Skeletal muscle, which constitutes 30-40% of adult
body mass and contains 50-75% of bodily proteins, is a dynamic and
plastic tissue system crucial for human physiology (1). It governs locomotive functions,
postural stability and systemic energy homeostasis while actively
participating in glucose use and lipid metabolism regulation.
Nevertheless, pathophysiological stressors, including senescence,
immobilisation, nutritional deprivation and chronic comorbidity,
can trigger the progressive degradation of muscle mass and
functional capacity, culminating in muscle atrophy (MA) (2). MA, which clinically manifests as
compromised mobility, metabolic dysregulation and elevated
susceptibility to chronic disease, substantially impairs quality of
life while imposing a heavy socioeconomic burden (3). The aforementioned factors
underscore the need for developing mechanistically targeted
interventions against this multifactorial disorder (4).
Synthetic glucocorticoids (GCs) are steroid hormones
that are used in clinical settings for the treatment of
inflammatory and autoimmune diseases due to their anti-inflammatory
and immunosuppressive effects (5). However, their prolonged use can
lead to adverse effects, one of which is MA (6). Patients typically exhibit a decline
in muscle strength, reduced exercise endurance and difficulties in
daily activities, such as climbing stairs and lifting heavy
objects, that affect their quality of life (7). Notably, GC-induced MA is one of the
most common drug-induced MA conditions, with muscle weakness
occurring in ~60% of patients with Cushing's syndrome (8). Given the notable population of GC
users and the non-negligible side effects of GCs, developing
clinical interventions targeting GC-induced MA is imperative. GCs
include ~20 common synthetic drugs, amongst which dexamethasone
(DEX) is widely used to construct MA models either through
subcutaneous injection or in vitro treatment (9,10). In addition to the stability and
reproducibility of models, DEX has a broad research base because of
its greater pharmacological potency and longer half-life compared
with other GCs (11,12). Several studies have identified
the signalling molecules and pathways involved in DEX-induced MA,
such as the Akt/mTOR pathway and mitophagy (13-15). DEX has a direct catabolic effect
on muscle by suppressing the synthesis and enhancing the breakdown
of muscle proteins. Consequently, DEX-induced MA is a crucial model
for studying the mechanisms and potential interventions of
drug-derived MA (15).
Forkhead box O (FoxO) is a key transcription factor
in mammals. It belongs to the forkhead family, which is involved in
the regulation of cell proliferation, differentiation, metabolism
and apoptosis (16). In humans,
four FoxO paralogues (FoxO1, FoxO3, FoxO4 and FoxO6) have been
identified, of which FoxO1 and FoxO3 are the most abundant and
functionally active isoforms in skeletal muscle (17). FoxO serves a key role in MA and
influences muscle mass primarily by regulating two notable protein
degradation pathways: The autophagy-lysosomal system (ALS) and the
ubiquitin-proteasome system (UPS). A growing body of evidence
indicates that DEX exacerbates MA by upregulating the activity of
FoxO, which promotes the expression of genes associated with muscle
protein breakdown (MPB) (18-20). Consequently, targeting FoxO may
be a promising strategy for developing clinical drugs to counteract
DEX-induced MA.
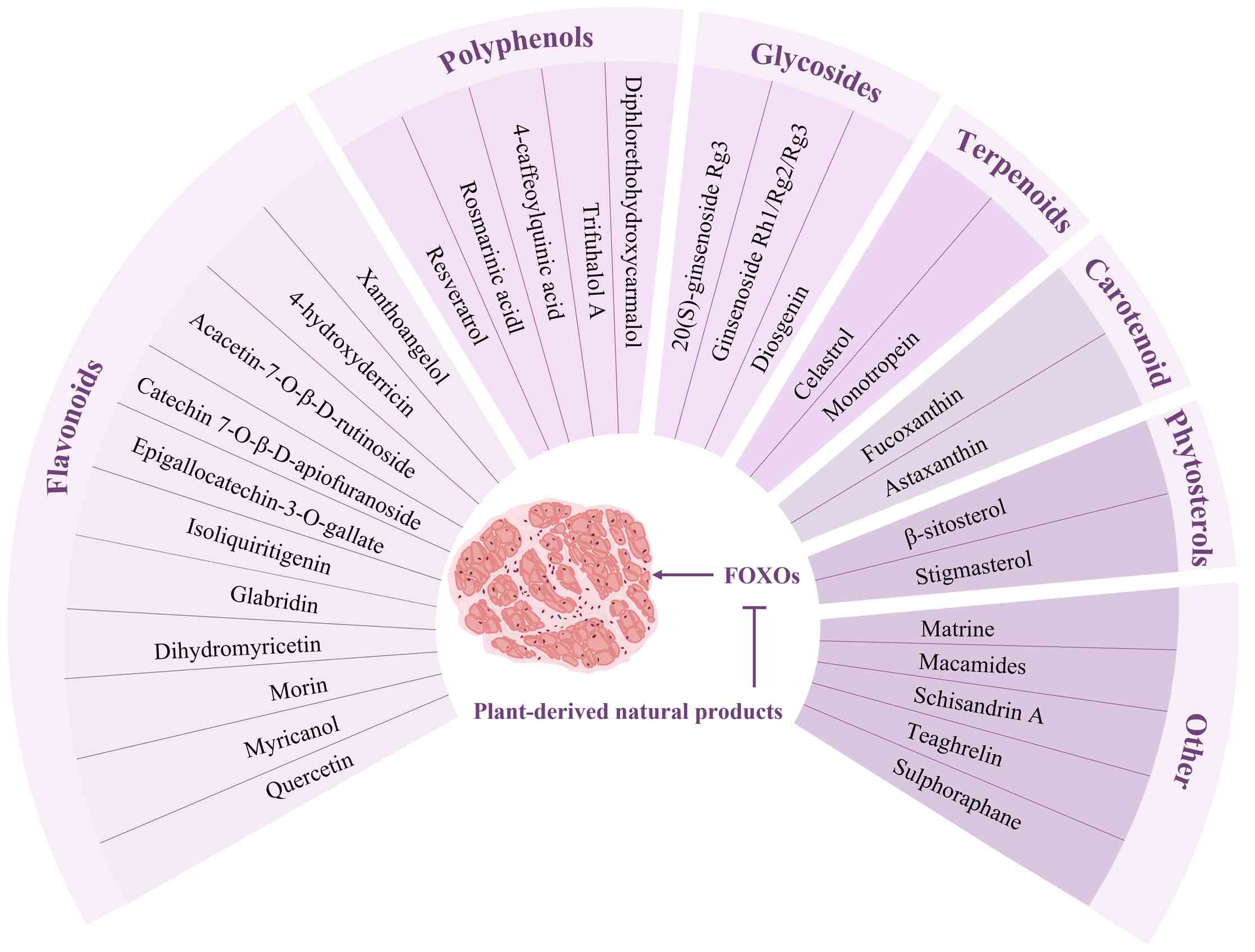
Natural products (NPs) are widely used in the design
and development of medicine for tumors, myocardial infarction, and
diabetic nephropathy) because of their diverse biological
activities and low side effects in humans (21-23). Numerous plant-derived NPs (e.g.,
ginsenosides, resveratrol, astaxanthin) markedly attenuate
proteolytic cascades in both animal and cellular models of
DEX-induced MA by modulating FoxO transcription (24-26). The present review aimed to
summarise the mechanism underlying DEX-induced MA and the key role
of FoxO and potential of plant-derived NPs to alleviate MA by
modulating FoxO activity. The present study aimed to provide a
theoretical foundation for the development of novel, safe and
effective therapeutic strategies against MA, as well as improved
clinical solutions for patients undergoing long-term GC
treatment.
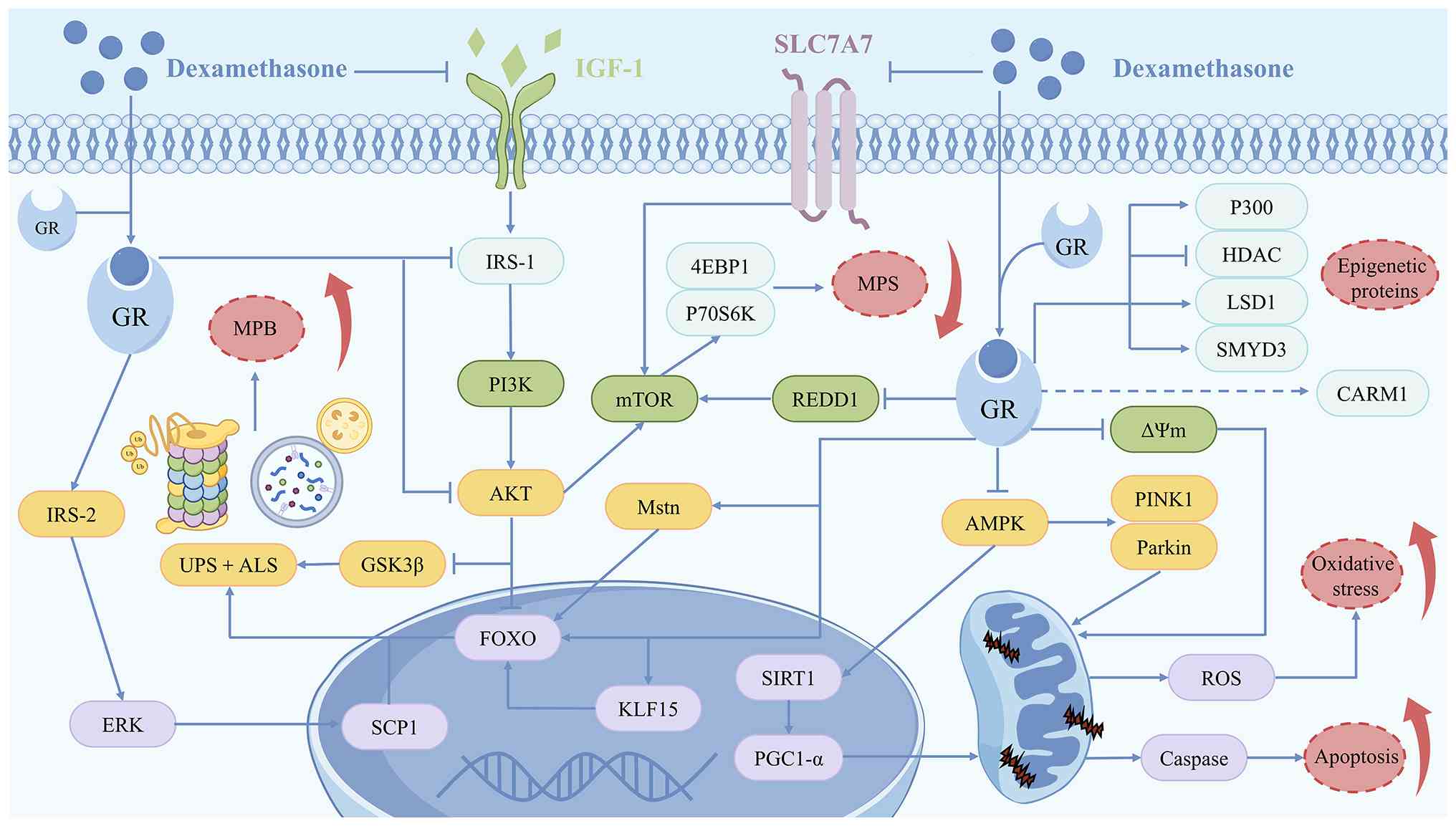
The pharmacological actions of DEX are mediated
through genomic and non-genomic mechanisms. In the canonical
genomic pathway, cytosolic glucocorticoid receptors (GRs) undergo
ligand-activated conformational changes following DEX binding,
dissociating from heat shock protein complexes and translocating to
the nucleus to modulate the transcriptional activity of target
genes via direct DNA binding or coregulator interactions (30). In parallel, non-genomic effects
are mediated by interaction with membrane proteins and alterations
in the activities of intracellular ion channels or signalling
molecules (31). These pathways
confer DEX with multifaceted therapeutic properties, such as
immunosuppression, cell metabolism regulation and oxidative stress
mitigation (32). Since its
initial synthesis in 1957 and subsequent approval by the US Food
and Drug Administration for clinical use in 1958, DEX has become a
key therapeutic agent for acute and chronic conditions (12), including cancer (33), rheumatism (34), eye disease (35) and asthma (36). Notably, its efficacy in reducing
mortality amongst critically ill patients with coronavirus disease
2019 has expanded its therapeutic applications (37). However, the prolonged use of DEX
is accompanied by numerous adverse effects, including hypertension,
gastrointestinal ulcers, hyperglycaemia, insomnia and anxiety and
MA (38). Therefore, it is
important to elucidate the mechanisms underlying GC-induced MA and
develop therapeutic strategies against this adverse effect.
GC dysregulation is a well-established driver of
skeletal MA, with clinical and experimental evidence (39-42). In 1932, Harvey Cushing documented
that endogenous GC excess caused muscle weakness in several
patients (39). Modern
interventions, including adrenalectomy, GR antagonists and
muscle-specific GR knockout models, have corroborated GC
pro-catabolic role in muscle (40,43). Chronic exogenous GC
administration induces steroid myopathy, which is clinically
characterised by progressive proximal muscle wasting, weakness and
pain (41). MA is a hallmark of
cachexia syndromes triggered by elevated cortisol levels and a
common side effect of treatment with synthetic GCs (39-41). A previous study demonstrated that
the exogenous supplementation of DEX, a typical representative of
synthetic GCs, rapidly induces MA in mice, as evidenced by weight
loss after 3 days and muscle loss after 7 days (42). Research suggests that these
phenomena may be driven by a combination of decreased muscle
glycogen reserves (44) and
increased muscle insulin resistance (45) and fat deposition (46). In addition, compared with
slow-twitch fibres, type IIB/IIB fast-twitch muscles show greater
atrophy, which is associated with their higher GR expression,
decreased myophosphorylase activity and limited capacity to use
alternative energy sources (47). DEX-induced MA is a complex
pathological process accompanied by an imbalance in the regulation
of multiple signalling molecules and pathways. DEX exacerbates the
loss of muscle mass and function by disrupting protein synthesis
and catabolism, impairing mitochondrial function and modulating the
expression of epigenetic regulatory proteins (Fig. 1). Therefore, an in-depth
understanding of these molecular mechanisms is essential for
developing effective prevention and treatment strategies.
The regulation of skeletal muscle mass depends on
the dynamic equilibrium between muscle protein synthesis (MPS) and
MPB, with MA arising when MPB persistently exceeds MPS (48). Protein synthesis encompasses
sequential steps, including amino acid transport, translation
initiation and peptide chain elongation. DEX inhibits the
expression of solute carrier family 7 member 7 transporter
proteins; this disrupts amino acid transport and translation
initiation, ultimately leading to the suppression of MPS (49). In mammals, PI3K/Akt is a major
pathway regulating muscle metabolism and promoting MPS. DEX
diminishes MPS by inhibiting Akt-mediated mTOR phosphorylation.
This effect decreases the expression of the downstream effectors
ribosomal protein 70 S6 kinase (P70S6K) and eIF4E-binding protein 1
(4EBP1) (15). Regulated in
development and DNA damage responses 1, a direct transcriptional
target of GR, regulates MPS by activating the mTOR signalling
pathway, thereby participating in the regulation of MA (50). The P13K/Akt pathway not only
activates the anabolic pathway but also effectively blocks the
catabolic pathway by inhibiting FoxO, thereby resisting DEX-induced
MA (51). Insulin-like growth
factor 1 (IGF-1) is a key upstream regulator of the PI3K/Akt
pathway, whereas DEX has a greater effect on the expression of
IGF-1 and insulin receptor substrate 1 (IRS-1) compared with other
GCs (52). DEX not only
decreases IGF-1 synthesis in muscle cells but also impairs insulin
signalling by accelerating the degradation of IRS-1 and lowering
its tyrosine phosphorylation levels (53,54). These changes also partially
explain the muscle insulin resistance that occurs during DEX
use.
The increase in MPB is a key driver of DEX-induced
MA and is primarily mediated by the activation of the UPS and ALS
(20). Ubiquitin ligases mediate
the specific degradation of target proteins by UPS through
substrate selection, including myogenic differentiation (MyoD),
myogenin (MyoG) and myosin heavy chain (MyHC), along with dynamic
modulation of ubiquitin chain formation, which drives MPB (55). Thus far, several ubiquitin
ligases mediating DEX-induced MA have been identified, including
muscle RING-finger protein 1 (MuRF1), MA F-box protein 1
(atrogin-1), casitas B-lineage lymphoma proto-oncogene B (CBL-B),
F-box and WD repeat domain-containing 7β and TNF
receptor-associated factor 6 (56-58). MuRF1 and atrogin-1 are
upregulated in MA models and are important in the study of MA
mechanisms (13,59). In DEX-induced MA, the deletion of
MuRF1 markedly protects muscle mass, whereas deletion of atrogin-1
does not show the same effect (60). FoxO is a key activator of both
ubiquitin ligases, and studies have indicated that myostatin (Mstn)
and Kruppel-like factor 15 regulate the expression of ubiquitin
ligases by acting on FoxO, thereby participating in the regulation
of MA (50,61,62). As a downstream target of the Akt
signalling pathway, glycogen synthase kinase 3 (GSK3)β not only
suppresses MPS by inhibiting the translation of eukaryotic
transcription factor 2B but also mediates the DEX-induced
expression of atrogin-1 and MuRF1, as demonstrated by knockdown
assays (63,64). Additionally, DEX activates the
ERK signalling pathway by upregulating IRS-2 expression, which
enhances the phosphorylation-dependent activity of the
transcription factor specificity protein 1, directly upregulating
ubiquitin gene expression and exacerbating MA (65). The ALS participates in MPB by
recognising and promoting the degradation of ubiquitinated proteins
in the lysosome (66). Notably,
the regulatory effect of DEX on autophagy remains controversial
(14,59,67,68). Troncoso et al (14) observed the conversion of the
autophagy marker microtubule-associated protein light chain 3
(LC3)-I into LC3-II and a decrease in the expression of the
autophagy substrate sequestosome 1 following addition of DEX to L6
and C2C12 cells; these changes are reversed by the addition of GR
small interfering (si)RNA, suggesting that DEX increases autophagic
flux through GR. In vivo and ex vivo, upregulation of
autophagy induced by the inhibition of DEX effectively alleviates
MA (67). This process is
regulated by DEX-induced changes in the phosphorylation status of
Akt and AMP-activated protein kinase (AMPK), which promote
FoxO-mediated autophagy gene expression (14,67). However, other studies have
reported that DEX administration inhibits muscle autophagic
activity; enhancing autophagy to promote the clearance of degraded
proteins contributes to the improvement of DEX-induced MA (59,68). Shen et al (59) suggested that DEX may exhibit a
dose-associated bidirectional effect in regulating autophagy. Low
doses of DEX promote autophagy through AMPK activation, whereas
high doses of DEX cause AMPK inactivation and inhibit autophagy.
However, concluding that DEX regulates autophagy in a
dose-dependent bidirectional manner may be an oversimplification.
Experimental results on autophagy changes under DEX treatment are
not entirely consistent with the aforementioned conclusion
(26,59,67-71). In in vitro studies, C2C12
cells treated with 10 μM DEX demonstrate autophagy
inhibition and plant-derived compounds (myricanol, fucoxanthin)
alleviate MA by activating autophagy (59,68,69). However, in in vivo
studies, autophagy activation is observed following treatment with
5.0 or 0.8 mg/kg DEX (26,67). Notably, similar autophagy-related
responses have been reported across studies using DEX doses that
differ by more than tenfold, suggesting autophagy regulation by DEX
may not only be dose-dependent but also affected by various
factors, such as animal strains, the in vitro and external
environment and the administration mode (26,67). To the best of our knowledge, few
studies have examined autophagy-related indicators (26,59,67-71). Therefore, further studies using
unified experimental models and standardized dosing regimens are
needed to clarify autophagy changes under different DEX doses.
Mitochondria serve as key subcellular targets for
the action of DEX, with independently expressed GRs capable of
sensing DEX and directly regulating mitochondrial DNA
transcription. This regulation affects biological processes,
including mitochondrial biogenesis, autophagy and respiratory chain
activity (72). DEX impairs
mitochondrial function through the regulation of AMPK activity,
thereby promoting MA (73).
Peroxisome proliferator-activated receptor γ coactivator 1-α
(PGC-1α), a master regulator of mitochondrial biogenesis,
demonstrates markedly decreased expression across multiple MA
models (74,75). PGC-1α overexpression in adult
muscle fibres suppresses FoxO-mediated atrogene transcription
(74). DEX inhibits
mitochondrial biogenesis and interferes with kinetic protein
expression by inhibiting AMPK and its downstream sirtuin 1 (SIRT1)
activity, impairing the PGC-1α deacetylation and its
transcriptional activity and inducing MA (59,76). In addition, the upregulation of
the AMPK-related mitochondrial autophagy factors PINK1 and parkin
is an important regulator of DEX-induced MA (76). A flow cytometric assay
demonstrated that DEX treatment notably decreases mitochondrial
membrane potential (ΔΨm) in myocytes, with ΔΨm restoration showing
therapeutic potential against MA (77). Concurrently, changes in ΔΨm are
accompanied by a marked increase in the BAX:Bcl-2 ratio and
activation of caspase cascade proteins in muscle tissue, with these
apoptotic signals exacerbating MA (77). Mitochondrial dysfunction
typically induces reactive oxygen species (ROS) accumulation,
triggering oxidative stress in DEX-treated muscle (78). Quantitative proteomics has
revealed impairment of the glutathione redox system in DEX-treated
muscle tissue, with mechanistic links to MAPK/haem oxygenase (HO)-1
pathway dysregulation (79,80).
