-STING
pathway in tumor cells can further induce ferroptosis (146). Conversely, in cancer stem
cells, the CPT1A/c-Myc feedback loop activates the NRF2/GPX4 axis
and downregulates ACSL4, thereby suppressing ferroptosis and
potentially diminishing immunotherapeutic response (147). Notably, NRF2 promotes
autophagosome formation and enhances autophagic activity, which in
turn inhibits apoptosis in NSCLC cells (148). These findings illustrate the
therapeutic potential of dual-pathway intervention, although the
complex interplay between autophagy, ferroptosis and various immune
cell populations requires further mechanistic elucidation to
optimize combination strategies. Fig. 3 depicts the involvement of
autophagy and ferroptosis across major therapeutic modalities in
LC. As illustrated in Table I,
most therapeutic strategies remain in the preclinical stage.
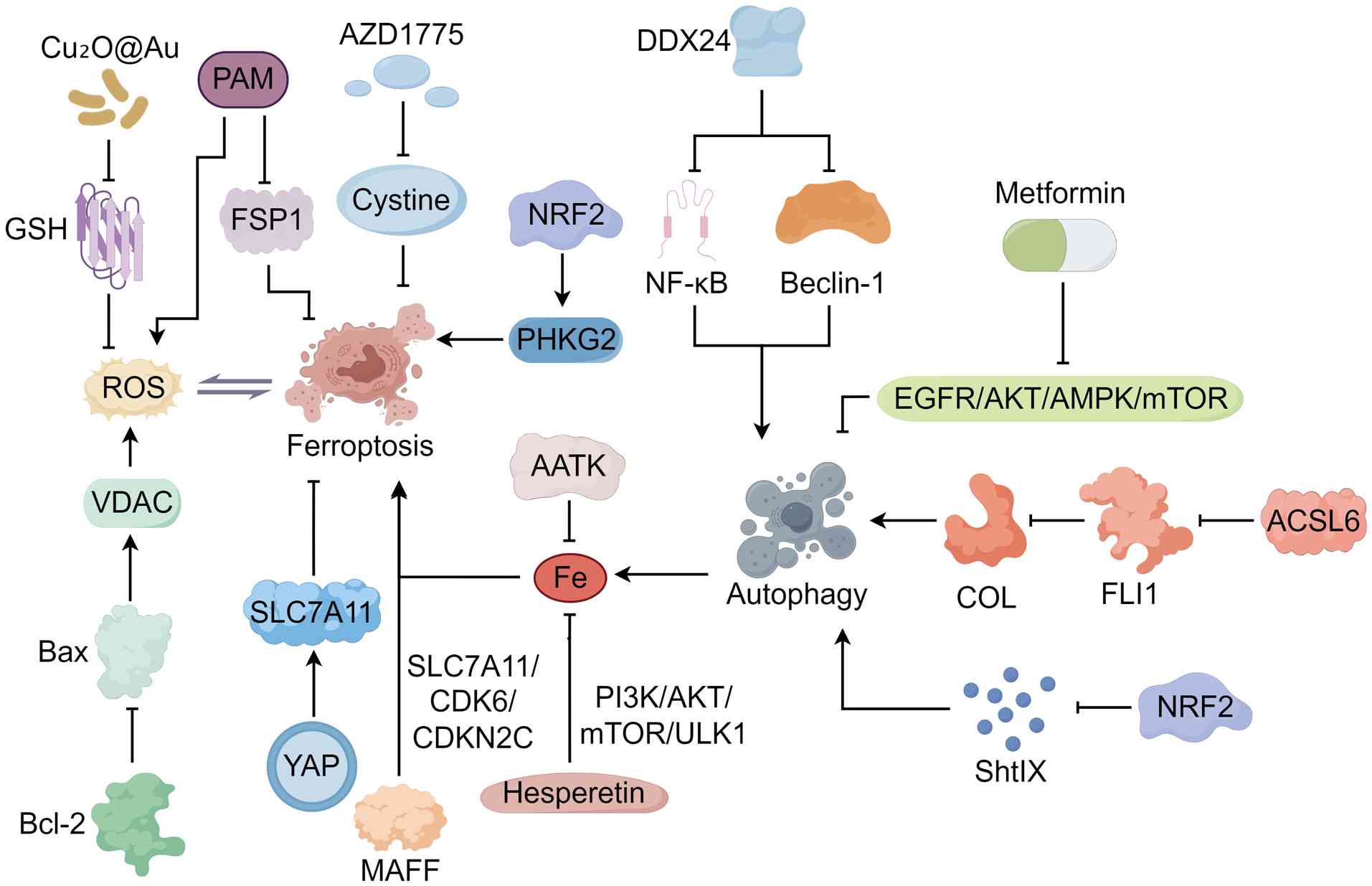 | Figure 3Mechanisms of autophagy and
ferroptosis in lung cancer gene therapy, targeted therapy and
immunotherapy. The figure was created by Figdraw (www.figdraw.com). GSH, glutathione; ROS, reactive
oxygen species; PAM, plasma-activated medium; FSP1, ferroptosis
suppressor protein 1; NRF2, nuclear factor, erythroid 2 like 2;
PHKG2, phosphorylase kinase catalytic subunit γ2; DDX24, DEAD-box
helicase 24; ACSL6, long-chain acyl-CoA synthase 6; COL, collagen;
AATK, apoptosis-associated tyrosine kinase; ShtIX,
S-3-hydroxy-7',2',4'-trimethoxyisoflavone; MAFF, MAF bZIP
transcription factor F; YAP, Yes-associated protein; SLC7A11,
solute carrier family 7 member 11; VDAC, voltage-dependent anion
channel. |
 | Table IRepresentative compounds targeting
autophagy-ferroptosis crosstalk in lung cancer: Mechanisms and
research stages. |
Table I
Representative compounds targeting
autophagy-ferroptosis crosstalk in lung cancer: Mechanisms and
research stages.
