Introduction
Following menopause, the risk of cardiovascular
disease (CVD) rises sharply compared with that in younger patients,
making it the primary cause of death in elderly female patients
(1,2). Patients who transition through
menopause at an age less than 45 years are at a 50% increased CVD
risk and they also have a 10% increased risk of fatal coronary
heart disease and all-cause mortality (1). In these patients, hyperlipidemia,
characterized by abnormally elevated LDL-C and TC, is common
(3,4). HDL-C levels are not associated with
menopause status, but higher LDL-C, TG, and TC levels are observed
in postmenopausal than in premenopausal women (3). Patients who experienced natural
menopause were at 3-times greater risk of atherosclerosis compared
with premenopausal patients and menopause is a risk factor for
age-related increase in arterial stiffness (5). Another peer study also showed a
higher rate of CVD events in postmenopausal patients (6). Age at menopause is not only a sign
of reproductive aging, but also a high-risk indicator of CVD
(7). In addition, the decline of
ovarian function and the decrease in estrogen can lead to physical
and psychological symptoms in older patients, including
osteoporosis, cognitive decline, night sweats, hot flashes,
insomnia, depression and metabolic abnormality (8,9).
These factors have a serious impact on the life and health of
elderly patients. Hormone therapy is commonly used for
postmenopausal patients at high risk of CVD, but studies have
reported that the disadvantages of hormone therapy may outweigh the
benefits and that the efficacy of treatment is influenced by
factors such as dosage used, mode of administration and time of
menopause (10,11). Consequently, devising safer and
more efficacious therapy for preventing and treating dyslipidemia
associated with menopause is important.
Polysaccharides can ameliorate metabolic
dysfunction-associated steatotic liver disease (MASLD), type 2
diabetes and obesity by modulating lipid metabolism, suppressing
inflammation, scavenging free radicals and reshaping the gut
microbiota (12-14). For example, hawthorn
polysaccharides significantly ameliorate high-fat diet
(HFD)-induced MASLD by modulating the gut microbiota and
ameliorating metabolic disorders in the liver of mice (15). Ganoderma lucidum has been
employed medicinally in China and across much of Asia (16). G. lucidum polysaccharides
(GLPs), the primary bioactive constituent, have demonstrated
antitumor (17), antioxidant,
immunomodulatory and lipid-metabolism-modulating activity (18). G. lucidum β-D-glucan forms
a hydrogen-bonded stable triple helix structure and its biological
activity depends on the β configuration (18). Differences in separation methods
and culture media lead to a diverse structure of polysaccharides,
including pure β-glucan, isosaccharides, heteromannans and their
peptide complexes (19). In
different animal models, GLP can induce the synthesis of superoxide
dismutase, succinate dehydrogenase and glutathione (GSH) and
decrease levels of TC, triglyceride (TG) and LDL-C, thereby
decreasing hyperlipidemia (18,20,21). GLP regulates glucose metabolism,
such as downregulating serum glucose and insulin (INS) levels,
inhibiting β cell apoptosis and promoting the regeneration of β
cells (22,23). In addition, GLP modulates
intestinal dysbiosis: In mice exposed to azoxymethane/dextran
sulfate sodium, GLP suppresses colitis, inflammation and
tumorigenesis, and simultaneously corrects microbial imbalances
while alleviating endotoxemia (24). In diabetic rats, GLP lowers
inflammation and enriches beneficial gut microbes, thereby
bolstering host defense against pathogens (25). Hence, investigating how GLP
ameliorates metabolic disorders in postmenopausal patients along
with its underlying mechanisms is imperative.
There is a connection between intestinal flora and
estrogen: A specific part of the intestinal flora uses
β-glucosidase to convert the bound form of estrogen into a free,
active state, which binds receptors and exerts effects, such as
stimulating the development of ovarian follicles, enhancing bone
density and providing cardiovascular protection (26,27). In postmenopausal patients, the
composition and quantity of intestinal microbiota change, which may
also be one of the mechanisms of metabolic disorders (28). Patients experiencing menopausal
vasomotor disorders tend to have a notable decline in the primary
representatives of Bifidobacterium and Lactobacillus,
meanwhile, there is an increase in the quantities of
Klebsiella and Clostridioides difficile (29,30). On the other hand, the liver, as
the hub of material metabolism, plays a key role in regulating
metabolism, including regulating the synthesis and metabolism of
lipids, sugar and proteins (31). A large number of clinical studies
support the role of gut microbiota in liver disease and an
imbalanced gut microbiota affects the metabolic and inflammatory
state of the liver, which affects the disease process, including
hepatocellular carcinoma, hepatitis and liver cirrhosis (32,33). Polysaccharides derived from G.
lucidum spores curb obesity and inflammation while attenuating
hyperlipidemia through modulation of the gut microbiota (24). Therefore, based on the gut-liver
axis theory, although GLP is not easily absorbed by the intestine,
it may regulate liver metabolism by affecting the intestinal flora,
thereby improving metabolic disorder.
The present study used a mouse model of ovariectomy
to simulate ovarian hormone deficiency in elderly female patients
and explored the regulatory effect and molecular mechanism of GLP
on body weight, liver lipids, circadian rhythm, INS resistance and
intestinal microbiota, providing a theoretical basis for the
potential use of GLP in the treatment of lipid metabolic disorder
of postmenopausal patients.
Materials and methods
Animal experiments
All the animal experiments were approved by the
Experimental Animal Ethics Committee of Guangdong Pharmaceutical
University (Guangzhou, China; approval no. gdpulac2020065) and
adhered to ARRIVE2 (34). In
total, 72 female C57BL/6 mice (weight, 17-19 g; age, 7 weeks)
purchased from Hunan Lex Jingda Laboratory Animal Co., Ltd. were
housed in a specific pathogen-free animal facility at 25°C, 60-65%
humidity, 12/12-h light/dark cycle, with free access to water and
food. To observe the anesthetic concentration and postoperative
condition of the mice, three mice were randomly selected, two for
ovariectomy and one for sham surgery. This was repeated once/day
for 3 consecutive days, confirming that a 3% isoflurane anesthetic
concentration was suitable for surgery and that the mice were in
good condition after the operation. Following 1 week of adaptive
feeding, mice were randomized and 63 mice were used for the
subsequent experiments. Groups were as follows: Sham operation
(n=10); bilateral ovariectomy (OVX, n=11); OVX + low-dose (100
mg/kg) GLP (OVX + LGLP, n=11); OVX + medium-dose (200 mg/kg) GLP
(OVX + MGLP, n=10); OVX + high-dose (400 mg/kg) GLP (OVX + HGLP,
n=10) and OVX + simvastatin (Siv; 10 mg/kg; n=11). Siv was chosen
as the positive reference drug. Mice were placed in an induction
chamber with 3% isoflurane in oxygen for anesthesia induction.
Following loss of consciousness, animals were maintained with 2%
isoflurane for bilateral ovariectomy. For the sham operation group,
ovaries were removed and replaced, with all other procedures
identical to the ovariectomy group. GLP (cat. no. Ksm083, Ciyuan
Biotechnology) was dissolved in sterile water and given once daily
by gavage. Every afternoon at 2.00, after weighing the mice in each
group, gavage was performed. Sham group and OVX group mice were
gavaged with the corresponding volume of sterile water. The body
weight of the mice was measured daily. Humane endpoints were as
follows: Weight loss >20%, complete refusal to eat/drink for 24
h, inability to stand for 48 h, severe depression/coma,
uncontrollable infection or severe respiratory distress. Animals
were monitored once daily. A total of 63 animals were used; no
animals reached these humane endpoints, and all animals were
euthanized at the planned experimental endpoints. Following 8 weeks
of gavage, the mice were fasted for 8 h. Animals were induced and
maintained with 3% isoflurane in oxygen until loss of consciousness
(absent righting reflex) and absent toe pinch reflex. Animals were
maintained at 3% isoflurane for 3-5 min for deep anesthesia.
Terminal blood collection (0.8-1.0 ml, ≤50% of total blood volume)
was performed via retro-orbital sinus puncture using heparinized
capillary tubes. Immediately after blood collection, mice were
euthanized by cervical dislocation while under anesthesia. Death
was confirmed by cessation of heartbeat and respiratory arrest.
Liver tissue was collected for biochemical, histological and
transcriptional testing. Stool samples of the mice were collected
in the last week of the experimental process and frozen at
−80°C.
GLP characterization
The monosaccharide composition was determined by
high performance anion electronic chromatography (HPAEC) with
electrochemical detectors (EDs). In brief, GLP (5 mg) was added to
3 M trifluoroacetic acid. After that, the solution was heated for 3
h at 120°C. The hydrolyzed products were thoroughly dissolved in 5
ml ddH2O with a vortex, then, 50 μl liquid was
diluted with 950 μl ddH2O. The solution was
centrifuged at a rate of 13,400 × g for 5 min at room temperature.
A membrane with a 0.22 μm pore size was used to filter the
supernatant before examination with an HPAEC-ED fitted with a
Dionex Carbopac PA20 column (Thermo Fisher Scientific, 150.0×3.0
mm). The mobile phase elution was performed at the flow rate of 0.3
ml/min using the following mobile phase: A, H2O; B, 15
mM NaOH; and C, 15 mM NaOH and 100 mM NaOAc. The injection volume
was 25 μl and the column temperature was 30°C. Elution
gradient was performed as follows: 0 min A/B/C (98.8:1.2:0.0), 18
min A/B/C (98.8:1.2:0.0, V/V), 20 min A/B/C (30:70:0,V/V), 30 min
A/B/C (30:70:0, V/V), 30.1 min A/B/C (0:0:100, V/V), 46 min A/B/C
(0:0:100, V/V), 46.1 min A/B/C (0:100:0, V/V), 50 min A/B/C
(0:100:0, V/V), 50.1 min A/B/C (98.8:1.2:0.0, V/V) and 80 min A/B/C
(98.8:1.2:0.0,V/V).
Polysaccharide infrared spectral
detection
A total of 2 mg GLP and 200 mg potassium bromide
were pressed into a pellet, with potassium bromide powder pressed
into a pellet as a blank control. Fourier Transform Infrared
Spectrometer FT-IR650 (Tianjin Gangdong Technology Development Co.,
Ltd.) was used for scanning and recording.
Blood biochemical profile assay
The levels of TC, TG, high-density lipoprotein
cholesterol (HDL-C), LDL-C, estradiol (E2), INS, free fatty acid
(FFA), alanine amino transferase (ALT) and aspartate
aminotransferase (AST) in serum and fasting blood glucose (Roche
Diabetes Care, Inc.; cat. nos. C8012 and P003874) were measured
according to the manufacturer's protocols using kits [cat. nos.
