Introduction
Systemic lupus erythematosus (SLE) is a prototypical
chronic autoimmune disease. It is characterized by dysregulated
adaptive immunity, leading to systemic inflammation and
immune-mediated damage affecting multiple organ systems, including
the mucocutaneous, musculoskeletal, hematological and renal
systems. Lupus nephritis (LN) is a severe complication of SLE that
can progress to glomerulosclerosis, interstitial fibrosis and renal
failure in untreated patients (1). Clinical management of LN primarily
relies on immunosuppressive therapies, such as corticosteroids
(2), cyclophosphamide (3), mycophenolate mofetil/mycophenolic
acid (3) and calcineurin
inhibitors (4). However, these
regimens achieve complete remission in 20-30% of patients after 6
months of standard treatment (5), highlighting a notable unmet
therapeutic need. This suboptimal response rate underscores the
need for novel pharmacotherapies that target the underlying
pathogenic mechanisms of LN.
SLE is driven by a cascade of aberrant immune
responses, including hyperactivation of B and T cells, overactive
type I IFN signaling and neutrophil dysfunction. The disease is
typically associated with the accumulation of endogenous
immunogenic chromatin, which activates innate immune sensors via
toll-like receptors and simultaneously promotes the expansion of
chromatin-reactive B cells (6).
This dual mechanism sustains B cell activation and leads to the
production of anti-DNA and antinuclear autoantibodies. In SLE,
elevated levels of soluble B cell activating factor (BAFF) drive B
cell activation and autoantibody production. Consequently,
therapeutic strategies targeting BAFF or depleting B cells have
demonstrated clinical efficacy by decreasing autoreactive B cell
populations. Notably, genetic and functional studies have revealed
conserved signaling defects in SLE B cells, marked by dysregulation
of key kinases (such as Lyn) (7,8)
and phosphatases (such as CD22 and Src homology 2 domain-containing
protein tyrosine phosphatase 1) (9). In lupus-prone mouse models, loss of
these regulatory molecules results in B cell hyperactivation,
autoantibody production and lupus-like nephritis (7-9),
whereas pharmacological restoration of their expression attenuates
disease progression (10,11).
RNA sequencing (RNA-seq) is a tool for profiling
gene expression in immune cells, enabling the identification of
dysregulated pathways, such as type I IFN and B cell receptor
signaling, and the discovery of druggable targets. Single-cell
(sc)RNA-seq improves resolution by identifying cell types within a
tissue and identifying rare pathogenic subsets, such as
IFN-responsive plasmablasts, which may serve as precision medicine
endpoints. Recent advances in scRNA-seq enable unbiased
characterization of immune cell diversity and revealing potential
therapeutic targets (12). For
example, Arazi et al (12) applied scRNA-seq to identify 21
distinct leukocyte subsets in patients with SLE, including diverse
populations of myeloid cells, T, natural killer and B cells. The
aforementioned study identified two broadly expressed chemokine
receptors as potential therapeutic nodes. Moreover, scRNA-seq
enables the reconstruction of functional intercellular networks by
resolving gene expression landscapes at single-cell resolution,
offering insights into the pathogenic roles of B cells and their
microenvironmental crosstalk.
Drug repurposing offers a cost-effective strategy to
accelerate drug development while decreasing the financial burden
of de novo discovery. Moreover, in silico analysis of
gene expression signatures is effective in identifying new
therapeutic applications for existing compounds (13). The Connectivity Map (CMAP;
clue.io/) is a platform that links disease transcriptomes with gene
perturbations and drug mechanisms of action. The CMAP database has
been utilized in drug repurposing for cancer, neurodegeneration and
metabolic disorders (14). The
present study analyzed transcriptomic profiles of peripheral B
lymphocytes and compared them with the CMAP database. As a
bioactive steroidal lactone extracted from Withania
somnifera, Withaferin A (WA) exhibits a broad spectrum of
pharmacological properties, most notably anti-inflammatory
(15), antioxidant (16), and anticancer activities
(17). Its relevance in
therapeutic interventions has been increasingly documented,
primarily through its ability to target inflammation (15) and oxidation (16). Therefore, characterization of its
effects on LN is vital to elucidate its full biomedical value.
Materials and methods
Reagents and antibodies
WA (cat. no. HY-N2065), DMSO (cat. no. HY-Y0320) and
lipopolysaccharide (cat. no. HY-D1056) were purchased from
MedChemExpress. The antibodies were as follows: Mouse α-smooth
muscle active (SMA; cat. no. ab7817), TNF-α (cat. no. ab183218),
IL-1β (cat. no. ab283818), kidney injury molecule-1 (KIM-1; cat.
no. ab47635), paraoxonase 1 (Pon1; cat. no. ab92466), peroxisome
proliferator-activated receptor (PPAR)-α (cat. no. ab126285),
PPAR-γ (cat. no. ab45036), Bax (cat. no. ab32503), BCL-2 (cat. no.
ab182858) and GAPDH (cat. no. ab8245; all Abcam). The mouse
anti-nuclear antibody (ANA)/extractable nuclear antigen (ENA) Igs
(total A + G + M; cat. no. 5210) and mouse anti-double-stranded DNA
Igs (total A+G+M) ELISA kit (cat. no. 5110) were obtained from
Alpha Diagnostic Intl Inc.
CMAP assay
To identify potential therapeutics for LN, scRNA-seq
data derived from human peripheral blood were obtained from the
Gene Expression Omnibus database. (ncbi.nlm.nih.gov/gds). Differentially expressed genes
(DEGs) in B lymphocytes from GSE135779, GSE162577 and GSE142016
were identified by comparing patients with SLE with healthy donors
(adjusted P<0.05). To represent the disease-specific expression
profile, the top 150 upregulated and top 150 downregulated genes
were selected as the gene signature for the CMAP/L1000 database
using the CLUE web platform (https://clue.io/),
based on their rank in log2 Fold Change magnitude.
Connectivity scores were calculated and the highest-ranking
compounds from the CMAP/L1000 analysis were selected for further
investigation.
Animal model and experimental design
MRL/lpr mice (n=10; weight, 27.71±0.86 g) were
purchased from Changzhou Cavens Laboratory Animal Co., Ltd. and
maintained under specific pathogen-free conditions at the Animal
Center of Gansu University of Chinese Medicine (Lanzhou, China).
Mice were housed at 22±2°C and relative humidity of 50±10%, with a
12/12-h light/dark cycle. Female mice (age, 8 weeks) were randomly
allocated into two groups (n=5 per group). The treatment group
received intraperitoneal injection of 2 mg/kg WA dissolved in DMSO
and the control group was treated with 2% DMSO once/day. All
animals had free access to food and water throughout the study.
After 8 weeks of treatment, mice were euthanized by CO2
asphyxiation with a chamber fill rate of 60% of total volume/min,
in accordance with the AVMA Guidelines for the Euthanasia of
Animals (18), followed by
sample collection for downstream analyses including spleen, kidney
and lymph gland. All experimental procedures were approved by the
Animal Experimental Ethical Inspection of Gansu University of
Chinese Medicine (approval no. SY2024-257) and performed in
compliance with institutional guidelines for animal welfare and
use.
Renal function
According to the manufacturer's instructions, serum
creatinine (cat. no. EIASCR) and urea nitrogen (cat. no. BC1535)
were measured using commercial kits from Thermo Fisher Scientific,
Inc. and Beijing Solarbio Science & Technology Co., Ltd.,
respectively. Urinary protein levels were determined using the
Pierce™ Bradford Plus Protein Assay kit (cat. no. 23236; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
protocol.
ELISA for ANAs and anti-double-stranded
(ds)DNA antibodies
The concentrations of ANA and dsDNA antibodies in
mouse serum were quantified using the aforementioned ELISA kits,
following the manufacturer's instructions. In brief, samples and
controls were incubated at 37°C for 60 min. HRP-conjugated
anti-mouse Ig horseradish peroxidase (1:100) and
tetramethylbenzidine substrate were added for 30 and 15 min at
37°C, respectively. Absorbance was measured at 450 nm using a
microplate reader.
Histology
Tissue samples were fixed in 4% paraformaldehyde
prepared in 1X phosphate-buffered saline (pH 7.5) at room
temperature overnight, dehydrated through a graded series of
ethanol and embedded in paraffin. 3-μm-thick sections were
stained with hematoxylin and eosin (H&E) and periodic
acid-Schiff (PAS) using commercial staining kits (cat. no. G1120
and G1281 for PAS, respectively; both Solarbio) at room temperature
(hematoxylin for 5 min and eosin for 2 min. For PAS staining,
sections were oxidized in periodic acid for 15 min, incubated with
Schiff's reagent for 15 min in the dark, and counterstained with
hematoxylin for 1 min. to evaluate renal structural damage and
pathological changes. The pathological changes in the tissue were
observed under the light microscope (Zeiss Axio Imager M2) and
analyzed by ImageJ (version 1.53, National Institutes of Health).
Glomerulosclerosis was assessed using a semi-quantitative scale as
follows: 0, normal; 1, mesangial expansion and slight glomerular
damage involving <25% of the glomerulus; 2, mild sclerosis
involving 25-49% of the glomerulus; 3, moderate sclerosis involving
50-74% of the glomerulus; and 4, severe sclerosis involving ≥75% of
the glomerulus (19).
Immunofluorescence
For the detection of glomerular immune complex
deposition, kidney tissues were embedded in OCT compound and frozen
at −20°C. Frozen sections with a thickness of 5 μm were
prepared. Following being blocking with 5% bovine serum albumin
(HY-D0842, MedChemExpress) for 1 h at room temperature, the
sections were incubated with primary antibodies, including rabbit
anti-complement C3 (1:200; cat. no. ab97462; Abcam), overnight at
4°C. Sections were incubated with secondary antibodies, such as
Cy5-conjugated goat anti-mouse IgG (H+L; 1:500; cat. no. 771421ES;
Yeasen) or Alexa Fluor-labeled antibodies (both 1:500; cat. no.
A-11012; Thermo Fisher Scientific), for 1 h at room temperature.
Images were captured using a microscope (LSM 800, Zeiss) and
analyzed using ImageJ (version 1.53, National IH). Semiquantitative
analysis of glomerular complement C3 deposition was performed as
follows: 0, no staining; 1, barely detectable at high
magnification; 2, moderately visible; and 3, strongly and clearly
visible (20).
Immunohistochemistry (IHC)
Renal α-SMA expression was evaluated via IHC.
Briefly, kidney tissues were fixed in 4% paraformaldehyde for 24 h
at 4°C and embedded in paraffin. Sections were cut at a thickness
of 3-5 μm. Following deparaffinization in xylene and
rehydration through a descending alcohol series, antigen retrieval
was performed by heating in 10 mM citrate buffer, pH 6.0 at 95°C
for 15 min. To block endogenous peroxidase activity, sections were
treated with 3% H2O2 for 10 min at room
temperature. Following blocking with 5% normal goat serum (Beijing
Solarbio) for 1 h at room temperature, sections were incubated with
primary anti-α-SMA antibodies (1:2,000; cat. no. ab124964; Abcam)
overnight at 4°C. Following four washes in PBS, sections were
incubated with HRP-conjugated secondary antibodies (1:500; cat. no.
GB23303; Servicebio) for 1 h at room temperature. The signal was
developed using a DAB substrate kit and counterstained with
hematoxylin for 2 min at room temperature. Images were acquired
using the light microscope (Zeiss Axio Imager M2). Positive
staining area was measured using ImageJ software (version
1.53).
RNA-seq
To prepare RNA-seq samples, kidneys were harvested
from the SLE model mice following treatment with either DMSO or WA.