| First author,
year | Compound/agent | Targeted
mechanism | Research stage | (Refs.) |
|---|
| Mauthe, 2018 | Chloroquine | Inhibits
autophagosome-lysosome fusion, blocks autophagic flux | In vitro and
in vivo | (82) |
| Li, 2018 | HCQ | Reduces lysosomal
drug sequestration and enhances CD8+ T-cell
responses | Random phase II
multicenter trial | (83) |
| Fang, 2024 | HFn@HCQ
(nanocarrier) | Targeted delivery
of HCQ, enhances autophagy inhibition | In vitro and
in vivo | (85) |
| Pan, 2019 | Erastin | Inhibits system
Xc−, depletes GSH, induces GPX4-mediated ferroptosis;
overcomes radioresistance in NSCLC | In
vitro | (87) |
| Tang, 2021 | Curcumin | Induces iron
overload, GSH depletion and lipid peroxidation | In
vitro | (75) |
| Liu, 2025 | Rabeprazole | Inhibits autophagy,
triggers pyroptosis and ferroptosis | In vitro and
in vivo | (90) |
| Xu, 2024 | ML385 | Inhibits NRF2,
enhances ferroptosis sensitivity | In
vitro | (101) |
| Fu, 2024 | Sanggenol L | Regulates
miR-26a-1-3p/MDM2/p53/SLC7A11 axis, induces ferroptosis | In vitro and
in vivo | (103) |
| Xia, 2025 | Metformin | Induces autophagy
via EGFR/AKT/AMPK/mTOR pathway in SCLC | In vitro and
in vivo | (92) |
| Zheng, 2024 | Hydrogen
sulfide | Regulates
homocysteine metabolism, enhances ferroptosis in NSCLC | In vitro and
in vivo | (94) |
| Gai, 2020 | Folate-modified
liposome (erastin + MT1DP) | Enhances
ferroptosis sensitivity via miR-365a-3p/NRF2 axis | In vitro and
in vivo | (105) |
| Tsai, 2024 | Iron-platinum alloy
nanoparticles | Reverses TKI
resistance by activating ferroptosis | In vitro and
in vivo | (113) |
| Luo, 2024 | Pseudolaric acid
B | Modulates
ROS/AMPK/mTOR/autophagy pathway | In vitro and
in vivo | (95) |
| Hou, 2024 | Corynoxine | Activates PP2A,
inhibits AKT-mTOR/GSK3β axes, induces autophagic cell death | In vitro and
in vivo | (98) |
| Bhatt, 2023 | HCQ +
trametinib | Inhibits autophagy
and blocks the MEK pathway, synergistically induces ferroptosis in
LKB1/KRAS co-mutant NSCLC | In vitro and
in vivo | (86) |
| Li, 2025 | ROR1-targeted CAR-T
cell therapy + ferroptosis inducers | Enhances lipid
peroxidation and antitumor efficacy | In vitro and
in vivo | (107) |
| Ma, 2024 | Xie-Bai-San | Inhibits autophagy
(via the mTOR/Beclin-1 pathway), reverses gefitinib resistance | In vitro and
in vivo | (134) |
| Liu, 2025 | Trametinib +
V-9302 | Regulates the
FOXO3a/FOXM1 axis and autophagy, induces pyroptosis/cell cycle
arrest | In vitro and
in vivo | (135) |
| Liu, 2024 | DTX2 knockdown | Stabilizes NCOA4,
promotes ferritinophagy and reverses cisplatin resistance in
NSCLC | In vitro and
in vivo | (121) |
| ALMatrafi,
2025 | TMEM164
overexpression | Activates
autophagy-dependent ferroptosis and synergizes with anti-PD-1 to
enhance antitumor immunity | In vitro and
in vivo | (13) |
| Yang, 2024 | Crizotinib + SIRT1
activation | Upregulates AMPK,
inhibits mTOR and induces autophagy to sensitize EML4-ALK mutant
cells | In
vitro | (137) |
Targeting the autophagy-ferroptosis axis represents
a rational and evolving approach to potentiate the efficacy of
immunotherapy in LC. Successful clinical translation will depend on
identifying predictive biomarkers, optimizing the timing and
sequence of combination therapies, and developing agents capable of
selectively modulating these pathways within the tumor
microenvironment.
Research challenges and future
perspectives
Current challenges
Notable challenges persist in translating the
autophagy-ferroptosis axis into clinical applications for LC,
despite growing mechanistic understanding. The primary hurdle is
the intrinsic complexity and context-dependency of this interplay.
Its regulatory network remains incompletely mapped, as it involves
numerous overlapping and often contradictory signaling pathways. A
critical gap is the identification of definitive molecular switches
that determine whether the crosstalk promotes tumor cell survival
or death in a given therapeutic setting, which severely impedes the
rational design of pathway-specific modulators.
Substantial inter- and intra-tumoral heterogeneity,
epitomized by the profound biological divergence between LUAD and
SCLC (149), remains a major
obstacle to developing universal treatment paradigms. Although
multimodal regimens combining radiotherapy, chemotherapy and
immunotherapy have achieved notable clinical benefits across LC
subtypes (150), the molecular
drivers of these differential responses remain poorly understood.
This knowledge gap represents a critical barrier to advancing truly
precision-based therapeutic strategies.
Most therapeutic interventions remain in preclinical
development, with clinical translation hindered by two key
barriers: The lack of validated biomarkers for patient
stratification and incomplete elucidation of synergistic
combination mechanisms. A central, unresolved question remains how
to precisely balance the modulation of autophagy and ferroptosis to
maximize therapeutic efficacy while minimizing the risks of tumor
promotion or off-target tissue damage.
Future directions and prospects
To tackle these barriers, future research should
prioritize the following key areas.
Deciphering context-specific regulatory
networks
Employing integrated multi-omics approaches across
diverse LC subtypes and dynamic treatment states is essential. This
will enable the construction of a high-resolution map of the
molecular landscape, the identification of subtype-specific
regulatory nodes and switches (151), and address the fundamental
knowledge gaps highlighted in preceding sections, thereby laying a
foundation for precise therapeutic targeting.
Development of predictive biomarkers
Predictive biomarkers, particularly those linked to
key regulatory axes such as NRF2/p62, COX7A1 or USP13, require
urgent discovery and clinical validation to reliably predict
sensitivity to ferroptosis inducers or autophagy modulators.
Correlating longitudinal multi-omics data from large,
well-annotated patient cohorts with clinical outcomes will thus
enable accurate patient stratification, guide therapeutic
decision-making, and increase the success rate of clinical
trials.
Leveraging artificial intelligence (AI)
and integrative analytics
Advanced computational approaches, including AI and
machine learning algorithms, are increasingly employed to decipher
the heterogeneity of autophagy-ferroptosis regulation and optimize
therapeutic decisions by integrating multimodal data (genomics,
transcriptomics, proteomics and clinical data) (152,153). Their core value is concentrated
in three key directions, as follows: i) Deep learning,
network-based machine learning and other algorithms are utilized to
mine patient-specific autophagy-ferroptosis molecular signatures,
enabling precise stratification of treatment responders and
non-responders, and providing a basis for individualized
therapeutic target selection (153). ii) AI models trained on
longitudinal clinical and molecular data integrate molecular
features and clinical variables to predict individual sensitivity
to specific therapeutic regimens, reducing reliance on therapeutic
trial and error, and optimizing drug dosage and sequence (154). iii) AI-driven real-time
analytical technologies monitor dynamic reprogramming in crosstalk
pathways during treatment, guiding timely adjustments to
therapeutic strategies and effectively overcoming inter-patient
variability in treatment responses among advanced patients
(155,156).
These technologies may accelerate the discovery of
novel therapeutic targets and the development of personalized
regimens, serving as critical cutting-edge tools to advance
autophagy-ferroptosis-targeted therapy from basic research to
clinical translation.
Advancing translational and clinical
research
Building upon the mechanistic foundations,
translational efforts must now focus on: i) Optimizing the
pharmacokinetics, biodistribution and safety profiles of advanced
delivery systems; ii) conducting large-scale, biomarker-driven
clinical trials to evaluate rational combination therapies; and
iii) exploring the underexplored role of the autophagy-ferroptosis
crosstalk in critical clinical processes such as metastasis, tumor
dormancy and therapy-resistant recurrence.
Conclusion
The dynamic crosstalk between autophagy and
ferroptosis represents a promising frontier for innovating LC
therapeutic strategies. While notable scientific and translational
challenges remain, concerted efforts to elucidate context-dependent
mechanisms, develop precision biomarkers and tools, and rigorously
evaluate targeted combinations in the clinic are poised to yield
transformative advances. Success in this endeavor may result in the
identification of effective, durable and safer therapeutic
paradigms, ultimately improving outcomes and quality of life for
patients with LC.
Availability of data and materials
Not applicable.
Authors' contributions
RS designed the research and analyzed the
literature. YZ drafted the manuscript. Data authentication is not
applicable. Both authors contributed to literature collection,
drafting and revising the manuscript, and have read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
No funding was received.
