Introduction
Metabolic diseases, such as obesity and diabetes,
represent major public health challenges worldwide due to their
increasing prevalence and associated complications (1). These diseases are complex and
multifactorial, involving intricate interactions between genetic,
environmental and lifestyle factors (2). One emerging area of interest in the
study of metabolic diseases is the role of primary cilia, which are
small, microtubule-based organelles that extend from the surface of
most vertebrate cells (3).
Historically viewed as vestigial, primary cilia are now recognized
as critical sensory and signaling hubs that regulate a variety of
cellular processes essential for maintaining homeostasis (4).
Primary cilia play a vital role in the function of
several key metabolic tissues, including the hypothalamus,
pancreatic islets, liver and adipose tissue (5-8).
In the hypothalamus, cilia are involved in sensing and integrating
hormonal signals that regulate appetite and energy balance.
Disruption of ciliary function in hypothalamic neurons can lead to
dysregulation of appetite control, resulting in conditions such as
hyperphagia and obesity (9). In
the pancreas, primary cilia are essential for the proper
functioning of beta cells, which are responsible for insulin
secretion (10). Ciliary
dysfunction in these cells can impair insulin release and
contribute to the development of diabetes. In addition to their
roles in hypothalamic and pancreatic function, primary cilia are
also crucial for the regulation of lipid metabolism (11). In adipose tissue, cilia-mediated
signaling pathways influence the storage and mobilization of fat.
Disruption of these pathways can lead to abnormal fat accumulation
and inflammation, which are hallmarks of obesity (12). Similarly, in the liver, primary
cilia are involved in regulating bile flow and cholesterol
metabolism (13). Impaired
ciliary function in hepatic cells can exacerbate metabolic
imbalances, further complicating the clinical management of
metabolic diseases.
Recent research has highlighted the significant
impact of ciliary dysfunction on bone health, particularly in the
context of diabetes (14).
Diabetes is known to impair bone healing and increase the risk of
fractures, with evidence suggesting that ciliary defects in
osteoblasts contribute to these complications. Ciliary dysfunction
in osteoblasts impairs bone formation and mineralization, leading
to weaker bones and delayed fracture healing (15). This underscores the broader
implications of ciliary dysfunction beyond traditional metabolic
tissues and highlights the interconnected nature of metabolic and
skeletal health.
Understanding the mechanisms by which primary cilia
influence metabolic processes is crucial for developing targeted
therapies for metabolic diseases. Current therapeutic approaches
often focus on managing symptoms rather than addressing underlying
causes. However, emerging treatments that aim to restore ciliary
function offer promising new avenues for intervention. These
include gene therapy techniques to correct genetic defects
affecting ciliogenesis, as well as small molecules that enhance
ciliary signaling pathways.
This review explored the multifaceted roles of
primary cilia in metabolic regulation and the impact of ciliary
dysfunction on metabolic health. The evidence linking ciliary
defects to obesity, diabetes and their complications were examined
and potential therapeutic strategies aimed at restoring ciliary
function were discussed. By shedding light on this underrecognized
aspect of metabolic disease, the study esteems to pave the way for
innovative treatments that improve clinical outcomes and enhance
the quality of life for individuals affected by these
conditions.
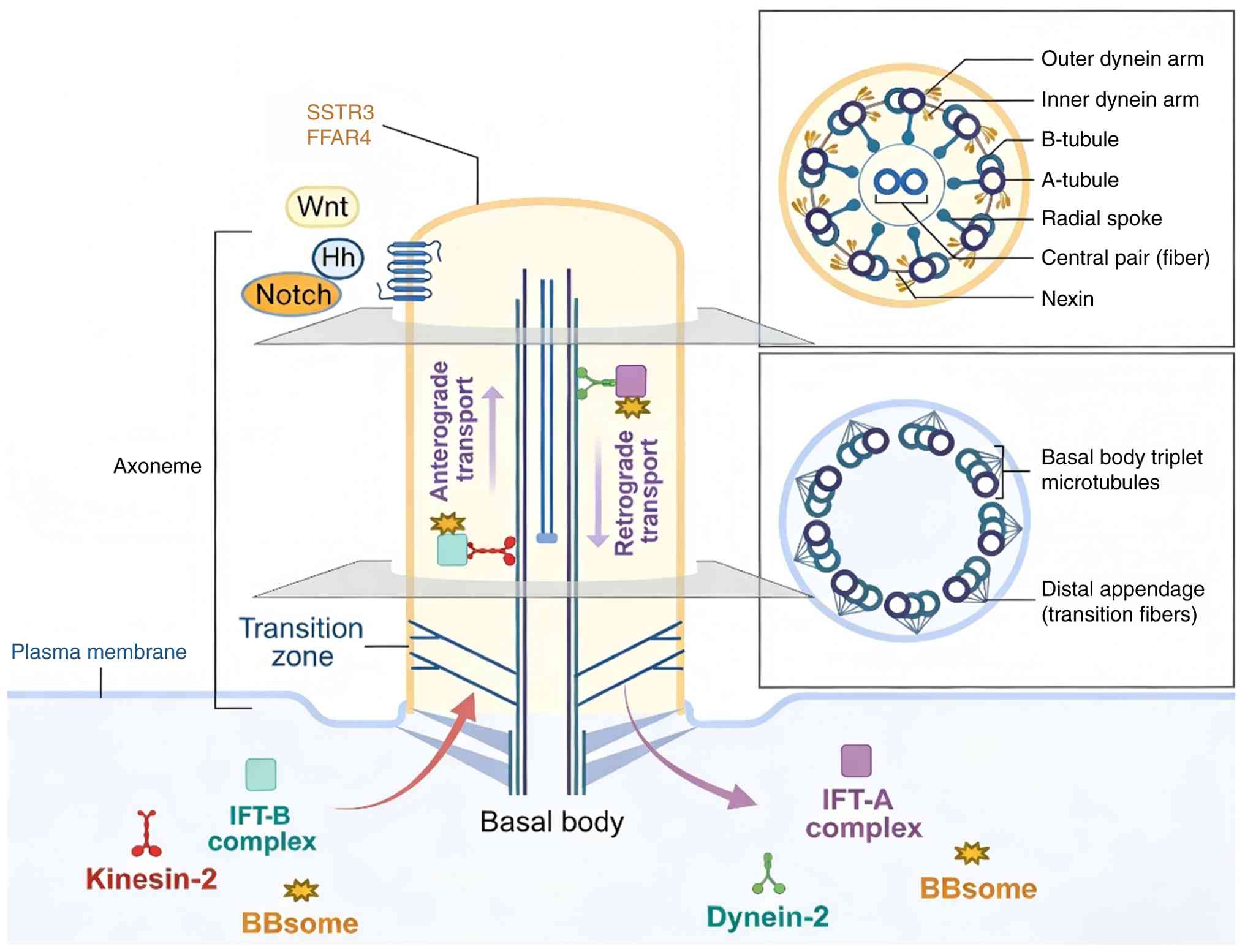
Cilia structure and function
Cilia are slender, microtubule-based projections
ubiquitously distributed on the surfaces of nearly all mammalian
cells (16). They are
categorized into two primary structural types: Motile cilia and
primary (or sensory) cilia (17). Motile cilia are found abundantly
present on specialized, multiciliated cells, where they facilitate
fluid movement or cell locomotion (18). Conversely, most cells feature a
single, non-motile primary cilium that plays a crucial role in
sensory signaling. Despite their lack of movement, primary cilia
are central to orchestrating key regulatory pathways involved in
growth, development and cellular homeostasis (19).
Architecture of primary and motile
cilia
The basic ciliary architecture consists of an
axoneme core of microtubule doublets surrounded by a membrane. The
axoneme extends from a modified centriole termed the basal body,
which migrates and docks at the cell surface to nucleate the cilium
(20). All cilia feature a ring
of nine peripheral microtubule doublets in the axoneme. In motile
cilia, the axoneme also contains a central pair of microtubules,
producing a 9+2 configuration. This central pair enables ATP-driven
motor protein dynein to generate motility for fluid flow or cell
movement (21). By contrast,
primary cilia lack the central pair, resulting in a 9+0 axoneme
that renders them immotile.
The transition zone, located at the ciliary base,
acts as a selective gateway that regulates protein access to the
ciliary compartment. Transition fibers anchor both the basal body
and the transition zone to the cellular membrane. This zone
features Y-shaped structures that connect the axoneme to the
ciliary membrane, playing a crucial role in controlling protein
trafficking (22). Notably, the
composition of the ciliary membrane is distinct from that of the
general cell surface. The importation of proteins into the cilium
is mediated by intraflagellar transport (IFT), which utilizes the
axoneme as a track. IFT is instrumental in the assembly and
maintenance of cilia, actively transporting structural components,
signaling molecules and receptors to and from the ciliary
compartment (Fig. 1) (23,24).
IFT and ciliogenesis
IFT is essential for the bidirectional movement of
cargo proteins along the axoneme, a process powered by the motor
proteins kinesin and dynein, which are part of multiprotein IFT
complexes (25,26). Anterograde transport, facilitated
by kinesin-2 family motors, moves cargos from the base to the tip
of the cilium, promoting ciliary assembly. Conversely, retrograde
transport, executed by cytoplasmic dynein, brings proteins back to
the cell body for recycling or degradation. The subunit, a crucial
scaffolding component of the IFT particle complex B, is
indispensable for ciliogenesis. The absence of IFT proteins halts
cilia formation and leads to ciliopathies (27).
The ciliogenesis process begins with the
transformation of the mother centriole into the basal body, which
then migrates and docks at the membrane to nucleate the axoneme
(28). Distal appendages on the
mature basal body facilitate membrane attachment and serve as
docking sites for transition fibers. The docking of the basal body
relies on the distal appendage centrosomal protein 164 (CEP164),
while appendage proteins CEP83 and are involved in recruiting
receptors that regulate vesicular trafficking to the ciliary base
(29-31). Following docking, transition
fibers extend from the basal body, setting the stage for axoneme
growth through IFT-driven processes.
Signaling pathways coordinated by primary
cilia
Primary cilia act as cellular antennas, extending
into the extracellular environment to detect mechanical and
chemical signals, including growth factors, nutrients, lipids,
odorants, light and fluid flow. These signals are then translated
into cellular responses, regulating vital developmental and
homeostatic pathways such as cell growth, proliferation,
differentiation, migration, polarity and tissue morphogenesis
(32). Notable signaling
pathways modulated by primary cilia include Hedgehog (Hh), Wnt,
Notch, Hippo, G protein-coupled receptors (GPCR), mTOR,
transforming growth factor (TGF)β, receptor tyrosine kinase (RTK)
and planar cell polarity pathways.
Hh signaling
The Hh signaling pathway is significantly modulated
by primary cilia (33). Cilia
provide a specialized compartment that aids in the processing of
Glioma-associated oncogene homolog (Gli) the family of Zn-finger
transcription factors, which are crucial for the transduction of Hh
signals. In the absence of Hh ligands such as Sonic Hh (Shh), Gli2
and Gli3 are processed into their activator and repressor forms via
proteolytic cleavage (34). This
processing is facilitated by the ciliary kinase protein kinase A
(PKA), the scaffolding protein kinesin family member 7 (Kif7) and a
protease complex. The binding of Shh to its receptor, patched 1
(PTCH1) located on the cilia, alleviates the inhibition of
Smoothened (SMO) by PTCH1, leading to the accumulation of SMO
within the cilia (35). This
accumulation prevents the proteolytic processing of Gli proteins,
enriching the Gli2/3 activator form and triggering the expression
of Hh target genes.
Wnt signaling
Primary cilia also influence Wnt signaling, a
pathway that involves the regulated proteolysis of the
transcription co-activator β-catenin (36). The ciliary protein Kif3a plays a
role in inhibiting β-catenin signaling by facilitating its
degradation (37). Consequently,
the loss of cilia results in enhanced Wnt signaling activity. The
interplay between Hh and Wnt signaling is pivotal in determining
cell fate during developmental processes. Furthermore, cilia
contribute to the orientation of the mitotic spindle and the
regulation of oriented cell divisions through planar cell polarity
signaling (38).
mTOR and Yes-associated protein
(YAP)/transcriptional coactivator with PDZ-binding motif (TAZ)
The master regulator of metabolism, mTOR, is
localized to primary cilia and plays a role in regulating ciliary
length (39). mTOR supports
ciliogenesis and the docking of ciliary vesicles. Of note, the
length of cilia can influence mTOR activity by modulating the
expression of its inhibitors, such as the DEP domain-containing
mTOR-interacting protein, an endogenous suppressor of mTOR
signaling. Additionally, the Hippo pathway effectors YAP/TAZ are
recruited to primary cilia under conditions of serum starvation
(40). This recruitment inhibits
the activity of YAP/TAZ, thereby limiting cell proliferation.
