According to epidemiological statistics from the
World Health Organization, primary liver cancer presently claims
the lives of ~700,000 individuals each year (1,2).
Hepatocellular carcinoma (HCC) is the most common type of primary
liver cancer, accounting for 75 to 90% of all cases. With a 5-year
overall survival (OS) rate of ~18%, its prognosis is often dismal
(3,4). Clinical data has shown that only
5-15% of patients with HCC meet the indications for surgical
resection at the time of diagnosis, and most of them are early
confirmed cases. In addition, <1/3 of patients benefit from
current conventional therapies due to characteristics such as tumor
heterogeneity and reduced liver regenerating ability (5). As a result, identifying more
effective and precise therapeutic strategies has become a central
priority in both scientific and clinical research.
Two major discoveries at the intersection of tumor
metabolism and epigenetics have generated novel opportunities to
elucidate the molecular processes behind HCC: Initially, Otto
Warburg's early 20th-century identification of the Warburg effect
revealed that tumor cells excessively absorb glucose, even in the
presence of oxygen, preferentially converting it into lactate
through glycolysis, thereby supplying the necessary materials and
energy for accelerated tumor growth (6); subsequently, Zhang et al's
(7) pioneering 2019 study first
demonstrated that lactate can function as an endogenous precursor
for the lysine lactylation (Kla) modification of histones.
Previous studies have established that lactylation
is crucial in liver cancer progression by modulating essential
biological processes, including liver cancer cell proliferation,
augmenting tumor invasion and metastatic potential, and
facilitating immune suppression within the tumor microenvironment
(TME) (8-10). Nonetheless, current research
demonstrates considerable limitations: On the one hand, research on
the mechanisms through which lactylation influences HCC regulation
is disjointed, lacking systematic integration from the molecular to
the microenvironmental level, hence complicating the establishment
of a full knowledge of the regulatory network. Conversely, clinical
translational research regarding lactate modification as a
diagnostic and prognostic biomarker for HCC, as well as the
formulation of focused therapy regimens, is yet nascent, with its
potential clinical uses not yet thoroughly clarified (9,11). The present review aimed at
systematically clarifying the molecular basis of lactylation, its
specific mechanisms in the development of HCC, and its potential
applications in the clinical diagnosis and treatment of liver
cancer, serving as a reference for future research and clinical
translation.
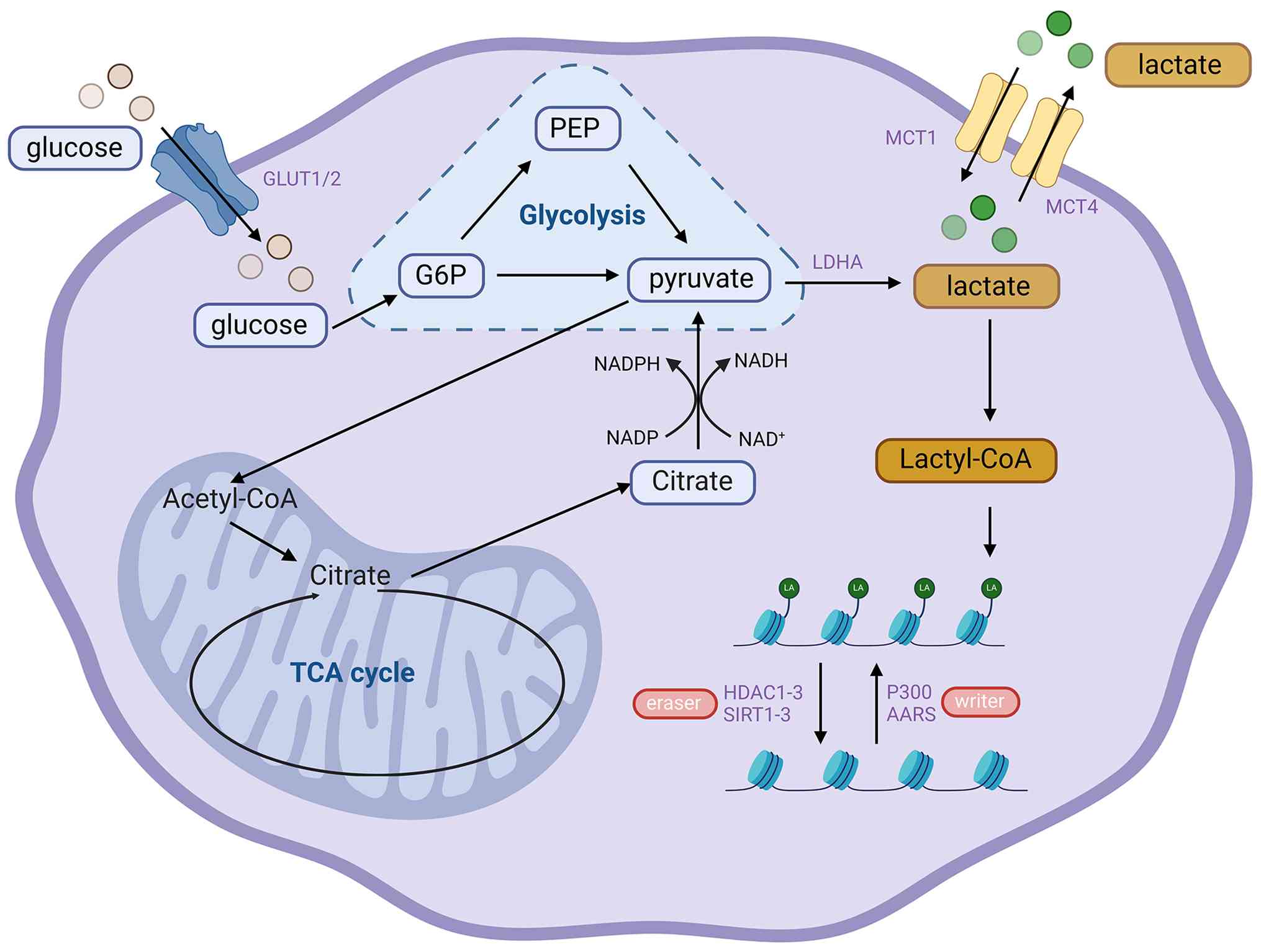
In physiological homeostasis, the liver functions as
the principal organ for lactate clearance in the human body,
sustaining lactate balance through the glucose-pyruvate-lactate
metabolic cycle. In the development of HCC, liver cancer cells are
often accompanied by the 'Warburg effect'. This metabolic pattern
will significantly increase the production of lactic acid and
reduce the clearance ability, leading to a large accumulation of
lactic acid within the hepatic TME (9). The buildup of lactate furnishes the
necessary substrate for the alteration of proteins by lactate,
hence substantially enhancing the likelihood of these modifications
taking place.
Lactylation is a recently identified important
post-translational modification (PTM). In the context of tissue
dynamic metabolic balance, some lactate generated by cells is
involved in energy metabolism, while another fraction aids in
epigenetic changes and non-histone lactylation processes (12). Specifically, lactylation is a
chemical process induced by increased lactate concentrations in the
TME, during which lactate groups are preferentially appended to
lysine residues on proteins. It has been identified as a
fundamental method for HCC cells to acclimatize to metabolic stress
and modulate the tumor immunological microenvironment (13).
In the process of enzyme-dependent protein acylation
modification, there is usually a classic regulatory mode of
'write-recognition-erase': The 'writers' are responsible for
transferring the acyl group in acyl-CoA to the side chain of amino
acid residues such as lysine, glycine, cysteine or serine in the
protein; the 'readers' recognize and bind to the modified amino
acid residues through specific structural domains, and initiates
the downstream functional effect; the 'erasers' can remove the acyl
group on the amino acid residues, so as to realize the dynamic
reversible regulation of the modification process (14).
Since both lactic acid and alanine are three-carbon
compounds with carboxylic acid groups, their molecular structures
are highly similar. The core difference lies only in that alanine
contains an amino group (-NH2), while lactic acid
contains a hydroxyl group (-OH). Consequently, it may be deduced
that lactylation adheres to the previously indicated
'write-recognize-erase' regulatory framework (Fig. 1). The alteration process is
predominantly governed by four principal variables, as indicated by
current study advancements (Table
I).
Lactate transferases function as the principal
enzymes that initiate enzymatic lactylation. The presently
recognized members predominantly belong to the histone
acetyltransferase (HAT) family and the aminoacyl-tRNA synthetase
(AALS) family. P300, as a member of the traditional HAT family, was
the first identified lactoyl-transferase capable of dynamically
modulating histone lactylation levels. Experimental results
indicated that the inhibition of p300 function through gene
knockdown or pharmacological inhibitors markedly decreases
intracellular histone lactylation levels (7,15,16). Subsequent investigations showed
that the inhibition of p300/CREB-binding protein (p300/CBP)
acetyltransferase activity or the suppression of p300/CBP gene
expression in macrophages significantly reduced the lactate-induced
lactylation of the high mobility group box 1, indicating that
p300/CBP also plays a role in non-histone lactylation (17). Of note, the evidence supporting
p300 as a lactylation writer is still largely based on correlative
and mechanistic studies, and its substrate selectivity in HCC
requires a deeper validation. AALS1/2, functioning as ATP-dependent
lactate transferases, specifically recognize and bind lactate
through designated domains. Utilizing ATP as an energy donor, they
transform lactate into the high-energy intermediate 'lactyl-CoA',
subsequently facilitating the lactylation modification of lysine
residues in substrate proteins (18-21). HAT binding to ORC1, a member of
the MOZ, Ybf2/Sas3, Sas2, Tip60 HAT family, operates as a
lactylating transferase. It aggregates at transcription start sites
and facilitates carcinogenesis by modulating histone H3 lysine 9
lactylation and oncogene expression (22).
Deacetylases are essential components that sustain
the dynamic balance of histone acetylation modifications. Previous
studies have preliminarily identified a variety of enzymes with
delactylase activity, mainly including members of the histone
deacetylase (HDAC) family and members of the silencing information
regulator (SIRT) family (23,24). Among them, studies have proved
that HDAC1-3 are able to remove the lactylation of histone H3
lysine 18 (H3K18) (24,25), among which HDAC3 has the
strongest delactylase activity (26). SIRT2 and SIRT3 are also
delactylases, but their activity is markedly lower than that of
HDAC3. Zessin et al (27)
found that the delactylase activity of SIRT2 is several order of
magnitude lower than that of HDAC3. However, in general, the
identification of delactylases is still incomplete, and their
specific role and regulatory network in liver cancer have not been
fully clarified. This remains a key area for future research.
The upstream metabolic regulation maintains the
balance between lactate production and clearance, hence ensuring a
consistent substrate supply for lactylation and acting as a vital
upstream regulator. This process is predominantly regulated by two
fundamental metabolic pathways: As tumor cells expand fast, the
requirement for cellular energy escalates. At this time, lactate
dehydrogenase (LDH) becomes a key regulatory enzyme of metabolic
flux.