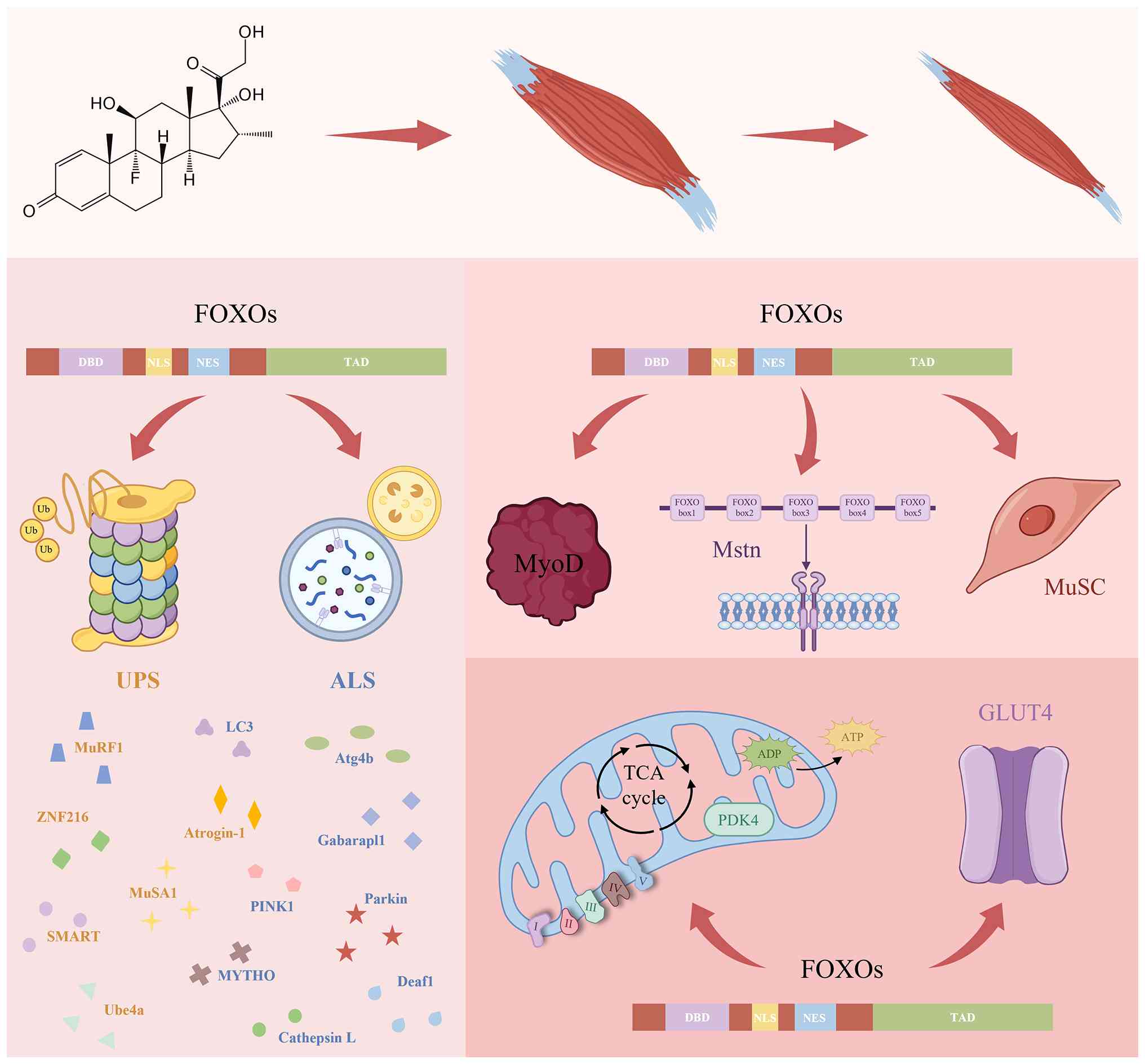
FoxO proteins are widely distributed in the
cytoplasm and nuclei of human tissue and their four isoforms
exhibit high structural similarity (97). The forkhead structural domain is
the most distinctive feature of FoxO family members. It is named
for its characteristic forked structure, which is formed by
classical α-helices and β-sheets. This domain enables FoxO to bind
DNA specifically, thereby regulating the expression of target genes
(98). The transactivation
domain (TAD), located in the C-terminus of FoxO proteins, not only
facilitates the transcription of target genes via interactions with
transcription factors or transcriptional regulators but also serves
as a key region for FoxO to undergo various posttranslational
modifications (PTMs) to regulate its activity, localisation and
stability. Additionally, each FoxO isoform contains nuclear
localisation signals (NLSs) and nuclear export signals, which
regulate the transport of FoxO proteins between the nucleus and
cytoplasm in response to intracellular and extracellular signals
(99). The high degree of
conservation of the forkhead structural domain is a key feature
that allows FoxO proteins to perform essential functions across
species. However, >70% of the FoxO region is predicted to be
disordered. In particular, the TAD is located exclusively within
intrinsically disordered regions (IDRs) (100). The IDR characteristic of TAD
confers a high degree of binding flexibility to FoxO, thereby
expanding its potential to interact with multiple coactivators and
revealing the molecular basis for isoform-specific functional
diversification (97).
In the physiological state, moderate FoxO activation
is key for maintaining homeostasis. FoxO proteins are regulators of
longevity, with their genetic association with human lifespan first
demonstrated through a genome-wide association study of Japanese
centenarians (101), which
identified FoxO3 as the only gene amongst five longevity candidates
showing a notable association with lifespan. The mechanism by which
FoxO proteins promote longevity is hypothesised to be associated
with activation of autophagy, resistance to oxidative stress and
maintenance of stem cell homeostasis (16). FoxO serves a key regulatory role
in several physiological processes, including glycolipid
metabolism, DNA repair, angiogenesis, inflammation and immune
regulation (102).
Consequently, FoxO is a promising clinical therapeutic target for a
range of human diseases, such as diabetes, cancer, cardiovascular
disease and neurodegenerative disorder (103). Although the four FoxO isoforms
originate from the same gene family, differences in their intrinsic
structures and environment-dependent mechanisms allow them to
exhibit functional differentiation in different biological
processes (97). Gene-targeting
studies have demonstrated that FoxO1 knockout impairs blood vessel
formation, FoxO3 deletion causes infertility in female mice and
FoxO6 deficiency disrupts memory consolidation (104,105). This functional differentiation
is key for the development of targeted therapeutic strategies
involving FoxO. For example, FoxO3 is implicated in slowing ageing,
whereas FoxO1 is a key target for diabetes intervention (103). MA induced by GC overdose causes
the pathologically aberrant activation of FoxOs, of which FoxO1 and
FoxO3 are the most important regulators (92,106,107).
ALS and UPS serve key roles in DEX-induced skeletal
muscle catabolism, with FoxO transcription factors serving as
master regulators of both proteolytic pathways (108). DEX-induced FoxO activation
promotes its nuclear translocation and leads to the upregulation of
ubiquitin ligase expression, which is suppressed by FoxO knockdown
(51). FoxO directly activates
atrogin-1 and MuRF1 transcription by binding multiple FoxO response
elements (FREs) in their promoter region rather than through
indirect pathways, such as affecting their translational efficiency
(19). The MuRF1 promoter
contains FREs and adjacent GREs, and their interaction constitutes
a key pathological mechanism underlying DEX-induced MA (93). Furthermore, atrogin-1 primarily
targets regulatory proteins, such as MyoD, whereas MuRF1 is
involved in the loss of MyHC induced by DEX (13,20). Notably, the FoxO regulatory
network extends to other atrophy-associated ligases, including
ubiquitination factor E4a (Ube4a) and muscle ubiquitin ligase of
the SCF complex in atrophy-1 (MUSA1). Ube4a is a U-box-type E3
ubiquitin ligase that, in addition to catalysing substrate
ubiquitination, serves as an E4 ubiquitin linker to extend the
ubiquitin chain. Its expression increases in MA, whereas this
increase is inhibited in the absence of FoxO (109). MUSA1, which is
transcriptionally regulated by Smad and FoxO, is upregulated in
various catabolic models, including MA models induced by DEX
administration, denervation and fasting (91,110). Beyond canonical E3 ligases,
Milan et al (91)
detected a ubiquitin ligase in atrophied muscle, specific of muscle
atrophy and regulated by transcription, and the expression of this
enzyme exhibits FoxO dependence. Similarly, ZNF216, an A20 zinc
finger-containing shuttle protein that directs ubiquitinated
substrates to proteasomes, is upregulated in cells treated with DEX
and transfected with constitutively active FoxO in vitro
(111). FoxO directly
upregulates the expression of macroautophagy- and mitochondrial
autophagy-associated proteins, upregulating the mRNA expression of
LC3, autophagy-related 4b (Atg4b), GABA(A) receptor-associated
protein like 1 (Gabarapl1), PINK1 and parkin (17,112). FoxO indirectly participates in
the regulation of autophagy in skeletal muscle through its
downstream transcription factors macroautophagy and youth optimizer
(MYTHO) (113) and deformed
epidermal autoregulatory factor-1 (DEAF1) (114). The knockdown of MYTHO, a
FoxO-dependent gene in muscle, causes autophagy dysfunction,
whereas the depletion of DEAF1 promotes autophagy hyperactivation.
Both of these effects lead to MA (113,114). In addition, the lysosomal
protein cathepsin L, regulated by FoxO, is a mediator of
DEX-induced MPB (115).
FoxO proteins indirectly regulate muscle growth
factors, including Myod and Mstn (116,117), through the ALS and UPS, and
directly affect their expression. FoxO exerts a dual effect on Myod
regulation. Kitamura et al (118) suggested that the loss of
function of FoxO1 removes the inhibitory effect on Notch
signalling, resulting in the upregulation of MyoD expression. Hu
et al (117) found
through in vitro electrophoretic mobility shift assays and ChIP
that FoxO3, but not FoxO1, binds two specific FREs in the Myod gene
and activates Myod transcription by recruiting RNA pol II. Negative
regulation by FoxO1 and direct activation by FoxO3 may reflect the
regulation of MyoD expression by FoxO at different stages of
differentiation or within specific microsignalling environments.
Notably, in cachexia-induced MA, the blockade of FoxO expression
concurrently elevates MyoD and decreases Mstn levels (94). The presence of five
FoxO1-specific binding sites within Mstn mediates its
transcriptional activation (116), and previous studies have
revealed a mechanism by which Mstn drives MA by enhancing FoxO
activity (119,120), suggesting that the a
bidirectional positive feedback regulation process that amplifies
muscle catabolic signalling. Furthermore, muscle satellite cells
(MuSCs) serve a critical role in muscle injury, repair and
regeneration by proliferating and differentiating into mature
muscle cells to maintain muscle structure and function. FoxO1 and
FoxO3 coordinate MuSC quiescence by upregulating
p27Kip1, a cell cycle inhibitor. Targeted FoxO
attenuation restores MuSC proliferative capacity, thereby offering
therapeutic potential for muscle regeneration and atrophy
prevention (121,122).
FoxO transcription factors serve as key regulators
of skeletal muscle energy homeostasis by coordinating mitochondrial
biogenesis and glycolipid metabolism (17). RNA sequencing shows that
following the dual knockdown of muscle insulin and IGF-1 receptors,
most of the altered genes, especially those associated with
mitochondrial oxidative phosphorylation (OXPHOS) and tricarboxylic
acid cycle, are dependent on the regulation of FoxO (123). Mechanistically, FoxO impairs
mitochondrial respiratory chain activity via the selective
suppression of complex I subunit expression (124). In ageing skeletal muscle and
amyotrophic lateral sclerosis models, FoxO gene deletion restores
OXPHOS capacity, thereby promoting ATP synthesis (109,125). In addition, FoxO promotes the
conversion of muscle metabolic substrates from glucose to lipids by
regulating pyruvate dehydrogenase kinase 4 (PDK4), lipoprotein
lipase and fatty acid translocase CD36 in skeletal muscle (126,127). PDK4, the most abundantly
expressed isoform of PDK in skeletal muscle, inhibits pyruvate
dehydrogenase complex activity via phosphorylation in the
mitochondrial matrix, thereby suppressing glucose oxidation. FoxO1
binds to the PDK4 promoter region to regulate energy under
starvation conditions (128).
Notably, PDK4 promotes the ubiquitinated degradation of atrogin-1
by enhancing MyoG phosphorylation, which promotes DEX-induced MA
(129). FoxO1 impairs systemic
glucose homeostasis and promotes muscle wasting through the
downregulation of glucose transporter type 4 expression in skeletal
muscle (130,131) (Fig. 2).
FoxO activity regulation involves a network of PTMs
and protein-protein interactions, resulting in precise functional
regulation at the levels of gene transcription and protein
activity. The phosphorylation, acetylation and ubiquitination of
FoxO are notable post-translational modifications involved in the
regulation of MA, and methylation is also a regulatory mechanism
associated with MA pathogenesis (132,133). Changes in the levels of
multiple PTMs of FoxO are observed in mouse and human myotubular
cells following DEX treatment, suggesting DEX-induced MA may be
partially mediated by altering the modification pattern of FoxO
(Table I) (134). Although previous reviews have
addressed FoxO regulatory mechanisms (102,103), the present review specifically
focuses on MA-associated pathways and proposes targeting upstream
regulators of FoxO signalling as a complementary approach to direct
FoxO modulation for MA intervention.
In DEX-treated muscle, MA-related transcription
factors (FoxO and NF-κB) are acetylated (143). The acetylation site of FoxO is
primarily located in the forkhead structural domain, regulating its
transcriptional activity by affecting the affinity of FoxO for DNA.
FoxO acetylation exerts precise and complex effects on
transcriptional activity, with activation or suppression dictated
by cell type and micro-environment (144). The acetyltransferase P300 was
identified as a FoxO regulator in skeletal muscle by Senf et
al (145); transfection of
a P300-deficient plasmid into rat skeletal muscle leads to a
4.6-fold increase in overall FoxO activity, with a 16-fold increase
in FoxO3-specific activity. In a DEX-induced MA model, P300
overexpression inhibits the transcriptional activity of FoxO and
differentially regulates the expression of its downstream target
genes, providing a potential strategy for the treatment of MA
(146). Mutants of FoxO3 have
been used to verify that P300-mediated Lys262 acetylation is key
for regulating FoxO3 subcellular localisation and transcriptional
activity (146). HDAC1 is also
a key factor regulating FoxO activity in muscle. HDAC1
overexpression (via plasmid delivery) or HDAC1 suppression (by
trichostatin A) demonstrate that the deacetylation function of
HDAC1 increases the activity of FoxO, its nuclear localisation, and
the transcription of atrophy-related target genes. IP experiments
with anti-acetylated lysine antibodies further demonstrate that
FoxO is a direct target of HDAC1 deacetylation activity in muscle
(147). SIRT1 is a class III
HDAC and can dually regulate FoxO through deacetylation. During MA,
SIRT1 directly inhibits FoxO transcriptional activity through
deacetylation, a finding confirmed by luciferase reporter gene
assay and ChIP (148).
Protein arginine methyltransferases (PRMTs) serve a
key role in MuSC function and phenotypical adaptive remodelling
(149). In mice, the
muscle-specific depletion of PRMT1 results in decreased muscle mass
and muscle weakness, accompanied by elevated levels of FoxO3
protein (133). While PRMT1
does not directly methylate FoxO3, its depletion triggers the
compensatory upregulation of PRMT6 (133). PRMT6 catalyses asymmetric
dimethylation at FoxO3 Arg188/Arg249 residues, inducing
transcriptional hyperactivation that exacerbates autophagy and
proteolysis, revealing the molecular mechanism by which PRMT1
deletion triggers MA (133).
FoxO activity is dynamically regulated by direct
interaction with various proteins; such regulation influences
muscle cell adaptability to atrophy-inducing stimuli. PGC-1α
promotes the conversion of myofibrils into an oxidative state in
skeletal muscle by regulating mitochondrial function and oxidative
metabolism, thereby affecting muscle size and resistance to atrophy
(74). The transfection of
persistently activated FoxO3 mutants into mouse tibialis anterior
muscle results in a marked decrease in muscle fibre cross-sectional
area (CSA) and PGC-1α co-expression abolishes this atrophy
(74). PGC-1α-mediated
protection occurs via the suppression of FoxO3-driven ubiquitin
ligase induction, independent of FoxO3 expression or
phosphorylation changes (74,150). By contrast, Smad not only
enhances the binding of FoxO3 to the MuRF1 promoter region, thereby
promoting the transcriptional activity of MuRF1, but also
dose-dependently increases FoxO3 protein levels by binding to
specific DNA sequences in MuRF1 (151). Therefore, the combined action
of Smad with FoxO may be required for the optimal activation of
ubiquitin ligases. However, FoxO and Smad act independently in the
regulation of Mstn expression and myoblast differentiation
(116).
FoxO activation drives MA through multiple
signalling pathways. DEX not only directly activates FoxO but also
regulates FoxO activity by affecting FoxO PTMs and protein
interactions. Notably, plant-derived NPs have garnered attention
because of their biological activities, particularly in terms of
anti-inflammatory, antioxidant, antiviral and immunomodulatory
aspects (21-23). At the same time, their safety
profile and low cost make them valuable for long-term treatment.
Numerous plant-derived NPs alleviate DEX-induced MA, with these
beneficial effects associated with FoxO activity modulation
(24-26) (Fig. 3; Table II).
Quercetin is a naturally occurring flavonoid found
in plants and is widely distributed in many vegetables and fruits,
such as onions, apples and asparagus. It accounts for >50% of
dietary flavonoid intake (152). Quercetin is a
3,3',4',5,7-pentahydroxyflavone featuring five phenolic hydroxyl
groups and a conjugated double bond in its molecular structure,
both of which confer powerful antioxidant activity (153). In addition, its
anti-inflammatory, antiviral, anticancer and antimicrobial
properties have led to a wide range of applications in dietary
supplements and pharmaceutical research (154). Quercetin ameliorates
DEX-induced C2C12 cell damage by scavenging ROS and restoring Δψm,
highlighting its potential against MA (77). Glycosylated derivatives are the
primary form of quercetin in plants, with rutin (quercetin
3-O-β-glucuronide) being the most abundant (155). In mice, rutin restores the
DEX-induced weight loss of the gastrocnemius, tibialis anterior and
extensor digitorum longus muscles by decreasing FoxO3 expression,
thereby suppressing atrogin-1 and MuRF1-mediated protein
degradation (106). In an ex
vivo model, the ethanolic extract of Ricinus communis
leaves alleviates DEX-induced MA by reducing oxidative stress and
enhancing FoxO3 phosphorylation, which inhibits UPS-mediated MPB
(156). Liquid
chromatography-tandem mass spectrometry reveals that the main
active component of the extract is rutin (156). In addition, the primary active
ingredient of the aqueous extract of lotus leaf is a quercetin
derivative, quercetin 3-O-β-glucosiduronic acid, which inhibits the
expression of FoxO3 and its downstream atrophy-related genes by
affecting the phosphorylation levels of AMPK and Akt, thereby
increasing muscle mass in DEX-induced MA mice (157). In mice, an isoquercetin
glycoside prepared from quercetin glycoside and bittersweet
ameliorates the DEX-induced decrease in the gastrocnemius muscle
wet weight-to-body weight ratio, as well as increasing Akt
phosphorylation levels and decreasing FoxO1 content on the third
day of administration (158).
Myricanol, a diarylheptanoid derived from parts of
Myrica species (such as Myrica rubra bark), is a plant secondary
metabolite. Although research on myricanol is limited, studies have
observed multiple biological effects, including anti-inflammatory,
antioxidant, apoptosis-inducing, ferroptosis-inhibiting and
neuroprotective activity, in models of diseases, such as lung
cancer, Alzheimer's disease and renal fibrosis (159-161). Myricetin is an AMPK activator.
In zebrafish fed a high-fat diet, it effectively inhibits lipid
accumulation by binding to the AMPK γ subunit (162). Shen et al (59) identified myricanol as a SIRT1
activator that directly inhibits downstream FoxO3 transcription by
enhancing SIRT1 deacetylase activity, thereby alleviating
DEX-induced MA and weakness. SIRT1 activation also improves
autophagy and apoptosis imbalance and restores mitochondrial
content/function in myotubes via the PGC-1α/FoxO3 and Akt/FoxO3
pathways (59). Given that AMPK
and SIRT1 are central regulators of energy metabolism (163), myricanol is hypothesised to
play a critical role in mitigating muscle metabolic dysfunction.
Metabolomics analysis reveals that myricanol notably rectifies
DEX-induced disturbances in glucose, lipid and protein metabolism,
consequently reversing muscle mass loss and strength decline in
mice (164). This effect may
involve irisin secretion from myotubes, which enhances metabolic
homeostasis and myogenesis (165). Recent evidence indicates that
myricanol ameliorates ageing-related sarcopenia by decreasing
oxidative stress (166),
solidifying its status as a clinically promising candidate for
regulating muscle metabolism and combating MA.
Morin, a natural pigment extracted from mulberry
plants and traditional Chinese herbs, is particularly abundant in
mulberry leaves. It has notable efficacy in the prevention and
treatment of organ damage, neurodegenerative disease, cancer,
diabetes and arthritis, primarily because of its potent antioxidant
and anti-inflammatory properties (167). Morin effectively counteracts
DEX-induced MA in vitro, as evidenced by increases in
myotube thickness and diameter and MyHC expression (78). Moreover, it exerts antioxidant
effects by inhibiting the DEX-induced upregulation of p66Shc (a
redox protein) and enhancing the expression of antioxidant enzymes,
such as SOD1 and catalase (78).
It decreases the expression of downstream atrogenes by elevating
the levels of PGC-1α and phosphorylated FoxO3 within myotubes,
thereby mitigating MPB (78).
Liquorice is one of the most widely used herbal
medicines globally and is employed in traditional Chinese medicine
to relieve pain, improve digestive function and alleviate cough.
Glabridin, a natural isoflavonoid isolated from liquorice root
extract, constitutes 0.08-0.35% of its dry weight (175). Glabridin has garnered notable
attention in the cosmetics industry, accounting for 70% of patents
in this field (176,177). As a potent tyrosinase
inhibitor, glabridin suppresses melanogenesis, thereby exhibiting
depigmentation efficacy (178).
Furthermore, its antioxidant, anti-inflammatory, anticancer and
antimicrobial activities have been extensively reported, providing
avenues for developing drugs for treating obesity, diabetes and
malignancy (179,180). Notably, glabridin serves as a
phyto-oestrogen with structural and lipophilic properties analogous
to those of oestradiol, suggesting its key regulatory role in bone
health (181). Glabridin
notably decreases MPB-related MuRF1 and CBL-B expression in C2C12
cells and mouse tibialis anterior muscle by inhibiting DEX binding
to GR, blocking GR nuclear translocation and suppressing FoxO3
phosphorylation (58). Moreover,
while changes in p38 phosphorylation are observed, siRNA
interference reveals that p38 is not involved in the regulation of
DEX-related MA (58).