All tissues were immediately snap-frozen in liquid nitrogen and
stored at −80°C. Total RNA was extracted using the GeneJET RNA
Purification kit (cat. no. K0731l Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions, followed by DNase I
treatment to eliminate genomic DNA contamination. RNA quality and
quantity were assessed using an Agilent 2100 Bioanalyzer (Agilent
Technologies) to ensure an RNA Integrity Number >7.0 prior to
cDNA library construction. Sequencing was performed on an DNBSEQ-T7
platform (MGI, Inc.) by Sangon Biotech Co., Ltd. The sequencing is
performed in 150 bp paired end using DNBSEQ DNB Reagent Kit (cat.
no. 1000028453; MGI Inc.). The loading concentration of the final
library is 18.6 pM. Transcript abundance was quantified using the
transcripts per million methods, which normalizes gene expression
based on exon models, enabling cross-gene and cross-experiment
comparability. Differential expression analysis was performed using
the DESeq2 R package (v1.12.4) (https://www.r-project.org), applying a significance
threshold of |fold-change|>2 and an adjusted P-value of
<0.05. Based on the differentially expressed genes (DEGs),
hierarchical clustering and heatmap visualization were performed
using R software gplots package (2.17.0). Functional annotation of
DEGs was performed using Gene Ontology (GO) (21) enrichment analysis using R
software topGO package (2.24.0) (22). Pathway enrichment analysis was
performed using the Kyoto Encyclopedia of Genes and Genomes (KEGG)
(21) via R software
clusterProfiler package (3.0.5) (23). Terms with a P-value <0.05 were
considered significantly enriched.
Primary B cell isolation
For primary B cell isolation, spleens were obtained
from MRL/lpr mice. Each spleen was placed in a 70-μm cell
strainer and dissociated using a syringe plunger while rinsing with
RPMI-1640 medium (cat. no. 11875093; Gibco; Thermo Fisher
Scientific, Inc.) to collect cells into a tube. Following
centrifugation at 300 × g for 5 min at 4°C, erythrocytes were lysed
by resuspending the pellet in erythrocyte lysis buffer (cat. no.
R1010; Solarbio) for 15 min at room temperature. The cells were
collected at 450 × g for 10 min at 4°C and then washed, resuspended
in PBS and adjusted to 1×107 cells/ml. Cells were
incubated with CD19 MicroBeads (cat. no. 130-121-301; Miltenyi
Bio.; 10 μl/1×107 cells) for 15 min at 4°C and
CD19+ B cells were isolated using a magnetic separation
column (cat. no. 130-042-401; Miltenyi Bio.) according to the
introductions.
Cell culture and treatment
Human proximal tubular human kidney-2 (HK-2) cells
(cat. no. CTCC-002-0018; MeisenCTCC) were cultured in DMEM
supplemented with 10% fetal bovine serum (both Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin streptomycin, under a
humidified atmosphere of 5% CO2 at 37°C. To investigate
the therapeutic potential of WA, HK-2 cells were pretreated with
2.5 μM WA for 1 h at 37°C prior to the induction of
inflammation using 40 μg/ml lipopolysaccharide (LPS) for 48
h at 37°C.
Cell transfection
To inhibit PON1 expression, HK-2 cells were
transfected with either PON1-specific small interfering RNA
(si-PON1; Table SI) or a PON1
overexpression plasmid (Ribobio). Cells were transfected with 1
pmol siRNA using Lipofectamine™ RNAiMAX (cat. no. 13778100; Thermo
Fisher Scientific, Inc.) at 37°C for 24 h before the medium was
replaced with fresh complete medium and treated with 1 μg/ml
LPS at 37°C for 24 h. For overexpression, the human PON1
full-length cDNA was cloned into the pcDNA3.1 plasmid vector
(Ruibiotech). Cells were transfected with 0.2 μg of either
the PON1 plasmid or the empty vector using Lipofectamine™ 3000
(Thermo Fisher Scientific, Inc.) at 37°C for 48 h. For PON1
activation, cells were exposed to 5 μM zinc phytate (ZnPA;
cat. no. 529505; Sigma-Aldrich) for 48 h at 37°C followed by 1
μg/ml LPS exposure for 24 h at 37°C. Cells were subsequently
harvested for downstream protein or RNA analysis.
Cell viability assay
HK-2 cells were seeded into 96-well plates at a
density of 1×104 cells/well. Following the respective
treatments, the culture medium was aspirated and replaced with 100
μl of fresh medium containing Cell Counting Kit-8 (CCK-8)
solution (cat. no. CA1210, Solarbio) in serum-free RPMI-1640. The
plates were then incubated at 37°C for 1-4 h in the dark, according
to the manufacturer's instructions. The absorbance at 450 nm was
measured using a microplate reader.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from kidney tissues or
cultured cells using the GeneJET RNA Purification kit (cat. no.
K0732; Thermo Fisher Scientific, Inc.). cDNA was synthesized from
500 ng of total RNA using the RevertAid First Strand cDNA Synthesis
kit (cat. no. K1622; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. qPCR was performed in 20 μl
reaction mixtures containing 2 μl of cDNA, 0.3 μM of
each gene-specific primer (Table
SII) and 10 μl of SYBR Green qPCR Master Mix (cat. no.
K0251; Thermo Fisher Scientific, Inc.). Amplification was carried
out on an ABI StepOnePlus™ Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.) under the following
thermocycling conditions: Initial denaturation at 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 30 sec. All
reactions were performed in triplicate. The relative levels of
target genes were calculated using the 2−ΔΔCq method
(24), normalized to GAPDH.
Measurement of reactive oxygen species
(ROS) levels
ROS levels were measured using a commercial kit
(cat. no. EEA019; Thermo Fisher Scientific, Inc.). Briefly,
following ZnPA treatments or PON1 overexpression, cells were loaded
with 10 μM DCFH-DA in serum-free DMEM (Gibco; Thermo Fisher
Scientific, Inc.) and incubated at 37°C for 30 min in the dark.
Cells were thoroughly washed with PBS to remove excess probe and
resuspended with serum-free medium for detection. The fluorescence
intensity was measured using a microplate reader
(excitation/emission, 488/525 nm). Results are expressed as a
percentage relative to the untreated control group.
Measurement of antioxidative enzyme
activity and malondialdehyde (MDA) levels
Lactate dehydrogenase (LDH) activity was assessed
using a commercial kit (cat. no. BC0625; Beijing Solarbio Science
& Technology Co., Ltd.) according to the manufacturer's
protocol. Following si-PON1 transfection and incubation with WA for
24 h at 37°C, the culture supernatant was mixed with the substrate
solution. The mixture was incubated in the dark at room temperature
for 30 min. After adding the stop solution, the absorbance was
measured at 450 nm using a microplate reader. LDH activity was
calculated based on the standard curve. Intracellular levels of MDA
and superoxide dismutase (SOD) were assessed using commercial assay
kits (cat. nos. BC6415 and BC5165, respectively; Beijing Solarbio
Science & Technology Co., Ltd.) according to the manufacturer's
instructions. The absorbance for each assay was measured at 532 and
600 nm for MDA or 450 nm for SOD using a microplate reader.
Western blotting
Total protein was extracted from kidney tissue or
cultured cells using RIPA lysis buffer (cat. no. R0010; Beijing
Solarbio) supplemented with protease and phosphatase inhibitors
(cat. no. 78440; Thermo Fisher Scientific), and the protein
concentration was quantified using the Bradford assay (cat. no.
PC0010; Solarbio). A total of 20 μg of protein lysate were
separated via 4-20% SDS-PAGE and transferred onto PVDF membranes.
The membranes were blocked with 5% non-fat milk for 1 h] at room
temperature and incubated overnight at 4°C with anti-PON1 (1:1,000;
cat. no. ab92466; Abcam), anti-TNF-α (1:1,000; cat. no. ab183218;
Abcam), anti-KIM-1 (1:1,000; cat. no. ab323414; Abcam), anti-IL-1β
(1:1,000; cat. no. ab283818; Abcam), anti-Bcl-2 (1:1,000; cat. no.
ab182858; Abcam), anti-Bax (1:1,000; cat. no. ab32503; Abcam),
anti-PPAR-α (1:1,000; cat. no. ab61182; Abcam), anti-PPARγ
(1:1,000; cat. no. ab45036; Abcam) and anti-GAPDH (1:2,000; cat.
no. ab8245; Abcam) as the internal reference control. After washing
three times with TBST with 0.1% Tween-20, the membranes were
incubated with HRP-conjugated secondary antibodies (1:5,000; cat.
no. HA1001; HuaBio) for 1 h at room temperature. Protein bands were
visualized using an ECL detection kit (cat. no. 32209; Thermo
Fisher Scientific, Inc.). Quantification of the protein bands was
performed using ImageJ software (version 1.54g).
Statistical analysis
Data are expressed as the mean ± standard error of
the mean of ≥3 independent experimental repeats. Data were analyzed
using one-way ANOVA followed by Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant difference.
All statistical analyses were performed using GraphPad Prism 8
(Dotmatics).
Results
WA is a candidate for treating
SLE-associated LN
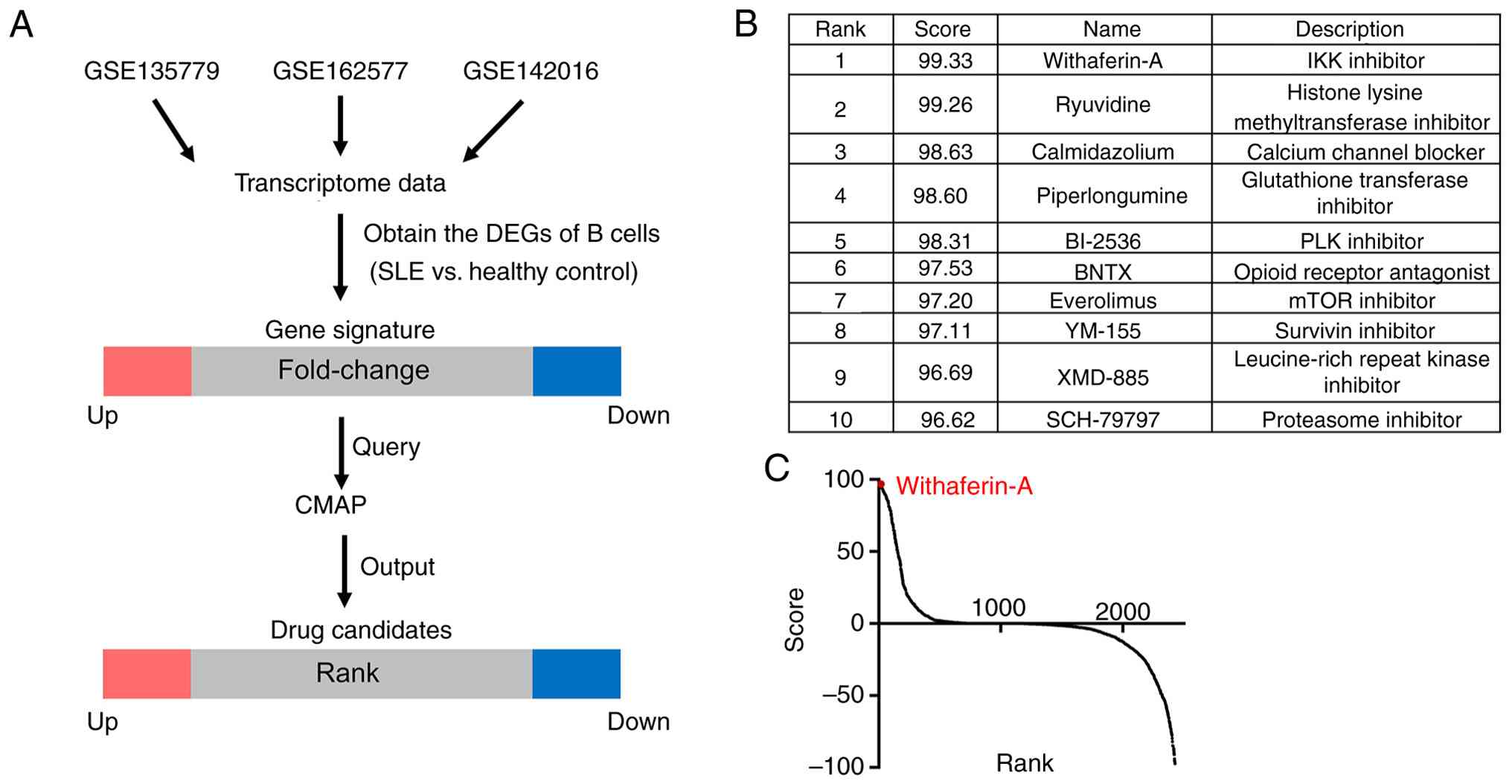
To identify potential therapeutic agents for
SLE-associated LN, the present study analyzed transcriptomic
profiles of peripheral immune cells from the public datasets
GSE135779, GSE162577 and GSE142016. Given the key role of B cells
in SLE pathogenesis (25), DEGs
were identified in B lymphocytes from patients with SLE compared
with those from healthy donors (adjusted P<0.05). Moreover, the
top 150 up-and downregulated genes, ranked by log2
fold-change, were selected as gene signatures (Table SIII) and submitted to the CMAP
platform to screen candidate compounds (Fig. 1A). All output compounds were
ranked by CMAP score and the top 10 candidates were identified
(Fig. 1B). Among these, WA
ranked first (Fig. 1C),
suggesting its potential as a therapeutic agent for SLE-associated
LN.