A111-1-1, A110-1-1, A112-1-1, A113-1-1 (all Nanjing Jiancheng
Bioengineering Institute), MM-0546M1, MM-0579M1 and MM-0326M1 (all
Meimian Bioengineering Company), C009-2-1 and C010-2-1 (both
Nanjing Jiancheng Bioengineering Institute), respectively].
Hematoxylin and eosin (H&E)
staining
The liver tissue was fixed in 4% paraformaldehyde at
4°C overnight for histological staining. The 4-μm-thick
paraffin sections were stained with hematoxylin (cat. no.
H9627-100G) for 3 min and eosin (cat. no. E4009-25G; both
Sigma-Aldrich; Merck KGaA) for 20 sec at room temperature. The
images were captured with the Olympus DP74 light microscope and
analyzed using Fijj (imagej.net/Contributors 2.14.0/1.54f).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was obtained from each mouse liver using
RNAiso plus reagent (cat. no. T9109, Takara Bio, Inc.) and
subjected to RT using the Prime Script™ RT Reagent kit (cat. no.
RR047A, Takara Bio, Inc.) at 37°C for 15 min and 85°C for 5 sec.
qPCR was performed using the SYBR Premix Ex Taq kit (cat. no.
RR820A, Takara Bio, Inc.) with the Light Cycler 480II System
(Roche, Inc.). Thermocycling conditions were as follows: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles of 95°C for
5 sec, 60°C for 20 sec and 65°C for 15 sec. GAPDH was used as the
internal reference. All primers used are listed in Table SI. Relative mRNA expression
levels were calculated using the 2^−ΔΔCq method (35).
16S rDNA gene analysis
The feces of the mice were collected the day before
tissue collection and stored in a −80°C freezer. Then, fecal
samples of 6 mice were randomly selected from each group for 16S
rDNA sequencing. Gene Denovo Biotechnology Co., Ltd. performed the
16S rDNA high-throughput sequencing analysis. Genomic DNA was
extracted using the HiPure Stool DNA kit (cat. no. D3141; Guangzhou
Meiji Biotechnology Co., Ltd.). The quality and integrity of the
processed DNA were verified using a NanoDrop 2000
micro-spectrophotometer (Thermo Fisher Scientific) to determine
purity and by 2% agarose gel electrophoresis to confirm intact
genomic DNA. The V3-V4 region of the 16S rDNA was amplified using
primers as follows: forward, 5'-CCTACGGGNGGCWGCAG-3'; and reverse,
5'-GGACTACHVGGGTATCTAAT-3'. The purified amplification products
were ligated to sequencing adapters to construct a sequencing
library using the Illumina DNA Prep Kit (cat. no. 20060059;
Illumina Inc.). PCR products were purified using AMPure XP Beads
(Beckman Coulter) and quantified with a Qubit 3.0 fluorometer
(Thermo Fisher Scientific). Library quality was validated using an
ABI StepOnePlus Real-Time PCR System (Applied Biosystems). The
pooled sequencing library (300 pM) was sequenced using the NovaSeq
6000 S2 Reagent Kit (cat. no. 20042038; Illumina Inc.) on an
Illumina NovaSeq 6000 platform in paired-end 250 bp (PE250) mode.
USEARCH (version 11.0.667; drive5.com/usearch/) was used for operational
taxonomic unit clustering (97% similarity) with the UPARSE (version
11.0.667; drive5.com/uparse/) algorithm, UCHIME
(version 11.0.667; drive5.com/usearch/manual/uchime_algo.html) for
chimera removal, and the SILVA database (version 138.2; arb-silva.
de/) for taxonomic annotation (36,37). Alpha diversity was assessed using
the Shannon diversity index (38). Beta diversity was evaluated by
principal coordinate analysis (PCoA) based on Bray-Curtis
dissimilarity matrices (39,40). H'=−∑i=1RpiIn(pi), where R is the total
number of species (richness), and p_i is the proportion of
individuals belonging to the i-th species. Higher Shannon values
indicate greater diversity within the community. Functional
prediction analysis. The functional profiles of the gut microbiota
were predicted using PICRUSt2 based on the 16S rRNA gene sequencing
data (41). The predicted
functional pathways were annotated against the KEGG (Kyoto
Encyclopedia of Genes and Genomes) database (42,43). Differential functional pathways
between groups were identified using Statistical Analysis of
Metagenomic Profiles; version 2.1.3) with two-sided Welch's t-test
and Benjamini-Hochberg FDR correction (q<0.05) (44).
Western blotting
The liver tissues of the mice of each group were
homogenized in RIPA lysis buffer (Dalian Meilun Biotechnology) and
centrifuged (13,400 × g, 4°C, 20 min) to collect the supernatant.
Protein concentration was determined with a BCA assay kit. Aliquots
(40 μg protein/lane) were resolved by 8-15% SDS-PAGE and
transferred to PVDF membranes. Following blocking with 5% skimmed
milk/TBST (0.1% Tween-20, 1.5 h, room temperature), the membranes
were probed with the primary antibodies (4°C, overnight) followed
by HRP-conjugated secondary antibodies (1 h, room temperature).
Protein signals were visualized using ECL and quantified with
ImageJ (v1.53k; National Institutes of Health). Antibody details
are provided in Table SII.
GAPDH, H3 and β-actin were used as internal controls.
Nuclear magnetic resonance (NMR) whole
body composition analysis
The lean, fat mass, and liquid mass were assessed
using a whole body composition analyzer (minispec LF90II; Bruker
BioSpin). Conscious mice were placed into a plastic holder and
inserted into the NMR magnet bore. Body composition was determined
by time-domain nuclear magnetic resonance.
Statistical analysis
The experimental data were statistically analyzed
and plotted using GraphPad Prism 8.0.2 (Dotmatics). One- or two-way
repeated measures ANOVA was used to determine the differences
between different groups, followed by Tukey's multiple comparisons
post hoc test. P<0.05 was considered to indicate a statistically
significant difference. All data are presented as the mean ± SEM.
The experiments were independently repeated ≥3 times.
Results
GLP improves abnormal serum indexes and
liver lipid accumulation of the OVX mice
GLP characterization was consistent with what is
described in the literature (45) (Fig. S1). During the 8 weeks of the
gavage, the OVX group showed a significantly faster rate of weight
gain compared with the sham group. Pharmacological intervention
moderated this weight gain, with all GLP-treated and the
simvastatin group having significantly lower body weight than the
OVX group (Figs. 1A and S2A). There was no significant
difference in the average food intake of the mice in each group
(Fig. S2B). Whole-body
time-domain-NMR revealed that OVX group displayed the lowest lean
mass and the highest fat mass across all groups (Fig. S2C and D). The lean mass of the
OVX + HGLP group was increased relative to the OVX group (Fig. S2C). The fat mass of the OVX +
MGLP group was decreased compared with the OVX group (Fig. S2D). The liquid mass of the OVX +
HGLP group was decreased compared with the OVX group (Fig. S2E). Consistent with successful
ovariectomy, serum E2 was markedly suppressed in every OVX group
compared with Sham controls (Fig.
1B).
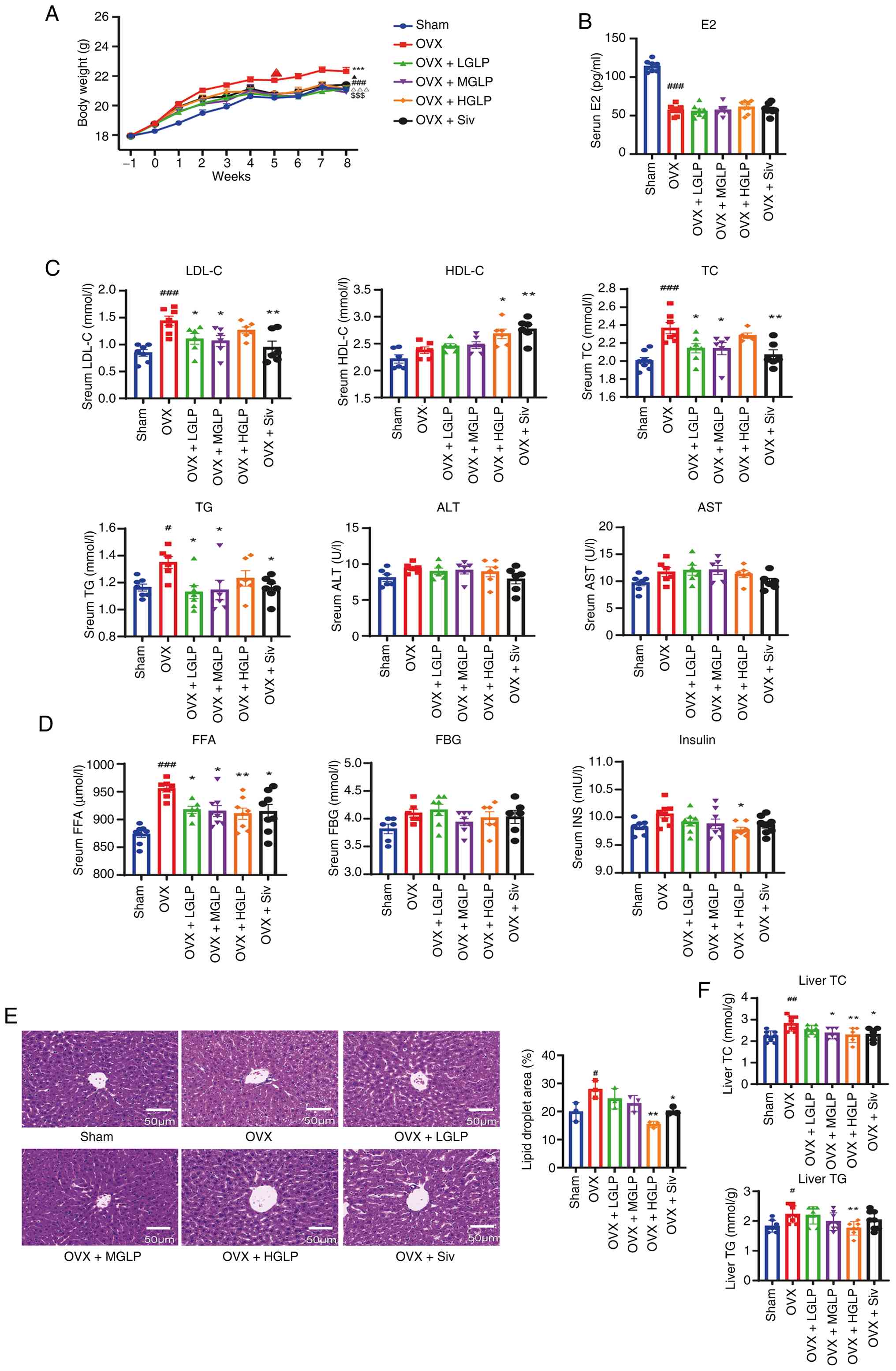 | Figure 1Effects of GLP on body weight, blood
biochemical indexes and liver lipid content. (A) Body weight
curves. ###P<0.005, Sham vs. OVX;
***P<0.005, OVX + LGLP vs. OVX;
$$$P<0.005, OVX + MGLP vs. OVX;
ΔΔΔP<0.005, OVX + HGLP vs. OVX;
▲P<0.05, OVX + Siv vs. OVX. Concentration of (B) E2
and (C) TG, TC, LDL-C, HDL-C, FFA, insulin and FBG in the serum.
