The functional diversity of the liver arises from
its spatially organized zonation architecture, where hepatocytes
progressively alter their metabolic and detoxification capacities
along the porto-central axis (1). This intricate partitioning,
sustained by blood-borne molecular gradients and intercellular
crosstalk, enables the liver to balance antagonistic biochemical
processes while maintaining systemic homeostasis (2). Paradoxically, this structural
stability is reconfigured during severe liver injury. Hepatocyte
necrosis disrupts zonal organization, which both limits remaining
liver function and triggers repair responses. Recent spatial
biology studies show that the liver adopts different regenerative
strategies depending on context, ranging from zonal self-renewal to
whole-lobule plasticity (3-5).
These processes are highly influenced by local microenvironmental
signals. New technologies have begun to reveal how surviving cells
use positional cues to rebuild functional structures.
Based on these studies and emerging technologies,
the concept of 'spatial hepatology' has gradually emerged. This
approach involves integrating multi-omics techniques in the context
of tissue in situ, such as spatial transcriptomics, spatial
proteomics and spatial metabolomics, to systematically analyze the
distribution, functional states and interactions of different cell
types at specific spatial locations within the liver (2). Compared with conventional
single-cell sequencing or bulk transcriptomic analyses, spatial
omics preserves tissue architecture while coupling molecular
information with spatial coordinates, enabling precise mapping of
functional liver regions and revealing dynamic molecular gradients
in local microenvironments. This strategy offers significant
advantages for investigating the remodeling of metabolic zonation,
heterogeneity of injury responses and the regulation of cell fate
during regeneration in liver disease, thereby providing a more
refined and dynamic framework for understanding the pathogenesis
and progression of liver disorders (4).
The liver lobule exhibits a clear functionally
compartmentalized structure. Traditional approaches to
characterizing liver zonation and metabolic compartmentalization
include quantitative PCR, immunohistochemistry and in situ
hybridization (6). These methods
reveal spatial differences in protein expression and enzymatic
activity between periportal and pericentral hepatocytes, forming
the basis of metabolic zonation. This spatial distribution arises
from sinusoidal blood flow, which generates gradients of oxygen
tension, nutrient concentrations and xenobiotics along the
porto-central axis (7). These
gradients of these substances are key determinants of liver
zonation.
Studies on zonal metabolic features show that the
periportal region predominantly carries out β-oxidation, urea
synthesis and gluconeogenesis, whereas the pericentral region is
primarily responsible for glycolysis, lipogenesis, and
detoxification processes (8-10). Although most metabolic processes
exhibit strict zonal segregation and serve as key indicators of
liver function, some pathways can operate independently in
different zones while remaining coordinated (11). A typical example is ammonia
metabolism; studies have shown that periportal hepatocytes
efficiently remove ammonia through glutaminase and urea cycle
enzymes, whereas pericentral hepatocytes detoxify residual ammonia
via glutamine synthetase (12,13). This spatial division of labor
ensures high metabolic efficiency. However, this compartmentalized
organization also concentrates cytochrome P450 activity in the
pericentral region, making it more susceptible to toxin-induced
necrosis (14).
Notably, liver zonation is not a static structure
but exhibits high dynamic plasticity through coordinated
transcriptional regulation and structural remodeling (15). Nutritional status is a key driver
of this dynamic change. For example, fasting enhances lobular
gluconeogenesis and promotes the extension of β-oxidation,
typically dominant in the periportal region, toward the midlobular
zone to meet systemic energy demands (16,17). During pregnancy, this adaptation
is further reinforced by the expansion of periportal gluconeogenic
hepatocytes, thereby maintaining maternal-fetal glucose homeostasis
(18). Together, these
regulatory mechanisms indicate that the liver can dynamically
reorganize its spatial architecture in response to physiological
states, enabling precise control of metabolic function.
Recent advances in single-cell sequencing and
multi-omics have improved the current understanding of liver
zonation (19,20). These approaches define metabolic
gradients, transcriptional networks and chromatin accessibility
across hepatocyte populations, refining traditional models of
lobular organization (21).
Evidence shows that liver zonation is not simply binary; >80% of
hepatocyte genes display continuous expression gradients along the
porto-central axis (19). Based
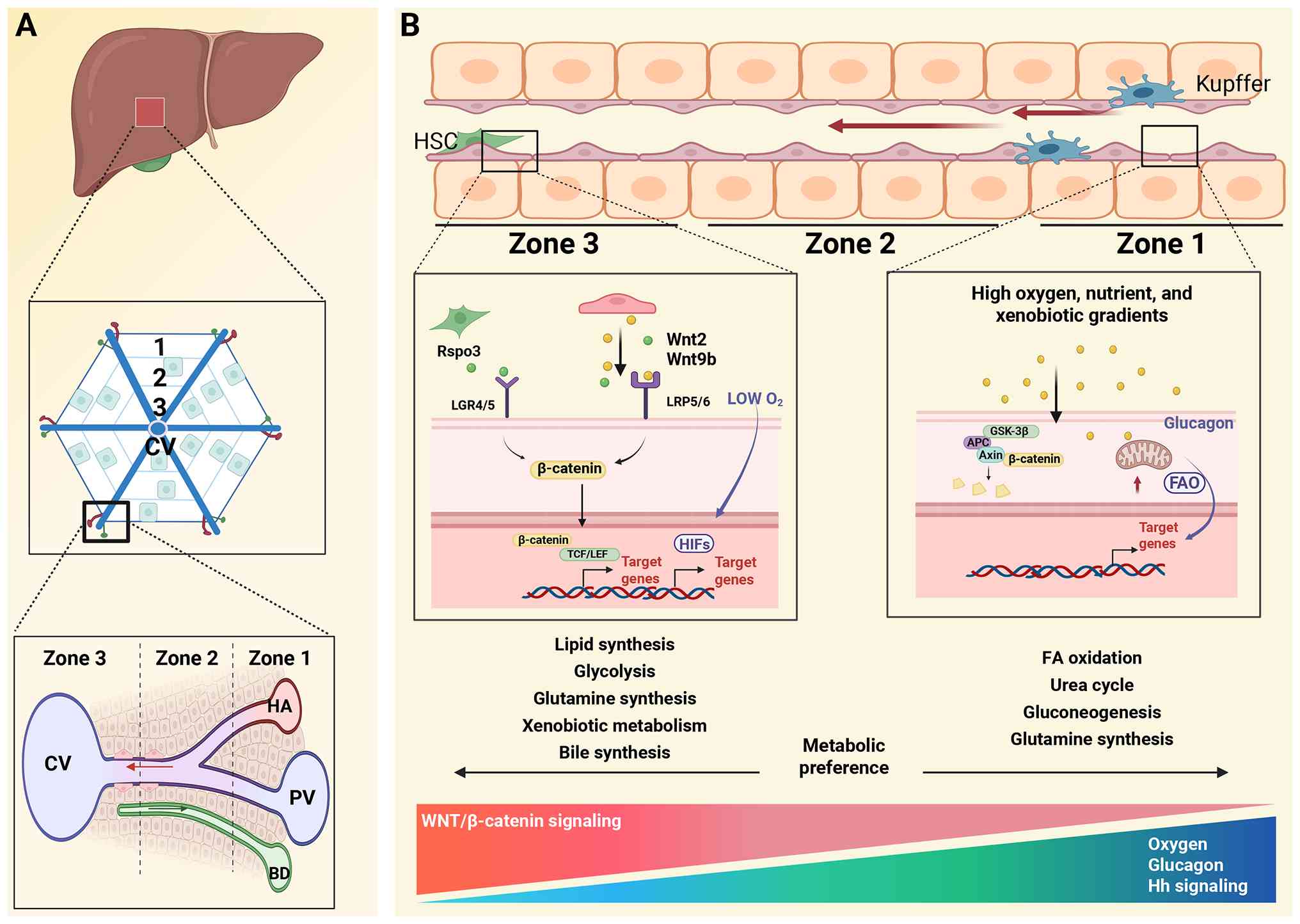
on this, the liver lobule is commonly divided into three zones: i)
Periportal (zone 1); ii) midlobular (zone 2); and iii) pericentral
(zone 3) (2). Higher-resolution
models further divide the lobule into nine layers, corresponding to
these three regions. Zone-specific markers have also been
identified. Periportal markers include ASS1, Cyp2f2, Sds, Asl, Gls2
and Arg1. Midlobular markers include hepcidin antimicrobial peptide
2 (Hamp2) and Insulin-like growth factor-binding protein 2
(Igfbp2). Pericentral markers include cytochrome P450 2E1 (Cyp2e1),
Cyp1a2, GS and Oat (Table I)
(2,19).
Trajectory analyses of liver development have
revealed the dynamic establishment of metabolic gradients. For
example, hepatocytes in neonatal mice initially exhibit pericentral
metabolic features, with zonation patterns progressively maturing
after birth (22). This
developmental plasticity is closely associated with hepatocyte
polyploidization. Regarding the spatial distribution of diploid
hepatocytes, early studies suggested that these cells are enriched
in the periportal and decline along the lobular axis, implying a
higher proliferative potential in this zone (23,24). However, subsequent investigations
employing alternative lineage-tracing strategies have reported a
more uniform distribution, with some even indicating relative
enrichment in the pericentral zone (25,26). These discrepancies may reflect
differences in species, age-related shifts in polyploidization and
methodological biases in nuclear isolation. Future studies
integrating spatially resolved single-cell omics will be essential
to clarify the interplay between ploidy status, zonation
characteristics and regenerative capacity. However, spatial
specialization is not unique to hepatocytes. Non-parenchymal cell
populations also exhibit pronounced zonal heterogeneity, which may
be a key factor in maintaining hepatic zonation (27-29). Non-parenchymal cells distributed
across different lobular regions likely perform context-dependent
functions, contributing to region-specific regulation during
disease progression and influencing regenerative processes. By
integrating single-cell atlases with spatial transcriptomics,
researchers have progressively constructed high-resolution maps of
both hepatocyte and non-parenchymal cell responses during injury
and regeneration, providing a theoretical foundation for
understanding liver repair mechanisms (30).
Liver zonation is established through the
coordinated action of systemic and local microenvironmental
signals. Circulating factors, including hormonal gradients, oxygen
tension and nutrient availability, activate spatially specific
signaling pathways to establish metabolic compartmentalization.
Among these, the Wnt/β-catenin pathway serves a central role, while
other signaling networks, including Hedgehog (Hh) and
oxygen-sensing pathways such as HIF, further coordinate epigenetic
remodeling and metabolic reprogramming, thereby restricting
hepatocyte functions to specific regions within the liver lobule
(Fig. 1) (31-33).
The oxygen gradient along the porto-central axis of
the hepatic lobule establishes a hypoxic microenvironment that
activates hypoxia-inducible factors (HIFs) (33). Among these, HIF-1α is
preferentially stabilized in pericentral hepatocytes, where it
directly regulates glycolytic gene programs through binding to
hypoxia-responsive elements (HREs) (33). This includes upregulation of
glucose transporters GLUT1 (SLC2A1) and GLUT3 (SLC2A3), as well as
key rate-limiting glycolytic enzymes such as HK2, PFK1, PKM2 and
LDHA. Concurrently, HIF-1α transcriptionally activates PDK1,
thereby inhibiting pyruvate entry into mitochondrial oxidative
metabolism, collectively reinforcing the glycolytic phenotype
characteristic of zone 3 hepatocytes (34,35). By contrast, HIF-2α primarily
governs antioxidant defense and lipid metabolic regulation,
directly inducing genes such as SOD2 and HMOX1 (36). However, sustained activation of
HIF-2α can impair lipid homeostasis by transcriptionally
suppressing PPARα-dependent fatty acid oxidation (FAO) genes,
including CPT1A and ACOX1, thereby promoting steatosis (37). HIF-3α splice variants function as
dominant-negative regulators by competing with HIF-1α for HRE
binding or forming low-activity heterodimers, ultimately
attenuating HIF-1α signaling (38,39).
Beyond oxygen gradients, the spatial distribution of
hormonal signals represents another key regulatory axis shaping
liver zonation. Although glucagon receptors are uniformly
distributed across the hepatic lobule, glucagon itself forms a
functional gradient along the porto-central axis (40). In glucagon-deficient mice,
expression of the periportal marker gene Gls2 is markedly reduced,
whereas the pericentral marker Glul, which is normally restricted
to 1 to 3 hepatocyte layers surrounding the central vein, expands
to 5 to 6 layers (41). This
abnormal pattern can be reversed by exogenous glucagon
supplementation. Functionally, glucagon differentially regulates
periportal urea cycle enzymes and pericentral xenobiotic
metabolizing enzymes, such as members of the Cyp450 family. In
addition, sexual dimorphisms further increases regulatory
complexity. The sex-specific patterns of growth hormone (GH)
secretion directly influence liver zonation by regulating the
activity of transcription factors in distinct regions. In males,
pulsatile GH in zone 1 activates SREBF1, promoting
gluconeogenesis-related pathways, whereas in females, continuous GH
in zone 3 supports TRIM24, enhancing hepatic progenitor
cell-related pathways (42).