WA improves renal function in
experimental LN
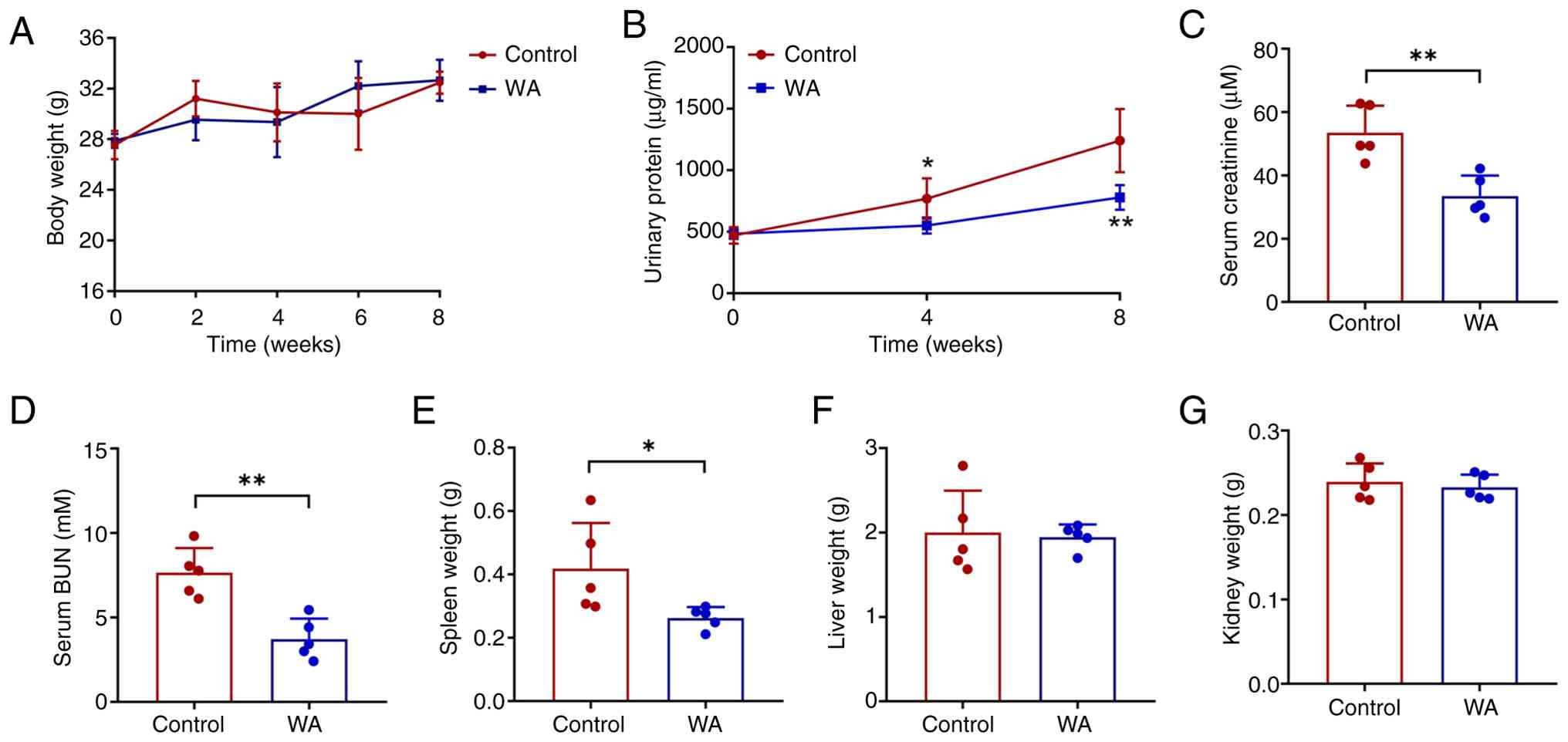
The present study assessed the effects of WA on
SLE-associated LN in MRL/lpr mice. Following 8 week treatment, mice
administered WA showed no significant difference in body weight
compared with that of the control group (Fig. 2A). However, WA significantly
improved renal function in lupus-prone mice, as demonstrated by
significant decreases in urinary protein excretion and serum
creatinine levels (Fig. 2B and
C). Additionally, a significant decrease in blood urea nitrogen
was observed in the WA-treated group (Fig. 2D). Notably, spleen weight was
significantly lower in WA-treated mice, whereas liver and kidney
weight remained comparable between the two groups (Fig. 2E-G).
WA ameliorates splenic immune cell
dysfunction in LN
SLE involves splenic immune cell dysfunction that
contributes to multi-organ pathology (26). SLE model mice displayed marked
splenomegaly and enlarged axillary lymph nodes, both of which were
attenuated by WA treatment (Fig.
3A). WA administration downregulated the splenic expression of
pro-inflammatory cytokines, including monocyte chemoattractant
protein (MCP)-1, TNF-α and IL-1β (Fig. 3B-D), while upregulating the
anti-inflammatory transcription factor forkhead box P3 (Fig. 3E). WA treatment significantly
decreased serum levels of ANA and dsDNA autoantibodies (Fig. 3F and G). Collectively, these
results demonstrated that WA ameliorates splenic immune dysfunction
in murine lupus.
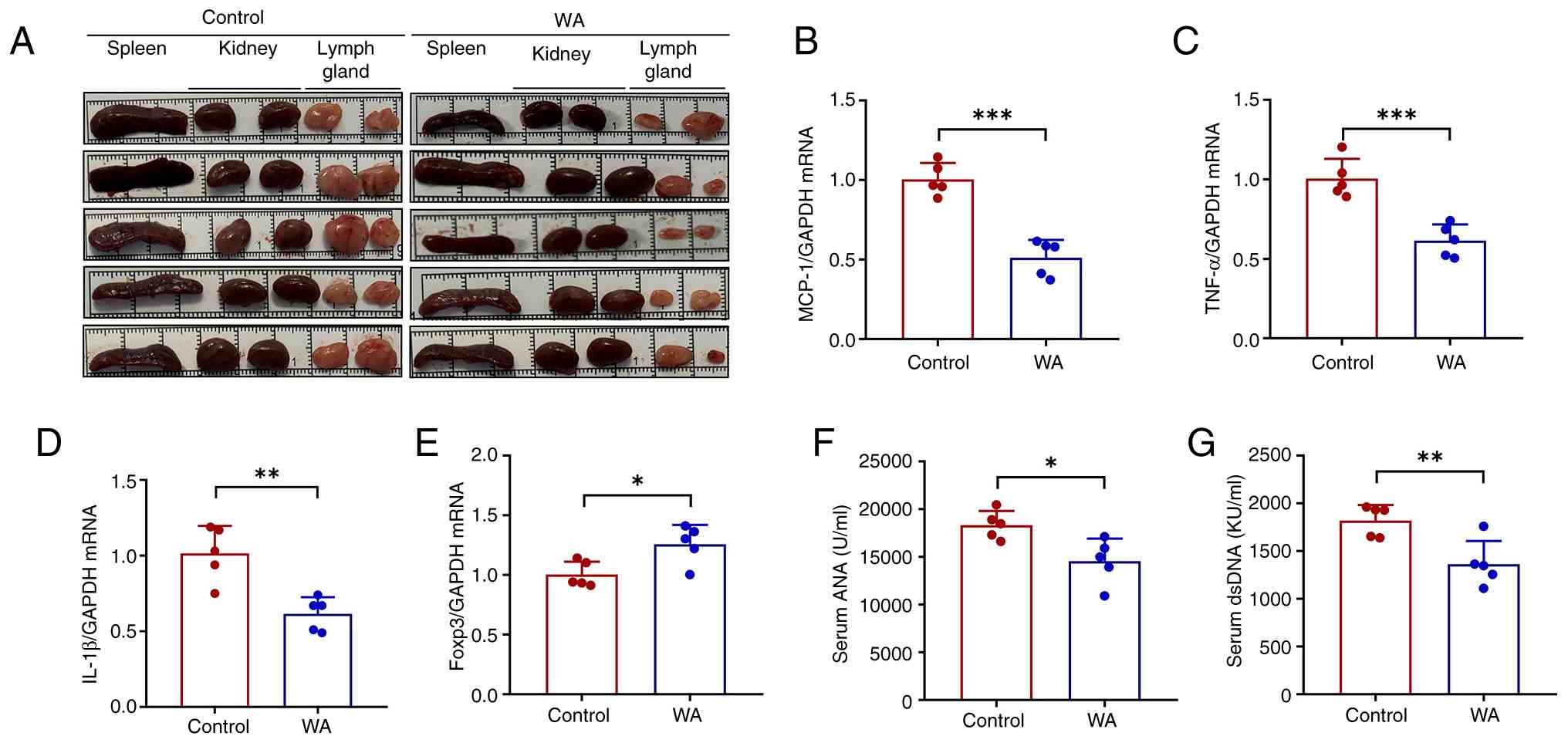 | Figure 3WA attenuates splenic immune cell
dysfunction in lupus nephritis. (A) Representative spleen, lymph
node and kidneys from lupus mice in the presence or absence of WA
treatment. Splenic mRNA expression of (B) MCP-1, (C) TNF-α, (D)
IL-1β and (E) Foxp3. Serum (F) ANA and (G) dsDNA levels.