Ciliogenesis is promoted, whereas deciliation activates
YAP/TAZ-dependent transcription, highlighting the complex interplay
between cilia and cellular signaling pathways (41).
RTKs
Primary cilia serve as a niche for growth factor
receptors such as platelet-derived growth factor (PDGF),
insulin-like growth factor (IGF) and fibroblast growth factor,
facilitating the initiation of intracellular signaling cascades
that govern cell proliferation and differentiation. Notably, the
PDGF receptor α is specifically localized to the primary cilia of
fibroblasts, where ligand binding triggers MEK/ERK and mTOR
signaling pathways (42).
Disruptions in PDGF signaling are linked to ciliopathy phenotypes,
highlighting the unique role of ciliary compartmentalization in
modulating RTK signaling, potentially eliciting distinct cellular
responses compared to activation at the cell surface (43).
GPCRs
GPCRs, pivotal in detecting extracellular signals,
are enriched within the membranes of cilia (44). An example includes the
somatostatin receptor 3 (SSTR3), which is preferentially localized
to neuronal cilia (45).
Activation of ciliary GPCRs can induce localized Ca2+
signaling, underscoring the importance of ciliary
compartmentalization in achieving signaling specificity through
subtype-selective GPCR localization (46). Furthermore, cilia facilitate the
integration of multiple signaling pathways, including Wnt, Hedgehog
and RTK, by housing components of these pathways and enabling
crosstalk with GPCRs (47).
TGFβ superfamily signaling
Cilia influence the TGFβ superfamily's signaling
mechanisms via receptor complexes comprising type I and II
serine/threonine kinases (48).
Localization of TGFβ receptors to cilia enhances ligand-mediated
activation, with ciliary loss leading to impaired signaling. This
cilia-mediated control is crucial for regulating developmental and
homeostatic processes. The ciliary protein Tectonic plays a
significant role in mediating TGFβ signals, interacting with Shh
and canonical Wnt pathways (49,50).
Notch signaling
Primary cilia enhance Notch signaling pathway
activation through the regulated intramembrane proteolysis of Notch
receptors, which releases an intracellular fragment that activates
gene transcription (51). Both
Notch receptors and their ligands, such as Delta, are localized to
cilia, emphasizing the cilia's role in facilitating Notch pathway
processing. The disruption of ciliogenesis adversely affects
Notch-dependent developmental processes, illustrating the
functional significance of ciliary regulation in signal
transduction (52).
Primary cilia in immune cells
Recent research has definitively identified
functional primary cilia within the immune system. A study utilized
an Arl13b - mCherry transgenic mouse model, where the ciliary
membrane protein Arl13b is fused to the red fluorescent protein
mCherry for specific labeling, revealed that 13.4±0.9% of spleen
cells and 20.3±0.9% of thymus cells possess primary cilia measuring
3-6 µm in length (53).
This challenges the long-standing paradigm that lymphocytes are
non-ciliated. While the specific immune cell subtypes that are
ciliated require further characterization, this finding implies a
conserved role for ciliary architecture and IFT in immune cells.
The presence of these signaling organelles suggests they may serve
as specialized sensory platforms in lymphoid tissues, potentially
detecting local cytokines, pathogens or metabolic signals to
modulate immune responses. This expands the physiological landscape
of cilia-mediated signaling into immunoregulation, with possible
implications for inflammatory aspects of metabolic diseases
(53,54).
Role of cilia in lipid metabolism
diseases
Cilia in obesity
Appetite regulation
Arcuate nucleus pro-opiomelanocortin (POMC) neurons
possess primary cilia interacting with melanocortin 4 receptor
(MC4R) to regulate satiety signaling (5,55). Ablating IFT88 in POMC neurons
caused cilia loss, reduced POMC firing and decreased α-melanocyte
stimulating hormone (α-MSH) release (5). Further analysis found altered
trafficking and dysfunctional signaling of the MC4R satiety
receptor in cilia-defective POMC neurons. This caused impaired POMC
neuron response to α-MSH and defective propagation of satiety
signals to downstream nuclei like the paraventricular nucleus. MC4R
activation normally induces membrane depolarization, action
potential firing and α-MSH secretion in POMC neurons (55). However, optogenetic and
electrophysiology experiments revealed loss of these MC4R responses
in POMC neurons lacking cilia. This MC4R/POMC signaling axis is
critical for appetite regulation (56). Accordingly, disrupting the axis
through cilia defects caused hyperphagia, reduced energy
expenditure and obesity in mice (Fig. 2).
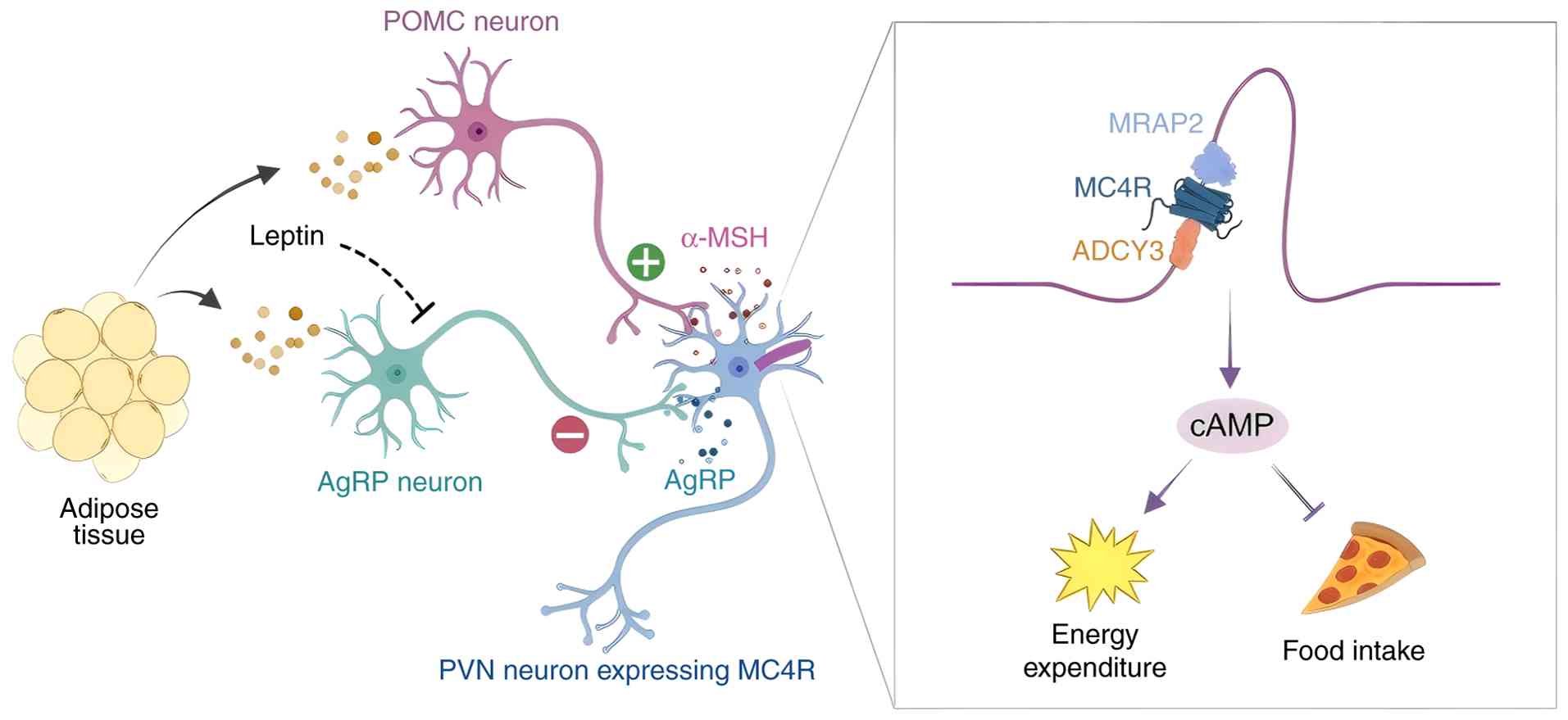 | Figure 2Role of cilia in regulation of food
intake and energy expenditure. The arcuate nucleus is composed of
different types of ciliary neurons, including anorexigenic neurons
expressing POMC and orexigenic neurons expressing AgRP. Leptin,
secreted by adipose tissue, activates leptin receptors in the
arcuate nucleus, where it increases the transcription of POMC and
decreases the transcription of AgRP. POMC-expressing neurons are
activated to accelerate the secretion of α-MSH, which activates
MC4R in the PVN, leading to decreased food intake and increased
energy expenditure. PVN, paraventricular nucleus; POMC neuron,
pro-opiomelanocortin neuron; AgRP neuron, agouti-related peptide
neuron; α-MSH, α-melanocyte-stimulating hormone; PVN,
paraventricular nucleus; MC4R, melanocortin 4 receptor; MRAP2,
melanocortin 2 receptor accessory protein 2; ADCY3, adenylate
cyclase 3; cAMP, cyclic adenosine monophosphate. |
Obesity is commonly linked with the aging process.
The MC4R is crucial in the hypothalamic leptin-melanocortin pathway
for combating obesity (57).
Research indicates that primary cilia containing MC4R in
hypothalamic neurons of rats progressively shorten with age, which
correlates with metabolic decline and increased fat accumulation.
This phenomenon, a condition termed 'age-associated ciliopathy' is
a progressive functional decline in a specific tissue or organ
system. This decline results from the deterioration of primary
cilia structure, which occurs as a direct consequence of the
biological aging process. It is driven by the upregulation of
leptin-melanocortin signaling due to excessive nutrient intake.
However, dietary restriction or inhibition of
ciliogenesis-associated kinase 1 can alleviate or reverse this
condition. Genetically induced shortening of MC4R-bearing cilia in
hypothalamic neurons results in decreased responsiveness to
melanocortin, leading to reduced thermogenesis in brown adipose
tissue, lower energy expenditure, increased appetite and subsequent
obesity and leptin resistance (58). Therefore, while leptin exerts
anti-obesity effects in the short term, prolonged
leptin-melanocortin signaling contributes to the age-related
shortening of MC4R cilia, heightening the risk of obesity.
Ventral medial hypothalamus SF1 neurons also require
primary cilia for regulating food intake (59). SF1 neuronal cilia modulate
activity and calcium dynamics through ATP/P2Y purinergic receptor
and γ-aminobutyric acid type B receptor signaling (60). Selectively ablating IFT88 in SF1
neurons abolished cilia and altered calcium oscillations. This
caused overactivity of SF1 neurons, hyperphagia, reduced energy
utilization and pronounced diet-induced obesity (61). Further characterization found
changes in genes regulating feeding and metabolism, such as agouti
related peptide (AGRP), melanin concentrating hormone and cocaine
and amphetamine regulated transcript (62). Thus, ciliary signaling regulates
hunger/satiety neuronal function to suppress overeating and weight
gain.
Olfactory cilia detect food odors, with their
ablation causing hyperphagia (63). Deleting IFT88 eliminates
olfactory cilia and receptors like cyclic nucleotide-gated channel
α2, a core ion channel essential for olfactory signal transduction,
disrupting odor detection and discrimination (64). Electro-olfactogram recordings
confirmed loss of odor-evoked responses. Further RNA sequencing
revealed reduced expression of adenylate cyclase 3 and other
olfactory signaling molecules. Behaviorally, mice lacking olfactory
cilia exhibited hyperphagia, glucose intolerance, increased
adiposity, liver steatosis and weight gain on a high-fat diet. This
demonstrates the key role of olfactory cilia in controlling food
seeking behaviors that impact energy balance and obesity
susceptibility (65).