Under the catalysis of LDH, pyruvate is reduced to
lactate, providing a core substrate for lactylation (28-30). In contrast to the role of LDH,
pyruvate dehydrogenase (PDH) can catalyze the oxidation and
decarboxylation of pyruvate to generate acetyl-CoA, which enters
the tricarboxylic acid (TCA) cycle for complete oxidation and
energy supply. This procedure diverts pyruvate from the lactate
generation pathway, facilitating indirect lactate clearance. The
PDH-catalyzed process is irreversibly functioning as a critical
'shunt valve' that preserves intracellular lactate homeostasis
(31).
In addition to LDH and PDH, the polyol pathway
constitutes a metabolic branch that indirectly modulates the
intracellular lactate pool available for lactylation. Under the
hyper-glycolytic conditions in HCC, aldose reductase (AR) competes
directly with the glycolytic pathway for the glucose (32). This competitive metabolic
shunting effectively diverts glucose flux, thereby limiting
pyruvate availability and the subsequent generation of lactate
(32,33). Through this mechanism, elevated
AR activity functions as a metabolic gate that attenuates
intracellular lactate accumulation and constrains the substrate
supply for downstream Kla. Concurrently, the consumption of NADPH
by AR may further perturb the redox environment required for the
optimal function of lactylation regulatory enzymes (34). Therefore, the polyol pathway
represents an indirect upstream modulator that governs lactylation
substrate availability in HCC.
Readers bind lactylated lysine residues and mediate
downstream transcriptional regulation. Recent studies have
identified several candidate readers. Brahma-related gene 1 (BRG1),
a core subunit of the SWItch/sucrose non-fermentable
chromatin-remodeling complex, recognizes histone H3 lysine 18
lactylation (H3K18la) and promotes transcriptional activation
through chromatin remodeling (35). Double PHD fingers 2
preferentially binds H3K14la and facilitates tumor-associated gene
expression, linking lactylation to oncogenic transcriptional
programs (36). In addition, the
bromodomain-containing protein tripartite motif-containing 33 can
directly recognize lactylated lysine residues, suggesting that
canonical acetyl-lysine reader domains may also accommodate
lactylation (37). However, the
number of identified lactylation readers remains limited, and their
functional roles in HCC require further investigation.
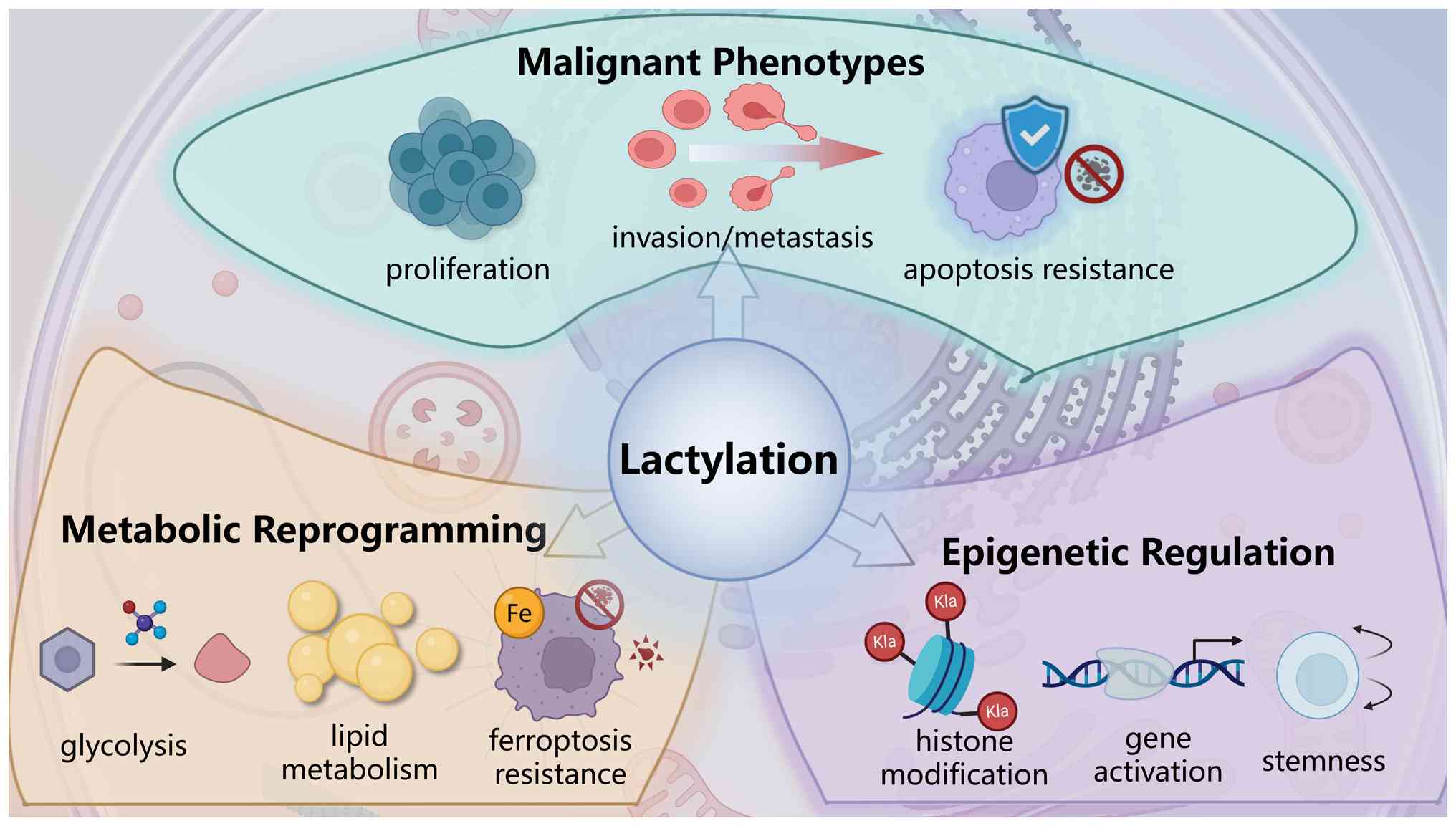
Lactylation plays a crucial role in the development,
progression and treatment resistance of HCC. This chapter focuses
on how lactylation drives the malignant phenotype of HCC through
multidimensional pathways and explains its underlying molecular
mechanisms in detail (Fig.
2).
Lactylation is essential for maintaining the
equilibrium between proliferation and death in HCC cells. Research
has shown that Kla of cyclin E2 (CCNE2) accelerates cell cycle
progression and promotes HCC cell proliferation. This process is
governed by SIRT3. Honokiol (HKL), acting as an activator of SIRT3,
enhances the delactylase activity of SIRT3 toward CCNE2, which
induces apoptosis in HCC cells and suppresses tumor growth in
vivo (50). Lactylation at
the K124 location of centromeric protein A similarly enhances its
transcriptional regulatory function. This modification synergizes
with transcription factor Yin Yang 1 to promote mitosis and
proliferation in liver cancer cells (71). In addition, certain natural
substances can demonstrate anticancer properties by inhibiting
lactylation. Demethyl zelasterol and royal jelly acid can reduce
the level of histone lactylation, thus inhibit the proliferation
and migration of liver cancer cells (13,72).
HCC cells depend on effective glycolysis to satisfy
their accelerated energy requirements for proliferation (76). Lactylation forms a positive
feedback loop in this process, promoting HCC initiation and
progression. On the one hand, lactate produced by glycolysis
provides ample substrate for lactylation. On the other hand,
lactylation, particularly histone lactylation, activates gene
expression of multiple key glycolytic enzymes, including hexokinase
2 (77,78), further enhancing lactate
production and sustaining high glycolysis.
Meanwhile, lipid metabolism in HCC cells is also
significantly reprogrammed to support cell membrane synthesis and
energy storage (79). It has
been found that in non-alcoholic fatty liver disease, the
lactylation of fatty acid synthase at the K673 site inhibits its
enzymatic activity, thereby mediating the downregulation of hepatic
lipid accumulation by mitochondrial pyruvate carrier (MPC) 1 and
reducing lipid accumulation (80). However, in certain cases,
lactylation can also promote lipid synthesis. Meng et al
(81) demonstrated that lactate
dehydrogenase A (LDHA)-induced mutations in the H3K18lac gene can
increase total cholesterol and triglyceride levels, as well as the
expression of fatty acid synthesis-related proteins stearoyl-CoA
desaturase 1, fatty acid synthase and sterol regulatory
element-binding protein 1, providing material and energy reserves
for the rapid growth of liver cancer cells. However, the
conclusions are mainly based on specific cell models, and their
universality in the complex metabolic environment of the human body
still needs to be verified.
Furthermore, lactylation is intricately linked to
resistance against ferroptosis. Ferroptosis depends on iron
molecules, which is a type of programmed cell death characterized
by lipid peroxidation (82). An
increasing number of studies have shown that ferroptosis can
closely influence the proliferation, metastasis and drug resistance
of HCC. It has been demonstrated that following sublethal thermal
stress, such as microwave ablation, lactate levels and histone
lactylation (particularly H3K18la) are significantly upregulated in
HCC cells. Histone lactylation promotes the expression of cysteine
desulfurase NFS1, thus enhancing the synthesis of iron-sulphur
clusters. Through this step, the resistance of HCC cells to iron
death is improved, which ultimately promotes their survival and
metastasis (75). This
elucidates a novel mechanism through which lactylation
modifications impart therapeutic resistance to HCC cells by
modulating the ferroptosis pathway.
In liver cancer cells, enhanced glycolysis leads to
a large accumulation of lactates, which acts as a substrate to
drive lactylation modification of histones, thereby triggering
changes in gene transcription programs related to tumor malignancy.
Numerous histone lactylation sites have been shown to be
intricately associated with malignant progression in HCC. H3K18la
is among the most thoroughly investigated lactylated modifications.
It primarily accumulates in the promoter regions of active genes
and is considered an activating epigenetic marker (7). Research demonstrates that O-linked
β-N-acetyl-glucosamin glycosylation of Y-box binding protein 1
(YBX1), a crucial regulator of glycolysis, stabilizes pyruvate
kinase M2 mRNA and increases glycolytic activity, consequently
leading to increased H3K18la levels. H3K18la enhances YBX1
expression, creating a positive feedback loop that stimulates
metabolic activation and malignant advancement in HCC (83). Furthermore, an elevated histone
H3 lysine 56 lactylation (H3K56la) has been demonstrated to be
closely associated with the tumorigenicity and stemness maintenance
of liver cancer stem cells (LCSCs) (84). In LCSCs, H3K56la levels are
significantly upregulated, and its enrichment directly activates
the transcription of multiple stemness-related genes, including the
oncogene MYC and the pro-angiogenic factor vascular endothelial
growth factor A (VEGFA), promoting the self-renewal and
tumorigenicity of LCSCs (85).