(D) Liver injury indexes including ALT and AST. (E) Hematoxylin and
eosin staining and quantitative analysis of liver lipid droplet
area of the liver tissue. Scale bar, 50 μm. (F) TG and TC
content in the liver (n≥6). #P<0.05,
##P<0.01, ###P<0.005 vs. Sham;
*P<0.05, **P<0.01 vs. OVX. GLP,
Ganoderma lucidum polysaccharides; OVX, ovariectomy; LGLP,
low-dose GLP; MGLP, medium-dose GLP; HGLP, high-dose GLP; Siv,
simvastatin; E2, estradiol; LDL-C, low-density lipoprotein
cholesterol; HDL-C, high-density lipoprotein cholesterol; TC, total
cholesterol; TG, triglyceride; ALT, alanine aminotransferase; AST,
aspartate transaminase; FFA, free fatty acid; FBG, fasting plasm
glucose. |
Serum lipid profiling showed that the LDL-C levels
in mice of the OVX group increased compared with Sham controls.
Serum LDL-C was decreased in the OVX + LGLP, MGLP and Siv groups
compared with OVX group (Fig.
1C). In addition, HDL-C was upregulated in the HGLP and Siv
groups compared with OVX group (Fig.
1C). The TC levels of the OVX group significantly increased
compared with Sham group, and the TC levels of the OVX + LGLP, MGLP
and Siv groups were decreased compared with OVX group (Fig. 1C). The TG levels increased in the
OVX compared with Sham controls, and decreased in the OVX + LGLP,
MGLP and Siv groups compared with the OVX group (Fig. 1C). Compared with the OVX group,
the OVX + HGLP group showed a decrease in serum TC and TG levels
although these were not significant (Fig. 1C). No inter-group differences
were observed for ALT and AST (Fig.
1C). The FFA levels increased in the OVX group compared with
the Sham controls, and decreased following administration of
different doses of GLP or Siv (Fig.
1D). The levels of INS decreased in the OVX + HGLP group
compared with the OVX group (Fig.
1D). There was no significant change in FBG between different
groups (Fig. 1D).
The H&E staining confirmed that the OVX mice had
more and larger lipid droplets in the liver, which demonstrated had
liver fat accumulation, and HGLP significantly attenuated this
lipid accumulation (Fig. 1E).
Moreover, hepatic TC and TG were elevated in OVX compared with the
Sham group, and the levels of TC in the livers of the OVX + MGLP
and Siv groups were downregulated and the levels of TC and TG in
the liver of the OVX + HGLP group were decreased compared with the
OVX group (Fig. 1F).
GLP improves abnormal cholesterol
synthesis of the OVX mice
Relative to Sham controls, hepatic sterol regulatory
element binding transcription factor 2 (Srebp2) and
3-hydroxy-3-methylglutaryl-CoA reductase (Hmgcr)
transcription was markedly elevated in the OVX group; this effect
was decreased by GLP or Siv. The OVX + LGLP, MGLP and HGLP groups
also showed effects similar to the OVX + Siv group in decreasing
the transcription of Srebp2 and Hmgcr (Fig. 2A). The mevalonate kinase
(Mvk) mRNA was increased in OVX group compared with the Sham
group, and this increase was decreased in the OVX + LGLP, HGLP and
Siv groups (Fig. 2A). The
diphosphomevalonate decarboxylase (Mvd) mRNA levels of the
OVX + LGLP, HGLP and Siv groups were decreased compared with the
OVX group (Fig. 2A). Mvk
mRNA and protein changes in the OVX + MGLP group were not
significant compared with the OVX + LGLP, HGLP and Siv groups
(Fig. 2A). In addition, compared
with the OVX group, only the OVX + HGLP group exhibited decreased
transcription of farnesyl-diphosphate farnesyltransferase 1 and
isopentenyl-diphosphate delta isomerase 1 (Idi1) (Fig. 2A). Hepatic farnesyl diphosphate
synthase (Fdps) transcription in the OVX group was
significantly upregulated compared with Sham controls, and this
effect was decreased in the OVX + LGLP, HGLP and Siv groups.
Compared with the OVX + Siv group, the OVX + MGLP group did not
show significant transcriptional change of Fdps (Fig. 2A). The lanosterol synthase
(Lss) mRNA expression decreased in the OVX + HGLP and Siv
groups compared with the OVX group. There was no significant
difference in transcript levels of Lss in the OVX + LGLP and
MGLP groups (Fig. 2A). Compared
with Sham controls, the expression of HMGCR protein in OVX mice was
upregulated, and this change was reversed by MGLP, HGLP or Siv
(Figs. 2B, C and S3A and B). LSS protein expression of
the OVX group was increased compared with the Sham controls, and
LSS protein expression of the OVX + HGLP group and Siv group was
decreased compared with the OVX group (Fig. 2B and C). Hepatic MVK protein
levels markedly increased in OVX mice compared with Sham controls,
and were significantly attenuated following treatment with either
HGLP or Siv (Figs. 2B, C and
S3A and B). Hepatic IDI1
protein expression was markedly elevated in OVX mice relative to
Sham controls and significantly reduced by HGLP or Siv (Figs. 2B, C and S3A and B). MVD protein expression
exhibited no marked variation between groups (Fig. 2B and C).
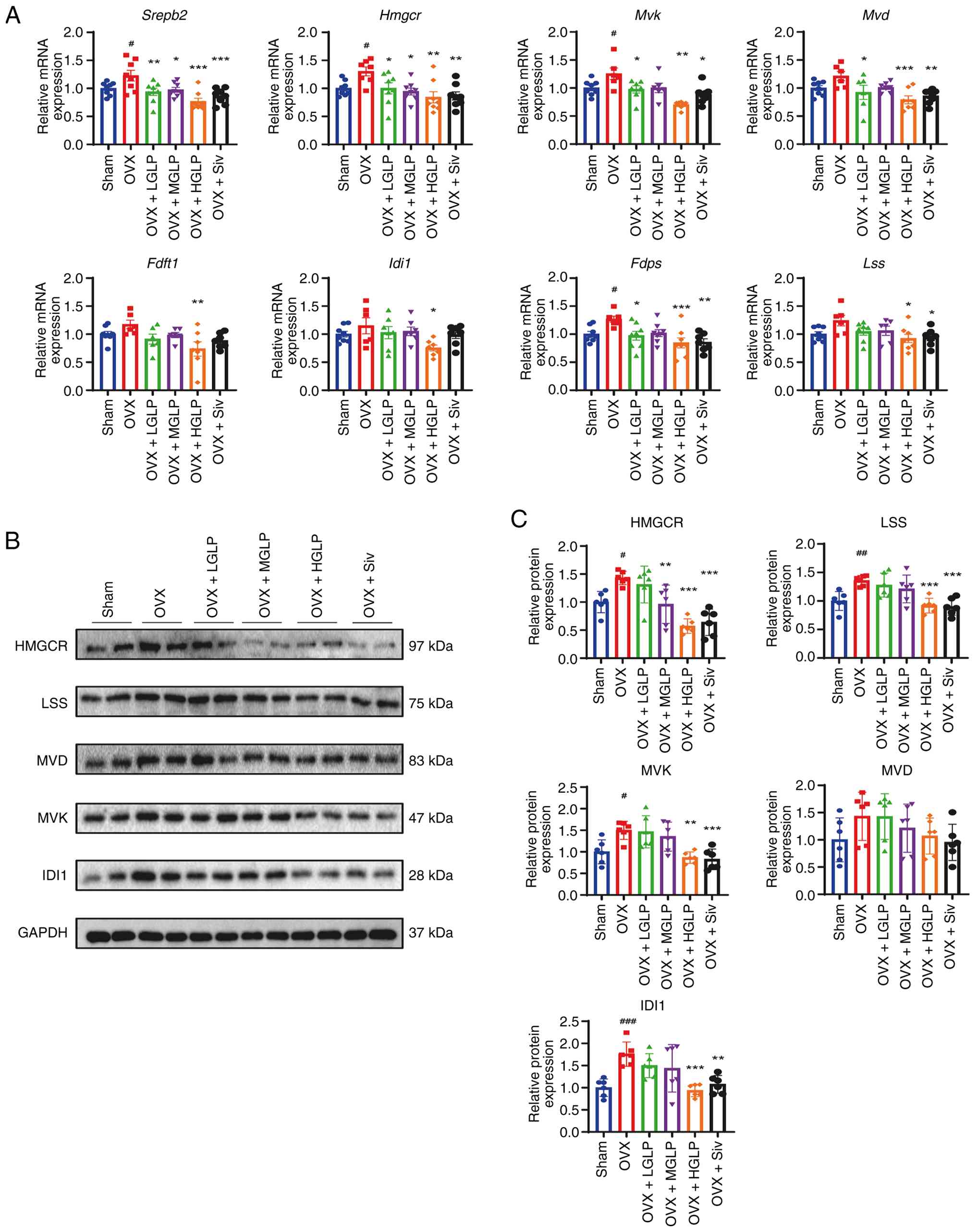 | Figure 2Effect of GLP on the expression of
cholesterol synthesis-associated genes in the liver tissue. (A)
Relative mRNA expression levels of Srebp2, Hmgcr,
Mvk, Mvd, Fdft1, Idi1, Fdps and
Lss in the liver tissue. Protein levels detected by (B)
western blot and (C) densitometry of HMGCR, LSS, MVK, MVD and IDI1
in the liver (n≥6). #P<0.05, ##P<0.01,
###P<0.005 vs. Sham; *P<0.05,
**P<0.01, ***P<0.005 vs. OVX. GLP,
Ganoderma lucidum polysaccharides; OVX, ovariectomy; LGLP,
low-dose GLP; MGLP, medium-dose GLP; HGLP, high-dose GLP; Siv,
simvastatin; Srebp2, sterol regulatory element-binding protein 2;
Hmgcr, 3-hydroxy-3-methylglutaryl-coenzyme A reductase; Mvk,
mevalonate kinase; Mvd, diphosphomevalonate decarboxylase; Fdft1,
farnesyl-diphosphate farnesyltransferase 1; Idi1,
isopentenyl-diphosphate delta isomerase 1; Fdps, farnesyl
diphosphate synthase; Lss, lanosterol synthase. |
GLP downregulates hepatic transcription
of FA biosynthetic enzymes in OVX mice
Hepatic Srebp1-c transcription of the OVX
group was upregulated compared with the Sham controls, and
Srebp1-c mRNA expression was downregulated by all doses GLP
or Siv (Fig. 3A). Peroxisome
proliferator-activated receptor α (Pparα) mRNA was markedly
elevated in the livers of the OVX + MGLP group and HGLP group
compared with the OVX group (Fig.