Isoliquiritigenin is a natural chalcone flavonoid
primarily isolated from liquorice root. This compound exhibits
notable anti-inflammatory and antioxidant properties (182,183). By modulating multiple
signalling pathways, including the Nrf2/HO-1, MAPK and SIRT1
pathways, it exerts notable protective effects in disease models,
such as myocardial ischaemic injury, diabetic cardiomyopathy, acute
kidney injury and Parkinson's disease (182,184). Isoliquiritigenin has been shown
to activate the transcription factors FoxO and Nrf2, which are key
regulators of cell antioxidant responses (185,186). In skeletal muscle,
isoliquiritigenin has also been shown to ameliorate MA by targeting
FoxO. In DEX-treated C2C12 cells and mice, isoliquiritigenin
supplementation alleviates atrophic changes; specifically, it
improves grip strength and exercise endurance in mice and reduces
the expression of atrophy-related genes in both models (187). Isoliquiritigenin activates the
Akt/mTOR signalling pathway, enhances FoxO1 and FoxO3
phosphorylation and thereby inhibits MPB and promotes MPS (187).
EGCG is the most potent catechin derivative in
green tea extract, where it accounts for 50-80% of the bioactive
components (188). Its 21
hydroxyl groups and stereochemical structure confer potent
antioxidant activity, which manifests as chelation of metal ions
and neutralisation of free radicals in redox reactions (189). Compared with other catechins,
EGCG demonstrates superior biological activity, including
anti-inflammatory, antiviral, antimicrobial and anticancer
properties, positioning it as a promising therapeutic agent for
chronic diseases, such as obesity, diabetes and cardiovascular
disease (190,191). Its antitumor efficacy has
progressed to clinical trials across multiple cancer types,
including breast and lung cancers (192,193), with the 67 kDa laminin receptor
identified as the primary mediator of its anticancer activity
(194). Notably, in C2C12
cells, EGCG counteracts the DEX-induced upregulation of FoxO3 and
MuRF1 via the aforementioned receptor, as evidenced by the
unaltered expression observed following siRNA-mediated receptor
silencing (195). High
concentrations of EGCG promote the phosphorylation of FoxO3 via the
activation of the PI3K/Akt signalling pathway, thereby inhibiting
FoxO activation and decreasing MPB (196). Although EGCG exerts regulatory
effects on multiple forms of MA through the modulation of the
NF-κB, MAPK and PI3K/Akt signalling pathways (197), the molecular mechanisms
underlying its protective effects against DEX-induced MA remain
largely unexplored.
AR, also known as linarin, is a flavonoid glycoside
abundantly found in plants of the Asteraceae, Lamiaceae and
Scrophulariaceae families. To the best of our knowledge, research
on AR remains limited, with its anti-inflammatory, antioxidant,
sedative and anti-osteoporotic activities preliminarily
characterised in animal models or in vitro studies, with
none having advanced to clinical investigation (202,203). Notably, molecular docking
simulation has revealed that the hydroxyl groups of AR forms robust
hydrogen bonding interactions with multiple residues of
acetylcholinesterase (AChE), suggesting its potential as an AChE
inhibitor and therapeutic candidate for Alzheimer's disease
(204). The traditional herb
Chrysanthemum zawadskii Herbich (CZH) exerts protective
effects against DEX-induced MA by suppressing GR nuclear
translocation, modulating the Akt/mTOR and FoxO3 pathways and
downregulating atrogenes (205). As the most abundant bioactive
constituent in CZH, AR partially mediates the aforementioned
antiatrophic effects through upregulating MyoD, MyoG and MyHC
expression, coupled with enhancing mitochondrial respiration in
myocytes (205).
4-HD and XAG are the two primary chalcone-derived
bioactive compounds isolated from Angelica keiskei (AK). Chalcones
are α,β-unsaturated carbonyl compounds and serve as a biological
precursor of plant flavonoids (206). Studies have characterised their
anti-inflammatory properties and regulatory effects on
glucose-lipid metabolism, highlighting their therapeutic potential
in diabetes and obesity management (207-209). A 12-week randomised
double-blind trial in obese populations demonstrated that 4-HD and
XAG supplementation decrease visceral fat area and waist
circumference in overweight individuals (210). Yoshioka et al (211) revealed that prolonged and acute
treatment with 4-HD/XAG attenuates DEX-induced MA by inhibiting GR
nuclear translocation, suppressing FoxO3 activity and
downregulating ubiquitin ligase expression. Changes in the
phosphorylation of p38 following 4-HD and XAG intervention are not
involved in the regulation of MA (211). However, in C2C12 cells,
treatment with 4-HD alone enhances myogenesis by activating the p38
signalling pathway (212).
Additionally, the ethanolic extract of AK partially ameliorates
DEX-induced muscle strength loss and CSA decreases, with improved
effects when combined with Morus alba L. extract (141). These findings collectively
suggest that the antiatrophic properties of AK are associated with
its primary bioactive constituents 4-HD and XAG (Fig. 4).
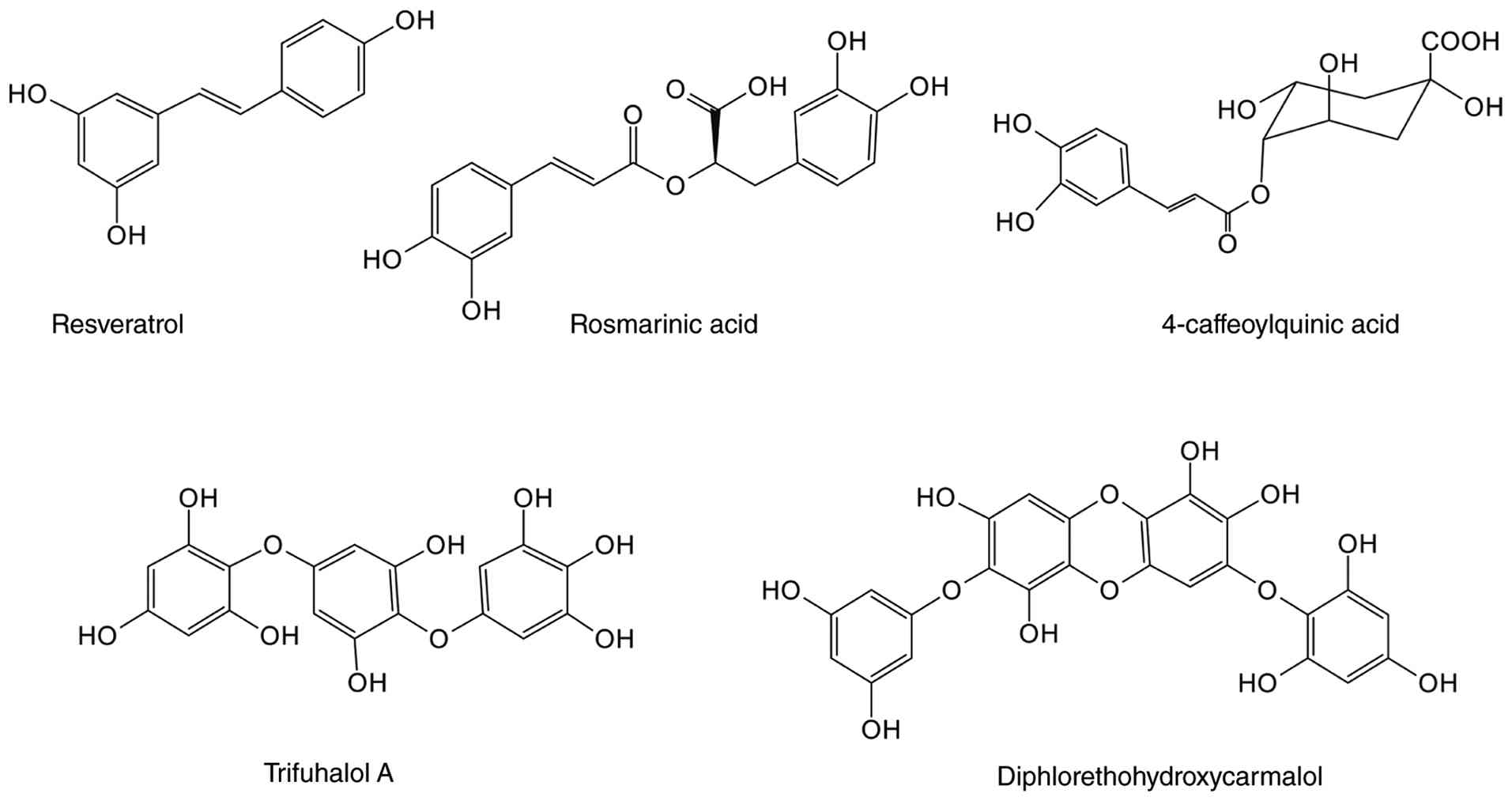
Resveratrol, one of the most widely studied
polyphenols, is produced by a range of plants in response to
pathogen attack and participates in plant self-defence responses as
an antitoxin (213,214). It is extracted from >70
plant species and is found at particularly high concentrations in
grape skin and wine products (215). Evidence from human clinical
trials has demonstrated resveratrol therapeutic potential across
pathologies, such as Alzheimer's disease, knee osteoarthritis, and
diabetic kidney disease, attributable to its pleiotropic biological
activity, including antioxidant, anti-inflammatory, antitumor,
antiproliferative and antiviral properties (216,217). Howitz (218) found that resveratrol mimics the
effects of caloric restriction by activating SIRT1, thereby
decelerating ageing. This promoted resveratrol in ageing research
and facilitated its commercialisation in nutraceutical formulations
(219,220). Although conclusive evidence
regarding its ability to extend human lifespan remains elusive, its
role as a potent SIRT1 agonist has been widely validated (216,218). It has demonstrated notable
efficacy in MA treatment through SIRT1 targeting (221-224). Resveratrol alleviates MA
induced by chronic kidney disease (221), ageing (222), cancer (223) and MAFLD (224) by modulating the SIRT1/FoxO1 and
AMPK/SIRT1 pathways. Experimental evidence from DEX-treated L6
myotubes reveals that resveratrol suppresses FoxO1 acetylation via
SIRT1 activation, consequently downregulating atrogenes, including
atrogin-1 and MuRF1, at the transcriptional level (25). The introduction of SIRT1 siRNA
further demonstrated that the inhibitory effect of resveratrol on
DEX-induced atrogene expression in myotubes is SIRT1-dependent
(25). Liu et al
(73) demonstrated that the
resveratrol targeting of AMPK/FoxO3 signalling not only restores
DEX-impaired mitochondrial respiration but also suppresses
ubiquitin ligase expression, ultimately reversing skeletal muscle
mass reduction in mice. Furthermore, resveratrol exhibits notable
myoprotective effects through the suppression of inflammatory
cascades, oxidative insult and insulin resistance (225), making it a commonly used herb
in MA treatment.
RA is a water-soluble phenolic acid compound found
in culinary herbs, such as rosemary, perilla, mint, sage and other
labiate plants (226). It
exhibits pharmacological effects similar to those of other
plant-derived NPs. These effects encompass anti-inflammatory,
antioxidant, antibacterial, anticancer and immunomodulatory
properties and confer potential for RA application in the
pharmaceutical, food and cosmetic industries (227). Clinical evidence highlights its
beneficial effects in ameliorating arthritis, cognitive impairment,
allergic disorder, dermatological conditions and metabolic syndrome
manifestations (228).
Ethanolic extract of the traditional herb Salvia plebeia R.
Br. is effective in reversing DEX-induced cell viability loss and
atrogene upregulation, thereby attenuating MA progression.
Centrifugal partition chromatography and high-performance liquid
chromatography analyses have identified RA as a major active
component in this extract (69).
The coadministration of DEX with RA mitigates myotube cytotoxicity
and atrophy (69). This effect
is mechanistically attributed to the RA-mediated activation of the
Akt/mTOR/p70S6K signalling pathway, suppression of ROS generation
and apoptotic pathways, inhibition of FoxO3-mediated ubiquitin
ligase expression, enhancement of autophagic flux and restoration
of mitochondrial content and functionality (69).
4-CQA, a naturally occurring phenolic acid
derivative, is synthesised by esterification between caffeic acid
and the C4 hydroxyl group of quinic acid. This compound is
abundantly distributed in Asteraceae and Lamiaceae
species, where it exerts anti-inflammatory and antioxidant effects
in conjunction with other quinic acid derivatives (229). 4-CQA is the predominant
bioactive constituent in Peucedanum japonicum Thunb.,
demonstrating notable therapeutic potential against DEX-induced MA
(230). In C2C12 myoblasts,
4-CQA regulates muscle protein metabolism by inhibiting the nuclear
translocation of GR and FoxO3 while simultaneously activating the
mTOR pathway, thereby promoting an increase in myotube diameter
(230).
Trifuhalol A is a phloroglucinol compound primarily
derived from the ethyl acetate extract of the edible brown alga
Agarum cribrosum. By contrast with polyphenols from
terrestrial plants, those from brown algae are polymers of
phloroglucinol (1,3,5-trihydroxybenzene) monomers, endowing them
with unique chemical structures and diverse biological activities
(231). As a representative
monomer, trifuhalol A has a low molecular weight and a simple
structure but notable pharmacological activity, including
anti-inflammatory, antiadipogenic and antidiabetic effects
(232-234). Yang et al (235) reported the protective role of
trifuhalol A in MA. In C2C12 cells and a zebrafish model,
trifuhalol A notably improves myotube diameter and fusion index and
increases muscle fibre CSA while also upregulating the locomotor
capacity of zebrafish. Mechanistically, trifuhalol A inhibits
protein degradation by suppressing the transcriptional activity of
FoxO3, thereby downregulating the expression of its downstream
targets MuRF1 and atrogin-1 (235). Concurrently, it activates the
Akt/mTOR pathway and upregulates the expression of molecules such
as MyoD, promoting MPS (235).
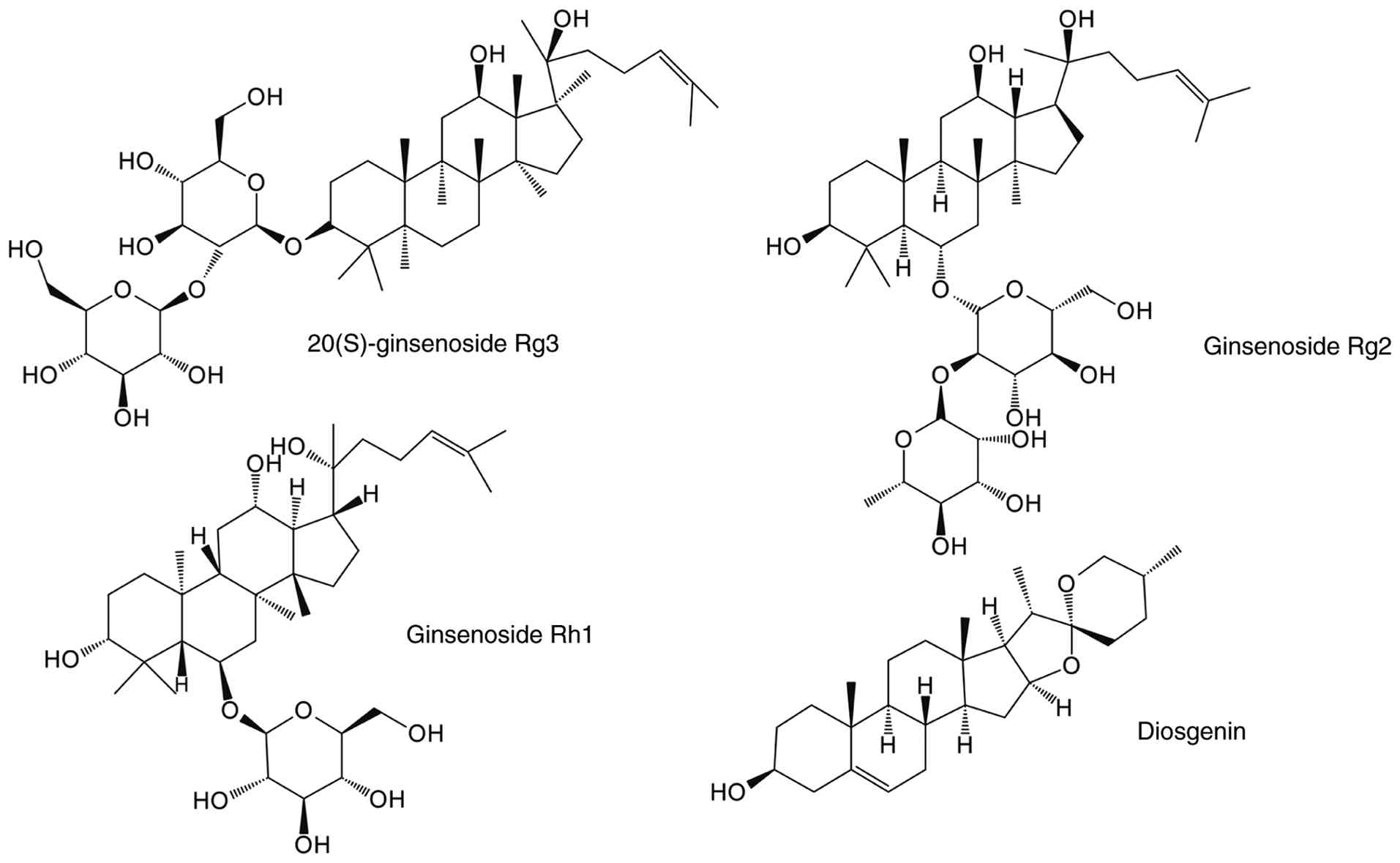
As members of the triterpenoid saponin family,
ginsenosides are predominantly derived from plants of the genus
Panax, which is used in traditional medicinal. To date, ~300
distinct ginsenosides have been isolated (241). They are classified into two
primary categories on the basis of their aglycone carbon skeletons:
Tetracyclic dammarane and pentacyclic oleanane types, with the
former including common ginsenosides, such as Rb, Rg and Re.
Ginsenosides have received extensive attention in pharmacology, as
exemplified by Rg3, which achieved regulatory approval by National
Medical Products Administration (NPMA) in 2000 as the first
monomeric antitumor drug derived from traditional Chinese medicine
(242,243). They demonstrate multifaceted
therapeutic properties through the modulation of key biological
processes, including cellular proliferation, apoptosis, oxidative
stress, inflammation and immune regulation, thereby exhibiting
clinical relevance in neurodegenerative disorder (244), cardiovascular diseases
(245), ageing (246) and metabolic syndrome (247). Zha et al (248) reported that ginsenosides
possess therapeutic potential for the treatment of sarcopenia by
promoting muscle repair and growth: Rh1, Rg1, Rg2, Rg3 and Rg5
improve DEX-induced MA (24,249-252). Mechanistic studies have
revealed that 20(S)-ginsenoside Rg3 (S-Rg3) substantially
attenuates mitochondrial structural abnormality and respiratory
dysfunction in a DEX-treated C2C12 in vitro atrophy model
via the modulation of the AMPK/FoxO3 signalling pathway,
concurrently suppressing atrogene and apoptotic protein expression
(251). S-Rg3 enhances C2C12
myoblast differentiation by regulating the Akt/mTOR/FoxO3 pathway
(251). This has been validated
by the use of the Akt inhibitor GDC-0068 and is consistent with the
findings of Men et al (24,249). Men et al (249) further demonstrated that
compared with DEX, Rh1, Rg2 and Rg3 administration notably improves
mitochondrial biogenesis through SIRT1/PGC-1α pathway
activation.
Diosgenin, a phytosteroidal saponin, is
predominantly found in fenugreek seeds and wild yam rhizomes. Its
medicinal value is derived from its hypoglycaemic, hypolipidaemic,
anti-inflammatory, antioxidant and immunomodulatory activity, which
show notable therapeutic potential in cancer, metabolic and
inflammatory disease (253).
Serving as a precursor for steroid hormone synthesis, diosgenin is
used in the production of hormone drugs via chemical conversion
into corticosteroids, oestrogens and androgens (254). As a steroid analogue molecule,
diosgenin competitively inhibits DEX and GR binding, thereby
effectively mitigating DEX-induced MA (255). The antiatrophic mechanisms of
diosgenin involve the upregulation of MyHC expression and
suppression of ubiquitin-proteasome activity and are mediated by
intracellular signalling alterations associated with GR nuclear
translocation inhibition, including the modulation of Akt
activation, FoxO3 Ser253 phosphorylation and P70S6K phosphorylation
levels (255) (Fig. 6).
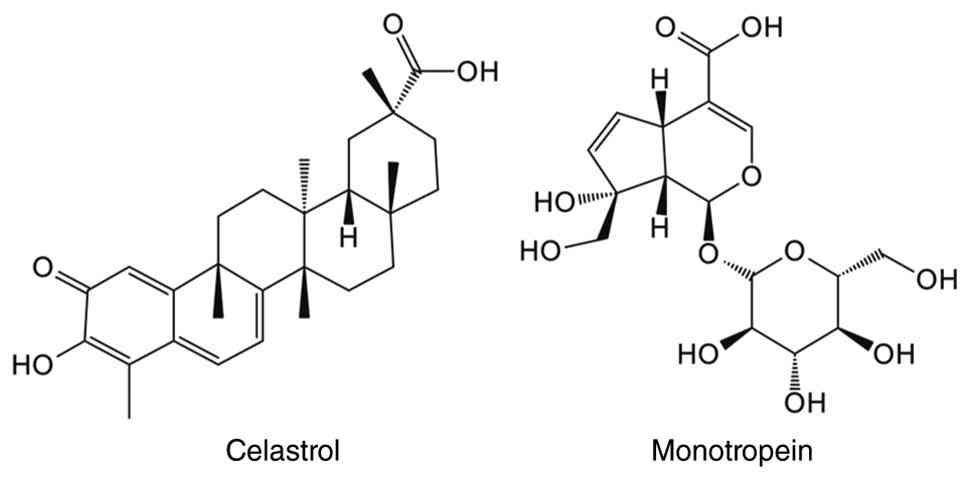
Celastrol, a pentacyclic triterpenoid predominantly
isolated from the root extract of Tripterygium wilfordii,
demonstrates pharmacological activities that are intrinsically
linked to its functional groups, notably its C-20 carboxyl, quinone
methide and C-3 hydroxyl groups. These structural features directly
mediate its antimicrobial, antiproliferative, antineoplastic and
cytotoxic properties (256).
Celastrol exhibits broad-spectrum protective effects in in
vitro and in vivo models of inflammatory disorder,
obesity, diabetes mellitus and neurodegenerative diseases,
establishing its status as a promising therapeutic candidate
(257,258). Research has indicated that
celastrol inhibits GR activity and promotes GR degradation by
enhancing the interaction between GR and Ube3a (259,260). This molecular mechanism
underpins its efficacy in alleviating DEX-induced MA in C2C12
myotubes. Celastrol modulates the DEX-induced downregulation of
FoxO3 phosphorylation via HSP70 activation, concurrently
suppressing MuRF1 expression and 26S proteasome overactivation
(261). Pharmacological
inhibition using tanespimycin (an HSP90 inhibitor) and U0126 (an
ERK1/2 inhibitor) have revealed that the Akt/mTOR and ERK1/2
signalling pathways mediate celastrol anti-MA effects through the
regulation of FoxO3 phosphorylation status (261). Celastrol exerts protective
effects in an in vitro model of MA induced by doxorubicin
(262).