These differential transcriptional programs restrict specific
hepatocyte functions to defined zones, creating a sex-biased
metabolic and regenerative landscape (42). Continuous GH infusion in males
can abolish this zonal dimorphism, indicating that the mode of
hormone secretion determines sex-specific liver zonation (43). These findings indicate that liver
zonation functions as a dynamic interface integrating metabolic,
endocrine and oxygen-dependent signals.
The Wnt/β-catenin pathway is a key regulator of
pericentral hepatocyte identity (44). Its activity depends on tight
regulation of the β-catenin destruction complex and phosphorylation
events. Under basal conditions, β-catenin is continuously degraded
by a destruction complex composed of Axin, adenomatous polyposis
coli (APC), CK1 and GSK3β. CK1 initiates phosphorylation at Ser45,
followed by sequential phosphorylation by GSK3β at Thr41, Ser37 and
Ser33, which marks β-catenin for recognition by the β-TrCP E3
ubiquitin ligase and subsequent proteasomal degradation (45). Upon Wnt stimulation, including
Wnt2 and Wnt9b secreted by central vein endothelial cells, the
binding of these ligands to Frizzled receptors and the co-receptors
LRP5/6 recruits CK1 and GSK3β to the membrane (46,47). This leads to phosphorylation of
LRP5/6, inhibition of GSK3β activity and inactivation of the
destruction complex (44).
Stabilized β-catenin accumulates and translocates into the nucleus,
where it binds T-cell factor/lymphoid enhancer-binding factor
transcription factors to activate pericentral gene programs such as
glutamine synthetase and Cyp2e1.
A critical determinant of zonal regulation is the
spatially restricted expression of APC (48). APC is largely absent in the
pericentral zone, permitting sustained nuclear accumulation of
β-catenin, whereas its high expression in periportal hepatocytes
constrains Wnt signaling (48).
Genetic studies demonstrate that loss of APC leads to panlobular
activation of β-catenin, accompanied by ectopic expression of
pericentral markers such as Cyp2e1 in periportal hepatocytes
(49). In addition, oxygen
gradients modulate this axis: HIF-1α suppresses APC transcription
in the pericentral zone, thereby enhancing β-catenin signaling,
while reactive oxygen species in the periportal region stabilize
APC, establishing a redox-dependent zonal boundary (50,51). Furthermore, Wnt5a in the
periportal region antagonizes β-catenin activity through
calcium-dependent signaling, which reinforces zone 1 identity
(52). The Ras/MAPK/ERK pathway
contributes additional modulation by promoting β-catenin
phosphorylation and proteasomal degradation in periportal
hepatocytes, thereby balancing regional signaling intensity
(53).
Non-canonical Wnt pathways further refine zonal
patterning. Hypoxia-induced Wnt11 in zone 3 activates planar cell
polarity signaling via the ROR2 receptor, regulating pericentral
gene expression independently of β-catenin (54-56). Wnt signaling defines hepatic
zonation through coordinated regulation of the β-catenin
destruction complex, spatial control of APC expression and
integration of canonical and non-canonical pathways. It also
interacts with multiple signaling networks to precisely specify
hepatocyte identity along the porto-central axis.
The Hh signaling pathway is also a key regulator of
hepatic metabolic zonation. It coordinates functional
specialization along the porto-central axis through spatially
restricted ligand-receptor interactions (57). Spatial transcriptomics analyses
have shown that Indian Hh is enriched in the pericentral region,
forming a morphogen gradient that is inversely correlated with
Wnt/β-catenin signaling activity in the periportal zone (31). This spatial relationship is
maintained through GLI-mediated inhibition of β-catenin
transcriptional activity, establishing a dynamic balance between
the two pathways (58). The Hh
gradient can also regulate components of the insulin-like growth
factor axis (IGF-I/IGFBP-1) to coordinate zonal metabolic
programming (59). Pericentral
hepatocytes exhibit GLI-dependent upregulation of gluconeogenic
enzymes, whereas periportal regions show enhanced lipid oxidation
(32). Furthermore, hepatic
stellate cells (HSCs) in the space of Disse act as spatial signal
integrators; they convert Hh/Wnt crosstalk into extracellular
matrix (ECM) remodeling, thereby physically maintaining the
structure of metabolic zonation (60).
Emerging evidence indicates that the spatially
restricted cellular niches of non-parenchymal cells, particularly
the periportal vs. pericentral zonation patterns, establish
microenvironments that directly modulate hepatocyte functional
specialization via ECM regulation, metabolic exchange and immune
surveillance, thereby driving the establishment of hepatic
metabolic zonation (61). Among
these, liver sinusoidal endothelial cells (LSECs) act as central
regulators of zonation. Central vein associated LSECs selectively
secrete Wnt2, Wnt9b and R-spondin 3, thereby activating β-catenin
signaling in adjacent hepatocytes and maintaining pericentral cell
identity (47,62). Consistently, combined deletion of
Wnt2 and Wnt9b abolishes zone 3 specific gene expression. In
addition, LSECs modulate sinusoidal blood flow and oxygen tension
through the secretion of vascular endothelial growth factors,
contributing to the establishment of the porto-central metabolic
axis (63). Kupffer cells
exhibit a preferential periportal distribution, forming an immune
zonation pattern (29). By
restricting infiltrating neutrophils to the periportal region, they
spatially confine pro-inflammatory responses, thereby protecting
the metabolically vulnerable pericentral hepatocytes. Disruption of
Kupffer cell localization leads to bacterial dissemination and
panlobular inflammatory injury (64-65). HSCs also display zonal
heterogeneity, comprising functionally distinct subpopulations
(66). Periportal-associated
stellate cells express markers such as Ngfr and Tagln and are
involved in rapid immune interactions, whereas
pericentral-associated stellate cells specifically express Rspo3,
which enhances Wnt signaling to sustain pericentral hepatocyte
homeostasis (67,68).
Beyond extrinsic cues, hepatic zonation is also
governed by intrinsic epigenetic programs (69). DNA methylation has been shown to
form a gradient along the porto-central axis, progressively
decreasing from the periportal to the pericentral zone. Key
transcription factors exhibit spatially restricted expression
patterns; USF1 is enriched in periportal hepatocytes, where it
exerts negative feedback on periportal-specific genes while
modulating lipid and glucose metabolism (70). By contrast, Nr2f2 is
progressively enriched in the pericentral zone and activates
pericentral identity genes during hepatocyte maturation (69,71). Although HNF4α is uniformly
expressed, its DNA-binding activity is gated by methylation status,
and its deletion leads to ectopic expression of pericentral genes
in periportal hepatocytes (71,72).
Histone modifications further reinforce zonal
specialization. H3K27ac, a histone acetylation marker of active
enhancers, is enriched at regulatory elements of
Wnt/β-catenin-associated pericentral metabolic genes, whereas
H3K4me3, a histone trimethylation marker of active promoters,
predominantly labels genes involved in periportal metabolic
pathways such as gluconeogenesis and the urea cycle (73). The SWI/SNF chromatin remodeling
component ARID1A maintains hepatocyte metabolic gene programs by
regulating nucleosome positioning and chromatin accessibility
(74); hepatocyte-specific loss
of ARID1A reduces accessibility at metabolic loci, impairs binding
of transcription factors such as PPARα and suppresses the
expression of genes involved in fatty acid β-oxidation (75).
MicroRNAs (miRs) also provide an additional layer of
fine-tuning. For example, liver-enriched miR-122 is required for
the maintenance of zonation, and its deletion results in expansion
of zone 3 characteristics (76,77). Collectively, DNA methylation,
histone modifications, non-coding RNAs and transcription factor
dynamics converge to form an integrated epigenetic-metabolic axis
that spatially encodes hepatocyte identity.
The spatial compartmentalization of hepatic function
constitutes a critical defense mechanism against metabolic stress.
Within this organizational framework, the Wnt/β-catenin pathway
serves a central role, not only in maintaining pericentral
hepatocyte identity but also in regulating hepatocyte turnover.
Early lineage-tracing studies proposed that Axin2+
hepatocytes located in the pericentral possess stem-like properties
and contribute to long-term hepatocyte renewal under homeostatic
conditions (78). However,
accumulating evidence suggests that the contribution of these cells
may have been overestimated due to labeling artifacts and
signal-dependent activation of reporter systems in Axin2-Cre models
(79,80). Multiple independent studies
indicate that hepatocyte renewal occurs broadly throughout the
liver lobule rather than being restricted to a discrete pericentral
progenitor population (81-83). Accordingly, Axin2+
hepatocytes are more likely to represent a Wnt responsive
hepatocyte subset primarily responsible for maintaining zonal
identity, rather than serving as a universal stem cell reservoir,
although they may acquire regenerative potential under specific
injury contexts (80). A similar
paradigm applies to Leucine-rich repeat-containing
G-protein-coupled receptor 5-positive (Lgr5+)
hepatocytes. These cells are exceedingly rare in the healthy liver
but can be induced following injury (84). Under physiological conditions,
the nuclear receptor farnesoid X receptor (FXR) maintains
Lgr5+ hepatocytes in a quiescent state; by contrast,
metabolic stress or tissue injury activates a PPARα-dependent
proliferative program, enabling their transient participation in
tissue repair (85). Notably,
aberrant activation of Lgr5 may compromise cellular identity and
contribute to tumor progression.
Based on the principles of zonated hepatocyte
renewal, pregnancy induces a spatiotemporally coordinated
remodeling of the liver. Hepatocyte proliferation progresses in a
zonal sequence, initiating in the periportal zone 1 during early
pregnancy, shifting to zone 2 in mid-gestation and culminating in
pericentral zone 3 expansion in late pregnancy. This dynamic
pattern parallels the progressive increase in fetal metabolic
demands (20). Lineage tracing
studies indicate that estrogen coordinates this hierarchical zonal
program by selectively promoting the proliferation of Ccnd1
positive hepatocytes in zone 2, whereas prolactin fine-tunes liver
mass through bile acid mediated hypertrophic regulation (90,91). This spatial proliferation program
is closely aligned with metabolic compartmentalization. Expansion
in zones 1 and 2 supports glucose and cofactor synthesis required
for fetal growth, while proliferation in zone 3 enhances
detoxification capacity to counteract pregnancy associated
oxidative stress (90,91). Despite these advances, critical
gaps remain in understanding how maternal liver zonation adapts and
synchronizes with key milestones of fetal development. This
highlights the need to elucidate how the placenta liver signaling
axis preserves zonation fidelity during pregnancy.
The spatial erosion of metabolic zonation is a
hallmark of liver pathology, where hepatocyte loss disrupts lobular
signaling topography and initiates self-amplifying injury circuits
(Fig. 2). Notably, zone-specific
hepatocyte necrosis, whether periportal oxidative stress or
pericentral toxin accumulation, triggers localized inflammatory
cascades that recruit neutrophils and monocytes (92,93). This inflammatory feed-forward
loop not only exacerbates regional damage but also propagates
zonation collapse.
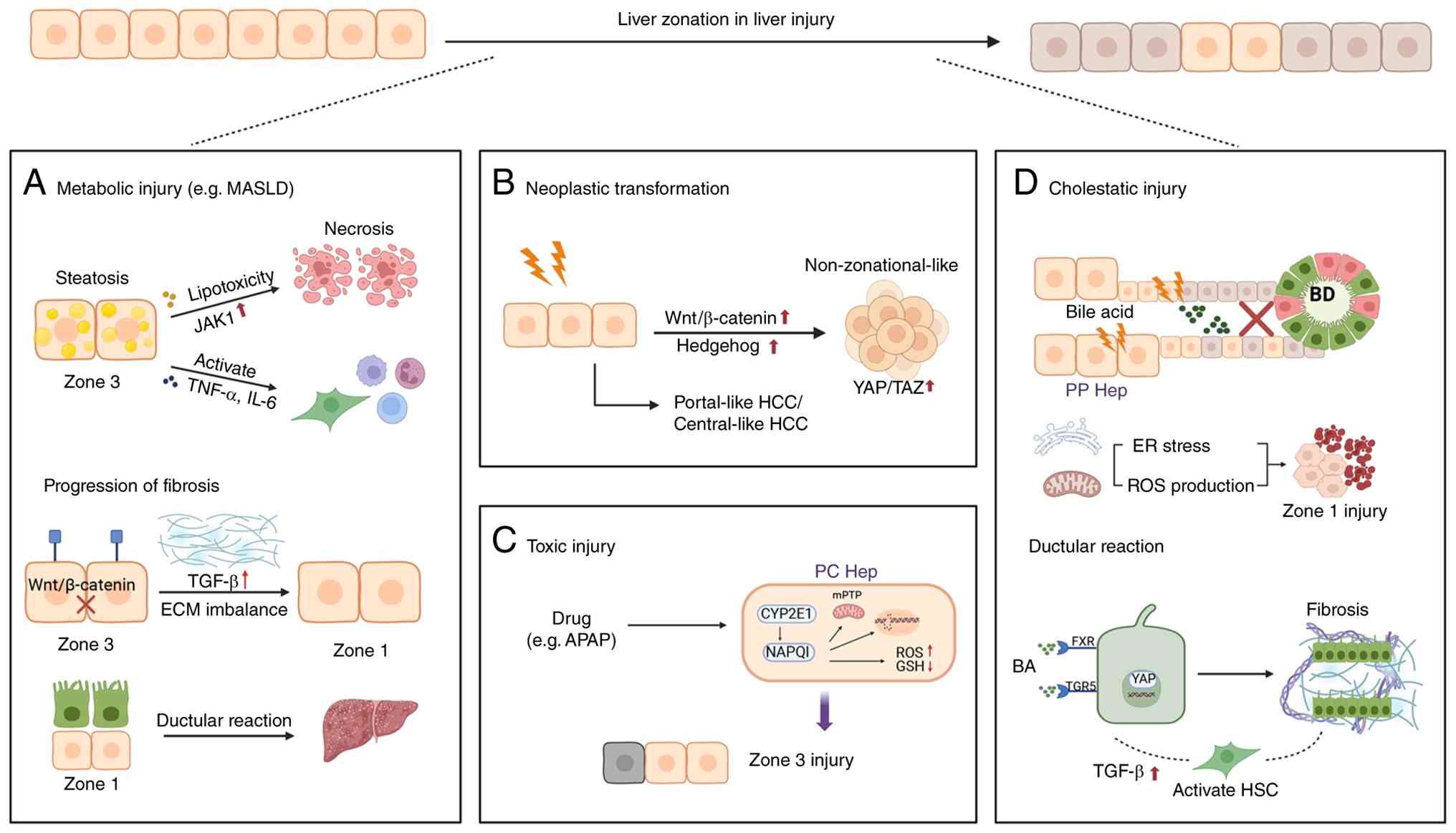
Metabolic dysfunction associated steatotic liver
disease (MASLD) is a liver disorder linked to metabolic dysfunction
and is characterized by a systemic collapse of lobular metabolic
zonation architecture (94).