Targeting lactate production or interfering with lactylation can
effectively reduce H3K56la levels, thereby inhibiting the malignant
phenotype of LCSCs and providing a new target for the treatment of
HCC.
The epigenetic regulation of lactylation also
transpires through non-histone mechanisms. The chromatin remodeling
protein BRG1 has been recognized as a 'reader' for histone
lactylation (86). It recognizes
and binds to H3K18la-enriched promoter regions, utilizing its
ATPase activity to remodel chromatin structure and alter its
accessibility. This mechanism enables BRG1 to mediate
lactylation-driven transcriptional reprogramming, profoundly
influencing cellular pluripotency and tumor progression (Fig. 2) (35). Beyond the aforementioned
examples, an expanding repertoire of lactylation targets has been
implicated in HCC pathogenesis and therapeutic resistance, which
are systematically summarized in Table II.
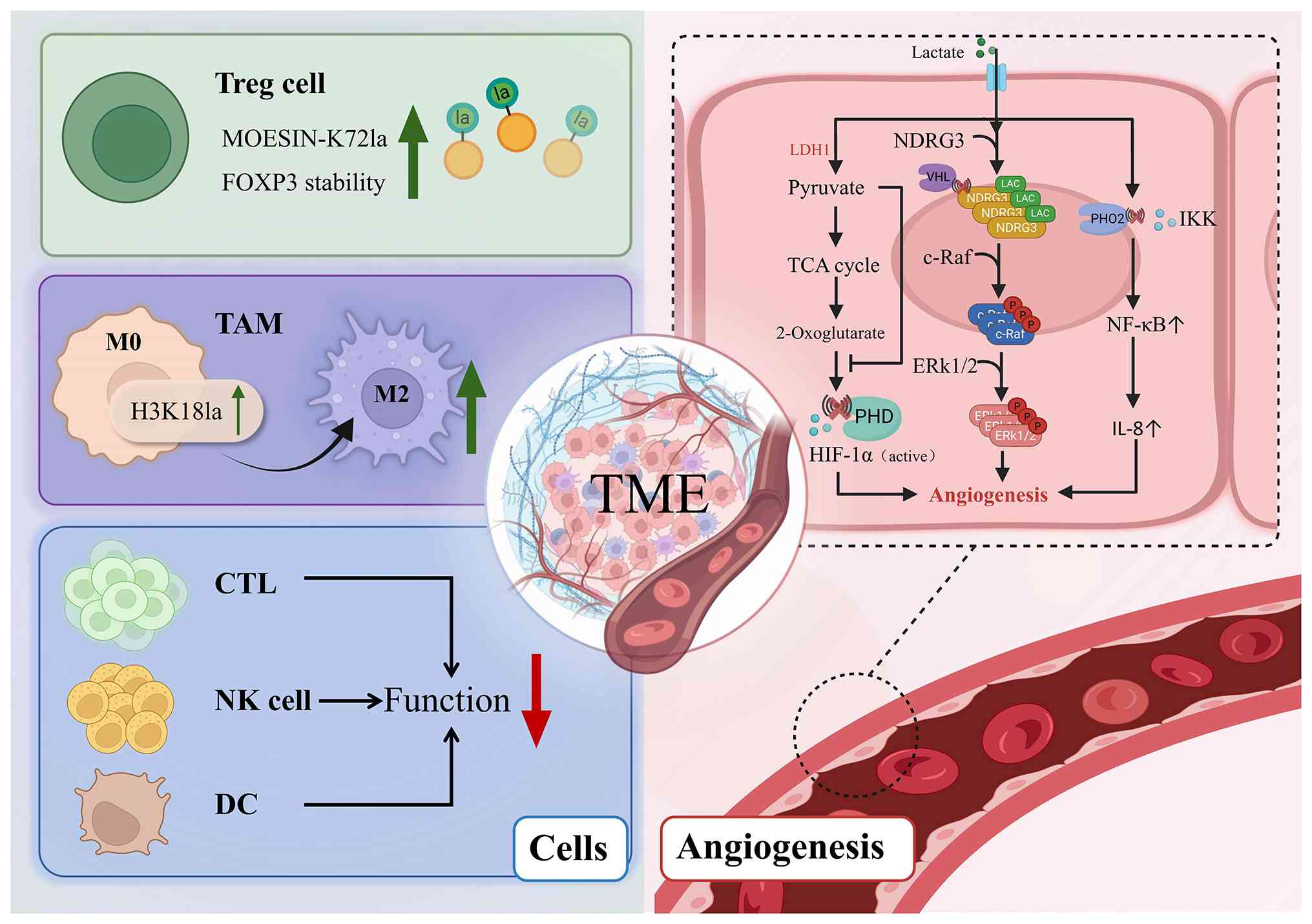
The TME of HCC is a complex ecosystem comprising
multiple components, including cancer cells, blood vessels, immune
cells and extracellular matrix (103). Dynamic interaction networks
form between cell populations through cell-cell contact, cytokine
exchange and metabolic product sharing, significantly influencing
tumor immune evasion and progression. Within this environment,
lactate functions as both the last result of glycolysis and an
essential signaling molecule and immunosuppressive agent (Fig. 3). It remodels the function of a
variety of immune cells and promotes angiogenesis through lactate
modification. This non-mutant epigenetic reprogramming and
phenotypic plasticity is a fundamental driver of malignancy and
provides a broader conceptual basis for how lactylation promotes
HCC progression (104).
Tregs are a crucial cell type that sustains
immunological tolerance and inhibits antitumor immune responses,
with their differentiation and function significantly influenced by
the tumor metabolic environment (105). In the high-lactic acid,
low-glucose microenvironment of HCC, Tregs actively absorb lactate
through the pronounced expression of monocarboxylate transporter 1
(MCT1). They convert lactate to pyruvate, which then enters the TCA
cycle to support mitochondrial oxidative phosphorylation (OXPHOS)
energy production. This allows Tregs to sustain functional
stability amid metabolic stress and assume a predominant role
within the tumor immunosuppressive network (106,107). Watson et al (106) conducted research indicating
that the Treg-specific deletion of MCT1 markedly diminished lactate
absorption and immunosuppressive function, hence reinforcing the
essential role of lactate metabolism in Treg homeostasis. However,
the complete mechanism by which it regulates immunosuppressive
molecules in the complex TME still needs further investigation.
Tumor-associated macrophages (TAMs) constitute a
significant population of immune cells within the HCC TME, serving
a pivotal function in tumor initiation, development and immune
evasion. TAMs can be traditionally classified into
pro-inflammatory, antitumor M1-type and anti-inflammatory,
pro-tumor M2-type based on their functional and phenotypic
properties (111). In the TME
of HCC, macrophages exhibit significant plasticity. After being
regulated by signals such as tumor cell metabolites, hypoxia and
cytokines, most are induced to polarize into the pro-tumor M2 TAMs
(112). Clinicopathological
analysis has shown that, in HCC tissues, the high infiltration of
M2 TAMs (cluster of differentiation CD206+ or
CD163+) is closely associated with tumor invasiveness,
angiogenesis and poor prognosis (113,114). Numerous evidence has confirmed
that high lactic acid accumulation in the TME can induce the
polarization of TAMs into the immunosuppressive M2 phenotypes
(115). The study by Zhang
et al (7) first revealed
that lactate from tumor cells can be taken up by macrophages
through MCTs and mediated by histone Kla under the catalysis of
acetyltransferase p300/CBP. This process exhibits a 'lactate
clock'-like dynamic amplification, progressively enhancing
M2-associated gene expression as lactate accumulates (7). Chromatin immunoprecipitation
sequencing analysis showed that H3K18la enriched in the promoter
regions of M2-associated genes such as arginase 1 and VEGFA,
directly promoting their transcriptional activation and TAM from M1
phenotype to M2 phenotype, enhancing the immune escape of tumors.
In addition, lactylation changes enhanced the stability of the M2
phenotype through the regulation of metabolic reprogramming.
Lactate increases tumor-associated macrophage reliance on OXPHOS
and the TCA cycle (116),
further stabilizing the M2-like metabolic phenotype and reinforcing
immunosuppressive functions. A recent study highlighted the
significant functional heterogeneity of TAMs in HCC. Emerging
evidence has suggested that specific subsets, such as
CD48+ macrophages, play an irreplaceable role in shaping
the immunosuppressive landscape and are priority targets for
next-generation immunotherapies (111).
The accumulation of lactate markedly diminishes the
anticancer efficacy of cytotoxic T lymphocytes (CTLs). It has been
shown that lactate can inhibit the activity of the CTL effect by
activating the c-Jun N-terminal kinase/c-Jun signaling pathway.
Simultaneously, lactate reduces the release of inflammatory
cytokines such as interferon (IFN)-γ and TNF-α, and downgrades the
expression of perforin and granzyme B, thus significantly weakening
the cytotoxic response mediated by CTL and weakening the antitumor
immune response (117).
Natural killer (NK) cells are also strongly
inhibited by lactate. Increased lactate concentration in tumor
tissues can weaken NK cell function through multiple pathways,
including by downregulating the expression of surface-activated
receptor NKp46, inhibiting mechanistic target of rapamycin
signaling and preventing the nuclear translocation of promyelocytic
leukemia zinc finger, thereby reducing metabolic activity and
cytotoxicity (76,118-120). In addition, the
lactate-mediated acidic environment can also directly induce the
apoptosis of liver-resident NK cells, further weakening innate
immune defense (76). However,
the aforementioned study mainly focuses on the CRLM model, and its
universality in HCC still needs further exploration.
Dendritic cells (DCs) are key antigen-presenting
cells that bridge innate and adaptive immunity. A recent study has
shown that lactate can activate the sterol regulatory
element-binding protein 2 pathway (121). Through the action of Toll-like
receptor (TLR), lactate can upregulate IL-10 and inhibit the
expression of IL-12, thus hindering the maturation and antigen
presentation function of DC. In melanoma and prostate cancer, there
are tumor-associated DCs, which are characterized by reduced IL-12
secretion and reduced CD1a expression. Gottfried et al
(122) found that lactate
accumulation can induce this DC phenotype (122). Plasmacytoid DCs (pDCs) can
produce type I interferon, which promotes tumors to avoid immune
attacks. On the contrary, lactate can activate the
G-protein-coupled receptor 81 (also known as HCAR1) receptor,
induce cytoplasmic Ca2+ mobilization and then downgrade
the IFN-I, thus weakening its antitumor immune activity (123). In addition, glycolysis
functions as the principal energy source for pDC IFN-γ generation
upon TLR stimulation; thus, lactate accumulation hinders pDC energy
metabolism by obstructing glycolysis, therefore further reducing
their antitumor efficacy (124).