References
|
1
|
Filho AM, Laversanne M, Ferlay J, Colombet
M, Piñeros M, Znaor A, Parkin DM, Soerjomataram I and Bray F: The
GLOBOCAN 2022 cancer estimates: Data sources, methods, and a
snapshot of the cancer burden worldwide. Int J Cancer.
156:1336–1346. 2025. View Article : Google Scholar
|
|
2
|
Wang H, Niu X, Jin Z, Zhang S, Fan R, Xiao
H and Hu SS: Immunotherapy resistance in non-small cell lung
cancer: From mechanisms to therapeutic opportunities. J Exp Clin
Cancer Res. 44:2502025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu X, Tuerxun H and Zhao Y, Li Y, Wen S,
Li X and Zhao Y: Crosstalk between ferroptosis and autophagy:
Broaden horizons of cancer therapy. J Transl Med. 23:182025.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Song YJ, Zhang SS, Guo XL, Sun K, Han ZP,
Li R, Zhao QD, Deng WJ, Xie XQ, Zhang JW, et al: Autophagy
contributes to the survival of CD133+ liver cancer stem cells in
the hypoxic and nutrient-deprived tumor microenvironment. Cancer
Lett. 339:70–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cui Y, Cao X, Zhang Y, Fu C, Li D, Sun Y,
Zhang Y, Xu T, Tsukamoto T, Cao D and Jiang J: Protein phosphatase
1 regulatory subunit 15 A (PPP1R15A) promoted the progression of
gastric cancer by activating cell autophagy under energy stress. J
Exp Clin Cancer Res. 44:522025. View Article : Google Scholar
|
|
6
|
Wu X, Wu J, Dai T, Wang Q, Cai S, Wei X,
Chen J and Jiang Z: β-elemene promotes miR-127-3p maturation,
induces NSCLCs autophagy, and enhances macrophage M1 polarization
through exosomal communication. J Pharm Anal. 14:1009612024.
View Article : Google Scholar
|
|
7
|
Fernandez-Acosta R, Vintea I, Koeken I,
Hassannia B and Vanden Berghe T: Harnessing ferroptosis for
precision oncology: Challenges and prospects. BMC Biol. 23:572025.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He Y, Lin Y, Song J, Song M, Nie X, Sun H,
Xu C, Han Z and Cai J: From mechanisms to medicine: Ferroptosis as
a Therapeutic target in liver disorders. Cell Commun Signal.
23:1252025. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu K, Vaughan AJ, Bossowski JP, Hao Y,
Ziogou A, Kim SM, Kim TH, Nakamura MN, Pillai R, Mancini M, et al:
Targeting FSP1 triggers ferroptosis in lung cancer. Nature.
649:487–495. 2026. View Article : Google Scholar :
|
|
10
|
Peng S, Chen G, Yu KN, Feng Y, Zhao L,
Yang M, Cao W, Almahi WAA, Sun M, Xu Y, et al: Synergism of
non-thermal plasma and low concentration RSL3 triggers ferroptosis
via promoting xCT lysosomal degradation through ROS/AMPK/mTOR axis
in lung cancer cells. Cell Commun Signal. 22:1122024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Z, Zhang Y, Zhong W, Wu K, Zhong T and
Jiang T: Targeting ferroptosis: A promising approach for treating
lung carcinoma. Cell Death Discov. 11:332025. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bian Y, Shan G, Liang J, Hu Z, Sui Q, Shi
H, Wang Q, Bi G and Zhan C: Retinoic acid receptor alpha inhibits
ferroptosis by promoting thioredoxin and protein phosphatase 1F in
lung adenocarcinoma. Commun Biol. 7:7512024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Al Matrafi TA: Deciphering the role of
TMEM164 in autophagy-mediated ferroptosis and immune modulation in
non-small cell lung cancer. Cell Immunol. 409-410:1049152025.
View Article : Google Scholar
|
|
14
|
Yu F, Zhang Q, Liu H, Liu J, Yang S, Luo
X, Liu W, Zheng H, Liu Q, Cui Y, et al: Dynamic O-GlcNAcylation
coordinates ferritinophagy and mitophagy to activate ferroptosis.
Cell Discov. 8:402022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jia M, Yue X, Sun W, Zhou Q, Chang C, Gong
W, Feng J, Li X, Zhan R, Mo K, et al: ULK1-mediated metabolic
reprogramming regulates Vps34 lipid kinase activity by its
lactylation. Sci Adv. 9:eadg49932023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pyo KE, Kim CR, Lee M, Kim JS, Kim KI and
Baek SH: ULK1 O-GlcNAcylation is crucial for activating VPS34 via
ATG14L during autophagy initiation. Cell Rep. 25:2878–2890.e4.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghazi PC, O'Toole KT, Srinivas Boggaram S,
Scherzer MT, Silvis MR, Zhang Y, Bogdan M, Smith BD, Lozano G,
Flynn DL, et al: Inhibition of ULK1/2 and KRAS(G12C) controls tumor
growth in preclinical models of lung cancer. Elife. 13:RP969922024.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Changotra H, Kaur S, Yadav SS, Gupta GL,
Parkash J and Duseja A: ATG5: A central autophagy regulator
implicated in various human diseases. Cell Biochem Funct.
40:650–667. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Runwal G, Stamatakou E, Siddiqi FH, Puri
C, Zhu Y and Rubinsztein DC: LC3-positive structures are prominent
in autophagy-deficient cells. Sci Rep. 9:101472019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gómez-Virgilio L, Silva-Lucero MD,
Flores-Morelos DS, Gallardo-Nieto J, Lopez-Toledo G,
Abarca-Fernandez AM, Zacapala-Gómez AE, Luna-Muñoz J, Montiel-Sosa
F, Soto-Rojas LO, et al: Autophagy: A key regulator of homeostasis
and disease: An overview of molecular mechanisms and modulators.
Cells. 11:22622022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Restrepo LJ and Baehrecke EH: Regulation
and functions of autophagy during animal development. J Mol Biol.
436:1684732024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fang C, Wu W, Ni Z, Liu Y, Luo J, Zhou Y,
Gong C, Hu D, Yao C, Chen X, et al: Ailanthone inhibits non-small
cell lung cancer growth and metastasis through targeting
UPF1/GAS5/ULK1 signaling pathway. Phytomedicine. 128:1553332024.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ding X, Li X, Jiang Y, Li Y, Li H, Shang
L, Feng G, Zhang H, Xu Z, Yang L, et al: RGS20 promotes non-small
cell lung carcinoma proliferation via autophagy activation and
inhibition of the PKA-Hippo signaling pathway. Cancer Cell Int.
24:932024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liang DM, Li YJ, Zhang JX, Shen HH, Wu CX,
Xie N, Liang Y, Li YM, Xue JN, Sun HF, et al: m6A-methylated
KCTD21-AS1 regulates macrophage phagocytosis through CD47 and cell
autophagy through TIPR. Commun Biol. 7:2152024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wen Y, Sun X, Zeng L, Liang S, Li D, Chen
X, Zeng F, Zhang C, Wang Q, Zhong Q, et al: CDK4/6 inhibitors
impede chemoresistance and inhibit tumor growth of small cell lung
cancer. Adv Sci (Weinh). 11:e24006662024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen J, Gou Z, Yang G, Zhou L, Kim AN, Shi
W and Zhou Y: Ferroptosis, a distinct form of cell death, and
research progress on its modulators. Pharmaceuticals (Basel).