This pathological cascade is driven by coordinated dysregulation of
the three core metabolic processes, uptake, synthesis and export,
ultimately disrupting the signaling gradients required to maintain
functional compartmentalization (95). Hepatic steatosis, a hallmark
feature of MASLD, shows a developmentally and spatially patterned
distribution. In adult patients, lipid accumulation typically
initiates in zone 3, the primary site of fatty acid synthesis
(96). Lipotoxicity in this
region triggers zone 3-predominant hepatocyte apoptosis and
pericentral fibrosis (97). As
the disease progresses to metabolic dysfunction-associated
steatohepatitis (MASH), the spatial pattern of fibrosis is
remodeled, with collagen deposition shifting toward zone 1 and
forming porto-central bridging fibrosis, a key precursor of
cirrhosis (98). But prepubertal
MASLD exhibits an opposite spatial pattern, characterized by zone
1-dominant steatosis and periportal fibrosis without pericentral
involvement, which is associated with developmental metabolic
programming and microenvironment-specific immune responses
(99).
Dysregulated zonal signaling is a key determinant of
disease severity. Elevated Hh ligand levels and yes-associated
protein (YAP) activation are associated with steatohepatitis and
fibrosis progression, while Wnt/β-catenin-mediated pericentral
homeostasis is disrupted (100,101). Kupffer cells, traditionally
localized to the periportal region, amplify zonation-restricted
inflammation in early MASLD by recruiting neutrophils and releasing
pro-fibrotic cytokines, thereby exacerbating zone 1 injury
(102). Meanwhile, periportal
ductular reactions characterized by hepatic progenitor cell
expansion may contribute to porto-portal fibrosis, although their
direct pro-fibrotic role remains to be fully validated (103,104). Mechanistically, early MASLD
retains lobular zonation, with preserved regional gene expression
and DNA methylation patterns, suggesting a therapeutic window for
zonation restoration (71). This
reversibility highlights the potential of targeting zone-specific
drivers, including Kupffer cell-mediated spatial inflammation,
HSC-derived ECM imbalance and liver sinusoidal endothelial
cell-driven Wnt signaling dysregulation, to block the
steatosis-fibrosis axis (105-107).
HCC arises from zonation-collapsed microenvironments
in cirrhotic livers, with chronic hepatitis and metabolic
dysfunction driving spatially biased oncogenesis (108). It is reported that >50% of
HCCs exhibit constitutive Wnt/β-catenin activation, a pathway
central to pericentral zonation, alongside Hh signaling
hyperactivation, constitutive YAP/TAZ activation and glycolytic
reprogramming, collectively hijacking zonation-maintaining circuits
to fuel tumor initiation (109,110). While HCC cellular origins
remain debated, clonal tracing reveals that regeneration-competent
hepatocytes within injury niches serve as dominant precursors,
acquiring malignant traits through niche-derived pressures
(83,111).
Hypoxia-mediated metabolic zonation collapse further
shapes HCC evolution. Pericentral hypoxia stabilizes HIF-1α,
synergizing with β-catenin to select for venous zone-adapted clones
with glutamine addiction and invasive potential (112). Notably, HCCs retain
zonation-like phenotypic diversity, as portal-like HCC with low
β-catenin expression mirrors periportal metabolism, exhibiting
indolent behavior and favorable post-resection outcomes due to
preserved immune surveillance. However, central-like HCC
recapitulates pericentral features with high β-catenin, marked by
immune-cold microenvironments and resistance to checkpoint
blockade, yet maintains vulnerability to Wnt/Hh-targeted therapies
(108). This zonation-based
classification system overcomes the limitations of conventional
histopathological subtyping and enables prognostic stratification
as well as compartment-specific therapeutic strategies (113). From a therapeutic perspective,
restoring Wnt signaling gradients or targeting the
hypoxia-HIF-β-catenin axis may reverse treatment resistance driven
by spatial dedifferentiation. In addition, portal vein-like HCC may
benefit from immunomodulatory approaches that exploit residual
zonal differentiation integrity (114,115).
DILI, exemplified by acetaminophen (APAP) and carbon
tetrachloride models, manifests zonation-biased damage, as
demonstrated by the studies that pericentral hepatocytes (zone 3)
with high cytochrome P450 activity (including Cyp2e1) metabolize
toxins into reactive intermediates such as NAPQI, triggering
oxidative stress and mitochondrial dysfunction that culminate in
zone 3-predominant necrosis (116-118). Spatial reprogramming at the
injury border initiates repair where damage-adjacent hepatocytes
transiently reactivate fetal genes and upregulate ribosomes and
proteasomes, enabling proteome remodeling to adopt pericentral
identities (82). Concurrently,
zonation-restricted regeneration unfolds where surviving
pericentral hepatocytes initiate proliferation via mTOR signaling,
followed by zone 2 hepatocyte expansion (3). A recent study demonstrated zone
2-derived hepatocytes as a main contributor to compartmentalized
replenishment, where Igfbp2+ hepatocytes migrate toward
necrotic zone 3 and restore metabolic zonation (89).
Experimental cholestasis models, including DDC diet
and bile duct ligation, precisely recapitulate human
cholangiopathies while unveiling zonation-defined injury patterns
(123). Cholestatic injury
predominantly targets zone 1 hepatocytes, as bile acids first
accumulate in the periportal region following impaired biliary
excretion. Hydrophobic bile acids induce membrane damage through
detergent-like effects, disrupt mitochondrial function, and trigger
oxidative stress and endoplasmic reticulum stress. In parallel,
bile acid-activated death receptor signaling, including Fas and
TRAIL pathways, further amplifies hepatocyte injury. Zone 1
hepatocytes are particularly susceptible due to their high
expression of bile acid uptake transporters such as NTCP and OATPs,
together with limited adaptive capacity for detoxification compared
with downstream zones (124).
In addition, cholestasis induces inflammatory signaling from portal
stromal and immune cells, further reinforcing periportal injury and
ECM remodeling.
Mechanistically, periportal hepatocytes exposed to
cholestatic stress can establish a self-limiting reparative
microenvironment. Within this setting, a distinct subpopulation of
hepatocytes with a unique phenotype has been identified in the
periportal niche. These cells express multiple biliary-enriched
genes, including low levels of Sox9, while retaining the canonical
hepatocyte marker HNF4α. This population, termed hybrid hepatocytes
(HybHPs), is capable of contributing to parenchymal regeneration
through proliferative expansion (125). However, the origin of HybHPs
remains controversial. In chronic injury models, HybHPs have been
proposed to arise from embryonic progenitors located in the ductal
plate, as these cells can upregulate biliary markers and
differentiate into cholangiocytes. However, studies using partial
hepatectomy models suggest that HybHPs originate from mature
hepatocytes, as they appear as early as 3 h after resection without
evidence of early activation of hepatic progenitor cells (126). Furthermore, in MASLD models,
analysis of fetal liver markers indicates that these biphenotypic
cells do not undergo dedifferentiation or redifferentiation
(127). Taken together, these
findings suggest that HybHPs most likely arise from mature
hepatocytes undergoing microenvironment-driven transdifferentiation
in response to alterations in the periportal niche during injury,
rather than from stem cell populations or developmental
progenitors. In addition, chronic cholestasis activates
cholangiocyte progenitor cells through the FXR/TGR5 signaling
pathway, which in turn induces collagen deposition from HSCs,
leading to periportal fibrosis and consequently disrupting the
architecture of zone 1 (128).
The regenerative capacity of the liver is deeply
rooted in its metabolically zonated architecture. Early
lineage-tracing studies suggested that Mfsd2a positive zone 1
hepatocytes migrate toward the pericentral region (129); however, more recent evidence
identifies Igfbp2 positive and Hamp2 positive zone 2 hepatocytes as
the primary drivers of regeneration. Their proliferative advantage
is mediated by mTOR-CCND1 signaling, a mechanism conserved with
pregnancy-induced liver growth (30). In parallel, Sox9 positive
hepatocytes derived from the periportal microenvironment acquire
progenitor-like features during regeneration, recapitulating
biliary repair programs and underscoring the dual role of zone 1 in
metabolic function and regenerative adaptation (130,131). Zone 1 mobilizes hepatic
progenitor cells and biliary transdifferentiation to compensate for
parenchymal loss, whereas zone 3 recruits Axin2+,
Lgr5+ and Cyp2e1+ hepatocytes to support
pericentral repair (132). This
hierarchical strategy ensures preservation of metabolic zonation
during tissue reconstruction. Importantly, this system operates
through interzonal crosstalk, with zone 2 derived Igfbp2 promoting
activation of zone 1 progenitor responses, while zone 3 Wnt
signaling suppresses ectopic biliary reactions (133).
In addition to the hepatocyte subpopulations with
regenerative potential as aforementioned, multiple liver injury
models have shown that hepatocytes at the injury interface can
dedifferentiate from mature hepatocytes into progenitor-like cells
(118). Beyond the
well-characterized HNF4α+Sox9+ population,
these also include AFP+ reprogrammed hepatocytes. These
cells retain lineage restriction while acquiring controllable
proliferative capacity. Their functional state is regulated through
AFP-dependent metabolic reprogramming and dynamically balanced
between proliferation and stress adaptation via the PPARγ signaling
axis (5). Furthermore, expansion
of this cell population is driven by TNF-α/AP-1 signaling
originating from neutrophils in the host liver, highlighting the
critical role of the immune microenvironment in regulating
regenerative cell states. Collectively, these findings indicate
that liver regeneration does not rely on a single progenitor pool
but instead involves coordinated responses from multiple hepatocyte
subpopulations with distinct zonal characteristics. This
coordinated response forms a dynamic and adaptive regenerative
network, and such cellular plasticity is essential for effective
tissue repair.