*P<0.05, **P<0.01,
***P<0.001. WA, withaferin A; MCP, monocyte
chemoattractant protein; KU, kilo unit; ANA, antinuclear antibody;
ds, double-stranded. |
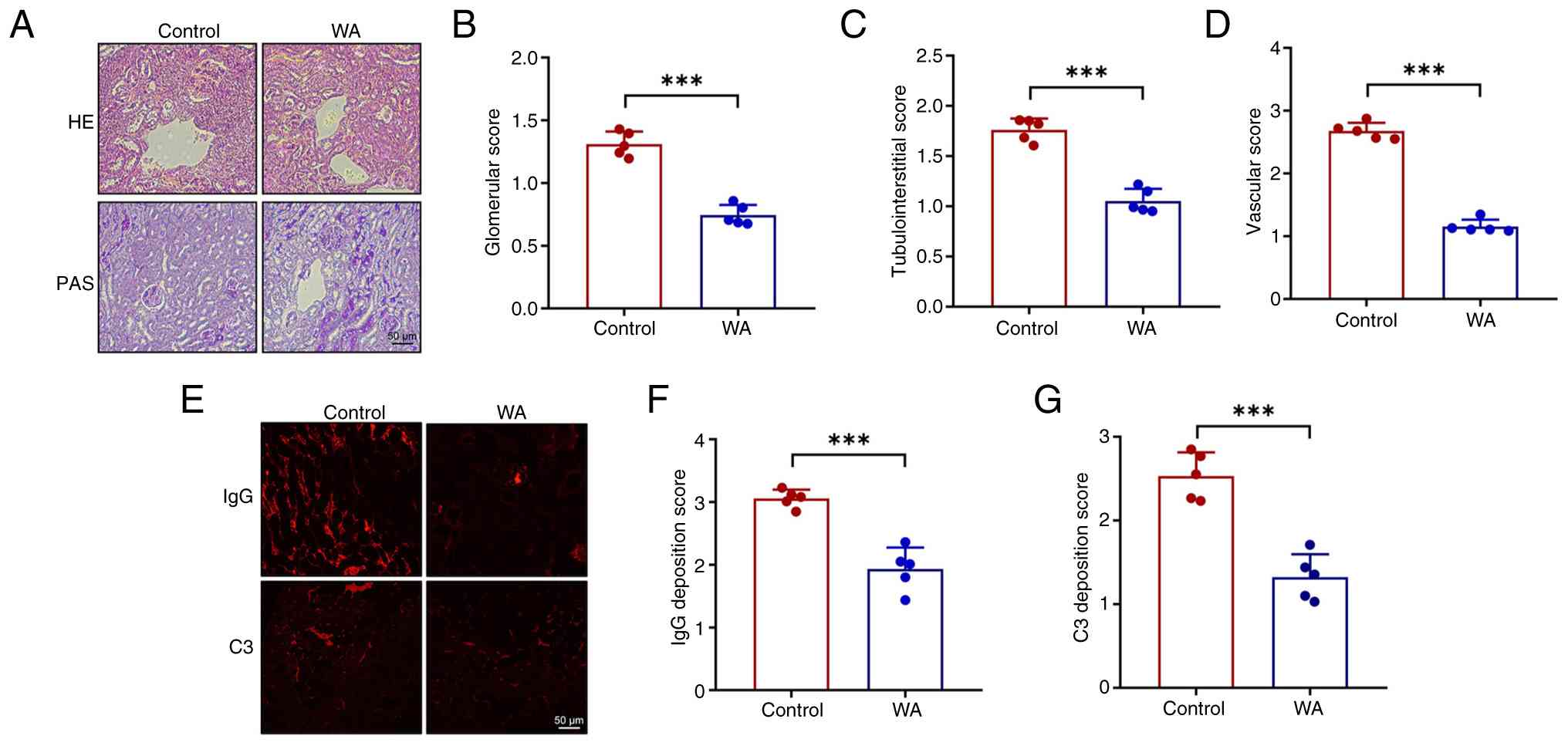
WA suppresses renal pathohistological
changes in experimental LN
Renal pathological changes were assessed using
H&E and PAS staining. Kidney sections from SLE model mice
exhibited glomerular abnormality, including diffuse endothelial
cell proliferation, segmental sclerosis and tubular atrophy. These
pathological features were markedly attenuated by WA, consistent
with the observed improvements in renal function (Fig. 4A-D). To evaluate glomerular
injury, immunofluorescence staining was performed, which revealed
that WA significantly decreased glomerular deposition of IgG and C3
in SLE mice (Fig. 4E-G).
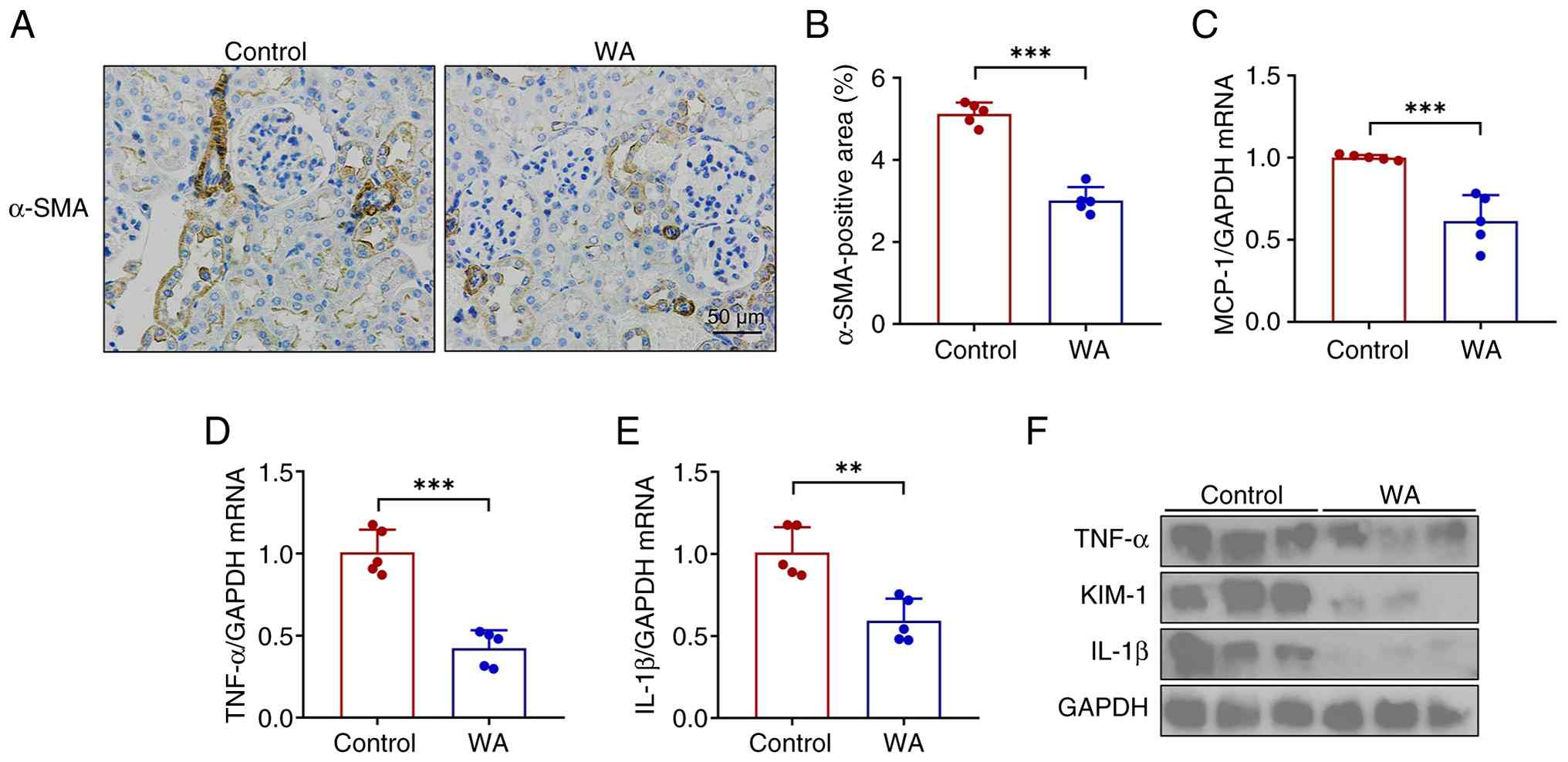
WA alleviates renal inflammation in
experimental LN
Immune cell-mediated sustained inflammation
contributes to the pathogenesis of renal injury in lupus (27). In the present study, WA
ameliorated lupus-induced renal pathology, as demonstrated by a
decrease in α-SMA positive areas (Fig. 5A and B). RT-qPCR and western
blotting demonstrated that WA abrogated the upregulation of key
proinflammatory mediators, including IL-1β, TNF-α and MCP-1, in the
kidney of SLE mice (Fig. 5C-F).
In addition, the tubular injury was also attenuated, demonstrated
by a decrease in the expression of tubular injury marker KIM-1
(Fig. 5F). Collectively, these
findings indicated that WA attenuated tubular injury by suppressing
the intrarenal inflammatory response.
Pon1 is a mediator of alleviated innate
immune responses induced by WA
The present study performed genome-wide
transcriptomic sequencing on renal tissue from SLE mice with or
without WA treatment to elucidate the molecular basis of its
immunosuppressive effects. Notably, WA markedly reshaped the renal
transcriptome (Fig. 6A),
enhancing the expression of lipid metabolism genes [perilipin
(PLIN)4, fatty acid-binding protein 4 (FABP4) and PLIN5] and
suppressing genes associated with immunity [lymphocyte antigen 6
complex locus D, immunoglobulin heavy variable 5-9, C-X-C motif
chemokine ligand (CXCL)10 and CXCL13], transcription (STAT4 and
transcription factor 7) and apoptosis (growth arrest and DNA
damage-inducible γ and serine/threonine kinase 17B; data not
shown). GO and KEGG analyses demonstrated that WA activated
lipid-centric and metabolic pathways while inhibiting
pro-inflammatory signaling (Fig. 6B
and C). Moreover, from the 161 DEGs identified by volcano plot
(18 up- and 143 downregulated), Pon1 emerged as a lead candidate as
the second most significantly upregulated transcript in the
WA-treated group (Fig. 6D and
E). Among the top upregulated genes identified by RNA-seq, Pon1
was selected for further functional validation due to its roles in
alleviating oxidative stress and systemic inflammation (28-31), both of which are hallmark
features of SLE-induced renal damage. Therefore, it was
hypothesized that induction of Pon1 is a critical mechanism through
which WA alleviates SLE pathology.