Monotropein is a natural iridoid glycoside that is
abundantly found in the root, stems and leaves of Morinda
officinalis How (263).
NF-κB is the most extensively studied downstream target of
monotropein (264,265). Molecular docking studies have
revealed that monotropein binds NF-κB, thereby inhibiting its
DNA-binding activity and decreasing the expression of inflammatory
factors (264,265). This anti-inflammatory action,
combined with antioxidant effects, suggests monotropein may have
therapeutic efficacy in osteoporosis, acute organ injury, asthma
and renal cancer (266). Its
antiosteoporotic effects involve oxidative stress suppression
through the activation of the Akt/mTOR pathway, which contributes
to MA mitigation (267,268). By modifying FoxO3
phosphorylation, monotropein-activated Akt/mTOR signalling
suppresses atrogene upregulation and enhances MyHC expression in
DEX-treated cells and animal (267). Therefore, focusing on the
Akt/mTOR pathway may provide a molecular basis for expanding the
pharmacological activity of monotropein and its potential clinical
application (Fig. 7).
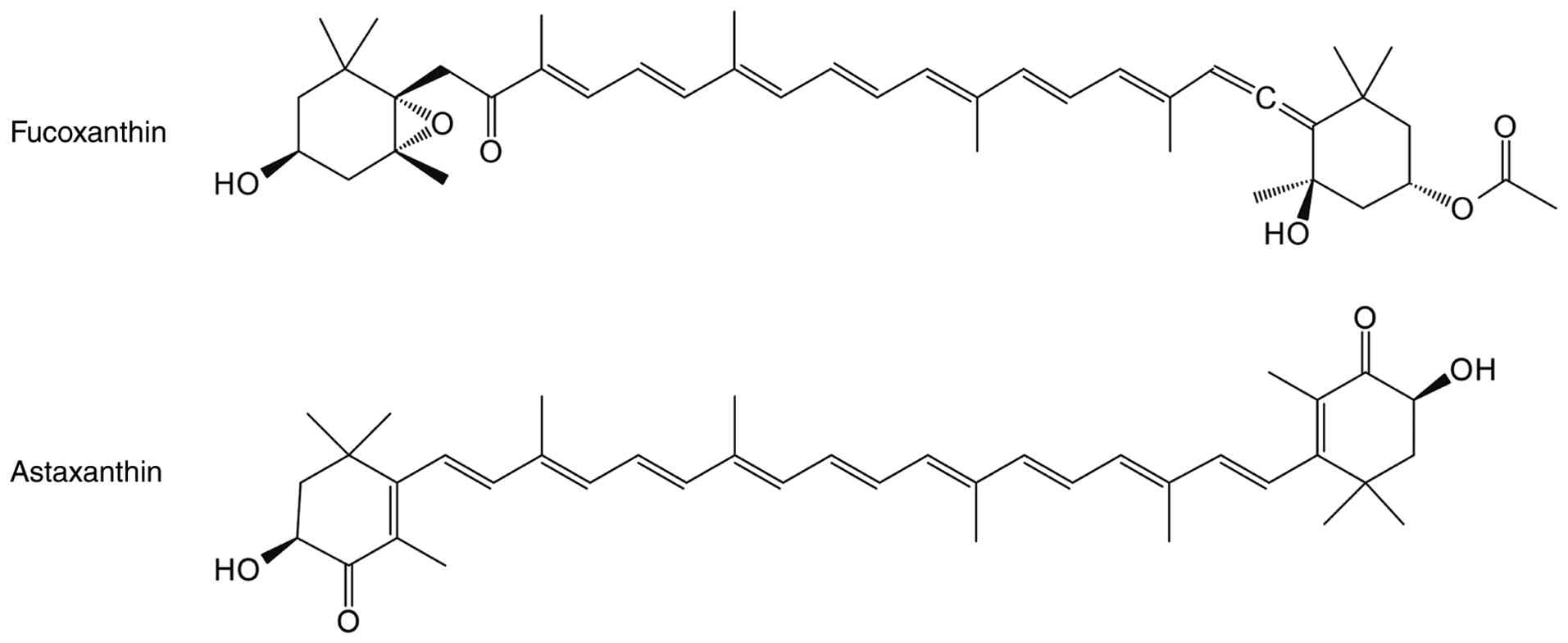
Fucoxanthin is abundantly distributed in algal
species, particularly brown algae, accounting for ~10% of all
naturally occurring carotenoids (269). As the primary photosynthetic
pigment in these organisms, fucoxanthin serves as a key component
of light-harvesting complexes, facilitating photon capture and
energy transfer (270).
Fucoxanthin is the first naturally isolated carotenoid containing
an allenic bond, a structure that is hypothesised to be the basis
for its multiple pharmacological effects, including anticancer,
anti-obesity, antibacterial and intestinal flora regulation effects
(271). The epoxy and
conjugated carbonyl groups in fucoxanthin confer potent antioxidant
capacity, predominantly via singlet oxygen quenching and free
radical scavenging mechanisms (272). Yoshikawa et al (70) demonstrated that this
antioxidative property of fucoxanthin exerts protective effects in
DEX-induced MA. Compared with DEX treatment, fucoxanthin
ameliorates mitochondrial dysfunction by suppressing AMPK pathway
activation and markedly decreases muscular malondialdehyde and ROS.
In an in vitro mechanistic study, DEX notably increased the
acetylation of FoxO3 and decreased its phosphorylation in C2C12
cells (68). These effects are
accompanied by a notable imbalance in ubiquitin ligase autophagy
and apoptosis (68). These
pathological alterations are reversed by fucoxanthin
supplementation via SIRT1 pathway activation; these protective
effects are attenuated by cotreatment with a SIRT1 inhibitor
(68).
Extensive clinical trials have established that
phytosterols are natural bioactive compounds capable of effectively
decreasing serum cholesterol levels (280,281). A daily intake of 2.0-2.5 g
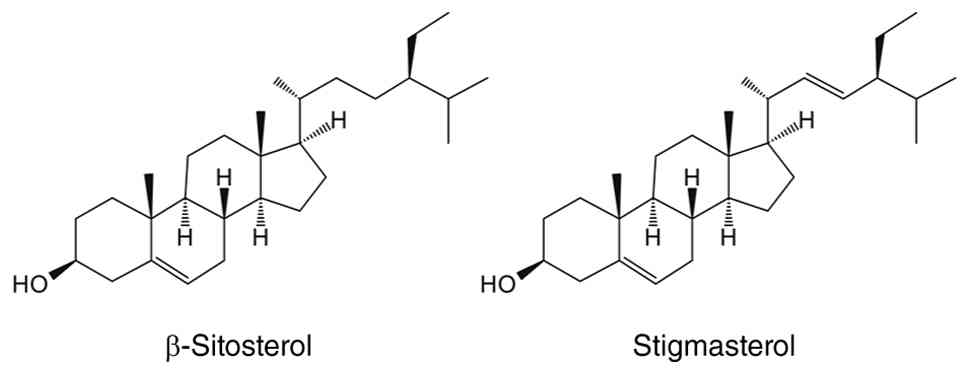
lowers the risk of cardiovascular disease (280). β-sitosterol, a structural
analogue of cholesterol, is one of the most abundant phytosterols
and is predominantly found in plant-derived foods, such as nuts,
seeds, leafy greens and vegetable oils (282). β-sitosterol is commonly
incorporated into dietary supplements, particularly those targeting
cardiovascular health and prostate function (283,284). Its cancer-preventive
properties, attributed to its anti-inflammatory, antioxidant,
proapoptotic and immunomodulatory activity, have been validated in
cell assays, animal models and clinical studies (285,286). β-sitosterol has demonstrated
beneficial effects on skeletal muscle mitochondrial function,
biogenesis and glucolipid metabolism in multiple biological
systems, including mice, C2C12 myotubes, chicken skeletal muscle
(287,288). Hah et al (107) used C57BL/6 mouse and C2C12 cell
models of DEX intervention to investigate the effects of
β-sitosterol on skeletal muscle catabolism. Treatment with
β-sitosterol ameliorates DEX-induced MA by increasing muscle mass,
enhancing physical performance and suppressing MPB (107). Mechanistically, FoxO1 is a
pivotal mediator of β-sitosterol catabolic inhibition, with its
activation associated with the decreased expression of atrogin-1,
MuRF1 and creatine kinase (107).
Stigmasterol is a common phytosterol whose chemical
structure is similar to that of β-sitosterol, differing only by an
additional double bond in its side chain (289). It is widely distributed in
plant cell membranes, such as those of soybeans and nuts, and has
analgesic, cholesterol-lowering and memory-improving properties
(289). Hah et al
(290) demonstrated the anti-MA
effect of stigmasterol. In vivo, oral administration of
stigmasterol notably reverses DEX-induced decreases in muscle mass,
myofibre CSA and bone mineral density. In vitro, 10
μM stigmasterol markedly increases the diameter and fusion
index of C2C12 cells, effectively ameliorating DEX-induced MA. From
a molecular mechanism perspective, stigmasterol alters the
subcellular distribution of FoxO3 in C2C12 cells (290). This manifests as notable
inhibition of the DEX-induced nuclear accumulation of FoxO3
(290). Furthermore,
stigmasterol concurrently restores mTORC1 activity, elevates the
phosphorylation levels of its downstream targets p70S6K and 4EBP1
and promotes MPS in skeletal muscle (290) (Fig. 9).
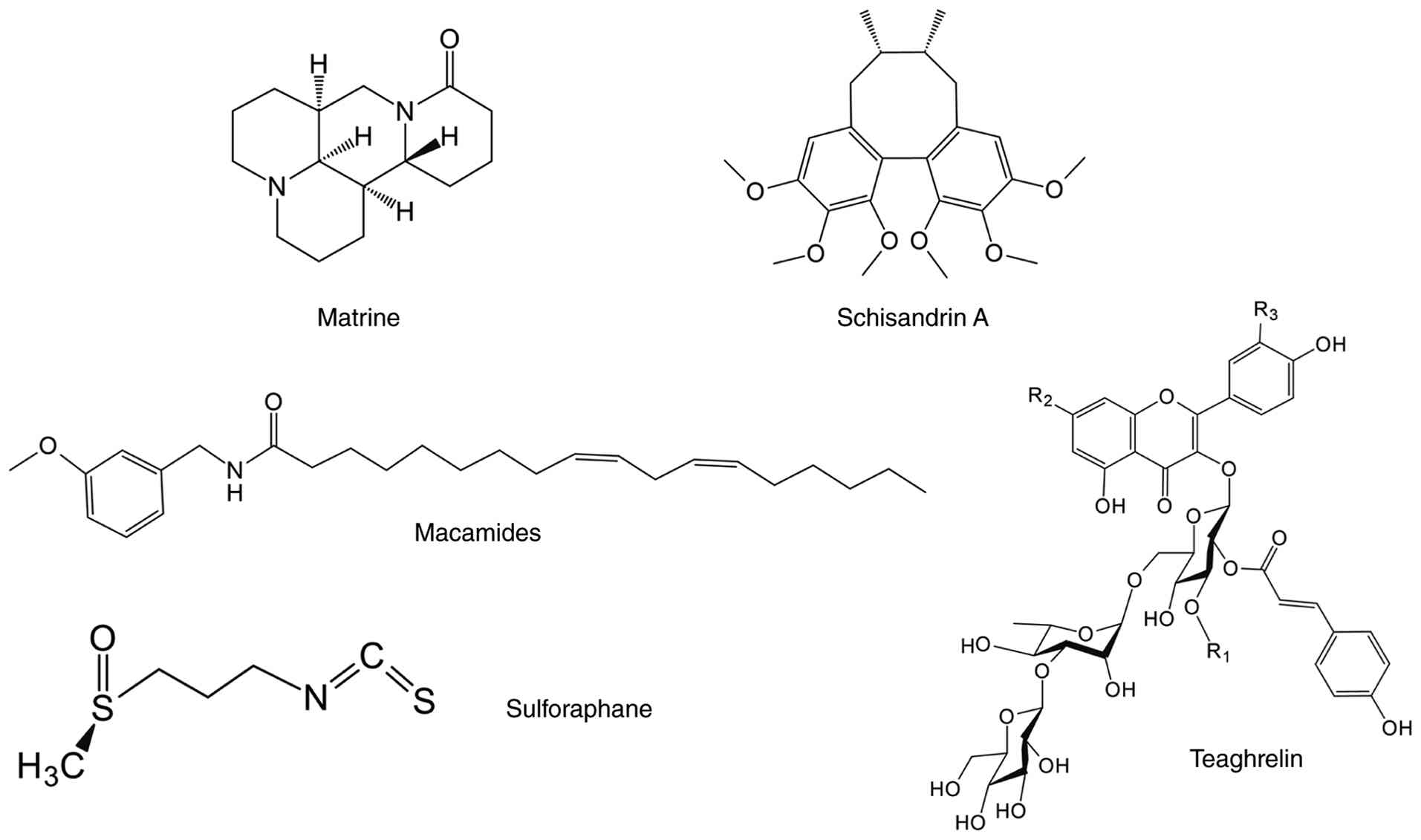
Matrine, an isoquinoline alkaloid compound derived
from the traditional Chinese medicinal herb ku shen, has
agricultural and pharmaceutical applications (291). In agriculture, matrine is used
as a low-toxicity and broad-spectrum insecticide with a good
preventive effect against crop pests, such as beetles and aphids
(292). In clinical practice,
matrine injection has been approved by the NMPA as an adjuvant
therapy to enhance chemotherapeutic outcomes in various types of
cancer, including pancreatic, lung, and breast cancer (293). Mechanistically, matrine exerts
its anticancer effects by inhibiting tumour cell proliferation and
metastasis, inducing apoptosis and mitigating drug resistance and
systemic toxicity associated with chemotherapeutic agents (294). Furthermore, its
anti-inflammatory, antibacterial, antiviral, cardioprotective,
neuroprotective and hepatorenal protective properties in in
vitro and in vivo models have been comprehensively
reviewed (293,295). The dual immunofluorescence
staining of MyHC and MyoD demonstrates that matrine considerably
ameliorates DEX- and TNF-α-induced MA by promoting myoblast
differentiation in a dose-dependent manner (296). Treatment with 0.1-0.4 mM
matrine, either alone or in combination with DEX, enhances myotube
differentiation, whereas treatment with 0.4-0.8 mM matrine induces
atypical myotube fusion and decreases MyoD expression (296). Mechanistically, treatment with
0.1 mM matrine for 48 h reverses the alteration of Akt/mTOR/FoxO3
pathway phosphorylation by DEX, thereby restoring the balance
between protein synthesis and degradation to mitigate MA (296).
Macamides are secondary metabolites unique to maca
root, with all 26 identified macamides belonging to the
N-benzylamide group (297).
Maca is known for its pharmacological effects on enhancing sexual
function and fertility, and macamides are the primary bioactive
compound mediating these benefits (298). Maca root comes in various
colours, with the extract from red maca root showing the most
potent muscle homeostatic modulation in promoting muscle
differentiation and inhibiting MPB. An in vitro intervention
experiment showed that N-(3-methoxybenzyl) linoleamide, a macamide
from Lepidium meyenii, inhibits the nuclear translocation of
GR and FoxO3 and downregulates MURF1 and atrogin-1 expression in a
concentration-dependent manner, thereby attenuating DEX-induced MA.
Molecular docking and dynamics simulation have identified multiple
binding sites between MAC (18:3) and Akt1, providing mechanistic
insight into its ability to enhance Akt phosphorylation during DEX
challenge (299).
Teaghrelin, an acylated flavonoid tetraglycoside
derived from oolong tea, regulates appetite and gastrointestinal
function while promoting growth hormone secretion (304). Its pharmacological activity is
associated with neuroprotection (305) and MA improvement (67,71). Hsieh et al (71) and Jhuo et al (67,71) demonstrated teaghrelin therapeutic
efficacy against DEX-induced MA in vitro and in vivo.
In C2C12 myotubes, teaghrelin mitigates DEX-induced MA by
decreasing E3 ubiquitin ligase expression, maintaining MyHC levels
and activating the Akt/mTOR signalling pathway (71). Teaghrelin regulates ubiquitin
ligase and autophagy marker expression associated with MPB in a
Sprague-Dawley rat model via Akt/FoxO1 pathway activation (67).
Sulphoraphane, an isothiocyanate abundant in
Brassicaceae vegetables, including broccoli, primarily exists in
plants in the form of its glucosinolate precursor glucoraphanin
(306). Its clinical
development as an Nrf2 activator stems from its covalent
modification of Keap1 cysteine sensors, which liberates Nrf2 for
activation (307). Through
Nrf2/NF-κB regulation and epigenetic modification (308), sulphoraphane exhibits
pharmacological activities, such as antioxidant, anti-inflammatory,
antitumor and neuroprotective activity (309), and is used to treat diabetes
(310). As a natural
phytochemical that promotes muscle health, sulphoraphane exerts a
skeletal muscle protective effect in models of muscle disease and
sports injury (311,312). Sulphoraphane monotherapy at
subtoxic concentrations enhances MyHCIb and MyHCIIx isoform
expression levels in C2C12 myotubes without cytotoxicity (313). When DEX is co-administered with
sulphoraphane, sulphoraphane promotes FoxO1 inactivation through
Akt activation, leading to the effective inhibition of the
DEX-induced upregulation of Mstn and atrogin-1 expression, showing
a potential protective effect against MA (313) (Fig. 10).
Plant-derived NPs primarily exert anti-MA effects
by modulating FoxO activity, with their regulatory mechanisms
exhibiting common molecular characteristics (Table II). Most compounds promote the
phosphorylation inactivation of FoxO1/FoxO3 or inhibit their
transcriptional activity via SIRT1-mediated deacetylation, thereby
preventing their nuclear translocation and downregulating the
expression of atrophy-associated genes. Concurrently, certain
compounds (myricanol, dihydromyricetin, fucoxanthin) indirectly
inhibit the abnormal activation of FoxO by improving mitochondrial
function, decreasing oxidative stress or restoring autophagy
homeostasis (59,68,70,174). In addition, GR is a direct
target of certain compounds (such as glabridin, AR and diosgenin)
that inhibit DEX-mediated FoxO activation by blocking GR nuclear
translocation, thereby exerting anti-MA effects (58,205,255).
In DEX-induced MA, existing studies have primarily
focused on indirectly affecting FoxO activity by regulating
upstream signalling pathways, such as PI3K/Akt, SIRT and GR
(58,59,187). To the best of our knowledge,
plant-derived compounds directly targeting FoxO have not yet been
reported. However, a limited number of studies in other disease
areas have begun to explore the direct effects of NPs on FoxO
(314-316). Kim et al (314) reported that syringaresinol
directly binds to the forkhead DNA-binding domain of FoxO3 and
enhances its transcriptional activity by stabilising the FoxO-DNA
complex. Through molecular docking, Gupta et al (315) identified NPs, such as fraxin,
as potential direct inhibitors of FoxO1. Guan et al
(316) demonstrated that
quercetin and catechin directly act on different sites of FoxO3 and
inhibit its transcriptional activity, thereby ameliorating acute
alcoholic liver injury. From a structural perspective, although
these compounds are derived from diverse sources, they are often
rich in aromatic rings and phenolic hydroxyl groups, exhibiting
certain conjugation and molecular rigidity. They may bind the
surfaces of FoxO proteins via hydrogen bonds and π-π interactions,
thereby modulating their conformation or DNA-binding state
(314-316). Nevertheless, to the best of our
knowledge, no systematic comparison has been performed to support
an association between the structure of plant-derived compounds and
FoxO-targeting activity.
The regulatory strategies for FoxO remain
predominantly indirect. For example, Orea-Soufi et al
(103) summarised clinical
drug-targeting strategies for FoxO across different disease
contexts and indicated that the majority of FoxO-targeting drug
strategies rely on indirect regulation through various signalling
pathways, with molecules directly acting on FoxO being limited.
This may be associated with structural characteristics of FoxO
proteins, namely, their lack of a classic small-molecule binding
pocket and their high proportion of IDRs, which make formation of a
stable drug-binding interface difficult (317). Targeted protein degradation
technologies offer new possibilities to overcome this limitation
(318,319). Among these technologies,
transcription factor-proteolysis-targeting chimera (PROTAC) has
garnered considerable attention (320,321). By contrast with traditional
PROTAC, which requires small-molecule ligands, transcription
factor-PROTAC directly uses the inherent DNA-binding capacity of
transcription factors. A transcription factor is selectively
degraded by conjugating the DNA-binding sequence of the target gene
with an E3 ubiquitin ligase ligand (320). This approach may be feasible in
the context of DEX-induced MA, as it does not require a
small-molecule ligand for FoxO. To the best of our knowledge,
however, none of the plant-derived compounds included in the
present review have been shown to bind FoxO directly. Therefore,
lead structures for the development of traditional PROTAC ligands
are lacking. FoxO proteins specifically recognise DNA sequences
through their forkhead domain, providing a directly usable
recognition element for the design of transcription factor-PROTACs.
Furthermore, certain NPs (syringaresinol, fraxin, quercetin)
possess direct affinity for FoxO (314-316), offering potential structural
templates for the development of FoxO degraders derived from NPs.
Therefore, developing highly efficient FoxO-targeting degraders by
integrating transcription factor-PROTAC with the structural
features of NPs may provide novel intervention strategies for
DEX-induced MA.
Although the transcriptional activity of FoxO is
regulated by PTMs, such as phosphorylation, acetylation and
methylation, the existing research on plant-derived NPs is
primarily restricted to the dynamic regulation of phosphorylation
modifications (134-136). Phosphorylation detection
techniques are mature and widely applied (134-136). By contrast, acetylation and
methylation detection relies on complex technical approaches, such
as co-IP, mass spectrometry and site-specific antibodies (133), with experimental complexity
limiting research. Moreover, current investigations have
predominantly focused on classical signalling pathways, such as the
PI3K/Akt and AMPK pathways, and their phosphorylation sites have
been extensively reported (134-136). The association between
phosphorylation and FoxO nuclear translocation, as well as
transcriptional activity, is established, making it readily
applicable for mechanistic interpretation (324). These factors collectively
render phosphorylation the most direct and commonly used indicator.