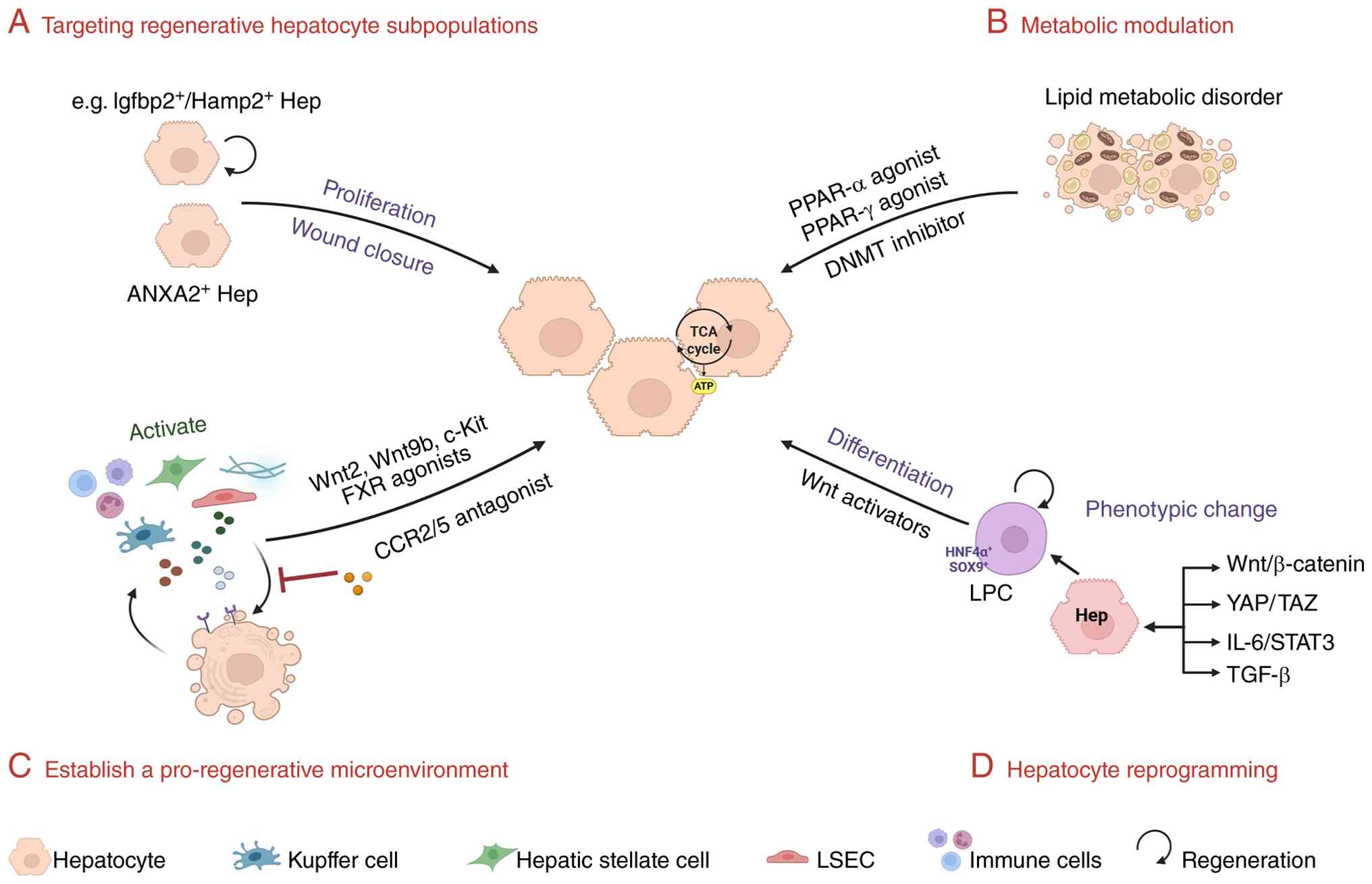
Emerging paradigms in liver regeneration
therapeutics converge on spatial orchestration of cellular
plasticity and microenvironmental reprogramming (1). At the cellular level, strategies
focus on modulating intrinsic plasticity thresholds through
epigenetic and metabolic circuit editing, enabling
context-dependent transdifferentiation while preserving functional
zonation (Fig. 3) (134,135). Concurrently, niche-directed
approaches recalibrate stromal-immune signaling gradients to
license region-specific progenitor activation (136).
Liver regeneration necessitates dynamic metabolic
adaptation, where zonation-defined specialization is transiently
repurposed to resolve the energy-biosynthesis paradox (135). In the early phase of
regeneration, HIF-1α-driven glycolytic flux predominates, while
AKT/mTOR-mediated suppression of gluconeogenesis and FAO redirects
substrates toward lipid droplet biogenesis, providing a strategic
energy reserve to support proliferative expansion (137). This metabolic triage creates
therapeutic opportunities through phased interventions. First, HIF
stabilizers boost zone 1-2 glycolytic priming. Concurrently, PPARγ
agonists sustain lipogenic competence, while mTOR modulators delay
catabolic restoration to preserve energy reservoirs (138). Post-proliferative metabolic
recovery, driven by PPARα-dependent FAO reactivation and
mitochondrial rejuvenation, is spatially coordinated to reinstate
functional zonation (139,140). This phase ensures pericentral
detoxification via CYP450 restoration and zonal energy flux
balancing, completing the metabolic reprogramming cycle essential
for functional restoration.
Epigenetic modifiers serve as spatial regulators of
metabolic reprogramming, offering druggable nodes to enhance
regenerative precision. DNA methyltransferases (DNMTs) enforce FAO
gene silencing during regeneration initiation, with pharmacological
DNMT inhibition extending the glycolytic proliferative window
(141). Conversely, histone
acetylase activators accelerate oxidative phosphorylation recovery
by decompacting chromatin at mitochondrial genes, enabling timely
metabolic restoration (142,143). Spatial multiomics have decoded
zone-specific metabolite-epigenome crosstalk, identifying serine
and ketone bodies as zone 2 resolved cofactors that prime chromatin
accessibility for plasticity transcription factors, a mechanism
that may be exploitable through dietary or metabolite
supplementation strategies (144). For instance, zone 1 targeted
HIF stabilizers amplify glycolytic flux, while zone 2 directed
serine supplementation reinforces epigenetic licensing of
plasticity genes. By mirroring the liver's innate spatiotemporal
hierarchy where metabolic priorities shift from proliferation fuel
to functional specialization, these strategies transform
regenerative bottlenecks into therapeutic windows.
Non-parenchymal cells have been recognized for their
roles in maintaining liver zonation, and also make critical
contributions to the regenerative microenvironment. Spatial
transcriptomics and single-cell analyses have shown that
pericentral HSC subsets enhance Wnt/β-catenin signaling through
secretion of Rspo3, thereby supporting hepatocyte proliferation
during regeneration (141). The
loss of HSCs or Rspo3 impairs regenerative capacity, while clinical
data indicate that higher Rspo3 expression is associated with
improved survival and a reduced risk of hepatocellular carcinoma,
supporting its potential as a therapeutic target (145).
LSECs are also key regulators of regeneration.
Previous studies demonstrate that LSECs promote hepatocyte
proliferation through secretion of Wnt2 and Wnt9b (47,146,147). Further work has identified
c-Kit signaling in LSECs as essential for this pro-regenerative
function. c-Kit+ LSECs upregulate Wnt2 expression
following injury, thereby stimulating proliferation of adjacent
hepatocytes, whereas disruption of c-Kit signaling reduces Wnt
production and significantly impairs liver regeneration (147,148). Recent advances have extended
these findings into therapeutic strategies. Engineered
exosome-based systems have been developed to target LSECs,
combining CRLTRKRGLK peptide-mediated targeting with CD47 mediated
evasion of mononuclear phagocyte clearance, enabling efficient
delivery of Wnt2 mRNA (149).
In APAP and dimethylnitrosamine-induced liver injury models, this
approach enhances Wnt signaling, promotes hepatocyte proliferation,
and improves tissue repair, demonstrating the feasibility of
modulating non-parenchymal cell-derived signals to reconstruct the
regenerative microenvironment (149).
In addition, a recent study has shown that
injury-induced downregulation of URI1 leads to glutamate
accumulation, which acts as a paracrine signal to recruit bone
marrow-derived monocytes. These immune cells subsequently secrete
Wnt3, activating YAP1 signaling in zone 2 hepatocytes and driving
their proliferation (150).
This metabolic immune regenerative axis follows a well-defined
spatiotemporal sequence and has been validated across multiple
models of acute and chronic liver injury. Clinical analyses further
suggest that this pathway is dysregulated in chronic liver disease,
characterized by reduced Wnt3 expression and disrupted metabolic
signaling (150).
Non-parenchymal cells thus establish a spatially organized
signaling network that governs liver regeneration within the
framework of liver zonation. This coordinated multicellular system
indicates that liver regeneration is not driven by a single cell
type but instead depends on the dynamic remodeling of
zonation-defined regenerative niches. Targeting non-parenchymal
cells and their spatial signaling networks therefore represents a
promising strategy for enhancing liver regeneration.
Hepatocyte dedifferentiation, a spatially
restricted process predominantly localized to zone 2, functions as
an evolutionary safeguard mechanism for hepatic repair. This
regenerative phenomenon involves surviving hepatocytes reactivating
fetal transcriptional programs through conserved developmental
pathways, particularly Wnt/β-catenin (151), YAP (152) and TGF-β signaling cascades
(153). Beyond the classical
Sox9+/HNF4α+ progenitor-like transition,
studies have identified multiple regenerative hepatocyte
subpopulations arising from dedifferentiation or enhanced cellular
plasticity of mature hepatocytes (154). For instance, AFP-positive
reprogrammed hepatocytes represent a state of partial
dedifferentiation, retaining lineage restriction while acquiring
proliferative capacity. Hepatocytes driven by YAP or Wnt/β-catenin
signaling exhibit a reversible progenitor-like phenotype (152). In addition, transitional
hepatocyte populations with high transcriptional plasticity, as
well as hepatocytes with migratory capacity involved in wound
repair, have been identified at injury borders (119). Collectively, these distinct
cellular states constitute a heterogeneous pool of regenerative
sources, indicating that liver regeneration relies on multilayered
cell fate reprogramming rather than a single progenitor
reservoir.
Critically, liver regeneration extends beyond
cell-autonomous reprogramming through spatially coordinated niche
interactions. For example, portal vein injury models reveal that
regionally activated Kupffer cells secrete IL-6, which induces
STAT3 activation in adjacent hepatocytes to directly drive
reprogramming-related gene re-expression (154). This spatial regulation is
further governed by metabolic zonation dynamics that
compartmentalize regenerative responses across hepatic lobules.
Therapeutic strategies for liver diseases have
expanded with the development of multiple targeted agents, several
of which directly exploit or modulate hepatic zonation features
(Table II). In MASH,
obeticholic acid, an agonist of the FXR, regulates bile acid
metabolism in close association with the functional specialization
of pericentral hepatocytes (155). Aldafermin (NGM282), a FGF19
analog, suppresses bile acid synthesis through FGFR4 signaling and
similarly targets metabolic regulation in the pericentral zone
(156). PRI-724 selectively
inhibits the interaction between β-catenin and CBP, thereby
directly modulating Wnt-dependent zonal signaling in pericentral
hepatocytes (157).
However, the clinical translation of these drugs
still faces considerable challenges. Cenicriviroc, by blocking the
CCR2/CCR5 signaling pathway, can reduce F4/80-positive macrophages
in the portal and necrotic regions, showing some anti-fibrotic
effects in a phase II trial (158). However, its efficacy was not
maintained in phase III trials (158). It could be considered that
single pathway blockade cannot overcome the redundancy of
chemotactic signals in liver fibrosis and the dual functional roles
of macrophages, nor does it directly target HSCs or ECM)
deposition, resulting in limited overall anti-fibrotic efficacy
(159). However, patients
exhibit heterogeneous fibrosis types; early, inflammation-dominant
fibrosis responds better to intervention, whereas ECM deposition
driven by sustained HSC activation is difficult to reverse.
Similarly, the failure of Belapectin is largely attributable to
insufficient patient stratification. Galectin-3 primarily regulates
HSC differentiation and serves a key role in ECM deposition
(160). As its inhibitor,
Belapectin can theoretically attenuate fibrosis, but without
accounting for the pathological heterogeneity of patients, the
overall efficacy was inadequate (161). This indicates that whether
targeting immune cells or non-parenchymal cells such as HSCs,
clinical success depends critically on precise patient
stratification and careful consideration of the spatial and
mechanistic heterogeneity of fibrosis.
Not only does the type of disease progression
affect treatment outcomes, but an increasing number of studies
indicate that factors such as sex, age and the gut microbiome can
reshape liver zonation through specific signaling pathways, thereby
influencing disease development and therapeutic responses. As
aforementioned, sex-dependent hormones, particularly GH can
dynamically regulate liver zonation (162). In MASLD, male GH pulses drive
STAT5 activation, exacerbating lipid metabolic disorders and
inflammation, making the centrilobular zone more prone to fat
accumulation and fibrosis. By contrast, estrogen promotes FAO and
inhibits HSC activation, providing protection in this region,
although this effect is markedly reduced after menopause (163). Age is also an key confounding
factor, as zonal characteristics become less distinct in the aging
liver, reducing the spatial precision of drug targeting (164). Furthermore, disruption of the
gut barrier allows microbes and their metabolites, such as bile
acids, to enter the portal circulation and form spatial gradients
along the hepatic lobule blood flow. Bile acids regulate metabolism
and inflammation through FXR and TGR5 receptors, which are
differentially expressed across liver zones, thereby driving
region-specific fibrotic progression (165). Thus, the gut microbiome
modulates pathological states in different liver regions via the
'bile acid-zonation signaling axis', influencing the efficacy of
zonation-targeted therapies. Taken together, future research should
focus on combination therapies, precise patient stratification and
spatially targeted delivery strategies based on liver zonation
biology.
With the rapid advancement of biotechnology, the
accumulation of high-throughput omics data is driving the deep
integration of bioinformatics and artificial intelligence (AI) in
liver disease research, providing new avenues for early diagnosis,
risk stratification and prognostic prediction (168,169). Traditional machine learning and
deep learning approaches can extract latent patterns from complex
multidimensional data and significantly outperform conventional
clinical parameters, highlighting the potential of multi-source
data integration as a key tool for precision hepatology. However,
current AI models largely remain at the level of prediction and
lack the ability to resolve spatial heterogeneity and underlying
mechanisms of disease (170).
Future efforts should shift from purely data-driven
prediction toward a new paradigm that integrates mechanistic
insights with spatial information. Incorporating spatial
transcriptomics and single-cell sequencing into AI models would
enable the modeling of zonation-specific signals within the hepatic
lobule, thereby revealing region-specific contributions to disease
initiation and progression. Overall, the convergence of AI and
spatial hepatology is poised to reshape liver disease research and
clinical management, providing a new theoretical and technological
foundation for precision therapy.