Lactate is a principal metabolic byproduct in the
TME, which functions both as an indicator of active glycolysis and
an essential signaling molecule. It promotes angiogenesis through
multiple mechanisms, hence endorsing persistent tumor cell
proliferation and invasion in hypoxic environments. Depending on
whether hypoxia-inducible factor (HIF) is involved, the
pro-angiogenic effect mainly involves the following two
mechanisms.
In a hypoxic environment, the accumulated lactate
can stabilize HIF-1α, thereby enhancing multiple
angiogenesis-related pathways and promoting neovascularization.
Lactate specifically competes with 2-ketoglutarate for binding and
decreases the action of pyruvate dehydrogenase E1 component subunit
beta (PDH2) (125). Decreased
PDH2 activity inhibits its degradation and then impairs HIF-1α
hydroxylation through the PHD2/von Hippel-Lindau pathway, thereby
increasing HIF-1α stability (126). Stable HIF-1α can increase the
expression of downstream related factors (including VEGFA and basic
fibroblast growth factor), which can promote the proliferation of
tumor-associated endothelial cells and angiogenesis (127).
In addition to stabilizing HIF-1α, lactate can also
promote angiogenesis independently of HIF signaling. After entering
tumor-associated endothelial cells through MCT1, lactate can
activate the NF-κB pathway by inhibiting the hydroxylation of IκB
kinase by PDH2, thereby enhancing IL-8 transcriptional expression
and promoting angiogenesis maturation and stabilization (128). Furthermore, lactate can also
inhibit the PHD2/VHL-dependent degradation of N-Myc downstream
regulatory gene 3 (NDRG3) by binding to it, further prolonging the
half-life of NDRG3. Under hypoxic conditions, stable NDRG3 and
cellular rapidly accelerated fibrosarcoma (C-Raf) jointly activate
the Raf-extracellular signal-regulated kinase pathway and promote
angiogenesis (129). This
constitutes the HIF-independent lactate-NDRG3-C-Raf axis, which is
an important compensatory pathway for tumors to maintain blood
supply in a hypoxic environment.
In conclusion, lactylation and lactate signaling
jointly participate in the multifaceted regulation of HCC
angiogenesis, providing a new metabolic target for anti-angiogenic
therapy.
As research into lactylation deepens, its crucial
role in the development and progression of HCC is becoming
increasingly clear, providing new directions for clinical
diagnosis, prognostic assessment and treatment. By linking
metabolic reprogramming with epigenetic regulation, lactylation not
only reveals the molecular mechanisms of metabolic adaptation in
HCC but also provides a theoretical basis for combined
'metabolic-epigenetic-immune' intervention.
Early diagnosis and recurrence prediction of HCC
still face significant challenges. Currently, commonly used
clinical biomarkers, such as α-fetoprotein, lack sufficient
specificity, and therefore new molecular indicators are urgently
needed. Zhang et al (130) established a dual-purpose model
based on lactate-associated genes based on multi-cohort data from
The Cancer Genome Atlas (TCGA; https://www.cancer.gov/ccg/research/genome-sequencing/tcga)
and Gene Expression Omnibus (GEO; https://www.ncbi.nlm.nih.gov/geo/). Their analysis
showed that specific markers, including basigin, lysine
acetyltransferase 2A and zinc finger E-box binding homeobox 1, not
only had a strong diagnostic performance (area under the curve of
>0.8) in distinguishing between HCC and normal tissue, but also
effectively predict patients' response to immunotherapy. Similarly,
Luan (131) also conducted
related studies. Using single-cell and spatial transcriptomics
techniques, they found that multiple histone lactylation-related
genes, such as EP300, SIRT2 and LDHA were significantly upregulated
in HCC tissues and were associated with CD8+ T cell
infiltration, providing a new direction for lactylation-modified
radiomics and liquid biopsy. Of note, lactylation modification can
also activate gene transcription by remodeling the epigenetic
state, promoting cell proliferation, invasion and immune escape
(132). In HCC, elevated
lactylation-related risk scores are associated with significantly
poorer OS and disease-free survival (131). Therefore, detecting serum or
tissue lactate levels, LDHA/MCT4 (SLC16A3) expression, and
lactylation modification markers may become a new method for
predicting the probability of HCC recurrence and progression.
Despite the promising diagnostic performance of these gene
signatures, the current evidence is predominantly limited by its
retrospective nature and heavy reliance on public databases, such
as TCGA and GEO. To transition into clinical practice, these
markers must undergo rigorous testing in large-scale, multi-center
prospective cohorts to account for the high inter-patient
heterogeneity of HCC.
However, it is noteworthy that the interpretation of
lactylation-related biomarkers in clinical studies can be
significantly interfered with by acetylation, as these two PTMs
share significant biochemical and regulatory similarities.
Structurally, the highly similar physicochemical properties and
small mass difference between lactyl and acetyl groups make
accurate differentiation using conventional mass spectrometry and
antibody-based detection methods difficult (14,26). Both lactylation and acetylation
can occur on lysine residues and are catalyzed by overlapping
enzyme systems, particularly HATs, such as p300/CBP (7,17,40). Furthermore, members of the HDAC
and SIRT families can also function as deacetylases and
delactylases, further increasing the overlap in their regulatory
networks (24,50,133,134). This leads to signals attributed
to lactylation in clinical settings potentially reflecting
acetylation or mixed acylation states, reducing the specificity of
biomarkers. Therefore, improving detection specificity and
developing reliable strategies to differentiate these modifications
are crucial steps in advancing the clinical translation of
lactylation in HCC.
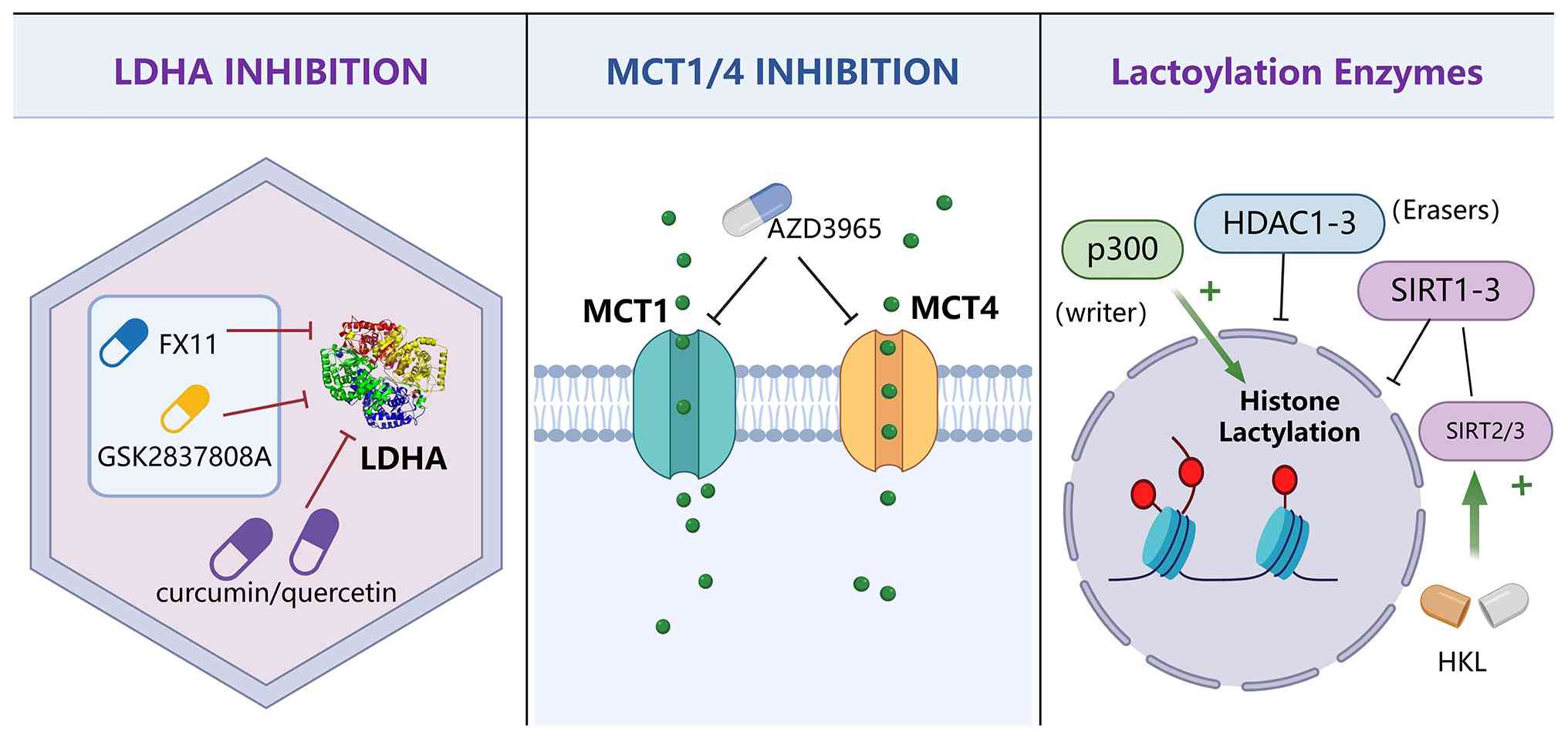
In HCC, lactylation can regulate metabolic
reprogramming and epigenetic regulation, providing a new
theoretical basis and breakthrough point for treatment. There are
two types of mainstream treatment methods. The first targets
lactate production, efflux and signaling pathways, reducing
immunosuppression and invasiveness by inhibiting tumor metabolism.
The second regulates the activity of lactylation-related enzymes to
intervene in lactate-mediated transcriptional reprogramming and
immune escape processes (Fig.
4).
LDHA is a key enzyme in the Warburg effect, which
can catalyze the reduction of pyruvate to lactate. It can be
inferred that inhibiting LDHA can effectively reduce the
accumulation of lactate, thus inhibiting lactylation in tumors.
Studies have shown that the high expression of LDHA is
significantly associated with the aggressive enhancement,
immunosuppression and poor prognosis of HCC. Some experimental
small molecules act as competitive LDHA inhibitors, such as FX11,
quinolin-3-sulfonamide derivatives, such as GSK2837808A, and
natural product derivatives, such as curcumin and quercetin, have
exhibited significant antitumor activity in various cancer models
(135). In the HCC model, LDHA
inhibitors can inhibit tumor proliferation and reduce the level of
histone lactylation, thus reducing the expression of oncogenes such
as VEGFA (136). It is worth
noting that the excessive inhibition of LDH may disrupt the normal
hepatocyte metabolism.
Lactate is transported between the tumor and its
microenvironment through MCTs. MCT1 is responsible for lactate
uptake, while MCT4 is involved in lactate efflux. MCT
overexpression promotes tumor metabolic symbiosis and the formation
of an immunosuppressive microenvironment. AZD3965, the first MCT1
inhibitor to enter clinical trials (NCT01791595), has demonstrated
favorable safety and tolerability in a variety of solid tumors
(137,138). In HCC cells, MCT1/MCT4
downregulation can prevent lactate efflux and increase the pH value
in the tumor, thus restoring the effect of T cells and DCs
(139).