3A). In addition, Pparγ transcription levels after
treatment with LGLP, HGLP or Siv were significantly lower compared
with the OVX group (Fig. 3A).
The mRNA expression level of Cd36 in the OVX mice was
increased compared with Sham controls. GLP and Siv reduced
Cd36 mRNA expression compared with the OVX group (Fig. 3A). The stearoyl-CoA desaturase 1
transcription level in the OVX + MGLP group was decreased compared
with the OVX group (Fig. 3A).
The Fasn and acetyl CoA carboxylase (Acc) mRNA
expression levels in the mice of the OVX group were higher than the
Sham controls, and this effect was decreased by all doses of GLP or
Siv (Fig. 3A). FASN protein
expression in the OVX group was increased compared with the Sham
controls, and decreased in the OVX + MGLP, HGLP and Siv groups
compared with OVX group (Fig. 3B and
C). Hepatic ACC protein abundance was markedly elevated in OVX
mice relative to Sham controls and was significantly attenuated by
HGLP (Fig. 3B and C).
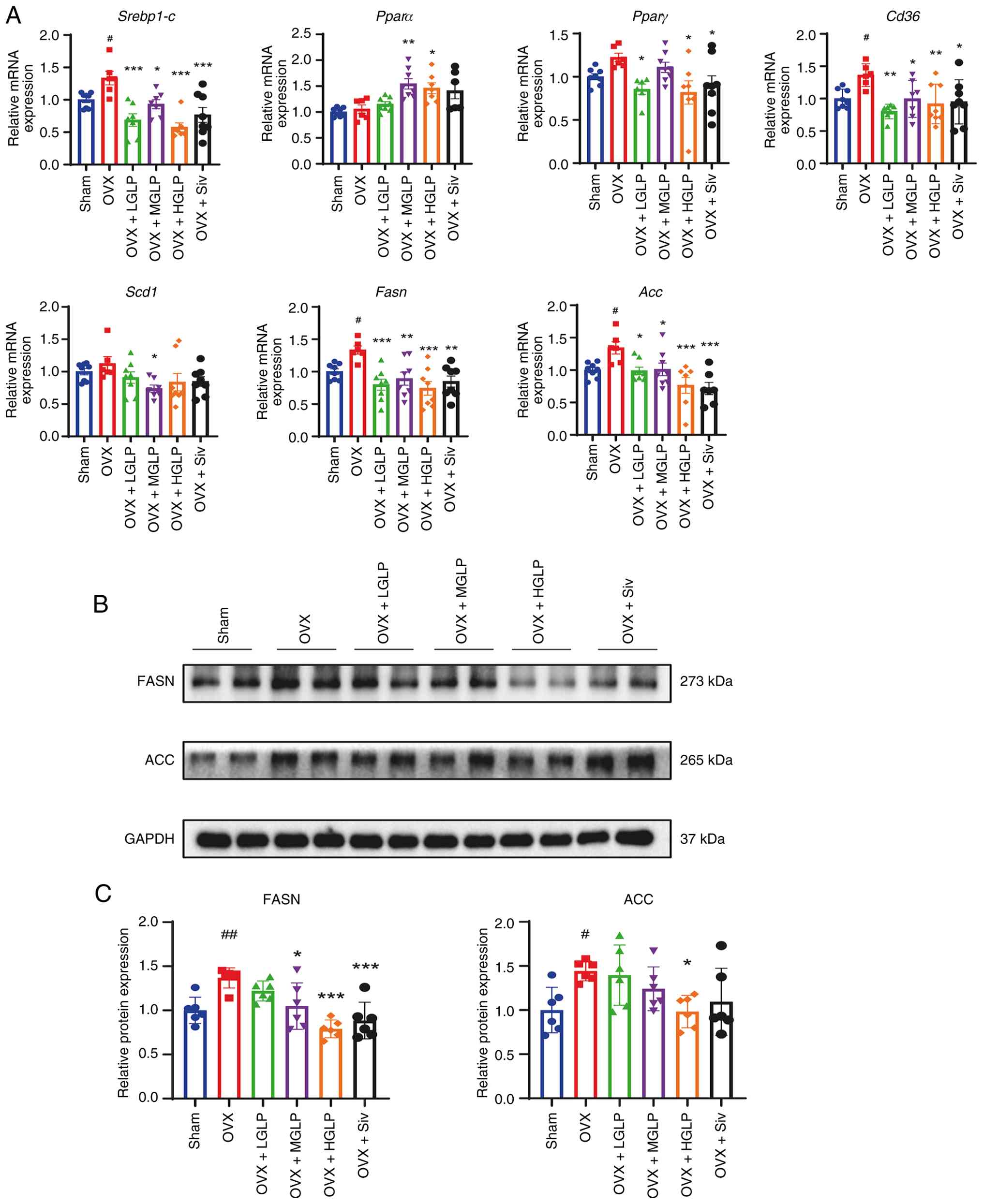 | Figure 3Effects of GLP on fatty acid
synthesis of liver tissue. (A) mRNA expression of Srebp1-c,
Pparα, Pparγ, Cd36, Scd1, Fasn
and Acc at in the liver. (B) Western blotting and (C)
densitometry of FASN and ACC in the liver (n≥6).
#P<0.05, ##P<0.01 vs. Sham;
*P<0.05, **P<0.01,
***P<0.005 vs. OVX. GLP, Ganoderma lucidum
polysaccharides; OVX, ovariectomy; LGLP, low-dose GLP; MGLP,
medium-dose GLP; HGLP, high-dose GLP; Siv, simvastatin; Srebp1-c,
sterol regulatory element-binding protein 1c; Pparα, peroxisome
proliferator-activated receptor α; Scd1, stearoyl-CoA desaturase 1;
Fasn, fatty acid synthase; Acc, acetyl CoA carboxylase. |
The ratio of phosphorylated (p-)glycogen
synthase/glycogen synthase in the OVX + MGLP group and the HGLP
group was significantly decreased compared with OVX group. There
were no significant difference in the ratio of p-IRS1 (insulin
receptor substrate (IRS1)/IRS1 and the ratio of p-mTOR/mTOR between
groups (Fig. 4A and B).
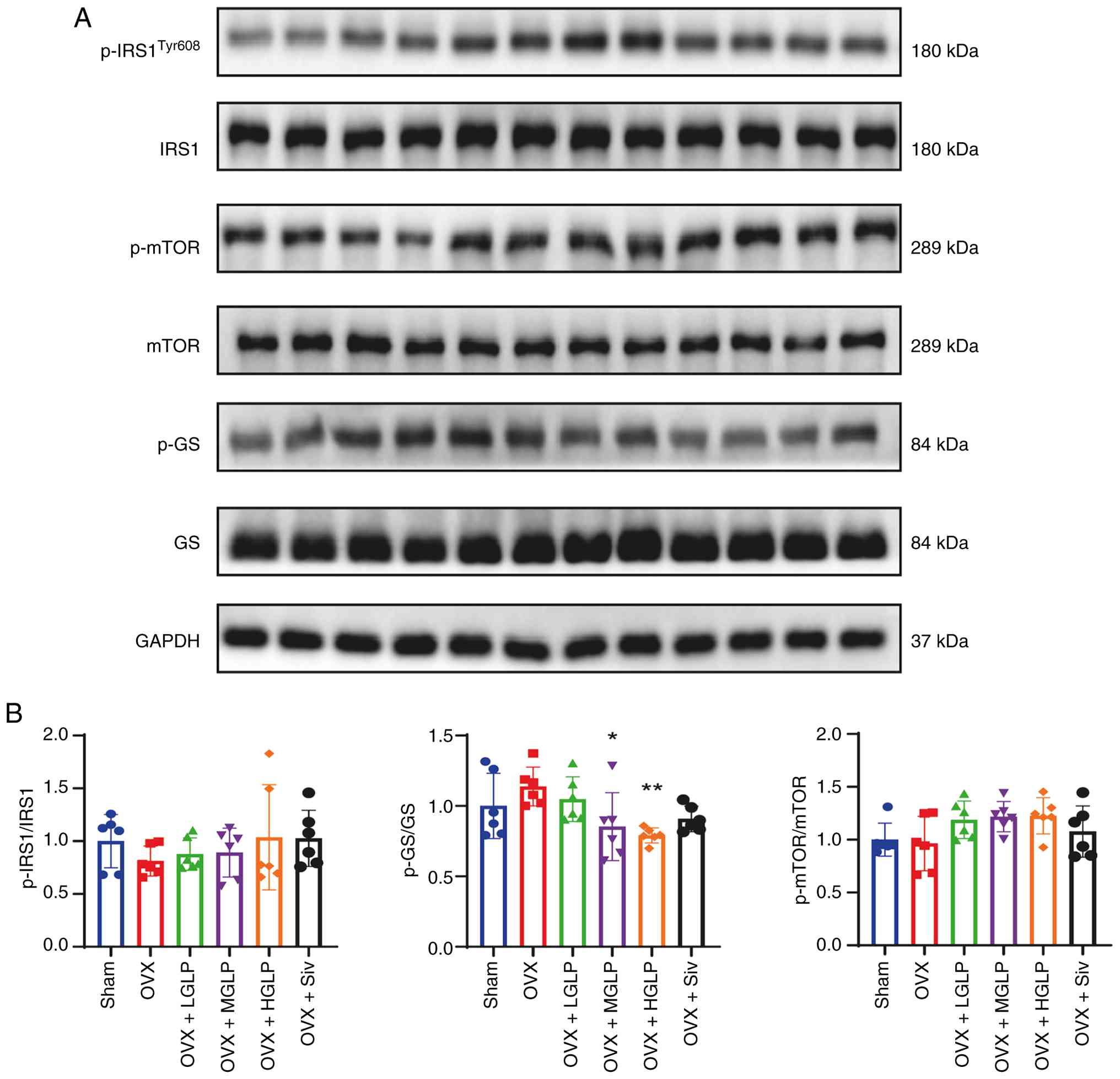 | Figure 4Effects of GLP on the expression of
proteins involved in the insulin signaling pathway in the liver.