Tetratricopeptide repeat domain 21B (THM1) (also
known as TTC21B or IFT139) encodes a component of the intraciliary
transport-A complex and mutations in this gene are present in 5% of
patients with ciliary diseases (66). Research has demonstrated that
adult mice lacking Thm1 develop obesity, diabetes, hypertension and
fatty liver, with notable sex differences in susceptibility to
weight gain and metabolic dysfunction (67). When Thm1 conditional knockout
mice were pair-fed with control mice, obesity and related disorders
were prevented, indicating that excessive food intake contributes
to the obesity phenotype. Thm1 knockdown leads to increased
localization of adenylyl cyclase III in primary cilia, resulting in
cilia with shortened, bulbous distal tips on neurons in the
hypothalamic arcuate nucleus, a key center for regulating feeding
and activity. In pre-obese Thm1 conditional knockout mice,
expression of the anorexigenic hormone POMC in the arcuate nucleus
was reduced by 50%, contributing to hyperphagia (68). Fasting in these mice did not
alter the expression of POMC or the Agrp, suggesting an impaired
ability to perceive peripheral signaling changes (69). Therefore, ciliary defects due to
THM1 gene mutations reduce sensitivity to ingestive signals,
disrupting appetite regulation and leading to hyperphagia, obesity
and metabolic disorders.
The PI3K-3 phosphoinositide dependent protein kinase
1 (PDK1)-forkhead box (Fox)O1 signaling pathway plays a role in
regulating energy homeostasis. When unphosphorylated by PDK1, FoxO1
is active and located in the nucleus, where it represses POMC
expression and stimulates Agrp expression (70). Activation of leptin and insulin
results in the phosphorylation of FoxO1 and its export from the
nucleus, allowing STAT3 to bind to the POMC and Agrp promoters,
which activates POMC neurons and inhibits AgRP neurons (71). In conditions of excess energy,
leptin signaling activates mTOR, promoting phosphorylation and
inhibiting adenosine monophosphate-activated protein kinase (AMPK),
thereby suppressing food intake (72).
Adipogenesis
Primary cilia play a pivotal role in suppressing
adipocyte differentiation by modulating Hh pathway signaling.
Specifically, the transcriptional repressor is a form of Gli3
(Gli3R) inhibits the expression of peroxisome
proliferator-activated receptor γ (PPARγ), a key regulator of
adipogenesis (73). Disruption
of ciliogenesis through the knockdown of intraflagellar transport
protein IFT88 or other ciliary genes results in the absence of
primary cilia, preventing the formation of Gli3R (74). Consequently, this leads to the
derepression of the PPARγ promoter. RNA interference targeting
IFT88 in 3T3-L1 preadipocytes has shown a significant upregulation
of PPARγ expression in the absence of cilia, facilitating enhanced
differentiation into mature adipocytes (75). This is evidenced by increased
lipid droplet formation, as indicated by Oil Red O staining, and
elevated expression of adipogenic markers such as fatty acid
binding protein 4, lipoprotein lipase, adiponectin and C/EBPα, as
confirmed by quantitative PCR and immunoblot analyses (8). Furthermore, the activation of Hh
signaling using the agonist purmorphamine or by expressing Gli3R
can mitigate the accelerated adipogenesis resulting from ciliary
loss.
Adult mesenchymal stem cells, including
preadipocytes, have a cellular sensory structure called primary
cilia. Preadipocytes with these cilia are prevalent around blood
vessels in adipose tissue and become active in response to high-fat
diets (76). Disruption of cilia
in mouse preadipocytes significantly affects the expansion of white
adipose tissue. Researchers have identified that the localization
of the omega-3 fatty acid receptor free fatty acid receptor 4/G
protein-coupled receptor 120 (FFAR4/GPR120) within cilia, which
promotes adipogenesis, depends on tubby-like protein 3, an
essential adaptor of the IFT-A complex that mediates the ciliary
entry of specific membrane proteins. FFAR4 agonists and omega-3
fatty acids, but not saturated fatty acids, stimulate mitogenesis
and adipogenesis by rapidly increasing cyclic (c)AMP production
within cilia, resulting in the phosphorylation and nuclear
translocation of CCCTC-binding factor (CTCF). Within the nucleus,
phosphorylated CTCF acts as a scaffold, recruiting the histone
acetyltransferase p300 to form an activation complex at the
regulatory regions of key adipogenic genes, such as PPARγ and
CCAAT/enhancer-binding protein α (CEBPα). P300 catalyzes local
histone hyperacetylation, opening the chromatin. This facilitates
the binding of the transcriptional machinery and adipogenic
factors, including PPARγ and CEBPα, creating a positive feedback
loop, which initiates adipogenesis (8). Dietary omega-3 fatty acids
specifically boost the number of adipocytes, facilitating the
creation of new adipocytes and promoting the storage of saturated
fatty acids to maintain a healthy balance of adipose tissue
(8) (Fig. 3).
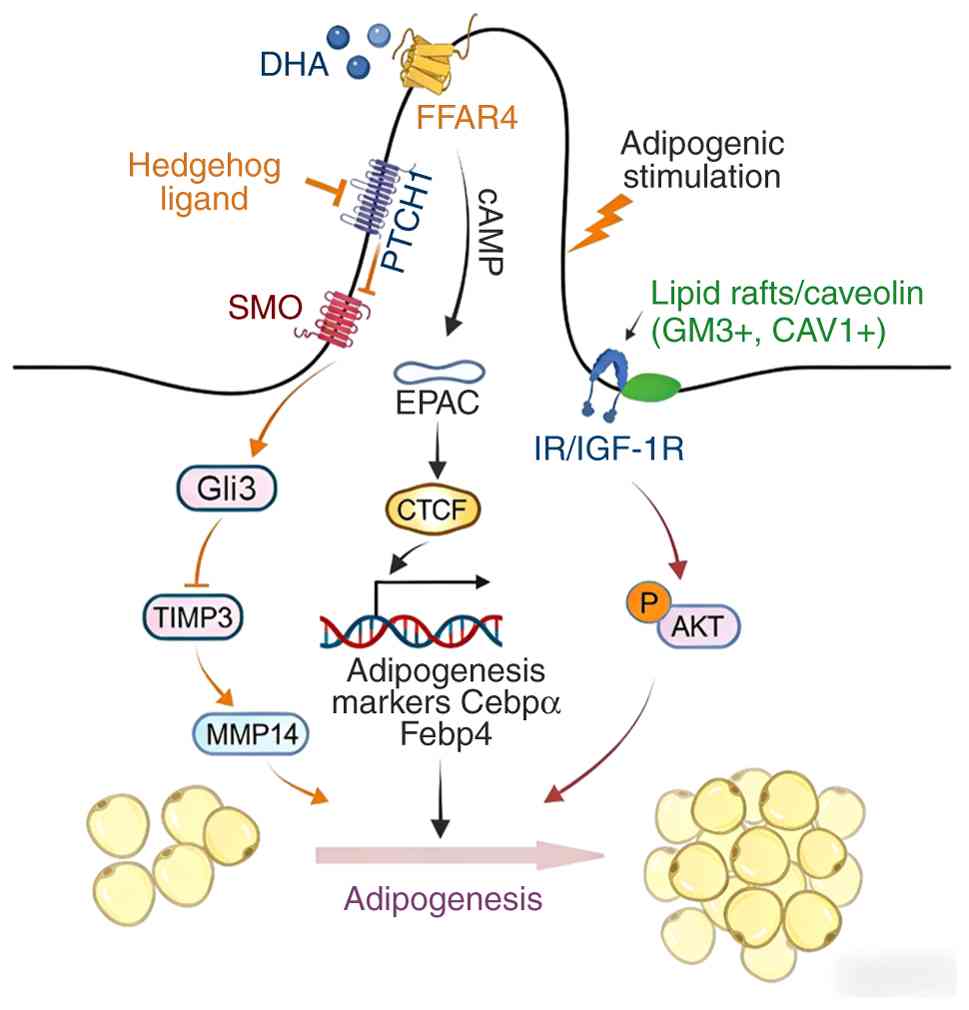 | Figure 3Role of cilia in adipogenesis. DHA
fatty acids activate FFAR4 on cilia, leading to an increase in cAMP
levels and the activation of EPAC signaling. EPAC signaling, in
turn, leads to CTCF-dependent recruitment of p300 to the PPARγ and
CEBPα loci, thereby facilitating chromatin remodeling. This process
results in the adipose transcriptional activation of PPARγ and
C/EBPα, which upregulate the expression of adipose synthesis genes
and promote the differentiation of preadipocytes into adipocytes.
In regions where IR (and possibly IGF1R) is present around the base
of the cilium, adipogenic stimuli induce the aggregation of lipid
rafts containing CAV1 and GM3. This aggregation activates the
IR/IGF1R-Akt cascade, thereby promoting adipogenesis. In addition,
the Hh signaling pathway regulates the expression of TIMP3 through
the transcription factor Ci/Gli. Gli3R is a transcriptional
repressor from the Gli family of proteins that can inhibit TIMP3,
allowing MMP14 to be active. MMP14 promotes adipogenesis through
mechanisms involving the activation of the master regulators PPARγ
and C/EBPα. DHA, docosahexaenoic acid; FFAR4, free fatty acid
receptor 4; PTCH1, patched 1; SMO, smoothened; GLI3, GLI family
zinc finger protein 3; TIMP3, tissue inhibitor of
metalloproteinases 3; MMP14, matrix metalloproteinase 14; cAMP,
cyclic adenosine monophosphate; EPAC, exchange protein directly
activated by cAMP; C/EBPα, CCAAT/enhancer-binding protein α; FABP4,
fatty acid-binding protein 4; IR/IGF-1R, insulin
receptor/insulin-like growth factor 1 receptor; GM3,
monosialodihexosylganglioside; CAV1, caveolin-1. |
Fas-binding factor 1 (FBF1) is a core component of
ciliary transition fibers that mediates stress-induced
cilium-to-promyelocytic leukemia protein-nuclear body signaling to
initiate cellular senescence (77). Of note, the absence of FBF1
results in a notable paradox: Fbf1tm1a/tm1a mice, which are a
genetically modified model carrying a 'knockout-first' tm1a allele
in the FbF1 locus, despite progressively becoming obese, do not
develop adverse metabolic complications throughout their lifespan.
The lack of FBF1 leads to the upregulation of the ciliary program
in white adipose tissue through an A-kinase anchoring protein 9
(AKAP9)-dependent, cilia-regulated PKA signaling pathway. AKAP9, a
scaffold protein enriched at the ciliary base, anchors PKA and
facilitates its compartmentalized activation (78). Additionally, it significantly
enhances adipogenesis through Hh signaling regulated by
Bardet-Biedl syndrome proteins. The combined effect of increased
ciliogenesis and expansion of adipose tissue highlights a potential
explanation for the phenomenon of 'healthy obesity' observed in
Fbf1tm1a/tm1a mice (78). This
study suggests that targeting preadipocyte cilia could represent a
promising strategy for combating metabolic disorders.
Injured skeletal muscle has the ability to
regenerate; however, with aging or in the case of muscular
dystrophies, muscle tissue tends to be replaced by fat. Following
injury, fibro/adipogenic progenitors (FAPs) within the muscle begin
to proliferate and differentiate into adipocytes (79). These FAPs actively form primary
cilia, which mediate intercellular communication, including Hh
signaling. Studies have shown that genetic ablation of cilia in
FAPs inhibits intramuscular fat formation both after injury and in
a mouse model of Duchenne muscular dystrophy. Furthermore,
preventing FAP ciliation not only improved myofiber regeneration
post-injury but also mitigated the reduction in myofiber size
observed in muscular dystrophy models (80). Hh signaling via FAP cilia
controls the expression of tissue inhibitor of metalloproteinases 3
(TIMP3), a secreted inhibitor of metalloproteinases (MMPs), which
in turn suppresses MMP14 activity to prevent adipogenesis.
Additionally, a pharmacological agent that mimics TIMP3 effectively
blocked the differentiation of FAPs into adipocytes, suggesting a
potential therapeutic approach to counteract the fatty degeneration
of skeletal muscle (80). Thus,
it may be concluded that Hh signaling through FAP cilia is crucial
in regulating the regenerative processes of skeletal muscle in
response to injury.
