Introduction
Erectile dysfunction (ED) is a prevalent male
disorder (1), with an estimated
332 million cases worldwide by 2026 (2), severely impairing the physical and
mental health of both patients and their partners. Although
phosphodiesterase type 5 inhibitors (PDE5is) represent a milestone
in ED treatment (3), their use
is limited by contraindications and suboptimal efficacy in
refractory ED (4). Therefore,
identifying modifiable risk factors and exploring novel therapeutic
strategies is urgently needed. In recent years, modern lifestyles,
including shift work, sleep deprivation, frequent jet lag and
excessive artificial light exposure, have become increasingly
common, and these factors can disrupt the body's internal timing
system, a condition termed circadian rhythm disruption (CRD)
(5,6). Emerging epidemiological evidence
indicates that CRD has become a growing concern in the field of
male reproductive health and is closely associated with the
development and progression of ED (7,8).
Although previous studies by the authors have confirmed that CRD
significantly impairs erectile function in rats (9,10), the underlying mechanisms remain
largely unclear.
Notably, CRD may promote the development of ED
through two distinct pathways. First, CRD can indirectly increase
the risk of metabolic and cardiovascular diseases, which are
well-established independent risk factors for ED (7,8).
Second, CRD may directly impair erectile function through
mechanisms independent of the aforementioned metabolic
complications, such as inducing endothelial CRD, oxidative stress,
and penile corpus cavernosum tissue damage (11,12). Although the indirect pathway has
been extensively studied, the direct pathogenic role of CRD in ED
remains poorly understood. Therefore, the present study aims to
focus on the direct pathogenic effects of CRD, using a rigorously
controlled animal model (excluding significant changes in body
weight and serum testosterone levels, as shown in Figs. S1 and S2) to minimize
confounding influences from systemic metabolic factors.
As a core regulator of systemic redox homeostasis,
the circadian rhythm controls the expression of antioxidant enzymes
[for example, superoxide dismutase (SOD), glutathione peroxidase
(GPx) and catalase (CAT)] through core clock genes (for example,
Clock, Bmal1, Per and Cry), generating rhythmic fluctuations in
antioxidant capacity to counterbalance the peaks of metabolically
produced reactive oxygen species (ROS) (9,13). CRD disrupts this synchrony,
leading to impaired ROS clearance and accumulation of oxidative
products, thereby inducing a state of 'rhythmic oxidative stress
dysregulation'. In the penile corpus cavernosum, this imbalance
directly damages endothelial cells, suppresses the endothelial
nitric oxide synthase (eNOS)-nitric oxide (NO)-cyclic guanosine
monophosphate (cGMP) pathway, and promotes fibrosis. More
critically, persistently elevated ROS are potent activators of the
NLR family pyrin domain containing 3 (NLRP3) inflammasome (14); its activation triggers
Caspase-1-mediated pyroptosis, resulting in cell membrane rupture
and the release of large amounts of pro-inflammatory cytokines (for
example, IL-1β and IL-18), which further amplify local inflammation
and tissue damage (15-18). Thus, oxidative stress not only
directly impairs endothelial function but also initiates a
'oxidative stress-pyroptosis' vicious cycle, exacerbating
structural damage to the corpus cavernosum. Ultimately, CRD
constitutes a core pathogenic axis from circadian disruption to ED
by disrupting the local rhythmic balance of oxidative stress and
activating the pyroptosis cascade.
Melatonin (MT), a key synchronizer of the endogenous
circadian rhythm, regulates circadian physiological processes such
as sleep-wake cycles through MT1/MT2 receptors (19), providing a theoretical basis for
intervening in CRD-related pathologies. Beyond its circadian
regulatory function, MT and its metabolites possess potent free
radical-scavenging abilities and can activate the Nrf2/HO-1 pathway
to enhance endogenous antioxidant defense (20), while also inhibiting NLRP3
inflammasome-mediated pyroptosis (21), which aligns closely with the
'oxidative stress-pyroptosis' axis investigated in the present
study. Previous studies have demonstrated that MT exerts protective
effects in various ED models, including diabetes-induced ED,
cavernous nerve injury, and age-related ED, as well as in
testicular ischemia-reperfusion injury and varicocele (22,23). These lines of evidence support
the therapeutic potential of MT against CRD-induced ED, providing a
solid rationale for the present study.
Previous studies by the authors have shown that CRD
induced by an altered light-dark cycle (2 h:2 h) significantly
impairs erectile function in rats, as evidenced by reduced
intracavernous pressure and decreased activity of the eNOS-NO-cGMP
pathway (9,10). However, these earlier
investigations were primarily observational and did not elucidate
the underlying molecular mechanisms. Specifically, it remains
unclear how CRD disrupts endothelial homeostasis, which downstream
signaling pathways mediate the damage, and whether MT can intervene
in this process. The present study was designed to fill these
mechanistic gaps by systematically investigating the role of
oxidative stress and pyroptosis in CRD-induced ED and by
delineating the molecular basis of MT's therapeutic effects.
Materials and methods
Animal experimental design
A total of 24 male Sprague-Dawley (SD) rats
(12-weeks old; weight, ~200 g) were obtained from the Experimental
Animal Center of Guizhou Medical University. The animal protocols
were approved by the Animal Ethics Committee of Guizhou Medical
University (approval no. 2402989; Guiyang, China). All rats were
acclimatized for 1 week under controlled temperature conditions at
22-25°C, 55-70% humidity, and received unrestricted access to food
and water. They were divided into one Control group and three CRD
groups randomly, 6 rats in each group. Two groups of CRD rats were
treated with MT (cat. no. M5250; MilliporeSigma) administered
intraperitoneally with the dosages of 5 and 10 mg/kg/d, as the low
dosage group (CRD + MT-LD) and high dosage group (CRD + MT-HD),
while the other groups were treated with vehicle (equivalent volume
of normal saline). To explore the association between CRD and
erectile function, a CRD model was constructed by altering daily
light-dark cycles in rats (24-26). Rats were housed under a 12/12-h
light/dark cycle (LD 12:12, lights on at Zeitgeber time (ZT) 0 and
lights off at ZT12). The lights for CRD group were switched to LD
2:2 for 4 weeks, the light source was a white, fluorescent lamp
(500 lux at cage level), and then weekly body weight monitoring was
conducted.
MT administration protocol and solution preparation:
MT (MilliporeSigma) was administered intraperitoneally (i.p.) once
daily for 4 consecutive weeks, starting from the first day of the
CRD modeling protocol. To maintain consistency with the circadian
rhythm experiment, MT was injected at a fixed Zeitgeber time (ZT1),
i.e., 1 h after lights on (9:00 AM under a 12:12 light-dark cycle).
The temporal relationship between MT injection and CRD modeling was
as follows: each day, the light cycle was first changed to the CRD
schedule (LD 2:2), and MT was administered intraperitoneally within
30 min thereafter. MT is a lipophilic compound with poor solubility
in aqueous vehicles. Therefore, a stock solution was first prepared
by dissolving 100 mg MT in 4.305 ml dimethyl sulfoxide (DMSO) to
obtain a 100 mM stock solution. The stock solution was aliquoted
into amber tubes and stored at −20°C protected from light. On each
day of administration, the stock solution was freshly diluted with
sterile normal saline (0.9% NaCl) to achieve the desired working
concentrations (5 mg/kg for low dose, 10 mg/kg for high dose). The
final concentration of DMSO in the working solution was ≤0.1%
(v/v), a concentration previously shown to have no adverse effects
on animal physiology or erectile function. The solution was
vortexed and, if necessary, briefly sonicated to ensure complete
dissolution before injection. Control animals received an
equivalent volume of vehicle (sterile normal saline containing 0.1%
DMSO).
Cell experimental design
The immortalized human umbilical vein endothelial
cell line HUVEC-SV40 (SUNNCELL; https://www.app17.com/c163183/products/b3383_p1.html)
was used in the present study. HUVECs were maintained in RPMI-1640
medium supplemented with 10% fetal bovine serum (both from Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin at
37°C under 5% CO2. To assess dose and time-dependent
effects of MT and LPS, HUVECs were seeded in 96-well plates at
1×104 cells/well and cultured for 24 h. Cells were then
treated with specified concentrations of MT or LPS.
Post-incubation, cell viability was measured using a Cell Counting
Kit-8 (CCK-8) kit (cat. no. AR1160; Wuhan Boster Biological
Technology, Ltd.) according to the manufacturer's protocol.
Briefly, 10 μl of CCK-8 solution was added to each well
containing 100 μl of culture medium. The plates were then
incubated at 37°C for 2 h in a humidified incubator with 5%
CO2. Absorbance was quantified at 450 nm using a
Multiskan FC microplate reader (Thermo Fisher Scientific, Inc.).
HUVECs were categorized into seven distinct experimental groups as
follows: Control, LPS (1 mg/l), LPS + MT (800 μM), LPS + MT
+ ML385 (20 μM), LPS + MCC950 (20 μM), LPS + MT +
BMS986299 (20 μM), and LPS + NAC (10 mM).
Evaluation of erectile function
Erectile function was measured from 10:00 to 12:00
a.m, and the penile tissue was subsequently dissected. Moreover,
erectile function was assessed by recording the maximum
intracavernous pressure (mICP) and mICP/mean arterial pressure
(MAP) ratio as previously described (27). Rats were anesthetized by
intraperitoneal injection of 3% pentobarbital sodium (30 mg/kg),
the carotid was carefully exposed and cannulated with a heparinized
detaining venipuncture (26 G) needle to monitor the arterial
pressure through a pressure transducer. The cavernous nerve (CN)
was carefully separated with a low midline abdominal incision, then
a heparinized scalp acupuncture needle was inserted into the penile
crus to record ICP through another pressure transducer. When the CN
was electrically stimulated (using a voltage of 5 V at frequency of
20 Hz, pulse width of 5 msec, and sustained for 60 sec) (27), the ICP and arterial pressure were
simultaneously recorded using a BL420 bio-function experiment
system (Chengdu TME Technology Co., Ltd.). The mICP and MAP were
analyzed, and the final mICP/MAP ratio and total ICP (area under
curve, AUC-ICP) were calculated. At the end of the experiment, all
rats were euthanized by intraperitoneal injection of excessive
pentobarbital sodium (150 mg/kg), and death was confirmed by
cardiac arrest, respiratory arrest and corneal reflex loss.
RNA-sequencing (RNA-seq) and
bioinformatics analysis
Penile corpus cavernosum tissues were obtained from
rats in the Control, CRD, and CRD + MT-HD groups. Total RNA was
isolated with TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA was extracted, and sequencing libraries were
prepared using TruSeq Stranded mRNA Library Prep Kit. The
concentration of the final libraries was measured using a Qubit 4.0
Fluorometer with the Qubit dsDNA HS Assay Kit, and the fragment
size distribution was assessed using an Agilent 2100 Bioanalyzer
with the High Sensitivity DNA Kit. Each library was loaded at a
final concentration of 10 nM onto the Illumina NovaSeq 6000
platform (Illumina, Inc.) according to the manufacturer's
instructions for paired-end sequencing (2×150 bp). Sufficient
double-stranded complementary DNA (dsDNA) for library preparation
was generated through SMART pre-amplification (cat. no.
634925/634926; Takara Bio USA, Inc.). The dsDNA was then fragmented
using dsDNA fragmentase, and fragments of the desired size were
selected with sample purification beads. The resulting ligation
product was amplified by PCR to construct the sequencing library.
Raw reads obtained from sequencing included adapter sequences and
low-quality bases. These were filtered using Cutadapt (version:
cutadapt-1.9) (28) to generate
high-quality clean reads. The clean data were then aligned to the
rat reference genome for gene expression quantification and
subsequent bioinformatic analyses.
Molecular docking and molecular dynamics
simulations
The predicted protein structures were generated
using AlphaFold (https://deepmind.google/technologies/alphafold/). Both
structures were subsequently prepared with AutoDockTools-1.5.7
(https://autodock.scripps.edu), including
manual removal of water molecules, addition of hydrogens, and other
structural refinements. Protein-protein docking was then carried
out using the GRAMM web server (29-31). The resulting complex was further
optimized in AutoDockTools-1.5.7. Finally, protein-protein
interaction (PPI) analysis and visualization were performed using
PyMOL (pymol.org).