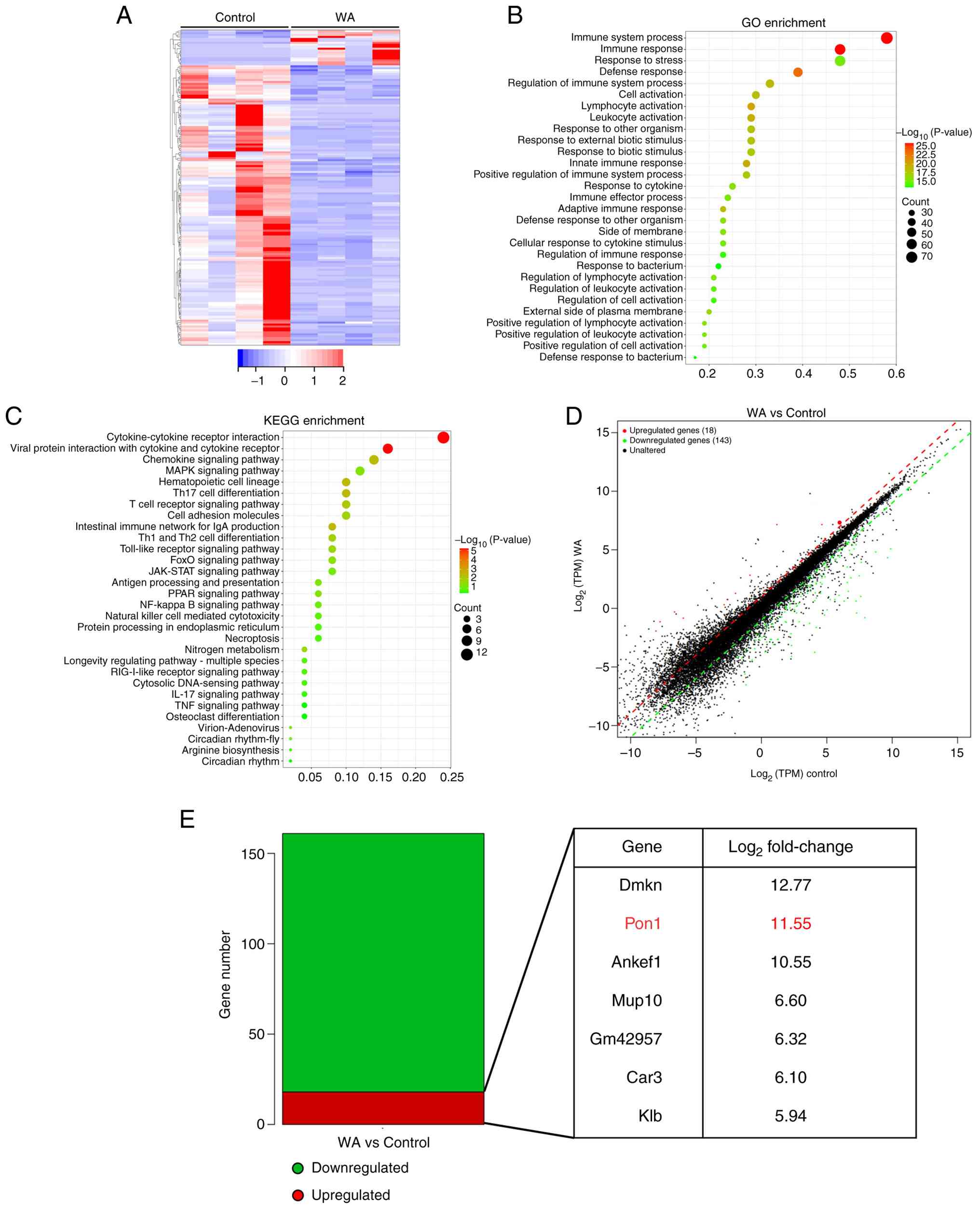 | Figure 6RNA sequencing analysis of kidney in
SLE mice after WA treatment. (A) Hierarchical clustered heatmap of
gene expression profiles for WA- or vehicle-treated SLE mice. (B)
GO enrichment analysis of DEGs. (C) KEGG pathway analysis of DEGs.
(D) Volcano plot of gene expression profiles. (E) Number of DEGs
(q<0.05). The top seven genes are presented. SLE, systemic lupus
erythematosus; WA, withaferin A; GO, gene ontology; DEG,
differentially expressed genes; KEGG, Kyoto encyclopedia of genes
and genomes; TPM, transcripts per kilobase; Dmkn, dermokine; Pon,
paraoxonase; Ankef, ankyrin repeat and EF-hand domain containing;
Mup, major urinary protein; Gm, predicted gene; Car, carbonic
anhydrase; Klb, klotho β. |
Inhibition of Pon1 reverses the effects
induced by WA in HK-2 cells under inflammatory conditions
To assess the role of Pon1, the present study
established an in vitro model of LN by stimulating HK-2
cells with LPS. WA significantly restored the impaired cell
viability induced by LPS (Fig.
7A) and suppressed the upregulation of proinflammatory
cytokines (IL-1β, TNF-α and MCP-1) at both the mRNA and protein
levels (Fig. 7B-E). Notably, WA
also upregulated Pon1 expression (Fig. 7F).
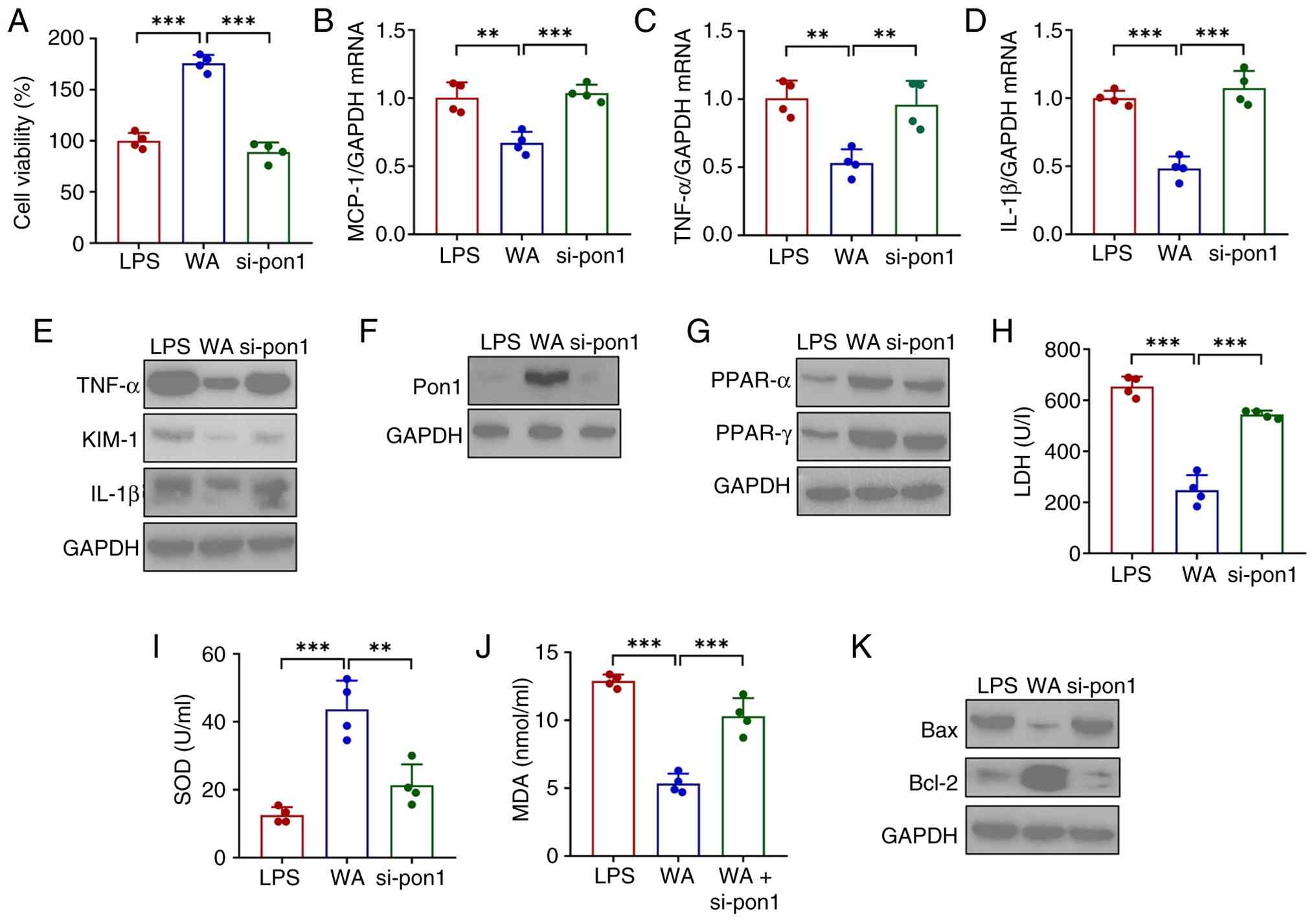 | Figure 7WA alleviates inflammation and
eliminates reactive oxygen species in HK-2 cells by regulating
Pon1/PPAR signaling under inflammatory conditions. (A) Viability of
HK-2 cells after WA treatment in the presence or absence of si-Pon1
transfection. All cells were pretreated with LPS. mRNA expression
of (B) MCP-1, (C) TNF-α and (D) IL-1β and (E) western blotting
analysis of the expression of MCP-1, TNF-α, IL-1β and KIM-1 in HK-2
cells. (F) Pon1 expression levels. (G) Protein expression of PPAR-α
and PPAR-γ. (H) LDH and (I) SOD activities. (J) MDA levels in HK-2
cells. (K) Expression of Bax and BCl-2. **P<0.01,
***P<0.001. WA, withaferin A; Pon, paraoxonase; si,
small interfering; LPS, lipopolysaccharide; MCP, monocyte
chemoattractant protein; LDH, lactate dehydrogenase; SOD,
superoxide dismutase; MDA, malondialdehyde. |
To determine whether Pon1 is essential for the
effects of WA, the present study performed loss-of-function
experiments using siRNA-mediated knockdown of Pon1. si-Pon1
effectively inhibited the expression of Pon1 (Fig. S1) and abrogated the protective
effects of WA, as demonstrated by a reversal of the improved cell
viability (Fig. 7A) and
increased proinflammatory cytokine expression (Fig. 7B-E). Given previous findings that
Pon1 mitigates cellular stress by activating PPARγ signaling
(32) and the transcriptomic
data in the present study indicating WA activates the PPAR pathway
in vivo, this pathway was investigated. WA enhanced the
expression of PPARα and PPARγ in LPS-treated HK-2 cells (Fig. 7G). This activation was associated
with attenuated oxidative stress, demonstrated by decreased levels
of LDH and MDA and an increase in SOD activity (Fig. 7H-J). WA modulated the expression
of apoptosis-related proteins, as indicated by the downregulated
expression of Bax and Bcl-2 (Fig.
7K). Notably, these beneficial regulatory effects of WA were
nullified by Pon1 knockdown (Fig.
7G-K).
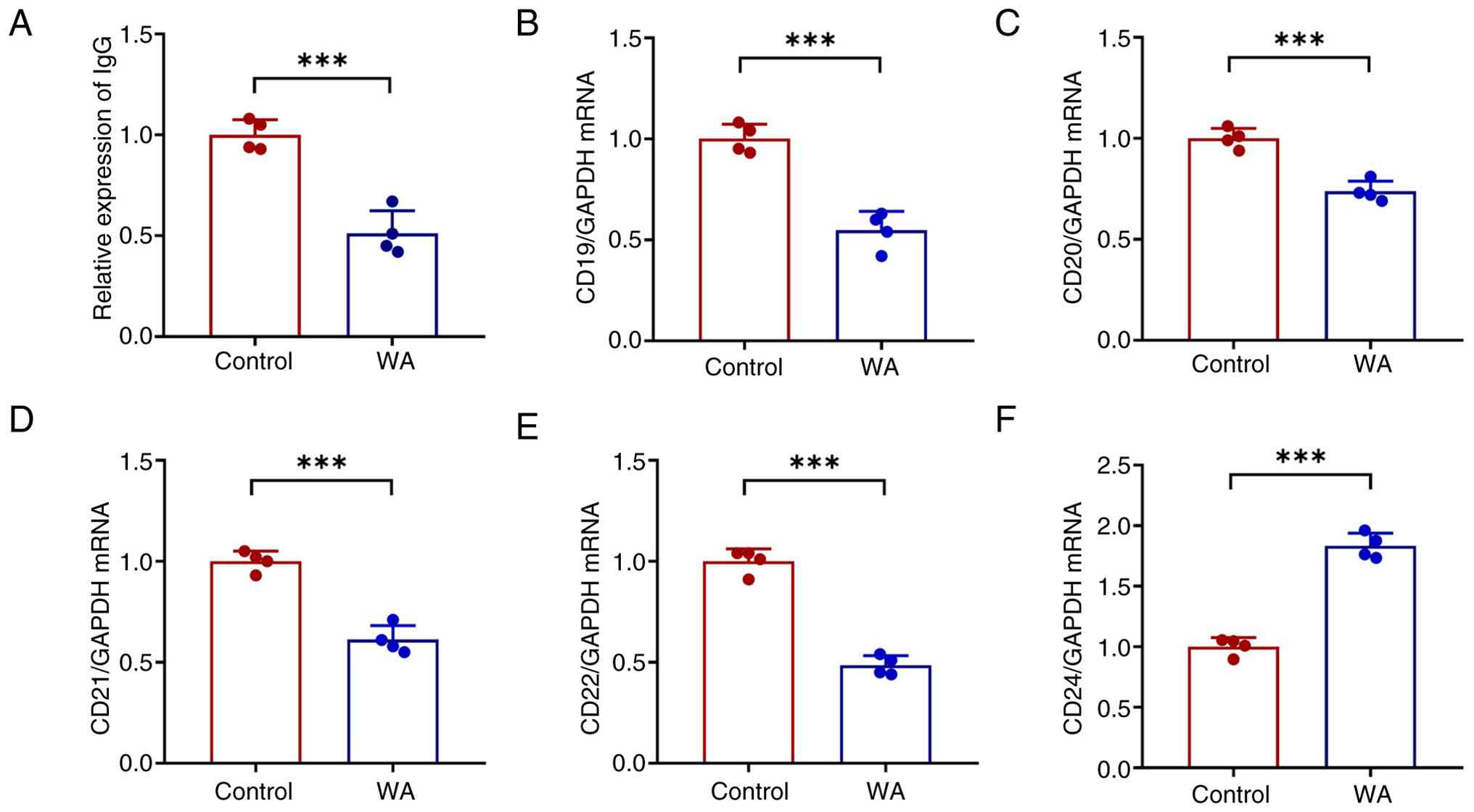
WA led to a decrease in supernatant IgG (Fig. 8A) and diminished expression of
the B cell surface markers CD19, CD20, CD21 and CD22 (Fig. 8B-E), suggesting suppressed B cell
differentiation and proliferation. Conversely, an increase in the
apoptosis-associated marker CD24 was observed (Fig. 8F), indicating promotion of B cell
apoptosis.