18:17852025. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Henning Y, Blind US, Larafa S, Matschke J
and Fandrey J: Hypoxia aggravates ferroptosis in RPE cells by
promoting the Fenton reaction. Cell Death Dis. 13:6622022.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ursini F and Maiorino M: Lipid
peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic
Biol Med. 152:175–185. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Griffiths K, Ida T, Morita M, Lamb RJ, Lee
JJ, Frenneaux MP, Fukuto JM, Akaike T, Feelisch M and Madhani M:
Cysteine hydropersulfide reduces lipid peroxidation and protects
against myocardial ischaemia-reperfusion injury-are endogenous
persulfides mediators of ischaemic preconditioning? Redox Biol.
60:1026052023. View Article : Google Scholar
|
|
30
|
Yao F, Zhao Y, Wang G, Zhao M, Hong X, Ye
Z, Dong F, Li W and Deng Q: Exosomal lncRNA ROR1-AS1 from
cancer-associated fibroblasts inhibits ferroptosis of lung cancer
cells through the IGF2BP1/SLC7A11 signal axis. Cell Signal.
120:1112212024. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yan RY, Liu D, Guo HJ, Liu M, Lv D,
Björkblom B, Wu M, Yu H, Leng H, Lu B, et al: LAPTM4B counteracts
ferroptosis via suppressing the ubiquitin-proteasome degradation of
SLC7A11 in non-small cell lung cancer. Cell Death Dis. 15:4362024.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li K, Yang B, Du Y, Ding Y, Shen S, Sun Z,
Liu Y, Wang Y, Cao S, Ren W, et al: The HOXC10/NOD1/ERK axis drives
osteolytic bone metastasis of pan-KRAS-mutant lung cancer. Bone
Res. 12:472024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma R, Yang D, Wang P, Zhang Z, Zhang X,
Song J, Liu H, Liu S, Zhang Y and Zou L: Oncogenic RIT1 mutations
confer ferroptosis vulnerability in lung adenocarcinoma. Biol
Direct. 20:192025. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim JW, Min DW, Kim D, Kim J, Kim MJ, Lim
H and Lee JY: GPX4 overexpressed non-small cell lung cancer cells
are sensitive to RSL3-induced ferroptosis. Sci Rep. 13:88722023.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu KY, Wei GX, Qi WH, Ye C, Liu Y, Wang S,
Yang F and Tang J: CircPOLA2 sensitizes non-small cell lung cancer
cells to ferroptosis and suppresses tumorigenesis via the
Merlin-YAP signaling pathway. iScience. 27:110832024. View Article : Google Scholar
|
|
36
|
Tang D, Chen X, Kang R and Kroemer G:
Ferroptosis: Molecular mechanisms and health implications. Cell
Res. 31:107–125. 2021. View Article : Google Scholar :
|
|
37
|
Wang B, Wang Y, Zhang J, Hu C, Jiang J, Li
Y and Peng Z: ROS-induced lipid peroxidation modulates cell death
outcome: mechanisms behind apoptosis, autophagy, and ferroptosis.
Arch Toxicol. 97:1439–1451. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liao H, Liu S, Ma Q, Huang H, Goel A,
Torabian P, Mohan CD and Duan C: Endoplasmic reticulum stress
induced autophagy in cancer and its potential interactions with
apoptosis and ferroptosis. Biochim Biophys Acta Mol Cell Res.
1872:1198692025. View Article : Google Scholar
|
|
39
|
Luo C, Liang H, Ji M, Ye C, Lin Y, Guo Y,
Zhang Z, Shu Y, Jin X, Lu S, et al: Autophagy induced by mechanical
stress sensitizes cells to ferroptosis by NCOA4-FTH1 axis.
Autophagy. 21:1263–1282. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang M, Zeng G, Xiong B, Zhu X, Guo J,
Chen D, Zhang S, Luo M, Guo L and Cai L: ALOX5 promotes
autophagy-dependent ferroptosis by activating the AMPK/mTOR pathway
in melanoma. Biochem Pharmacol. 212:1155542023. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang Y, Hu Y, Xu R, Jin X and Jiao W:
Plin2 inhibits autophagy via activating AKT/mTOR pathway in
non-small cell lung cancer. Exp Cell Res. 435:1139552024.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen S, Cai D, Zhao Q, Wu J, Zhou X, Xu H,
Li X, Zhang R, Peng W, Li G and Nan A: NSUN2-mediated m5C
modification of circFAM190B promotes lung cancer progression by
inhibiting cellular autophagy. Int J Biol Macromol. 306:1415282025.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li XQ, Cheng XJ, Wu J, Wu KF and Liu T:
Targeted inhibition of the PI3K/AKT/mTOR pathway by
(+)-anthrabenzoxocinone induces cell cycle arrest, apoptosis, and
autophagy in non-small cell lung cancer. Cell Mol Biol Lett.
29:582024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee S, Hwang N, Seok BG, Lee S, Lee SJ and
Chung SW: Autophagy mediates an amplification loop during
ferroptosis. Cell Death Dis. 14:4642023. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Su Y, Jin W, Niu J, Lyu X, Hao Q, Lyu Q,
Sheng N, Liu Z and Yu X: Harnessing an MMP-independent NIR probe
unveiling the different mitochondrial cristae changes during
mitophagy and ferroptosis under STED microscopy. Anal Chem.
97:2906–2913. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu S, Chen JH, Li LC, Ye ZP, Liu JN, Chen
YH, Hu BX, Tang JH, Feng GK, Li ZM, et al: Susceptibility of
mitophagy-deficient tumors to ferroptosis induction by relieving
the suppression of lipid peroxidation. Adv Sci (Weinh).
12:e24125932025. View Article : Google Scholar :
|
|
47
|
Yan Y, Ran X, Zhou Z, Gu Y, Wang R, Qiu C,
Sun Y and Wang J, Xiao J, Lu Y and Wang J: FGF21 inhibits
ferroptosis caused by mitochondrial damage to promote the repair of
peripheral nerve injury. Front Pharmacol. 15:13586462024.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yamashita SI, Sugiura Y, Matsuoka Y, Maeda
R, Inoue K, Furukawa K, Fukuda T, Chan DC and Kanki T: Mitophagy
mediated by BNIP3 and NIX protects against ferroptosis by
downregulating mitochondrial reactive oxygen species. Cell Death
Differ. 31:651–661. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Han Z, Wang B, Wen YQ, Li YN, Feng CX,
Ding XS, Shen Y, Yang Q and Gao L: Acteoside alleviates lipid
peroxidation by enhancing Nrf2-mediated mitophagy to inhibit
ferroptosis for neuroprotection in Parkinson's disease. Free Radic
Biol Med. 223:493–505. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gao G and Zhang X: Broadening horizons:
Research on ferroptosis in lung cancer and its potential
therapeutic targets. Front Immunol. 16:15428442025. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li Y, Du Y, Zhou Y, Chen Q, Luo Z, Ren Y,
Chen X and Chen G: Iron and copper: Critical executioners of
ferroptosis, cuproptosis and other forms of cell death. Cell Commun
Signal. 21:3272023. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Subburayan K, Thayyullathil F,
Pallichankandy S, Cheratta AR, Alakkal A, Sultana M, Drou N, Arshad
M, Palanikumar L, Magzoub M, et al: Tumor suppressor Par-4
activates autophagy-dependent ferroptosis. Commun Biol. 7:7322024.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Forcina GC and Dixon SJ: GPX4 at the
crossroads of lipid homeostasis and ferroptosis. Proteomics.