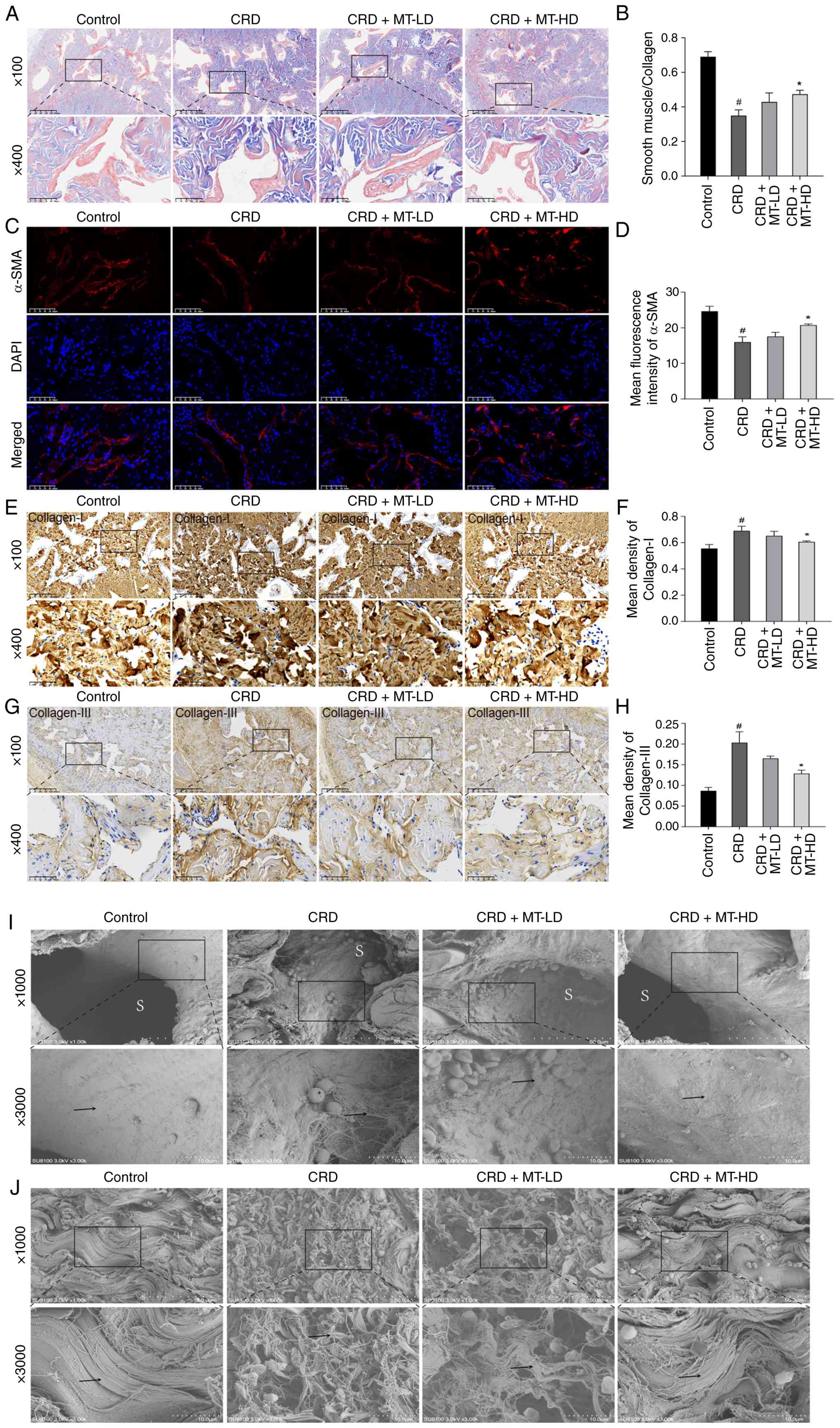
Masson trichrome staining
Penile corpus cavernosum was fixed in 4%
paraformaldehyde, paraffin-embedded, and sectioned at 5 μm.
Histological analysis included Masson trichrome staining per
standard protocols. Smooth muscle and collagen content in penile
corpus cavernosum were assessed by calculating their area ratio
using Image-Pro Plus software (ver. 6.0; Media Cybernetics,
Inc.).
Immunofluorescence (IF) staining
The paraffin-embedded sections of penile corpus
cavernosum tissues and HUVECs slides were used for IF to explore
the expression of target proteins. The primary antibodies used for
incubating sections overnight at 4°C were rabbit anti-eNOS (1:100;
cat. no. AF0096; Affinity Biosciences), anti-Nrf2 (1:200; cat. no.
33123-1-AP; Proteintech Group, Inc.), anti-HO-1 (1:100; cat. no.
10701-1-AP; Proteintech Group, Inc.), anti-α-SMA (1:100; cat. no.
14395-1-AP; Proteintech Group, Inc.) and anti-NLRP3 (1:100; cat.
no. DF7438; Affinity Biosciences). Then, appropriate secondary
antibodies [Alexa Fluor 488-conjugated goat anti-rabbit IgG (H+L);
1:200; cat. no. A32731; Invitrogen; Thermo Fisher Scientific, Inc.]
were selected for further incubation. A total of five random fields
per group were imaged. Semi-quantitative analysis of relative
fluorescence intensity was conducted using Image-Pro Plus software
(v6.0).
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) staining for pyroptosis
detection
To detect pyroptotic cells, TUNEL staining was
performed using a commercial kit according to the manufacturer's
instructions, with modifications for paraffin-embedded sections.
Briefly, penile corpus cavernosum tissue sections were dewaxed and
rehydrated. After washing with PBS (three times, 5 min each),
sections were incubated with proteinase K working solution (1
μl of 100X proteinase K in 99 μl PBS) for 20 min at
37°C to permeabilize the tissue. Following three additional PBS
washes, sections were incubated with TdT equilibration buffer for
20 min at 37°C. The buffer was then removed, and labeling working
solution (35 μl TdT equilibration buffer+10 μl
labeling solution + 5 μl TdT enzyme) was added to each
section. Sections were incubated in a humidified chamber for 60 min
at 37°C in the dark. After three PBS washes, nuclei were
counterstained with DAPI working solution [4 μl DAPI reagent
(25 μg/ml) in 96 μl PBS] for 5 min at room
temperature in the dark. Sections were washed four times with PBS,
mounted with anti-fade mounting medium, and immediately observed
under a fluorescence microscope.
Double IF for Caspase-1 and TUNEL
For simultaneous detection of Caspase-1 and TUNEL,
sections were first processed for TUNEL staining as aforementioned,
followed by IF staining for Caspase-1 using rabbit anti-Caspase-1
primary antibody (1:100; cat. no. 31020-1-AP; Proteintech Group,
Inc.) and appropriate Alexa Fluor-conjugated secondary antibody.
Nuclei were counterstained with DAPI. Co-localization of Caspase-1
and TUNEL signals was assessed to identify pyroptotic cells
(18,32). The percentage of
Caspase-1+/TUNEL+ double-positive cells was
calculated from at least five random fields per section.
Immunohistochemistry (IHC) staining
The paraffin-embedded sections of penile corpus
cavernosum tissues were fixed in 4% paraformaldehyde at 4°C for 24
h, embedded in paraffin, and cut into sections as aforementioned.
Sections were incubated with rabbit anti-eNOS (1:100; Affinity
Biosciences), anti-Nrf2 (1:200; Proteintech Group, Inc.), anti-HO-1
(1:100; Proteintech Group, Inc.), anti-Collagen-I (1:200; Abcam)
and anti-Collagen-III (1:200; Abcam) overnight at 4°C. Then the
sections were washed by PBS and incubated by relevant
HRP-conjugated goat anti-rabbit IgG (H+L) secondary antibodies
(1:500; cat. no. 31460; Thermo Fisher Scientific, Inc.) for 1 h at
room temperature. Five random fields per group were imaged.
Semi-quantitative analysis of relative fluorescence intensity was
conducted using Image-Pro Plus software (v6.0).
Scanning electron microscope (SEM)
Fresh penile corpus cavernosum tissues were
dissected into ≤1 mm3 fragments and fixed in 2.5%
glutaraldehyde for 4 h. Samples were post-fixed in 1% osmium
tetroxide for 2 h at room temperature. After dehydration through a
graded ethanol series and transition in propylene oxide, tissues
were embedded in resin and sectioned into 60-80 nm ultrathin
slices. Sections were dual-stained with uranyl acetate and lead
citrate, then observed and imaged using a SEM.
Measurement of NO and cGMP levels
NO and cGMP levels were quantified in fresh penile
corpus cavernosum tissues and HUVECs. NO Assay Kit (cat. no.
A013-2-1; Nanjing Jiancheng Bioengineering Research Institute) was
applied to detect the level of NO. cGMP ELISA Kit (cat. no.
JM-01434R2; JINGMEI; http://jsjmsw.com/) was used to measure the
concentration of cGMP. They were performed according to the
manufacturer's protocols. The levels of NO and cGMP were normalized
to the protein concentration.
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from rat penile corpus
cavernosum tissues and HUVECs using a Rapid RNA Isolation Kit (cat.
no. G3013; Wuhan Servicebio Technology Co., Ltd.). cDNA was
synthesized with a PrimeScript™ RT Reagent Kit (cat. no. RR047A;
Takara Bio, Inc.) according to the manufacturer's instructions.
RT-qPCR was performed on a CFX96 Real-Time System (Bio-Rad
Laboratories, Inc.) using TB Green Premix Ex Taq II (cat. no.
RR820A; Takara Bio, Inc.). The thermocycling conditions were as
follows: Initial denaturation at 95°C for 30 sec, followed by 40
cycles of denaturation at 95°C for 5 sec and annealing/extension at
60°C for 30 sec. A melting curve analysis was performed after
amplification to verify the specificity of the PCR products (95°C
for 15 sec, 60°C for 1 min, then gradually increasing to 95°C). The
relative expression levels of target genes were calculated using
the 2-ΔΔCq method (33), where ΔCt=Ct(target)-Ct(GAPDH) and
ΔΔCt=ΔCt(treatme nt)-ΔCt(control). The specific primers used in
RT-qPCR are shown in Tables SI and
SII.
Western blotting
Penile corpus cavernosum tissues and HUVECs were
lysed in RIPA buffer (cat. no. R0010; Beijing Solarbio Science
& Technology Co., Ltd.) containing protease inhibitor cocktail
(cat. no. HY-K0010; MedChemExpress), followed by sonication and
centrifugation (12,000 × g, 15 min, 4°C) to collect supernatants.
Total protein concentration was determined by BCA assay (cat. no.
AR0146; Boster Biological Technology). Protein samples (30
μg) were separated by 10% SDS-PAGE and transferred to PVDF
membranes. Membranes were blocked with 5% BSA (http://www.genenode.com)/TBST (0.1% Tween-20) for 1 h
at room temperature, then incubated overnight at 4°C with primary
antibodies: Rabbit anti-Nrf2 (1:2,000) and anti-HO-1 (1:1,000).
After primary incubation, membranes were probed with
species-matched HRP-conjugated secondary antibodies (1:5,000) for 1
h at room temperature. Band intensities were quantified using
ImageJ software (version 1.53t; National Institutes of Health) and
normalized to GAPDH.
Analysis of ROS and mitochondrial
ROS
Cellular ROS and mitochondrial ROS (mtROS) levels
were assessed using the fluorescent probes H2DCFH-DA (cat. no.
D6470; Beijing Solarbio Science & Technology Co., Ltd.) and
MitoSOX™ Red Mitochondrial Superoxide Indicator (cat. no. HY-D1055;
MedChemExpress), respectively.
For tissue sample preparation, penile corpus
cavernosum tissue fragments were enzymatically digested at 37°C for
30 min, filtered through a 70-μm nylon mesh, and centrifuged
at 500 × g. The pellet was washed twice with PBS and subsequently
incubated with 10 μM H2DCFH-DA at 37°C for 1 h. After
incubation, the samples were centrifuged at 1,000 × g and washed
again twice with PBS. Fluorescence intensity was measured using a
microplate reader at Ex/Em=488/525 nm.
For the cell-based assay, HUVECs cultured on
adherent slides were incubated with 10 μM H2DCFH-DA at 37°C
for 30 min under light-protected conditions, following the
manufacturer's instructions. Fluorescence images were acquired
immediately using a fluorescence microscope, with five random
fields captured per sample. Semi-quantitative analysis of
fluorescence intensity was performed using Image-Pro Plus software
(v6.0). For mtROS detection, the MitoSOX™ Red probe was used
according to the manufacturer's protocol, and fluorescence was
detected at Ex/Em=510/580 nm under light-protected microscopy.
Detection of malondialdehyde (MDA),
reduced glutathione (GSH), SOD and total antioxidant capacity
(T-AOC) levels
MDA, GSH, SOD and T-AOC levels were quantified in
rat CC tissue homogenates and HUVECs lysates using commercial kits:
MDA (cat. no. BC0025), GSH (cat. no. BC1175), SOD (cat. no. BC5165)
and T-AOC (cat. no. BC1315; all from Beijing Solarbio Science &
Technology Co., Ltd.). Assays were performed strictly according to
manufacturer's protocols. All values were normalized to total
protein concentration.
Detection of mitochondrial membrane
potential (MMP)
MMP was assessed using JC-1 staining (cat. no.
E-CK-A301; Elabscience Biotechnology, Inc.) and analyzed via
fluorescence microscopy.
Statistical analysis
All results were presented as mean ± standard
deviation and analyzed with GraphPad Prism software (version 9.5.0;
Dotmatics). Shapiro-Wilk test was used for normality and Levene's
test was used for homogeneity of variance before applying
parametric tests. For data that did not meet normality or equal
variance assumptions, Kruskal-Wallis H test followed by Dunnett's
multiple comparisons test was utilized. It was clarified that
Dunnett's test was used for comparisons against the control group
(primary aim), while Tukey's or Dunnett's tests were applied for
all pairwise comparisons or non-parametric data, respectively.
P<0.05 was considered to indicate a statistically significant
difference. The number of independent biological replicates were as
follows: n=6 per group for animal experiments; n=3 independent
culture batches for cell experiments, each with technical
triplicates.
Results
MT protects CRD-induced ED and preserves
eNOS-NO-cGMP pathway in penile corpus cavernosum and HUVECs
As was shown, the CRD group (44.91±7.51 mmHg) showed
significantly lower mICP than Control group (76.82±3.81 mmHg,
P<0.0001); however, the CRD + MT-LD (57.03±2.03 mmHg, P=0.0008)
and CRD + MT-HD (66.15±2.49 mmHg, P<0.0001) groups significantly
increased mICP than the CRD group (Fig. 1A and C); the total ICP also
showed a similar trend (Fig. 1A and
D). Meanwhile, the CRD group significantly reduced mICP/MAP
ratio (0.38±0.07) than Control group (0.64±0.03, P<0.0001),
while the CRD + MT-LD (0.50±0.02, P=0.0013) and CRD + MT-HD
(0.59±0.05, P<0.0001) significantly increased it (Fig. 1A and F). Moreover, the bICP and
MAP were consistent among the four groups (P>0.05) (Fig. 1A, B and E). These results
suggested that CRD impaired rats' normal erectile function, while
MT demonstrated a protective effect for CRD-induced ED. In
addition, these procedures had no impact on rats' mean body weight
(P>0.05) (Fig. S1), which
excluded potential confounding such as systemic metabolic
effects.