Pon1 activator alleviates innate immune
responses in HK-2 cells
As Pon1 demonstrated a role in WA-induced effects,
the present study assessed whether Pon1 activator showed similar
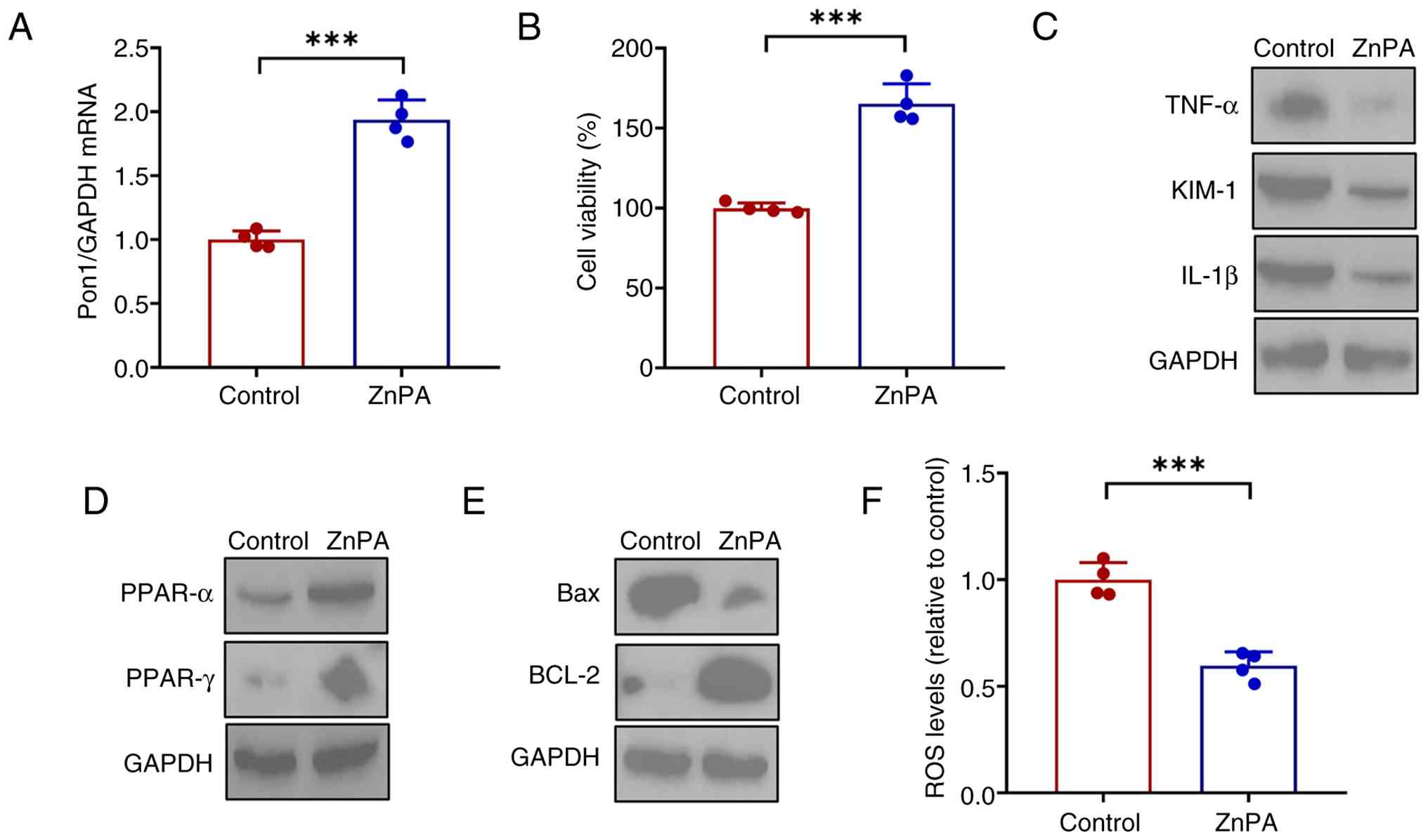
effects induced by WA. ZnPA significantly enhanced the expression
of Pon1 (Fig. 9A) in HK-2 cells.
ZnPA increased the viability of HK-2 cells treated with LPS
(Fig. 9B), which was accompanied
by decreased expression of the proinflammatory cytokines IL-1β,
TNF-α and MCP-1 (Fig. 9C).
Notably, PPAR signaling, oxidative stress and apoptosis induced by
LPS were markedly attenuated in HK-2 cells following ZnPA
administration (Fig. 9D-F). Pon1
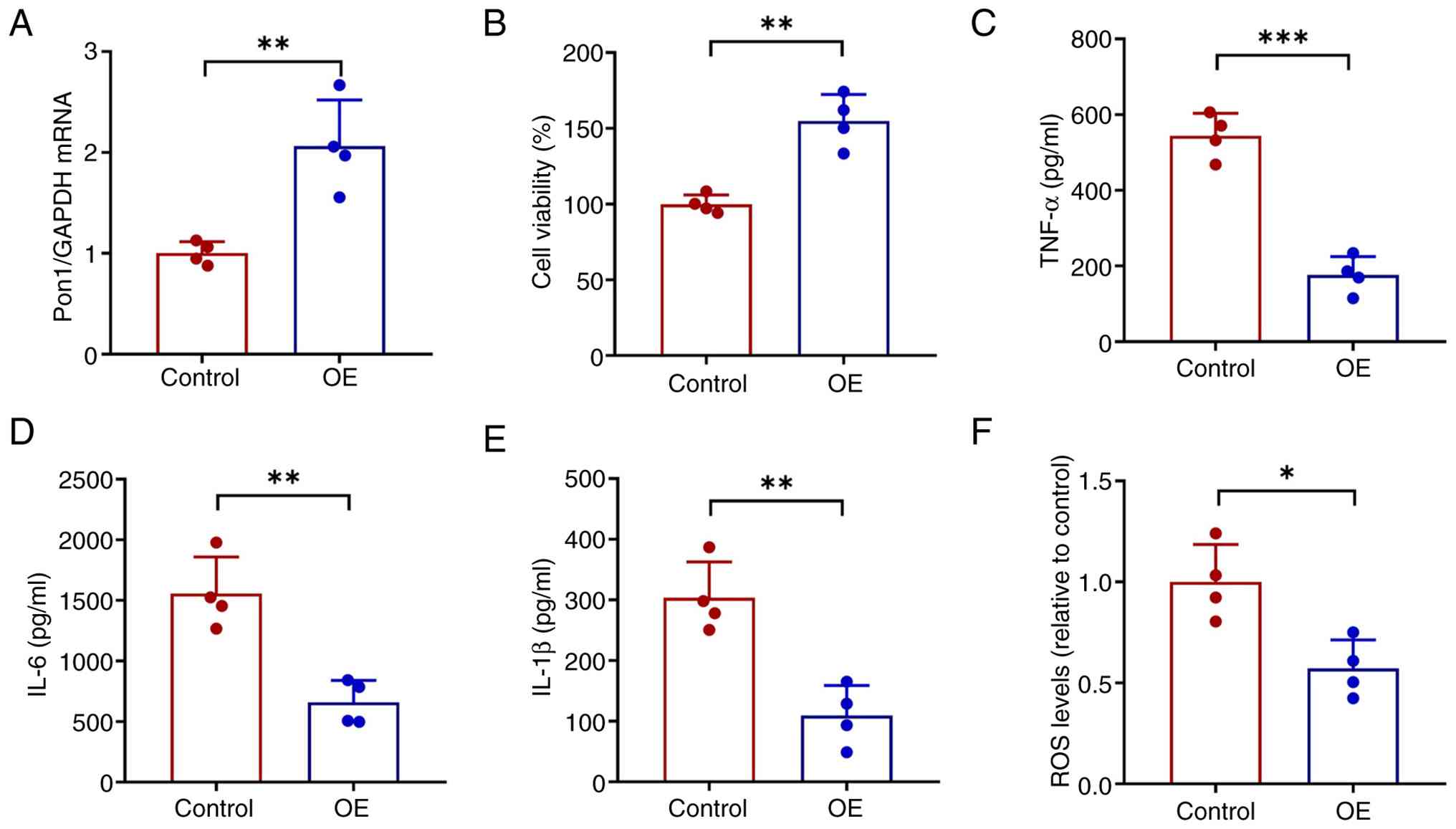
was overexpressed in HK-2 cells to evaluate whether higher levels
of Pon1 led to similar kidney protection (Fig. 10A). Pon1 overexpression resulted
in increased cell viability (Fig.
10B) and decreased inflammation (Fig. 10C-E) and ROS levels (Fig. 10F). Relative mRNA expression of
Pon1 was compared between healthy donors and patients with LN in
the public dataset GSE121239. Pon1 expression was significantly
lower in the patients with LN than in healthy donors (Fig. S2), supporting the hypothesis
that augmenting Pon1 expression represents a promising therapeutic
strategy for LN.
Discussion
SLE is a chronic autoimmune disorder marked by
diverse clinical manifestations that impact multiple organ systems.
Therapeutic strategies for SLE primarily aim to suppress the immune
response. However, the complexity of SLE pathophysiology highlights
the need to elucidate the underlying molecular mechanisms and key
signaling pathways involved in disease progression, as well as to
develop novel, targeted therapies with enhanced efficacy and safety
profiles.
The present study identified WA as a promising
therapeutic agent capable of alleviating SLE symptoms both in
vivo and in vitro. Mechanistically, WA upregulated Pon1
expression in the kidney and decreased splenic and renal
inflammation, oxidative stress and apoptosis through activation of
the PPAR signaling pathway. ZnPA, a Pon1 activator, countered
LPS-induced cytotoxicity, suppressed excessive immune activation
and diminished oxidative stress and the expression of
apoptosis-associated proteins in HK-2 cells, underscoring the
potential of Pon1-targeted interventions for the treatment of
SLE.