Research indicates that the acetyltransferase p300
may act as a potential 'acylation-writing' enzyme, facilitating the
acetylation of histone lysine residues (109). A recent study indicated that
HDAC1-3 and SIRT1-3 are capable of removing lactyl modifications
from both histones and non-histones (131). In HCC models, SIRT3 has been
demonstrated to remove lactylation from CCNE2, consequently
inhibiting cell cycle progression (50). HKL activates SIRT2 and SIRT3,
demonstrating the inhibition of HCC cell proliferation in
preclinical studies. Of note, HKL treatment significantly reduced
the liver tumor burden in mice without causing pathological damage
to the kidneys and spleen (140). However, its actual impact on
clinical treatment requires further research.
Lactate accumulation can promote tumor immune
escape, which provides a new opportunity for 'metabolism-immune
combination therapy'. A high-lactate environment promotes the
polarization of TAMs towards an immunosuppressive phenotype, and
influences PD-L1 expression through lactate, consequently reducing
the effectiveness of immune checkpoint inhibitors (141).
Furthermore, lactate is closely associated with stem
cell-like phenotypes and tyrosine kinase inhibitor (TKI) resistance
(142). Numerous studies have
demonstrated that lactylation facilitates epithelial-mesenchymal
transition and activates pathways associated with drug tolerance
(143-145). The combined use of lactylation
inhibitors with multi-target TKIs, such as sorafenib or lenvatinib
can simultaneously inhibit drug resistance mechanisms at the
metabolic and epigenetic levels (146). In addition, combining HDAC
inhibitors or DNA methylation inhibitors can synergistically affect
the open state of chromatin and further enhance the epigenetic
effect of lactylation inhibition (131). These strategies are expected to
be key breakthroughs in overcoming the challenges of precision
treatment for HCC.
While intervention strategies targeting lactylation
in HCC have shown antitumor activity in preclinical studies, their
clinical translation still faces two major challenges: Insufficient
target selectivity and potential toxicity (147). These drugs often do not only
act on tumor cells but may also affect normal tissues. Therefore,
lactylation-targeted therapy cannot simply adopt a
'one-size-fits-all' strategy. Taking LDHA inhibitors as an example,
although existing studies have confirmed that they can enhance the
antitumor effect of drugs such as sorafenib on HCC by inhibiting
lactate production and inhibiting tumor cell proliferation
(148,149), the direct toxicological
evidence on normal hepatocytes is still limited. Considering that
the healthy liver undertakes most of the body's lactate clearance
function, systemic LDHA inhibition may theoretically interfere with
the function of normal hepatocytes, thereby bringing metabolic
toxicity risks. Based on this, a more feasible direction in the
future could be to combine tumor-targeted delivery, combination
therapy, and lactylation feature stratification to improve the
therapeutic index and minimize the impact on normal liver
tissue.
Lactylation is an emerging research epigenetic
modification, which affects gene transcription, factor secretion
and cellular phenotypic remodeling through histone and non-histone
mechanisms, thus playing a role in the progression of HCC. As our
understanding of the regulatory mechanisms of lactylation deepens,
an increasing number of regulatory factors are being
discovered.
However, several key scientific questions still
remain to be addressed. First, the regulatory system of lactylation
is not fully defined, including whether tissue-or context-specific
enzymes exist. Secondly, the association between lactylation and
other acylation modifications, particularly acetylation, requires
clarification due to shared substrates and enzymes. Thirdly, the
role of lactylation in the TME, especially in immune regulation,
remains incompletely understood. Finally, the way to precisely and
safely target lactylation in HCC remains challenging, given the
essential role of lactate metabolism and the limited clinical
validation of related biomarkers. Addressing these questions will
be essential for bridging the gap between mechanistic insights and
the clinical application of lactylation in HCC.
Not applicable.
XF and QC conceptualized the review. LC and FG wrote
the draft of the manuscript. QM reviewed the literature. YL revised
the manuscript. All authors read and approved the final version of
the manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by the National Natural Science
Foundation of China (grant no. 82500760), the Zhejiang Provincial
Natural Science Foundation of China (grant no. LQN25H070003) and
the Ningbo Yongjiang Talent Program (grant no. 2024A-422-G).
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Singal AG, Kanwal F and Llovet JM: Global
trends in hepatocellular carcinoma epidemiology: implications for
screening, prevention and therapy. Nat Rev Clin Oncol. 20:864–884.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Llovet JM, Kelley RK, Villanueva A, Singal
AG, Pikarsky E, Roayaie S, Lencioni R, Koike K, Zucman-Rossi J and
Finn RS: Hepatocellular carcinoma. Nat Rev Dis Primers. 7:62021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim DY: Changing etiology and epidemiology
of hepatocellular carcinoma: Asia and worldwide. J Liver Cancer.
24:62–70. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan W, Adebowale K, Váncza L, Li Y, Rabbi
MF, Kunimoto K, Chen D, Mozes G, Chiu DK, Li Y, et al: Matrix
viscoelasticity promotes liver cancer progression in the
pre-cirrhotic liver. Nature. 626:635–642. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang D, Tang Z, Huang H, Zhou G, Cui C,
Weng Y, Liu W, Kim S, Lee S, Perez-Neut M, et al: Metabolic
regulation of gene expression by histone lactylation. Nature.
574:575–580. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang H, Wang X, Wang W, Qu X, Song X,
Zhang Y, Zhong L, Yang DP, Dong X and Zhao Y: Injectable hydrogel
for postoperative synergistic photothermal-chemodynamic tumor and
anti-infection therapy. Biomaterials. 280:1212892022. View Article : Google Scholar
|
|
9
|
Chen J, He G, Cai D, Giovannetti E,
Inamura K, Liu S and Ma W: Lactic acid: a narrative review of a
promoter of the liver cancer microenvironment. J Gastrointest
Oncol. 15:1282–1296. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zaib S, Hayyat A, Ali N, Gul A, Naveed M
and Khan I: Role of mitochondrial membrane potential and lactate
dehydrogenase A in apoptosis. Anticancer Agents Med Chem.
22:2048–2062. 2022. View Article : Google Scholar
|
|
11
|
Wang D, Du G, Chen X, Wang J, Liu K, Zhao
H, Cheng C, He Y, Jing N, Xu P, et al: Zeb1-controlled metabolic
plasticity enables remodeling of chromatin accessibility in the
development of neuroendocrine prostate cancer. Cell Death Differ.
31:779–791. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu Y, Hao X, Ren Y, Xu Q, Liu X, Song S
and Wang Y: Research progress of abnormal lactate metabolism and
lactate modification in immunotherapy of hepatocellular carcinoma.
Front Oncol. 12:10634232023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pan L, Feng F, Wu J, Fan S, Han J, Wang S,
Yang L, Liu W, Wang C and Xu K: Demethylzeylasteral targets lactate
by inhibiting histone lactylation to suppress the tumorigenicity of
liver cancer stem cells. Pharmacol Res. 181:1062702022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xie Y, Hu H, Liu M, Zhou T, Cheng X, Huang
W and Cao L: The role and mechanism of histone lactylation in
health and diseases. Front Genet. 13:9492522022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cui H, Xie N, Banerjee S, Ge J, Jiang D,
Dey T, Matthews QL, Liu RM and Liu G: Lung myofibroblasts promote
macrophage profibrotic activity through lactate-induced histone
lactylation. Am J Respir Cell Mol Biol. 64:115–125. 2021.
View Article : Google Scholar
|
|
16
|
Xiong J, He J, Zhu J, Pan J, Liao W, Ye H,
Wang H, Song Y, Du Y, Cui B, et al: Lactylation-driven
METTL3-mediated RNA m(6)A modification promotes immunosuppression
of tumor-infiltrating myeloid cells. Mol Cell. 82:1660–1677.e10.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang K, Fan M, Wang X, Xu J, Wang Y, Tu F,
Gill PS, Ha T, Liu L, Williams DL and Li C: Lactate promotes
macrophage HMGB1 lactylation, acetylation, and exosomal release in
polymicrobial sepsis. Cell Death Differ. 29:133–146. 2022.
View Article : Google Scholar :
|
|
18
|
Mao Y, Zhang J, Zhou Q, He X, Zheng Z, Wei
Y, Zhou K, Lin Y, Yu H, Zhang H, et al: Hypoxia induces
mitochondrial protein lactylation to limit oxidative
phosphorylation. Cell Res. 34:13–30. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ju J, Zhang H, Lin M, Yan Z, An L, Cao Z,
Geng D, Yue J, Tang Y, Tian L, et al: The alanyl-tRNA synthetase
AARS1 moonlights as a lactyltransferase to promote YAP signaling in
gastric cancer. J Clin Invest. 134:e1745872024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zong Z, Xie F, Wang S, Wu X, Zhang Z, Yang
B and Zhou F: Alanyl-tRNA synthetase, AARS1, is a lactate sensor
and lactyltransferase that lactylates p53 and contributes to
tumorigenesis. Cell. 187:2375–2392.e33. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H, Liu C, Li R, Zhou L, Ran Y, Yang Q,
Huang H, Lu H, Song H, Yang B, et al: AARS1 and AARS2 sense
l-lactate to regulate cGAS as global lysine lactyltransferases.
Nature. 634:1229–1237. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Niu Z, Chen C, Wang S, Lu C, Wu Z, Wang A,
Mo J, Zhang J, Han Y, Yuan Y, et al: HBO1 catalyzes lysine
lactylation and mediates histone H3K9la to regulate gene
transcription. Nat Commun. 15:35612024. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan Z, Liu Z, Zhang N, Wei W, Cheng K, Sun
H and Hao Q: Identification of SIRT3 as an eraser of H4K16la.
iScience. 26:1077572023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moreno-Yruela C, Zhang D, Wei W, Bæk M,
Liu W, Gao J, Danková D, Nielsen AL, Bolding JE, Yang L, et al:
Class I histone deacetylases (HDAC1-3) are histone lysine
delactylases. Sci Adv. 8:eabi66962022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dai SK, Liu PP, Li X, Jiao LF, Teng ZQ and
Liu CM: Dynamic profiling and functional interpretation of histone
lysine crotonylation and lactylation during neural development.
Development. 149:dev2000492022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Moreno-Yruela C, Bæk M, Monda F and Olsen
CA: Chiral post-translational modification to Lysine ε-Amino
Groups. Acc Chem Res. 55:1456–1466. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zessin M, Meleshin M, Praetorius L, Sippl
W, Bařinka C and Schutkowski M: Uncovering robust delactoylase and
depyruvoylase activities of HDAC isoforms. ACS Chem Biol.