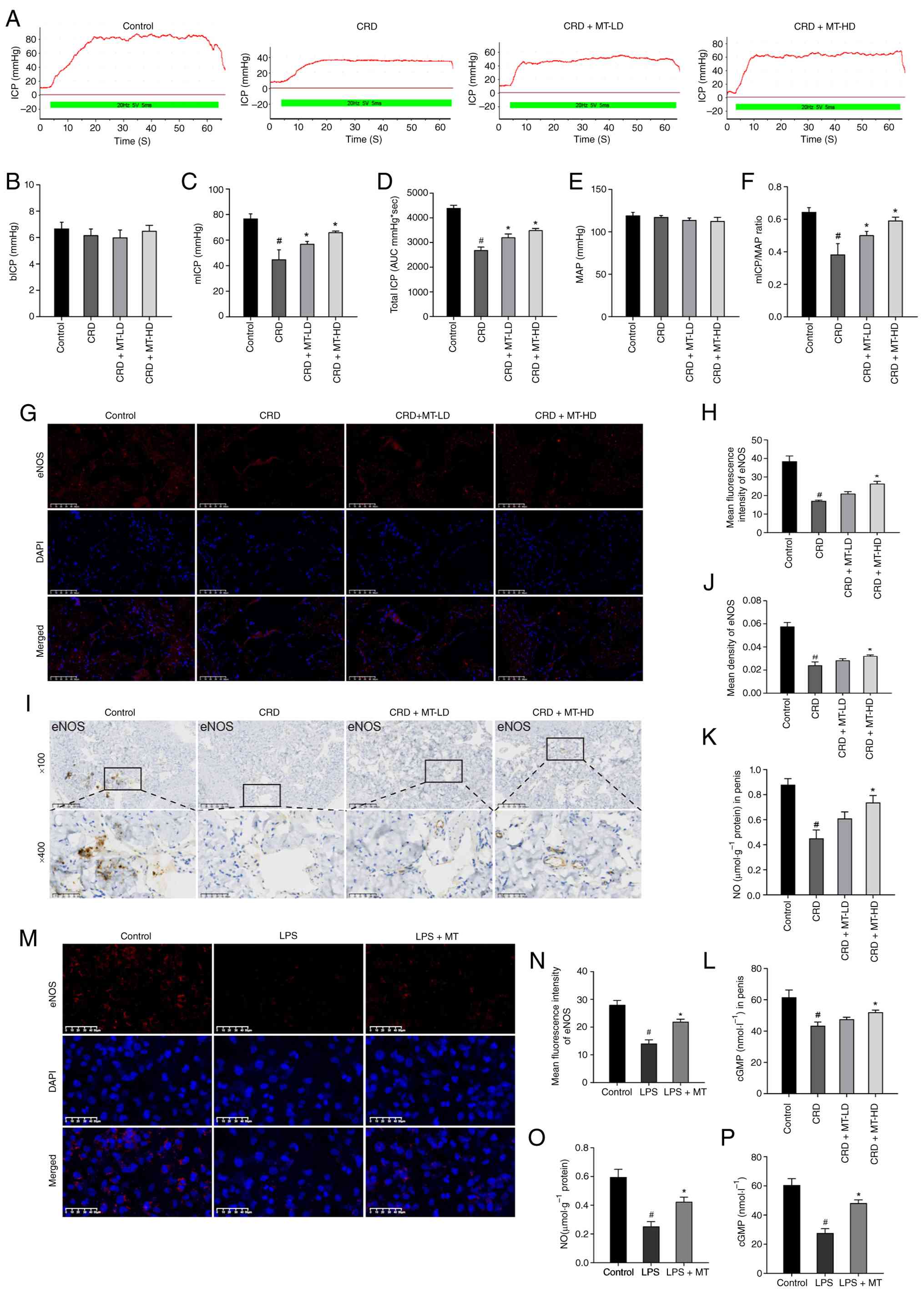 | Figure 1MT protects CRD-induced ED and
preserves eNOS-NO-cGMP pathway in penile corpus cavernosum and
HUVECs. (A) Representative ICP with the electrical stimulation of
the CN. (B-F) The erectile function was evaluated by (B) bICP, (C)
mICP, (D) Total ICP (represented by area under the curve), (E) MAP
and (F) mICP/MAP. (G and I) Representative images of
immunofluorescence staining and IHC staining for eNOS in penile
corpus cavernosum, respectively. (H and J) Semi-quantitative data
of eNOS expression as relative fluorescence intensity and density
in penile corpus cavernosum, respectively. There were three
replicates in this experiment (n=3). (K and L) NO content and cGMP
content in penile corpus cavernosum, respectively. (M)
Representative images of IHC staining for eNOS in HUVECs. (N)
Semi-quantitative data of eNOS expression as relative fluorescence
intensity in HUVECs. (O and P) NO content and cGMP content in
HUVECs, respectively. #P<0.05 compared with the
control group; *P<0.05 compared with the CRD or LPS
group. MT, melatonin; CRD, circadian rhythm disruption; ED,
erectile dysfunction; eNOS, endothelial nitric oxide synthase; NO,
nitric oxide; cGMP, cyclic guanosine monophosphate; HUVECs, human
umbilical vein endothelial cells; ICP, intracavernous pressure; CN,
cavernous nerve; mICP, maximal ICP; MAP, mean arterial pressure;
IHC, immunohistochemistry; LD, low dosage; HD, high dosage. |
The IF and IHC staining of penile corpus cavernosum
revealed that CRD significantly reduced eNOS expression, which was
partially preserved by MT-LD and MT-HD (Fig. 1G-J). Meanwhile, the penile corpus
cavernosum of CRD group also demonstrated significantly decreased
NO and cGMP concentrations, which was also inhibited by MT-LD and
MT-HD (Fig. 1K and L). These
in vivo results also suggested that MT could preserve the
CRD-induced ED.
To further confirm these findings, LPS-treated
HUVECs were selected to imitate CRD stimulation in vitro
(34,35) to verify the underlying mechanism,
while the specific concentrations and exposure times of LPS and MT
for HUVECs were provided in the following statement. Firstly,
HUVECs were treated with different concentrations of LPS and MT for
different time intervals to evaluate their cytotoxicity. It was
found that the concentration of LPS up to 0.5 mg/l and MT up to
1,600 μM had significant effects on the viability of HUVECs
for 24 h (Fig. S3A and C).
However, LPS with 1.0 mg/l for 24 h reduced HUVECs viability to
52.03% than the control group, indicating a cytotoxic effect with
an approximate IC50 of 1.0 mg/l, whereas MT with 800
μM was well-tolerated with no significant effect on
viability even after 48 h (Fig. S3B
and D). Thus, HUVECs treated with LPS at 1.0 mg/l were used to
establish a CRD stimulation model in vivo. Therefore, cells
were incubated with 400 or 800 μM MT for 2 h before exposure
to 1.0 mg/l LPS. The protective effect of 800 μM MT was
significantly more pronounced, a finding consistently corroborated
by both CCK-8 and phase-contrast microscopy assay (Fig. S3E and F). As was shown, LPS
could significantly reduce eNOS expression while decreased NO and
cGMP levels, confirming that CRD could impair HUVECs; however,
these damages were reversed by MT (Fig. 1M-P).
Collectively, these in vivo and in
vitro results suggested that CRD impaired rats' erectile
function and injured HUVECs by disrupting eNOS-NO-cGMP pathway,
however, MT treatment could reduce these alterations to ameliorate
CRD-induced ED.
Verification of CRD model establishment
at the molecular level
To confirm that photoperiodic manipulation
successfully induced circadian rhythm disorders, the expression of
core clock genes was first analyzed in the corpus cavernosum tissue
of a model rat by sequencing data. As revealed in Fig. S4, firstly, RNA quality was
evaluated using the Affy package in R software (version 4.2.1;
https://www.R-project.org) and gene expression
levels were normalized using limma package (https://bioconductor.org/packages/limma/) and
StringTie software. (https://github.com/gpertea/stringtie). Visual analysis
was conducted using a box plot of relative logarithmic expression
levels, and the results demonstrated favorable consistency among
samples within each group (Fig.
S4A). Secondly, the limma package and Wilcoxon test were used
to screen differentially expressed genes (DEGs), with a screening
threshold set at |logFC|>0.5 and FDR<0.05. The results showed
that a total of 872 DEGs were identified between the Control group
and the CRD group, of which 389 genes were significantly
downregulated, and 483 genes were significantly upregulated
(Fig. S4B). Further
differential expression analysis was conducted on the core clock
genes in the selected DEGs. The heatmap results identified that 12
core clock genes, including Per1/2/3, Npas2, Csnk1e, Cry1/2,
Nr1d1/2, Clock, Rorα and Timeless, were generally upregulated in
the CRD group, indicating the successful construction of a CRD
model by changing the light cycle (Fig. S4C). Secondly, through IF, IHC,
WB and RT-qPCR experiments, it was found that compared with the
control group, the protein and mRNA expression levels of the core
circadian clock gene Per1 in the corpus cavernosum tissue of CRD
group rats were significantly increased (Fig. S5A-G). This result is consistent
with RNA-seq data, and a CRD model was successfully established at
the functional and molecular levels.
Rationale for using LPS-induced HUVECs to
mimic CRD in vitro
Haspel et al (34) demonstrated that LPS exposure
in vitro and in vivo leads to reprogramming of the
circadian clock, including changes in the expression of core clock
genes such as Per1, Per2 and Bmal1. Ryzhikov et al (35) further demonstrated that LPS
induces a CRD-like state in cultured cells, characterized by loss
of rhythmic expression of clock control genes. In the authors'
preliminary experiments, LPS treatment significantly upregulated
the expression of Per1 in HUVECs (Fig. S6), simulating the molecular
clock disruption observed in the corpus cavernosum of rats with
CRD. This validates the use of LPS treated HUVEC as a relevant
in vitro model for studying the mechanism of CRD-induced
endothelial injury.
MT reduces microstructural damage of
penile corpus cavernosum
For the microstructural changes of penile corpus
cavernosum, Masson trichrome staining showed that SMC/Collagen
ratio in CRD group was significantly reduced, which was increased
in CRD + MT-LD and CRD + MT-HD groups (Fig. 2A and B). IF staining revealed
that the reduced α-SMA in CRD group was ameliorated in CRD + MT-LD
and CRD + MT-HD groups (Fig. 2C and
D), while IHC staining presented that CRD + MT-LD and CRD +
MT-HD significantly attenuated Collagen-I and III deposition than
CRD group (Fig. 2E-H). For the
SEM of penile corpus cavernosum, CRD group presented that collagen
fibers (→) in the outer layers of tunica albuginea were irregularly
lined and sinusoidal (S) spaces were significantly narrowed;
however, these collagen fibers (→) were regularly lined and
sinusoidal (S) spaces were widened in CRD + MT-HD group (Fig. 2I and J). All these demonstrated
that CRD impaired the microstructural integrity of penile corpus
cavernosum, which were partially reversed through MT
intervention.
Oxidative stress is a core pathway in the
treatment of ED with MT
To elucidate the therapeutic mechanisms of MT on
CRD-induced ED, a systematic bioinformatics analysis was performed.
Firstly, bioinformatic screening across multiple databases
[including GeneCards (https://www.genecards.org), STRING (https://string-db.org), DAVID, PubChem (https://pubchem.ncbi.nlm.nih.gov), CTD
(https://ctdbase.org) and OMIM (https://www.omim.org)] identified 1,500 ED-associated
genes and 825 predicted MT target genes, with 284 overlapping
targets selected for further analysis (Fig. 3A). PPI network construction and
analysis of these overlapping targets (Fig. 3B and C) identified 63 core
targets (Fig. 3D), which notably
included oxidative stress-related genes (OSRGs). These findings
establish oxidative stress as a core pathway through which MT
exerts its therapeutic effects in ED.