Nevertheless, with technological advancements, future research may
investigate the regulation of multiple PTMs of FoxO, thereby
enabling study into the mechanisms underlying the regulatory
effects of plant-derived NPs on FoxO.
As a potent synthetic GC, DEX is used to induce
skeletal MA,. DEX mediates the pathological process of skeletal MA
through multiple pathways, with the key mechanism being the dynamic
imbalance of muscle protein metabolism, which manifests as the
bidirectional dysregulation of anabolic inhibition and catabolic
activation. FoxO is associated with the MPB pathway, activating the
ALS and UPS to degrade key proteins involved in muscle growth and
differentiation while disrupting the homeostasis of muscle energy
metabolism. DEX intervention alters FoxO transcriptional activity
in myocytes, particularly FoxO1 and FoxO3. Therefore, the FoxO
family may serve as the central molecular hub in DEX-induced MA,
integrating catabolic signalling with energy metabolism networks to
promote the progressive loss of muscle mass and function. FoxO is
evolutionarily highly conserved and stable, with gene mutations and
deletions being rare. Therefore, the pharmacological modulation of
its inhibition or activation represents a promising therapeutic
strategy for various diseases, such as cancer, diabetes and MA. NPs
exhibit the advantages of low toxicity, multitarget effects and
broad-spectrum pharmacological activity, with herbal compounds
having been proven to be effective (21-23). The present review focused on
drug-induced MA caused by DEX, summarising the roles of
plant-derived monomeric compounds in ameliorating this type of MA.
Plant-derived NPs have potential in alleviating DEX-induced MA by
modulating FoxO. The present review not only expands the
application prospects of NPs in drug-induced MA but also provides
insights into the pathological mechanisms and intervention
strategies associated with drug-induced MA.
WD conceived the study and wrote the manuscript. JW
conceived the study and edited the manuscript. XM, LK, MW and HG
edited the manuscript. WW and WX supervised the study and edited
the manuscript. Data authentication is not applicable. All authors
have read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by the National Natural Science
Foundation of China (grant no. 32371185), the Shanghai Science and
Technology Plan Project (grant no. 23010504200), the Key Lab of
Exercise and Health Sciences of Ministry of Education (Shanghai
University of Sport; grant no. 2022KF001), the Shanghai Key Lab of
Human Performance (Shanghai University of Sport; grant no.
11DZ2261100) and Shanghai Oriental Talents Program (Youth
Project).
|
1
|
Frontera WR and Ochala J: Skeletal muscle:
A brief review of structure and function. Calcif Tissue Int.
96:183–195. 2015. View Article : Google Scholar
|
|
2
|
Mukund K and Subramaniam S: Skeletal
muscle: A review of molecular structure and function, in health and
disease. Wiley Interdiscip Rev Syst Biol Med. 12:e14622020.
View Article : Google Scholar
|
|
3
|
Ebert SM, Al-Zougbi A, Bodine SC and Adams
CM: Skeletal muscle atrophy: Discovery of mechanisms and potential
therapies. Physiology (Bethesda). 34:232–239. 2019.PubMed/NCBI
|
|
4
|
Yin L, Li N, Jia W, Wang N, Liang M, Yang
X and Du G: Skeletal muscle atrophy: From mechanisms to treatments.
Pharmacol Res. 172:1058072021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cain DW and Cidlowski JA: Immune
regulation by glucocorticoids. Nat Rev Immunol. 17:233–247. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Permpoon U, Moon J, Kim CY and Nam TG:
Glucocorticoid-mediated skeletal muscle atrophy: Molecular
mechanisms and potential therapeutic targets. Int J Mol Sci.
26:76162025. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miernik S, Matusiewicz A and Olesińska M:
Drug-induced myopathies: A comprehensive review and update.
Biomedicines. 12:9872024. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gupta A and Gupta Y:
Glucocorticoid-induced myopathy: Pathophysiology, diagnosis, and
treatment. Indian J Endocrinol Metab. 17:913–916. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang BYH, Hsiao AWT, Wong N, Chen YF, Lee
CW and Lee WYW: Is dexamethasone-induced muscle atrophy an
alternative model for naturally aged sarcopenia model? J Orthop
Translat. 39:12–20. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Son YH, Lee SJ, Lee KB, Lee JH, Jeong EM,
Chung SG, Park SC and Kim IG: Dexamethasone downregulates
caveolin-1 causing muscle atrophy via inhibited insulin signaling.
J Endocrinol. 225:27–37. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Czock D, Keller F, Rasche FM and Häussler
U: Pharmacokinetics and pharmacodynamics of systemically
administered glucocorticoids. Clin Pharmacokinet. 44:61–98. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vazquez S, Gold J, Spirollari E, Akmal S
and Hanft SJ: The story of dexamethasone and how it became one of
the most widely used drugs in neurosurgery. J Neurosurg.
140:1191–1197. 2024. View Article : Google Scholar
|
|
13
|
Clarke BA, Drujan D, Willis MS, Murphy LO,
Corpina RA, Burova E, Rakhilin SV, Stitt TN, Patterson C, Latres E
and Glass DJ: The E3 ligase MuRF1 degrades myosin heavy chain
protein in dexamethasone-treated skeletal muscle. Cell Metab.
6:376–385. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Troncoso R, Paredes F, Parra V, Gatica D,
Vásquez-Trincado C, Quiroga C, Bravo-Sagua R, López-Crisosto C,
Rodriguez AE, Oyarzún AP, et al: Dexamethasone-induced autophagy
mediates muscle atrophy through mitochondrial clearance. Cell
Cycle. 13:2281–2295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klein AJ and Vaughan RA: A systematic
review and user reference of phenotypic and molecular
characteristics of Dexamethasone-mediated C2C12 muscle atrophy. J
Cachexia Sarcopenia Muscle. 17:e701272026. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Martins R, Lithgow GJ and Link W: Long
live FOXO: Unraveling the role of FOXO proteins in aging and
longevity. Aging Cell. 15:196–207. 2016. View Article : Google Scholar :
|
|
17
|
Sanchez AMJ, Candau RB and Bernardi H:
FoxO transcription factors: Their roles in the maintenance of
skeletal muscle homeostasis. Cell Mol Life Sci. 71:1657–1671. 2014.
View Article : Google Scholar
|
|
18
|
Poulsen RC, Carr AJ and Hulley PA:
Protection against glucocorticoid-induced damage in human tenocytes
by modulation of ERK, Akt, and forkhead signaling. Endocrinology.
152:503–514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sandri M, Sandri C, Gilbert A, Skurk C,
Calabria E, Picard A, Walsh K, Schiaffino S, Lecker SH and Goldberg
AL: Foxo transcription factors induce the atrophy-related ubiquitin
ligase atrogin-1 and cause skeletal muscle atrophy. Cell.
117:399–412. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schakman O, Kalista S, Barbé C, Loumaye A
and Thissen JP: Glucocorticoid-induced skeletal muscle atrophy. Int
J Biochem Cell Biol. 45:2163–2172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu J, Li K, Zhou M, Gao H, Wang W and Xiao
W: Natural compounds improve diabetic nephropathy by regulating the
TLR4 signaling pathway. J Pharm Anal. 14:1009462024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tabrizi FB, Yarmohammadi F, Hayes AW and
Karimi G: The modulation of SIRT1 and SIRT3 by natural compounds as
a therapeutic target in doxorubicin-induced cardiotoxicity: A
review. J Biochem Mol Toxicol. 36:e229462022. View Article : Google Scholar
|
|
23
|
Zhao W, Li B, Hao J, Sun R, He P, Lv H, He
M, Shen J and Han Y: Therapeutic potential of natural products and
underlying targets for the treatment of aortic aneurysm. Pharmacol
Ther. 259:1086522024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang M, Ren J, Chen X, Liu J, Xu X, Li X,
Zhao D and Sun L: 20(S)-ginsenoside Rg3 promotes myoblast
differentiation and protects against myotube atrophy via regulation
of the Akt/mTOR/FoxO3 pathway. Biochem Pharmacol. 180:1141452020.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Alamdari N, Aversa Z, Castillero E, Gurav
A, Petkova V, Tizio S and Hasselgren PO: Resveratrol prevents
dexamethasone-induced expression of the muscle atrophy-related
ubiquitin ligases atrogin-1 and MuRF1 in cultured myotubes through
a SIRT1-dependent mechanism. Biochem Biophys Res Commun.
417:528–533. 2012. View Article : Google Scholar
|
|
26
|
Yue H, Huan Y, Ren P, Yu X, Tang Q, Xue C
and Xu J: Astaxanthin ameliorates dexamethasone-induced skeletal
muscle atrophy and disorders of glucolipid metabolism. Biochem
Biophys Res Commun. 743:1511382025. View Article : Google Scholar
|
|
27
|
Urbańska J, Karewicz A and Nowakowska M:
Polymeric delivery systems for dexamethasone. Life Sci. 96:1–6.
2014. View Article : Google Scholar
|
|
28
|
Benedek TG: History of the development of
corticosteroid therapy. Clin Exp Rheumatol. 29(5 Suppl 68):
S-5–S-12. 2011.
|
|
29
|
Cole TJ: Glucocorticoid action and the
development of selective glucocorticoid receptor ligands.
Biotechnol Annu Rev. 12:269–300. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Madamsetty VS, Mohammadinejad R, Uzieliene
I, Nabavi N, Dehshahri A, García-Couce J, Tavakol S, Moghassemi S,
Dadashzadeh A, Makvandi P, et al: Dexamethasone: Insights into
pharmacological aspects, therapeutic mechanisms, and delivery
systems. ACS Biomater Sci Eng. 8:1763–1790. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Haller J, Mikics E and Makara GB: The
effects of non-genomic glucocorticoid mechanisms on bodily
functions and the central neural system. A critical evaluation of
findings. Front Neuroendocrinol. 29:273–291. 2008. View Article : Google Scholar
|
|
32
|
Williams DM: Clinical pharmacology of
corticosteroids. Respir Care. 63:655–670. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Richardson PG, Trudel S, Popat R, Mateos
MV, Vangsted AJ, Ramasamy K, Martinez-Lopez J, Quach H, Orlowski
RZ, Arnao M, et al: Mezigdomide plus dexamethasone in relapsed and
refractory multiple myeloma. N Engl J Med. 389:1009–1022. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jick H, Pinals RS, Ullian R, Slone D and
Muench H: Dexamethasone and dexamethasone-aspirin in the treatment
of chronic rheumatoid arthritis. A controlled trial. Lancet.
2:1203–1205. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin MX, Guo L, Saldanha IJ, VanCourt S,
Zeng J, Karakus S, Hessen M, Li G and Akpek EK: Dexamethasone
intracanalicular insert for clinically significant
Aqueous-Deficient dry eye: A randomized controlled trial.
Ophthalmology. 131:1033–1044. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li J, Panganiban R, Kho AT, McGeachie MJ,
Farnam L, Chase RP, Weiss ST, Lu Q and Tantisira KG: Circulating
MicroRNAs and treatment response in childhood asthma. Am J Respir
Crit Care Med. 202:65–72. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li G, Hilgenfeld R, Whitley R and De
Clercq E: Therapeutic strategies for COVID-19: Progress and lessons
learned. Nat Rev Drug Discov. 22:449–475. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Oray M, Abu Samra K, Ebrahimiadib N, Meese
H and Foster CS: Long-term side effects of glucocorticoids. Expert
Opin Drug Saf. 15:457–465. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cushing H: The basophil adenomas of the
pituitary body and their clinical manifestations (Pituitary
basophilism). 1932. Obes Res. 2:486–508. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tiao G, Fagan J, Roegner V, Lieberman M,
Wang JJ, Fischer JE and Hasselgren PO: Energy-ubiquitin-dependent
muscle proteolysis during sepsis in rats is regulated by
glucocorticoids. J Clin Invest. 97:339–348. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
De Oliveira GS, Ahmad S, Fitzgerald PC,
Marcus RJ, Altman CS, Panjwani AS and McCarthy RJ: Dose ranging
study on the effect of preoperative dexamethasone on postoperative
quality of recovery and opioid consumption after ambulatory
gynaecological surgery. Br J Anaesth. 107:362–371. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Habibian JS, Bolino M, Qian A, Woolsey R,
Quilici D, Petereit J and Ferguson BS: Class I HDAC inhibitors
attenuate dexamethasone-induced muscle atrophy via increased
protein kinase C (PKC) delta phosphorylation. Cell Signal.
110:1108152023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Braun TP, Zhu X, Szumowski M, Scott GD,
Grossberg AJ, Levasseur PR, Graham K, Khan S, Damaraju S, Colmers
WF, et al: Central nervous system inflammation induces muscle
atrophy via activation of the hypothalamic-pituitary-adrenal axis.
J Exp Med. 208:2449–2463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Barel M, Perez OAB, Giozzet VA, Rafacho A,
Bosqueiro JR and do Amaral SL: Exercise training prevents
hyperinsulinemia, muscular glycogen loss and muscle atrophy induced
by dexamethasone treatment. Eur J Appl Physiol. 108:999–1007. 2010.
View Article : Google Scholar
|
|
45
|
Ewart HS, Somwar R and Klip A:
Dexamethasone stimulates the expression of GLUT1 and GLUT4 proteins
via different signalling pathways in L6 skeletal muscle cells. FEBS
Lett. 425:179–183. 1998.PubMed/NCBI
|
|
46
|
Wang XJ, Song ZG, Jiao HC and Lin H:
Dexamethasone facilitates lipid accumulation in chicken skeletal
muscle. Stress. 15:443–456. 2012. View Article : Google Scholar
|
|
47
|
Livingstone I, Johnson MA and Mastaglia
FL: Effects of dexamethasone on fibre subtypes in rat muscle.
Neuropathol Appl Neurobiol. 7:381–398. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schiaffino S, Dyar KA, Ciciliot S, Blaauw
B and Sandri M: Mechanisms regulating skeletal muscle growth and
atrophy. FEBS J. 280:4294–4314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu WJ, Guo K, Shi JL, Guo CT, Xu JL, Zheng
R, Jiang SW and Chai J: Glucocorticoid regulates the synthesis of
porcine muscle protein through m6A modified amino acid transporter
SLC7A7. Int J Mol Sci. 23:6612022. View Article : Google Scholar
|
|
50
|
Shimizu N, Yoshikawa N, Ito N, Maruyama T,
Suzuki Y, Takeda S, Nakae J, Tagata Y, Nishitani S, Takehana K, et
al: Crosstalk between glucocorticoid receptor and nutritional
sensor mTOR in skeletal muscle. Cell Metab. 13:170–182. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stitt TN, Drujan D, Clarke BA, Panaro F,
Timofeyva Y, Kline WO, Gonzalez M, Yancopoulos GD and Glass DJ: The
IGF-1/PI3K/Akt pathway prevents expression of muscle
atrophy-induced ubiquitin ligases by inhibiting FOXO transcription
factors. Mol Cell. 14:395–403. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fappi A, Neves J, de C, Sanches LN,
Massaroto E, Silva PV, Sikusawa GY, Brandão TPC, Chadi G and
Zanoteli E: Skeletal muscle response to deflazacort, dexamethasone
and methylprednisolone. Cells. 8:4062019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Koh A, Lee MN, Yang YR, Jeong H, Ghim J,
Noh J, Kim J, Ryu D, Park S, Song P, et al: C1-Ten is a protein
tyrosine phosphatase of insulin receptor substrate 1 (IRS-1),
regulating IRS-1 stability and muscle atrophy. Mol Cell Biol.
33:1608–1620. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Li XL, Wang L, He MC, Li WX, Zhang JL, Fu
YF and Zhang Y: A clinical herbal prescription Gu-Shu-Kang capsule
exerted beneficial effects on the musculoskeletal system of
dexamethasone-treated mice by acting on tissue IGF-1 signalling
pathway. Pharm Biol. 60:2098–2109. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Foletta VC, White LJ, Larsen AE, Léger B
and Russell AP: The role and regulation of MAFbx/atrogin-1 and
MuRF1 in skeletal muscle atrophy. Pflugers Arch. 461:325–335. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Shin K, Ko YG, Jeong J and Kwon H: Fbxw7β
is an inducing mediator of dexamethasone-induced skeletal muscle
atrophy in vivo with the axis of Fbxw7β-myogenin-atrogenes. Mol
Biol Rep. 45:625–631. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sun H, Gong Y, Qiu J, Chen Y, Ding F and
Zhao Q: TRAF6 inhibition rescues dexamethasone-induced muscle
atrophy. Int J Mol Sci. 15:11126–11141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yoshioka Y, Kubota Y, Samukawa Y,
Yamashita Y and Ashida H: Glabridin inhibits dexamethasone-induced
muscle atrophy. Arch Biochem Biophys. 664:157–166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shen S, Liao Q, Liu J, Pan R, Lee SMY and
Lin L: Myricanol rescues dexamethasone-induced muscle dysfunction
via a sirtuin 1-dependent mechanism. J Cachexia Sarcopenia Muscle.
10:429–444. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Baehr LM, Furlow JD and Bodine SC: Muscle
sparing in muscle RING finger 1 null mice: Response to synthetic
glucocorticoids. J Physiol. 589:4759–4776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kang SH, Lee HA, Kim M, Lee E, Sohn UD and
Kim I: Forkhead box O3 plays a role in skeletal muscle atrophy
through expression of E3 ubiquitin ligases MuRF-1 and atrogin-1 in
Cushing's syndrome. Am J Physiol Endocrinol Metab. 312:E495–E507.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ma K, Mallidis C, Bhasin S, Mahabadi V,
Artaza J, Gonzalez-Cadavid N, Arias J and Salehian B:
Glucocorticoid-induced skeletal muscle atrophy is associated with
upregulation of myostatin gene expression. Am J Physiol Endocrinol
Metab. 285:E363–E371. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Verhees KJP, Schols AMWJ, Kelders MCJM,
Opden Kamp CMH, van der Velden JLJ and Langen RCJ: Glycogen
synthase kinase-3β is required for the induction of skeletal muscle
atrophy. Am J Physiol Cell Physiol. 301:C995–C1007. 2011.
View Article : Google Scholar
|
|
64
|
Li BG, Hasselgren PO and Fang CH:
Insulin-like growth factor-I inhibits dexamethasone-induced
proteolysis in cultured L6 myotubes through PI3K/Akt/GSK-3beta and
PI3K/Akt/mTOR-dependent mechanisms. Int J Biochem Cell Biol.
37:2207–2216. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zheng B, Ohkawa S, Li H, Roberts-Wilson TK
and Price SR: FOXO3a mediates signaling crosstalk that coordinates
ubiquitin and atrogin-1/MAFbx expression during
glucocorticoid-induced skeletal muscle atrophy. FASEB J.
24:2660–2669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Pankiv S, Clausen TH, Lamark T, Brech A,
Bruun JA, Outzen H, Øvervatn A, Bjørkøy G and Johansen T:
p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of
ubiquitinated protein aggregates by autophagy. J Biol Chem.
282:24131–24145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Jhuo CF, Hsieh SK, Chen WY and Tzen JTC:
Attenuation of skeletal muscle atrophy induced by dexamethasone in
rats by teaghrelin supplementation. Molecules. 28:6882023.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhiyin L, Jinliang C, Qiunan C, Yunfei Y
and Qian X: Fucoxanthin rescues dexamethasone induced C2C12
myotubes atrophy. Biomed Pharmacother. 139:1115902021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kim JY, Kim HM, Kim JH, Guo S, Lee DH, Lim
GM, Kim W and Kim CY: Salvia plebeia R.Br. and rosmarinic acid
attenuate Dexamethasone-induced muscle atrophy in C2C12 myotubes.
Int J Mol Sci. 24:18762023. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yoshikawa M, Hosokawa M, Miyashita K,
Nishino H and Hashimoto T: Effects of fucoxanthin on the inhibition
of Dexamethasone-induced skeletal muscle loss in mice. Nutrients.
13:10792021. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hsieh SK, Lin HY, Chen CJ, Jhuo CF, Liao
KY, Chen WY and Tzen JTC: Promotion of myotube differentiation and
attenuation of muscle atrophy in murine C2C12 myoblast cells
treated with teaghrelin. Chem Biol Interact. 315:1088932020.
View Article : Google Scholar
|
|
72
|
Luan G, Li G, Ma X, Jin Y, Hu N, Li J,
Wang Z and Wang H: Dexamethasone-Induced mitochondrial dysfunction
and insulin Resistance-Study in 3T3-L1 adipocytes and mitochondria
isolated from mouse liver. Molecules. 24:19822019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu J, Peng Y, Wang X, Fan Y, Qin C, Shi
L, Tang Y, Cao K, Li H, Long J and Liu J: Mitochondrial dysfunction
launches Dexamethasone-induced skeletal muscle atrophy via
AMPK/FOXO3 signaling. Mol Pharm. 13:73–84. 2016. View Article : Google Scholar
|
|
74
|
Sandri M, Lin J, Handschin C, Yang W,
Arany ZP, Lecker SH, Goldberg AL and Spiegelman BM: PGC-1alpha
protects skeletal muscle from atrophy by suppressing FoxO3 action
and atrophy-specific gene transcription. Proc Natl Acad Sci USA.
103:16260–16265. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Cannavino J, Brocca L, Sandri M,
Bottinelli R and Pellegrino MA: PGC1-α over-expression prevents
metabolic alterations and soleus muscle atrophy in hindlimb
unloaded mice. J Physiol. 592:4575–4589. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Yeon MH, Seo E, Lee JH and Jun HS:
Bavachin and corylifol a improve muscle atrophy by enhancing
mitochondria quality control in type 2 diabetic mice. Antioxidants
(Basel). 12:1372023. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Chen C, Yang JS, Lu CC, Chiu YJ, Chen HC,
Chung MI, Wu YT and Chen FA: Effect of quercetin on
Dexamethasone-Induced C2C12 skeletal muscle cell injury. Molecules.
25:32672020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ulla A, Uchida T, Miki Y, Sugiura K,
Higashitani A, Kobayashi T, Ohno A, Nakao R, Hirasaka K, Sakakibara
I and Nikawa T: Morin attenuates dexamethasone-mediated oxidative
stress and atrophy in mouse C2C12 skeletal myotubes. Arch Biochem
Biophys. 704:1088732021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Malkawi AK, Masood A, Shinwari Z, Jacob M,
Benabdelkamel H, Matic G, Almuhanna F, Dasouki M, Alaiya AA and
Rahman AMA: Proteomic analysis of morphologically changed tissues
after prolonged dexamethasone treatment. Int J Mol Sci.