19:e18003112019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen L, Ning J, Linghu L, Tang J, Liu N,
Long Y, Sun J, Lv C, Shi Y, Tao T, et al: USP13 facilitates a
ferroptosis-to-autophagy switch by activation of the
NFE2L2/NRF2-SQSTM1/p62-KEAP1 axis dependent on the KRAS signaling
pathway. Autophagy. 21:565–582. 2025. View Article : Google Scholar :
|
|
55
|
Zhu L, Zhu D, Ran J, Li M, Lai Z, Zhou Y,
Luo L, Liu X, Mao K and Tian K: Autophagy aggravates multi-walled
carbon nanotube-induced ferroptosis by suppressing PGC-1
dependent-mitochondrial biogenesis in lung epithelial cells. Chem
Biol Interact. 400:1111582024. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cao Z, Tian K, Ran Y, Zhou H, Zhou L, Ding
Y and Tang X: Beclin-1: A therapeutic target at the intersection of
autophagy, immunotherapy, and cancer treatment. Front Immunol.
15:15064262024. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Feng Y, Xu J, Shi M, Liu R, Zhao L, Chen
X, Li M, Zhao Y, Chen J, Du W and Liu P: COX7A1 enhances the
sensitivity of human NSCLC cells to cystine deprivation-induced
ferroptosis via regulating mitochondrial metabolism. Cell Death
Dis. 13:9882022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hill SM, Wrobel L and Rubinsztein DC:
Post-translational modifications of Beclin 1 provide multiple
strategies for autophagy regulation. Cell Death Differ. 26:617–629.
2019. View Article : Google Scholar
|
|
59
|
Fujiwara N, Shibutani S, Ohama T and Sato
K: Protein phosphatase 6 dissociates the Beclin 1/Vps34 complex and
inhibits autophagy. Biochem Biophys Res Commun. 552:191–195. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhu L, He S, Huang L, Ren D, Nie T, Tao K,
Xia L, Lu F, Mao Z and Yang Q: Chaperone-mediated autophagy
degrades Keap1 and promotes Nrf2-mediated antioxidative response.
Aging Cell. 21:e136162022. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yang Y, Willis TL, Button RW, Strang CJ,
Fu Y, Wen X, Grayson PRC, Evans T, Sipthorpe RJ, Roberts SL, et al:
Cytoplasmic DAXX drives SQSTM1/p62 phase condensation to activate
Nrf2-mediated stress response. Nat Commun. 10:37592019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhou Y, Shen Y, Chen C, Sui X, Yang J,
Wang L and Zhou J: The crosstalk between autophagy and ferroptosis:
What can we learn to target drug resistance in cancer? Cancer Biol
Med. 16:630–646. 2019. View Article : Google Scholar
|
|
63
|
Ikeda F: Ubiquitin conjugating enzymes in
the regulation of the autophagy-dependent degradation pathway.
Matrix Biol. 100-101:23–29. 2021. View Article : Google Scholar
|
|
64
|
Kuang E, Qi J and Ronai Z: Emerging roles
of E3 ubiquitin ligases in autophagy. Trends Biochem Sci.
38:453–460. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cajee UF, Hull R and Ntwasa M:
Modification by ubiquitin-like proteins: Significance in apoptosis
and autophagy pathways. Int J Mol Sci. 13:11804–11831. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu L, Chen X, Wu L, Huang K, Wang Z,
Zheng Y, Zheng C, Zhang Z, Chen J, Wei J, et al: Ubiquitin ligase
subunit FBXO9 inhibits V-ATPase assembly and impedes lung cancer
metastasis. Exp Hematol Oncol. 13:322024. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Park E and Chung SW: ROS-mediated
autophagy increases intracellular iron levels and ferroptosis by
ferritin and transferrin receptor regulation. Cell Death Dis.
10:8222019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wu SY, Chen JW, Liu HY, Wang YC, Chu YS,
Huang CY, Lan KY, Liu HS and Lan SH: Secretory autophagy promotes
Rab37-mediated exocytosis of tissue inhibitor of metalloproteinase
1. J Biomed Sci. 29:1032022. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Koundouros N and Poulogiannis G:
Reprogramming of fatty acid metabolism in cancer. Br J Cancer.
122:4–22. 2020. View Article : Google Scholar :
|
|
70
|
Guo JY, Karsli-Uzunbas G, Mathew R, Aisner
SC, Kamphorst JJ, Strohecker AM, Chen G, Price S, Lu W, Teng X, et
al: Autophagy suppresses progression of K-ras-induced lung tumors
to oncocytomas and maintains lipid homeostasis. Genes Dev.
27:1447–1461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang Z, Sun J, Jin C, Zhang L, Wu L and
Tian G: Identification and validation of a fatty acid metabolism
gene signature for the promotion of metastasis in liver cancer.
Oncol Lett. 26:4572023. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wang K, Wang S, Zhang Y, Xie L and Song X
and Song X: SNORD88C guided 2'-O-methylation of 28S rRNA regulates
SCD1 translation to inhibit autophagy and promote growth and
metastasis in non-small cell lung cancer. Cell Death Differ.
30:341–355. 2023. View Article : Google Scholar
|
|
73
|
Khilwani R and Singh S: Traversing through
the mechanistic event analysis in IL-6 and IL-17 signaling for a
new therapeutic paradigm in NSCLC. Int J Mol Sci. 25:12162024.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Xanthopoulou E, Lamprou I, Mitrakas AG,
Michos GD, Zois CE, Giatromanolaki A, Harris AL and Koukourakis MI:
Autophagy blockage up-regulates HLA-class-I molecule expression in
lung cancer and enhances anti-PD-L1 immunotherapy efficacy. Cancers
(Basel). 16:32722024. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tang X, Ding H, Liang M, Chen X, Yan Y,
Wan N, Chen Q, Zhang J and Cao J: Curcumin induces ferroptosis in
non-small-cell lung cancer via activating autophagy. Thorac Cancer.
12:1219–1230. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Jo A, Bae JH, Yoon YJ, Chung TH, Lee EW,
Kim YH, Joh HM and Chung JW: Plasma-activated medium induces
ferroptosis by depleting FSP1 in human lung cancer cells. Cell
Death Dis. 13:2122022. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang L, Li Q, Liu H and Li L: HPCAL1 is a
novel driver of autophagy-dependent ferroptosis. J Zhejiang Univ
Sci B. 24:1053–1056. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Debnath J, Gammoh N and Ryan KM: Autophagy
and autophagy-related pathways in cancer. Nat Rev Mol Cell Biol.
24:560–575. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Niu X, You Q, Hou K, Tian Y, Wei P, Zhu Y,
Gao B, Ashrafizadeh M, Aref AR, Kalbasi A, et al: Autophagy in
cancer development, immune evasion, and drug resistance. Drug
Resist Updat. 78:1011702025. View Article : Google Scholar
|
|
80
|
Guo D, Cai S, Deng L, Xu W, Fu S, Lin Y,
Jiang T, Li Q, Shen Z, Zhang J, et al: Ferroptosis in pulmonary
disease and lung cancer: Molecular mechanisms, crosstalk
regulation, and therapeutic strategies. MedComm (2020).
6:e701162025. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Li Y, Hahn T, Garrison K, Cui ZH, Thorburn
A, Thorburn J, Hu HM and Akporiaye ET: The vitamin E analogue α-TEA
stimulates tumor autophagy and enhances antigen cross-presentation.