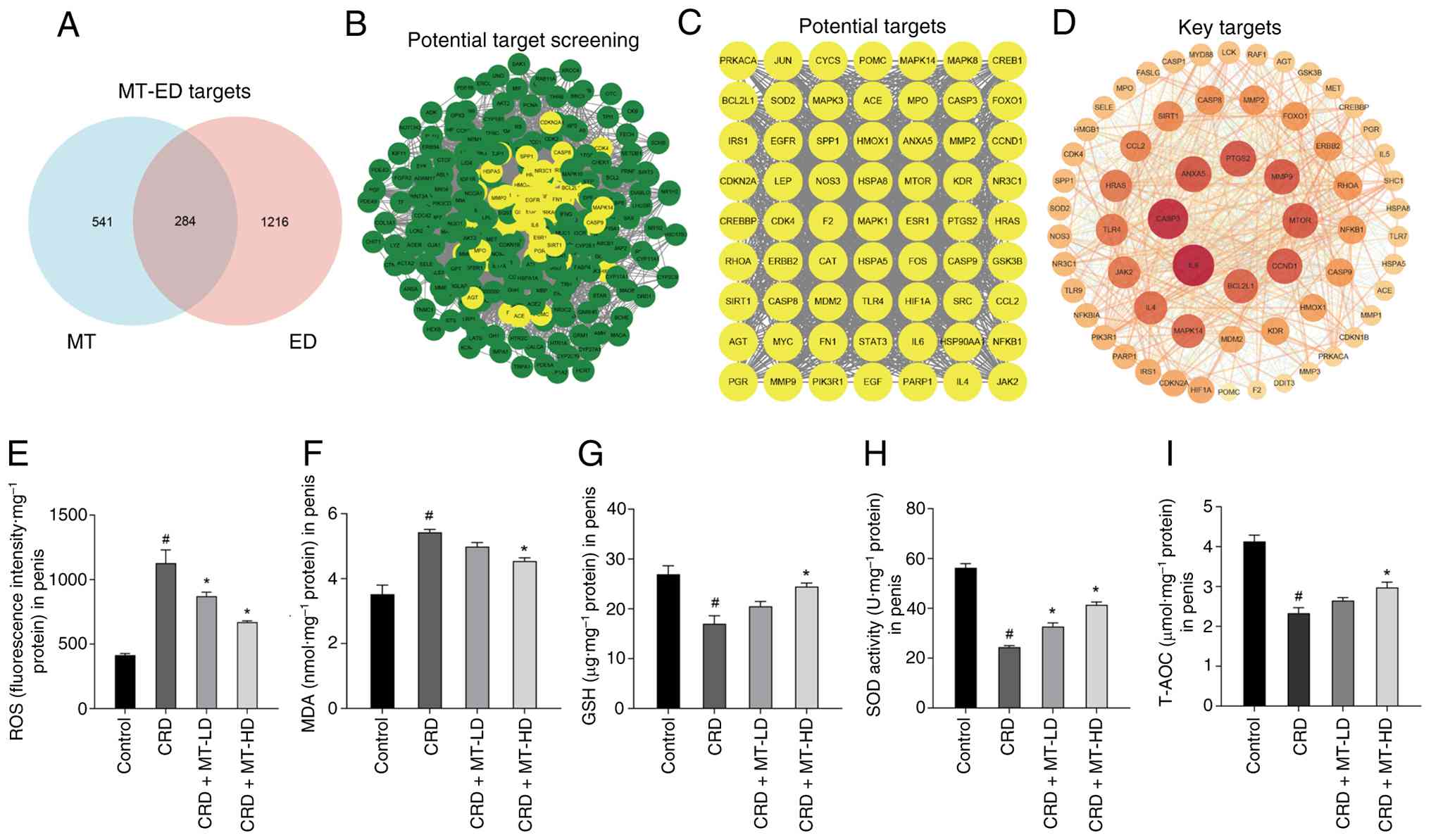 | Figure 3Oxidative stress is a core pathway in
the treatment of ED with MT. (A) Overlapping targets between MT and
ED. (B) Screening process of potential therapeutic targets. (C)
Final set of candidate targets. (D) Core targets derived from
protein-protein interaction network analysis. (E-I) Levels of ROS,
MDA and GSH, as well as SOD activity and T-AOC, were measured in
the penile corpus cavernosum of CRD rats, respectively.
#P<0.05 compared with the control group;
*P<0.05 compared with the CRD group. ED, erectile
dysfunction; MT, melatonin; ROS, reactive oxygen species; MDA,
malondialdehyde; GSH, reduced glutathione; SOD, superoxide
dismutase; T-AOC, total antioxidant capacity; CRD, circadian rhythm
disruption; LD, low dosage; HD, high dosage. |
The oxidative stress level of rats' penile corpus
cavernosum was thus evaluated. The oxidative stress markers (ROS
and MDA) in CRD group were significantly increased and antioxidant
parameters (GSH, SOD and T-AOC) were decreased, all of which were
partially reversed by MT-LD and MT-HD (Fig. 3E-I). All these inferred that
oxidative imbalance is an essential pathological process for
CRD-induced ED, which is also a core therapeutical mechanism for
MT.
MT inhibits oxidative stress via
Nrf2/HO-1 pathway in penile corpus cavernosum and HUVECs
Sequencing for penile corpus cavernosum was further
performed, which identified 1,320 DEGs between CRD and CRD + MT-HD
groups (Fig. 4A); all the top 20
OSRGs exhibited significantly differential expression. Notably, the
core Nrf2 and HO-1 of OSRGs in CRD + MT-HD group were significantly
upregulated compared with CRD group (Fig. 4B), suggesting their crucial role
in mediating the therapeutic effect of MT. To further investigate
whether MT directly interacts with Nrf2 and HO-1, molecular docking
was performed. The results showed binding energies of -5.7 kcal/mol
for Nrf2 and -6.4 kcal/mol for HO-1, indicating stable binding
between MT and both proteins (Fig.
4C and D). Given the core role of Nrf2 (36) and HO-1 (37) in oxidative stress response, it
was hypothesized that MT primarily exerted antioxidant effects
through activating Nrf2/HO-1 pathway.
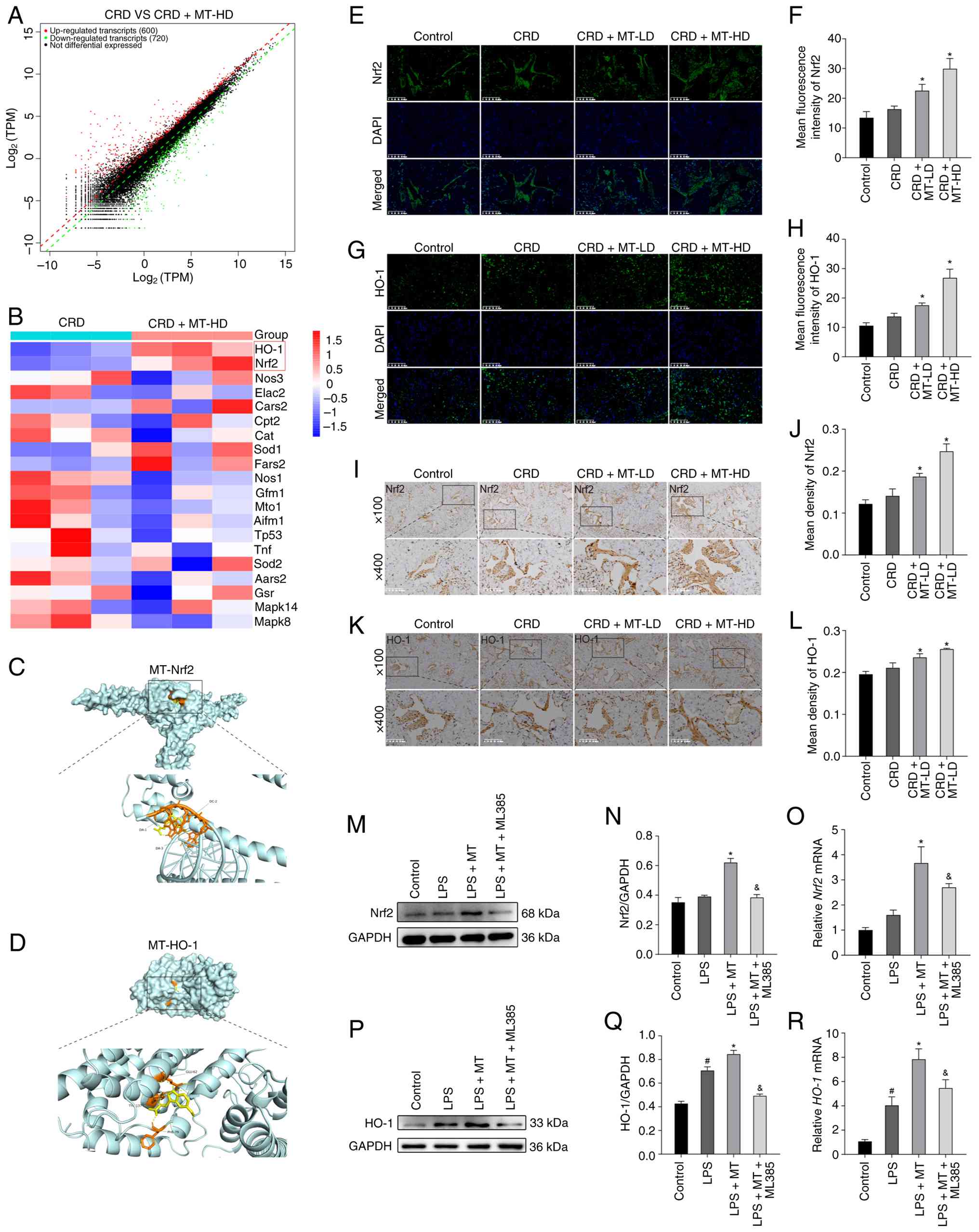 | Figure 4Identification of key targets and
analysis of pathway effects in MT-mediated amelioration of ED. (A)
Volcano plot showing the differentially expressed genes between the
CRD and CRD + MT-HD groups. (B) Differential expression of
oxidative stress-related genes in penile corpus cavernosum: a
comparison between CRD and CRD + MT-HD groups. (C and D)
Three-dimensional molecular docking of MT with Nrf2 and HO-1. (E
and G) Representative images of immunofluorescence staining for
Nrf2 and HO-1 in penile corpus cavernosum, respectively. (F and H)
Semi-quantitative data of Nrf2 and HO-1 expression as relative
fluorescence intensity in penile corpus cavernosum, respectively.
(I and K) Representative images of immunohistochemistry staining
for Nrf2 and HO-1 in penile corpus cavernosum, respectively. (J and
L) Semi-quantitative data of Nrf2 and HO-1 expression as relative
density in penile corpus cavernosum, respectively. (M and P)
Representative western blot bands for Nrf2 and HO-1 in HUVECs,
respectively. (N and Q) Semi-quantitative analysis of protein
levels of Nrf2 and HO-1 in HUVECs, respectively. (O and R)
Semi-quantitative analysis of mRNA levels of Nrf2 and HO-1 in
HUVECs, respectively. There were three replicates in these
experiments. #P<0.05 compared with the control group;
*P<0.05 compared with the CRD or LPS group;
&P<0.05 compared with the LPS + MT group. MT,
melatonin; ED, erectile dysfunction; HD, high dosage; Nrf2, nuclear
factor erythroid 2-related factor 2; HO-1, heme oxygenase 1;
HUVECs, human umbilical vein endothelial cells; LPS,
lipopolysaccharide; LD, low dosage. |
In vivo analyses (IF and IHC of penile corpus
cavernosum) revealed that Nrf2 and HO-1 protein expression levels
were slightly increased in CRD rats, whereas MT intervention
significantly upregulated their levels (Fig. 4E-L); this pattern was
corroborated in BCNI rat model, that the disease itself did not
alter Nrf2 or HO-1 expression, yet antioxidant treatment elevated
them (38). To validate these
findings, the LPS-treated HUVECs were selected to imitate CRD
stimulation in vitro (34,35). Consistent with the in vivo
data, western blotting and RT-qPCR analysis demonstrated that the
levels of Nrf2 and HO-1 in the LPS + MT group were significantly
higher than LPS group, while Nrf2 inhibitor (ML385) significantly
reduced Nrf2 and HO-1 expression compared with LPS + MT group
(Fig. 4M-R).
It has been previously proved by the authors that
oxidative stress was involved in the therapeutic effects of MT on
CRD-induced ED; whether this occurs through Nrf2/HO-1 pathway
remains unclear. The present in vitro experiments
demonstrated that MT reduced the LPS-induced elevation of oxidative
stress markers (ROS, mtROS and MDA) while rescued reduction of
antioxidant parameters (MMP, GSH, SOD and T-AOC); notably, these
beneficial effects were partially reversed by Nrf2 inhibitor
(ML385), indicating that MT exerted antioxidant effects through
activating Nrf2/HO-1 pathway (Fig.
5A-J).