20:31222019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Huang M, Yan Y, Deng Z, Zhou L, She M,
Yang Y, Zhang M and Wang D: Saikosaponin A and D attenuate skeletal
muscle atrophy in chronic kidney disease by reducing oxidative
stress through activation of PI3K/AKT/Nrf2 pathway. Phytomedicine.
114:1547662023. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Luo L, Martin SC, Parkington J, Cadena SM,
Zhu J, Ibebunjo C, Summermatter S, Londraville N, Patora-Komisarska
K, Widler L, et al: HDAC4 Controls muscle homeostasis through
deacetylation of myosin heavy chain, PGC-1α, and Hsc70. Cell Rep.
29:749–763.e12. 2019. View Article : Google Scholar
|
|
82
|
Choi J, Jang H, Kim H, Kim ST, Cho EJ and
Youn HD: Histone demethylase LSD1 is required to induce skeletal
muscle differentiation by regulating myogenic factors. Biochem
Biophys Res Commun. 401:327–332. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Scionti I, Hayashi S, Mouradian S, Girard
E, Esteves de Lima J, Morel V, Simonet T, Wurmser M, Maire P,
Ancelin K, et al: LSD1 controls timely MyoD expression via MyoD
core enhancer transcription. Cell Rep. 18:1996–2006. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Clark EA, Wu F, Chen Y, Kang P, Kaiser UB,
Fang R and Shi YG: GR and LSD1/KDM1A-targeted gene activation
requires selective H3K4me2 demethylation at enhancers. Cell Rep.
27:3522–3532.e3. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Cai Q, Sahu R, Ueberschlag-Pitiot V,
Souali-Crespo S, Charvet C, Silem I, Cottard F, Ye T, Taleb F,
Metzger E, et al: LSD1 inhibition circumvents
glucocorticoid-induced muscle wasting of male mice. Nat Commun.
15:35632024. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Araki H, Hino S, Anan K, Kuribayashi K,
Etoh K, Seko D, Takase R, Kohrogi K, Hino Y, Ono Y, et al: LSD1
defines the fiber type-selective responsiveness to environmental
stress in skeletal muscle. Elife. 12:e846182023. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Qin J, Du R, Yang YQ, Zhang HQ, Li Q, Liu
L, Guan H, Hou J and An XR: Dexamethasone-induced skeletal muscle
atrophy was associated with upregulation of myostatin promoter
activity. Res Vet Sci. 94:84–89. 2013. View Article : Google Scholar
|
|
88
|
Proserpio V, Fittipaldi R, Ryall JG,
Sartorelli V and Caretti G: The methyltransferase SMYD3 mediates
the recruitment of transcriptional cofactors at the myostatin and
c-Met genes and regulates skeletal muscle atrophy. Genes Dev.
27:1299–1312. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Liu Y, Li J, Shang Y, Guo Y and Li Z:
CARM1 contributes to skeletal muscle wasting by mediating FoxO3
activity and promoting myofiber autophagy. Exp Cell Res.
374:198–209. 2019. View Article : Google Scholar
|
|
90
|
Senf SM, Dodd SL and Judge AR: FOXO
signaling is required for disuse muscle atrophy and is directly
regulated by Hsp70. Am J Physiol Cell Physiol. 298:C38–C45. 2010.
View Article : Google Scholar
|
|
91
|
Milan G, Romanello V, Pescatore F, Armani
A, Paik JH, Frasson L, Seydel A, Zhao J, Abraham R, Goldberg AL, et
al: Regulation of autophagy and the ubiquitin-proteasome system by
the FoxO transcriptional network during muscle atrophy. Nat Commun.
6:66702015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Lützner N, Kalbacher H, Krones-Herzig A
and Rösl F: FOXO3 is a glucocorticoid receptor target and regulates
LKB1 and its own expression based on cellular AMP levels via a
positive autoregulatory loop. PLoS One. 7:e421662012. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Waddell DS, Baehr LM, van den Brandt J,
Johnsen SA, Reichardt HM, Furlow JD and Bodine SC: The
glucocorticoid receptor and FOXO1 synergistically activate the
skeletal muscle atrophy-associated MuRF1 gene. Am J Physiol
Endocrinol Metab. 295:E785–E797. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Reed SA, Sandesara PB, Senf SM and Judge
AR: Inhibition of FoxO transcriptional activity prevents muscle
fiber atrophy during cachexia and induces hypertrophy. FASEB J.
26:987–1000. 2012. View Article : Google Scholar
|
|
95
|
Langendorf EK, Rommens PM, Drees P,
Mattyasovszky SG and Ritz U: Detecting the effects of the
glucocorticoid dexamethasone on primary human skeletal muscle
Cells-differences to the murine cell line. Int J Mol Sci.
21:24972020. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Qin W, Pan J, Qin Y, Lee DN, Bauman WA and
Cardozo C: Identification of functional glucocorticoid response
elements in the mouse FoxO1 promoter. Biochem Biophys Res Commun.
450:979–983. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Santos BF, Grenho I, Martel PJ, Ferreira
BI and Link W: FOXO family isoforms. Cell Death Dis. 14:7022023.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Weigel D and Jäckle H: The fork head
domain: A novel DNA binding motif of eukaryotic transcription
factors? Cell. 63:455–456. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Obsil T and Obsilova V: Structure/function
relationships underlying regulation of FOXO transcription factors.
Oncogene. 27:2263–2275. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Wang F, Marshall CB and Ikura M: Forkhead
followed by disordered tail: The intrinsically disordered regions
of FOXO3a. Intrinsically Disord Proteins. 3:e10569062015.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Willcox BJ, Donlon TA, He Q, Chen R, Grove
JS, Yano K, Masaki KH, Willcox DC, Rodriguez B and Curb JD: FOXO3A
genotype is strongly associated with human longevity. Proc Natl
Acad Sci USA. 105:13987–13992. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Calissi G, Lam EWF and Link W: Therapeutic
strategies targeting FOXO transcription factors. Nat Rev Drug
Discov. 20:21–38. 2021. View Article : Google Scholar
|
|
103
|
Orea-Soufi A, Paik J, Bragança J, Donlon
TA, Willcox BJ and Link W: FOXO transcription factors as
therapeutic targets in human diseases. Trends Pharmacol Sci.
43:1070–1084. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Hosaka T, Biggs WH, Tieu D, Boyer AD,
Varki NM, Cavenee WK and Arden KC: Disruption of forkhead
transcription factor (FOXO) family members in mice reveals their
functional diversification. Proc Natl Acad Sci USA. 101:2975–2980.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Salih DA, Rashid AJ, Colas D, de la
Torre-Ubieta L, Zhu RP, Morgan AA, Santo EE, Ucar D, Devarajan K,
Cole CJ, et al: FoxO6 regulates memory consolidation and synaptic
function. Genes Dev. 26:2780–2801. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Hah YS, Lee WK, Lee SJ, Lee SY, Seo JH,
Kim EJ, Choe YI, Kim SG and Yoo JI: Rutin prevents
Dexamethasone-induced muscle loss in C2C12 myotube and mouse model
by controlling FOXO3-Dependent signaling. Antioxidants (Basel).
12:6392023. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Hah YS, Lee WK, Lee S, Kim EJ, Lee JH, Lee
SJ, Ji YH, Kim SG, Lee HH, Hong SY and Yoo JI: β-Sitosterol
attenuates Dexamethasone-induced muscle atrophy via regulating
FoxO1-dependent signaling in C2C12 cell and mice model. Nutrients.
14:28942022. View Article : Google Scholar
|
|
108
|
Smith IJ, Alamdari N, O'Neal P, Gonnella
P, Aversa Z and Hasselgren PO: Sepsis increases the expression and
activity of the transcription factor Forkhead Box O 1 (FOXO1) in
skeletal muscle by a glucocorticoid-dependent mechanism. Int J
Biochem Cell Biol. 42:701–711. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Penniman CM, Bhardwaj G, Nowers CJ, Brown
CU, Junck TL, Boyer CK, Jena J, Fuqua JD, Lira VA and O'Neill BT:
Loss of FoxOs in muscle increases strength and mitochondrial
function during aging. J Cachexia Sarcopenia Muscle. 14:243–259.
2023. View Article : Google Scholar :
|
|
110
|
Yu R, Chen JA, Xu J, Cao J, Wang Y, Thomas
SS and Hu Z: Suppression of muscle wasting by the plant-derived
compound ursolic acid in a model of chronic kidney disease. J
Cachexia Sarcopenia Muscle. 8:327–341. 2017. View Article : Google Scholar :
|
|
111
|
Hishiya A, Iemura S, Natsume T, Takayama
S, Ikeda K and Watanabe K: A novel ubiquitin-binding protein ZNF216
functioning in muscle atrophy. EMBO J. 25:554–564. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhao J, Brault JJ, Schild A, Cao P, Sandri
M, Schiaffino S, Lecker SH and Goldberg AL: FoxO3 coordinately
activates protein degradation by the autophagic/lysosomal and
proteasomal pathways in atrophying muscle cells. Cell Metab.
6:472–483. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Leduc-Gaudet JP, Franco-Romero A, Cefis M,
Moamer A, Broering FE, Milan G, Sartori R, Chaffer TJ, Dulac M,
Marcangeli V, et al: MYTHO is a novel regulator of skeletal muscle
autophagy and integrity. Nat Commun. 14:11992023. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Goh KY, Lee WX, Choy SM, Priyadarshini GK,
Chua K, Tan QH, Low SY, Chin HS, Wong CS, Huang SY, et al:
FOXO-regulated DEAF1 controls muscle regeneration through
autophagy. Autophagy. 20:2632–2654. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Komamura K, Shirotani-Ikejima H, Tatsumi
R, Tsujita-Kuroda Y, Kitakaze M, Miyatake K, Sunagawa K and Miyata
T: Differential gene expression in the rat skeletal and heart
muscle in glucocorticoid-induced myopathy: Analysis by microarray.
Cardiovasc Drugs Ther. 17:303–310. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Allen DL and Unterman TG: Regulation of
myostatin expression and myoblast differentiation by FoxO and SMAD
transcription factors. Am J Physiol Cell Physiol. 292:C188–C199.
2007. View Article : Google Scholar
|
|
117
|
Hu P, Geles KG, Paik JH, DePinho RA and
Tjian R: Codependent activators direct myoblast-specific MyoD
transcription. Dev Cell. 15:534–546. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Kitamura T, Kitamura YI, Funahashi Y,
Shawber CJ, Castrillon DH, Kollipara R, DePinho RA, Kitajewski J
and Accili D: A Foxo/Notch pathway controls myogenic
differentiation and fiber type specification. J Clin Invest.
117:2477–2485. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Lokireddy S, Mouly V, Butler-Browne G,
Gluckman PD, Sharma M, Kambadur R and McFarlane C: Myostatin
promotes the wasting of human myoblast cultures through promoting
ubiquitin-proteasome pathway-mediated loss of sarcomeric proteins.
Am J Physiol Cell Physiol. 301:C1316–C1324. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
McFarlane C, Plummer E, Thomas M, Hennebry
A, Ashby M, Ling N, Smith H, Sharma M and Kambadur R: Myostatin
induces cachexia by activating the ubiquitin proteolytic system
through an NF-κB-independent, FoxO1-dependent mechanism. J Cell
Physiol. 209:501–514. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Lees SJ, Childs TE and Booth FW:
Age-dependent FOXO regulation of p27Kip1 expression via a conserved
binding motif in rat muscle precursor cells. Am J Physiol Cell
Physiol. 295:C1238–C1246. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Rathbone CR, Booth FW and Lees SJ: FoxO3a
preferentially induces p27Kip1 expression while impairing muscle
precursor cell-cycle progression. Muscle Nerve. 37:84–89. 2008.
View Article : Google Scholar
|
|
123
|
Bhardwaj G, Penniman CM, Klaus K,
Weatherford ET, Pan H, Dreyfuss JM, Nair KS, Kahn CR and O'Neill
BT: Transcriptomic regulation of muscle mitochondria and calcium
signaling by Insulin/IGF-1 receptors depends on FoxO transcription
factors. Front Physiol. 12:7791212021. View Article : Google Scholar
|
|
124
|
Bhardwaj G, Penniman CM, Jena J, Suarez
Beltran PA, Foster C, Poro K, Junck TL, Hinton AO Jr, Souvenir R,
Fuqua JD, et al: Insulin and IGF-1 receptors regulate complex
I-dependent mitochondrial bioenergetics and supercomplexes via
FoxOs in muscle. J Clin Invest. 131:e1464152021. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zufiría M, Pikatza-Menoio O,
Garciandia-Arcelus M, Bengoetxea X, Jiménez A, Elicegui A, Levchuk
M, Arnold-García O, Ondaro J, Iruzubieta P, et al: Dysregulated
FOXO1 activity drives skeletal muscle intrinsic dysfunction in
amyotrophic lateral sclerosis. Acta Neuropathol. 148:432024.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Kamei Y, Mizukami J, Miura S, Suzuki M,
Takahashi N, Kawada T, Taniguchi T and Ezaki O: A forkhead
transcription factor FKHR up-regulates lipoprotein lipase
expression in skeletal muscle. FEBS Lett. 536:232–236. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Bastie CC, Nahlé Z, McLoughlin T, Esser K,
Zhang W, Unterman T and Abumrad NA: FoxO1 stimulates fatty acid
uptake and oxidation in muscle cells through CD36-dependent and
-independent mechanisms. J Biol Chem. 280:14222–14229. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Furuyama T, Kitayama K, Yamashita H and
Mori N: Forkhead transcription factor FOXO1 (FKHR)-dependent
induction of PDK4 gene expression in skeletal muscle during energy
deprivation. Biochem J. 375:365–371. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Sinam IS, Chanda D, Thoudam T, Kim MJ, Kim
BG, Kang HJ, Lee JY, Baek SH, Kim SY, Shim BJ, et al: Pyruvate
dehydrogenase kinase 4 promotes ubiquitin-proteasome
system-dependent muscle atrophy. J Cachexia Sarcopenia Muscle.
13:3122–3136. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Lundell LS, Massart J, Altıntaş A, Krook A
and Zierath JR: Regulation of glucose uptake and inflammation
markers by FOXO1 and FOXO3 in skeletal muscle. Mol Metab. 20:79–88.
2019. View Article : Google Scholar :
|
|
131
|
Kamei Y, Miura S, Suzuki M, Kai Y,
Mizukami J, Taniguchi T, Mochida K, Hata T, Matsuda J, Aburatani H,
et al: Skeletal muscle FOXO1 (FKHR) transgenic mice have less
skeletal muscle mass, down-regulated Type I (slow twitch/red
muscle) fiber genes, and impaired glycemic control. J Biol Chem.
279:41114–41123. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Hasselgren PO: Ubiquitination,
phosphorylation, and acetylation-triple threat in muscle wasting. J
Cell Physiol. 213:679–689. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Choi S, Jeong HJ, Kim H, Choi D, Cho SC,
Seong JK, Koo SH and Kang JS: Skeletal muscle-specific Prmt1
deletion causes muscle atrophy via deregulation of the PRMT6-FOXO3
axis. Autophagy. 15:1069–1081. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Cid-Díaz T, Santos-Zas I, González-Sánchez
J, Gurriarán-Rodríguez U, Mosteiro CS, Casabiell X,
García-Caballero T, Mouly V, Pazos Y and Camiña JP: Obestatin
controls the ubiquitin-proteasome and autophagy-lysosome systems in
glucocorticoid-induced muscle cell atrophy. J Cachexia Sarcopenia
Muscle. 8:974–990. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Jaitovich A, Angulo M, Lecuona E, Dada LA,
Welch LC, Cheng Y, Gusarova G, Ceco E, Liu C, Shigemura M, et al:
High CO2 levels cause skeletal muscle atrophy via
AMP-activated kinase (AMPK), FoxO3a protein, and muscle-specific
Ring finger protein 1 (MuRF1). J Biol Chem. 290:9183–9194. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Tong JF, Yan X, Zhu MJ and Du M:
AMP-activated protein kinase enhances the expression of
muscle-specific ubiquitin ligases despite its activation of
IGF-1/Akt signaling in C2C12 myotubes. J Cell Biochem. 108:458–468.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Wei B, Dui W, Liu D, Xing Y, Yuan Z and Ji
G: MST1, a key player, in enhancing fast skeletal muscle atrophy.
BMC Biol. 11:122013. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo
P, Hu LS, Anderson MJ, Arden KC, Blenis J and Greenberg ME: Akt
promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Saline M, Badertscher L, Wolter M, Lau R,
Gunnarsson A, Jacso T, Norris T, Ottmann C and Snijder A: AMPK and
AKT protein kinases hierarchically phosphorylate the N-terminus of
the FOXO1 transcription factor, modulating interactions with 14-3-3
proteins. J Biol Chem. 294:13106–13116. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Greer EL, Oskoui PR, Banko MR, Maniar JM,
Gygi MP, Gygi SP and Brunet A: The energy sensor AMP-activated
protein kinase directly regulates the mammalian FOXO3 transcription
factor. J Biol Chem. 282:30107–30119. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Hwangbo H, Kim MY, Ji SY, Kim DH, Park BS,
Jeong SU, Yoon JH, Kim TH, Kim GY and Choi YH: A Mixture of morus
alba and Angelica keiskei leaf extracts improves muscle atrophy by
activating the PI3K/Akt/mTOR signaling pathway and inhibiting
FoxO3a in vitro and in vivo. J Microbiol Biotechnol. 33:1635–1647.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Fernando P, Kelly JF, Balazsi K, Slack RS
and Megeney LA: Caspase 3 activity is required for skeletal muscle
differentiation. Proc Natl Acad Sci USA. 99:11025–11030. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Chamberlain W, Gonnella P, Alamdari N,
Aversa Z and Hasselgren PO: Multiple muscle wasting-related
transcription factors are acetylated in dexamethasone-treated
muscle cells. Biochem Cell Biol. 90:200–208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Wang Z, Yu T and Huang P:
Post-translational modifications of FOXO family proteins (review).
Mol Med Rep. 14:4931–4941. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Senf SM, Sandesara PB, Reed SA and Judge
AR: p300 Acetyltransferase activity differentially regulates the
localization and activity of the FOXO homologues in skeletal
muscle. Am J Physiol Cell Physiol. 300:C1490–C1501. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Enrico B, Coletto L and Sandri M:
Posttranslational modifications control FoxO3 activity during
denervation. Am J Physiol Cell Physiol. 302:C587–C596. 2012.
View Article : Google Scholar
|
|
147
|
Beharry AW, Sandesara PB, Roberts BM,
Ferreira LF, Senf SM and Judge AR: HDAC1 activates FoxO and is both
sufficient and required for skeletal muscle atrophy. J Cell Sci.
127:1441–1453. 2014.PubMed/NCBI
|
|
148
|
Lee D and Goldberg AL: SIRT1 protein, by
blocking the activities of transcription factors FoxO1 and FoxO3,
inhibits muscle atrophy and promotes muscle growth. J Biol Chem.
288:30515–30526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
vanLieshout TL and Ljubicic V: The
emergence of protein arginine methyltransferases in skeletal muscle
and metabolic disease. Am J Physiol Endocrinol Metab.
317:E1070–E1080. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Brault JJ, Jespersen JG and Goldberg AL:
Peroxisome proliferator-activated receptor gamma coactivator 1alpha
or 1beta overexpression inhibits muscle protein degradation,
induction of ubiquitin ligases, and disuse atrophy. J Biol Chem.
285:19460–19471. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Bollinger LM, Witczak CA, Houmard JA and
Brault JJ: SMAD3 augments FoxO3-induced MuRF-1 promoter activity in
a DNA-binding-dependent manner. Am J Physiol Cell Physiol.
307:C278–C287. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Di Petrillo A, Orrù G, Fais A and Fantini
MC: Quercetin and its derivates as antiviral potentials: A
comprehensive review. Phytother Res. 36:266–278. 2022. View Article : Google Scholar
|
|
153
|
Deepika and Maurya PK: Health benefits of
quercetin in Age-related diseases. Molecules. 27:24982022.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Georgiou N, Kakava MG, Routsi EA, Petsas
E, Stavridis N, Freris C, Zoupanou N, Moschovou K, Kiriakidi S and
Mavromoustakos T: Quercetin: A potential polydynamic drug.
Molecules. 28:81412023. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Zaplatic E, Bule M, Shah SZA, Uddin MS and
Niaz K: Molecular mechanisms underlying protective role of
quercetin in attenuating Alzheimer's disease. Life Sci.
224:109–119. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Lee H, Kim YI, Kim MJ, Hahm JH, Seo HD, Ha
TY, Jung CH and Ahn J: Castor oil plant (Ricinus communis L.)
Leaves improve Dexamethasone-Induced muscle atrophy via Nrf2
activation. Front Pharmacol. 13:8917622022. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Park SH, Oh J, Jo M, Kim JK, Kim DS, Kim
HG, Yoon K, Yang Y, Geum JH, Kim JE, et al: Water extract of lotus
leaf alleviates Dexamethasone-induced muscle atrophy via regulating
protein Metabolism-Related pathways in mice. Molecules.
25:45922020. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Otsuka Y, Egawa K, Kanzaki N, Izumo T,
Rogi T and Shibata H: Quercetin glycosides prevent
Dexamethasone-induced muscle atrophy in mice. Biochem Biophys Rep.
18:1006182019.PubMed/NCBI
|
|
159
|
Dai GH, Meng GM, Tong YL, Chen X, Ren ZM,
Wang K and Yang F: Growth-inhibiting and apoptosis-inducing
activities of Myricanol from the bark of Myrica rubra in human lung
adenocarcinoma A549 cells. Phytomedicine. 21:1490–1496. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Zheng M, Jiang Q, You J, Gao B, Cui W, Yao
W, Su F, Sun X and La L: Myricanol represses renal fibrosis by
activating TFAM and ZNRF1 to inhibit tubular epithelial cells
ferroptosis. Eur J Pharmacol. 984:1769992024. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Jones JR, Lebar MD, Jinwal UK, Abisambra
JF, Koren J III, Blair L, O'Leary JC, Davey Z, Trotter J, Johnson
AG, et al: The diarylheptanoid (+)-aR, 11S-myricanol and two
flavones from bayberry (Myrica cerifera) destabilize the
microtubule-associated protein tau. J Nat Prod. 74:38–44. 2011.
View Article : Google Scholar
|
|
162
|
Shen S, Liao Q, Feng Y, Liu J, Pan R, Lee
SMY and Lin L: Myricanol mitigates lipid accumulation in 3T3-L1
adipocytes and high fat diet-fed zebrafish via activating
AMP-activated protein kinase. Food Chem. 270:305–314. 2019.