Liver zonation is not merely a manifestation of
metabolic compartmentalization but represents a dynamic regulatory
framework that integrates physiological function with disease
pathogenesis. Along the porto-central axis, a continuous metabolic
gradient is established through coordinated signaling interactions
between zonation-specific hepatocyte subpopulations and
non-parenchymal cells, thereby shaping region-specific metabolic
programs and functional specialization. This highly ordered spatial
organization supports efficient hepatic metabolism under
homeostatic conditions, while simultaneously conferring
region-specific vulnerability under pathological states. Spatial
multi-omics studies further demonstrate that different hepatic
zones exhibit distinct responses to injury, indicating that liver
disease fundamentally arises from disruption of zonal architecture
and imbalance of spatial signaling networks.
Building on this concept, the integration of
spatial biology and regenerative medicine is driving a paradigm
shift in therapeutic strategies from 'cell-targeted' to
'zonation-targeted' approaches. Emerging evidence shows that
adeno-associated virus-mediated expression of R-spondin3 can
restore Wnt/β-catenin signaling gradients, thereby re-establishing
pericentral metabolic identity and ameliorating fibrosis in
preclinical models. However, clinical translation of such
strategies remains challenged by the difficulty of achieving
precise spatial control of zonal signaling and by substantial
inter-individual variability. Recently developed organoid and
bioengineered systems incorporate physiologically relevant oxygen
and nutrient gradients, enabling reconstruction of liver
lobule-like spatial architecture in vitro and providing
powerful platforms for dissecting zonal regulatory mechanisms and
screening compartment-specific interventions. In the future, the
integration of spatial omics, AI, controllable delivery systems and
personalized models are expected to bridge mechanistic insights
with therapeutic development, thereby advancing zonation-based
precision interventions toward clinical application.
Not applicable.
DY conceived the idea and supervised the project.
DY and GC wrote the manuscript. HJ, YL, CG and HZ helped with
figure preparation. DY, GC and KZ revised the manuscript. HJ and YL
participated in editing the manuscript. CG, HZ and YC contributed
to revision of the manuscript. KZ, YC and DY finalized the
manuscript and made the conceptual evaluation of the manuscript.
Data authentication is not applicable. All authors read and
approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present was supported by grants from the Key Laboratory Open
Project Foundation of Jiangsu University (grant no. 8161280002),
the Key Laboratory Open Project Foundation of Jiangsu University
(Outstanding Research Project) (grant no. JSKLM-Z-2024-011),
Science and Technology Plan (Apply Basic Research) of Changzhou
City (grant no. CJ20210006), Science and Technology Plan (Apply
Basic Research) of Changzhou City (grant no. CJ20230002), Science
and Technology Plan (Apply Basic Research) of Changzhou City (grant
no. CJ20241041), Natural Science Foundation of China (grant no.
82100666), the National Natural Science Foundation for Young
Scientists of Jiangsu Province (grant no. BK20200906) and the Young
Scientists Initiative Foundation of Jiangsu University (grant no.
20JDG49).
|
1
|
Ben-Moshe S and Itzkovitz S: Spatial
heterogeneity in the mammalian liver. Nat Rev Gastroenterol
Hepatol. 16:395–410. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Halpern KB, Shenhav R, Matcovitch-Natan O,
Toth B, Lemze D, Golan M, Massasa EE, Baydatch S, Landen S, Moor
AE, et al: Single-cell spatial reconstruction reveals global
division of labour in the mammalian liver. Nature. 542:352–356.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang S, Wang X, Shan Y, Tan Z, Su Y, Cao
Y, Wang S, Dong J, Gu J and Wang Y: Region-specific cellular and
molecular basis of liver regeneration after acute pericentral
injury. Cell Stem Cell. 31:341–358. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Watson BR, Paul B, Rahman RU,
Amir-Zilberstein L, Segerstolpe Å, Epstein ET, Murphy S,
Geistlinger L, Lee T, Shih A, et al: Spatial transcriptomics of
healthy and fibrotic human liver at single-cell resolution. Nat
Commun. 16:3192025. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fang T, Yang C, Qiu H, Du Y, Wang X, Li Y,
Xu M, Liu C, Li X, Guo N, et al: Conversion of transplanted mature
hepatocytes into Afp+ reprogrammed cells for liver
regeneration after injury. Adv Sci (Weinh). 13:e171262026.
View Article : Google Scholar
|
|
6
|
Gebhardt R: Metabolic zonation of the
liver: Regulation and implications for liver function. Pharmacol
Ther. 53:275–354. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Braeuning A, Ittrich C, Kohle C,
Hailfinger S, Bonin M, Buchmann A and Schwarz M: Differential gene
expression in periportal and perivenous mouse hepatocytes. FEBS J.
273:5051–5061. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jungermann K and Katz N: Functional
specialization of different hepatocyte populations. Physiol Rev.
69:708–764. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jungermann K and Katz N: Functional
hepatocellular heterogeneity. Hepatology. 2:385–395. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hijmans BS, Grefhorst A, Oosterveer MH and
Groen AK: Zonation of glucose and fatty acid metabolism in the
liver: Mechanism and metabolic consequences. Biochimie. 96:121–129.
2014. View Article : Google Scholar
|
|
11
|
Jungermann K and Kietzmann T: Zonation of
parenchymal and nonparenchymal metabolism in liver. Annu Rev Nutr.
16:179–203. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Haussinger D and Gerok W: Hepatocyte
heterogeneity in ammonia metabolism: Impairment of glutamine
synthesis in CCL4 induced liver cell necrosis with no effect on
urea synthesis. Chem Biol Interact. 48:191–194. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Berndt N, Kolbe E, Gajowski R, Eckstein J,
Ott F, Meierhofer D, Holzhutter HG and Matz-Soja M: Functional
consequences of metabolic zonation in murine livers: Insights for
an old story. Hepatology. 73:795–810. 2021. View Article : Google Scholar
|
|
14
|
Zanger UM and Schwab M: Cytochrome p450
enzymes in drug metabolism: Regulation of gene expression, enzyme
activities, and impact of genetic variation. Pharmacol Ther.
138:103–141. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Parlakgul G, Arruda AP, Pang S, Cagampan
E, Min N, Guney E, Lee GY, Inouye K, Hess HF, Xu CS and
Hotamışlıgil GS: Regulation of liver subcellular architecture
controls metabolic homeostasis. Nature. 603:736–742. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Parlakgul G, Pang S, Artico LL, Min N,
Cagampan E, Villa R, Goncalves R, Lee GY, Xu CS, Hotamisligil GS
and Arruda AP: Spatial mapping of hepatic ER and mitochondria
architecture reveals zonated remodeling in fasting and obesity. Nat
Commun. 15:39822024. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Plata-Gomez AB, de Prado-Rivas L, Sanz A,
Deleyto-Seldas N, Garcia F, de la Calle AC, Silva C, Caleiras E,
Grana-Castro O, Pineiro-Yanez E, et al: Hepatic nutrient and
hormone signaling to mTORC1 instructs the postnatal metabolic
zonation of the liver. Nat Commun. 15:18782024. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kozuki S, Kabata M, Sakurai S, Iwaisako K,
Nishimura T, Toi M, Yamamoto T and Toyoshima F: Periportal
hepatocyte proliferation at midgestation governs maternal glucose
homeostasis in mice. Commun Biol. 6:12262023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rosenberger FA, Thielert M, Strauss MT,
Schweizer L, Ammar C, Madler SC, Metousis A, Skowronek P, Wahle M,
Madden K, et al: Spatial single-cell mass spectrometry defines
zonation of the hepatocyte proteome. Nat Methods. 20:1530–1536.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He S, Guo Z, Zhou M, Wang H, Zhang Z, Shi
M, Li X, Yang X and He L: Spatial-temporal proliferation of
hepatocytes during pregnancy revealed by genetic lineage tracing.
Cell Stem Cell. 30:1549–1558. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hildebrandt F, Andersson A, Saarenpaa S,
Larsson L, Van Hul N, Kanatani S, Masek J, Ellis E, Barragan A,
Mollbrink A, et al: Spatial transcriptomics to define
transcriptional patterns of zonation and structural components in
the mouse liver. Nat Commun. 12:70462021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liang Y, Kaneko K, Xin B, Lee J, Sun X,
Zhang K and Feng GS: Temporal analyses of postnatal liver
development and maturation by single-cell transcriptomics. Dev
Cell. 57:398–414. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang L, Wang X, Zheng JX, Xu ZR, Li LC,
Xiong YL, Zhou BC, Gao J and Xu CR: Determination of key events in
mouse hepatocyte maturation at the single-cell level. Dev Cell.
58:1996–2010. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matsumoto T, Wakefield L, Tarlow BD and
Grompe M: In vivo lineage tracing of polyploid hepatocytes reveals
extensive proliferation during liver regeneration. Cell Stem Cell.
26:34–47. 2020. View Article : Google Scholar
|
|
25
|
Katsuda T, Hosaka K, Matsuzaki J, Usuba W,
Prieto-Vila M, Yamaguchi T, Tsuchiya A, Terai S and Ochiya T:
Transcriptomic dissection of hepatocyte heterogeneity: Linking
ploidy, zonation, and stem/progenitor cell characteristics. Cell
Mol Gastroenterol Hepatol. 9:161–183. 2020. View Article : Google Scholar
|
|
26
|
Richter ML, Deligiannis IK, Yin K, Danese
A, Lleshi E, Coupland P, Vallejos CA, Matchett KP, Henderson NC,
Colome-Tatche M and Martinez-Jimenez CP: Single-nucleus rna-seq2
reveals functional crosstalk between liver zonation and ploidy. Nat
Commun. 12:42642021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Su T, Yang Y, Lai S, Jeong J, Jung Y,
Mcconnell M, Utsumi T and Iwakiri Y: Single-cell transcriptomics
reveals zone-specific alterations of liver sinusoidal endothelial
cells in cirrhosis. Cell Mol Gastroenterol Hepatol. 11:1139–1161.
2021. View Article : Google Scholar
|
|
28
|
Payen VL, Lavergne A, Alevra SN, Colonval
M, Karim L, Deckers M, Najimi M, Coppieters W, Charloteaux B, Sokal
EM, et al: Single-cell RNA sequencing of human liver reveals
hepatic stellate cell heterogeneity. JHEP Rep. 3:1002782021.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gola A, Dorrington MG, Speranza E, Sala C,
Shih RM, Radtke AJ, Wong HS, Baptista AP, Hernandez JM, Castellani
G, et al: Commensal-driven immune zonation of the liver promotes
host defence. Nature. 589:131–136. 2021. View Article : Google Scholar
|
|
30
|
Wu B, Shentu X, Nan H, Guo P, Hao S, Xu J,
Shangguan S, Cui L, Cen J, Deng Q, et al: A spatiotemporal atlas of
cholestatic injury and repair in mice. Nat Genet. 56:938–952. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kolbe E, Aleithe S, Rennert C, Spormann L,
Ott F, Meierhofer D, Gajowski R, Stopel C, Hoehme S, Kucken M, et
al: Mutual zonated interactions of Wnt and Hh signaling are
orchestrating the metabolism of the adult liver in mice and human.
Cell Rep. 29:4553–4567. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Matz-Soja M, Hovhannisyan A and Gebhardt
R: Hedgehog signalling pathway in adult liver: A major new player
in hepatocyte metabolism and zonation? Med Hypotheses. 80:589–594.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jungermann K and Kietzmann T: Oxygen:
Modulator of metabolic zonation and disease of the liver.
Hepatology. 31:255–260. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Semba H, Takeda N, Isagawa T, Sugiura Y,
Honda K, Wake M, Miyazawa H, Yamaguchi Y, Miura M, Jenkins DM, et
al: HIF-1α-PDK1 axis-induced active glycolysis plays an essential
role in macrophage migratory capacity. Nat Commun. 7:116352016.
View Article : Google Scholar
|
|
35
|
Rius J, Guma M, Schachtrup C, Akassoglou
K, Zinkernagel AS, Nizet V, Johnson RS, Haddad GG and Karin M:
NF-kappaB links innate immunity to the hypoxic response through
transcriptional regulation of HIF-1alpha. Nature. 453:807–811.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Scortegagna M, Ding K, Oktay Y, Gaur A,
Thurmond F, Yan LJ, Marck BT, Matsumoto AM, Shelton JM, Richardson
JA, et al: Multiple organ pathology, metabolic abnormalities and
impaired homeostasis of reactive oxygen species in Epas1−/− mice.
Nat Genet. 35:331–340. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rankin EB, Rha J, Selak MA, Unger TL,
Keith B, Liu Q and Haase VH: Hypoxia-inducible factor 2 regulates
hepatic lipid metabolism. Mol Cell Biol. 29:4527–4538. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Makino Y, Kanopka A, Wilson WJ, Tanaka H
and Poellinger L: Inhibitory pas domain protein (IPAS) is a
hypoxia-inducible splicing variant of the hypoxia-inducible
factor-3alpha locus. J Biol Chem. 277:32405–32408. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhan L, Huang C, Meng XM, Song Y, Wu XQ,
Yang Y and Li J: Hypoxia-inducible factor-1alpha in hepatic
fibrosis: A promising therapeutic target. Biochimie. 108:1–7. 2015.