(A) Expression of (B) p-IRS1, Tyr609, IRS1, p-mTOR, mTOR, p-GS and
GS proteins in the liver (n≥6). *P<0.05,
**P<0.01 vs. OVX. GLP, Ganoderma lucidum
polysaccharides; OVX, ovariectomy; LGLP, low-dose GLP; MGLP,
medium-dose GLP; HGLP, high-dose GLP; Siv, simvastatin; p-,
phosphorylated; IRS1, insulin receptor substrate 1; GS, glycogen
synthase. |
GLP restores the abnormal expression of
genes related to circadian rhythm in the OVX mice
Hepatic clock circadian regulator (Clock)
transcription was significantly elevated in OVX + MGLP and HGLP
livers relative to the OVX group (Fig. 5A). Basic helix-loop-helix ARNT
like 1 (Bmal1) mRNA expression was markedly suppressed in
the OVX group relative to Sham controls, and this change was
restored by MGLP or HGLP (Fig.
5A). Hepatic period circadian regulator 1 transcription was
significantly upregulated in OVX group relative to Sham controls,
and administration of LGLP or HGLP effectively reversed this
elevation (Fig. 5A). The
Per2 mRNA of the OVX + HGLP group was downregulated relative
to the OVX group (Fig. 5A).
Cryptochrome circadian regulator 2 (Cry2) mRNA expression in
the OVX group was increased compared with the Sham controls.
However, compared with the OVX group, Cry2 mRNA expression
was decreased in the OVX + HGLP group (Fig. 5A). Nuclear receptor subfamily 1
group D member 1 (Nr1d1) mRNA expression in the OVX group
was higher relative to Sham controls. Compared with the OVX group,
the Nr1d1 mRNA expression was reduced in the OVX + MGLP and
HGLP groups (Fig. 5A). Basic
helix-loop-helix family member e41 (Bhlhe41) transcription
in the OVX + HGLP group was lower than in the OVX group (Fig. 5A). As key regulators of the
circadian rhythm (46), CLOCK
and BMAL1 proteins were measured. Expression of CLOCK was increased
in the OVX + MGLP group and HGLP group compared with the OVX group
(Figs. 5B, C and S4A and B). The BMAL1 protein
expression in the OVX group was decreased relative to Sham
controls, and increased in the OVX + MGLP group and HGLP group
compared with the OVX group (Figs.
5B, C and S4A and B).
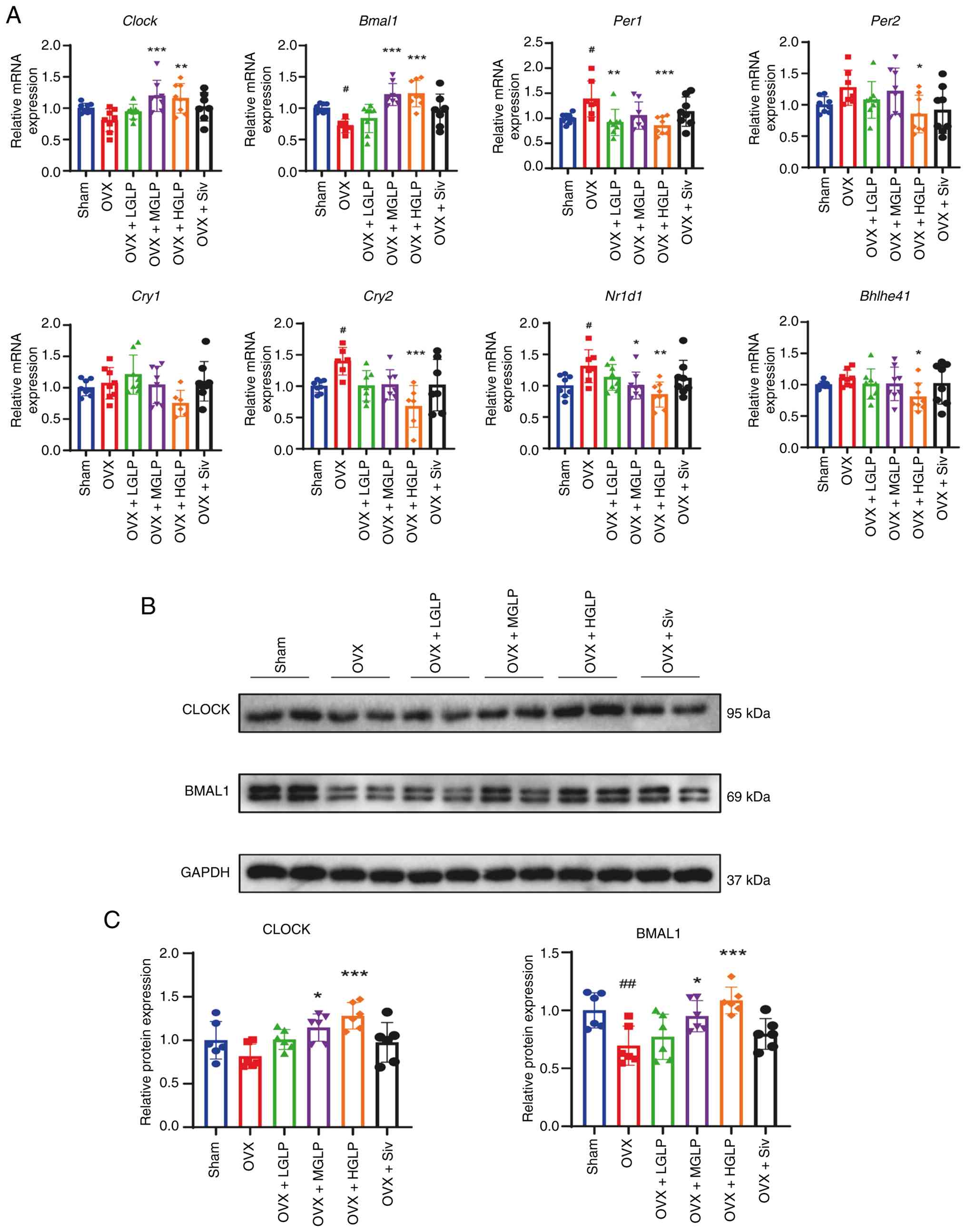 | Figure 5GLP improves the abnormal expression
of genes associated with circadian rhythm. (A) mRNA expression
levels of Clock, Bmal1, Per1, Per2,
Cry1, Cry2, Nr1d1 and Bhlhe41 in the
liver. (B) Protein expression levels of (C) CLOCK and BMAL1 in the
liver. #P<0.05, ##P<0.01 vs. Sham;
*P<0.05, **P<0.01,
***P<0.005 vs. OVX. GLP, Ganoderma lucidum
polysaccharides; OVX, ovariectomy; LGLP, low-dose GLP; MGLP,
medium-dose GLP; HGLP, high-dose GLP; Siv, simvastatin; Bmal1,
brain and muscle arnt-like protein 1; Per1, period circadian
regulator 1; Cry1, cryptochrome circadian regulator 1; Nr1d1,
nuclear receptor subfamily 1 group D member 1; Bhlhe41, basic
helix-loop-helix family member e41. |
GLP improves intestinal microbiota
disorders of OVX mice
To explore the mechanism of GLP in improving the
abnormal metabolism of lipids, the transcription of several genes
associated with the estrogen receptor (ER) pathway was assessed.
Compared with the OVX group, the mRNA expression of estrogen
receptor 1 (Esr1), Esr2), HNF1 homeobox A
(Hnf1a), and carnitine palmitoyltransferase 1A
(Cpt1a) was significantly reduced in the OVX + Siv group.
The mRNA expression of carnitine O-acetyltransferase (Crat)
was significantly reduced in the OVX+ HGLP and Siv groups than the
OVX group. The mRNA expression of family 27, subfamily a,
polypeptide 1 (Cyp27a1) significantly increased in the
OVX+HGLP group than the OVX group. The mRNA expression levels of
the genes fatty acid desaturase 2 (Fads2), acyl-CoA
synthetase long chain family member 5 (Acsl5), and ATP
citrate lyase (Acly) showed no significant changes among the
groups. Compared with the OVX group, the mRNA expression level of
apolipoprotein A-II (Apoa2) was significantly increased in
the OVX+LGLP group. The mRNA expression level of ATP binding
cassette subfamily G member 8 (Abcg8) was significantly
increased in the OVX+MGLP group than the OVX group. The mRNA
expression of glycerol-3-phosphate acyltransferase, mitochondrial
(Gpam) was significantly decreased in the OVX +LGLP, HGLP,
and Siv groups compared with the OVX group (Fig. S5).
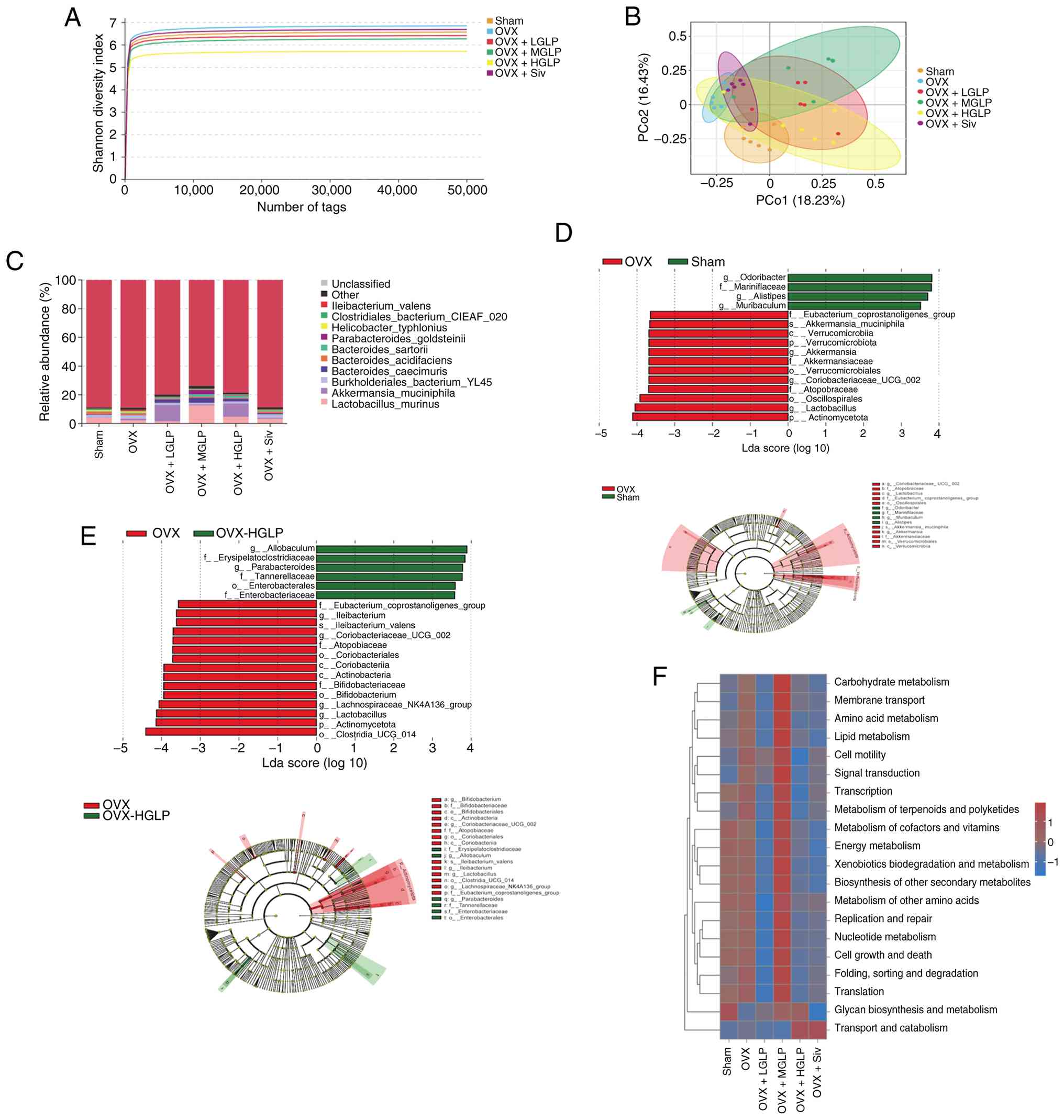
16S rDNA sequencing of the intestinal microbiota was
performed. The Shannon diversity index of the OVX group showed an
increase compared with the Sham group, while the Shannon diversity
index of the OVX + HGLP group was decreased compared with the OVX
group (Fig. 6A). PCoA showed
that the microbiota of the OVX group was distinct from the Sham
group indicating the structure of the microbiota was different
between them, while the intestinal microbiota of the OVX + HGLP
group was closer to that of the Sham group (Fig. 6B). The abundance of
Verrucomicrobiota of the OVX + LGLP group was increased compared
with the OVX group (Fig. S6).