View Article : Google Scholar
|
|
163
|
Chen J, Liu B, Yao X, Yang X, Sun J, Yi J,
Xue F, Zhang J, Shen Y, Chen B and Sun H: AMPK/SIRT1/PGC-1α
Signaling pathway: Molecular mechanisms and targeted strategies
from energy homeostasis regulation to disease therapy. CNS Neurosci
Ther. 31:e706572025. View Article : Google Scholar
|
|
164
|
Li T, Hu X, Fan L, Yang Y and He K:
Myricanol improves metabolic profiles in dexamethasone induced
lipid and protein metabolism disorders in mice. Biomed
Pharmacother. 174:1165572024. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Shen S, Liao Q, Zhang T, Pan R and Lin L:
Myricanol modulates skeletal muscle-adipose tissue crosstalk to
alleviate high-fat diet-induced obesity and insulin resistance. Br
J Pharmacol. 176:3983–4001. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Shen S, Liao Q, Lyu P, Wang J and Lin L:
Myricanol prevents aging-related sarcopenia by rescuing
mitochondrial dysfunction via targeting peroxiredoxin 5. MedComm
(2020). 5:e5662024. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Rajput SA, Wang XQ and Yan HC: Morin
hydrate: A comprehensive review on novel natural dietary bioactive
compound with versatile biological and pharmacological potential.
Biomed Pharmacother. 138:1115112021. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Wu RR, Li X, Cao YH, Peng X, Liu GF, Liu
ZK, Yang Z, Liu ZY and Wu Y: China medicinal plants of the
ampelopsis grossedentata-A review of their botanical
characteristics, Use, phytochemistry, active pharmacological
components, and toxicology. Molecules. 28:71452023. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Liu D, Mao Y, Ding L and Zeng XA:
Dihydromyricetin: A review on identification and quantification
methods, biological activities, chemical stability, metabolism and
approaches to enhance its bioavailability. Trends Food Sci Technol.
91:586–597. 2019. View Article : Google Scholar
|
|
170
|
Chen S, Zhao X, Wan J, Ran L, Qin Y, Wang
X, Gao Y, Shu F, Zhang Y, Liu P, et al: Dihydromyricetin improves
glucose and lipid metabolism and exerts anti-inflammatory effects
in nonalcoholic fatty liver disease: A randomized controlled trial.
Pharmacol Res. 99:74–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Ran L, Wang X, Lang H, Xu J, Wang J, Liu
H, Mi M and Qin Y: Ampelopsis grossedentata supplementation
effectively ameliorates the glycemic control in patients with type
2 diabetes mellitus. Eur J Clin Nutr. 73:776–782. 2019. View Article : Google Scholar
|
|
172
|
You L: Dihydromyricetin inhibits
ferroptosis to attenuate Cisplatin-induced muscle atrophy. Physiol
Res. 73:405–413. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Hou L, Jiang F, Huang B, Zheng W, Jiang Y,
Cai G, Liu D, Hu CY and Wang C: Dihydromyricetin resists
inflammation-induced muscle atrophy via ryanodine
receptor-CaMKK-AMPK signal pathway. J Cell Mol Med. 25:9953–9971.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Huang Y, Chen K, Ren Q, Yi L, Zhu J, Zhang
Q and Mi M: Dihydromyricetin attenuates dexamethasone-induced
muscle atrophy by improving mitochondrial function via the PGC-1α
pathway. Cell Physiol Biochem. 49:758–779. 2018. View Article : Google Scholar
|
|
175
|
Hayashi H, Hattori S, Inoue K,
Khodzhimatov O, Ashurmetov O, Ito M and Honda G: Field survey of
Glycyrrhiza plants in Central Asia (3). Chemical characterization
of G. glabra collected in Uzbekistan. Chem Pharm Bull (Tokyo).
51:1338–1340. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Simmler C, Pauli GF and Chen SN:
Phytochemistry and biological properties of glabridin. Fitoterapia.
90:160–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Liu J, Xu X, Jian M, Guo Y, Zhai L, Sun G,
Sun L and Jiang R: Glycyrrhiza glabra extract as a skin-whitening
Agent: Identification of active components and CRTC1/MITF
pathway-inhibition mechanism. J Ethnopharmacol. 349:1199482025.
View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Pan C, Liu X, Zheng Y, Zhang Z, Li Y, Che
B, Liu G, Zhang L, Dong C, Aisa HA, et al: The mechanisms of
melanogenesis inhibition by glabridin: Molecular docking, PKA/MITF
and MAPK/MITF pathways. Food Sci Hum Wellness. 12:212–222. 2023.
View Article : Google Scholar
|
|
179
|
Zhang J, Wu X, Zhong B, Liao Q, Wang X,
Xie Y and He X: Review on the diverse biological effects of
glabridin. Drug Des Devel Ther. 17:15–37. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Kang MR, Park KH, Oh SJ, Yun J, Lee CW,
Lee MY, Han SB and Kang JS: Cardiovascular protective effect of
glabridin: Implications in LDL oxidation and inflammation. Int
Immunopharmacol. 29:914–918. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Wang L, Yang R, Yuan B, Liu Y and Liu C:
The antiviral and antimicrobial activities of licorice, a
widely-used Chinese herb. Acta Pharm Sin B. 5:310–315. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Qiu M, Ma K, Zhang J, Zhao Z, Wang S, Wang
Q and Xu H: Isoliquiritigenin as a modulator of the Nrf2 signaling
pathway: Potential therapeutic implications. Front Pharmacol.
15:13957352024. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Chen Z, Ding W, Yang X, Lu T and Liu Y:
Isoliquiritigenin, a potential therapeutic agent for treatment of
inflammation-associated diseases. J Ethnopharmacol. 318:1170592024.
View Article : Google Scholar
|
|
184
|
Tang S, Deng Z, Ou M and Wang Z:
Pharmacological potentials and delivery strategies of
isoliquiritigenin: Challenges and advances in enhancing
bioavailability. Drug Des Devel Ther. 19:9879–9911. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Link P and Wink M: Isoliquiritigenin
exerts antioxidant activity in Caenorhabditis elegans via
insulin-like signaling pathway and SKN-1. Phytomedicine.
55:119–124. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Hayes JD, Dinkova-Kostova AT and Tew KD:
Oxidative stress in cancer. Cancer Cell. 38:167–197. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Jiao X, Zhang Y, Wang Z, Chang H, Li Y,
Wang B, Gan Y and Gu D: Isoliquiritigenin, an extract from
licorice, attenuates dexamethasone-induced muscle atrophy via
Akt/mTOR pathway. Mol Nutr Food Res. 69:e700002025. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Khan N, Afaq F, Saleem M, Ahmad N and
Mukhtar H: Targeting multiple signaling pathways by green tea
polyphenol (−)-epigallocatechin-3-gallate. Cancer Res.
66:2500–2505. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Farhan M: Green tea catechins: Nature's
Way of preventing and treating cancer. Int J Mol Sci. 23:107132022.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Chowdhury A, Sarkar J, Chakraborti T,
Pramanik PK and Chakraborti S: Protective role of
epigallocatechin-3-gallate in health and disease: A perspective.
Biomed Pharmacother. 78:50–59. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Kaihatsu K, Yamabe M and Ebara Y:
Antiviral mechanism of action of Epigallocatechin-3-O-gallate and
its fatty acid esters. Molecules. 23:24752018. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Zhao H, Zhu W, Zhao X, Li X, Zhou Z, Zheng
M, Meng X, Kong L, Zhang S, He D, et al: Efficacy of
Epigallocatechin-3-gallate in preventing dermatitis in patients
with breast cancer receiving postoperative radiotherapy: A
Double-Blind, Placebo-controlled, phase 2 randomized clinical
trial. JAMA Dermatol. 158:779–786. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Zhao H, Jia L, Chen G, Li X, Meng X, Zhao
X, Xing L and Zhu W: A prospective, three-arm, randomized trial of
EGCG for preventing radiation-induced esophagitis in lung cancer
patients receiving radiotherapy. Radiother Oncol. 137:186–191.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Tachibana H, Koga K, Fujimura Y and Yamada
K: A receptor for green tea polyphenol EGCG. Nat Struct Mol Biol.
11:380–381. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Murata M, Shimizu Y, Marugame Y, Nezu A,
Fujino K, Yamada S, Kumazoe M, Fujimura Y and Tachibana H: EGCG
down-regulates MuRF1 expression through 67-kDa laminin receptor and
the receptor signaling is amplified by eriodictyol. J Nat Med.
74:673–679. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Bartholome A, Kampkötter A, Tanner S, Sies
H and Klotz LO: Epigallocatechin gallate-induced modulation of FoxO
signaling in mammalian cells and C. elegans: FoxO stimulation is
masked via PI3K/Akt activation by hydrogen peroxide formed in cell
culture. Arch Biochem Biophys. 501:58–64. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Wang T, Xu H, Wu S, Guo Y, Zhao G and Wang
D: Mechanisms underlying the effects of the green tea polyphenol
EGCG in sarcopenia prevention and management. J Agric Food Chem.
71:9609–9627. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Lyu J, Kim BJ and Kim H: Anti-allergic
effect of ulmus davidiana cortex on contact dermatitis induced by
dinitrofluoro-Benzene in mice. J Pharmacopuncture. 16:41–45. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Rho J, Seo CS, Park HS, Wijerathne CU,
Jeong HY, Moon OS, Seo YW, Son HY, Won YS and Kwun HJ: Ulmus
macrocarpa Hance improves benign prostatic hyperplasia by
regulating prostatic cell apoptosis. J Ethnopharmacol. 233:115–122.
2019. View Article : Google Scholar
|
|
200
|
Kim MS, Park S, Kwon Y, Kim T, Lee CH,
Jang H, Kim EJ, Jung JI, Min S, Park KH and Choi SE: Effects of
Ulmus macrocarpa extract and catechin 7-O-β-D-apiofuranoside on
Muscle loss and muscle atrophy in C2C12 murine skeletal muscle
cells. Curr Issues Mol Biol. 46:8320–8339. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Lee CH, Kwon Y, Park S, Kim T, Kim MS, Kim
EJ, Jung JI, Min S, Park KH, Jeong JH and Choi SE: The impact of
Ulmus macrocarpa extracts on a model of Sarcopenia-induced C57BL/6
Mice. Int J Mol Sci. 25:61972024. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Mottaghipisheh J, Taghrir H, Boveiri
Dehsheikh A, Zomorodian K, Irajie C, Mahmoodi Sourestani M and
Iraji A: Linarin, a glycosylated flavonoid, with potential
therapeutic attributes: A comprehensive review. Pharmaceuticals
(Basel). 14:11042021. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Zhen ZG, Ren SH, Ji HM, Ma JH, Ding XM,
Feng FQ, Chen SL, Zou P, Ren JR and Jia L: Linarin suppresses
glioma through inhibition of NF-κB/p65 and up-regulating p53
expression in vitro and in vivo. Biomed Pharmacother. 95:363–374.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Pan H, Zhang J, Wang Y, Cui K, Cao Y, Wang
L and Wu Y: Linarin improves the dyskinesia recovery in Alzheimer's
disease zebrafish by inhibiting the acetylcholinesterase activity.
Life Sci. 222:112–116. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Lee H, Kim YI, Nirmala FS, Jeong HY, Seo
HD, Ha TY, Jung CH and Ahn J: Chrysanthemum zawadskil Herbich
attenuates dexamethasone-induced muscle atrophy through the
regulation of proteostasis and mitochondrial function. Biomed
Pharmacother. 136:1112262021. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Rudrapal M, Khan J, Dukhyil AAB, Alarousy
RM II, Attah EI, Sharma T, Khairnar SJ and Bendale AR: Chalcone
scaffolds, bioprecursors of flavonoids: Chemistry, bioactivities,
and pharmacokinetics. Molecules. 26:71772021. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Li Y, Goto T, Ikutani R, Lin S, Takahashi
N, Takahashi H, Jheng HF, Yu R, Taniguchi M, Baba K, et al:
Xanthoangelol and 4-hydroxyderrcin suppress obesity-induced
inflammatory responses. Obesity (Silver Spring). 24:2351–2360.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Odongo K, Abe A, Kawasaki R, Kawabata K
and Ashida H: Two Prenylated Chalcones, 4-hydroxyderricin, and
xanthoangelol prevent postprandial hyperglycemia by promoting GLUT4
translocation via the LKB1/AMPK signaling pathway in skeletal
muscle cells. Mol Nutr Food Res. 68:e23005382024. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Enoki T, Ohnogi H, Nagamine K, Kudo Y,
Sugiyama K, Tanabe M, Kobayashi E, Sagawa H and Kato I:
Antidiabetic activities of chalcones isolated from a Japanese Herb,
Angelica keiskei. J Agric Food Chem. 55:6013–6017. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Watanabe N, Inoue K, Hara H, Midorikawa M,
Ohta M and Ohkura N: Randomised, double-blind, parallel group
comparison of Ashitaba (Angelica keiskei) chalcone effects on
visceral fat areas and waist circumference of overweight persons.
Int J Food Sci Nutr. 75:426–435. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Yoshioka Y, Samukawa Y, Yamashita Y and
Ashida H: 4-Hydroxyderricin and xanthoangelol isolated from
Angelica keiskei prevent dexamethasone-induced muscle loss. Food
Funct. 11:5498–5512. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Kweon M, Lee H, Park C, Choi YH and Ryu
JH: A Chalcone from ashitaba (Angelica keiskei) Stimulates myoblast
differentiation and inhibits Dexamethasone-induced muscle atrophy.
Nutrients. 11:24192019. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Frémont L: Biological effects of
resveratrol. Life Sci. 66:663–673. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Rauf A, Imran M, Butt MS, Nadeem M, Peters
DG and Mubarak MS: Resveratrol as an anti-cancer agent: A review.
Crit Rev Food Sci Nutr. 58:1428–1447. 2018. View Article : Google Scholar
|
|
215
|
Ren B, Kwah MX, Liu C, Ma Z, Shanmugam MK,
Ding L, Xiang X, Ho PC, Wang L, Ong PS and Goh BC: Resveratrol for
cancer therapy: Challenges and future perspectives. Cancer Lett.
515:63–72. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Singh AP, Singh R, Verma SS, Rai V,
Kaschula CH, Maiti P and Gupta SC: Health benefits of resveratrol:
Evidence from clinical studies. Med Res Rev. 39:1851–1891. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Brown K, Theofanous D, Britton RG, Aburido
G, Pepper C, Sri Undru S and Howells L: Resveratrol for the
management of human health: How Far have we come? A systematic
review of resveratrol clinical trials to highlight gaps and
opportunities. Int J Mol Sci. 25:7472024. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Howitz KT, Bitterman KJ, Cohen HY, Lamming
DW, Lavu S, Wood JG, Zipkin RE, Chung P, Kisielewski A, Zhang LL,
et al: Small molecule activators of sirtuins extend Saccharomyces
cerevisiae lifespan. Nature. 425:191–196. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Pezzuto JM: Resveratrol: Twenty years of
growth, development and controversy. Biomol Ther (Seoul). 27:1–14.
2019. View Article : Google Scholar :
|
|
220
|
Huhn S, Beyer F, Zhang R, Lampe L, Grothe
J, Kratzsch J, Willenberg A, Breitfeld J, Kovacs P, Stumvoll M, et
al: Effects of resveratrol on memory performance, hippocampus
connectivity and microstructure in older adults-A randomized
controlled trial. Neuroimage. 174:177–190. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Wang R, Yuan W, Li L, Lu F, Zhang L, Gong
H and Huang X: Resveratrol ameliorates muscle atrophy in chronic
kidney disease via the axis of SIRT1/FoxO1. Phytother Res.
36:3265–3275. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Liao ZY, Chen JL, Xiao MH, Sun Y, Zhao YX,
Pu D, Lv AK, Wang ML, Zhou J, Zhu SY, et al: The effect of
exercise, resveratrol or their combination on Sarcopenia in aged
rats via regulation of AMPK/Sirt1 pathway. Exp Gerontol.
98:177–183. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Penedo-Vázquez A, Duran X, Mateu J,
López-Postigo A and Barreiro E: Curcumin and resveratrol improve
muscle function and structure through attenuation of proteolytic
markers in experimental Cancer-induced cachexia. Molecules.
26:49042021. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Liu CW, Huang CC, Hsu CF, Li TH, Tsai YL,
Lin MW, Tsai HC, Huang SF, Yang YY, Hsieh YC, et al:
SIRT1-dependent mechanisms and effects of resveratrol for
amelioration of muscle wasting in NASH mice. BMJ Open
Gastroenterol. 7:e0003812020. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Yang L, Liu D, Jiang S, Li H, Chen L, Wu
Y, Essien AE, Opoku M, Naranmandakh S, Liu S, et al: SIRT1
signaling pathways in sarcopenia: Novel mechanisms and potential
therapeutic targets. Biomed Pharmacother. 177:1169172024.
View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Kowalczyk A, Tuberoso CIG and Jerković I:
The role of rosmarinic acid in cancer prevention and therapy:
Mechanisms of antioxidant and anticancer activity. Antioxidants
(Basel). 13:13132024. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Guan H, Luo W, Bao B, Cao Y, Cheng F, Yu
S, Fan Q, Zhang L, Wu Q and Shan M: A comprehensive review of
rosmarinic acid: From phytochemistry to pharmacology and its new
insight. Molecules. 27:32922022. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Hitl M, Kladar N, Gavarić N and Božin B:
Rosmarinic Acid-human pharmacokinetics and health benefits. Planta
Med. 87:273–282. 2021. View Article : Google Scholar
|
|
229
|
Liang N and Kitts DD: Amelioration of
oxidative stress in Caco-2 cells treated with Pro-inflammatory
proteins by chlorogenic acid isomers via activation of the
Nrf2-Keap1-ARE-signaling pathway. J Agric Food Chem.
66:11008–11017. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Kim YI, Lee H, Kim MJ, Jung CH, Kim YS and
Ahn J: Identification of Peucedanum japonicum Thunb. Extract
components and their protective effects against
dexamethasone-induced muscle atrophy. Phytomedicine.
128:1554492024. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Heffernan N, Brunton NP, FitzGerald RJ and
Smyth TJ: Profiling of the molecular weight and structural isomer
abundance of macroalgae-derived phlorotannins. Mar Drugs.
13:509–528. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Xiao D, Fu J, Kim AT, Liyanage NM, Yang F
and Jeon YJ: Multi-targeted Anti-diabetic effects of trifuhalol a
from edible seaweed via glycation inhibition, glucose uptake
enhancement and pancreatic islet protection. Biofactors.
52:e700812026. View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Kim AT and Park Y: Trifuhalol A, a
phlorotannin from the brown algae Agarum cribrosum, reduces
adipogenesis of human primary adipocytes through Wnt/β-catenin and
AMPK-dependent pathways. Curr Res Food Sci. 7:1006462023.
View Article : Google Scholar
|
|
234
|
Bong SK, Park NJ, Lee SH, Lee JW, Kim AT,
Liu X, Kim SM, Yang MH, Kim YK and Kim SN: Trifuhalol A suppresses
allergic inflammation through dual inhibition of TAK1 and MK2
mediated by IgE and IL-33. Int J Mol Sci. 23:101632022. View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Yang F, Yang HW, Xiao D, Kim AT, Liu X,
Kim SM, Kim D and Jeon YJ: Protective effects of Trifuhalol A, a
Phlorotannin derived from edible Brown seaweed Agarum cribrosum, on
dexamethasone-induced muscle atrophy in muscle cells and zebrafish
models. Food Res Int. 209:1163032025. View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Kang NJ, Han SC, Kang GJ, Koo DH, Koh YS,
Hyun JW, Lee NH, Ko MH, Kang HK and Yoo ES:
Diphlorethohydroxycarmalol inhibits interleukin-6 production by
regulating NF-κB, STAT5 and SOCS1 in lipopolysaccharide-stimulated
RAW264.7 cells. Mar Drugs. 13:2141–2157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Piao MJ, Kang KA, Kim KC, Chae S, Kim GO,
Shin T, Kim HS and Hyun JW: Diphlorethohydroxycarmalol attenuated
cell damage against UVB radiation via enhancing antioxidant effects
and absorbing UVB ray in human HaCaT keratinocytes. Environ Toxicol
Pharmacol. 36:680–688. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
238
|
Cha S-H, Hwang Y, Heo SJ and Jun HS:
Diphlorethohydroxycarmalol attenuates Palmitate-induced hepatic
lipogenesis and inflammation. Mar Drugs. 18:4752020. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Zhen AX, Piao MJ, Hyun YJ, Kang KA,
Madushan Fernando PDS, Cho SJ, Ahn MJ and Hyun JW:
Diphlorethohydroxycarmalol attenuates fine particulate
Matter-induced subcellular skin dysfunction. Mar Drugs. 17:952019.
View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Ryu B, Oh S, Yang HW, Sosorburam B, Chung
DM, Seo M, Park SJ, Byun K and Jeon YJ: Diphlorethohydroxycarmalol
derived from Ishige okamurae improves behavioral and physiological
responses of muscle atrophy induced by dexamethasone in an In-Vivo
model. Pharmaceutics. 14:7192022. View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Hou M, Wang R, Zhao S and Wang Z:
Ginsenosides in Panax genus and their biosynthesis. Acta Pharm Sin
B. 11:1813–1834. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Xu W, Lyu W, Duan C, Ma F, Li X and Li D:
Preparation and bioactivity of the rare ginsenosides Rg3 and Rh2:
An updated review. Fitoterapia. 167:1055142023. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Bai X, Duan Z, Deng J, Zhang Z, Fu R, Zhu
C and Fan D: Ginsenoside Rh4 inhibits colorectal cancer via the
modulation of gut microbiota-mediated bile acid metabolism. J Adv
Res. 72:37–52. 2025. View Article : Google Scholar :
|
|
244
|
Zarneshan SN, Fakhri S and Khan H:
Targeting Akt/CREB/BDNF signaling pathway by ginsenosides in
neurodegenerative diseases: A mechanistic approach. Pharmacol Res.
177:1060992022. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Karmazyn M, Moey M and Gan XT: Therapeutic
potential of ginseng in the management of cardiovascular disorders.
Drugs. 71:1989–2008. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
246
|
Cheng Y, Shen LH and Zhang JT:
Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and
its mechanism of action. Acta Pharmacol Sin. 26:143–149. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
247
|
Liu P, Zhang Z, Cai Y, Yang Y, Yuan J and
Chen Q: Inhibition of the pyroptosis-associated inflammasome
pathway: The important potential mechanism of ginsenosides in
ameliorating diabetes and its complications. Eur J Med Chem.