View Article : Google Scholar
|
|
40
|
Aggarwal SR, Lindros KO and Palmer TN:
Glucagon stimulates phosphorylation of different peptides in
isolated periportal and perivenous hepatocytes. FEBS Lett.
377:439–443. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kinugasa A and Thurman RG: Differential
effect of glucagon on gluconeogenesis in periportal and pericentral
regions of the liver lobule. Biochem J. 236:425–430. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Saito K, Negishi M and James SE: Sexual
dimorphisms in zonal gene expression in mouse liver. Biochem
Biophys Res Commun. 436:730–735. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Goldfarb CN, Karri K, Pyatkov M and Waxman
DJ: Interplay between GH-regulated, Sex-biased liver transcriptome
and hepatic zonation revealed by single-nucleus RNA sequencing.
Endocrinology. 163:bqac0592022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Annunziato S, Sun T and Tchorz JS: The
RSPO-LGR4/5-ZNRF3/RNF43 module in liver homeostasis, regeneration,
and disease. Hepatology. 76:888–899. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun T, Annunziato S, Bergling S, Sheng C,
Orsini V, Forcella P, Pikiolek M, Kancherla V, Holwerda S, Imanci
D, et al: ZNRF3 and RNF43 cooperate to safeguard metabolic liver
zonation and hepatocyte proliferation. Cell Stem Cell.
28:1822–1837. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Torre C, Perret C and Colnot S: Molecular
determinants of liver zonation. Prog Mol Biol Transl Sci.
97:127–150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hu S, Liu S, Bian Y, Poddar M, Singh S,
Cao C, Mcgaughey J, Bell A, Blazer LL, Adams JJ, et al: Single-cell
spatial transcriptomics reveals a dynamic control of metabolic
zonation and liver regeneration by endothelial cell Wnt2 and Wnt9b.
Cell Rep Med. 3:1007542022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Benhamouche S, Decaens T, Godard C,
Chambrey R, Rickman DS, Moinard C, Vasseur-Cognet M, Kuo CJ, Kahn
A, Perret C and Colnot S: Apc tumor suppressor gene is the
'zonation-keeper' of mouse liver. Dev Cell. 10:759–770. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gerbal-Chaloin S, Dume A, Briolotti P,
Klieber S, Raulet E, Duret C, Fabre J, Ramos J, Maurel P and
Daujat-Chavanieu M: The Wnt/β-catenin pathway is a transcriptional
regulator of CYP2E1, CYP1A2, and aryl hydrocarbon receptor gene
expression in primary human hepatocytes. Mol Pharmacol. 86:624–634.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kietzmann T: Liver zonation in health and
disease: Hypoxia and hypoxia-inducible transcription factors as
concert masters. Int J Mol Sci. 20:23472019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mazumdar J, O'Brien WT, Johnson RS,
Lamanna JC, Chavez JC, Klein PS and Simon MC: O2 regulates stem
cells through Wnt/β-catenin signalling. Nat Cell Biol.
12:1007–1013. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mori H, Yao Y, Learman BS, Kurozumi K,
Ishida J, Ramakrishnan SK, Overmyer KA, Xue X, Cawthorn WP, Reid
MA, et al: Induction of WNT11 by hypoxia and hypoxia-inducible
factor-1α regulates cell proliferation, migration and invasion. Sci
Rep. 6:215202016. View Article : Google Scholar
|
|
53
|
Braeuning A, Menzel M, Kleinschnitz EM,
Harada N, Tamai Y, Kohle C, Buchmann A and Schwarz M: Serum
components and activated Ha-ras antagonize expression of perivenous
marker genes stimulated by beta-catenin signaling in mouse
hepatocytes. FEBS J. 274:4766–4777. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mcenerney L, Duncan K, Bang BR, Elmasry S,
Li M, Miki T, Ramakrishnan SK, Shah YM and Saito T: Dual modulation
of human hepatic zonation via canonical and non-canonical Wnt
pathways. Exp Mol Med. 49:e4132017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wakizaka K, Kamiyama T, Kakisaka T, Orimo
T, Nagatsu A, Aiyama T, Shichi S and Taketomi A: Expression of
Wnt5a and ROR2, components of the noncanonical Wnt-signaling
pathway, is associated with tumor differentiation in hepatocellular
carcinoma. Ann Surg Oncol. 31:262–271. 2024. View Article : Google Scholar
|
|
56
|
Yang J, Cusimano A, Monga JK, Preziosi ME,
Pullara F, Calero G, Lang R, Yamaguchi TP, Nejak-Bowen KN and Monga
SP: Wnt5a inhibits hepatocyte proliferation and concludes
beta-catenin signaling in liver regeneration. Am J Pathol.
185:2194–2205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Omenetti A, Choi S, Michelotti G and Diehl
AM: Hedgehog signaling in the liver. J Hepatol. 54:366–373. 2011.
View Article : Google Scholar
|
|
58
|
Varnat F, Zacchetti G and Ruiz IAA:
Hedgehog pathway activity is required for the lethality and
intestinal phenotypes of mice with hyperactive Wnt signaling. Mech
Dev. 127:73–81. 2010. View Article : Google Scholar
|
|
59
|
Matz-Soja M, Aleithe S, Marbach E, Bottger
J, Arnold K, Schmidt-Heck W, Kratzsch J and Gebhardt R: Hepatic
hedgehog signaling contributes to the regulation of IGF1 and IGFBP1
serum levels. Cell Commun Signal. 12:112014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sicklick JK, Li YX, Choi SS, Qi Y, Chen W,
Bustamante M, Huang J, Zdanowicz M, Camp T, Torbenson MS, et al:
Role for hedgehog signaling in hepatic stellate cell activation and
viability. Lab Invest. 85:1368–1380. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Saviano A, Henderson NC and Baumert TF:
Single-cell genomics and spatial transcriptomics: Discovery of
novel cell states and cellular interactions in liver physiology and
disease biology. J Hepatol. 73:1219–1230. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Rocha AS, Vidal V, Mertz M, Kendall TJ,
Charlet A, Okamoto H and Schedl A: The angiocrine factor rspondin3
is a key determinant of liver zonation. Cell Rep. 13:1757–1764.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Halpern KB, Shenhav R, Massalha H, Toth B,
Egozi A, Massasa EE, Medgalia C, David E, Giladi A, Moor AE, et al:
Paired-cell sequencing enables spatial gene expression mapping of
liver endothelial cells. Nat Biotechnol. 36:962–970. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Macparland SA, Liu JC, Ma XZ, Innes BT,
Bartczak AM, Gage BK, Manuel J, Khuu N, Echeverri J, Linares I, et
al: Single cell RNA sequencing of human liver reveals distinct
intrahepatic macrophage populations. Nat Commun. 9:43832018.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wen Y, Lambrecht J, Ju C and Tacke F:
Hepatic macrophages in liver homeostasis and diseases-diversity,
plasticity and therapeutic opportunities. Cell Mol Immunol.
18:45–56. 2021. View Article : Google Scholar
|
|
66
|
Trinh VQ, Lee TF, Lemoinne S, Ray KC,
Ybanez MD, Tsuchida T, Carter JK, Agudo J, Brown BD, Akat KM, et
al: Hepatic stellate cells maintain liver homeostasis through
paracrine neurotrophin-3 signaling that induces hepatocyte
proliferation. Sci Signal. 16:eadf66962023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Dobie R, Wilson-Kanamori JR, Henderson B,
Smith JR, Matchett KP, Portman JR, Wallenborg K, Picelli S,
Zagorska A, Pendem SV, et al: Single-cell transcriptomics uncovers
zonation of function in the mesenchyme during liver fibrosis. Cell
Rep. 29:1832–1847. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Khan MA, Fischer J, Harrer L, Schwiering
F, Groneberg D and Friebe A: Hepatic stellate cells in zone 1
engage in capillarization rather than myofibroblast formation in
murine liver fibrosis. Sci Rep. 14:188402024. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Bravo GC, Matetovici I, Hillen H, Taskiran
II, Vandepoel R, Christiaens V, Sansores-Garcia L, Verboven E,
Hulselmans G, Poovathingal S, et al: Single-cell spatial
multi-omics and deep learning dissect enhancer-driven gene
regulatory networks in liver zonation. Nat Cell Biol. 26:153–167.
2024. View Article : Google Scholar
|
|
70
|
Chen H, Lu S, Zhou J, Bai Z, Fu H, Xu X,
Yang S, Jiao B and Sun Y: An integrated approach for the
identification of USF1-centered transcriptional regulatory networks
during liver regeneration. Biochim Biophys Acta. 1839:415–423.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Brosch M, Kattler K, Herrmann A, von
Schonfels W, Nordstrom K, Seehofer D, Damm G, Becker T, Zeissig S,
Nehring S, et al: Epigenomic map of human liver reveals principles
of zonated morphogenic and metabolic control. Nat Commun.
9:41502018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Stanulovic VS, Kyrmizi I, Kruithof-De JM,
Hoogenkamp M, Vermeulen JL, Ruijter JM, Talianidis I, Hakvoort TB
and Lamers WH: Hepatic hnf4alpha deficiency induces periportal
expression of glutamine synthetase and other pericentral enzymes.
Hepatology. 45:433–444. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lee J, Wang C, Xu S, Cho Y, Wang L, Feng
X, Baldridge A, Sartorelli V, Zhuang L, Peng W, et al: H3K4 mono-
and di-methyltransferase MLL4 is required for enhancer activation
during cell differentiation. Elife. 2:e15032013. View Article : Google Scholar
|
|
74
|
Moore A, Wu L, Chuang J, Sun X, Luo X,
Gopal P, Li L, Celen C, Zimmer M and Zhu H: Arid1a loss drives
nonalcoholic steatohepatitis in mice through epigenetic
dysregulation of hepatic lipogenesis and fatty acid oxidation.
Hepatology. 69:1931–1945. 2019. View Article : Google Scholar
|
|
75
|
Qu Y, Deng C, Luo Q, Shang X, Wu J, Shi Y,
Wang L and Han Z: Arid1a regulates insulin sensitivity and lipid
metabolism. EBioMedicine. 42:481–493. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hsu SH, Delgado ER, Otero PA, Teng KY,
Kutay H, Meehan KM, Moroney JB, Monga JK, Hand NJ, Friedman JR, et
al: Microrna-122 regulates polyploidization in the murine liver.
Hepatology. 64:599–615. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ben-Moshe S, Shapira Y, Moor AE, Manco R,
Veg T, Bahar Halpern K and Itzkovitz S: Spatial sorting enables
comprehensive characterization of liver zonation. Nat Metab.
1:899–911. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wang B, Zhao L, Fish M, Logan CY and Nusse
R: Self-renewing diploid Axin2(+) cells fuel homeostatic renewal of
the liver. Nature. 524:180–185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Sun T, Pikiolek M, Orsini V, Bergling S,
Holwerda S, Morelli L, Hoppe PS, Planas-Paz L, Yang Y, Ruffner H,
et al: Axin2+ pericentral hepatocytes have limited
contributions to liver homeostasis and regeneration. Cell Stem
Cell. 26:97–107.e6. 2020. View Article : Google Scholar
|
|
80
|
May S, Muller M, Livingstone CR, Skalka
GL, Walsh PJ, Nixon C, Hedley A, Shaw R, Clark W, Vande VJ, et al:
Absent expansion of AXIN2+ hepatocytes and altered physiology in
Axin2CreERT2 mice challenges the role of pericentral hepatocytes in
homeostatic liver regeneration. J Hepatol. 78:1028–1036. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Lin S, Nascimento EM, Gajera CR, Chen L,
Neuhofer P, Garbuzov A, Wang S and Artandi SE: Distributed
hepatocytes expressing telomerase repopulate the liver in
homeostasis and injury. Nature. 556:244–248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Ben-Moshe S, Veg T, Manco R, Dan S,
Papinutti D, Lifshitz A, Kolodziejczyk AA, Bahar HK, Elinav E and
Itzkovitz S: The spatiotemporal program of zonal liver regeneration
following acute injury. Cell Stem Cell. 29:973–989. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
He L, Pu W, Liu X, Zhang Z, Han M, Li Y,
Huang X, Han X, Li Y, Liu K, et al: Proliferation tracing reveals
regional hepatocyte generation in liver homeostasis and repair.
Science. 371:eabc43462021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Prior N, Hindley CJ, Rost F, Melendez E,
Lau W, Gottgens B, Rulands S, Simons BD and Huch M:
Lgr5+ stem and progenitor cells reside at the apex of a
heterogeneous embryonic hepatoblast pool. Development.