Abundance of Akkermansia muciniphila in the OVX + LGLP group
was increased compared with the OVX group (Fig. 6C). Abundance of Lactobacillus
murinus in the OVX + MGLP group was increased compared with the
OVX group (Fig. 6C). There were
16 different bacterial groups between the OVX group and the Sham
group: 12 bacterial groups were specific to the OVX group,
including Verrucomicrobiota and Akkermansiaceae; the Sham group
included four distinct bacterial groups, including Alistipes
and Muribaculum (Fig.
6D). A total of 21 bacterial groups with differential abundance
were detected between the OVX group and the OVX + HGLP group, and
15 bacterial groups, including Actinomycetota and
lachnospiraceae_nk4a136_group, were identified as biomarkers
enriched in the OVX group, moreover, there were six specific
bacterial groups in the OVX + HGLP group, including
Allobaculum and Erysipelatoclostridiaceae (Fig. 6E). Ovarian resection
significantly enhanced the function of certain pathways compared
with the Sham group, including 'amino acid metabolism', 'membrane
transport', 'carbohydrate metabolism', 'cell motility', 'lipid
metabolism', 'signal transduction' and 'metabolism of terpenoids
and polyketides'. Compared with the OVX group, the functions of
certain metabolic pathways of the OVX + LGLP group, HGLP and Siv
groups were significantly inhibited, including 'carbohydrate
metabolism', 'membrane transport', 'amino acid metabolism', 'cell
motility', 'lipid metabolism', 'signal transduction', 'metabolism
of terpenoids and polyketides', 'transcription', 'metabolism of
cofactors and vitamins', 'energy metabolism', 'xenobiotics
biodegradation and metabolism', 'biosynthesis of other secondary
metabolites', 'cell growth and death', 'metabolism of other amino
acids', 'replication and repair', 'nucleotide metabolism',
'folding, sorting and degradation', 'glycan biosynthesis and
metabolism' and 'translation' (Fig.
6F).
Discussion
The present study treated OVX mice with L-, M- and
HGLP for 8 weeks. GLP decreased the serum TC, TG and LDL-C levels,
expression of genes associated with cholesterol and FA biosynthesis
and lipid accumulation in the hepatic tissues of the OVX mice.
Medium and high doses of GLP upregulate the expression of CLOCK and
BMAL1 mRNA and protein, which are decreased due to ovariectomy.
Finally, GLP influenced the intestinal microbiota disorder in mice
caused by ovariectomy, regulated the intestinal microbial structure
and increased the abundance of beneficial bacteria A.
muciniphila and L. murinus.
With the onset of menopause, patients lack the
protective effect of estrogen, resulting in changes in glucose and
lipid metabolism; therefore, the risk of CVD increases, and by the
age of 65 years, the incidence of CVD in female patients is the
same as that in males (1,2,47).
Moreover, the menopausal transition accelerates the accumulation of
CVD risk factors, such as abdominal adiposity, INS resistance,
arterial hypertension and atherogenic dyslipidemia including
changes in serum LDL-C, TC, TG and apolipoprotein B (48). Two studies concluded that hormone
therapy does not provide heart protection (2,49). Therefore, hormone therapy is not
recommended to prevent primary or secondary cardiovascular disease
and it is necessary to find more safe and effective drugs.
Polysaccharides serve a key role in metabolism by
influencing lipid transport and distribution, regulating endogenous
lipid synthesis and metabolism, inhibiting lipid peroxidation and
affecting intestinal microbiota (12,50,51). GLP, the active component of G.
lucidum, exhibits pharmacological characteristics in
Traditional Chinese Medicine (52). For example, in a mouse model of
diabetes, GLP shows a dose-dependent hypoglycemic effect, improving
fasting blood glucose levels; the underlying mechanism may be
associated with the activation of adenosine monophosphate-activated
protein kinases and key glucose-metabolizing enzymes that regulate
glycogenolysis or gluconeogenesis (53,54). Furthermore, GLP rectifies
HFD-induced intestinal dysbiosis by decreasing the number of
harmful bacteria while concomitantly suppressing pathobionts,
thereby preserving gut-barrier integrity and attenuating metabolic
endotoxemia (55). GLP decreases
serum levels of TG, plasma TC and LDL-C and increase HDL-C levels,
thereby regulating blood lipids and improving metabolic disorder
(55,56). To the best of our knowledge,
however, no studies have reported the role of GLP in regulating
lipid metabolism in patients with declining ovarian function, and
its specific role and mechanism need to be further explored. Here,
GLP effectively decreased the weight gain and body fat content of
the OVX mice. In dyslipidemia caused by ovarian resection, GLP may
serve an ameliorating role, reducing the serum levels of LDL-C, TC,
TG and FFA. HDL-C reverses transport of cholesterol and decreases
excessive peripheral cholesterol accumulation (57). The present study demonstrated
that HGLP could also increase serum HDL-C levels.
The liver, the key organ of material metabolism in
the body, is responsible for regulating the metabolism of
cholesterol and FAs. When intracellular cholesterol levels
decrease, Srebp2 is activated to enter the nucleus and
initiate the expression of downstream genes associated with
cholesterol synthesis to maintain cholesterol homeostasis (58). Hepatic Srebp1-c is a key
transcription factor driving FA synthesis and Pparγ
primarily promotes lipid storage and adipose tissue development;
when these regulators of FA synthesis and storage are inhibited, FA
synthesis and storage are decreased. Pparα mainly regulates
FA oxidation, and its upregulation promotes metabolism of FAs
(59). FASN and ACC are key
enzymes in FA biosynthesis and their action forms the core engine
of de novo lipogenesis, serving as a key hub connecting
nutritional status with cellular lipid homeostasis (60). Hepatic steatosis is interrelated
with insulin resistance; and insulin resistance, liver fibrosis,
and gut-liver axis disorders are all key factors leading to the
progression of MASLD (61-63). Here, GLP inhibited cholesterol
synthesis in OVX mice by downregulating the expression of
Srebp2 and Hmgcr, key genes for cholesterol
synthesis, at the mRNA level. In addition, GLP downregulated the
mRNA expression of FA synthesis genes such as Srebp1-c,
Pparγ, Cd36, Fasn and Acc and
upregulated Pparα expression in the liver of the OVX mice.
GLP regulates insulin signaling pathways and modulates FA
metabolism through multiple mechanisms. Although these
metabolism-related genes are regulated by the estrogen pathway,
their expression levels are influenced by regulation from other
pathways (64-66). However, the mRNA levels of most
estrogen target genes showed no significant change in the liver
tissues among the five groups. It was hypothesized that GLP may not
primarily exert its effects through the estrogen pathway in the
hepatic tissue. Moreover, within the peripheral circadian clock
network, circadian rhythm genes are coupled to lipid, glucose and
cholesterol homeostasis (67).
Circadian rhythm disorder in the liver cause metabolic disorder,
including metabolic disease such as hypertension, obesity and fatty
liver disease (46,68,69), which threaten health. Here,
ovariectomy downregulated the expression of Clock and
Bmal1, while GLP increased the expression of Clock
and Bmal1 in OVX mice, thereby regulating circadian rhythm
disorders.
GLP mitigates dextran sulfate sodium-induced
colitis in rats by restructuring the colonic microbiota, moreover,
GLP consumption regulated 11 genes, including six upregulated [C-C
motif chemokine ligand 5 (Ccl5), CD3 epsilon subunit of
T-cell receptor complex (Cd3e), CD8a molecule (Cd8a),
interleukin 21 receptor (Il21r), lck proto-oncogene, Src
family tyrosine kinase (Lck), and T cell receptor beta
variable (Trbv)] and five downregulated [C-C motif chemokine
ligand 3 (Ccl3), C-X-C motif chemokine ligand 1
(Gro), IL-11, major histocompatibility complex, class II
(Mhc2), and prostaglandin-endoperoxide synthase
(Ptgs)] genes resulting in enhancement of immunity and
reduction of inflammatory response and colonic cancer risk
(70). Ingestion of GLP reverses
gut microbiota and metabolic disorder in rats with type 2 diabetes
(25). The present results
suggested GLP administration can effectively improve the richness
and diversity of microbiota and regulate the composition of
microbial communities following ovarian resection. A.
muciniphila is a prominent member of Verrucomicrobiota; its
depletion is associated with obesity, type 2 diabetes, hepatic
steatosis and associated metabolic disorders (71), and it is a candidate
next-generation probiotic. Levels of A. muciniphila in the
intestine of obese and type 2 diabetic mice decrease, causing an
imbalance of the intestinal microbiota; the abundance of A.
muciniphila in the intestine is restored by supplementation
with prebiotic preparations (72). In addition, therapeutic
supplementation with A. muciniphila reverses HFD-induced
metabolic dysfunction, manifested as excess adiposity, metabolic
endotoxemia, adipose tissue inflammation and INS resistance
(72). Similarly, L.
murinus improves glucose tolerance and INS sensitivity by
enhancing FA oxidation (73).