WA, a steroidal lactone derived from the plant W.
somnifera, is utilized as a natural compound in traditional
medicine due to its broad-spectrum efficacy in several disorders,
such as Parkinson's disease (33) and obesity (34). Additionally, WA is a potent
antioxidant (35),
anti-inflammatory (36),
anti-angiogenic (37),
anti-epithelial-mesenchymal transition (38) and immunomodulatory agent
(39). WA can mitigate oxidative
stress-induced apoptotic death in cardiomyocytes (40) and protect against free fatty
acid-induced inflammatory responses and oxidative stress by
inhibiting IL-6 production and suppressing NF-κB signaling in
endothelial cells (41). Kanak
et al (42) reported that
administering WA, an NF-κB inhibitor, inhibits endoplasmic
reticulum stress and inflammasome activity to moderate chronic
pancreatitis in a mouse model. Moreover, the present study
identified WA as a potential therapeutic agent for SLE through the
CMAP approach. WA significantly improved renal function in a murine
model of SLE. Furthermore, WA ameliorated splenic immune cell
dysregulation, attenuated renal pathohistological abnormality and
decreased renal inflammation. Integrated RNA-seq analysis and siRNA
transfection revealed that Pon1 serves a pivotal role in mediating
the protective effects of WA by mitigating oxidative stress and
apoptosis. The present findings also highlight the multi-organ
protective profile of WA, showing no obvious changes in body weight
and liver weight. Pires et al (43) reported clinical adverse effects
in patients with advanced-stage high-grade osteosarcoma treated
with WA, including fever, fatigue, gastrointestinal reaction and
elevated liver enzymes. These may be disease-related complications
or sequelae of the advanced clinical stage rather than direct
toxicities of WA. Advanced high-grade osteosarcoma is typically
associated with systemic inflammatory responses and
hypoalbuminemia, which predispose patients to fever and edema, both
of which are common clinical hallmarks of disease progression.
Furthermore, these symptoms were also observed in the control
group. The incidence of these events did not exhibit a clear
dose-response association, further suggesting they were independent
of WA administration (43).
Consequently, further research is warranted to delineate the
clinical effects attributable to WA treatment. The aforementioned
study found WA to be well-tolerated by patients at doses up to 216
mg/day (43). The dose in the
present study (2 mg/kg/day, equivalent to 9.8 mg/day for a 60 kg
human) was substantially lower. Therefore, it was unlikely to cause
observable toxicity and was safe for the regular use. Nevertheless,
the lack of comprehensive, longitudinal pharmacokinetic profiles
and human preclinical data remains a limitation of the present
study. It is necessary to determine the association between
systemic exposure of WA and clinical complications such as fatigue
or hepatotoxicity.
As a circulating calcium-dependent esterase and
lactonase, Pon1 is primarily synthesized in the liver, kidney and
intestine (44), and is
predominantly associated with the high-density lipoprotein (HDL)
fraction in circulation (45).
This contributes to the antioxidant and antiatherogenic properties
of HDL (44,46). HDL is a key mediator of reverse
cholesterol transport (47,48), facilitating the removal of excess
cholesterol from peripheral tissue. Accumulating evidence has
revealed the functional complexity of HDL in several pathological
conditions, including infection, neoplasms and autoimmune diseases
(49-52). The pleiotropic functions of HDL
include anti-inflammatory, antioxidant, antithrombotic and
antiapoptotic effects and the promotion of nitric oxide synthesis
(49). In SLE, numerous patients
exhibit elevated levels of very-low-density lipoprotein and
low-density lipoprotein, along with decreased HDL levels, a profile
commonly referred to as the lupus lipoprotein pattern (50-52). Low levels of HDL-cholesterol
represent one of the most prevalent dyslipidemia markers in SLE,
including among pediatric populations. Notably, additional
HDL-associated abnormalities have been reported, such as diminished
Pon1 activity (53), suggesting
that modulation of HDL function via Pon1 regulation may serve as an
effective therapeutic strategy for SLE. In the present study, Pon1
was identified as a potential molecular target of WA in the
treatment of SLE. Upregulated Pon1 expression in both kidney tissue
and HK-2 cells following WA administration was demonstrated and
siRNA-mediated knockdown and pharmacological activation assays
suggested that Pon1 mediates the beneficial effects of WA in SLE
via the PPAR signaling pathway. These findings support Pon1 as a
promising therapeutic target for SLE intervention. RNA-seq revealed
activation of the PPAR signaling pathway and upregulation of key
lipid metabolism-associated genes, including PLIN4, FABP4 and
PLIN5. PPAR signaling transcriptionally regulates energy metabolism
and inflammatory responses. Patients with LN typically exhibit a
distinct lupus lipoprotein pattern, characterized by systemic and
intrarenal lipid dysregulation (54). The present study indicated that
WA not only enhanced the expression of PPARα and PPARγ in HK-2
cells but also promoted the expression of proteins essential for
lipid droplet formation and fatty acid transport. These metabolic
modulations may contribute to the reinforcement of energy
homeostasis within renal proximal tubular cells by mitigating
ectopic lipid deposition and its associated lipotoxicity, thereby
alleviating renal injury. Furthermore, Pon1 is key in maintaining
lipid and redox stability. Pon1 activation can mitigate cell stress
by triggering PPARγ signaling (32). The present study further
demonstrated this regulatory axis, as the beneficial effects of WA
on PPAR protein levels were abolished upon Pon1 knockdown. The
activation of the Pon1/PPAR pathway may be key for the suppression
of ROS. Mechanistically, the PPAR pathway inhibits pro-inflammatory
cascades, such as NF-κB, while bolstering antioxidant defenses,
effectively breaking the cycle of intrarenal inflammation and
oxidative damage (55).
B cell dysfunction serves a key role in the
pathogenesis of SLE (56). Under
physiological conditions, B cell activation is regulated by several
factors, including ROS (57).
ROS contribute to B cell maturation (58-60). For example, hydrogen peroxide
enhances the DNA-binding activity of paired box 5, and exposure of
B cells to H2O2 induces rapid nuclear
translocation of the cytoplasmic redox factor apurinic/apyrimidinic
endonuclease 1/redox effector factor-1, thereby promoting early B
cell development and maturation (58,59). In addition, ROS regulate B cell
activation through phosphorylation of spleen tyrosine kinase and
other membrane-proximal B cell receptor signaling effectors
(60). ROS levels are elevated
in activated B cells, and oxidative processes are key for B cell
differentiation. ROS production increases progressively during B
cell activation and differentiation (57). The RNA-seq analysis in the
present study demonstrated downregulation of several Ig and IL
genes, including Ig heavy variable 8-12, immunoglobulin heavy
constant ε and IL-21 (data not shown), suggesting suppressed B cell
activation. As WA decreased ROS levels, it was hypothesized that
this reduction contributes to inhibition of B cell activation in
SLE mice. WA acts on HK-2 cells to decrease ROS generation via the
Pon1/PPAR signaling pathway, thereby inhibiting B cell activation
and ameliorating SLE symptoms. Furthermore, RNA-seq revealed
upregulation of several lipid metabolism-associated factors in
WA-treated mice such as plin5, pnpla3 and plin4. These changes may
also be associated with PPAR signaling as this pathway suppresses
ROS production (61), although
the underlying mechanisms require further investigation.
The present study has certain limitations. First,
although Pon1 was identified as a potential therapeutic target for
SLE, additional evidence is needed to establish its functional role
in SLE models. The present study did not assess whether Pon1
overexpression protects against LN-induced kidney injury. Second,
the present study did not determine whether WA directly targets B
cells to modulate their activation and correct B cell dysfunction
in SLE. The present findings indicated that WA enhances Pon1
expression and suppresses ROS production through PPAR signaling,
leading to inhibition of B cell activation and alleviation of
SLE-associated LN in mice.
Supplementary Data
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the Sequence Read Archive dataset
under accession no. PRJNA1393487 or at the following URL:
ncbi.nlm.nih.gov/bioproject/PRJNA1393487.
Authors' contributions
MX and XM conceived and designed the study. MX and
JF performed the language editing, analyzed data and constructed
figures. MX performed bioinformatics analysis. All authors have
read and approved the final manuscript. MX, JF and XM confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the National Institutes of Health Guide for the Care and Use
of Laboratory Animals and were approved by the Animal Experimental
Ethical Inspection of Gansu University of Chinese Medicine
(approval no. SY2024-257; Lanzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural Science Fund
Project of Gansu (grant no. 25JRRA1011) and Science and Technology
Plan Project of Gansu Province (grant no. 24YFFA037).
References
|
1
|
Morales E, Galindo M, Trujillo H and Praga
M: Update on lupus nephritis: Looking for a new vision. Nephron.
145:1–13. 2021. View Article : Google Scholar
|
|
2
|
Mohan C, Zhang T and Putterman C:
Pathogenic cellular and molecular mediators in lupus nephritis. Nat
Rev Nephrol. 19:491–508. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Appel GB, Contreras G, Dooley MA, Ginzler
EM, Isenberg D, Jayne D, Li LS, Mysler E, Sánchez-Guerrero J,
Solomons N, et al: Mycophenolate mofetil versus cyclophosphamide
for induction treatment of lupus nephritis. J Am Soc Nephrol.
20:1103–1112. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ponticelli C and Podestà MA: Calcineurin
inhibitors in lupus nephritis. J Nephrol. 34:399–402. 2021.
View Article : Google Scholar
|
|
5
|
Yu C, Li P, Dang X, Zhang X, Mao Y and
Chen X: Lupus nephritis: New progress in diagnosis and treatment. J
Autoimmun. 132:1028712022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Anders HJ, Saxena R, Zhao MH, Parodis I,
Salmon JE and Mohan C: Lupus nephritis. Nat Rev Dis Primers.
6:72020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Flores-Borja F, Kabouridis PS, Jury EC,
Isenberg DA and Mageed RA: Decreased Lyn expression and
translocation to lipid raft signaling domains in B lymphocytes from
patients with systemic lupus erythematosus. Arthritis Rheum.
52:3955–3965. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liossis SN, Solomou EE, Dimopoulos MA,
Panayiotidis P, Mavrikakis MM and Sfikakis P: B-cell kinase lyn
deficiency in patients with systemic lupus erythematosus. J
Investig Med. 49:157–165. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zouali M and Sarmay G: B lymphocyte
signaling pathways in systemic autoimmunity: Implications for
pathogenesis and treatment. Arthritis Rheum. 50:2730–2741. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Geh D and Gordon C: Epratuzumab for the
treatment of systemic lupus erythematosus. Expert Rev Clin Immunol.
14:245–258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wallace DJ and Goldenberg DM: Epratuzumab
for systemic lupus erythematosus. Lupus. 22:400–405. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Arazi A, Rao DA, Berthier CC, Davidson A,
Liu Y, Hoover PJ, Chicoine A, Eisenhaure TM, Jonsson AH, Li S, et
al: The immune cell landscape in kidneys of patients with lupus
nephritis. Nat Immunol. 20:902–914. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin K, Li L, Dai Y, Wang H, Teng S, Bao X,
Lu ZJ and Wang D: A comprehensive evaluation of connectivity
methods for L1000 data. Brief Bioinform. 21:2194–2205. 2020.