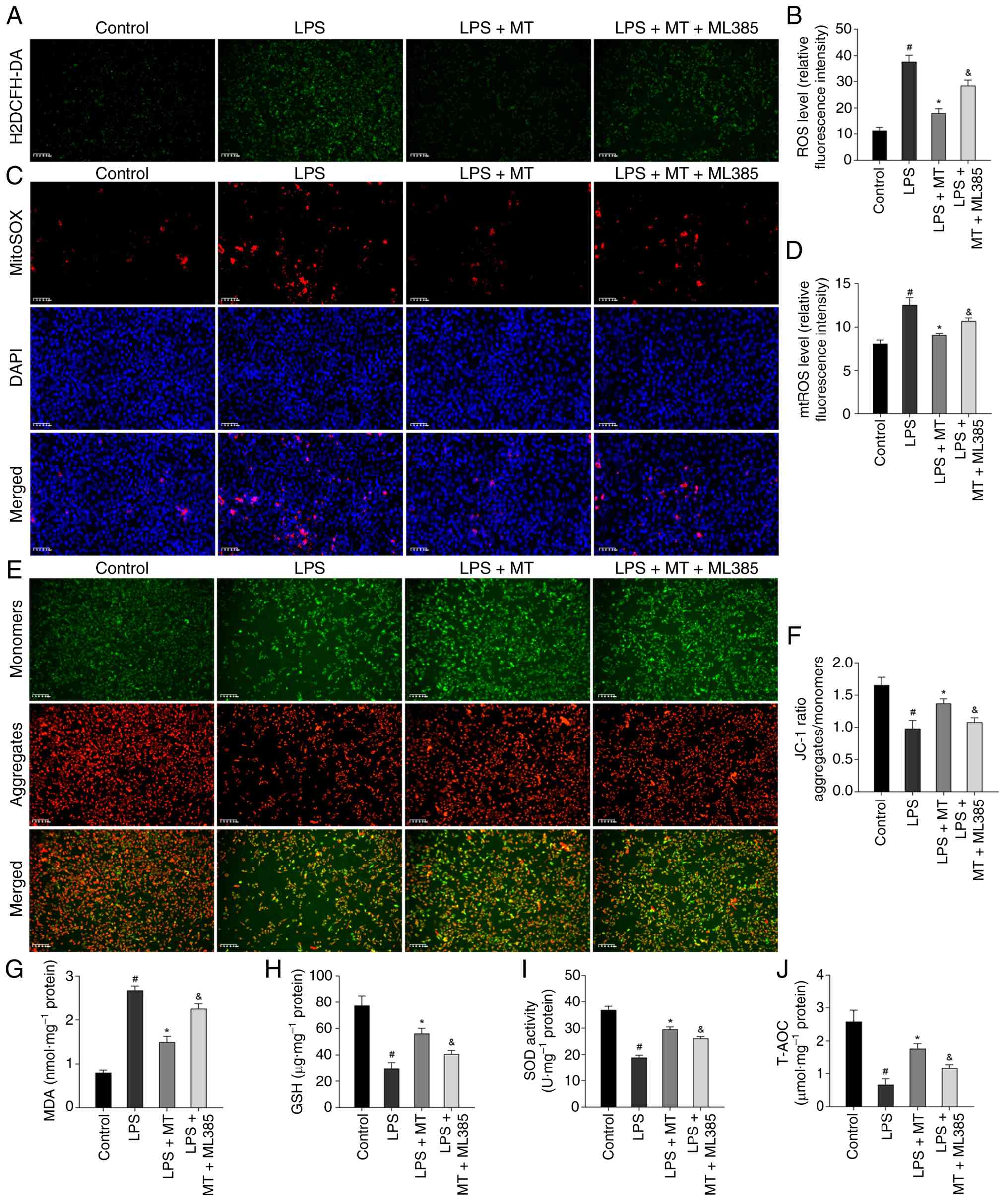 | Figure 5MT inhibits oxidative stress via
Nrf2/HO-1 pathway in HUVECs. (A) Representative images of H2DCFH-DA
staining for ROS in HUVECs. (B) Semi-quantitative data of ROS level
as relative fluorescence intensity in HUVECs. (C) Representative
images of MitoSOX Red staining for mtROS in HUVECs. (D)
Semi-quantitative data of mtROS level as relative fluorescence
intensity in HUVECs. (E) Representative images of JC-1 staining for
MMP in HUVECs. (F) Semi-quantitative data of MMP as a relative JC-1
ratio in HUVECs. (G-J) Levels of MDA, GSH, as well as SOD activity
and T-AOC were measured in HUVECs, respectively.
#P<0.05 compared with the control group;
*P<0.05 compared with the CRD or LPS group;
&P<0.05 compared with the LPS + MT group. MT,
melatonin; Nrf2, nuclear factor erythroid 2-related factor 2; HO-1,
heme oxygenase 1; HUVECs, human umbilical vein endothelial cells;
ROS, reactive oxygen species; mtROS, mitochondrial ROS; MMP,
mitochondrial membrane potential; MDA, malondialdehyde; GSH,
reduced glutathione; SOD, superoxide dismutase; T-AOC, total
antioxidant capacity; LPS, lipopolysaccharide. |
MT suppresses NLRP3-mediated pyroptosis
in penile corpus cavernosum and HUVECs
To further explore the downstream mechanism of how
MT regulates CRD-induced ED through oxidative stress, sequencing
analysis was subsequently performed on penile corpus cavernosum
tissues from CRD rat model groups. A total of 37 pyroptosis-related
DEGs were identified by intersecting RNA-seq data from CRD and CRD
+ MT-HD groups with pyroptosis-related genes from GeneCard
(Fig. 6A). PPI network analysis
was constructed using STRING and visualized in Cytoscape (version
3.10.3; https://web.cytoscape.org). Functional
enrichment analysis (DAVID; https://davidbioinformatics.nih.gov) showed
significant involvement of the 'NOD-like receptor signaling
pathway' (Fig. 6B). Based on
these findings, along with the established link between NLRP3 and
pyroptosis (39) (Fig. 6C), it was therefore hypothesized
that NLRP3-mediated pyroptosis may contribute to the development of
CRD-induced ED.
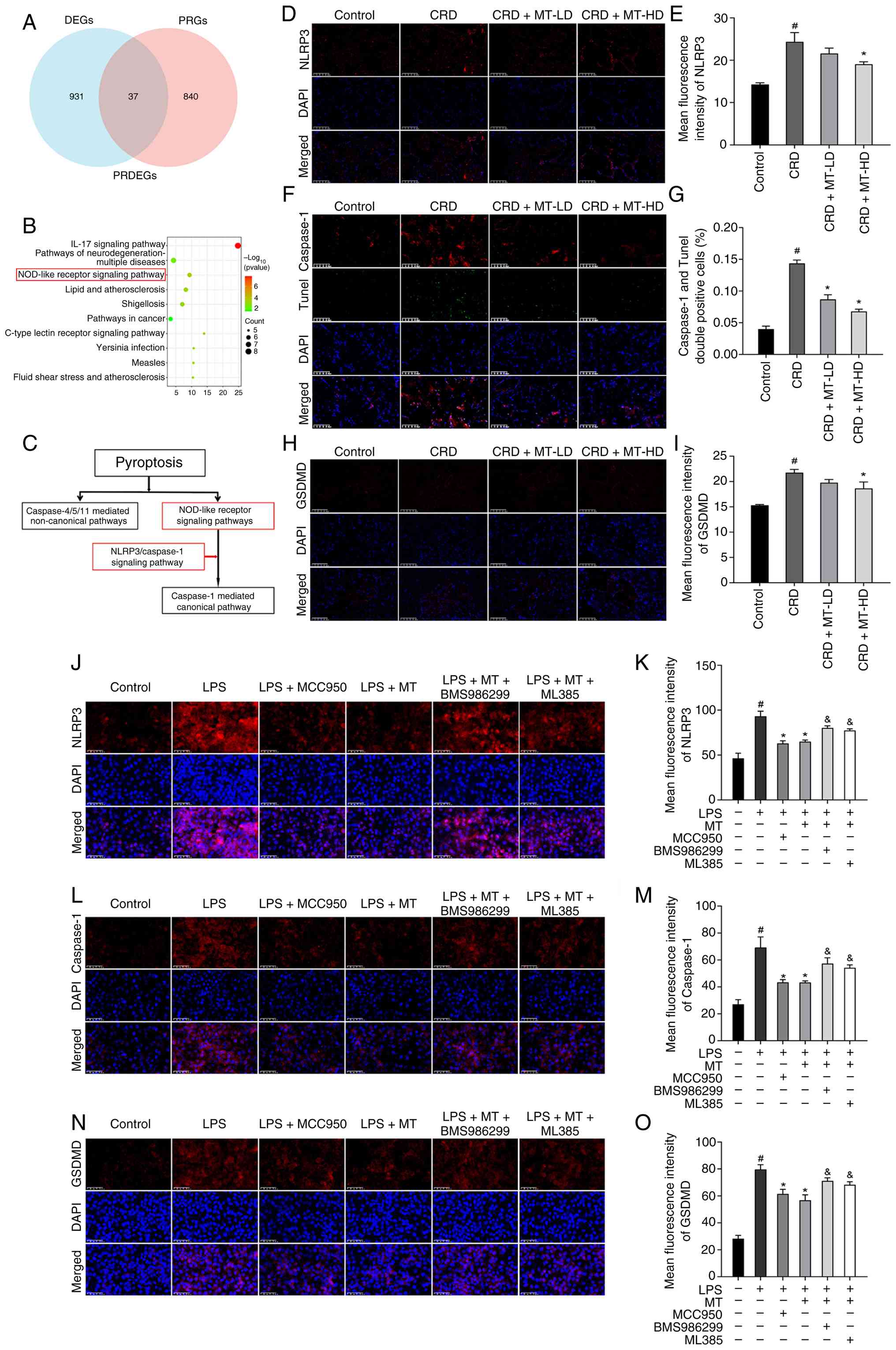 | Figure 6MT suppresses NLRP3-mediated
pyroptosis in penile corpus cavernosum and HUVECs. (A) PRDEGs were
identified from DEGs and PRGs using Venn analysis. (B) Kyoto
Encyclopedia of Genes and Genomes pathway enrichment analysis of
PRDEGs was visualized using a bubble plot. (C) The NLRP3 pathway
was a canonical pyroptosis pathway. (D, F and H) Representative
images of IF staining for NLRP3,
caspase-1+/TUNEL+, and GSDMD in penile corpus
cavernosum, respectively. (E, G and I) Semi-quantitative data of
NLRP3, caspase-1+/TUNEL+ index and GSDMD
expression as relative fluorescence intensity in penile corpus
cavernosum, respectively. (J, L and N) Representative images of IF
staining for NLRP3, caspase-1 and GSDMD in HUVECs, respectively.
(K, M and O) Semi-quantitative data of NLRP3, caspase-1 and GSDMD
expression as relative fluorescence intensity in HUVECs,
respectively. There were three replicates in these experiments.
#P<0.05 compared with the Control group.
*P<0.05 compared with the CRD or LPS group.
&P<0.05 compared with the LPS + MT group. MT,
melatonin; NLRP3, NLR family pyrin domain containing 3; HUVECs,
human umbilical vein endothelial cells; DEGS, differentially
expressed genes; PRDEGs, pyroptosis-related DEGs; IF,
immunofluorescence; GSDMD, gasdermin D; CRD, circadian rhythm
disruption; LPS, lipopolysaccharide; LD, low dosage; HD, high
dosage. |
The IF staining of penile corpus cavernosum was
performed to evaluate the core protein of pyroptosis (NLRP3,
Caspase-1 and GSDMD). As was demonstrated, NLRP3 protein expression
in rats with CRD was significantly increased, which was markedly
attenuated following CRD + MT-LD and CRD + MT-HD groups (Fig. 6D and E). Meanwhile, both the
caspase-1+/TUNEL+ index [identified as
pyroptosis (18,32)] (Fig. 6F and G) and GSDMD expression
(Fig. 6H and I) revealed
consistent alterations of NLRP3, rising under CRD and declining
after MT. The in vitro LPS-treated HUVECs also presented
similar consequences. LPS significantly elevated the key pyroptosis
protein levels (NLRP3, caspase-1 and GSDMD); however, all these
alterations were partially reversed by MT treatment (Fig. 6J-O). These demonstrated that CRD
promoted pyroptosis, whereas MT treatment alleviated this
process.
To further elucidate the mechanism by which MT
mitigates LPS-induced pyroptosis in HUVECs, a dual-intervention
strategy was employed using the NLRP3 inhibitor (MCC950) and
agonist (BMS986299). IF analysis demonstrated that both the NLRP3
inhibitor (MCC950) and MT similarly suppressed the LPS-induced
upregulation of key pyroptosis markers (NLRP3, Caspase-1 and
GSDMD). By contrast, the NLRP3 agonist (BMS986299) attenuated the
inhibitory effect of MT on these LPS-induced pyroptosis markers
(Fig. 6J-O). The present
findings suggested that CRD could lead ED through inducing
pyroptosis, whereas MT counteracted this effect through inhibiting
NLRP3 activation.
MT alleviates NLRP3-mediated pyroptosis
through suppression of oxidative stress
To further explore whether MT alleviates
NLRP3-mediated pyroptosis through oxidative stress, we incorporated
the Nrf2 inhibitor (ML385) and NLRP3 inhibitor/agonist
(MCC950/BMS986299) to enable a systematic comparison across these
targeted interventions. IF analysis revealed that MT significantly
suppressed the LPS-induced up-regulation of pyroptosis markers, an
effect that was also found by NLRP3 inhibitor (MCC950). Meanwhile,
Nrf2 inhibitor (ML385) completely abolished the protective effect
of MT, an effect that was also found by NLRP3 agonist (BMS986299)
(Fig. 6J-O). All these proved
the core mechanism that MT alleviates NLRP3-mediated pyroptosis
through suppression of oxidative stress.
Anti-oxidative stress reduces
NLRP3-mediated pyroptosis in HUVECs
N-Acetyl-L-cysteine (NAC), an amino acid derivative
of glutathione and cysteine, serves as a potent scavenger of ROS
(40) and plays a crucial role
in cellular antioxidant stress (41). To evaluate the inhibitory effect
of NAC on LPS-induced NLRP3 activation, 10 mM NAC was pretreated
for 3 h before LPS intervention in HUVECs. Initially, the impact of
NAC on LPS-treated HUVECs was assessed. LPS induced a significant
redox imbalance, that significantly elevated oxidative stress
markers (ROS, mtROS and MDA) and decreased antioxidant parameters
(MMP, GSH, SOD and T-AOC), all of which were partially reversed by
NAC treatment (Fig. 7A-J),
suggesting the ability of anti-oxidative stress.