146:dev1745572019. View Article : Google Scholar
|
|
85
|
Qin D, Liu S, Lu Y, Yan Y, Zhang J, Cao S,
Chen M, Chen N, Huang W, Wang L, et al: Lgr5+ cell fate
regulation by coordination of metabolic nuclear receptors during
liver repair. Theranostics. 12:6130–6142. 2022. View Article : Google Scholar
|
|
86
|
Chen F, Jimenez RJ, Sharma K, Luu HY, Hsu
BY, Ravindranathan A, Stohr BA and Willenbring H: Broad
distribution of hepatocyte proliferation in liver homeostasis and
regeneration. Cell Stem Cell. 26:27–33. 2020. View Article : Google Scholar
|
|
87
|
Wei Y, Wang YG, Jia Y, Li L, Yoon J, Zhang
S, Wang Z, Zhang Y, Zhu M, Sharma T, et al: Liver homeostasis is
maintained by midlobular zone 2 hepatocytes. Science.
371:eabb16252021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kubota N, Kubota T and Kadowaki T:
Midlobular zone 2 hepatocytes: A gatekeeper of liver homeostasis.
Cell Metab. 33:855–856. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lin YH, Wei Y, Zeng Q, Wang Y, Pagani CA,
Li L, Zhu M, Wang Z, Hsieh MH, Corbitt N, et al: IGFBP2 expressing
midlobular hepatocytes preferentially contribute to liver
homeostasis and regeneration. Cell Stem Cell. 30:665–676.e4. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Q BA, Vesco KK, Purnell JQ, Francisco M,
Goddard E, Guan X, Debarber A, Leo MC, Baetscher E, Rooney W, et
al: Pregnancy and weaning regulate human maternal liver size and
function. Proc Natl Acad Sci USA. 118:e21072691182021. View Article : Google Scholar
|
|
91
|
Milona A, Owen BM, van Mil S, Dormann D,
Mataki C, Boudjelal M, Cairns W, Schoonjans K, Milligan S, Parker
M, et al: The normal mechanisms of pregnancy-induced liver growth
are not maintained in mice lacking the bile acid sensor Fxr. Am J
Physiol Gastrointest Liver Physiol. 298:G151–G158. 2010. View Article : Google Scholar
|
|
92
|
Jaeschke H and Ramachandran A:
Acetaminophen hepatotoxicity: Paradigm for understanding mechanisms
of drug-induced liver injury. Ann Rev Pathol. 19:453–478. 2024.
View Article : Google Scholar
|
|
93
|
Li M, Cai S and Boyer JL: Mechanisms of
bile acid mediated inflammation in the liver. Mol Aspects Med.
56:45–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Younossi Z and Henry L: Contribution of
alcoholic and nonalcoholic fatty liver disease to the burden of
liver-related morbidity and mortality. Gastroenterology.
150:1778–1785. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Ibrahim SH, Hirsova P and Gores GJ:
Non-alcoholic steatohepatitis pathogenesis: Sublethal hepatocyte
injury as a driver of liver inflammation. Gut. 67:963–972. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Chalasani N, Wilson L, Kleiner DE,
Cummings OW, Brunt EM and Unalp A: Relationship of steatosis grade
and zonal location to histological features of steatohepatitis in
adult patients with non-alcoholic fatty liver disease. J Hepatol.
48:829–834. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Sheka AC, Adeyi O, Thompson J, Hameed B,
Crawford PA and Ikramuddin S: Nonalcoholic steatohepatitis: A
review. JAMA. 323:1175–1183. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Leow WQ, Chan AW, Mendoza P, Lo R, Yap K
and Kim H: Non-alcoholic fatty liver disease: The Pathologist's
perspective. Clin Mol Hepatol. 29(Suppl): S302–S318. 2023.
View Article : Google Scholar
|
|
99
|
Carter-Kent C, Yerian LM, Brunt EM, Angulo
P, Kohli R, Ling SC, Xanthakos SA, Whitington PF,
Charatcharoenwitthaya P, Yap J, et al: Nonalcoholic steatohepatitis
in children: A multi-center clinicopathological study. Hepatology.
50:1113–1120. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Swiderska-Syn M, Suzuki A, Guy CD,
Schwimmer JB, Abdelmalek MF, Lavine JE and Diehl AM: Hedgehog
pathway and pediatric nonalcoholic fatty liver disease. Hepatology.
57:1814–1825. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Machado MV, Michelotti GA, Pereira TA, Xie
G, Premont R, Cortez-Pinto H and Diehl AM: Accumulation of duct
cells with activated yap parallels fibrosis progression in
non-alcoholic fatty liver disease. J Hepatol. 63:962–970. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Gadd VL, Skoien R, Powell EE, Fagan KJ,
Winterford C, Horsfall L, Irvine K and Clouston AD: The portal
inflammatory infiltrate and ductular reaction in human nonalcoholic
fatty liver disease. Hepatology. 59:1393–1405. 2014. View Article : Google Scholar
|
|
103
|
Richardson MM, Jonsson JR, Powell EE,
Brunt EM, Neuschwander-Tetri BA, Bhathal PS, Dixon JB, Weltman MD,
Tilg H, Moschen AR, et al: Progressive fibrosis in nonalcoholic
steatohepatitis: Association with altered regeneration and a
ductular reaction. Gastroenterology. 133:80–90. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Ko S, Russell JO, Molina LM and Monga SP:
Liver progenitors and adult cell plasticity in hepatic injury and
repair: Knowns and unknowns. Annu Rev Pathol. 15:23–50. 2020.
View Article : Google Scholar
|
|
105
|
Hammoutene A and Rautou P: Role of liver
sinusoidal endothelial cells in non-alcoholic fatty liver disease.
J Hepatol. 70:1278–1291. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhou Y, Zhao Y, Carbonaro M, Chen H,
Germino M, Adler C, Ni M, Zhu YO, Kim SY, Altarejos J, et al:
Perturbed liver gene zonation in a mouse model of non-alcoholic
steatohepatitis. Metabolism. 154:1558302024. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Kostallari E, Wei B, Sicard D, Li J,
Cooper SA, Gao J, Dehankar M, Li Y, Cao S, Yin M, et al: Stiffness
is associated with hepatic stellate cell heterogeneity during liver
fibrosis. Am J Physiol Gastrointest Liver Physiol. 322:G234–G246.
2022. View Article : Google Scholar
|
|
108
|
Llovet JM, Pinyol R, Kelley RK,
El-Khoueiry A, Reeves HL, Wang XW, Gores GJ and Villanueva A:
Molecular pathogenesis and systemic therapies for hepatocellular
carcinoma. Nat Cancer. 3:386–401. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Barker N and Clevers H: Mining the Wnt
pathway for cancer therapeutics. Nat Rev Drug Discov. 5:997–1014.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Sia D, Villanueva A, Friedman SL and
Llovet JM: Liver cancer cell of origin, molecular class, and
effects on patient prognosis. Gastroenterology. 152:745–761. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Holczbauer A, Wangensteen KJ and Shin S:
Cellular origins of regenerating liver and hepatocellular
carcinoma. JHEP Rep. 4:1004162022. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Perugorria MJ, Olaizola P, Labiano I,
Esparza-Baquer A, Marzioni M, Marin J, Bujanda L and Banales JM:
Wnt-beta-catenin signalling in liver development, health and
disease. Nat Rev Gastroenterol Hepatol. 16:121–136. 2019.
View Article : Google Scholar
|
|
113
|
Zhang T, Gu J, Wang X, Lu Y, Cai K, Li H,
Nie Y, Chen X and Wang J: A novel liver zonation
phenotype-associated molecular classification of hepatocellular
carcinoma. Front Immunol. 14:11402012023. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Desert R, Rohart F, Canal F, Sicard M,
Desille M, Renaud S, Turlin B, Bellaud P, Perret C, Clement B, et
al: Human hepatocellular carcinomas with a periportal phenotype
have the lowest potential for early recurrence after curative
resection. Hepatology. 66:1502–1518. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Ng C, Piscuoglio S and Terracciano LM:
Molecular classification of hepatocellular carcinoma: The view from
metabolic zonation. Hepatology. 66:1377–1380. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Stravitz RT and Lee WM: Acute liver
failure. Lancet. 394:869–881. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Anundi I, Lahteenmaki T, Rundgren M,
Moldeus P and Lindros KO: Zonation of acetaminophen metabolism and
cytochrome P450 2E1-mediated toxicity studied in isolated
periportal and perivenous hepatocytes. Biochem Pharmacol.
45:1251–1259. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Avasarala S, Yang L, Sun Y, Leung AW, Chan
W, Cheung W and Lee SS: A temporal study on the histopathological,
biochemical and molecular responses of CCL(4)-induced
hepatotoxicity in Cyp2e1-null mice. Toxicology. 228:310–322. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Matchett KP, Wilson-Kanamori JR, Portman
JR, Kapourani CA, Fercoq F, May S, Zajdel E, Beltran M, Sutherland
EF, Mackey J, et al: Multimodal decoding of human liver
regeneration. Nature. 630:158–165. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Kpetemey M, Dasgupta S, Rajendiran S, Das
S, Gibbs LD, Shetty P, Gryczynski Z and Vishwanatha JK: MIEN1, a
novel interactor of annexin A2, promotes tumor cell migration by
enhancing AnxA2 cell surface expression. Mol Cancer. 14:1562015.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Gouw ASH, Clouston AD and Theise ND:
Ductular reactions in human liver: Diversity at the interface.
Hepatology. 54:1853–1863. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Miyajima A, Tanaka M and Itoh T:
Stem/progenitor cells in liver development, homeostasis,
regeneration, and reprogramming. Cell Stem Cell. 14:561–574. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Fickert P, Stoger U, Fuchsbichler A,
Moustafa T, Marschall HU, Weiglein AH, Tsybrovskyy O, Jaeschke H,
Zatloukal K, Denk H and Trauner M: A new xenobiotic-induced mouse
model of sclerosing cholangitis and biliary fibrosis. Am J Pathol.
171:525–536. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Kullak-Ublick GA, Stieger B and Meier PJ:
Enterohepatic bile salt transporters in normal physiology and liver
disease. Gastroenterology. 126:322–342. 2004. View Article : Google Scholar
|
|
125
|
Font-Burgada J, Shalapour S, Ramaswamy S,
Hsueh B, Rossell D, Umemura A, Taniguchi K, Nakagawa H, Valasek MA,
Ye L, et al: Hybrid periportal hepatocytes regenerate the injured
liver without giving rise to cancer. Cell. 162:766–779. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Shao C, Jing Y, Zhao S, Yang X, Hu Y, Meng
Y, Huang Y, Ye F, Gao L, Liu W, et al: LPS/Bcl3/YAP1 signaling
promotes Sox9+HNF4α+ hepatocyte-mediated liver regeneration after
hepatectomy. Cell Death Dis. 13:2772022. View Article : Google Scholar
|
|
127
|
Gribben C, Galanakis V, Calderwood A,
Williams EC, Chazarra-Gil R, Larraz M, Frau C, Puengel T, Guillot
A, Rouhani FJ, et al: Acquisition of epithelial plasticity in human
chronic liver disease. Nature. 630:166–173. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Lu W, Bird TG, Boulter L, Tsuchiya A, Cole
AM, Hay T, Guest RV, Wojtacha D, Man TY, Mackinnon A, et al:
Hepatic progenitor cells of biliary origin with liver repopulation
capacity. Nat Cell Biol. 17:971–983. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Pu W, Zhang H, Huang X, Tian X, He L, Wang
Y, Zhang L, Liu Q, Li Y, Li Y, et al: Mfsd2a+ hepatocytes
repopulate the liver during injury and regeneration. Nat Commun.
7:133692016. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Tarlow BD, Finegold MJ and Grompe M:
Clonal tracing of Sox9+ liver progenitors in mouse oval cell
injury. Hepatology. 60:278–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Tarlow BD, Pelz C, Naugler WE, Wakefield
L, Wilson EM, Finegold MJ and Grompe M: Bipotential adult liver
progenitors are derived from chronically injured mature
hepatocytes. Cell Stem Cell. 15:605–618. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Raven A, Lu WY, Man TY, Ferreira-Gonzalez
S, O'Duibhir E, Dwyer BJ, Thomson JP, Meehan RR, Bogorad R,
Koteliansky V, et al: Cholangiocytes act as facultative liver stem
cells during impaired hepatocyte regeneration. Nature. 547:350–354.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Pradhan-Sundd T, Kosar K, Saggi H, Zhang
R, Vats R, Cornuet P, Green S, Singh S, Zeng G, Sundd P, et al:
Wnt/β-catenin signaling plays a protective role in the Mdr2
knockout murine model of cholestatic liver disease. Hepatology.
71:1732–1749. 2020. View Article : Google Scholar
|
|
134
|
Macchi F and Sadler KC: Unraveling the
epigenetic basis of liver development, regeneration and disease.
Trends Genet. 36:587–597. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Caldez MJ, Van Hul N, Koh H, Teo XQ, Fan
JJ, Tan PY, Dewhurst MR, Too PG, Talib S, Chiang BE, et al:
Metabolic remodeling during liver regeneration. Dev Cell.
47:425–438.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Li W, Li L and Hui L: Cell plasticity in
liver regeneration. Trends Cell Biol. 30:329–338. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Lambrecht R, Rudolf F, Uckert A, Sladky
VC, Phan TS, Jansen J, Naim S, Kaufmann T, Keogh A, Kirschnek S, et
al: Non-canonical BIM-regulated energy metabolism determines
drug-induced liver necrosis. Cell Death Differ. 31:119–131. 2024.