Here, GLP significantly affected the abundance of beneficial A.
muciniphila and L. murinus bacteria in the intestinal
tract of the OVX mice. Our previous correlation analysis between
the gut microbiota of OVX mice and liver transcriptomes showed that
Akkermansia is positively associated with Cyp7a1, while
Lactobacillus is negatively associated with bile acid transport
proteins [solute carrier family 10 (sodium/bile acid cotransporter
family), member 1 (Ntcp), ATP-binding cassette, sub-family B
member 11 (Bsep)], which also supports that the hypothesis
that the mechanism of GLP may partially involve the gut-liver axis
(74). Therefore, GLP may
improve liver lipid metabolism disorder caused by decreased ovarian
function by increasing the abundance of gut probiotics and
partially acting through the gut-liver axis. Due to the
experimental design, the present study did not explore the causal
association between GLP and the gut-liver axis in depth, and lacked
correlation analysis between specific microbial populations and key
metabolic parameters, as well as functional metagenomics and fecal
microbiota transplantation experiments.
In summary, GLP influences the structure of the gut
microbiota and increases the abundance of probiotics (Fig. 7), thereby improving liver glucose
and lipid metabolic disorder caused by decreased or absent ovarian
function. The present study demonstrated that GLP and Siv have
similar effects in improving liver lipid metabolism in OVX mice,
indicating that GLP has certain potential for the prevention and
treatment of lipid metabolic disorder in menopausal patients.
However, GLP offers distinct clinical advantages over Siv. Siv
primarily inhibits 3-hydroxy-3-methylglutaryl-Coenzyme A reductase
(75). Compared with Siv, GLP is
more effective at increasing the proportion of probiotics, such as
A. muciniphila. Since GLP is not easily absorbed by the gut,
it acts directly on the intestinal microbiota, enhancing the
structure and function of the microbiome, which may regulate liver
glucose and lipid metabolism through the gut-liver axis. As a
natural plant polysaccharide, GLP may have a superior safety
profile and tolerability compared with statins, which are
associated with myopathy, hepatotoxicity and new-onset diabetes
(76-78). This renders GLP suitable for
long-term preventive use in postmenopausal patients with
dyslipidemia and metabolic comorbidities. Furthermore, the distinct
mechanisms suggest potential for combination therapy, where GLP
could enhance statin efficacy while mitigating adverse effects. As
a natural product, GLP may also offer cost-effectiveness
advantages. Collectively, GLP matches Siv in lipid-lowering
efficacy while providing added value through its
microbiota-centered benefits and favorable safety profile,
positioning it as a candidate for managing postmenopausal metabolic
disorder. In addition, GLP can also affect the expression of
hepatic circadian rhythm genes, thereby influencing glucose and
lipid metabolism in the liver by modulating the expression of genes
associated with these metabolic processes. Given the high incidence
of cardiovascular disease in elderly patients, GLP has potential
efficacy in treating metabolic disorder caused by decreased ovarian
function.
Supplementary Data
Availability of data and materials
The data generated in the present study may be
found in the Sequence read archive under accession no. PRJNA1279352
or at the following URL: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1279352.
Authors' contributions
XJ conceived and designed the study and analyzed
data. XL analyzed data, constructed figures and wrote the
manuscript. YJW designed the experiments. ZLL and YHY conceived the
study, performed the experiments and edited the manuscript. XJ and
XL confirm the authenticity of all the raw dat. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
All the animal experiments were approved by the
Experimental Animal Ethics Committee of Guangdong Pharmaceutical
University (Guangzhou, China; approval no. gdpulac2020065).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 82171855) and the Key Field Special
Project for Colleges and Universities of Guangdong Province
(Biomedicine and Health; grant no. 2023ZDZX2030).
References
|
1
|
Gatenby C and Simpson P: Menopause:
Physiology, definitions, and symptoms. Best Pract Res Clin
Endocrinol Metab. 38:1018552024. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gersh FL, O'Keefe JH and Lavie CJ:
Postmenopausal hormone therapy for cardiovascular health: The
evolving data. Heart. 107:1115–1122. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li H, Sun R, Chen Q, Guo Q, Wang J, Lu L
and Zhang Y: Association between HDL-C levels and menopause: A
meta-analysis. Hormones (Athens). 20:49–59. 2021. View Article : Google Scholar :
|
|
4
|
Chen Y, Wang A, Zhang X, Xia F and Zhao X:
Effect of age at menopause and menopause itself on high sensitivity
C-reactive protein, pulse wave velocity, and carotid intima-media
thickness in a Chinese population. Medicine (Baltimore).
102:e356292023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Witteman JC, Grobbee DE, Kok FJ, Hofman A
and Valkenburg HA: Increased risk of atherosclerosis in women after
the menopause. BMJ. 298:642–644. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zaydun G, Tomiyama H, Hashimoto H, Arai T,
Koji Y, Yambe M, Motobe K, Hori S and Yamashina A: Menopause is an
independent factor augmenting the age-related increase in arterial
stiffness in the early postmenopausal phase. Atherosclerosis.
184:137–142. 2006. View Article : Google Scholar
|
|
7
|
Colditz GA, Willett WC, Stampfer MJ,
Rosner B, Speizer FE and Hennekens CH: Menopause and the risk of
coronary heart disease in women. N Engl J Med. 316:11051987.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Talaulikar V: Menopause transition:
Physiology and symptoms. Best Pract Res Clin Obstet Gynaecol.
81:3–7. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee E, Anselmo M, Tahsin CT, Vanden Noven
M, Stokes W, Carter JR and Keller-Ross ML: Vasomotor symptoms of
menopause, autonomic dysfunction, and cardiovascular disease. Am J
Physiol Heart Circ Physiol. 323:H1270–H1280. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y and Li C: Hormone therapy and
biological aging in postmenopausal women. JAMA Netw Open.
7:e24308392024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Innes KE, Selfe TK and Vishnu A: Mind-body
therapies for menopausal symptoms: A systematic review. Maturitas.
66:135–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu Y, Shen M, Song Q and Xie J: Biological
activities and pharmaceutical applications of polysaccharide from
natural resources: A review. Carbohydr. Polym. 183:91–101. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cao Y, Xu X, Liu S, Huang L and Gu J:
Ganoderma: A cancer immunotherapy review. Front Pharmacol.
9:12172018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Liu Y, Tang Q, Zhou S, Feng J and
Chen H: Polysaccharide of Ganoderma and its bioactivities. Adv Exp
Med Biol. 1181:107–134. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou Y, Wang M, Wang Z, Qiu J, Wang Y, Li
J, Dong F, Huang X, Zhao J and Xu T: Polysaccharides from hawthorn
fruit alleviate high-fat diet-induced NAFLD in mice by improving
gut microbiota dysbiosis and hepatic metabolic disorder.
Phytomedicine. 139:1564582025. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shiao MS: Natural products of the
medicinal fungus Ganoderma lucidum: occurrence, biological
activities, and pharmacological functions. Chem Rec. 3:172–180.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sohretoglu D and Huang S: Ganoderma
lucidum polysaccharides as an anti-cancer agent. Anticancer Agents
Med Chem. 18:667–674. 2018. View Article : Google Scholar
|
|
18
|
Seweryn E, Ziała A and Gamian A:
Health-promoting of polysaccharides extracted from Ganoderma
lucidum. Nutrients. 13:27252021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li J, Niu D, Zhang Y and Zeng XA:
Physicochemical properties, antioxidant and antiproliferative
activities of polysaccharides from Morinda citrifolia L. (Noni)
based on different extraction methods. Int J Biol Macromol.
150:114–121. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zeng P, Guo Z, Zeng X, Hao C, Zhang Y,
Zhang M, Liu Y, Li H, Li J and Zhang L: Chemical, biochemical,
preclinical and clinical studies of Ganoderma lucidum
polysaccharide as an approved drug for treating myopathy and other
diseases in China. J Cell Mol Med. 22:3278–3297. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Tang J, Gao H, Xu Y, Han Y, Shang H,
Lu Y and Qin C: Ganoderma lucidum triterpenoids and polysaccharides
attenuate atherosclerotic plaque in high-fat diet rabbits. Nutr
Metab Cardiovasc Dis. 31:1929–1938. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vitak T, Yurkiv B, Wasser S, Nevo E and
Sybirna N: Effect of medicinal mushrooms on blood cells under
conditions of diabetes mellitus. World J Diabetes. 8:187–201. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng Y, Bai L, Zhou Y, Tong R, Zeng M, Li
X and Shi J: Polysaccharides from Chinese herbal medicine for
anti-diabetes recent advances. Int J Biol Macromol. 121:1240–1253.
2019. View Article : Google Scholar
|
|
24
|
Sang T, Guo C, Guo D, Wu J, Wang Y, Wang
Y, Chen J, Chen C, Wu K, Na K, et al: Suppression of obesity and
inflammation by polysaccharide from sporoderm-broken spore of
Ganoderma lucidum via gut microbiota regulation. Carbohydr Polym.
256:1175942021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen M, Xiao D, Liu W, Song Y, Zou B, Li
L, Li P, Cai Y, Liu D, Liao Q and Xie Z: Intake of Ganoderma
lucidum polysaccharides reverses the disturbed gut microbiota and
metabolism in type 2 diabetic rats. Int J Biol Macromol.
155:890–902. 2020. View Article : Google Scholar
|
|
26
|
Baker JM, Al-Nakkash L and
Herbst-Kralovetz MM: Estrogen-gut microbiome axis: Physiological
and clinical implications. Maturitas. 103:45–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang M, Wen S, Zhang J, Peng J, Shen X and
Xu L: Systematic review and meta-analysis: Changes of gut
microbiota before and after menopause. Dis Markers.
2022:37673732022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu Y, Zhou Y, Mao T, Huang Y, Liang J,
Zhu M, Yao P, Zong Y, Lang J and Zhang Y: The relationship between
menopausal syndrome and gut microbes. BMC Womens Health.
22:4372022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pavlovska OM, Pavlovska KM, Heryak SM,
Khmil SV and Khmil MS: Vasomotor menopausal disorders as a possible
result of dysfunction of the microbiota-intestine-brain axis. J Med
Life. 15:234–240. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu S, Ding Q, Zhang W, Kang M, Ma J and
Zhao L: Gut microbial beta-glucuronidase: A vital regulator in
female estrogen metabolism. Gut Microbes. 15:22367492023.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Trefts E, Gannon M and Wasserman DH: The
liver. Curr Biol. 27:R1147–R1151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo X, Okpara ES, Hu W, Yan C, Wang Y,
Liang Q, Chiang JYL and Han S: Interactive relationships between
intestinal flora and bile acids. Int J Mol Sci. 23:83432022.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu S and Yang X: Intestinal flora plays a
role in the progression of hepatitis-cirrhosis-liver cancer. Front
Cell Infect Microbiol. 13:11401262023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lilley E, Stanford SC, Kendall DE,
Alexander SPH, Cirino G, Docherty JR, George CH, Insel PA, Izzo AA,
Ji Y, et al: ARRIVE 2.0 and the British journal of pharmacology:
Updated guidance for 2020. Br J Pharmacol. 177:3611–3616. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
36
|
Edgar RC: Search and clustering orders of
magnitude faster than BLAST. Bioinformatics. 26:2460–2461. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Edgar RC, Haas BJ, Clemente JC, Quince C
and Knight R: UCHIME improves sensitivity and speed of chimera
detection. Bioinformatics. 27:2194–2200. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shannon CE: A mathematical theory of
communication. Bell Syst Tech J. 27:379–423. 623–656. 1948.