Research has shown that C3H10T1/2 mesenchymal
progenitor cells, crucial for cilia formation, exhibit
significantly longer cilia compared to control cells and are unable
to differentiate into adipocytes (81). The elongated cilia hinder the
accumulation of caveolin-1 and/or monosialodihexosylganglioside
(GM3)-positive lipid rafts surrounding the ciliary base. Caveolin-1
is a scaffold protein essential for caveolae formation and insulin
receptor stabilization, and GM3 is a ganglioside enriched in lipid
rafts that promotes insulin receptor retention, which are all
critical components involved in the compartmentalization of insulin
signaling. Impaired accumulation of these components at the ciliary
base disrupts local lipid raft microdomains, leading to the
accumulation of insulin receptor proteins at the ciliary base and
subsequent inhibition of insulin-Akt signaling (82). In trichostatin knockout mice,
adipogenic progenitor cells have elongated cilia, which disrupts
lipid raft dynamics. Knockout mice fed a chronically high-fat diet
displayed reduced body fat and smaller adipocytes compared to
wild-type mice (82). Therefore,
primary cilia may play a role in regulating adipogenic signaling by
controlling lipid raft dynamics around the cilia (Fig. 3).
Glucose homeostasis
Primary cilia on pancreatic islet cells are pivotal
for the maintenance of normal glucose homeostasis, with defects in
cilia contributing to impaired insulin secretion and the
exacerbation of metabolic dysfunction associated with obesity
(83,84). These non-motile cilia are present
on all types of islet endocrine cells, including insulin-producing
β-cells, glucagon-secreting α-cells, somatostatin-releasing
δ-cells, and pancreatic polypeptide-containing PP cells. PP cells
are a distinct islet endocrine subtype that regulates exocrine and
endocrine pancreatic activity via autocrine and paracrine
signaling. These cilia project into the central lumen of the
pancreatic ducts (85).
Positioned strategically, cilia play a critical role
in sensing fluid flow and modulating Ca2+ signaling
dynamics, crucial for stimulus-secretion coupling in β-cells
(86). The influx of
Ca2+ in response to glucose metabolism acts as a
fundamental trigger for the biphasic secretion of insulin.
Experimental deletion of the intraflagellar transport protein
IFT88, or disruption of other genes involved in ciliogenesis
specifically within β-cells, has been shown to impair
glucose-induced Ca2+ influx. Such perturbation in
Ca2+ signaling leads to the loss of both first and
second-phase insulin releases in response to glucose, as
demonstrated in static islet incubations (87). This diminished insulin secretion
capacity was corroborated in vivo through observed impaired
glucose tolerance tests in conditional IFT88 knockout mice.
Furthermore, islet cilia enhance insulin secretion
by facilitating the proper localization of insulin secretory
granules at the plasma membrane (83). The absence of cilia disrupts the
normal clustering of insulin granules near the ciliary base, a
region where they are primed for glucose-responsive exocytosis.
Super-resolution imaging techniques have highlighted the
preferential localization of granules in proximity to ciliary bases
(88). The absence of cilia
interrupts the trafficking of these granules along microtubules,
leading to aberrant docking and compromised insulin release.
Additionally, primary cilia are implicated in supporting exosome
secretion, a process that aids in the trafficking of secretory
granules, thereby optimizing insulin secretion (80,89). Cilia have been shown to support
exosome secretion from β-cells by acting as the trigger for a
specific paracrine signaling cascade. They ensure that the specific
signaling receptors, particularly Ephrin type-A receptor 2 (EphA2),
are selectively packaged into exosomes to regulate the function of
neighboring cells. Cilia defects prevent EphA2 from ever reaching
the endosomal sorting complexes required for transport pathway, a
conserved membrane trafficking system that mediates the sorting,
degradation, or recycling of ubiquitinated membrane proteins
(87). This impairment in an
auxiliary secretion route likely further aggravates deficits in
granule docking and release.
In summary, primary cilia orchestrate multiple
aspects of stimulus-secretion coupling, including Ca2+
signaling dynamics, the trafficking and docking of insulin granules
and exosome functionality. The disruption of these processes by
diabetes and obesity-induced cilia defects significantly
compromises insulin release and glucose regulation, propagating a
detrimental cycle where hyperglycemia induces further cilia loss,
exacerbating insulin secretion impairment and amplifying metabolic
disease challenges. Unraveling these intricate mechanisms sheds
light on new avenues for understanding β-cell dysfunction and its
relevance to metabolic diseases.
Inflammation and metabolic
homeostasis
Primary cilia play a crucial role in suppressing
inflammatory pathways, the dysregulation of which can disrupt
metabolic homeostasis and contribute to the development of obesity
(90). Notably, specific
hypothalamic neurons, equipped with primary cilia, modulate
inflammation through signaling mediated by somatostatin and leptin
receptors (91). The SSTR3,
selectively localized to neuronal cilia, is a Gai-coupled receptor,
inhibits adenylyl cyclase, and reduces cAMP levels and PKA
activity. The suppression of the cAMP/PKA pathway which alters the
availability and function of the transcriptional co - activator
CBP/p300. CBP/p300 is a pair of highly homologous histone
acetyltransferases that facilitate gene activation by acetylating
histones and recruiting transcription machinery. This suppression
inhibits NF-κB - mediated cytokine production (92,93). The absence of SSTR3 in
hypothalamic arcuate nucleus POMC neurons amplifies the
inflammatory response to lipopolysaccharides, leading to increased
release of cytokines such as IL-6, IL-1β and TNFα. Furthermore,
SSTR3 activation within POMC neuron cilia curtails
inflammation-induced ceramide production and endoplasmic reticulum
stress, underscoring the anti-inflammatory effects vital for
preserving the normal function and satiety signaling of POMC
neurons. Disruption of this ciliary SSTR3 pathway perturbs satiety
signaling and feeding behaviors, impacting energy balance (94).
In the context of obesity, hyperleptinemia triggers
the relocalization of the leptin receptor (LepR) away from neuronal
cilia within the mediobasal hypothalamus (MBH), impairing leptin
sensing (58). This alteration
in LepR trafficking, evidenced through immunofluorescence staining
and live cell imaging, leads to diminished leptin pathway
activation, as indicated by reduced STAT3 phosphorylation (95). Mice exhibiting disrupted ciliary
trafficking of LepR develop leptin resistance, characterized by
overeating, decreased energy expenditure and exacerbated
dietary-induced obesity. However, restoring the proper ciliary
localization of LepR in MBH neurons can reestablish leptin
sensitivity and ameliorate obesity-related phenotypes, highlighting
the significance of LepR signaling integrity in hypothalamic
neuronal cilia for maintaining normal appetite regulation (58).
Adipocyte cilia also mitigate inflammation by
modulating responses to cytokines such as TNF (78). In obese mice on a high-fat diet,
adipocyte cilia become shortened, losing SSTR3 localization.
Compared to their lean counterparts, obese visceral fat exhibits
reduced cilia prevalence and length, with cilia loss intensifying
TNFα-induced inflammation (75).
This is characterized by increased NF-κB pathway activity, elevated
production of pro-inflammatory cytokines and higher levels of
macrophage markers like F4/80, establishing the role of adipocyte
cilia in curbing inflammation during obesity (96).
Beyond these tissue-specific mechanisms, the chronic
low-grade inflammation that drives increased fat storage and fatty
liver disease involves critical inter-organ communication. The
progression of metabolic dysfunction-associated steatotic liver
disease (MASLD) is supported by a spleen-liver axis that modulates
systemic inflammation. This process is characterized by an
enrichment of splenic myeloid-derived suppressor cells (MDSCs) and
natural killer T (NKT) cells, along with a loss of hepatic T and B
cells. Correlation analysis confirms a selective, strong positive
correlation between the distribution of spleen and liver MDSC and
NKT cells, indicating that the spleen-liver axis modulates
obesity-induced immune dysregulation in a cell-specific manner, as
demonstrated by recent research (97). Furthermore, spleen-derived
signals can directly amplify hepatic inflammation. Leukotriene B4
(LTB4) released by the spleen enhances liver TNF production both
in vivo and in vitro, identifying LTB4 as a
spleen-derived endocrine signal that promotes hepatic TNF
production during systemic inflammation (98). All these findings focusing on
pro-inflammatory processes lend credence to the significance of the
liver-spleen axis in non-alcoholic fatty liver disease/MASLD
pathogenesis (99). This axis
represents a critical systemic circuit through which inflammatory
signals are amplified and disseminated, contributing to the
metabolic dysfunction driven by ciliary defects in metabolic
tissues.
Overall, by regulating key inflammatory, metabolic
and endocrine pathways in the hypothalamus and adipose tissue,
primary cilia suppress local and systemic abnormalities that can
disturb energy homeostasis and promote obesity. Further studies are
needed to elucidate the complex molecular intersections between
ciliary dysfunction and inflammation in metabolic disease.
Nutrient digestion/absorption
When cells face nutrient deprivation, they encounter
an energetic crisis that is managed through metabolic
reorganization and adjustments in organelle functions. Primary
cilia, which are microtubule-based organelles on the cell surface,
have the capability to integrate various metabolic and signaling
cues. Primary cilia have been shown to sense glutamine availability
and respond by elongating via an asparagine synthetase (ASNS),
dependent mechanism that promotes ciliary tubulin acetylation and
stabilization, thereby enhancing ciliary length under nutrient
replete conditions (100).
During nutrient deprivation, cilia elongate, which occurs
independently of mTOR complex 1 (mTORC1), even when mitochondrial
function, ATP supply and AMPK activation are compromised. Of note,
the removal or supplementation of glutamine, under both in
vivo and in vitro nutritional stress, is necessary and
sufficient to regulate mitochondrial mitophagy. This process
depends on the production of glutamate from ASNS, leading to either
the elongation or retraction of cilia. Cells with IFT88 gene
mutations that lack cilia show glutamate-dependent mitochondrial
anaphase during metabolic stress, due to reduced ASNS expression
and activity at the ciliary base (100). These findings suggest that
primary cilia are responsive to cellular glutamine levels through
ASNS during periods of metabolic stress.
Given that certain commensal microbes metabolize
nutrients in manners protective against obesity, the regulation of
their populations through cilia-mediated antimicrobial secretion
directly impacts nutritional absorption. Additionally, cilia within
the biliary tract facilitate the circulation of bile necessary for
the digestion and absorption of lipids and fat-soluble vitamins.
Disruptions to motile cilia in the biliary system can alter bile
flow and contribute to gallstone disease (101).
In summary, through the coordinated regulation of
epithelial barrier function, antimicrobial release, microbiome
composition, bile flow and nutrient absorption, intestinal cilia
exert a profound influence on metabolism related to obesity.
Elucidating these intricate roles underscores the importance of
intestinal cilia in maintaining metabolic health and offers
potential avenues for targeting obesity and its associated
metabolic disorders.
Type 2 diabetes (T2D)
Insufficient adaptive β-cell compensation is a
hallmark of T2D (102). Primary
cilia, which are versatile sensory appendages, play a crucial role
in regulating diverse cellular functions. Studies have identified a
notably higher number of downregulated genes associated with cilia
in the pancreatic islets of diabetes-susceptible New Zealand Obese
mice compared to diabetes-resistant B6-ob/ob mice (103). Among the 327 mouse cilia genes
with altered expression, 81 had human homologs that were also
impacted in islets from diabetic individuals. In non-diabetic mouse
and human islets, there was a significant overlap in the
upregulation of ciliary genes linked to cell-cycle progression.
Suppression of KIF3A, a critical gene for ciliogenesis, through
short hairpin RNA, disrupted cell division in MIN6 β-cells and
primary pancreatic islet cells in both mice and humans, as shown by
decreased bromodeoxyuridine incorporation (103). Collectively, these results
highlight the vital role of ciliary gene regulation in islet
functionality and the risk of developing T2D.
Kidney epithelial cell metabolic
adaptation
Organs and cells need to adapt to the shear stress
induced by biofluids. Studies have demonstrated that shear stress
promotes mitochondrial biogenesis and metabolic reprogramming in
renal epithelial cells to ensure energy production and cellular
adaptation (104). This stress
also stimulates lipophagy, which facilitates the production of
fatty acids that mitochondria use to produce ATP through
β-oxidation (105). This
flow-induced process relies on primary cilia located at the tips of
epithelial cells (104).