17:1364–1375. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: Metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yao S, Xu MD, Wang Y, Zhao ST, Wang J,
Chen GF, Chen WB, Liu J, Huang GB, Sun WJ, et al: Astrocytic
lactate dehydrogenase A regulates neuronal excitability and
depressive-like behaviors through lactate homeostasis in mice. Nat
Commun. 14:7292023. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Comandatore A, Franczak M, Smolenski RT,
Morelli L, Peters GJ and Giovannetti E: Lactate Dehydrogenase and
its clinical significance in pancreatic and thoracic cancers. Semin
Cancer Biol. 86(Pt): 93–100. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vayakkattil AB, Pazhanchery AS, Andrews S
and Vayakkattil U: The warburg effect redefined: A kinetic and
regulatory perspective. Cureus. 17:e933312025.PubMed/NCBI
|
|
32
|
Nagini S, Kallamadi PR, Tanagala KKK and
Reddy GB: Aldo-keto reductases: Role in cancer development and
theranostics. Oncol Res. 32:1287–1308. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Khayami R, Hashemi SR and Kerachian MA:
Role of aldo-keto reductase family 1 member B1 (AKR1B1) in the
cancer process and its therapeutic potential. J Cell Mol Med.
24:8890–8902. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang T, Ye Z, Li Z, Jing DS, Fan GX, Liu
MQ, Zhuo QF, Ji SR, Yu XJ, Xu XW and Qin Y: Lactate-induced protein
lactylation: A bridge between epigenetics and metabolic
reprogramming in cancer. Cell Prolif. 56:e134782023. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu X, Huang X, Yang Y, Sun Y, Zhao Y,
Zhang Z, Qiu D, Wu Y, Wu G and Lei L: Dux activates
metabolism-lactylation-MET network during early iPSC reprogramming
with Brg1 as the histone lactylation reader. Nucleic Acids Res.
52:5529–5548. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhai G, Niu Z, Jiang Z, Zhao F, Wang S,
Chen C, Zheng W, Wang A, Zang Y, Han Y and Zhang K: DPF2 reads
histone lactylation to drive transcription and tumorigenesis. Proc
Natl Acad Sci USA. 121:e24214961212024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nunez R, Sidlowski PFW, Steen EA,
Wynia-Smith SL, Sprague DJ, Keyes RF and Smith BC: The TRIM33
bromodomain recognizes histone lysine lactylation. ACS Chem Biol.
19:2418–2428. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ogryzko VV, Schiltz RL, Russanova V,
Howard BH and Nakatani Y: The transcriptional coactivators p300 and
CBP are histone acetyltransferases. Cell. 87:953–959. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Narita T, Ito S, Higashijima Y, Chu WK,
Neumann K, Walter J, Satpathy S, Liebner T, Hamilton WB, Maskey E,
et al: Enhancers are activated by p300/CBP activity-dependent PIC
assembly, RNAPII recruitment, and pause release. Mol Cell.
81:2166–2182.e6. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen Q, Yang B, Liu X, Zhang XD, Zhang L
and Liu T: Histone acetyltransferases CBP/p300 in tumorigenesis and
CBP/p300 inhibitors as promising novel anticancer agents.
Theranostics. 12:4935–4948. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li F, Si W, Xia L, Yin D, Wei T, Tao M,
Cui X, Yang J, Hong T and Wei R: Positive feedback regulation
between glycolysis and histone lactylation drives oncogenesis in
pancreatic ductal adenocarcinoma. Mol Cancer. 23:902024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang X, Fan W, Li N, Ma Y, Yao M, Wang G,
He S, Li W, Tan J, Lu Q and Hou S: YY1 lactylation in microglia
promotes angiogenesis through transcription activation-mediated
upregulation of FGF2. Genome Biol. 24:872023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li L, Jiang D, Liu H, Guo C, Zhao R, Zhang
Q, Xu C, Qin Z, Feng J, Liu Y, et al: Comprehensive proteogenomic
characterization of early duodenal cancer reveals the
carcinogenesis tracks of different subtypes. Nat Commun.
14:17512023. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen Z, Mei K, Xiao Y, Xiong Y, Long W,
Wang Q, Zhong J, Di D, Ge Y, Luo Y, et al: Prognostic assessment of
oxidative stress-related genes in colorectal cancer and new
insights into tumor immunity. Oxid Med Cell Longev.
2022:25183402022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu L, Gao J, Liu X, Zhang F, Hu B, Zhang
H, Wang Z, Tang H, Shi JH and Zhang S: AARS2 as a novel biomarker
for prognosis and its molecular characterization in pan-cancer.
Cancer Med. 12:21531–21544. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhu Z, Hou Q, Wang B, Li C, Liu L, Gong W,
Chai J and Guo H: A novel mitochondria-related gene signature for
controlling colon cancer cell mitochondrial respiration and
proliferation. Hum Cell. 35:1126–1139. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ishihama K, Yamakawa M, Semba S, Takeda H,
Kawata S, Kimura S and Kimura W: Expression of HDAC1 and CBP/p300
in human colorectal carcinomas. J Clin Pathol. 60:1205–1210. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Eichner LJ, Curtis SD, Brun SN, McGuire
CK, Gushterova I, Baumgart JT, Trefts E, Ross DS, Rymoff TJ and
Shaw RJ: HDAC3 is critical in tumor development and therapeutic
resistance in Kras-mutant non-small cell lung cancer. Sci Adv.
9:eadd32432023. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang H, Fu C, Du J, Wang H, He R, Yin X,
Li H, Li X, Wang H, Li K, et al: Enhanced histone H3 acetylation of
the PD-L1 promoter via the COP1/c-Jun/HDAC3 axis is required for
PD-L1 expression in drug-resistant cancer cells. J Exp Clin Cancer
Res. 39:292020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jin J, Bai L, Wang D, Ding W, Cao Z, Yan
P, Li Y, Xi L, Wang Y, Zheng X, et al: SIRT3-dependent
delactylation of cyclin E2 prevents hepatocellular carcinoma
growth. EMBO Rep. 24:e560522023. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zu H, Li C, Dai C, Pan Y, Ding C, Sun H,
Zhang X, Yao X, Zang J and Mo X: SIRT2 functions as a histone
delactylase and inhibits the proliferation and migration of
neuroblastoma cells. Cell Discov. 8:542022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jena BC, Das CK, Banerjee I, Bharadwaj D,
Majumder R, Das S, Biswas A, Kundu M, Roy PK, Kundu CN and Mandal
M: TGF-β1 induced autophagy in cancer associated fibroblasts during
hypoxia contributes EMT and glycolysis via MCT4 upregulation. Exp
Cell Res. 417:1131952022. View Article : Google Scholar
|
|
53
|
Hu X, Huang Z and Li L: LDHB mediates
histone lactylation to activate PD-L1 and promote ovarian cancer
immune escape. Cancer Invest. 43:70–79. 2025. View Article : Google Scholar
|
|
54
|
Huang Y, Zhang W, Hao J and Zhu R: The
application value of lactate dehydrogenase in gynecological
malignant tumors. Front Oncol. 15:17311872026. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Jha MK, Lee IK and Suk K: Metabolic
reprogramming by the pyruvate dehydrogenase kinase-lactic acid
axis: Linking metabolism and diverse neuropathophysiologies.
Neurosci Biobehav Rev. 68:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bennis Y, Bodeau S, Batteux B,
Gras-Champel V, Masmoudi K, Maizel J, De Broe ME, Lalau JD and
Lemaire-Hurtel AS: A study of associations between plasma metformin
concentration, lactic acidosis, and mortality in an emergency
hospitalization context. Crit Care Med. 48:e1194–e1202. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Arce-Molina R, Cortés-Molina F, Sandoval
PY, Galaz A, Alegría K, Schirmeier S, Barros LF and San Martín A: A
highly responsive pyruvate sensor reveals pathway-regulatory role
of the mitochondrial pyruvate carrier MPC. Elife. 9:e539172020.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schwab A, Siddiqui MA, Ramesh V,
Gollavilli PN, Turtos AM, Møller SS, Pinna L, Havelund JF, Rømer
AMA, Ersan PG, et al: Polyol pathway-generated fructose is
indispensable for growth and survival of non-small cell lung
cancer. Cell Death Differ. 32:587–597. 2025. View Article : Google Scholar :
|
|
59
|
Liu Y, Zhou C, Tang Y, Lei H, Aihemaiti A,
Liu H, Zou P, Xie J, Guo X, Xia R, et al: Targeting AKR1B1
reprograms tumor-associated macrophages to enhance antitumor
immunity. J Immunother Cancer. 14:e0140432026. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu Z, Yuan J, Su S, Han J, Zeng N, Ma Y,
Chen N and Lv T: AARS1-mediated AKR1B10 lactylation stabilizes an
aerobic glycolysis-positive feedback loop to drive lenvatinib
resistance in hepatocellular carcinoma. Clin Transl Med.
16:e705612026. View Article : Google Scholar
|
|
61
|
Miranda-Gonçalves V, Gonçalves CS, Granja
S, Vieira de Castro J, Reis RM, Costa BM and Baltazar F: MCT1 is a
new prognostic biomarker and its therapeutic inhibition boosts
response to temozolomide in human glioblastoma. Cancers (Basel).
13:34682021. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Payen VL, Mina E, Van Hée VF, Porporato PE
and Sonveaux P: Monocarboxylate transporters in cancer. Mol Metab.
33:48–66. 2020. View Article : Google Scholar :
|
|
63
|
Singh M, Afonso J, Sharma D, Gupta R and
Kumar V, Rani R, Baltazar F and Kumar V: Targeting monocarboxylate
transporters (MCTs) in cancer: How close are we to the clinics?
Semin Cancer Biol. 90:1–14. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Clem B, Telang S, Clem A, Yalcin A, Meier
J, Simmons A, Rasku MA, Arumugam S, Dean WL, Eaton J, et al:
Small-molecule inhibition of 6-phosphofructo-2-kinase activity
suppresses glycolytic flux and tumor growth. Mol Cancer Ther.
7:110–120. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Feng Y, Xiong Y, Qiao T, Li X, Jia L and
Han Y: Lactate dehydrogenase A: A key player in carcinogenesis and
potential target in cancer therapy. Cancer Med. 7:6124–6136. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Basheeruddin M and Qausain S:
Hypoxia-inducible factor 1-Alpha (HIF-1α): An essential regulator
in cellular metabolic control. Cureus. 16:e638522024.
|
|
67
|
Gordan JD, Thompson CB and Simon MC: HIF
and c-Myc: sibling rivals for control of cancer cell metabolism and
proliferation. Cancer Cell. 12:108–113. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Dang CV, Hamaker M, Sun P, Le A and Gao P:
Therapeutic targeting of cancer cell metabolism. J Mol Med (Berl).