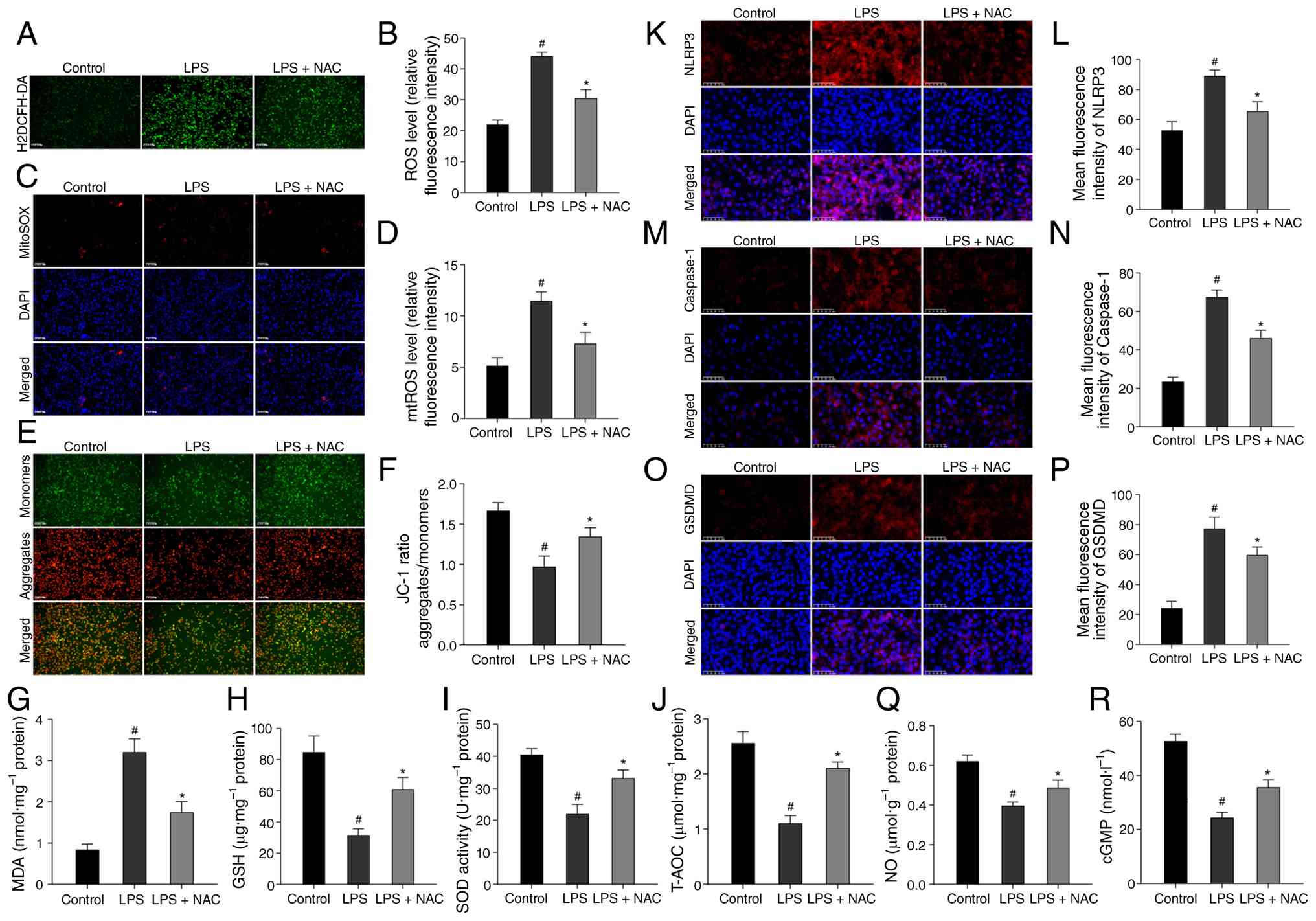 | Figure 7Antioxidative stress reduces
NLRP3-mediated pyroptosis in HUVECs. (A) Representative images of
H2DCFH-DA staining for ROS in HUVECs. (B) Semi-quantitative data of
ROS level as relative fluorescence intensity in HUVECs. (C)
Representative images of MitoSOX Red staining for mtROS in HUVECs.
(D) Semi-quantitative data of mtROS level as relative fluorescence
intensity in HUVECs. (E) Representative images of JC-1 staining for
MMP in HUVECs. (F) Semi-quantitative data of MMP as a relative JC-1
ratio in HUVECs. (G-J) Levels of MDA, GSH, as well as SOD activity
and T-AOC were measured in HUVECs, respectively. (K, M and O)
Representative images of immunofluorescence staining for NLRP3,
caspase-1 and GSDMD in HUVECs, respectively. (L, N and P)
Semi-quantitative data of NLRP3, caspase-1 and GSDMD expression as
relative fluorescence intensity in HUVECs, respectively. (Q and R)
NO content and cGMP content in HUVECs, respectively. There were
three replicates in these experiments. #P<0.05
compared with the control group; *P<0.05 compared
with the LPS group. NLRP3, NLR family pyrin domain containing 3;
HUVECs, human umbilical vein endothelial cells; ROS, reactive
oxygen species; mtROS, mitochondrial ROS; MMP, mitochondrial
membrane potential; MDA, malondialdehyde; GSH, reduced glutathione;
SOD, superoxide dismutase; T-AOC, total antioxidant capacity;
GSDMD, gasdermin D; NO, nitric oxide; cGMP, cyclic guanosine
monophosphate; LPS, lipopolysaccharide. |
Subsequently, it was explored whether the
anti-oxidative effect of NAC could influence CRD-induced
pyroptosis. As expected, LPS-treated HUVECs presented elevated
pyroptosis activation, as indicated by increased expression of
NLRP3, caspase-1 and GSDMD; however, this activation was partially
reversed by NAC (Fig. 7K-P),
suggesting that NAC reduced CRD-induced pyroptosis by inhibiting
oxidative stress.
Furthermore, the endothelial function was also
evaluated, that the endothelial function indicators of NO and cGMP
in LPS-treated HUVECs were significantly decreased, however, these
reductions were partially reversed by NAC (Fig. 7Q and R).
These data confirmed that NAC mitigated LPS-induced
endothelial impairment by reducing oxidative stress to curb
NLRP3-mediated pyroptosis, highlighting the central role of the
oxidative stress-pyroptosis axis. In conclusion, all these proved
that CRD-induced ED by inhibiting oxidative stress mediated
pyroptosis via Nrf2/HO-1 axis, which was also the core therapeutic
mechanism of MT.
Discussion
The present study aimed to elucidate the
pathological mechanisms underlying CRD-induced ED and to further
reveal the molecular basis of MT-mediated amelioration of this
process. The principal findings of the present study demonstrated
that CRD impaired erectile function in rats, suppressed the key
eNOS-NO-cGMP signaling pathway, and caused severe microstructural
damage to the penile corpus cavernosum. These structural and
functional deficits were closely associated with elevated oxidative
stress and increased pyroptosis. MT treatment successfully reversed
all of the aforementioned changes. Mechanistically, it was
demonstrated that CRD impaired endothelial function and induced
pyroptosis through an oxidative stress-triggered cascade, leading
to ED. By contrast, MT interrupted this cascade by activating the
Nrf2/HO-1 signaling axis, thereby alleviating oxidative stress and
pyroptosis, restoring endothelial integrity, and rescuing erectile
function (Fig. 8). Based on
these findings, the following sections integrate the current
results with the existing literature to clarify the
interrelationships among CRD, ED, oxidative stress and pyroptosis,
while exploring the molecular mechanisms underlying the action of
MT.
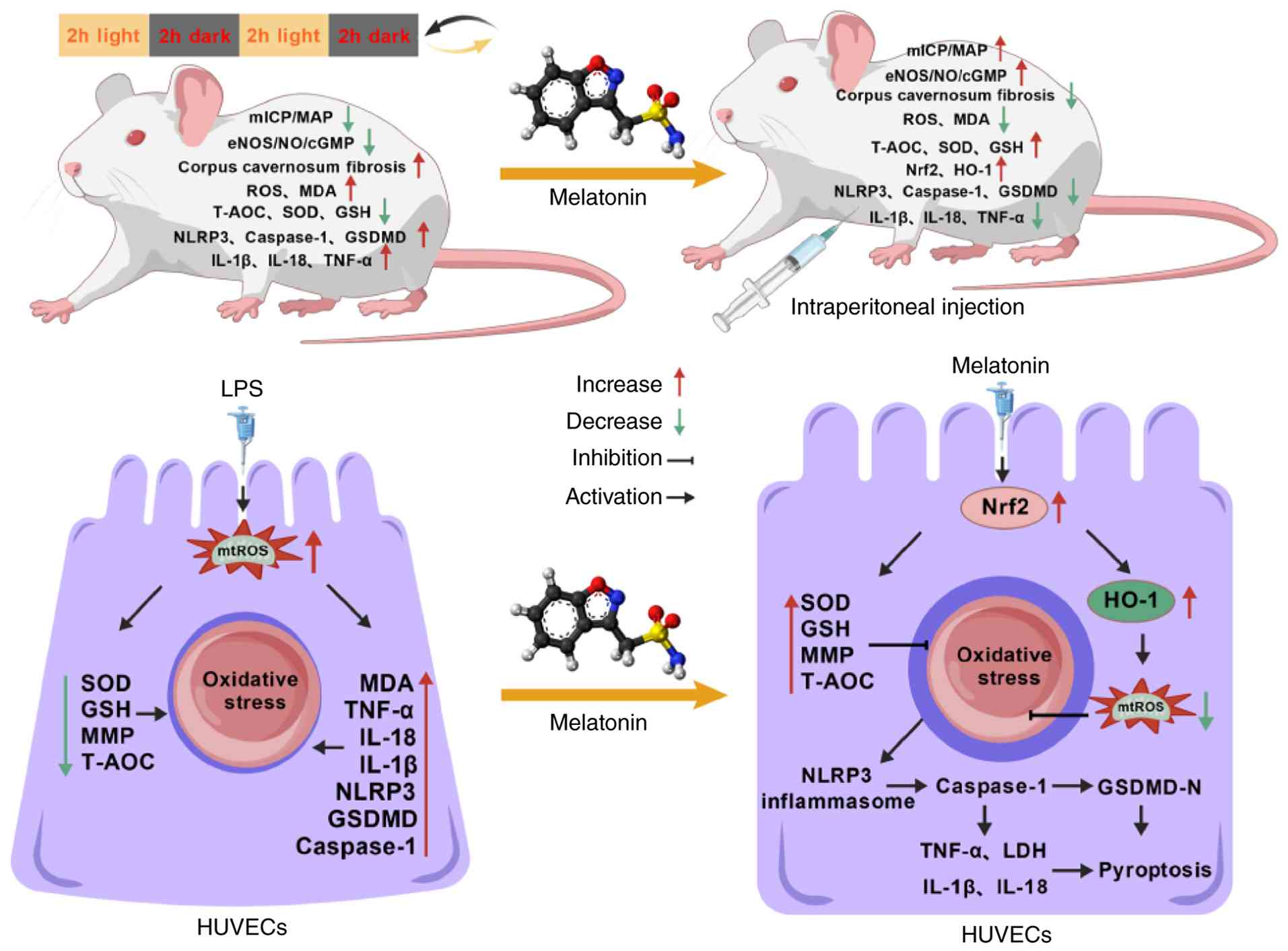 | Figure 8Summary of the therapeutic effect of
MT on CRD-induced ED via suppression of oxidative stress-mediated
pyroptosis through the Nrf2/HO-1 axis. MT, melatonin; CRD,
circadian rhythm disruption; ED, erectile dysfunction; Nrf2,
nuclear factor erythroid 2-related factor 2; HO-1, heme oxygenase
1; ICP, intracavernous pressure; mICP, maximum intracavernous
pressure; MAP, mean arterial pressure; MDA, malondialdehyde; GSH,
reduced glutathione; SOD, superoxide dismutase; T-AOC, total
antioxidant capacity; LPS, lipopolysaccharide; ROS, reactive oxygen
species; mtROS, mitochondrial ROS; MMP, mitochondrial membrane
potential; NLRP3, NLR family pyrin domain containing 3; GSDMD,
gasdermin D; eNOS, endothelial nitric oxide synthase; NO, nitric
oxide; cGMP, cyclic guanosine monophosphate. |
CRD and ED
The circadian rhythm represents a fundamental
intrinsic timekeeping system (42). Epidemiological and clinical
studies have consistently shown that CRD exposures, such as shift
work (43), frequent time-zone
changes (44) and chronic sleep
loss (45), are strongly
associated with an increased risk of ED, a finding also confirmed
by the authors' previous studies (9,10). Based on direct in vivo and
in vitro evidence, the present study establishes CRD as an
independent risk factor for ED, thereby providing direct
implications for understanding the high prevalence of ED among
shift workers and other high-risk populations. The physiological
process of erection critically depends on the NO-mediated
vasodilatory signaling pathway within penile endothelial cells
(11). The current investigation
reveals that CRD substantially impairs the activity of this central
eNOS-NO-cGMP pathway, which constitutes the direct molecular
foundation of ED pathogenesis (46). Notably, beyond the disruption of
this crucial signaling axis, the deeper pathophysiological
consequences of CRD extend to substantial tissue structural damage
and programmed cell death. These observations prompted a crucial
investigation into the specific downstream pathways through which
CRD mediates its damaging effects.
CRD and oxidative stress
The circadian rhythm system and cellular redox state
maintain a sophisticated bidirectional interplay (47). CRD disrupts this coordination,
resulting in both overproduction and inefficient clearance of ROS
(48). At the molecular level,
core circadian components directly regulate the expression of
numerous genes involved in antioxidant defense; however, CRD leads
to the dysregulation of these genes, thereby increasing
susceptibility to oxidative stress (49). Concurrently, oxidative stress
products (for example, ROS) can feed back and impair the function
of circadian rhythm proteins (for example, PER and EZH2), forming a
vicious cycle (50). At the
physiological level, CRD disrupts metabolism under the control of
the circadian rhythm (for example, glycolysis and lipophagy). This
metabolic reprogramming elevates oxidative byproducts (for example,
ROS), while the capacity for their clearance is compromised due to
CRD, collectively promoting oxidative stress (51). In the present study, penile
corpus cavernosum tissue from rats with CRD displayed clear signs
of oxidative stress; the current data strongly support a model in
which oxidative stress acts as a critical link between CRD and
downstream pathological injury. Furthermore, having identified
oxidative stress as the central pathway in the therapeutic effect
of MT on CRD-induced ED through transcriptomic analysis, it was
next sought to elucidate how MT mediates its antioxidant protection
at the molecular level.