Evidence from a unilateral ureteral obstruction mouse model
revealed that the absence of urinary flow disrupts cilia-dependent
lipophagy and mitochondrial biogenesis, culminating in lipid
droplet accumulation, thereby demonstrating that fluid flow is a
critical driver of lipid metabolism and metabolic adaptation in
renal epithelial cells in vivo (104). Shear stress activates two
signaling pathways originating from primary cilia. The first
pathway increases mitochondrial mass by upregulating two key
regulators of mitochondrial protein expression: PPARγ coactivator
1α, a master transcriptional coactivator of mitochondrial
biogenesis, and mitochondrial transcription factor A, a
nuclear-encoded protein essential for mitochondrial DNA
transcription and replication. The second pathway involves the
activation of lipophagy, a form of selective autophagy, to degrade
lipid droplets (104). Fatty
acids generated from lipophagy and cytoplasmic lipolysis are then
used to enhance ATP production in mitochondria (106). Primary cilia-dependent
lipophagy and mitochondrial biogenesis are essential for supporting
energy-demanding cellular processes such as glucose reabsorption,
gluconeogenesis and cytoskeletal remodeling (104). These findings underscore the
crucial role of primary cilia and autophagy in converting
mechanical forces into biological and physiological responses.
Impact of lipid metabolism abnormalities on
cilia
Metabolic abnormalities set in motion a
self-perpetuating cycle wherein ciliary defects, induced by a
high-fat diet and subsequent metabolic disturbances, further
aggravate weight gain and associated health issues (107). The disruption of anorexigenic
signaling from hypothalamic neuronal cilia leads to unchecked
hyperphagia. Dysfunctional pancreatic islet cilia hinder insulin
secretion, aggravating glucose intolerance. Furthermore, shortened
adipocyte cilia amplify inflammatory responses, driving excessive
adipose tissue growth and ectopic fat storage. Impaired biliary
cilia affect bile circulation and cholesterol metabolism,
highlighting the tissue-specific consequences of obesity on cilia
and their contribution to systemic metabolic imbalances.
Neuronal cilia
Emerging research indicates that obesity itself
compromises ciliary structure and function through various
mechanisms, setting off a deleterious cycle that exacerbates
metabolic dysfunction. In neurons critical for appetite and glucose
regulation, consumption of a high-fat diet has been shown to
adversely affect cilia prevalence and morphology in the
hypothalamus, cortex and hippocampus (96,108). Specifically, palmitic acid
exposure led to a reduction in axoneme length and a decrease in the
incidence of cilia within hypothalamic POMC and AgRP neurons
(109). A similar deterioration
was observed in hippocampal neurons subjected to saturated fatty
acids in vitro. The hyperactivation of inflammatory pathways
is a likely culprit behind neuronal cilia disruption. In POMC
neurons, saturated fatty acids were found to prompt ceramide
production and activate NF-κB, processes mediated by the ciliary
somatostatin receptor SSTR3 (110). Counteracting this inflammation
was shown to mitigate the loss of POMC cilia. Furthermore, a
prolonged high-fat diet contributed to the mislocalization of the
leptin receptor and disrupted leptin signaling specifically within
MBH neuronal cilia that are instrumental in appetite regulation
(91,111).
Pancreatic islet cilia
Pancreatic islet cilia are also vulnerable to the
deleterious effects of chronic hyperglycemia and elevated free
fatty acids characteristic of obesity (10). Hyperglycemia induced the
accumulation of oxidative stress markers like 4-hydroxynonenal, a
major bioactive aldehyde end-product of lipid peroxidation in
β-cell cilia. Elevated glucose levels impaired ciliary Hh
signaling, while palmitate hindered intraflagellar transport,
compounding disruptions from glucotoxicity and lipotoxicity that
impair insulin secretion by affecting ciliary signaling involved in
Ca2+ dynamics and exocytosis.
Adipocyte cilia
Adipocyte cilia exhibited notable structural and
functional impairments in the context of obesity. Cilia in obese
visceral fat were shortened, with compromised motility and
mechanosensation (108).
Proteomic analyses identified a reduction in structural and
signaling proteins, such as IFT88, smoothened and riboflavin
kinase, in cilia from obese adipocytes, suggesting that alterations
in ciliary signaling contribute to pathological adipose tissue
expansion and inflammation. Cilia in cholangiocytes also displayed
shortened, sparse axonemes in models of biliary cirrhosis (112). The collective disruption of
cilia functions across diverse tissues due to obesity-induced
damage plays a critical role in amplifying metabolic
abnormalities.
Osteoblast cilia
Fracture risk and impaired healing in diabetes pose
significant health concerns. Type 1 diabetes mellitus (T1DM) is
known to affect osteoblasts and their progenitor cells, making it a
critical risk factor for compromised fracture repair (14). Primary cilia, essential
organelles, play a key role in cell differentiation, proliferation
and function during tissue development and homeostasis (113). Proper cilia formation and
function depend on an efficient IFT system, with IFT80 being a core
transport protein vital for cilia assembly and bone development
(27,114,115). Studies have demonstrated that
diabetes lowers IFT80 expression and disrupts cilia formation in
osteoblasts, leading to defective fracture healing (14). Researchers examined the impact of
T1DM on primary cilia in a streptozotocin (STZ)-induced diabetes
mouse model and assessed the role of cilia in osteoblastic fracture
healing by deleting IFT80 in an osteoblastic cell line using
osterix (OSX)-cre (OSXcre/tTAIFT80f/f). Findings indicated that
diabetes inhibited cilia gene expression and primary cilia
formation to a similar extent as IFT80 deletion in normoglycemic
mice. Both diabetic mice and normoglycemic mice with
cilia-deficient osteoblasts (OSXcre/tTAIFT80f/f) exhibited delayed
fracture healing, significantly reduced bone density and mechanical
strength, lower expression of osteoblast markers, diminished
angiogenesis and decreased proliferation of bone-lining cells at
the fracture site. In vitro studies revealed that advanced
glycation end products (AGEs) decrease IFT80 expression in
osteoblast progenitor cells. The absence of AGEs and IFT80
significantly reduced the number and length of cilia, thus
inhibiting the differentiation of primary osteoblast precursors
(14). Increased Foxo1
expression was linked to defective fracture healing under diabetic
conditions. Researchers discovered that STZ-induced T1D markedly
increased Foxo1 expression in the femoral fracture callus,
resulting in a significant decrease in IFT80 expression and primary
cilia number (14,116). Ablation of Foxo1 in osteoblasts
of OSXcre/tTAFoxo1f/f mice restored IFT80 expression and
ciliogenesis, improving bone formation and mechanical strength in
diabetic fracture callus. In vitro, AGEs impaired cilia
formation in osteoblasts and reduced mineralized matrix production,
which was rescued by Foxo1 gene deletion. Mechanistically, AGEs
increased Foxo1 expression and transcriptional activity, inhibiting
IFT80 expression and cilia formation (102). Thus, diabetes impairs fracture
healing through Foxo1-mediated inhibition of ciliary IFT80
expression and primary cilia formation, resulting in compromised
osteogenesis. Inhibition of Foxo1 and/or restoration of cilia
formation could enhance fracture healing in diabetes.
Chronic hyperglycemia and metabolic intermediates,
including inflammatory factors, reactive oxygen species (ROS) and
AGEs, are recognized as major contributors to bone complications
(117). Through the use of a
diabetic Sprague Dawley rat model and an in vitro cellular
model of diabetic bone loss, researchers have shown that diabetes
impairs primary cilia, leading to bone deterioration. However, the
addition of Icariin (ICA) has been found to mitigate this negative
impact. ICA acts by scavenging ROS and preserving mitochondrial and
primary cilia homeostasis in osteoblasts. Functional primary cilia
provide anchoring and modification sites for Gli2, which in turn
activates the primary cilia/Gli2/osteocalcin signaling pathway,
enhancing osteoblast differentiation (118). Thus, ICA shows promise as a
therapeutic agent for preventing diabetes-induced bone loss.
Metabolic abnormalities lead to ciliary dysfunction,
which in turn exacerbates metabolic abnormalities, and an
understanding of this interrelationship is critical to the
development of therapeutic interventions. This creates a vicious
cycle: Ciliary dysfunction in osteoblasts impairs bone formation
and reduces the synthesis and bioactivity of bone-derived hormones
like osteocalcin. Given that osteocalcin is known to enhance
pancreatic β-cell proliferation and insulin secretion, its
deficiency contributes directly to worsening systemic glucose
homeostasis, thereby exacerbating the underlying metabolic
dysfunction (119). Exploring
strategies to restore ciliary function and break this cycle opens
promising avenues for novel antimetabolic disease therapies. In the
future, investigating the subtle effects of metabolic abnormalities
on cilia in a variety of metabolic diseases will be critical to
identify timely opportunities for tissue-specific
interventions.
Cilia as therapeutic targets for treating
obesity
The development of small molecules that stabilize
the cytoskeleton presents a promising strategy to combat
obesity-induced ciliary loss in neurons crucial for appetite
regulation. Despite the clinical limitations of epothilones due to
their side effects, the search for microtubule-stabilizing agents
with favorable safety profiles remains a high priority.
Ciliobrevins, known for their specific inhibition of cytoplasmic
dynein motors, offer another avenue for modulating cilia length and
Hh signaling (120).
Ciliobrevin D, at low doses, has been shown to prevent the
excessive elongation of adipocyte cilia triggered by mTORC1
activation, thereby normalizing lipid accumulation (120,121). The exploration of this
compound, along with related dynein inhibitors, could provide
insights into normalizing cilia length and adipose tissue
homeostasis in the context of obesity.
Gene therapy strategies aiming to overexpress
depleted ciliary proteins offer a method to restore structural
integrity. The disruption of ciliogenesis through knockdown of the
IFT protein IFT88 can be counteracted by adeno-associated virus
(AAV)-mediated overexpression of IFT88, which rescues cilia
formation and corrects aberrant Hh signaling in deficient cells
(122). Employing similar
approaches for AAV delivery of axonemal proteins, which are
downregulated in obesity, such as SSTR3, might reinstate normal
ciliary assembly and sensory function in metabolic tissues
(123). Furthermore,
therapeutic modulation of autophagic pathways appears promising for
correcting the truncation of neuronal cilia associated with
obesity. The mTOR inhibitor rapamycin has been shown to normalize
shortened neuronal cilia, likely by promoting autophagic flux via
transcription factor EB (TFEB) activation (124). Small molecule agonists of TFEB
could potentially replicate this effect, offering a novel
therapeutic strategy (125).
Lastly, targeting receptors concentrated in ciliary
membranes offers another therapeutic avenue. The serotonin 2c
receptor (HT2cR), localized in neuronal cilia, plays a vital role
in regulating appetite circuits (126). Agonists that enhance HT2cR
activity specifically within cilia may potentiate satiety
signaling. Furthermore, biased agonists that directly activate MC4R
in POMC cilia (127),
irrespective of α-MSH levels, could address defective satiety
signaling resulting from ciliary dysfunction (55,56). Pharmacologically amplifying the
activity of appetite-suppressing receptors in the remaining
neuronal cilia may provide a means to mitigate hyperphagia and
combat obesity despite structural ciliary deficits.
Significant work remains to translate basic findings
on ciliary dysfunction in obesity into clinical therapies. Cilia
are widespread, so cell type-specific manipulation will be
imperative. For example, selectively stabilizing POMC neuronal
cilia vs. pancreatic islet cilia may produce different therapeutic
benefits. Additionally, distinct approaches may be needed to
correct defects in motile cilia vs. non-motile primary cilia.
Improved imaging methods and biomarkers would help stratify obese
patients by the contribution of ciliary defects vs. other
mechanisms, to select appropriate subgroups for cilia-targeted
therapies.
Studies should optimize treatment regimens based on
disease stage, since cilia dysfunction may exert more pronounced
effects in initiating vs. perpetuating obesity. For example,
enhancing ciliogenesis may have a greater impact early on, while
stabilizing remaining cilia or augmenting ciliary signaling could
give greater benefit in established obesity. Despite current
challenges, rapid progress elucidating cilia biology in metabolic
control continues, offering hope for novel therapeutic inroads
against obesity by targeting this intricate signaling nexus.
Conclusions and outlook
This review synthesizes emerging evidence supporting
a bidirectional relationship between primary cilia defects and
obesity pathogenesis. Cilia are microscopic sensory organelles
projecting from nearly all mammalian cell types. They serve vital
signaling roles during development and homeostasis by detecting
mechanical, chemical and biological cues and transducing these into
intracellular signaling cascades (22). Cilia accomplish this through
compartmentalization of receptors, ion channels and components of
pathways like Hh, Wnt, receptor tyrosine kinase, mTOR and GPCR
(16,19). Accordingly, specialized cilia in
neuronal, endocrine and metabolic tissues help coordinate systemic
energy balance. For example, hypothalamic neuronal cilia regulate
appetite circuits through satiety hormone receptors like MC4R
(58). Pancreatic islet cilia
control insulin secretion dynamics (10). Gastrointestinal cilia modulate
nutrient digestion and absorption. Adipocyte cilia restrain lipid
accumulation and inflammation (128). Dysfunction of these
metabolically relevant cilia promotes weight gain and
complications. Conversely, obesity damages cilia integrity through
inflammation, lipotoxicity, glucotoxicity and other stresses.