89:205–212. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Mossmann D, Park S and Hall MN: mTOR
signalling and cellular metabolism are mutual determinants in
cancer. Nat Rev Cancer. 18:744–757. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Allen E, Miéville P, Warren CM, Saghafinia
S, Li L, Peng MW and Hanahan D: Metabolic symbiosis enables
adaptive resistance to anti-angiogenic therapy that is dependent on
mTOR signaling. Cell Rep. 15:1144–1160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liao J, Chen Z, Chang R, Yuan T, Li G, Zhu
C, Wen J, Wei Y, Huang Z, Ding Z, et al: CENPA functions as a
transcriptional regulator to promote hepatocellular carcinoma
progression via cooperating with YY1. Int J Biol Sci. 19:5218–5232.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Xu H, Li L, Wang S, Wang Z, Qu L, Wang C
and Xu K: Royal jelly acid suppresses hepatocellular carcinoma
tumorigenicity by inhibiting H3 histone lactylation at H3K9la and
H3K14la sites. Phytomedicine. 118:1549402023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yang Z, Yan C, Ma J, Peng P, Ren X, Cai S,
Shen X, Wu Y, Zhang S, Wang X, et al: Lactylome analysis suggests
lactylation-dependent mechanisms of metabolic adaptation in
hepatocellular carcinoma. Nat Metab. 5:61–79. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wu X: In-depth discovery of protein
lactylation in hepatocellular carcinoma. Proteomics.
23:e23000032023. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Huang J, Xie H, Li J, Huang X, Cai Y, Yang
R, Yang D, Bao W, Zhou Y, Li T and Lu Q: Histone lactylation drives
liver cancer metastasis by facilitating NSF1-mediated ferroptosis
resistance after microwave ablation. Redox Biol. 81:1035532025.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Harmon C, Robinson MW, Hand F, Almuaili D,
Mentor K, Houlihan DD, Hoti E, Lynch L, Geoghegan J and O'Farrelly
C: Lactate-mediated acidification of tumor microenvironment induces
apoptosis of liver-resident NK cells in colorectal liver
metastasis. Cancer Immunol Res. 7:335–346. 2019. View Article : Google Scholar
|
|
77
|
Cheng S, Xiao X, Wang D, Wang X and Yang
M: Lactate and lactylation in liver diseases: Energy metabolism,
inflammatory immunity and tumor microenvironment. Front Immunol.
16:15815822025. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Shangguan X, He J, Ma Z, Zhang W, Ji Y,
Shen K, Yue Z, Li W, Xin Z, Zheng Q, et al: SUMOylation controls
the binding of hexokinase 2 to mitochondria and protects against
prostate cancer tumorigenesis. Nat Commun. 12:18122021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Tan Q, Liu M and Tao X: Targeting
lactylation: From metabolic reprogramming to precision therapeutics
in liver diseases. Biomolecules. 15:11782025. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Gao R, Li Y, Xu Z, Zhang F, Xu J, Hu Y,
Yin J, Yang K, Sun L, Wang Q, et al: Mitochondrial pyruvate carrier
1 regulates fatty acid synthase lactylation and mediates treatment
of nonalcoholic fatty liver disease. Hepatology. 78:1800–1815.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Meng J, Yan C and Liu J: LDHA-mediated
histone lactylation promotes the nonalcoholic fatty liver disease
progression through targeting The METTL3/YTHDF1/SCD1 m6A axis.
Physiol Res. 73:985–999. 2024. View Article : Google Scholar
|
|
82
|
Jiang X, Stockwell BR and Conrad M:
Ferroptosis: Mechanisms, biology and role in disease. Nat Rev Mol
Cell Biol. 22:266–282. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ji Y, Xu Z, Tang L, Huang T, Mu X, Ni C,
Tang B, Lu H, Zhang C, Yang S and Wang X: O-GlcNAcylation of YBX1
drives a glycolysis-histone lactylation feedback loop in
hepatocellular carcinoma. Cancer Lett. 631:2179572025. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Feng F, Wu J, Chi Q, Wang S, Liu W, Yang
L, Song G, Pan L, Xu K and Wang C: Lactylome analysis unveils
lactylation-dependent mechanisms of stemness remodeling in the
liver cancer stem cells. Adv Sci (Weinh). 11:e24059752024.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ma F and Yu W: The roles of lactate and
lactylation in diseases related to mitochondrial dysfunction. Int J
Mol Sci. 26:71492025. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Liu X, Wang J, Lao M, Liu F, Zhu H, Man K
and Zhang J: Study on the effect of protein lysine lactylation
modification in macrophages on inhibiting periodontitis in rats. J
Periodontol. 95:50–63. 2024. View Article : Google Scholar
|
|
87
|
Wei X, Zou L, Huang Y, Qiu C, Cheng G,
Chen Y and Rao J: LDHA-mediated YAP lactylation promotes the tumor
progression of hepatocellular carcinoma by inducing YAP
dephosphorylation and activation. Biol Direct. 20:642025.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wang L, Zeng T, Wang Y, Wang G, Yu W,
Zhang J, Shi Y, Li J and Ding J: K90 lactylation orchestrates YAP
nuclear sequestration by impairing binding with exportin CRM1 and
enhances HCC malignancy. Cancer Lett. 611:2173862024. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hong H, Han H, Wang L, Cao W, Hu M, Li J,
Wang J, Yang Y, Xu X, Li G, et al: ABCF1-K430-Lactylation promotes
HCC malignant progression via transcriptional activation of HIF1
signaling pathway. Cell Death Differ. 32:613–631. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Lu Y, Zhu J, Zhang Y, Li W, Xiong Y, Fan
Y, Wu Y, Zhao J, Shang C, Liang H and Zhang W: Lactylation-Driven
IGF2BP3-Mediated serine metabolism reprogramming and RNA
m6A-modification promotes lenvatinib resistance in HCC. Adv Sci
(Weinh). 11:e24013992024. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Han H, Wang S, Ma L, Yin H, Cheng X, Wang
Y, Xia S, Zhang Y, Zhang Y, Zhu R, et al: ASH2L-K312-Lac stimulates
angiogenesis in tumors to expedite the malignant progression of
hepatocellular carcinoma. Adv Sci (Weinh). 12:e094772025.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Jiang C, He X, Chen X, Huang J, Liu Y,
Zhang J, Chen H, Sui X, Lv X, Zhao X, et al: Lactate accumulation
drives hepatocellular carcinoma metastasis through facilitating
tumor-derived exosome biogenesis by Rab7A lactylation. Cancer Lett.
627:2176362025. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Sun Y, Luo C, Yang H, Ye J, Song F, Yi Q,
Zou W, Huang Y, Fan X, Wang L, et al: Lactylation Converts ABHD6
into a mitochondrial regulator that drives lenvatinib resistance in
hepatocellular carcinoma. Cancer Res. Mar 20–2026.Epub ahead of
print.
|
|
94
|
Li Q, Yu K, Zhou S, Fang C, Zhang L, Jin
F, Hou X, Yao F, Tang Y, Xu J, et al: JOSD1 drives hepatocellular
carcinoma malignancy by modulating the ubiquitination-lactylation
switch on PGAM1. Gut. Apr 28–2026.Epub ahead of print. View Article : Google Scholar
|
|
95
|
Liu S, Pan Y, Liu W, Bu X, Shao R, Wang Q,
Wu J, Wu C, Hu W, Xu J, et al: Lactylation-driven MVP upregulation
boosts immunotherapy resistance by inhibiting PD-L1 degradation in
hepatocellular carcinoma. J Immunother Cancer. 13:e0122302025.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Chen S, Zhao L, Han P, Liu J, Hou J, Liu
X, Zhao Q, Wang R, Wang F and Li J: Lactate drives immune
resistance via a pharmaceutically reversible
H3K18la-KIF20A-c-Myc-PD-L1 axis in hepatocellular carcinoma. Cancer
Biol Med. Apr 2–2026.Epub ahead of print. View Article : Google Scholar
|
|
97
|
Ye J, Gao X, Huang X, Huang S, Zeng D, Luo
W, Zeng C, Lu C, Lu L, Huang H, et al: Integrating single-cell and
spatial transcriptomics to uncover and elucidate GP73-Mediated
pro-angiogenic regulatory networks in hepatocellular carcinoma.
Research (Wash DC). 7:03872024.
|
|
98
|
Wei E, Ji D, Jia Y, Sun Z, Gao C, Zeng C,
Wang C, Yu M, Shang G, Xie L, et al: BRD9 recognizes
lactate-induced H3K18 lactylation to drive oncogenic chromatin
remodeling in hepatocellular carcinoma. Cell Death Differ. Mar
7–2026.Epub ahead of print. View Article : Google Scholar
|
|
99
|
Cai J, Zhang P, Cai Y, Zhu G, Chen S, Song
L, Du J, Wang B, Dai W, Zhou J, et al: Lactylation-Driven NUPR1
promotes immunosuppression of tumor-infiltrating macrophages in
hepatocellular carcinoma. Adv Sci (Weinh). 12:e24130952025.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Cai J, Song L, Zhang F, Wu S, Zhu G, Zhang
P, Chen S, Du J, Wang B, Cai Y, et al: Targeting SRSF10 might
inhibit M2 macrophage polarization and potentiate anti-PD-1 therapy
in hepatocellular carcinoma. Cancer Commun (Lond). 44:1231–1260.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zou X, Wang N, Guan C, Gao J, Shi W, Yang
C, Bi S, Zhang T and Zhong X: PRC1 promotes MAFLD progression by
regulating glycolysis/lactylation axis and forming a positive
feedback loop. J Nutr Biochem. 154:1103782026. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Xie J, Pan X and Xia Y: PDP1 drives
hepatocellular carcinoma progression by regulating senescence
through the cAMP/Ca(2+) signaling pathway. Biochem Pharmacol.
247:1177722026. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Sangro B, Sarobe P, Hervás-Stubbs S and
Melero I: Advances in immunotherapy for hepatocellular carcinoma.
Nat Rev Gastroenterol Hepatol. 18:525–543. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Hanahan D: Hallmarks of cancer: New
dimensions. Cancer Discov. 12:31–46. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Kasagi S, Zhang P, Che L, Abbatiello B,
Maruyama T, Nakatsukasa H, Zanvit P, Jin W, Konkel JE and Chen W:
In vivo-generated antigen-specific regulatory T cells treat
autoimmunity without compromising antibacterial immune response.
Sci Transl Med. 6:241ra782014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Watson MJ, Vignali PDA, Mullett SJ,
Overacre-Delgoffe AE, Peralta RM, Grebinoski S, Menk AV,
Rittenhouse NL, DePeaux K, Whetstone RD, et al: Metabolic support
of tumour-infiltrating regulatory T cells by lactic acid. Nature.
591:645–651. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Watson M, Vignali P, Mullet S,
Overacre-Delgoffe A, Peralta R, Grebinoski S, Menk A, Rittenhouse
N, DePeaux K, Whetstone R, et al: 517 Regulatory T cell functional
identity is sustained by a glucose:lactate axis that is exploited
in the tumor microenvironment. J ImmunoTher Cancer. 8:A337–A655.