MT inhibits oxidative stress via the
Nrf2/HO-1 pathway
Nrf2 is a master transcriptional regulator of
cellular antioxidant responses (52). Under physiological conditions,
Nrf2 is constitutively bound to Keap1, localizes in the cytoplasm,
and undergoes continuous proteasomal degradation (53). During oxidative stress, Nrf2
dissociates from Keap1 and translocates to the nucleus (54), where it binds to antioxidant
response elements (AREs) (55)
and initiates the transcription of antioxidant genes, including the
critical cytoprotective enzyme HO-1 (56). HO-1 exerts potent antioxidant and
cytoprotective effects by catalyzing the degradation of heme into
biliverdin, carbon monoxide and free iron (57). Core circadian rhythm proteins
regulate the expression and activity of Nrf2, generating circadian
oscillations in cellular antioxidant capacity (58); Nrf2 knockout attenuates the
rhythmicity of circadian genes while inducing aberrant oscillations
in stress-response genes (59).
Moreover, CRD impairs the time-giving function of the circadian
rhythm, preventing pre-activation of antioxidant defenses prior to
peak oxidative stress, thereby compromising defensive capacity when
it is most needed (58).
Ultimately, a vicious cycle emerges: CRD causes Nrf2 dysfunction
and impairs antioxidant capacity, while accumulated ROS further
disrupts circadian rhythm operation (60). Exogenous MT has been shown to
activate the Nrf2 pathway and reduce oxidative stress in various
models (61-63). The current data demonstrate that
MT mediates its antioxidant activity by activating the Nrf2/HO-1
axis, the central transcriptional regulator of endogenous
antioxidant responses, thus rebuilding cellular antioxidant
capacity and counteracting CRD-induced oxidative damage.
Oxidative stress and pyroptosis:
Downstream effector targets
If oxidative stress acts as an upstream event, what
is its downstream effector target? In recent years, pyroptosis has
been recognized as a key mechanism in the pathogenesis of multiple
organ injuries (64). Pyroptosis
is a novel form of inflammatory programmed cell death. The present
study is the first, to the best of our knowledge to show the
activation of NLRP3-mediated pyroptosis in a CRD-induced ED model.
The mechanisms to establish the causal relationship between
oxidative stress and pyroptosis in the present model were
systematically analyzed. First, it was observed that MT alleviates
pyroptosis by suppressing oxidative stress. This suggests an
association between these processes. When Nrf2 was inhibited, the
protection was abolished. This confirmed that Nrf2-dependent
antioxidant activity is indispensable for preventing pyroptosis.
The present data support that MT inhibits NLRP3-mediated pyroptosis
mainly by activating Nrf2/HO-1 and reducing oxidative stress. This
is shown by the reversal of MT's effects by the ROS scavenger NAC.
However, Nrf2 may also directly suppress NLRP3 inflammasome
activation through non-antioxidant mechanisms. Some studies have
shown that Nrf2 can downregulate NLRP3 transcriptionally by binding
to AREs in its promoter (65-67).
Nrf2 has also been reported to interact physically
with NLRP3 or its upstream regulators (for example, TXNIP) and
inhibit inflammasome assembly (68,69). In the present study, the
possibility that MT-activated Nrf2 directly represses NLRP3
activation cannot be excluded. Future studies with Nrf2 mutants
that keep transcriptional activity but lack antioxidant capacity,
or chromatin immunoprecipitation assays to examine Nrf2 binding to
the NLRP3 promoter, would clarify the roles of
antioxidant-dependent and direct pathways.
Core innovations and clinical
translational value
The core innovations of the present study are
reflected in the following three aspects: i) First report of the
therapeutic effect of MT on CRD-induced ED: Although MT has shown
protective effects in diabetic ED and neurogenic ED models, its
effect on CRD-related ED has never been investigated. The present
study is the first to demonstrate that MT significantly improves
erectile function in CRD rats, restoring the ICP/MAP ratio and
preserving the eNOS-NO-cGMP pathway. i) First revelation of the
'oxidative stress-pyroptosis' axis in CRD-related ED: Previous
mechanistic studies on ED have not linked CRD with the oxidative
stress-pyroptosis axis. The present study is the first to establish
that CRD triggers NLRP3-mediated pyroptosis through oxidative
stress, and that MT interrupts this cascade via Nrf2/HO-1
activation, representing a completely new molecular pathway in iii)
CRD-induced ED. Clinical translational value: CRD-related ED is a
modern lifestyle-associated condition affecting shift workers,
frequent travelers, and individuals with chronic sleep deprivation.
These patients often have contraindications to PDE5is (for example,
due to nitrate use for cardiovascular diseases) or suboptimal
responses because of endothelial dysfunction. As an endogenous,
low-toxicity, multi-target agent, MT represents a promising
alternative or adjunctive therapy. The present study provides a
strong preclinical rationale for future clinical trials evaluating
MT supplementation in CRD-exposed populations with ED.
There are certain limitations to the present study.
First, the in vitro experiments were performed using HUVECs
rather than corpus cavernosum endothelial cells (CCECs). These two
cell types differ in tissue origin, phenotype and physiological
function; HUVECs may not fully recapitulate the in vivo
pathophysiological state of the corpus cavernosum. Future studies
should use primary CCECs for validation.
Second, while the present study measured
mitochondrial ROS and membrane potential, key indicators of
mitochondrial oxidative stress, it is recognized that a more
comprehensive assessment of mitochondrial homeostasis would
strengthen the mechanistic understanding. Mitochondrial quality
control involves not only redox balance but also dynamic changes in
morphology (fusion/fission), biogenesis and mitophagy. Future
studies should examine mitochondrial ultrastructure using
transmission electron microscopy to visualize cristae integrity and
swelling. Additionally, markers of mitochondrial biogenesis
(PGC-1α, NRF1 and TFAM) and mitophagy (Parkin, PINK1 and LC3B)
would help determine whether MT promotes the removal of damaged
mitochondria or enhances the generation of healthy ones. It has
been previously demonstrated that MT preserves mitochondrial
function in various oxidative stress models by promoting
PINK1/Parkin-dependent mitophagy (70). It would be of great interest to
test whether similar mechanisms operate in CRD-induced ED. The
current findings provide a strong foundation for such future
investigations, which has been added as a key direction in the
authors' ongoing studies.
Third, although programmed cell death pathways
(pyroptosis, apoptosis and necroptosis) dynamically interact and
may lead to integrated processes such as PANoptosis, pyroptosis was
only assessed in endothelial cells. Markers of apoptosis
(caspase-3), ferroptosis (GPX4), or necroptosis (RIPK3) were not
systematically detected. Therefore, a comprehensive understanding
of the individual contributions and crosstalk among different cell
death modalities under pathological conditions is lacking.
Fourth, regarding the detection of pyroptosis,
total GSDMD protein expression and caspase-1/TUNEL double staining
were only assessed, without measuring the gold-standard indicators,
including the cleaved N-terminal fragment of GSDMD (GSDMD-N). Thus,
the conclusions are based primarily on NLRP3 and caspase-1
activation and morphological changes. Future studies should include
these additional endpoints to provide more definitive evidence of
pyroptosis.
In conclusion, it was demonstrated that CRD-induced
ED by triggering an oxidative stress-pyroptosis cascade.
Conversely, MT treatment effectively counteracts this pathology by
activating the Nrf2/HO-1 pathway to suppress oxidative stress,
thereby attenuating NLRP3-mediated pyroptosis and ultimately
restoring erectile function. These results provide the first
systematic evidence for the central role of the oxidative
stress-pyroptosis axis in CRD-induced ED, establishing a solid
theoretical foundation for MT as a promising therapeutic strategy
for CRD-related ED.
Supplementary Data
Availability of data and materials
The data generated in the present study may be
found in the Gene Expression Omnibus under accession number
PRJNA1466509 or at the following URL: http://www.ncbi.nlm.nih.gov/bioproject/1466509.
Authors' contributions
QYa, WL and QYu conceptualized the study, wrote the
original draft, and wrote, reviewed and edited the manuscript. JS
and LY contributed to data curation and formal analysis. JQ and XW
were involved in methodology. CG participated in investigation. FS
and TL contributed to study conceptualization, were responsible for
funding acquisition and reviewed and edited the manuscript and
confirm the authenticity of all the raw data. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The animal protocols were approved by the Animal
Ethics Committee of Guizhou Medical University (approval no.
2402989; Guiyang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Nature Science
Foundation of China (grant nos. 82360295 and 82560562), the Guizhou
Provincial Basic Research Program (Natural Science) program [grant
no. QianKeHeJiChu-zk(2025)MianShang457], the Science and Technology
Foundation Project of Guizhou Provincial Health Commission (grant
no. gzwkj2024-150), the Doctor Start-up Fund of Affiliated Hospital
of Guizhou Medical University (grant no. gyfybsky-2023-03) and the
Affiliated Hospital of Guizhou Medical University 2025 Research
Education and Research Feedback in Teaching Project (grant no.
gyfykj-2025-y24).
References
|
1
|
Maestre-Lorén F, Castillo-Garayoa JA,
López-I-Martín X, Sarquella-Geli J, Andrés A and Cifre I:
Psychological distress in erectile dysfunction: The moderating role
of attachment. Sex Med. 9:1004362021.PubMed/NCBI
|
|
2
|
Salonia A, Bettocchi C, Boeri L,
Capogrosso P, Carvalho J, Cilesiz NC, Cocci A, Corona G,
Dimitropoulos K, Gül M, et al: European association of urology
guidelines on sexual and reproductive health-2021 update: Male
sexual dysfunction. Eur Urol. 80:333–357. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Samidurai A, Xi L, Das A and Kukreja RC:
Beyond erectile dysfunction: cGMP-specific phosphodiesterase 5
inhibitors for other clinical disorders. Annu Rev Pharmacol
Toxicol. 63:585–615. 2023. View Article : Google Scholar
|
|
4
|
Brock GB, McMahon CG, Chen KK, Costigan T,
Shen W, Watkins V, Anglin G and Whitaker S: Efficacy and safety of
tadalafil for the treatment of erectile dysfunction: Results of
integrated analyses. J Urol. 168:1332–1336. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ruan W, Yuan X and Eltzschig HK: Circadian
rhythm as a therapeutic target. Nat Rev Drug Discov. 20:287–307.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McAlpine CS and Swirski FK: Circadian
influence on metabolism and inflammation in atherosclerosis. Circ
Res. 119:131–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Terentes-Printzios D, Ioakeimidis N,
Rokkas K and Vlachopoulos C: Interactions between erectile
dysfunction, cardiovascular disease and cardiovascular drugs. Nat
Rev Cardiol. 19:59–74. 2022. View Article : Google Scholar
|
|
8
|
Nguyen V, McGovern AM, Rojanasarot S,
Patel DP, Bhattacharyya S, Hargens LM, Aworunse O and Hsieh TC:
Patient out-of-pocket costs for guideline-recommended treatments
for erectile dysfunction: A medicare cost modeling analysis. Int J
Impot Res. 37:45–50. 2025. View Article : Google Scholar :
|
|
9
|
Li T, Jiang YT, Qi XZ, Chen P, Zhang JH,
Luo F, Qiao J, Gu J, Du GS and Wang Q: Circadian disturbance
induces erectile dysfunction by impairing endothelial function.
Asian J Androl. 26:205–211. 2024. View Article : Google Scholar :
|
|
10
|
Li T, Bai Y, Jiang Y, Jiang K, Tian Y,
Wang Z, Ban Y, Liang X, Luo G and Sun F: Potential effect of the
circadian clock on erectile dysfunction. Aging Dis. 13:8–23. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mao Y, Zha Y, Zang Y, Gao Y, Sun J, Liu Y,
Wang Z, Wei Z, Wang M and Yang Y: Isorhamnetin improves
diabetes-induced erectile dysfunction in rats through activation of
the PI3K/AKT/eNOS signaling pathway. Biomed Pharmacother.
177:1169872024. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oyama Y, Bartman CM, Bonney S, Lee JS,
Walker LA, Han J, Borchers CH, Buttrick PM, Aherne CM, Clendenen N,
et al: Intense light-mediated circadian cardioprotection via
transcriptional reprogramming of the endothelium. Cell Rep.
28:1471–1484.e11. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maiese K: Diabetes mellitus and glymphatic
dysfunction: Roles for oxidative stress, mitochondria, circadian
rhythm, artificial intelligence, and imaging. World J Diabetes.