High-fat diet and obesity disrupt cilia in appetite-regulating
neurons, reducing satiety signaling (108). Islet cilia are impaired by
hyperglycemia and lipotoxicity, compromising insulin secretion
(83). Shortened adipocyte cilia
exacerbate inflammation and ectopic fat deposition. These
compounding hits on diverse tissue cilia precipitate a vicious
cycle where cilia defects beget worsened metabolic dysfunction.
Clarifying this bidirectional relationship reveals exciting new
perspectives on obesity pathogenesis and highlights novel targets
for intervention. Approaches targeting neuronal cilia may boost
satiety signaling to reduce hyperphagia. Stabilizing islet cilia
could enhance glucose-responsive insulin secretion (84). Normalizing adipocyte cilia length
and signaling may ameliorate inflammation from obesity (129). Elucidating such intersections
between ciliopathies and metabolic disease creates new frontiers
for obesity research and therapy development.
Progressing from these foundational insights will
require developing new tools to assess cilia dysfunction in human
obesity. The current understanding is derived largely from animal
models, so improved imaging modalities and biomarkers to analyze
cilia non-invasively in obese patients could enable translation to
the clinic. Defining the specific contributions of distinct tissue
cilia populations is crucial, as neuronal, endocrine and metabolic
cilia likely fulfill unique roles (90), and differences between motile and
primary cilia must be delineated. With obesity's heterogeneous
nature, identifying patient subgroups with predominant cilia
defects is imperative to select candidates for targeted treatment.
Considerable heterogeneity likely exists in obese individuals
regarding causal mechanisms. Biomarkers stratifying those with
major cilia deficits could enable personalized therapy (1,2).
Exploring pharmacological, gene or cell therapy approaches to
prevent obesity-induced cilia deterioration or rebuild ciliary
integrity represents an important direction (130). Small molecules stabilizing
microtubules may protect neuronal cilia, while IFT protein
overexpression could enable ciliogenesis (114,115). However, considering cilia's
ubiquitous roles, potential unintended consequences of globally
manipulating cilia require investigation. Understanding cell
type-specific differences will be critical to avoid developmental
or neurological impacts. Furthermore, defining optimal timing and
duration of cilia-focused therapies based on disease stage requires
further elucidation, as early intervention for prevention may
differ from the treatment of established obesity. In summary, rapid
discovery continues in this domain, bringing hope that leveraging
intricate cilia biology could yield innovative solutions against
the intractable obesity epidemic.
Availability of data and materials
Not applicable.
Authors' contributions
XW and YT conceived and designed the study. YJ, LS
and PY collected and organized references. XW wrote the original
draft. KX and LH processed the images. YT and YJ critically
reviewed the manuscript. All authors read and approved the final
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
This work was supported by the Shandong Provincial Natural
Science Foundation (grant no. ZR2023QC058).
References
|
1
|
Wu X, Xu M, Geng M, Chen S, Little PJ, Xu
S and Weng J: Targeting protein modifications in metabolic
diseases: molecular mechanisms and targeted therapies. Signal
Transduct Target Ther. 8:2202023. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fan Y and Pedersen O: Gut microbiota in
human metabolic health and disease. Nat Rev Microbiol. 19:55–71.
2021. View Article : Google Scholar
|
|
3
|
Kasahara K and Nagaki M: Primary ciliary
signaling: links with the cell cycle. Trends Cell Biol. 31:954–964.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang DJ, Hong J and Kim KW: Hypothalamic
primary cilium: A hub for metabolic homeostasis. Exp Mol Med.
53:1109–1115. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee CH, Song DK, Park CB, Choi J, Kang GM,
Shin SH, Kwon I, Park S, Kim S, Kim JY, et al: Primary cilia
mediate early life programming of adiposity through lysosomal
regulation in the developing mouse hypothalamus. Nat Commun.
11:57722020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu CT, Hilgendorf KI, Bevacqua RJ, Hang Y,
Demeter J, Kim SK and Jackson PK: Discovery of ciliary G
protein-coupled receptors regulating pancreatic islet insulin and
glucagon secretion. Genes Dev. 35:1243–1255. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hong R, Tan Y, Tian X, Huang Z, Wang J, Ni
H, Yang J, Bu W, Yang S, Li T, et al: XIAP-mediated degradation of
IFT88 disrupts HSC cilia to stimulate HSC activation and liver
fibrosis. EMBO Rep. 25:1055–1074. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hilgendorf KI, Johnson CT, Mezger A, Rice
SL, Norris AM, Demeter J, Greenleaf WJ, Reiter JF, Kopinke D and
Jackson PK: Omega-3 Fatty Acids Activate Ciliary FFAR4 to control
adipogenesis. Cell. 179:1289–1305 e21. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Bernard A, Comblain F, Yue X,
Paillart C, Zhang S, Reiter JF and Vaisse C: Melanocortin 4
receptor signals at the neuronal primary cilium to control food
intake and body weight. J Clin Invest. 131:e1420642021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee EY and Hughes JW: Rediscovering
primary cilia in pancreatic islets. Diabetes Metab J. 47:454–469.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gohlke S, Mancini C, Garcia-Carrizo F and
Schulz TJ: Loss of the ciliary gene Bbs4 results in defective
thermogenesis due to metabolic inefficiency and impaired lipid
metabolism. FASEB J. 35:e219662021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vaisse C, Reiter JF and Berbari NF: Cilia
and obesity. Cold Spring Harb Perspect Biol. 9:a0282172017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim YZ, Zhu M, Wang Y, Sharma T, Kelley S,
Oertling E, Zhu H and Corbitt N: Pkd1l1-deficiency drives biliary
atresia through ciliary dysfunction in biliary epithelial cells. J
Hepatol. 81:62–75. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chinipardaz Z, Yuan G, Liu M, Graves DT
and Yang S: Diabetes impairs fracture healing through Foxo1
mediated disruption of ciliogenesis. Cell Death Discov. 9:2992023.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chinipardaz Z, Liu M, Graves D and Yang S:
Diabetes impairs fracture healing through disruption of cilia
formation in osteoblasts. Bone. 153:1161762021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tu HQ, Li S, Xu YL, Zhang YC, Li PY, Liang
LY, Song GP, Jian XX, Wu M, Song ZQ, et al: Rhythmic cilia changes
support SCN neuron coherence in circadian clock. Science.
380:972–979. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu CT, Lidsky PV, Xiao Y, Cheng R, Lee IT,
Nakayama T, Jiang S, He W, Demeter J, Knight MG, et al: SARS-CoV-2
replication in airway epithelia requires motile cilia and
microvillar reprogramming. Cell. 186:112–130 e20. 2023. View Article : Google Scholar
|
|
18
|
Price ME and Sisson JH: Redox regulation
of motile cilia in airway disease. Redox Biol. 27:1011462019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Silva DF and Cavadas C: Primary cilia
shape hallmarks of health and aging. Trends Mol Med. 29:567–579.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Beeby M, Ferreira JL, Tripp P, Albers SV
and Mitchell DR: Propulsive nanomachines: The convergent evolution
of archaella, flagella and cilia. FEMS Microbiol Rev. 44:253–304.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nachury MV and Mick DU: Establishing and
regulating the composition of cilia for signal transduction. Nat
Rev Mol Cell Biol. 20:389–405. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mill P, Christensen ST and Pedersen LB:
Primary cilia as dynamic and diverse signalling hubs in development
and disease. Nat Rev Genet. 24:421–441. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hesketh SJ, Mukhopadhyay AG, Nakamura D,
Toropova K and Roberts AJ: IFT-A structure reveals carriages for
membrane protein transport into cilia. Cell. 185:4971–4985.e16.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
van den Hoek H, Klena N, Jordan MA,
Alvarez Viar G, Righetto RD, Schaffer M, Erdmann PS, Wan W, Geimer
S, Plitzko JM, et al: In situ architecture of the ciliary base
reveals the stepwise assembly of intraflagellar transport trains.
Science. 377:543–548. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang M, Palicharla VR, Miller D, Hwang
SH, Zhu H, Hixson P, Mukhopadhyay S and Sun J: Human IFT-A complex
structures provide molecular insights into ciliary transport. Cell
Res. 33:288–298. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Meleppattu S, Zhou H, Dai J, Gui M and
Brown A: Mechanism of IFT-A polymerization into trains for ciliary
transport. Cell. 185:4986–4998.e12. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Deepak V, Yang ST, Li Z, Li X, Ng A, Xu D,
Li YP, Oursler MJ and Yang S: IFT80 negatively regulates osteoclast
differentiation via association with Cbl-b to disrupt TRAF6
stabilization and activation. Proc Natl Acad Sci USA.
119:e22014901192022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Katoh TA, Omori T, Mizuno K, Sai X,
Minegishi K, Ikawa Y, Nishimura H, Itabashi T, Kajikawa E, Hiver S,
et al: Immotile cilia mechanically sense the direction of fluid
flow for left-right determination. Science. 379:66–71. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Magistrati E, Maestrini G, Nino CA,
Lince-Faria M, Beznoussenko G, Mironov A, Maspero E,
Bettencourt-Dias M and Polo S: Myosin VI regulates ciliogenesis by
promoting the turnover of the centrosomal/satellite protein OFD1.
EMBO Rep. 23:e541602022. View Article : Google Scholar :
|
|
30
|
Lo CH, Lin IH, Yang TT, Huang YC, Tanos
BE, Chou PC, Chang CW, Tsay YG, Liao JC and Wang WJ:
Phosphorylation of CEP83 by TTBK2 is necessary for cilia
initiation. J Cell Biol. 218:3489–3505. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee H, Moon KH, Song J, Je S, Bok J and Ko
HW: Tissue-specific requirement of sodium channel and clathrin
linker 1 (Sclt1) for ciliogenesis during limb development. Front
Cell Dev Biol. 10:10588952022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gopalakrishnan J, Feistel K, Friedrich BM,
Grapin-Botton A, Jurisch-Yaksi N, Mass E, Mick DU, Muller RU,
May-Simera H, Schermer B, et al: Emerging principles of primary
cilia dynamics in controlling tissue organization and function.
EMBO J. 42:e1138912023. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Yang Y, Li X, Li G, Mizukami T,
Liu Y, Wang Y, Xu G, Roder H, Zhang L and Yang ZJ: PDLIM3 supports
hedgehog signaling in medulloblastoma by facilitating cilia
formation. Cell Death Differ. 30:1198–1210. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Somatilaka BN, Hwang SH, Palicharla VR,
White KA, Badgandi H, Shelton JM and Mukhopadhyay S: Ankmy2
prevents smoothened-independent hyperactivation of the hedgehog
pathway via cilia-regulated adenylyl cyclase signaling. Dev Cell.
54:710–726.e8. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang ZW, Teng X, Zhao F, Ma C, Zhang J,
Xiao LF, Wang Y, Chang M, Tian Y, Li C, et al: METTL3 regulates
m(6)A methylation of PTCH1 and GLI2 in Sonic hedgehog signaling to
promote tumor progression in SHH-medulloblastoma. Cell Rep.
41:1115302022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang K, Da Silva F, Seidl C,
Wilsch-Brauninger M, Herbst J, Huttner WB and Niehrs C: Primary
cilia are WNT-transducing organelles whose biogenesis is controlled
by a WNT-PP1 axis. Dev Cell. 58:139–154.e8. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu S, Jin R, Zheng G, Wang Y, Li Q, Jin
F, Li Y, Li T, Mao N, Wei Z, et al: Ac-SDKP promotes KIF3A-mediated
beta-catenin suppression through a ciliary mechanism to constrain
silica-induced epithelial-myofibroblast transition. Biomed
Pharmacother. 166:1154112023. View Article : Google Scholar
|
|
38
|
Zhou H, Wu S, Ling H, Zhang C and Kong Y:
Primary cilia: A cellular regulator of articular cartilage
degeneration. Stem Cells Int. 2022:25604412022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gerhards J, Maerz LD, Matthees ESF, Donow
C, Moepps B, Premont RT, Burkhalter MD, Hoffmann C and Philipp M:
Kinase activity is not required for G protein-coupled receptor
kinase 4 restraining mTOR signaling during cilia and kidney
development. J Am Soc Nephrol. 34:590–606. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rausch V and Hansen CG: The hippo pathway,
YAP/TAZ, and the plasma membrane. Trends Cell Biol. 30:32–48. 2020.