View Article : Google Scholar
|
|
14
|
Subramanian A, Narayan R, Corsello SM,
Peck DD, Natoli TE, Lu X, Gould J, Davis JF, Tubelli AA, Asiedu JK,
et al: A next generation connectivity map: L1000 platform and the
first 1,000,000 profiles. Cell. 171:1437–1452.e17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang Y, Jiang Q, Zou H, Zhao J, Zhang J
and Ren L: Withaferin A: A potential selective glucocorticoid
receptor modulator with anti-inflammatory effect. Food Chem
Toxicol. 179:1139492023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kumar S, Mathew SO, Aharwal RP, Tulli HS,
Mohan CD, Sethi G, Ahn KS, Webber K, Sandhu SS and Bishayee A:
Withaferin A: A pleiotropic anticancer agent from the indian
medicinal plant Withania somnifera (L.) Dunal. Pharmaceuticals
(Basel). 16:1602023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nisar MF, Wan C, Büsselberg D, Calina D
and Sharifi-Rad J: Current mechanistic insights into Withaferin A:
A promising potential adjuvant anticancer agent from Withania
somnifera. Naunyn Schmiedebergs Arch Pharmacol. 398:3573–3593.
2025. View Article : Google Scholar
|
|
18
|
Underwood W and Anthony R: AVMA Guidelines
for the Euthanasia of Animals: 2020 Edition. American Veterinary
Medical Association; 2020
|
|
19
|
Yang C, Xue J, An N, Huang XJ, Wu ZH, Ye
L, Li ZH, Wang SJ, Pan QJ, Liang D and Liu HF: Accelerated
glomerular cell senescence in experimental lupus nephritis. Med Sci
Monit. 24:6882–6891. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen XC, Wu D, Wu HL, Li HY, Yang C, Su
HY, Liu ZJ, Huang XR, Lu X, Huang LF, et al: Metformin improves
renal injury of MRL/lpr lupus-prone mice via the AMPK/STAT3
pathway. Lupus Sci Med. 9:e0006112022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mao C, Jin W, Dou L, Guo T, Huang J, Wang
Y, Liu X, Wu S, Qiao W, Xiang Y, et al: Bioengineered ROS-tolerant
probiotic reshapes gut microbiota-host axis to ameliorate type 2
diabetes in male mice. Nat Commun. 17:33392026. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ackermann M and Strimmer K: A general
modular framework for gene set enrichment analysis. BMC
Bioinformatics. 10:472009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Parodis I, Gatto M and Sjöwall C: B cells
in systemic lupus erythematosus: Targets of new therapies and
surveillance tools. Front Med (Lausanne). 9:9523042022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ahmed A, Li S, Yu JJ and Shao WH:
Immunopathogenesis of systemic lupus erythematosus: Interplay of
innate and adaptive immunity, microbiome dysbiosis, and emerging
therapeutic targets. Pathophysiology. 32:612025. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dong C, Wu M, Liu H, Yang K, Ma S, Guo Y,
Lan Y and Lu X: Breaking the cycle: Immune complexes, complement
activation, and novel immunotherapies in lupus nephritis. Front
Immunol. 16:16248502025. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dornas W and Silva M: Modulation of the
antioxidant enzyme paraoxonase-1 for protection against
cardiovascular diseases. Nutr Metab Cardiovasc Dis. 34:2611–2622.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jakubowski H: The molecular bases of
Anti-oxidative and Anti-inflammatory properties of paraoxonase 1.
Antioxidants (Basel). 13:12922024. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rozenberg O, Shih DM and Aviram M:
Paraoxonase 1 (PON1) attenuates macrophage oxidative status:
Studies in PON1 transfected cells and in PON1 transgenic mice.
Atherosclerosis. 181:9–18. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sikora M and Jakubowski H: Changes in
redox plasma proteome of Pon1-/-mice are exacerbated by a
hyperhomocysteinemic diet. Free Radic Biol Med. 169:169–180. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang M, Yu X, Liu C and Liu Y:
Overexpression of PON1 reduces high glucose induced renal tubular
epithelial cell injury by activating PPARgamma signaling pathway to
alleviate diabetes nephropathy. Arch Endocrinol Metab.
69:e2403772025. View Article : Google Scholar
|
|
33
|
Zhao M, Wang B, Zhang C, Su Z, Guo B, Zhao
Y and Zheng R: The DJ1-Nrf2-STING axis mediates the neuroprotective
effects of Withaferin A in Parkinson's disease. Cell Death Differ.
28:2517–2535. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo B, Liu J, Wang B, Zhang C, Su Z, Zhao
M, Qin L, Zhang W and Zheng R: Withaferin A promotes white adipose
browning and prevents obesity through sympathetic Nerve-activated
Prdm16-FATP1 axis. Diabetes. 71:249–263. 2022. View Article : Google Scholar
|
|
35
|
Oh JH, Lee TJ, Park JW and Kwon TK:
Withaferin A inhibits iNOS expression and nitric oxide production
by Akt inactivation and down-regulating LPS-induced activity of
NF-kappaB in RAW 264.7 cells. Eur J Pharmacol. 599:11–17. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Martorana F, Guidotti G, Brambilla L and
Rossi D: Withaferin A inhibits nuclear Factor-κB-Dependent
Pro-Inflammatory and stress response pathways in the astrocytes.
Neural Plast. 2015:3819642015. View Article : Google Scholar
|
|
37
|
Mohan R, Hammers HJ, Bargagna-Mohan P,
Zhan XH, Herbstritt CJ, Ruiz A, Zhang L, Hanson AD, Conner BP,
Rougas J and Pribluda VS: Withaferin A is a potent inhibitor of
angiogenesis. Angiogenesis. 7:115–122. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee J, Hahm ER, Marcus AI and Singh SV:
Withaferin A inhibits experimental epithelial-mesenchymal
transition in MCF-10A cells and suppresses vimentin protein level
in vivo in breast tumors. Mol Carcinog. 54:417–429. 2015.
View Article : Google Scholar
|
|
39
|
Rasool M and Varalakshmi P:
Immunomodulatory role of Withania somnifera root powder on
experimental induced inflammation: An in vivo and in vitro study.
Vascul Pharmacol. 44:406–410. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yan Z, Guo R, Gan L, Lau WB, Cao X, Zhao
J, Ma X, Christopher TA, Lopez BL and Wang Y: Withaferin A inhibits
apoptosis via activated Akt-mediated inhibition of oxidative
stress. Life Sci. 211:91–101. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Batumalaie K, Amin MA, Murugan DD, Sattar
MZ and Abdullah NA: Withaferin A protects against palmitic
acid-induced endothelial insulin resistance and dysfunction through
suppression of oxidative stress and inflammation. Sci Rep.
6:272362016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kanak MA, Shahbazov R, Yoshimatsu G, Levy
MF, Lawrence MC and Naziruddin B: A small molecule inhibitor of
NFκB blocks ER stress and the NLRP3 inflammasome and prevents
progression of pancreatitis. J Gastroenterol. 52:352–365. 2017.
View Article : Google Scholar
|
|
43
|
Pires N, Gota V, Gulia A, Hingorani L,
Agarwal M and Puri A: Safety and pharmacokinetics of Withaferin-A
in advanced stage high grade osteosarcoma: A phase I trial. J
Ayurveda Integr Med. 11:68–72. 2020. View Article : Google Scholar :
|
|
44
|
Furlong CE, Marsillach J, Jarvik GP and
Costa LG: Paraoxonases-1, -2 and -3: What are their functions? Chem
Biol Interact. 259:51–62. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gugliucci A and Menini T: Paraoxonase 1
and HDL maturation. Clin Chim Acta. 439:5–13. 2015. View Article : Google Scholar
|
|
46
|
Précourt LP, Amre D, Denis MC, Lavoie JC,
Delvin E, Seidman E and Levy E: The three-gene paraoxonase family:
Physiologic roles, actions and regulation. Atherosclerosis.
214:20–36. 2011. View Article : Google Scholar
|
|
47
|
Diaz L and Bielczyk-Maczynska E:
High-density lipoprotein cholesterol: How studying the 'good
cholesterol' could improve cardiovascular health. Open Biol.
15:2403722025. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yazdanyar A, Yeang C and Jiang XC: Role of
phospholipid transfer protein in high-density lipoprotein-mediated
reverse cholesterol transport. Curr Atheroscler Rep. 13:242–248.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Besler C, Lüscher TF and Landmesser U:
Molecular mechanisms of vascular effects of High-density
lipoprotein: Alterations in cardiovascular disease. EMBO Mol Med.
4:251–268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gaál K, Tarr T, Lőrincz H, Borbás V, Seres
I, Harangi M, Fülöp P and Paragh G: High-density lipopoprotein
antioxidant capacity, subpopulation distribution and paraoxonase-1
activity in patients with systemic lupus erythematosus. Lipids
Health Dis. 15:602016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lilleby V, Haugen M, Mørkrid L, Frey
Frøslie K, Holven KB and Førre O: Body composition, lipid and
lipoprotein levels in childhood-onset systemic lupus erythematosus.
Scand J Rheumatol. 36:40–47. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yuan J, Li LI, Wang Z, Song W and Zhang Z:
Dyslipidemia in patients with systemic lupus erythematosus:
Association with disease activity and B-type natriuretic peptide
levels. Biomed Rep. 4:68–72. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kiss E, Seres I, Tarr T, Kocsis Z, Szegedi
G and Paragh G: Reduced paraoxonase1 activity is a risk for
atherosclerosis in patients with systemic lupus erythematosus. Ann
N Y Acad Sci. 1108:83–91. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Guleria A, Phatak S, Dubey D, Kumar S,
Zanwar A, Chaurasia S, Kumar U, Gupta R, Aggarwal A, Kumar D and
Misra R: NMR-Based serum metabolomics reveals reprogramming of
lipid dysregulation following Cyclophosphamide-Based induction
therapy in lupus nephritis. J Proteome Res. 17:2440–2448. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu H, Wu L, Luo L, Wu YT, Zhang QX, Li HY
and Zhang BF: Corrigendum to 'Quercetin inhibits mitophagy-mediated
apoptosis and inflammatory response by targeting the
PPARγ/PGC-1α/NF-κB axis to improve acute liver failure' [Int
Immunopharmacol 143P2 (2024) 113444]. Int Immunopharmacol.
143:1136792024. View Article : Google Scholar
|
|
56
|
Santana-Sánchez P, Vaquero-García R,
Legorreta-Haquet MV, Chávez-Sánchez L and Chávez-Rueda AK: Hormones
and B-cell development in health and autoimmunity. Front Immunol.
15:13855012024. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang H, Wang L and Chu Y: Reactive oxygen
species: The signal regulator of B cell. Free Radic Biol Med.
142:16–22. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Merluzzi S, Moretti M, Altamura S, Zwollo
P, Sigvardsson M, Vitale G and Pucillo C: CD40 stimulation induces
Pax5/BSAP and EBF activation through a APE/Ref-1-dependent redox
mechanism. J Biol Chem. 279:1777–1786. 2004. View Article : Google Scholar
|
|
59
|
Tell G, Zecca A, Pellizzari L, Spessotto
P, Colombatti A, Kelley MR, Damante G and Pucillo C: An
'environment to nucleus' signaling system operates in B
lymphocytes: Redox status modulates BSAP/Pax-5 activation through
Ref-1 nuclear translocation. Nucleic Acids Res. 28:1099–1105. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Polikowsky HG, Wogsland CE, Diggins KE,
Huse K and Irish JM: Cutting Edge: Redox signaling hypersensitivity
distinguishes human germinal center B cells. J Immunol.
195:1364–1367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wang M, Zhou F, Luo Y, Deng X, Chen X and
Yi Q: The transcription factor PPARA mediates SIRT1 regulation of
NCOR1 to protect damaged heart cells. Cardiovasc Diagn Ther.
14:832–847. 2024. View Article : Google Scholar : PubMed/NCBI
|