Cancer Res. 72:3535–3545. 2012.PubMed/NCBI
|
|
82
|
Mauthe M, Orhon I, Rocchi C, Zhou X, Luhr
M, Hijlkema KJ, Coppes RP, Engedal N, Mari M and Reggiori F:
Chloroquine inhibits autophagic flux by decreasing
autophagosome-lysosome fusion. Autophagy. 14:1435–1455. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Li Y, Cao F, Li M, Li P, Yu Y, Xiang L, Xu
T, Lei J, Tai YY, Zhu J, et al: Hydroxychloroquine induced lung
cancer suppression by enhancing chemo-sensitization and promoting
the transition of M2-TAMs to M1-like macrophages. J Exp Clin Cancer
Res. 37:2592018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Lee SM, Hewish M, Ahmed S,
Papadatos-Pastos D, Karapanagiotou E, Blackhall F, Ford A, Young R,
Garcia A, Arora A, et al: Hydroxychloroquine in combination with
platinum doublet chemotherapy as first-line treatment for
extensive-stage small cell lung cancer (study 15): A randomised
phase II multi-centre trial. Eur J Cancer. 215:1151622025.
View Article : Google Scholar
|
|
85
|
Fang X, Zeng J, Li Y, Yu H, Wu Z and Qi X:
Hydroxychloroquine loaded hollow apoferritin nanocages for cancer
drug repurposing and autophagy inhibition. Eur J Pharm Biopharm.
203:1144732024. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Bhatt V, Lan T, Wang W, Kong J, Lopes EC,
Wang J, Khayati K, Raju A, Rangel M, Lopez E, et al: Inhibition of
autophagy and MEK promotes ferroptosis in Lkb1-deficient
Kras-driven lung tumors. Cell Death Dis. 14:612023. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Pan X, Lin Z, Jiang D, Yu Y, Yang D, Zhou
H, Zhan D, Liu S, Peng G, Chen Z and Yu Z: Erastin decreases
radioresistance of NSCLC cells partially by inducing GPX4-mediated
ferroptosis. Oncol Lett. 17:3001–3008. 2019.PubMed/NCBI
|
|
88
|
Xiong C, Ling H, Huang YD, Dong H, Xie B,
Hao Q and Zhou X: AZD1775 synergizes with SLC7A11 inhibition to
promote ferroptosis. Sci China Life Sci. 68:204–218. 2025.
View Article : Google Scholar
|
|
89
|
Bow YD, Ko CC, Chang WT, Chou SY, Hung CT,
Huang JL, Tseng CH, Chen YL, Li RN and Chiu CC: A novel quinoline
derivative, DFIQ, sensitizes NSCLC cells to ferroptosis by
promoting oxidative stress accompanied by autophagic dysfunction
and mitochondrial damage. Cancer Cell Int. 23:1712023. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Liu C, Sun RL, Wang HM, Xia YH and Wang
YJ: Rabeprazole inhibits lung cancer progression by triggering
NLRP3/CASP-1/caspase-dependent pyroptosis. Int Immunopharmacol.
146:1138062025. View Article : Google Scholar
|
|
91
|
Cao W, Sun MY, Yu KN, Zhao L, Feng Y, Tan
C, Yang M, Wang Y, Zhu F, Chen L, et al: Exogenous carbon monoxide
promotes GPX4-dependent ferroptosis through ROS/GSK3β axis in
non-small cell lung cancer. Cell Death Discov. 10:422024.
View Article : Google Scholar
|
|
92
|
Xia H, Tai XJ, Cheng W, Wu Y, He D, Wang
LF, Liu H, Zhang SY, Sun YT, Liu HZ, et al: Metformin inhibits the
growth of SCLC cells by inducing autophagy and apoptosis via the
suppression of EGFR and AKT signalling. Sci Rep. 15:60812025.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Liu X and Sun Y, Lv X, Song M, Sun J and
Sun Y: Targeting lipid peroxidation-associated ferroptosis
suppresses lung carcinoma progression by regulating cell cycle
arrest. Int Immunopharmacol. 138:1125182024. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zheng H, Chen H, Cai Y, Shen M, Li X, Han
Y, Deng X, Cao H, Liu J, Li H, et al: Hydrogen sulfide-mediated
persulfidation regulates homocysteine metabolism and enhances
ferroptosis in non-small cell lung cancer. Mol Cell.
84:4016–4030.e6. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Luo D, He F, Liu J, Dong X, Fang M, Liang
Y, Chen M, Gui X, Wang W, Zeng L, et al: Pseudolaric acid B
suppresses NSCLC progression through the ROS/AMPK/mTOR/autophagy
signalling pathway. Biomed Pharmacother. 175:1166142024. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Liu MH, Liu ZK and Liu F: An anti-tumor
protein PFAP specifically interacts with cholesterol-enriched
membrane domains of A549 cells and induces paraptosis and
endoplasmic reticulum stress. Int J Biol Macromol. 264:1306902024.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
O'Neill EJ, Sze NSK, Macpherson REK and
Tsiani E: Carnosic acid against lung cancer: Induction of autophagy
and activation of sestrin-2/LKB1/AMPK signalling. Int J Mol Sci.
25:19502024. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hou G, Hu W, Sang Y, Gan X, Xu H, Hu Q and
Cao X: Corynoxine triggers cell death via activating PP2A and
regulating AKT-mTOR/GSK3β axes in NSCLC. Biochem Pharmacol.
222:1161102024. View Article : Google Scholar
|
|
99
|
Yin J, Ding L, Yao S, Huang J, Xiao Y,
Wang Y, Zhang B, Rehmutulla M, Gu L, Tong Q and Zhang Y: Y9, a
Gboxin analog, displays anti-tumor effect in non-small cell lung
cancer by inducing lysosomal dysfunction and apoptosis. Bioorg
Chem. 153:1078202024. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Song C, Wang Z, Cao J, Dong Y and Chen Y:
Hesperetin alleviates aflatoxin B1 induced liver toxicity in mice:
Modulating lipid peroxidation and ferritin autophagy. Ecotoxicol
Environ Saf. 284:1168542024. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xu C, Chen Y, Zhou Z, Yan Y, Fu W, Zou P
and Ni D: ML385, an Nrf2 inhibitor, synergically enhanced celastrol
triggered endoplasmic reticulum stress in lung cancer cells. ACS
Omega. 9:43697–43705. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Liu J, Zhou S, Chen J, Lin H, Li Y, Zhang
X, Chen S, Lv X and Zhao H: Nrf2 inhibition and NCOA4-mediated
ferritinophagy activation synergistically exacerbated
S-3'-hydroxy-7', 2', 4'-trimethoxyisoxane induced ferroptosis in
lung cancer cells. Chem Biol Interact. 406:1113532025. View Article : Google Scholar
|
|
103
|
Fu R, You Y, Wang Y, Wang J, Lu Y, Gao R,
Pang M, Yang P and Wang H: Sanggenol L induces ferroptosis in
non-small cell lung cancer cells via regulating the
miR-26a-1-3p/MDM2/p53 signaling pathway. Biochem Pharmacol.