View Article : Google Scholar
|
|
41
|
Wang L, Paudyal SC, Kang Y, Owa M, Liang
FX, Spektor A, Knaut H, Sanchez I and Dynlacht BD: Regulators of
tubulin polyglutamylation control nuclear shape and cilium
disassembly by balancing microtubule and actin assembly. Cell Res.
32:190–209. 2022. View Article : Google Scholar
|
|
42
|
Tsai YC, Kuo TN, Chao YY, Lee PR, Lin RC,
Xiao XY, Huang BM and Wang CY: PDGF-AA activates AKT and ERK
signaling for testicular interstitial Leydig cell growth via
primary cilia. J Cell Biochem. 124:89–102. 2023. View Article : Google Scholar
|
|
43
|
Schwark K, Messinger D, Cummings JR,
Bradin J, Kawakibi A, Babila CM, Lyons S, Ji S, Cartaxo RT, Kong S,
et al: Receptor tyrosine kinase (RTK) targeting in pediatric
high-grade glioma and diffuse midline glioma: Pre-clinical models
and precision medicine. Front Oncol. 12:9229282022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wachten D and Mick DU: Signal transduction
in primary cilia-analyzing and manipulating GPCR and second
messenger signaling. Pharmacol Ther. 224:1078362021. View Article : Google Scholar
|
|
45
|
Shinde SR, Nager AR and Nachury MV:
Ubiquitin chains earmark GPCRs for BBSome-mediated removal from
cilia. J Cell Biol. 219:e2020030202020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Carey RM, Hariri BM, Adappa ND, Palmer JN
and Lee RJ: HSP90 modulates T2R bitter taste receptor nitric oxide
production and innate immune responses in human airway epithelial
cells and macrophages. Cells. 11:14782022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hansen CG, Moroishi T and Guan KL: YAP and
TAZ: A nexus for Hippo signaling and beyond. Trends Cell Biol.
25:499–513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chinipardaz Z, Liu M, Graves DT and Yang
S: Role of primary cilia in bone and cartilage. J Dent Res.
101:253–260. 2022. View Article : Google Scholar
|
|
49
|
Truong HM, Cruz-Colón KO, Martínez-Márquez
JY, Willer JR, Travis AM, Biswas SK, Lo WK, Bolz HJ and Pearring
JN: The tectonic complex regulates membrane protein composition in
the photoreceptor cilium. Nat Commun. 14:56712023. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li S, Sampson C, Liu C, Piao HL and Liu
HX: Integrin signaling in cancer: Bidirectional mechanisms and
therapeutic opportunities. Cell Commun Signal. 21:2662023.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li X, Lu Q, Peng Y, Geng F, Shao X, Zhou
H, Cao Y and Zhang R: Primary cilia mediate Klf2-dependant Notch
activation in regenerating heart. Protein Cell. 11:433–445. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu Z, Tu H, Kang Y, Xue Y, Ma D, Zhao C,
Li H, Wang L and Liu F: Primary cilia regulate hematopoietic stem
and progenitor cell specification through Notch signaling in
zebrafish. Nat Commun. 10:18392019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li X and Yang S, Deepak V, Chinipardaz Z
and Yang S: Identification of cilia in different mouse tissues.
Cells. 10:16232021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Finetti F, Paccani SR, Riparbelli MG,
Giacomello E, Perinetti G, Pazour GJ, Rosenbaum JL and Baldari CT:
Intraflagellar transport is required for polarized recycling of the
TCR/CD3 complex to the immune synapse. Nat Cell Biol. 11:1332–1339.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Varela L and Horvath TL: Neuronal cilia:
Another player in the melanocortin system. Trends Mol Med.
24:333–334. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Quarta C, Claret M, Zeltser LM, Williams
KW, Yeo GSH, Tschop MH, Diano S, Bruning JC and Cota D: POMC
neuronal heterogeneity in energy balance and beyond: an integrated
view. Nat Metab. 3:299–308. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lotta LA, Mokrosinski J, Mendes de
Oliveira E, Li C, Sharp SJ, Luan J, Brouwers B, Ayinampudi V,
Bowker N, Kerrison N, et al: Human gain-of-function MC4R variants
show signaling bias and protect against obesity. Cell. 177:597–607
e9. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Oya M, Miyasaka Y, Nakamura Y, Tanaka M,
Suganami T, Mashimo T and Nakamura K: Age-related ciliopathy:
Obesogenic shortening of melanocortin-4 receptor-bearing neuronal
primary cilia. Cell Metab. 36:1044–1058.e10. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Radecki KC, Ford MJ, Phillipps HR,
Lorenson MY, Grattan DR, Yamanaka Y and Walker AM: Multiple cell
types in the oviduct express the prolactin receptor. FASEB Bioadv.
4:485–504. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sanchez GM, Incedal TC, Prada J,
O'Callaghan P, Dyachok O, Echeverry S, Dumral O, Nguyen PM, Xie B,
Barg S, et al: The β-cell primary cilium is an autonomous Ca2+
compartment for paracrine GABA signaling. J Cell Biol.
222:e2021081012023. View Article : Google Scholar
|
|
61
|
Rouabhi M, Guo DF, Morgan DA, Zhu Z, Lopez
M, Zingman L, Grobe JL and Rahmouni K: BBSome ablation in SF1
neurons causes obesity without comorbidities. Mol Metab.
48:1012112021. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Quinones M, Al-Massadi O, Folgueira C,
Bremser S, Gallego R, Torres-Leal L, Haddad-Tovolli R,
Garcia-Caceres C, Hernandez-Bautista R, Lam BYH, et al: p53 in AgRP
neurons is required for protection against diet-induced obesity via
JNK1. Nat Commun. 9:34322018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Blaess S and Wachten D: The BBSome: A
nexus controlling energy metabolism in the brain. J Clin Invest.
131:e1489032021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Koike K, Yoo SJ, Bleymehl K, Omura M,
Zapiec B, Pyrski M, Blum T, Khan M, Bai Z, Leinders-Zufall T, et
al: Danger perception and stress response through an olfactory
sensor for the bacterial metabolite hydrogen sulfide. Neuron.
109:2469–2484.e7. 2021. View Article : Google Scholar
|
|
65
|
Engle SE, Bansal R, Antonellis PJ and
Berbari NF: Cilia signaling and obesity. Semin Cell Dev Biol.
110:43–50. 2021. View Article : Google Scholar
|
|
66
|
Allard BA, Wang W, Pottorf TS, Mumtaz H,
Jack BM, Wang HH, Silva LM, Jacobs DT, Wang J, Bumann EE and Tran
PV: Thm2 interacts with paralog, Thm1, and sensitizes to Hedgehog
signaling in postnatal skeletogenesis. Cell Mol Life Sci.
78:3743–3762. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Pottorf TS, Fagan MP, Burkey BF, Cho DJ,
Vath JE and Tran PV: MetAP2 inhibition reduces food intake and body
weight in a ciliopathy mouse model of obesity. JCI Insight.
5:e1342782020. View Article : Google Scholar :
|
|
68
|
Jacobs DT, Allard BA, Pottorf TS, Silva
LM, Wang W, Al-Naamani A, Agborbesong E, Wang T, Carr DA and Tran
PV: Intraflagellar-transport A dysfunction causes
hyperphagia-induced systemic insulin resistance in a pre-obese
state. FASEB J. 34:148–160. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Jacobs DT, Silva LM, Allard BA, Schonfeld
MP, Chatterjee A, Talbott GC, Beier DR and Tran PV: Dysfunction of
intraflagellar transport-A causes hyperphagia-induced obesity and
metabolic syndrome. Dis Model Mech. 9:789–798. 2016.PubMed/NCBI
|
|
70
|
Kitamura T, Feng Y, Kitamura YI, Chua SC
Jr, Xu AW, Barsh GS, Rossetti L and Accili D: Forkhead protein
FoxO1 mediates Agrp-dependent effects of leptin on food intake. Nat
Med. 12:534–540. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
71
|
Iskandar K, Cao Y, Hayashi Y, Nakata M,
Takano E, Yada T, Zhang C, Ogawa W, Oki M, Chua S Jr, et al:
PDK-1/FoxO1 pathway in POMC neurons regulates Pomc expression and
food intake. Am J Physiol Endocrinol Metab. 298:E787–E798. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Dagon Y, Hur E, Zheng B, Wellenstein K,
Cantley LC and Kahn B: p70S6 kinase phosphorylates AMPK on serine
491 to mediate leptin's effect on food intake. Cell Metab.
16:104–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lim GE, Albrecht T, Piske M, Sarai K, Lee
JTC, Ramshaw HS, Sinha S, Guthridge MA, Acker-Palmer A, Lopez AF,
et al: 14-3-3ζ coordinates adipogenesis of visceral fat. Nat
Commun. 6:76712015. View Article : Google Scholar
|
|
74
|
Chang CF, Chang YT, Millington G and
Brugmann SA: Craniofacial ciliopathies reveal specific requirements
for GLI proteins during development of the facial midline. PLoS
Genet. 12:e10063512016. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Oh S, Kim HM, Batsukh S, Sun HJ, Kim T,
Kang D, Son KH and Byun K: High-intensity focused ultrasound
induces adipogenesis via control of cilia in adipose-derived stem
cells in subcutaneous adipose tissue. Int J Mol Sci. 23:88662022.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tian JL and Gomeshtapeh FI: Potential
roles of O-GlcNAcylation in primary cilia-mediated energy
metabolism. Biomolecules. 10:15042020. View Article : Google Scholar
|
|
77
|
Ma X, Zhang Y, Zhang Y, Zhang X, Huang Y,
He K, Chen C, Hao J, Zhao D, LeBrasseur NK, et al: A stress-induced
cilium-to-PML-NB route drives senescence initiation. Nat Commun.
14:18402023. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang Y, Hao J, Tarrago MG, Warner GM,
Giorgadze N, Wei Q, Huang Y, He K, Chen C, Peclat TR, et al: FBF1
deficiency promotes beiging and healthy expansion of white adipose
tissue. Cell Rep. 36:1094812021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Giuliani G, Rosina M and Reggio A:
Signaling pathways regulating the fate of fibro/adipogenic
progenitors (FAPs) in skeletal muscle regeneration and disease.
FEBS J. 289:6484–6517. 2022. View Article : Google Scholar
|
|
80
|
Yamakawa D, Tsuboi J, Kasahara K, Matsuda
C, Nishimura Y, Kodama T, Katayama N, Watanabe M and Inagaki M:
Cilia-Mediated Insulin/Akt and ST2/JNK signaling pathways regulate
the recovery of muscle injury. Adv Sci (Weinh). 10:e22026322022.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Fu S, Zhang C, Yan X, Li D, Wang Y, Dong
C, Cao Z, Ning Y, Shao C and Yang T: Primary cilia as a biomarker
in mesenchymal stem cells senescence: Influencing osteoblastic
differentiation potency associated with hedgehog signaling
regulation. Stem Cells Int. 2021:88501142021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Yamakawa D, Katoh D, Kasahara K, Shiromizu
T, Matsuyama M, Matsuda C, Maeno Y, Watanabe M, Nishimura Y and
Inagaki M: Primary cilia-dependent lipid raft/caveolin dynamics
regulate adipogenesis. Cell Rep. 34:1088172021. View Article : Google Scholar
|
|
83
|
Cho JH, Li ZA, Zhu L, Muegge BD, Roseman
HF, Lee EY, Utterback T, Woodhams LG, Bayly PV and Hughes JW: Islet
primary cilia motility controls insulin secretion. Sci Adv.
8:eabq84862022. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hughes JW, Cho JH, Conway HE, DiGruccio
MR, Ng XW, Roseman HF, Abreu D, Urano F and Piston DW: Primary
cilia control glucose homeostasis via islet paracrine interactions.
Proc Natl Acad Sci USA. 117:8912–8923. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Pablos M, Casanueva-Alvarez E,
Gonzalez-Casimiro CM, Merino B, Perdomo G and Cozar-Castellano I:
Primary cilia in pancreatic β- and α-cells: Time to revisit the
role of insulin-degrading enzyme. Front Endocrinol (Lausanne).