View Article : Google Scholar
|
|
138
|
Gazit V, Huang J, Weymann A and Rudnick
DA: Analysis of the role of hepatic ppargamma expression during
mouse liver regeneration. Hepatology. 56:1489–1498. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Jiao M, Ren F, Zhou L, Zhang X, Zhang L,
Wen T, Wei L, Wang X, Shi H, Bai L, et al: Peroxisome
proliferator-activated receptor alpha activation attenuates the
inflammatory response to protect the liver from acute failure by
promoting the autophagy pathway. Cell Death Dis. 5:e13972014.
View Article : Google Scholar
|
|
140
|
Fan S, Gao Y, Qu A, Jiang Y, Li H, Xie G,
Yao X, Yang X, Zhu S, Yagai T, et al: YAP-TEAD mediates PPAR
α-induced hepatomegaly and liver regeneration in mice. Hepatology.
75:74–88. 2022. View Article : Google Scholar
|
|
141
|
Kaji K, Factor VM, Andersen JB, Durkin ME,
Tomokuni A, Marquardt JU, Matter MS, Hoang T, Conner EA and
Thorgeirsson SS: Dnmt1 is a required genomic regulator for murine
liver histogenesis and regeneration. Hepatology. 64:582–598. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Snykers S, Henkens T, De Rop E, Vinken M,
Fraczek J, De Kock J, De Prins E, Geerts A, Rogiers V and Vanhaecke
T: Role of epigenetics in liver-specific gene transcription,
hepatocyte differentiation and stem cell reprogrammation. J
Hepatol. 51:187–211. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Shi Y, Sun H, Bao J, Zhou P, Zhang J, Li L
and Bu H: Activation of inactive hepatocytes through histone
acetylation: A mechanism for functional compensation after massive
loss of hepatocytes. Am J Pathol. 179:1138–1147. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Grimsrud PA, Carson JJ, Hebert AS, Hubler
SL, Niemi NM, Bailey DJ, Jochem A, Stapleton DS, Keller MP,
Westphall MS, et al: A quantitative map of the liver mitochondrial
phosphoproteome reveals posttranslational control of Ketogenesis.
Cell Metab. 16:672–683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Sugimoto A, Saito Y, Wang G, Sun Q, Yin C,
Lee KH, Geng Y, Rajbhandari P, Hernandez C, Steffani M, et al:
Hepatic stellate cells control liver zonation, size and functions
via R-spondin 3. Nature. 640:752–761. 2025. View Article : Google Scholar
|
|
146
|
Preziosi M, Okabe H, Poddar M, Singh S and
Monga SP: Endothelial Wnts regulate β-catenin signaling in murine
liver zonation and regeneration: A sequel to the Wnt-Wnt situation.
Hepatol Commun. 2:845–860. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Duan JL, Zhou ZY, Ruan B, Fang ZQ, Ding J,
Liu JJ, Song P, Xu H, Xu C, Yue ZS, et al: Notch-regulated
c-kit-positive liver sinusoidal endothelial cells contribute to
liver zonation and regeneration. Cell Mol Gastroenterol Hepatol.
13:1741–1756. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Li HY, Gao YX, Wu JC, Li JZ, Fu SW and Xu
MY: Single-cell transcriptome reveals a novel mechanism of
C-kit+-liver sinusoidal endothelial cells in NASH. Cell
Biosci. 14:312024. View Article : Google Scholar
|
|
149
|
Du W, Chen C, Liu Y, Quan H, Xu M, Liu J,
Song P, Fang Z, Yue Z, Xu H, et al: A combined 'eat me/don't eat
me' strategy based on exosome for acute liver injury treatment.
Cell Rep Med. 6:1020332025. View Article : Google Scholar
|
|
150
|
Rigual MDM, Angulo-Aguado M, Zagorac S,
Álvarez-Díaz R, Benítez-Mondéjar M, Yi F, Martínez-Garay C,
Santos-De-Frutos K, Kim E, Campos-Olivas R, et al: Macrophages
harness hepatocyte glutamate to boost liver regeneration. Nature.
641:1005–1016. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Hu M, Kurobe M, Jeong YJ, Fuerer C, Ghole
S, Nusse R and Sylvester KG: Wnt/beta-catenin signaling in murine
hepatic transit amplifying progenitor cells. Gastroenterology.
133:1579–1591. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Yimlamai D, Christodoulou C, Galli GG,
Yanger K, Pepe-Mooney B, Gurung B, Shrestha K, Cahan P, Stanger BZ
and Camargo FD: Hippo pathway activity influences liver cell fate.
Cell. 157:1324–1338. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Oh SH, Swiderska-Syn M, Jewell ML, Premont
RT and Diehl AM: Liver regeneration requires Yap1-TGFβ-dependent
epithelial-mesenchymal transition in hepatocytes. J Hepatol.
69:359–367. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Li L, Cui L, Lin P, Liu Z, Bao S, Ma X,
Nan H, Zhu W, Cen J, Mao Y, et al: Kupffer-cell-derived Ll-6 is
repurposed for hepatocyte dedifferentiation via activating
progenitor genes from injury-specific enhancers. Cell Stem Cell.
30:283–299. 2023. View Article : Google Scholar
|
|
155
|
Younossi ZM, Ratziu V, Loomba R, Rinella
M, Anstee QM, Goodman Z, Bedossa P, Geier A, Beckebaum S, Newsome
PN, et al: Obeticholic acid for the treatment of non-alcoholic
steatohepatitis: Interim analysis from a multicentre, randomised,
placebo-controlled phase 3 trial. Lancet. 394:2184–2196. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Sanyal AJ, Ling L, Beuers U, Depaoli AM,
Lieu HD, Harrison SA and Hirschfield GM: Potent suppression of
hydrophobic bile acids by aldafermin, an FGF19 analogue, across
metabolic and cholestatic liver diseases. JHEP Rep. 3:1002552021.
View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Kimura K, Ikoma A, Shibakawa M, Shimoda S,
Harada K, Saio M, Imamura J, Osawa Y, Kimura M, Nishikawa K, et al:
Safety, tolerability, and preliminary efficacy of the anti-fibrotic
small molecule PRI-724, a CBP/β-catenin inhibitor, in patients with
Hepatitis C Virus-related cirrhosis: A single-center, open-label,
dose escalation phase 1 trial. EBioMedicine. 23:79–87. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Francque SM, Hodge A, Boursier J, Younes
ZH, Rodriguez-Araujo G, Park GS, Alkhouri N and Abdelmalek MF:
Phase 2, open-label, rollover study of cenicriviroc for liver
fibrosis associated with metabolic dysfunction-associated
steatohepatitis. Hepatol Commun. 8:e3352024. View Article : Google Scholar
|
|
159
|
Geervliet E, Karkdijk E and Bansal R:
Inhibition of intrahepatic monocyte recruitment by cenicriviroc and
extracellular matrix degradation by mmp1 synergistically attenuate
liver inflammation and fibrogenesis in vivo. Sci Rep. 14:168972024.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Lurje I, Biegel DK, Zhang IW and Tacke F:
The role of galectin-3 in liver inflammation and fibrosis. J
Inflamm Res. 19:5726372026. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Chalasani N, Abdelmalek MF, Garcia-Tsao G,
Vuppalanchi R, Alkhouri N, Rinella M, Noureddin M, Pyko M, Shiffman
M, Sanyal A, et al: Effects of belapectin, an inhibitor of
galectin-3, in patients with nonalcoholic steatohepatitis with
cirrhosis and portal hypertension. Gastroenterology. 158:1334–1345.
2020. View Article : Google Scholar
|
|
162
|
Dhir RN, Dworakowski W, Thangavel C and
Shapiro BH: Sexually dimorphic regulation of hepatic isoforms of
human cytochrome p450 by growth hormone. J Pharmacol Exp Ther.
316:87–94. 2006. View Article : Google Scholar
|
|
163
|
Matz-Soja M, Berg T and Kietzmann T:
Sex-related variations in liver homeostasis and disease: From
zonation dynamics to clinical implications. J Hepatol. 84:181–193.
2026. View Article : Google Scholar
|
|
164
|
Sinha S, Ali Q, Zhang T, T-Nguyen DH,
Hanna S, Sethiadi J, Hissong E and Schwartz RE: Aging disrupts
hepatocyte zonation homeostasis in mice and humans. Hepatology.
83:1143–1157. 2026. View Article : Google Scholar
|
|
165
|
Zhang Y, Li Z, Gou H, Song X and Zhang L:
The gut microbiota-bile acid axis: A potential therapeutic target
for liver fibrosis. Front Cell Infect Microbiol. 12:9453682022.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Rizvi F, Everton E, Smith AR, Liu H, Osota
E, Beattie M, Tam Y, Pardi N, Weissman D and Gouon-Evans V: Murine
liver repair via transient activation of regenerative pathways in
hepatocytes using lipid nanoparticle-complexed nucleoside-modified
mrna. Nat Commun. 12:6132021. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Debacker AJ, Voutila J, Catley M, Blakey D
and Habib N: Delivery of oligonucleotides to the liver with GalNAc:
From research to registered therapeutic drug. Mol Ther.
28:1759–1771. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Yang Z, Song L, Chen H, Chen Y, Xie Y and
Xie J: Exploring the potential anticancer effects of lobelia
chinensis lour in liver cancer via multiomics analysis. Med Res.
1:483–488. 2025.
|
|
169
|
Dana J, Venkatasamy A, Saviano A,
Lupberger J, Hoshida Y, Vilgrain V, Nahon P, Reinhold C, Gallix B
and Baumert TF: Conventional and artificial intelligence-based
imaging for biomarker discovery in chronic liver disease. Hepatol
Int. 16:509–522. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Ghosh S, Zhao X, Alim M, Brudno M and Bhat
M: Artificial intelligence applied to 'omics data in liver disease:
Towards a personalised approach for diagnosis, prognosis and
treatment. Gut. 74:295–311. 2025. View Article : Google Scholar
|
|
171
|
Bradley CA: Liver: Spatial division of
hepatic metabolic labour. Nat Rev Gastroenterol Hepatol.
14:1392017. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Younossi ZM, Stepanova M, Nader F, Loomba
R, Anstee QM, Ratziu V, Harrison S, Sanyal AJ, Schattenberg JM,
Barritt AS, et al: Obeticholic acid impact on quality of life in
patients with nonalcoholic steatohepatitis: REGENERATE 18-month
interim analysis. Clin Gastroenterol Hepatol. 20:2050–2058. 2022.
View Article : Google Scholar
|
|
173
|
Kessoku T, Kobayashi T, Ozaki A, Iwaki M,
Honda Y, Ogawa Y, Imajo K, Saigusa Y, Yamamoto K, Yamanaka T, et
al: Rationale and design of a randomised, double-blind,
placebo-controlled, parallel-group, investigator-initiated phase 2a
study to investigate the efficacy and safety of elobixibat in
combination with cholestyramine for non-alcoholic fatty liver
disease. BMJ Open. 10:e379612020. View Article : Google Scholar
|
|
174
|
Noureddin M, Rinella M, Taub R, Labriola
D, Camacho RC, Alkhouri N, Loomba R and Bansal MB: Effects of
resmetirom on metabolic-dysfunction associated steatohepatitis in
patients with weight loss and/or diabetes taking glucagon-like
peptide-1 receptor agonists and other diabetes therapies: A
secondary analysis of the MAESTRO-NASH trial. Aliment Pharmacol
Ther. 62:1089–1099. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Nakajima A, Eguchi Y, Yoneda M, Imajo K,
Tamaki N, Suganami H, Nojima T, Tanigawa R, Iizuka M, Iida Y, et
al: Randomised clinical trial: Pemafibrate, a novel selective
peroxisome proliferator-activated receptor alpha modulator
(SPPARMα), versus placebo in patients with non-alcoholic fatty
liver disease. Aliment Pharmacol Ther. 54:1263–1277. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Cooreman MP, Butler J, Giugliano RP,
Zannad F, Dzen L, Huot-Marchand P, Baudin M, Beard DR, Junien J,
Broqua P, et al: The pan-PPAR agonist lanifibranor improves
cardiometabolic health in patients with metabolic
dysfunction-associated steatohepatitis. Nat Commun. 15:39622024.
View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Cheng HS, Tan WR, Low ZS, Marvalim C, Lee
JYH and Tan NS: Exploration and development of PPAR modulators in
health and disease: An update of clinical evidence. Int J Mol Sci.
20:50552019. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Nedrud MA, Chaudhry M, Middleton MS,
Moylan CA, Lerebours R, Luo S, Farjat A, Guy C, Loomba R,
Abdelmalek MF, et al: MRI quantification of placebo effect in
nonalcoholic steatohepatitis clinical trials. Radiology.
306:e2207432023. View Article : Google Scholar
|
|
179
|
Yoo C, Fabregat-Franco C, Lin C, Qvortrup
C, Oh D, Yau T, Kim H, Castet F, Ponz Sarvise M, Hsu C, et al:
First-in-human phase 1 study of RO7119929, an oral TLR7 agonist
prodrug, in patients with advanced primary or metastatic liver
cancers. J Immunother Cancer. 14:e127832026. View Article : Google Scholar
|