View Article : Google Scholar
|
|
39
|
Gower JC: Some distance properties of
latent root and vector methods used in multivariate analysis.
Biometrika. 53:325–338. 1966. View Article : Google Scholar
|
|
40
|
Bray JR and Curtis JT: An ordination of
the upland forest communities of southern wisconsin. Ecol Monogr.
27:325–349. 1957. View Article : Google Scholar
|
|
41
|
Douglas GM, Maffei VJ, Zaneveld JR, Yurgel
SN, Brown JR, Taylor CM, Huttenhower C and Langille MGI: PICRUSt2
for prediction of metagenome functions. Nat Biotechnol. 38:685–688.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar
|
|
43
|
Kanehisa M, Goto S, Sato Y, Kawashima M,
Furumichi M and Tanabe M: Data, information, knowledge and
principle: Back to metabolism in KEGG. Nucleic Acids Res.
42(Database Issue): D199–D205. 2014. View Article : Google Scholar :
|
|
44
|
Parks DH, Tyson GW, Hugenholtz P and Beiko
RG: STAMP: Statistical analysis of taxonomic and functional
profiles. Bioinformatics. 30:3123–3124. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liang XL, Liang YW, Tian J, Mo FF, Pan TM,
Chen YF, Shao XQ and Li KP: Structural characterization and
oxidative stress modulation activity of an acidic
Heteropolysaccharide from Microctis folium. Carbohydr Polym.
369:1242612025. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Reppert SM and Weaver DR: Coordination of
circadian timing in mammals. Nature. 418:935–941. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Otto CM: Heartbeat: Is postmenopausal
hormone therapy a risk factor or preventative therapy for
cardiovascular disease in women? Heart. 107:1103–1105. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Anagnostis P and Stevenson JC:
Cardiovascular health and the menopause, metabolic health. Best
Pract Res Clin Endocrinol Metab. 38:1017812024. View Article : Google Scholar
|
|
49
|
Rossouw JE, Anderson GL, Prentice RL,
LaCroix AZ, Kooperberg C, Stefanick ML, Jackson RD, Beresford SA,
Howard BV, Johnson KC, et al: Risks and benefits of estrogen plus
progestin in healthy postmenopausal women: Principal results from
the women's health initiative randomized controlled trial. JAMA.
288:321–333. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nai J, Zhang C, Shao H, Li B, Li H, Gao L,
Dai M, Zhu L and Sheng H: Extraction, structure, pharmacological
activities and drug carrier applications of Angelica sinensis
polysaccharide. Int J Biol Macromol. 183:2337–2353. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Fang S, Wang T, Li Y, Xue H, Zou J, Cai J,
Shi R, Wu J and Ma Y: Gardenia jasminoides Ellis polysaccharide
ameliorates cholestatic liver injury by alleviating gut microbiota
dysbiosis and inhibiting the TLR4/NF-κB signaling pathway. Int J
Biol Macromol. 205:23–36. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ahmad R, Riaz M, Khan A, Aljamea A,
Algheryafi M, Sewaket D and Alqathama A: Ganoderma lucidum (Reishi)
an edible mushroom; a comprehensive and critical review of its
nutritional, cosmeceutical, mycochemical, pharmacological,
clinical, and toxicological properties. Phytother Res. 35:60302021.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Xiao C, Wu QP, Cai W, Tan JB, Yang XB and
Zhang JM: Hypoglycemic effects of Ganoderma lucidum polysaccharides
in type 2 diabetic mice. Arch Pharm Res. 35:1793–1801. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xiao C, Wu Q, Zhang J, Xie Y, Cai W and
Tan J: Antidiabetic activity of Ganoderma lucidum polysaccharides
F31 down-regulated hepatic glucose regulatory enzymes in diabetic
mice. J Ethnopharmacol. 196:47–57. 2017. View Article : Google Scholar
|
|
55
|
Chang CJ, Lin CS, Lu CC, Martel J, Ko YF,
Ojcius DM, Tseng SF, Wu TR, Chen YY, Young JD and Lai HC: Ganoderma
lucidum reduces obesity in mice by modulating the composition of
the gut microbiota. Nat Commun. 6:74892015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhu K, Nie S, Li C, Lin S, Xing M, Li W,
Gong D and Xie M: A newly identified polysaccharide from Ganoderma
atrum attenuates hyperglycemia and hyperlipidemia. Int J Biol
Macromol. 57:142–150. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jung E, Kong SY, Ro YS, Ryu HH and Shin
SD: Serum cholesterol levels and risk of cardiovascular death: A
systematic review and a dose-response meta-analysis of prospective
cohort studies. Int J Environ Res Public Health. 19:82722022.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Xiao MY, Pei WJ, Li S, Li FF, Xie P, Luo
HT, Hyun Yoo H and Piao XL: Gypenoside L inhibits hepatocellular
carcinoma by targeting the SREBP2-HMGCS1 axis and enhancing immune
response. Bioorg Chem. 150:1075392024. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhao R, Chen Q and He YM: The effect of
Ganoderma lucidum extract on immunological function and identify
its anti-tumor immunostimulatory activity based on the biological
network. Sci Rep. 8:126802018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu W, Qiu J, Chen Y, Che X and Li X:
Chrysanthemum morifolium extract improves metabolic
dysfunction-associated fatty liver disease by regulating lipid
metabolism. Sci Rep. 15:400692025. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Romanos M, Garcia Cordova JM, Villamarin
J, Pazmino Zurita JD and Acosta A: Obesity and metabolic
dysfunction-associated steatotic liver disease (MASLD): A
literature review on pathophysiology and treatment. Diabetes Obes
Metab. 28(Suppl2): S19–S30. 2026. View Article : Google Scholar
|
|
62
|
Cuthbertson DJ, Whyte M, Henney AE, Alam
U, Goff L, Fielding BA and Umpleby AM: Differential
pathophysiological drivers of susceptibility to type 2 diabetes and
metabolic dysfunction-associated steatotic liver disease: Ethnic
differences in insulin dynamics, whole-body fat metabolism, and
organ-specific lipid deposition. Obes Rev. 2026:e701042026.
View Article : Google Scholar
|
|
63
|
Servin-Uribe RI, Pérez-Jiménez J, Castaño
Tostado E, Hernández-Saavedra D and Reynoso Camacho R: Hepatic
lipids as molecular targets in obesity-associated insulin
resistance in rodent models: A systematic review and meta-analysis.
Lipids. Apr 28–2026.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Fuentes N and Silveyra P: Estrogen
receptor signaling mechanisms. Adv Protein Chem Struct Biol.
116:135–170. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
MacLeod B, Wang C, Brown LH, Borkowski E,
Nakamura MT, Wells KR, Brunt KR, Harasim-Symbor E, Chabowski A and
Mutch DM: Fads2 knockout mice reveal that ALA prevention of hepatic
steatosis is dependent on delta-6 desaturase activity. J Lipid Res.
65:1006422024. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li Z, Zheng D, Zhang T, Ruan S, Li N, Yu
Y, Peng Y and Wang D: The roles of nuclear receptors in cholesterol
metabolism and reverse cholesterol transport in nonalcoholic fatty
liver disease. Hepatol Commun. 8:e03432023.PubMed/NCBI
|
|
67
|
Tseng HL, Yang SC, Yang SH and Shieh KR:
Hepatic circadian-clock system altered by insulin resistance,
diabetes and insulin sensitizer in mice. PLoS One. 10:e01203802015.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Eckel-Mahan K and Sassone-Corsi P:
Metabolism control by the circadian clock and vice versa. Nat
Struct Mol Biol. 16:462–467. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Gachon F, Bugianesi E, Castelnuovo G,
Oster H, Pendergast JS and Montagnese S: Potential bidirectional
communication between the liver and the central circadian clock in
MASLD. NPJ Metab Health Dis. 3:152025. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xie J, Liu Y, Chen B, Zhang G, Ou S, Luo J
and Peng X: Ganoderma lucidum polysaccharide improves rat
DSS-induced colitis by altering cecal microbiota and gene
expression of colonic epithelial cells. Food Nutr Res. 63:
View Article : Google Scholar : 2019.
|
|
71
|
Cani PD, Depommier C, Derrien M, Everard A
and de Vos WM: Akkermansia muciniphila: Paradigm for
next-generation beneficial microorganisms. Nat Rev Gastroenterol
Hepatol. 19:625–637. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Everard A, Belzer C, Geurts L, Ouwerkerk
JP, Druart C, Bindels LB, Guiot Y, Derrien M, Muccioli GG, Delzenne
NM, et al: Cross-talk between Akkermansia muciniphila and
intestinal epithelium controls diet-induced obesity. Proc Natl Acad
Sci USA. 110:9066–9071. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Long J, Shi Z, Miao Z, Dong L and Yan D:
Lactobacillus murinus alleviates insulin resistance via promoting
L-citrulline synthesis. J Endocrinol Invest. 48:1005–1015. 2025.
View Article : Google Scholar
|
|
74
|
Jin X, Liu X, Wang Y, Li X, Zhang T, Li J,
Lei Z and Yang Y: The mechanism by which melatonin improves the
dysregulation of glucose and lipid metabolism in castrated female
mice. J Pineal Res. 77:e700822025. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lennernäs H and Fager G: Pharmacodynamics
and pharmacokinetics of the HMG-CoA reductase inhibitors.
Similarities and differences. Clin Pharmacokinet. 32:403–425. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ramsey LB, Johnson SG, Caudle KE, Haidar
CE, Voora D, Wilke RA, Maxwell WD, McLeod HL, Krauss RM, Roden DM,
et al: The clinical pharmacogenetics implementation consortium
guideline for SLCO1B1 and simvastatin-induced myopathy: 2014
update. Clin Pharmacol Ther. 96:423–428. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhou L, Wu B, Bian Y, Lu Y, Zou Y, Lin S,
Li Q and Liu C: Hepatotoxicity associated with statins: A
retrospective pharmacovigilance study based on the FAERS database.
PLoS One. 20:e03275002025. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wang X, Li J, Wang T, Zhang Z, Li Q, Ma D,
Chen Z, Ju J, Xu H and Chen K: Associations between statins and
adverse events in secondary prevention of cardiovascular disease:
Pairwise, network, and dose-response meta-analyses of 47 randomized
controlled trials. Front Cardiovasc Med. 9:9290202022. View Article : Google Scholar : PubMed/NCBI
|