13:9228252022. View Article : Google Scholar
|
|
86
|
Saternos H, Ley S and AbouAlaiwi W:
Primary cilia and calcium signaling interactions. Int J Mol Sci.
21:71092020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Volta F, Scerbo MJ, Seelig A, Wagner R,
O'Brien N, Gerst F, Fritsche A, Haring HU, Zeigerer A, Ullrich S
and Gerdes JM: Glucose homeostasis is regulated by pancreatic
beta-cell cilia via endosomal EphA-processing. Nat Commun.
10:56862019. View Article : Google Scholar
|
|
88
|
Polino AJ, Sviben S, Melena I, Piston DW
and Hughes JW: Scanning electron microscopy of human islet cilia.
Proc Natl Acad Sci USA. 120:e23026241202023. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Maurya DK, Berghard A and Bohm S: A
multivesicular body-like organelle mediates stimulus-regulated
trafficking of olfactory ciliary transduction proteins. Nat Commun.
13:68892022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Picon-Galindo E, Latz E and Wachten D:
Primary cilia and their effects on immune cell functions and
metabolism: A model. Trends Immunol. 43:366–378. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yang DL, Tran LT, Yoon SG, Seong JK, Shin
DM, Choi YH and Kim KW: Primary cilia regulate adaptive responses
to fasting. Metabolism. 135:1552732022. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Zhang B, Feng H, Lin H and Li R:
Somatostatin-SSTR3-GSK3 modulates human T-cell responses by
inhibiting OXPHOS. Front Immunol. 15:13226702024. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Green JA and Mykytyn K: Neuronal primary
cilia: An underappreciated signaling and sensory organelle in the
brain. Neuropsychopharmacology. 39:244–245. 2014. View Article : Google Scholar
|
|
94
|
Berbari NF, Lewis JS, Bishop GA, Askwith
CC and Mykytyn K: Bardet-Biedl syndrome proteins are required for
the localization of G protein-coupled receptors to primary cilia.
Proc Natl Acad Sci USA. 105:4242–4246. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wang L, Liu Y, Stratigopoulos G, Panigrahi
S, Sui L, Zhang Y, Leduc CA, Glover HJ, De Rosa MC, Burnett LC, et
al: Bardet-Biedl syndrome proteins regulate intracellular signaling
and neuronal function in patient-specific iPSC-derived neurons. J
Clin Invest. 131:e1462872021. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Baek H, Shin HJ, Kim JJ, Shin N, Kim S, Yi
MH, Zhang E, Hong J, Kang JW, Kim Y, et al: Primary cilia modulate
TLR4-mediated inflammatory responses in hippocampal neurons. J
Neuroinflammation. 14:1892017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Brummer C, Singer K, Renner K, Bruss C,
Hellerbrand C, Dorn C, Reichelt-Wurm S, Gronwald W, Pukrop T, Herr
W, et al: The spleen-liver axis supports obesity-induced systemic
and fatty liver inflammation via MDSC and NKT cell enrichment. Mol
Cell Endocrinol. 601:1125182025. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Fonseca MT, Moretti EH, Marques LMM,
Machado BF, Brito CF, Guedes JT, Komegae EN, Vieira TS, Festuccia
WT, Lopes NP and Steiner AA: A leukotriene-dependent spleen-liver
axis drives TNF production in systemic inflammation. Sci Signal.
14:eabb09692021. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Tarantino G, Citro V and Balsano C:
Liver-spleen axis in nonalcoholic fatty liver disease. Expert Rev
Gastroenterol Hepatol. 15:759–769. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Steidl ME, Nigro EA, Nielsen AK,
Pagliarini R, Cassina L, Lampis M, Podrini C, Chiaravalli M,
Mannella V, Distefano G, et al: Primary cilia sense glutamine
availability and respond via asparagine synthetase. Nat Metab.
5:385–397. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Ogawa M, Jiang JX, Xia S, Yang D, Ding A,
Laselva O, Hernandez M, Cui C, Higuchi Y, Suemizu H, et al:
Generation of functional ciliated cholangiocytes from human
pluripotent stem cells. Nat Commun. 12:65042021. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Xourafa G, Korbmacher M and Roden M:
Inter-organ crosstalk during development and progression of type 2
diabetes mellitus. Nat Rev Endocrinol. 20:27–49. 2024. View Article : Google Scholar
|
|
103
|
Kluth O, Stadion M, Gottmann P, Aga H,
Jahnert M, Scherneck S, Vogel S, Krus U, Seelig A, Ling C, et al:
Decreased expression of cilia genes in pancreatic islets as a risk
factor for type 2 diabetes in mice and humans. Cell Rep.
26:3027–3036 e3. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Miceli C, Roccio F, Penalva-Mousset L,
Burtin M, Leroy C, Nemazanyy I, Kuperwasser N, Pontoglio M,
Friedlander G, Morel E, et al: The primary cilium and lipophagy
translate mechanical forces to direct metabolic adaptation of
kidney epithelial cells. Nat Cell Biol. 22:1091–1102. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Claude-Taupin A, Isnard P, Bagattin A,
Kuperwasser N, Roccio F, Ruscica B, Goudin N, Garfa-Traore M,
Regnier A, Turinsky L, et al: The AMPK-Sirtuin 1-YAP axis is
regulated by fluid flow intensity and controls autophagy flux in
kidney epithelial cells. Nat Commun. 14:80562023. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Hoogstraten CA, Hoenderop JG and de Baaij
JHF: Mitochondrial dysfunction in kidney tubulopathies. Annu Rev
Physiol. 86:379–403. 2024. View Article : Google Scholar
|
|
107
|
Qiu N, Fang WJ, Li HS, He ZM, Xiao ZS and
Xiong Y: Impairment of primary cilia contributes to visceral
adiposity of high fat diet-fed mice. J Cell Biochem. 119:1313–1325.
2018. View Article : Google Scholar
|
|
108
|
Oh S, Son M, Jang JT, Park CH, Son KH and
Byun K: Pyrogallol-phloroglucinol-6, 6-bieckol restored primary
cilia length, which was decreased by high-fat diet in visceral
adipose tissue, and decreased adipogenesis. Int J Endocrinol.
2022:84869652022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Avalos Y, Hernandez-Caceres MP, Lagos P,
Pinto-Nunez D, Rivera P, Burgos P, Diaz-Castro F, Joy-Immediato M,
Venegas-Zamora L, Lopez-Gallardo E, et al: Palmitic acid control of
ciliogenesis modulates insulin signaling in hypothalamic neurons
through an autophagy-dependent mechanism. Cell Death Dis.
13:6592022. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Parker AK, Le MM, Smith TS, Hoang-Minh LB,
Atkinson EW, Ugartemendia G, Semple-Rowland S, Coleman JE and
Sarkisian MR: Neonatal seizures induced by pentylenetetrazol or
kainic acid disrupt primary cilia growth on developing mouse
cortical neurons. Exp Neurol. 282:119–127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Lee CH, Kang GM and Kim MS: Mechanisms of
weight control by primary cilia. Mol Cells. 45:169–176. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Chen J, Cheng NC, Boland JA, Liu K, Kench
JG, Watkins DN, Ferreira-Gonzalez S, Forbes SJ and McCaughan GW:
Deletion of kif3a in CK19 positive cells leads to primary cilia
loss, biliary cell proliferation and cystic liver lesions in
TAA-treated mice. Biochim Biophys Acta Mol Basis Dis.
1868:1663352022. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Maglione M, Tosco A, Borrelli M and
Santamaria F: Primary ciliary dyskinesia treatment: time for a new
approach? Lancet Respir Med. 12:2–3. 2024. View Article : Google Scholar
|
|
114
|
Jeon HH, Kang J, Li JM, Kim D, Yuan G,
Almer N, Liu M and Yang S: The Effect of IFT80 deficiency in
osteocytes on orthodontic loading-induced and physiologic bone
remodeling: In vivo study. Life (Basel). 12:11472022.PubMed/NCBI
|
|
115
|
Li X and Yang S, Han L, Mao K and Yang S:
Ciliary IFT80 is essential for intervertebral disc development and
maintenance. FASEB J. 34:6741–6756. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Liu J, Xie X, Yan D, Wang Y, Yuan H, Cai
Y, Luo J, Xu A, Huang Y, Cheung CW, et al: Up-regulation of FoxO1
contributes to adverse vascular remodelling in type 1 diabetic
rats. J Cell Mol Med. 24:13727–13738. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Li D, Chen K, Tang H, Hu S, Xin L, Jing X,
He Q, Wang S, Song J, Mei L, et al: A Logic-based diagnostic and
therapeutic hydrogel with multistimuli responsiveness to
orchestrate diabetic bone regeneration. Adv Mater. 34:e21084302022.
View Article : Google Scholar
|
|
118
|
Liu J, Cheng Q, Wu X, Zhu H, Deng X, Wang
M, Yang S, Xu J, Chen Q, Li M, et al: Icariin treatment rescues
diabetes induced bone loss via scavenging ROS and activating
primary Cilia/Gli2/osteocalcin signaling pathway. Cells.
11:40912022. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wei J, Shimazu J, Makinistoglu MP, Maurizi
A, Kajimura D, Zong H, Takarada T, Lezaki T, Pessin JE, Hinoi E and
Karsenty G: Glucose uptake and Runx2 synergize to orchestrate
osteoblast differentiation and bone formation. Cell. 161:1576–1591.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Girardet L, Bernet A, Calvo E, Soulet D,
Joly-Beauparlant C, Droit A, Cyr DG and Belleannee C: Hedgehog
signaling pathway regulates gene expression profile of epididymal
principal cells through the primary cilium. FASEB J. 34:7593–7609.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Dores C, Alpaugh W, Su L, Biernaskie J and
Dobrinski I: Primary cilia on porcine testicular somatic cells and
their role in hedgehog signaling and tubular morphogenesis in
vitro. Cell Tissue Res. 368:215–223. 2017. View Article : Google Scholar
|
|
122
|
Hoi KK, Xia W, Wei MM, Ulloa Navas MJ,
Garcia Verdugo JM, Nachury MV, Reiter JF and Fancy SPJ: Primary
cilia control oligodendrocyte precursor cell proliferation in white
matter injury via Hedgehog-independent CREB signaling. Cell Rep.
42:1132722023. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Barbeito P and Garcia-Gonzalo FR: HTR6 and
SSTR3 targeting to primary cilia. Biochem Soc Trans. 49:79–91.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Yuan S, Li J, Diener DR, Choma MA,
Rosenbaum JL and Sun Z: Target-of-rapamycin complex 1 (Torc1)
signaling modulates cilia size and function through protein
synthesis regulation. Proc Natl Acad Sci USA. 109:2021–2026. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Finetti F, Cassioli C, Cianfanelli V,
Onnis A, Paccagnini E, Kabanova A and Baldari CT: The
intraflagellar transport protein IFT20 controls lysosome biogenesis
by regulating the post-Golgi transport of acid hydrolases. Cell
Death Differ. 27:310–328. 2020. View Article : Google Scholar
|
|
126
|
Guo DF, Lin Z, Wu Y, Searby C, Thedens DR,
Richerson GB, Usachev YM, Grobe JL, Sheffield VC and Rahmouni K:
The BBSome in POMC and AgRP neurons is necessary for body weight
regulation and sorting of metabolic receptors. Diabetes.
68:1591–1603. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Tao YX: Constitutive activity in
melanocortin-4 receptor: Biased signaling of inverse agonists. Adv
Pharmacol. 70:135–154. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Drygalski K, Higos R, Merabtene F, Mojsak
P, Grubczak K, Ciborowski M, Razak H, Clement K and Dugail I:
Extracellular matrix hyaluronan modulates fat cell differentiation
and primary cilia dynamics. Biochim Biophys Acta Mol Cell Biol
Lipids. 1869:1594702024. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Ritter A, Friemel A, Kreis NN, Hoock SC,
Roth S, Kielland-Kaisen U, Brüggmann D, Solbach C, Louwen F and
Yuan J: Primary cilia are dysfunctional in obese adipose-derived
mesenchymal stem cells. Stem Cell Reports. 10:583–599. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Xun Y, Jiang Y, Xu B, Tang M, Ludwig S,
Nakamura K, Mukhopadhyay S, Liu C, Beutler B and Zhang Z: GPR45
modulates Gαs at primary cilia of the paraventricular
hypothalamus to control food intake. Science. 388:adp39892025.
View Article : Google Scholar
|