226:1163452024. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Lin JW, Zhou Y, Xiao HP, Wu LL, Li PC,
Huang MD, Xie D, Xu P, Li XX and Li ZX: Antitumor effects of a
Sb-rich polyoxometalate on non-small-cell lung cancer by inducing
ferroptosis and apoptosis. Chem Sci. 15:15367–15376. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Gai C, Liu C, Wu X, Yu M, Zheng J, Zhang
W, Lv S and Li W: MT1DP loaded by folate-modified liposomes
sensitizes erastin-induced ferroptosis via regulating
miR-365a-3p/NRF2 axis in non-small cell lung cancer cells. Cell
Death Dis. 11:7512020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhang C, Liu H, Li X, Xiao N, Chen H, Feng
H, Li Y, Yang Y, Zhang R, Zhao X, et al: Cold atmospheric plasma
enhances SLC7A11-mediated ferroptosis in non-small cell lung cancer
by regulating PCAF mediated HOXB9 acetylation. Redox Biol.
75:1032992024. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Li D, Zhang W, Wang R, Xie S, Wang Y, Guo
W, Huang Z, Lu C, Shan L, Liu H, et al: ROR1 CAR-T cells and
ferroptosis inducers orchestrate tumor ferroptosis via PC-PUFA2.
Biomark Res. 13:172025. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Huang F, Pang J, Xu L, Niu W, Zhang Y, Li
S and Li X: Hedyotis diffusa injection induces ferroptosis via the
Bax/Bcl2/VDAC2/3 axis in lung adenocarcinoma. Phytomedicine.
104:1543192022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhang F, Huang B, Xu Y, Cao G, Shen M, Liu
C and Luo J: MISP suppresses ferroptosis via MST1/2 kinases to
facilitate YAP activation in non-small cell lung cancer. Adv Sci
(Weinh). 12:e24158142025. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Sun S, Jing X, Tong G, Chen C, Xie S, Wang
C, Chen D, Zhao J, Qi Y, Zhang W, et al: Loss of DDX24 inhibits
lung cancer progression by stimulating IKBKG splicing-mediated
autophagy. Theranostics. 15:1879–1895. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhang L, Lou W and Wang J: Advances in
nucleic acid therapeutics: Structures, delivery systems, and future
perspectives in cancer treatment. Clin Exp Med. 24:2002024.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Li X, Zhang X, Song L, Li Y, Liu A, Li L,
Nešić MD, Li D, Peng L, Wang C and Lin Q: Nanozyme as tumor energy
homeostasis disruptor to augment cascade catalytic therapy. ACS
Nano. 18:34656–34670. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Tsai T, Wu S, Lai Y, Wang H, Hou P, Huang
Y, Chen HH and Su W: CD44-hyaluronan mediating endocytosis of
iron-platinum alloy nanoparticles induces ferroptotic cell death in
mesenchymal-state lung cancer cells with tyrosine kinase inhibitor
resistance. Acta Biomater. 186:396–410. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Shu L, Luo P, Chen Q, Liu J, Huang Y, Wu
C, Pan X and Huang Z: Fibroin nanodisruptor with
ferroptosis-autophagy synergism is potent for lung cancer
treatment. Int J Pharm. 664:1245822024. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Nie Q, Chen W, Zhang T, Ye S, Ren Z, Zhang
P and Wen J: Iron oxide nanoparticles induce ferroptosis via the
autophagic pathway by synergistic bundling with paclitaxel. Mol Med
Rep. 28:1982023. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Li S, Lv J, Li Z, Zhang Q, Lu J, Huo X,
Guo M, Liu X, Li C, Wang J, et al: Overcoming multi-drug resistance
in SCLC: A synergistic approach with venetoclax and
hydroxychloroquine targeting the lncRNA LYPLAL1-DT/BCL2/BECN1
pathway. Mol Cancer. 23:2432024. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Zhao Y, Dhani S, Gogvadze V and
Zhivotovsky B: The crosstalk between SND1 and PDCD4 is associated
with chemoresistance of non-small cell lung carcinoma cells. Cell
Death Discov. 11:342025. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Rao S, Yang H, Penninger JM and Kroemer G:
Autophagy in non-small cell lung carcinogenesis: A positive
regulator of antitumor immunosurveillance. Autophagy. 10:529–531.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Deng J, Lin X, Qin J, Li Q, Zhang Y, Zhang
Q, Ji C, Shen S, Li Y, Zhang B and Lin N: SPTBN2 suppresses
ferroptosis in NSCLC cells by facilitating SLC7A11 membrane
trafficking and localization. Redox Biol. 70:1030392024. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ma J, Zhang P, Wang Y, Lu M, Cao K, Wei S,
Qi C, Ling X and Zhu J: LncRNA HAR1A inhibits non-small cell lung
cancer growth by downregulating c-MYC transcripts and facilitating
its proteasomal degradation. Int Immunopharmacol. 142:1132642024.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Liu Z, Liu C, Fan C, Li R, Zhang S, Liu J,
Li B, Zhang S, Guo L, Wang X, et al: E3 ubiquitin ligase DTX2
fosters ferroptosis resistance via suppressing NCOA4-mediated
ferritinophagy in non-small cell lung cancer. Drug Resist Updat.
77:1011542024. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Zhang L, Xu Y, Cheng Z, Zhao J, Wang M,
Sun Y, Mi Z, Yuan Z and Wu Z: The EGR1/miR-139/NRF2 axis
orchestrates radiosensitivity of non-small-cell lung cancer via
ferroptosis. Cancer Lett. 595:2170002024. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Han F, Chen S, Zhang K, Zhang K, Wang M
and Wang P: Targeting Nrf2/PHKG2 axis to enhance radiosensitivity
in NSCLC. NPJ Precis Oncol. 8:1832024. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Liang J, Bi G, Huang Y, Zhao G, Sui Q,
Zhang H, Bian Y, Yin J, Wang Q, Chen Z and Zhan C: MAFF confers
vulnerability to cisplatin-based and ionizing radiation treatments
by modulating ferroptosis and cell cycle progression in lung
adenocarcinoma. Drug Resist Updat. 73:1010572024. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Ding W, Bao S, Zhao Q, Hao W, Fang K, Xiao
Y, Lin X, Zhao Z, Xu X, Cui X, et al: Blocking ACSL6 compromises
autophagy via FLI1-mediated downregulation of COLs to
radiosensitize lung cancer. Adv Sci (Weinh). 11:e24032022024.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Wen J, Zheng W, Zeng L, Wang B, Chen D,
Chen Y, Lu X, Shao C, Chen J and Fan M: LTF induces radioresistance
by promoting autophagy and forms an AMPK/SP2/NEAT1/miR-214-5p
feedback loop in lung squamous cell carcinoma. Int J Biol Sci.
19:1509–1527. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Lv S, Pan Q, Lu W, Zhang W, Wang N, Huang
L, Li L, Liu J, Ma J, Li Z, et al: Tenovin 3 induces apoptosis and
ferroptosis in EGFR 19del non small cell lung cancer cells. Sci
Rep. 14:76542024. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Lee WC, Moi SH, Yang SF, Tseng HH and Liu
YP: Downregulation of AATK enhances susceptibility to ferroptosis
by promoting endosome recycling in gefitinib-resistant lung cancer
cells. J Pathol. 265:422–436. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Jiang Z, Sun X, Li Y, Wang J, Wang C, Pan
Z and Yang Y: Anlotinib induced ferroptosis through the
p53/xCT/GPX4 pathway in non-small cell lung cancer. Transl Oncol.
53:1022892025. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Huang CY, Chen LJ, Chen CS, Wang CY and
Hong SY: MCL1 inhibition: A promising approach to augment the
efficacy of sorafenib in NSCLC through ferroptosis induction. Cell
Death Discov. 10:1372024. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zhao L, Deng H, Zhang J, Zamboni N, Yang
H, Gao Y, Yang Z, Xu D, Zhong H, van Geest G, et al: Lactate
dehydrogenase B noncanonically promotes ferroptosis defense in
KRAS-driven lung cancer. Cell Death Differ. 32:632–645. 2025.