253:1153362023. View Article : Google Scholar : PubMed/NCBI
|
|
248
|
Zha W, Sun Y, Gong W, Li L, Kim W and Li
H: Ginseng and ginsenosides: Therapeutic potential for sarcopenia.
Biomed Pharmacother. 156:1138762022. View Article : Google Scholar : PubMed/NCBI
|
|
249
|
Men X, Han X, Lee SJ, Oh G, Im JH, Bae KS,
Seong GS, La IJ, Lee DS, Choi SI and Lee OH: Ginsenosides Rh1, Rg2,
and Rg3 ameliorate dexamethasone-induced muscle atrophy in C2C12
myotubes. Food Sci Biotechnol. 33:1233–1243. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
250
|
Kim R, Kim JW, Choi H, Oh JE, Kim TH, Go
GY, Lee SJ and Bae GU: Ginsenoside Rg5 promotes muscle regeneration
via p38MAPK and Akt/mTOR signaling. J Ginseng Res. 47:726–734.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Wang M, Jiang R, Liu J, Xu X, Sun G, Zhao
D and Sun L: 20(s)-ginseonside-Rg3 modulation of AMPK/FoxO3
signaling to attenuate mitochondrial dysfunction in a
dexamethasone-injured C2C12 myotube-based model of skeletal atrophy
in vitro. Mol Med Rep. 23:3062021. View Article : Google Scholar
|
|
252
|
Go GY, Lee SJ, Jo A, Lee J, Seo DW, Kang
JS, Kim SK, Kim SN, Kim YK and Bae GU: Ginsenoside Rg1 from Panax
ginseng enhances myoblast differentiation and myotube growth. J
Ginseng Res. 41:608–614. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
253
|
Jesus M, Martins APJ, Gallardo E and
Silvestre S: Diosgenin: Recent highlights on pharmacology and
analytical methodology. J Anal Methods Chem. 2016:41562932016.
View Article : Google Scholar
|
|
254
|
Chen Y, Tang YM, Yu SL, Han YW, Kou JP,
Liu BL and Yu BY: Advances in the pharmacological activities and
mechanisms of diosgenin. Chin J Nat Med. 13:578–587.
2015.PubMed/NCBI
|
|
255
|
Yoshioka Y, Oishi S, Onoda K, Shibata K
and Miyoshi N: Diosgenin prevents dexamethasone-induced myotube
atrophy in C2C12 cells. Arch Biochem Biophys. 747:1097592023.
View Article : Google Scholar
|
|
256
|
Hou W, Liu B and Xu H: Celastrol:
Progresses in structure-modifications, structure-activity
relationships, pharmacology and toxicology. Eur J Med Chem.
189:1120812020. View Article : Google Scholar : PubMed/NCBI
|
|
257
|
Ng SW, Chan Y, Chellappan DK, Madheswaran
T, Zeeshan F, Chan YL, Collet T, Gupta G, Oliver BG, Wark P, et al:
Molecular modulators of celastrol as the keystones for its diverse
pharmacological activities. Biomed Pharmacother. 109:1785–1792.
2019. View Article : Google Scholar
|
|
258
|
Xu S, Feng Y, He W, Xu W, Xu W, Yang H and
Li X: Celastrol in metabolic diseases: Progress and application
prospects. Pharmacol Res. 167:1055722021. View Article : Google Scholar : PubMed/NCBI
|
|
259
|
Trott A, West JD, Klaić L, Westerheide SD,
Silverman RB, Morimoto RI and Morano KA: Activation of heat shock
and antioxidant responses by the natural product celastrol:
Transcriptional signatures of a Thiol-targeted molecule. Mol Biol
Cell. 19:1104–1112. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
260
|
Tan Q, Liu Z, Gao X, Wang Y, Qiu X, Chen
J, Liang L, Guo H, Huang S, Wu D, et al: Celastrol recruits UBE3A
to recognize and degrade the DNA binding domain of steroid
receptors. Oncogene. 41:4754–4767. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
261
|
Gwag T, Park K, Kim E, Son C, Park J,
Nikawa T and Choi I: Inhibition of C2C12 myotube atrophy by a novel
HSP70 inducer, celastrol, via activation of Akt1 and ERK1/2
pathways. Arch Biochem Biophys. 537:21–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
262
|
Wastag M, Bieżuńska-Kusiak K, Szewczyk A,
Szlasa W, Grimling B and Kulbacka J: Celastrol and Rhynchophylline
in the mitigation of simulated muscle atrophy under in vitro. Saudi
Pharm J. 30:1387–1395. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
263
|
Kang L, Zhang Y, Zhou L, Yang J, He Y,
Yang S, Li G, Hao Q, Yu Y and Guo L: Structural characterization
and discrimination of Morinda officinalis and processing Morinda
officinalis based on metabolite profiling analysis. Front Chem.
9:8035502021. View Article : Google Scholar
|
|
264
|
Zhao S, Guo L, Cui W, Zhao Y, Wang J, Sun
K, Zhang H, Sun Y, Zhao D, Hu X, et al: Monotropein protects
mesenchymal stem cells from Lipopolysaccharide-induced impairments
and promotes fracture healing in an ovariectomized mouse model.
Calcif Tissue Int. 113:558–570. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
265
|
Shin JS, Yun KJ, Chung KS, Seo KH, Park
HJ, Cho YW, Baek NI, Jang D and Lee KT: Monotropein isolated from
the roots of Morinda officinalis ameliorates proinflammatory
mediators in RAW 264.7 macrophages and dextran sulfate sodium
(DSS)-induced colitis via NF-κB inactivation. Food Chem Toxicol.
53:263–271. 2013. View Article : Google Scholar
|
|
266
|
Wu M, Lai H, Peng W, Zhou X, Zhu L, Tu H,
Yuan K and Yang Z: Monotropein: A comprehensive review of
biosynthesis, physicochemical properties, pharmacokinetics, and
pharmacology. Front Pharmacol. 14:11099402023. View Article : Google Scholar : PubMed/NCBI
|
|
267
|
Wang P, Kang SY, Kim SJ, Park YK and Jung
HW: Monotropein improves Dexamethasone-induced muscle atrophy via
the AKT/mTOR/FOXO3a signaling pathways. Nutrients. 14:18592022.
View Article : Google Scholar : PubMed/NCBI
|
|
268
|
Shi Y, Liu XY, Jiang YP, Zhang JB, Zhang
QY, Wang NN and Xin HL: Monotropein attenuates oxidative stress via
Akt/mTOR-mediated autophagy in osteoblast cells. Biomed
Pharmacother. 121:1095662020. View Article : Google Scholar
|
|
269
|
Dembitsky VM and Maoka T: Allenic and
cumulenic lipids. Prog Lipid Res. 46:328–375. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
270
|
Guglielmi G, Lavaud J, Rousseau B, Etienne
AL, Houmard J and Ruban AV: The light-harvesting antenna of the
diatom Phaeodactylum tricornutum. Evidence for a
diadinoxanthin-binding subcomplex. FEBS J. 272:4339–4348. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
271
|
Pajot A, Hao Huynh G, Picot L, Marchal L
and Nicolau E: Fucoxanthin from algae to human, an extraordinary
bioresource: Insights and advances in up and downstream processes.
Mar Drugs. 20:2222022. View Article : Google Scholar : PubMed/NCBI
|
|
272
|
Miyashita K, Nishikawa S, Beppu F, Tsukui
T, Abe M and Hosokawa M: The allenic carotenoid fucoxanthin, a
novel marine nutraceutical from brown seaweeds. J Sci Food Agric.
91:1166–1174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Cui H, Zhu X, Yu X, Li S, Wang K, Wei L,
Li R and Qin S: Advancements of astaxanthin production in
Haematococcus pluvialis: Update insight and way forward.
Biotechnology Adv. 79:1085192025. View Article : Google Scholar
|
|
274
|
Nishida Y, Nawaz A, Hecht K and Tobe K:
Astaxanthin as a novel mitochondrial regulator: A new aspect of
carotenoids, beyond antioxidants. Nutrients. 14:1072021. View Article : Google Scholar
|
|
275
|
Donoso A, González-Durán J, Muñoz AA,
González PA and Agurto-Muñoz C: 'Therapeutic uses of natural
astaxanthin: An evidence-based review focused on human clinical
trials'. Pharmacol Res. 166:1054792021. View Article : Google Scholar
|
|
276
|
Jabarpour M, Aleyasin A, Shabani Nashtaei
M and Amidi F: Astaxanthin supplementation impact on insulin
resistance, lipid profile, blood pressure, and oxidative stress in
polycystic ovary syndrome patients: A triple-blind randomized
clinical trial. Phytother Res. 38:321–330. 2024. View Article : Google Scholar
|
|
277
|
EFSA Panel on Nutrition Novel Foods and
Food Allergens (NDA); Turck D, Castenmiller J, de Henauw S,
Hirsch-Ernst KI, Kearney J, Maciuk A, Mangelsdorf I, McArdle HJ, et
al: Safety of astaxanthin for its use as a novel food in food
supplements. EFSA J. 18:e059932020.PubMed/NCBI
|
|
278
|
Shah MMR, Liang Y, Cheng JJ and Daroch M:
Astaxanthin-producing green microalga Haematococcus pluvialis: From
single cell to high value commercial products. Front Plant Sci.
7:5312016. View Article : Google Scholar : PubMed/NCBI
|
|
279
|
Wong SK, Ima-Nirwana S and Chin KY:
Effects of astaxanthin on the protection of muscle health (review).
Exp Ther Med. 20:2941–2952. 2020.PubMed/NCBI
|
|
280
|
Lauer MS and Fontanarosa PB: Updated
guidelines for cholesterol management. JAMA. 285:2508–2509. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
281
|
Lin X, Racette SB, Lefevre M, Ma L,
Spearie CA, Steger-May K and Ostlund RE: Combined effects of
ezetimibe and phytosterols on cholesterol metabolism: A randomized,
controlled feeding study in humans. Circulation. 124:596–601. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
282
|
Khan Z, Nath N, Rauf A, Emran TB, Mitra S,
Islam F, Chandran D, Barua J, Khandaker MU, Idris AM, et al:
Multifunctional roles and pharmacological potential of
β-sitosterol: Emerging evidence toward clinical applications. Chem
Biol Interact. 365:1101172022. View Article : Google Scholar
|
|
283
|
Makhmudova U, Schulze PC, Lütjohann D and
Weingärtner O: Phytosterols and Cardiovascular Disease. Curr
Atheroscler Rep. 23:682021. View Article : Google Scholar : PubMed/NCBI
|
|
284
|
Pagano E, Laudato M, Griffo M and Capasso
R: Phytotherapy of benign prostatic hyperplasia. A minireview.
Phytother Res. 28:949–955. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
285
|
Wang H, Wang Z, Zhang Z, Liu J and Hong L:
β-Sitosterol as a promising anticancer agent for chemoprevention
and chemotherapy: Mechanisms of action and future prospects. Adv
Nutr. 14:1085–1110. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
286
|
Nandi S, Nag A, Khatua S, Sen S,
Chakraborty N, Naskar A, Acharya K, Calina D and Sharifi-Rad J:
Anticancer activity and other biomedical properties of
β-sitosterol: Bridging phytochemistry and current pharmacological
evidence for future translational approaches. Phytother Res.
38:592–619. 2024. View Article : Google Scholar
|
|
287
|
Li Q, Yang H, Song S, Liu J, Wang Z and
Wang J: Bioactive components in whole grains for the regulation of
skeletal muscle function. Foods. 11:27522022. View Article : Google Scholar : PubMed/NCBI
|
|
288
|
Cheng Y, Chen Y, Li J, Qu H, Zhao Y, Wen C
and Zhou Y: Dietary β-Sitosterol improves growth performance, meat
quality, antioxidant status, and mitochondrial biogenesis of breast
muscle in broilers. Animals. 9:712019. View Article : Google Scholar
|
|
289
|
Valitova J, Renkova A, Beckett R and
Minibayeva F: Stigmasterol: An enigmatic plant stress sterol with
versatile functions. Int J Mol Sci. 25:81222024. View Article : Google Scholar : PubMed/NCBI
|
|
290
|
Hah YS, Lee SJ, Ji YH, Hwang J, Kim HG, Ju
YT, Yoo JI and Kwag SJ: Stigmasterol protects against
Dexamethasone-induced muscle atrophy by modulating the
FoxO3-MuRF1/MAFbx signaling pathway in C2C12 myotubes and mouse
skeletal muscle. Biomolecules. 15:15512025. View Article : Google Scholar : PubMed/NCBI
|
|
291
|
Li J, Wei S, Marabada D, Wang Z and Huang
Q: Research progress of natural matrine compounds and synthetic
matrine derivatives. Molecules. 28:57802023. View Article : Google Scholar : PubMed/NCBI
|
|
292
|
Ali S, Zhang C, Wang Z, Wang XM, Wu JH,
Cuthbertson AGS, Shao Z and Qiu BL: Toxicological and biochemical
basis of synergism between the entomopathogenic fungus
Lecanicillium muscarium and the insecticide matrine against Bemisia
tabaci (Gennadius). Sci Rep. 7:465582017. View Article : Google Scholar : PubMed/NCBI
|
|
293
|
Li X, Tang Z, Wen L, Jiang C and Feng Q:
Matrine: A review of its pharmacology, pharmacokinetics, toxicity,
clinical application and preparation researches. J Ethnopharmacol.
269:1136822021. View Article : Google Scholar
|
|
294
|
Zhang H, Chen L, Sun X, Yang Q, Wan L and
Guo C: Matrine: A promising natural product with various
pharmacological activities. Front Pharmacol. 11:5882020. View Article : Google Scholar : PubMed/NCBI
|
|
295
|
Lin Y, He F, Wu L, Xu Y and Du Q: Matrine
exerts pharmacological effects through multiple signaling pathways:
A comprehensive review. Drug Des Devel Ther. 16:533–569. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
296
|
Chen L, Chen L, Wan L, Huo Y, Huang J, Li
J, Lu J, Xin B, Yang Q and Guo C: Matrine improves skeletal muscle
atrophy by inhibiting E3 ubiquitin ligases and activating the
Akt/mTOR/FoxO3α signaling pathway in C2C12 myotubes and mice. Oncol
Rep. 42:479–494. 2019.PubMed/NCBI
|
|
297
|
Yi F, Tan X, Yan X and Liu H: In silico
profiling for secondary metabolites from Lepidium meyenii (maca) by
the pharmacophore and ligand-shape-based joint approach. Chin Med.
11:422016. View Article : Google Scholar : PubMed/NCBI
|
|
298
|
Zheng BL, He K, Kim CH, Rogers L, Shao Y,
Huang ZY, Lu Y, Yan SJ, Qien LC and Zheng QY: Effect of a lipidic
extract from lepidium meyenii on sexual behavior in mice and rats.
Urology. 55:598–602. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
299
|
Chae J, Hahn D and Nam JO: Macamide, a
component of maca (Lepidium meyenii Walp) lipophilic extract,
enhances myogenic differentiation via AKT/p38 signaling and
attenuates dexamethasone-induced muscle atrophy. Biomed
Pharmacother. 172:1162492024. View Article : Google Scholar : PubMed/NCBI
|
|
300
|
Yang K, Qiu J, Huang Z, Yu Z, Wang W, Hu H
and You Y: A comprehensive review of ethnopharmacology,
phytochemistry, pharmacology, and pharmacokinetics of Schisandra
chinensis (Turcz.) Baill. and Schisandra sphenanthera Rehd. et
Wils. J Ethnopharmacol. 284:1147592022. View Article : Google Scholar
|
|
301
|
Fu K, Zhou H, Wang C, Gong L, Ma C, Zhang
Y and Li Y: A review: Pharmacology and pharmacokinetics of
Schisandrin A. Phytother Res. 36:2375–2393. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
302
|
Kim CH, Shin JH, Hwang SJ, Choi YH, Kim DS
and Kim CM: Schisandrae fructus enhances myogenic differentiation
and inhibits atrophy through protein synthesis in human myotubes.
Int J Nanomedicine. 11:2407–2415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
303
|
Yeon M, Choi H and Jun HS: Preventive
effects of schisandrin A, A bioactive component of Schisandra
Chinensis, on Dexamethasone-induced muscle atrophy. Nutrients.
12:12552020. View Article : Google Scholar : PubMed/NCBI
|
|
304
|
Lo YH, Chen YJ, Chang CI, Lin YW, Chen CY,
Lee MR, Lee VS and Tzen JT: Teaghrelins, unique acylated flavonoid
tetraglycosides in Chin-shin oolong tea, are putative oral agonists
of the ghrelin receptor. J Agric Food Chem. 62:5085–5091. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
305
|
Jhuo CF, Hsieh SK, Chen CJ, Chen WY and
Tzen JTC: Teaghrelin protects SH-SY5Y cells against MPP+-induced
neurotoxicity through activation of AMPK/SIRT1/PGC-1α and ERK1/2
pathways. Nutrients. 12:36652020. View Article : Google Scholar
|
|
306
|
Mahn A and Castillo A: Potential of
sulforaphane as a natural immune system enhancer: A review.
Molecules. 26:7522021. View Article : Google Scholar :
|
|
307
|
Cuadrado A, Pajares M, Benito C,
Jiménez-Villegas J, Escoll M, Fernández-Ginés R, Garcia Yagüe AJ,
Lastra D, Manda G, Rojo AI and Dinkova-Kostova AT: Can activation
of NRF2 Be a strategy against COVID-19? Trends Pharmacol Sci.
41:598–610. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
308
|
Tortorella SM, Royce SG, Licciardi PV and
Karagiannis TC: Dietary sulforaphane in cancer chemoprevention: The
role of epigenetic regulation and HDAC inhibition. Antioxid Redox
Signal. 22:1382–1424. 2015. View Article : Google Scholar :
|
|
309
|
Otoo RA and Allen AR: Sulforaphane's
multifaceted potential: From neuroprotection to anticancer action.
Molecules. 28:69022023. View Article : Google Scholar : PubMed/NCBI
|
|
310
|
Mthembu SXH, Mazibuko-Mbeje SE, Moetlediwa
MT, Muvhulawa N, Silvestri S, Orlando P, Nkambule BB, Muller CJF,
Ndwandwe D, Basson AK, et al: Sulforaphane: A nutraceutical against
diabetes-related complications. Pharmacol Res. 196:1069182023.
View Article : Google Scholar : PubMed/NCBI
|
|
311
|
Vargas-Mendoza N, Madrigal-Santillán E,
Álvarez-González I, Madrigal-Bujaidar E, Anguiano-Robledo L,
Aguilar-Faisal JL, Morales-Martínez M, Delgado-Olivares L,
Rodríguez-Negrete EV, Morales-González Á and Morales-González JA:
Phytochemicals in skeletal muscle health: Effects of curcumin (from
Curcuma longa Linn) and Sulforaphane (from Brassicaceae) on muscle
function, recovery and therapy of muscle atrophy. Plants (Basel).
11:25172022.PubMed/NCBI
|
|
312
|
Lilley T, Camera DM and Kwa FAA: Repairing
muscle with broccoli-derived sulforaphane: A preclinical evaluation
for the treatment of mitochondrial myopathies. Drug Discovery
Today. 30:1042832025. View Article : Google Scholar
|
|
313
|
Son YH, Jang EJ, Kim YW and Lee JH:
Sulforaphane prevents dexamethasone-induced muscle atrophy via
regulation of the Akt/Foxo1 axis in C2C12 myotubes. Biomed
Pharmacother. 95:1486–1492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
314
|
Kim J, Lee SH, Cho M, Lee JY, Choi DH, Lee
HY, Cho S, Min KJ and Suh Y: Small molecule from natural
phytochemical mimics dietary restriction by modulating FoxO3a and
metabolic reprogramming. Adv Biosyst. 4:e19002482020. View Article : Google Scholar : PubMed/NCBI
|
|
315
|
Gupta A, Haldhar R, Agarwal V, Rajput DS,
Chun KS, Han SB, Raj V and Lee S: Exploring the potential of
natural products as FoxO1 inhibitors: An in silico approach. Biomol
Ther (Seoul). 32:390–398. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
316
|
Guan H, Zhang W, Liu H, Jiang Y, Li F,
Wang D, Liu Y, He F, Wu M, Ivan Neil Waterhouse G, et al:
Simultaneous binding of quercetin and catechin to FOXO3 enhances
IKKα transcription inhibition and suppression of oxidative
stress-induced acute alcoholic liver injury in rats. J Adv Res.
67:71–92. 2025. View Article : Google Scholar :
|
|
317
|
Patel MN, Halling-Brown MD, Tym JE,
Workman P and Al-Lazikani B: Objective assessment of cancer genes
for drug discovery. Nat Rev Drug Discov. 12:35–50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
318
|
Chen S, Zheng Y, Liang B, Yin Y, Yao J,
Wang Q, Liu Y and Neamati N: The application of PROTAC in HDAC. Eur
J Med Chem. 260:1157462023. View Article : Google Scholar : PubMed/NCBI
|
|
319
|
Cornu M, Lemaitre T, Kieffer C and
Voisin-Chiret AS: PROTAC 2.0: Expanding the frontiers of targeted
protein degradation. Drug Discovery Today. 30:1043762025.
View Article : Google Scholar : PubMed/NCBI
|
|
320
|
Liu J, Chen H, Kaniskan HÜ, Xie L, Chen X,
Jin J and Wei W: TF-PROTACs Enable targeted degradation of
transcription factors. J Am Chem Soc. 143:8902–8910. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
321
|
Gao P, Zhong Y, Luo K and Jin J: Bridged
PROTAC, DNA-based PROTAC and hydrophobic tag technologies for
targeted protein degradation. Adv Drug Deliv Rev. 232:1158192026.
View Article : Google Scholar : PubMed/NCBI
|
|
322
|
He A, Koszegi B, Uzun S, Bilgic A, Bozca
BC, Yang B, Daneshpazhooh M, Boziou M, Patsatsi A, Kakuta R, et al:
Autoimmune blistering diseases treated with glucocorticoids: An
international study of steroid-induced myopathy. J Eur Acad
Dermatol Venereol. 39:340–349. 2025. View Article : Google Scholar
|
|
323
|
Nakao R, Yamamoto S, Yasumoto Y and Oishi
K: Dosing schedule-dependent attenuation of dexamethasone-induced
muscle atrophy in mice. Chronobiol Int. 31:506–514. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
324
|
Van Der Heide LP, Hoekman MFM and Smidt
MP: The ins and outs of FoxO shuttling: Mechanisms of FoxO
translocation and transcriptional regulation. Biochem J.
380:297–309. 2004. View Article : Google Scholar : PubMed/NCBI
|