2020.
|
|
108
|
Gu J, Zhou J, Chen Q, Xu X, Gao J, Li X,
Shao Q, Zhou B, Zhou H, Wei S, et al: Tumor metabolite lactate
promotes tumorigenesis by modulating MOESIN lactylation and
enhancing TGF-beta signaling in regulatory T cells. Cell Rep.
39:1109862022. View Article : Google Scholar
|
|
109
|
Xue Q, Peng W, Zhang S, Wei X, Ye L, Wang
Z, Xiang X, Liu Y, Wang H and Zhou Q: Lactylation-driven TNFR2
expression in regulatory T cells promotes the progression of
malignant pleural effusion. J Immunother Cancer. 12:e0100402024.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lopez Krol A, Nehring HP, Krause FF, Wempe
A, Raifer H, Nist A, Stiewe T, Bertrams W, Schmeck B, Luu M, et al:
Lactate induces metabolic and epigenetic reprogramming of
pro-inflammatory Th17 cells. EMBO Rep. 23:e546852022. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Han X and Jin R: Rewiring tumor-associated
macrophages in hepatocellular carcinoma. Front Immunol.
17:17756032026. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Mills CD, Kincaid K, Alt JM, Heilman MJ
and Hill AM: M-1/M-2 macrophages and the Th1/Th2 paradigm. J
Immunol. 164:6166–6173. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Huang Y, Ge W, Zhou J, Gao B, Qian X and
Wang W: The role of tumor associated macrophages in hepatocellular
carcinoma. J Cancer. 12:1284–1294. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Liu YT, Mao ZW, Ding Y and Wang WL:
Macrophages as targets in hepatocellular carcinoma therapy. Mol
Cancer Ther. 23:780–790. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Xu B, Liu Y, Li N and Geng Q: Lactate and
lactylation in macrophage metabolic reprogramming: Current progress
and outstanding issues. Front Immunol. 15:13957862024. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Noe JT, Rendon BE, Geller AE, Conroy LR,
Morrissey SM, Young LEA, Bruntz RC, Kim EJ, Wise-Mitchell A,
Barbosa de Souza Rizzo M, et al: Lactate supports a
metabolic-epigenetic link in macrophage polarization. Sci Adv.
7:eabi86022021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Mendler AN, Hu B, Prinz PU, Kreutz M,
Gottfried E and Noessner E: Tumor lactic acidosis suppresses CTL
function by inhibition of p38 and JNK/c-Jun activation. Int J
Cancer. 131:633–640. 2012. View Article : Google Scholar
|
|
118
|
Brand A, Singer K, Koehl GE, Kolitzus M,
Schoenhammer G, Thiel A, Matos C, Bruss C, Klobuch S, Peter K, et
al: LDHA-Associated lactic acid production blunts tumor
immunosurveillance by T and NK cells. Cell Metab. 24:657–671. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Husain Z, Huang Y, Seth P and Sukhatme VP:
Tumor-derived lactate modifies antitumor immune response: Effect on
myeloid-derived suppressor cells and NK cells. J Immunol.
191:1486–1495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Xie D, Zhu S and Bai L: Lactic acid in
tumor microenvironments causes dysfunction of NKT cells by
interfering with mTOR signaling. Sci China Life Sci. 59:1290–1296.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Plebanek MP, Xue Y, Nguyen YV, DeVito NC,
Wang X, Holtzhausen A, Beasley GM, Theivanthiran B and Hanks BA: A
lactate-SREBP2 signaling axis drives tolerogenic dendritic cell
maturation and promotes cancer progression. Sci Immunol.
9:eadi41912024. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Gottfried E, Kunz-Schughart LA, Ebner S,
Mueller-Klieser W, Hoves S, Andreesen R, Mackensen A and Kreutz M:
Tumor-derived lactic acid modulates dendritic cell activation and
antigen expression. Blood. 107:2013–2021. 2006. View Article : Google Scholar
|
|
123
|
Raychaudhuri D, Bhattacharya R, Sinha BP,
Liu CSC, Ghosh AR, Rahaman O, Bandopadhyay P, Sarif J, D'Rozario R,
Paul S, et al: Lactate induces pro-tumor reprogramming in
intratumoral plasmacytoid dendritic cells. Front Immunol.
10:18782019. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Dong H and Bullock TN: Metabolic
influences that regulate dendritic cell function in tumors. Front
Immunol. 5:242014. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Sonveaux P, Copetti T, De Saedeleer CJ,
Végran F, Verrax J, Kennedy KM, Moon EJ, Dhup S, Danhier P, Frérart
F, et al: Targeting the lactate transporter MCT1 in endothelial
cells inhibits lactate-induced HIF-1 activation and tumor
angiogenesis. PLoS One. 7:e334182012. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Lu H, Dalgard CL, Mohyeldin A, McFate T,
Tait AS and Verma A: Reversible inactivation of HIF-1 prolyl
hydroxylases allows cell metabolism to control basal HIF-1. J Biol
Chem. 280:41928–41939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Hirschhaeuser F, Sattler UG and
Mueller-Klieser W: Lactate: A metabolic key player in cancer.
Cancer Res. 71:6921–6925. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Végran F, Boidot R, Michiels C, Sonveaux P
and Feron O: Lactate influx through the endothelial cell
monocarboxylate transporter MCT1 supports an NF-κB/IL-8 pathway
that drives tumor angiogenesis. Cancer Res. 71:2550–2560. 2011.
View Article : Google Scholar
|
|
129
|
Lee DC, Sohn HA, Park ZY, Oh S, Kang YK,
Lee KM, Kang M, Jang YJ, Yang SJ, Hong YK, et al: A lactate-induced
response to hypoxia. Cell. 161:595–609. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zhang J, Dong C, Wu L, Chen L, Zhang L and
Shi L: Diagnostic value of a lactylation-related gene signature in
hepatocellular carcinoma. Transl Cancer Res. 14:296–312. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Luan S: The role of histone lactylation
genes in hepatocellular carcinoma prognostic models and their
immune cell infiltration features: A comprehensive analysis of
single-cell, spatial transcriptome, Mendelian randomization and
experiment. Discov Oncol. 16:292025. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Zhang Q, Cao L and Xu K: Role and
mechanism of lactylation in cancer. Zhongguo Fei Ai Za Zhi.
27:471–479. 2024.In Chinese. PubMed/NCBI
|
|
133
|
Asmamaw MD, He A, Zhang LR, Liu HM and Gao
Y: Histone deacetylase complexes: Structure, regulation and
function. Biochim Biophys Acta Rev Cancer. 1879:1891502024.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Abeywardana MY, Whedon SD, Lee K, Nam E,
Dovarganes R, DuBois-Coyne S, Haque IA, Wang ZA and Cole PA:
Multifaceted regulation of sirtuin 2 (Sirt2) deacetylase activity.
J Biol Chem. 300:1077222024. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Byun JK: Tumor lactic acid: A potential
target for cancer therapy. Arch Pharm Res. 46:90–110. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Zhao Y, Li M, Luo Z, Su M and Yu C: Recent
progress in the therapeutic modulation of lactate for cancer
treatment. Adv Ther. 6:22002542023. View Article : Google Scholar
|
|
137
|
Halford S, Veal GJ, Wedge SR, Payne GS,
Bacon CM, Sloan P, Dragoni I, Heinzmann K, Potter S, Salisbury BM,
et al: A Phase I dose-escalation study of AZD3965, an oral
monocarboxylate transporter 1 inhibitor, in patients with advanced
cancer. Clin Cancer Res. 29:1429–1439. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Benyahia Z, Blackman MCNM, Hamelin L,
Zampieri LX, Capeloa T, Bedin ML, Vazeille T, Schakman O and
Sonveaux P: In vitro and in vivo characterization of MCT1 inhibitor
AZD3965 confirms preclinical safety compatible with breast cancer
treatment. Cancers (Basel). 13:5692021. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Beloueche-Babari M, Casals Galobart T,
Delgado-Goni T, Wantuch S, Parkes HG, Tandy D, Harker JA and Leach
MO: Monocarboxylate transporter 1 blockade with AZD3965 inhibits
lipid biosynthesis and increases tumour immune cell infiltration.
Br J Cancer. 122:895–903. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Xiang F, Zhang Z, Li Y, Xia B, Liao Y,
Yang C, He Z, Lin L and Xie J: Honokiol targeting SIRT3: From
molecular mechanisms to therapeutic opportunities. FASEB J.
39:e707982025. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Gu XY, Yang JL, Lai R, Zhou ZJ, Tang D, Hu
L and Zhao LJ: Impact of lactate on immune cell function in the
tumor microenvironment: mechanisms and therapeutic perspectives.
Front Immunol. 16:15633032025. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Papadaki S and Magklara A: Regulation of
metabolic plasticity in cancer stem cells and implications in
cancer therapy. Cancers (Basel). 14:59122022. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Zang Y, Wang A, Zhang J, Xia M, Jiang Z,
Jia B, Lu C, Chen C, Wang S, Zhang Y, et al: Hypoxia promotes
histone H3K9 lactylation to enhance LAMC2 transcription in
esophageal squamous cell carcinoma. iScience. 27:1101882024.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Zhao Y, Jiang J, Zhou P, Deng K, Liu Z,
Yang M, Yang X, Li J, Li R and Xia J: H3K18 lactylation-mediated
VCAM1 expression promotes gastric cancer progression and metastasis
via AKT-mTOR-CXCL1 axis. Biochem Pharmacol. 222:1161202024.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Wang F, Chen L, Kong D, Zhang X, Xia S,
Liang B, Li Y, Zhou Y, Zhang Z, Shao J, et al: Canonical Wnt
signaling promotes HSC glycolysis and liver fibrosis through an
LDH-A/HIF-1α transcriptional complex. Hepatology. 79:606–623. 2024.
View Article : Google Scholar :
|
|
146
|
Heuser C, Renner K, Kreutz M and Gattinoni
L: Targeting lactate metabolism for cancer immunotherapy-a matter
of precision. Semin Cancer Biol. 88:32–45. 2023. View Article : Google Scholar
|
|
147
|
Li S, Lu X and Zhang L: Lactylation: the
malignant playbook of hepatocellular carcinoma cells and their
roadmap to therapy resistance. Front Immunol. 16:17524042026.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Billiard J, Dennison JB, Briand J, Annan
RS, Chai D, Colón M, Dodson CS, Gilbert SA, Greshock J, Jing J, et
al: Quinoline 3-sulfonamides inhibit lactate dehydrogenase A and
reverse aerobic glycolysis in cancer cells. Cancer Metab. 1:192013.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Fiume L, Vettraino M, Manerba M and Di
Stefano G: Inhibition of lactic dehydrogenase as a way to increase
the anti-proliferative effect of multi-targeted kinase inhibitors.
Pharmacol Res. 63:328–334. 2011. View Article : Google Scholar
|