View Article : Google Scholar :
|
|
132
|
Li N, Zuo R, He Y, Gong W, Wang Y, Chen L,
Luo Y, Zhang C, Liu Z, Chen P and Guo H: PD-L1 induces autophagy
and primary resistance to EGFR-TKIs in EGFR-mutant lung
adenocarcinoma via the MAPK signaling pathway. Cell Death Dis.
15:5552024. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Lyu X, Zeng L, Shi J, Ming Z, Li W, Liu B,
Chen Y, Yuan B, Sun R, Yuan J, et al: Essential role for
STAT3/FOXM1/ATG7 signaling-dependent autophagy in resistance to
icotinib. J Exp Clin Cancer Res. 41:2002022. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Ma C, Zhang X, Mo X, Yu Y, Xiao Z, Wu J,
Ding L, Lei C, Zhu Y and Zhang H: Xie-Bai-San increases NSCLC cells
sensitivity to gefitinib by inhibiting beclin-1 mediated
autophagosome formation. Phytomedicine. 125:1553512024. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Liu Q, Hu J, Li X, Gao H, Kong D and Jin
M: Glutamine transporter inhibitor enhances the sensitivity of
NSCLC to trametinib through GSDME-dependent pyroptosis. Biochem
Pharmacol. 233:1167962025. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Schläfli AM, Tokarchuk I, Parejo S, Jutzi
S, Berezowska S, Engedal N and Tschan MP: ALK inhibition activates
LC3B-independent, protective autophagy in EML4-ALK positive lung
cancer cells. Sci Rep. 11:90112021. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Yang Q, Sun K, Gao T, Gao Y, Yang Y, Li Z
and Zuo D: SIRT1 silencing promotes EMT and Crizotinib resistance
by regulating autophagy through AMPK/mTOR/S6K signaling pathway in
EML4-ALK L1196M and EML4-ALK G1202R mutant non-small cell lung
cancer cells. Mol Carcinog. 63:2133–2144. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Wu L, Liu J, Wang S, Bai M, Wu M, Gao Z,
Li J, Yu J, Liu J and Meng X: Negative correlation between
18F-RGD uptake via PET and tumoral PD-L1 expression in
non-small cell lung cancer. Front Endocrinol (Lausanne).
13:9136312022. View Article : Google Scholar
|
|
139
|
Gou Q, Che S, Chen M, Chen H, Shi J and
Hou Y: PPARγ inhibited tumor immune escape by inducing PD-L1
autophagic degradation. Cancer Sci. 114:2871–2881. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Young MJ, Wang SA, Chen YC, Liu CY, Hsu
KC, Tang SW, Tseng YL, Wang YC, Lin SM and Hung JJ: USP24-i-101
targeting of USP24 activates autophagy to inhibit drug resistance
acquired during cancer therapy. Cell Death Differ. 31:574–591.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Yuan Z, Yu T, Wang X, Meng K, Wang T, Wang
B, Xi Y, Wang C, Zeng C, Hu S, et al: Glutamine deprivation confers
immunotherapy resistance by inhibiting IFN-γ signaling in cancer
cells. Pharmacol Res. 213:1076432025. View Article : Google Scholar
|
|
142
|
Luo L and Xu G: Fascaplysin induces
apoptosis and ferroptosis, and enhances anti-PD-1 immunotherapy in
non-small cell lung cancer (NSCLC) by promoting PD-L1 expression.
Int J Mol Sci. 23:137742022. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Ke ZH, Chen Y, Yu T, Zhang Q, Xiang Y and
Lu KH: LRP1B suppresses immunotherapy efficacy in lung
adenocarcinoma by preventing ferroptosis. Cancer Med.
13:e704862024. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Diao JD, Jia YY, Dai EY, Liu J, Kang R,
Tang D, Han L, Zhong Y and Meng L: Ferroptotic therapy in cancer:
benefits, side effects, and risks. Mol Cancer. 23:892024.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Zhao X, Hu S, Zeng L, Liu X, Song Y, Zhang
Y, Chen Q, Bai Y, Zhang J, Zhang H, et al: Irradiation combined
with PD-L1−/− and autophagy inhibition enhances the
antitumor effect of lung cancer via cGAS-STING-mediated T cell
activation. iScience. 25:1046902022. View Article : Google Scholar
|
|
146
|
Li C, Zhao W, Geng D, Jin Y and Guan W:
Targeting the interplay of cGAS-STING and ferroptosis by
nanomedicine in the treatment of cancer. J Exp Clin Cancer Res.
44:2492025. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Ma L, Chen C, Zhao C, Li T, Ma L, Jiang J,
Duan Z, Si Q, Chuang TH, Xiang R and Luo Y: Targeting carnitine
palmitoyl transferase 1A (CPT1A) induces ferroptosis and synergizes
with immunotherapy in lung cancer. Signal Transduct Target Ther.
9:642024. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Wang J, Liu Z, Hu T, Han L, Yu S, Yao Y,
Ruan Z, Tian T, Huang T, Wang M, et al: Nrf2 promotes progression
of non-small cell lung cancer through activating autophagy. Cell
Cycle. 16:1053–1062. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Gardner EE, Earlie EM, Li K, Thomas J,
Hubisz MJ, Stein BD, Zhang C, Cantley LC, Laughney AM and Varmus H:
Lineage-specific intolerance to oncogenic drivers restricts
histological transformation. Science. 383:eadj14152024. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Liu D, Wu L, Xu X, Yang S, Liu M, Hu M,
Ren S and Xu Y: Regimens combining radiation and immunotherapy for
cancer: Latest updates from 2024 ASCO annual meeting. J Hematol
Oncol. 17:842024. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Cheng Y, Bai L and Cui J: Harnessing
multi-omics approaches to decipher tumor evolution and improve
diagnosis and therapy in lung cancer. Biomark Res. 13:1402025.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Hsu CY, Askar S, Alshkarchy SS, Nayak PP,
Attabi KAL, Khan MA, Mayan JA, Sharma MK, Islomov S and Soleimani
Samarkhazan H: AI-driven multi-omics integration in precision
oncology: Bridging the data deluge to clinical decisions. Clin Exp
Med. 26:292025. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Li X, Yin D, Geng J, Xu Y, Xu Z, Yang X,
Li Q, Shang Z, Yang Z, Xu Z, et al: Machine learning-driven
multi-omics analysis identifies a prognostic gene signature
associated with programmed cell death and metabolism in
hepatocellular carcinoma. Biol Proced Online. 27:292025. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Kale M, Wankhede N, Pawar R, Ballal S,
Kumawat R, Goswami M, Khalid M, Taksande B, Upaganlawar A, Umekar
M, et al: AI-driven innovations in Alzheimer's disease: Integrating
early diagnosis, personalized treatment, and prognostic modelling.
Ageing Res Rev. 101:1024972024. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Yu G and Gong J: Targeting CSC-immune cell
crosstalk to overcome chemoresistance and enhance immunotherapy
efficacy. Front Immunol. 16:16208072025. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Fang C, Zhou P, Zhang X, He Y and Yang Q:
Artificial intelligence in oncology drug development and
management: A precision medicine perspective. Front Oncol.
15:16098272025. View Article : Google Scholar : PubMed/NCBI
|