16:989482025. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang X, Bian Y, Zhang R, Liu X, Ni L, Ma
B, Zeng R, Zhao Z, Song X and Liu C: Melatonin alleviates cigarette
smoke-induced endothelial cell pyroptosis through inhibiting
ROS/NLRP3 axis. Biochem Biophys Res Commun. 519:402–408. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fang Y, Tian S, Pan Y, Li W, Wang Q, Tang
Y, Yu T, Wu X, Shi Y, Ma P and Shu Y: Pyroptosis: A new frontier in
cancer. Biomed Pharmacother. 121:1095952020. View Article : Google Scholar
|
|
16
|
Zhang KJ, Wu Q, Jiang SM, Ding L, Liu CX,
Xu M, Wang Y, Zhou Y and Li L: Pyroptosis: A new frontier in kidney
diseases. Oxid Med Cell Longev. 2021:66866172021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akbal A, Dernst A, Lovotti M, Mangan MSJ,
McManus RM and Latz E: How location and cellular signaling combine
to activate the NLRP3 inflammasome. Cell Mol Immunol. 19:1201–1214.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Y, Yuan H, Shen D, Liu S, Kong W,
Zheng K, Yang J and Ge L: Artemisinin attenuated ischemic stroke
induced pyroptosis by inhibiting ROS/TXNIP/NLRP3/Caspase-1
signaling pathway. Biomed Pharmacother. 177:1168942024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bhattacharya S, Patel KK, Dehari D,
Agrawal AK and Singh S: Melatonin and its ubiquitous anticancer
effects. Mol Cell Biochem. 462:133–155. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pandi-Perumal SR, BaHammam AS, Brown GM,
Spence DW, Bharti VK, Kaur C, Hardeland R and Cardinali DP:
Melatonin antioxidative defense: Therapeutical implications for
aging and neurodegenerative processes. Neurotox Res. 23:267–300.
2013. View Article : Google Scholar
|
|
21
|
Ashrafizadeh M, Najafi M, Kavyiani N,
Mohammadinejad R, Farkhondeh T and Samarghandian S:
Anti-inflammatory activity of melatonin: A Focus on the role of
NLRP3 inflammasome. Inflammation. 44:1207–1222. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sahan A, Akbal C, Tavukcu HH, Cevik O,
Cetinel S, Sekerci CA, Sener TE, Sener G and Tanidir Y: Melatonin
prevents deterioration of erectile function in
streptozotocin-induced diabetic rats via sirtuin-1 expression.
Andrologia. 52:e136392020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Z, Zhai J, Ma J, Chen P, Lin W, Zhang
W, Xiong J, Zhang C and Wei H: Melatonin-primed mesenchymal stem
cells-derived small extracellular vesicles alleviated neurogenic
erectile dysfunction by reversing phenotypic modulation. Adv
Healthc Mater. 12:e22030872023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schwartz MD, Wotus C, Liu T, Friesen WO,
Borjigin J, Oda GA and de la Iglesia HO: Dissociation of circadian
and light inhibition of melatonin release through forced
desynchronization in the rat. Proc Natl Acad Sci USA.
106:17540–17545. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ogo FM, Siervo GEML, de Moraes AMP,
Machado KGB, Scarton SRDS, Guimarães ATB, Cecchini AL, Simão ANC,
Mathias PCF and Fernandes GSA: Extended light period in the
maternal circadian cycle impairs the reproductive system of the rat
male offspring. J Dev Orig Health Dis. 12:595–602. 2021. View Article : Google Scholar
|
|
26
|
Moustafa A: Effect of light-dark cycle
misalignment on the hypothalamic-pituitary-gonadal axis, testicular
oxidative stress, and expression of clock genes in adult male rats.
Int J Endocrinol. 2020:14268462020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yuan J, Lin H, Li P, Zhang R, Luo A,
Berardinelli F, Dai Y and Wang R: Molecular mechanisms of vacuum
therapy in penile rehabilitation: A novel animal study. Eur Urol.
58:773–780. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kechin A, Boyarskikh U, Kel A and
Filipenko M: cutPrimers: A new tool for accurate cutting of primers
from reads of targeted next generation sequencing. J Comput Biol.
24:1138–1143. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Morris GM, Huey R and Olson AJ: Using
AutoDock for ligand-receptor docking. Curr Protoc Bioinformatics
Chapter. 8:Unit 8.14. 2008.
|
|
30
|
Katchalski-Katzir E, Shariv I, Eisenstein
M, Friesem AA, Aflalo C and Vakser IA: Molecular surface
recognition: Determination of geometric fit between proteins and
their ligands by correlation techniques. Proc Natl Acad Sci USA.
89:2195–2199. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vakser IA: Long-distance potentials: An
approach to the multiple-minima problem in ligand-receptor
interaction. Protein Eng. 9:37–41. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou C, Li J, Wu X and Liu F: Activation
of spleen tyrosine kinase (SYK) contributes to neuronal pyroptosis
and cognitive impairment in diabetic mice via the
NLRP3/Caspase-1/GSDMD signaling pathway. Exp Gerontol.
198:1126262024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
34
|
Haspel JA, Chettimada S, Shaik RS, Chu JH,
Raby BA, Cernadas M, Carey V, Process V, Hunninghake GM, Ifedigbo
E, et al: Circadian rhythm reprogramming during lung inflammation.
Nat Commun. 5:47532014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ryzhikov M, Ehlers A, Steinberg D, Xie W,
Oberlander E, Brown S, Gilmore PE, Townsend RR, Lane WS, Dolinay T,
et al: Diurnal rhythms spatially and temporally organize autophagy.
Cell Rep. 26:1880–1892.e6. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yu Z, Wu D, Shi T, Chen D, Feng H, Chen H,
Lin H, Sun L and Liu W: Melatonin attenuates spinal cord injury by
regulating ferroptosis through the Nrf2/HO-1/GPX4 pathway. Mol
Neurobiol. 62:15530–15549. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Asghar MA, Wan B, Li L, Zhang J, Tang S,
Han H, Yang Y, Chu L, Zhang Q, Zhang X and Zhao Q: Micronutrient
antioxidant supplementation alleviates valproic acid-induced
oxidative stress and male infertility via the NRF2/HO-1 pathway.
Redox Biol. 85:1036852025. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Song G, Wang J, Liu J and Ruan Y: Dimethyl
fumarate ameliorates erectile dysfunction in bilateral cavernous
nerve injury rats by inhibiting oxidative stress and NLRP3
inflammasome-mediated pyroptosis of nerve via activation of
Nrf2/HO-1 signaling pathway. Redox Biol. 68:1029382023. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu P, Lu Z, Liu L, Li R, Liang Z, Shen M,
Xu H, Ren D, Ji M, Yuan S, et al: NOD-like receptor signaling in
inflammation-associated cancers: From functions to targeted
therapies. Phytomedicine. 64:1529252019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wood JPM, Chidlow G, Wall GM and Casson
RJ: N-acetylcysteine amide and di-N-acetylcysteine amide protect
retinal cells in culture via an antioxidant action. Exp Eye Res.
248:1100742024. View Article : Google Scholar
|
|
41
|
Santus P, Signorello JC, Danzo F,
Lazzaroni G, Saad M and Radovanovic D: Anti-inflammatory and
anti-oxidant properties of N-acetylcysteine: A fresh perspective. J
Clin Med. 13:41272024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dollish HK, Tsyglakova M and McClung CA:
Circadian rhythms and mood disorders: Time to see the light.
Neuron. 112:25–40. 2024. View Article : Google Scholar
|
|
43
|
Zheng Z, Pan J, Chen Z, Gao P, Gao J,
Jiang H and Zhang X: The association between shift work, shift work
sleep disorders and premature ejaculation in male workers. BMC
Public Health. 24:17722024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Desmet L, Thijs T, Segers A, Verbeke K and
Depoortere I: Chronodisruption by chronic jetlag impacts metabolic
and gastrointestinal homeostasis in male mice. Acta Physiol (Oxf).
233:e137032021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Petzke F, Heppner C, Mbulamberi D,
Winkelmann W, Chrousos GP, Allolio B and Reincke M: Hypogonadism in
Rhodesian sleeping sickness: Evidence for acute and chronic
dysfunction of the hypothalamic-pituitary-gonadal axis. Fertil
Steril. 65:68–75. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Smimmo M, Casale V, Casillo GM, Mitidieri
E, d'Emmanuele di Villa Bianca R, Bello I, Schettino A, Montanaro
R, Brancaleone V, Indolfi C, et al: Hydrogen sulfide dysfunction in
metabolic syndrome-associated vascular complications involves cGMP
regulation through soluble guanylyl cyclase persulfidation. Biomed
Pharmacother. 174:1164662024. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Paeng SK, Wi SD, Chae HB, Bae SB, Phan
KAT, Kim MG, Yun DJ, Kim WY, McClung CR and Lee SY: NTRC mediates
the coupling of chloroplast redox rhythm with nuclear circadian
clock in plant cells. Mol Plant. 18:468–484. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang S, Zhang W, Ba M, Xuan S, Huang D,
Qi D, Pei X, Lu D and Li Z: Chronic jet lag disrupts circadian
rhythms and induces hyperproliferation in murine lacrimal glands
via ROS accumulation. Invest Ophthalmol Vis Sci. 66:122025.
|
|
49
|
Wang H, Guo M, Ren B, Zhang H, Zhang J,
Qiao R, Qian L, Zhu J, Zhang S, Su W, et al: Circadian control of
hepatic ischemia/reperfusion injury via HSD17B13-mediated autophagy
in hepatocytes. J Hepatol. 83:750–767. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang HY, Li KY, Wang YL, Wei CJ, Gao YX,
Ren-Zhou, Zhong YB, Yin ZJ and Ren DL: ROS regulates circadian
rhythms by modulating Ezh2 interactions with clock proteins. Redox
Biol. 81:1035262025. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhou R, Li K, Hu X, Fan S, Gao Y, Xue X,
Bu Y, Zhang H, Wang Y, Wei C, et al: Sleep deprivation activates a
conserved lactate-H3K18la-RORα axis driving neutrophilic
inflammation across species. Adv Sci (Weinh). 12:e040282025.
View Article : Google Scholar
|
|
52
|
Nishikawa S, Inoue Y, Hori Y, Miyajima C,
Morishita D, Ohoka N, Hida S, Makino T and Hayashi H:
Anti-inflammatory activity of kurarinone involves induction of HO-1
via the KEAP1/Nrf2 pathway. Antioxidants (Basel). 9:8422020.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Xian P, Hei Y, Wang R, Wang T, Yang J, Li
J, Di Z, Liu Z, Baskys A, Liu W, et al: Mesenchymal stem
cell-derived exosomes as a nanotherapeutic agent for amelioration
of inflammation-induced astrocyte alterations in mice.
Theranostics. 9:5956–5975. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang T, Wu P, Budbazar E, Zhu Q, Sun C,
Mo J, Peng J, Gospodarev V, Tang J, Shi H and Zhang JH: Mitophagy
reduces oxidative stress via Keap1 (kelch-like
epichlorohydrin-associated protein 1)/Nrf2 (nuclear
factor-E2-related factor 2)/PHB2 (prohibitin 2) pathway after
subarachnoid hemorrhage in rats. Stroke. 50:978–988. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kopacz A, Klóska D, Proniewski B, Cysewski
D, Personnic N, Piechota-Polańczyk A, Kaczara P, Zakrzewska A,
Forman HJ, Dulak J, et al: Keap1 controls protein S-nitrosation and
apoptosis-senescence switch in endothelial cells. Redox Biol.
28:1013042020. View Article : Google Scholar
|
|
56
|
Wu CT, Deng JS, Huang WC, Shieh PC, Chung
MI and Huang GJ: Salvianolic acid C against acetaminophen-induced
acute liver injury by attenuating inflammation, oxidative stress,
and apoptosis through inhibition of the Keap1/Nrf2/HO-1 signaling.
Oxid Med Cell Longev. 2019:90568452019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
O'Rourke SA, Shanley LC and Dunne A: The
Nrf2-HO-1 system and inflammaging. Front Immunol. 15:14570102024.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pekovic-Vaughan V, Gibbs J, Yoshitane H,
Yang N, Pathiranage D, Guo B, Sagami A, Taguchi K, Bechtold D,
Loudon A, et al: The circadian clock regulates rhythmic activation
of the NRF2/glutathione-mediated antioxidant defense pathway to
modulate pulmonary fibrosis. Genes Dev. 28:548–560. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Horton NS, Copple IM, Vasilaki A, Jackson
MJ, McArdle A and Pekovic-Vaughan V: P 089-NRF2/KEAP1-mediated
antioxidant defence pathway regulates skeletal muscle circadian
clock function. Free Radic Biol Med. 108:S482017. View Article : Google Scholar
|
|
60
|
Tamaru T, Hattori M, Ninomiya Y, Kawamura
G, Varès G, Honda K, Mishra DP, Wang B, Benjamin I, Sassone-Corsi
P, et al: ROS stress resets circadian clocks to coordinate
pro-survival signals. PLoS One. 8:e820062013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Jing H, Sun X, Li M, Peng J, Gu X and
Xiong J: Exogenous melatonin activating nuclear factor E2-related
factor 2 (Nrf2) pathway via melatonin receptor to reduce oxidative
stress and apoptosis in antler mesenchymal stem cells. Molecules.
27:25152022. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Guo S, Yang J, Qin J, Qazi IH, Pan B, Zang
S, Lv T, Deng S, Fang Y and Zhou G: Melatonin promotes in vitro
maturation of vitrified-warmed mouse germinal vesicle oocytes,
potentially by reducing oxidative stress through the Nrf2 pathway.
Animals (Basel). 11:23242021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang Y, Cong P, Tong C, Jin H, Liu Y and
Hou M: Melatonin pretreatment alleviates blast-induced oxidative
stress in the hypothalamic-pituitary-gonadal axis by activating the
Nrf2/HO-1 signaling pathway. Life Sci. 280:1197222021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhao X, Xie J, Duan C, Wang L, Si Y, Liu
S, Wang Q, Wu D, Wang Y, Yin W, et al: ADAR1 protects pulmonary
macrophages from sepsis-induced pyroptosis and lung injury through
miR-21/A20 signaling. Int J Biol Sci. 20:464–485. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tao W, Hu Y, Chen Z, Dai Y, Hu Y and Qi M:
Magnolol attenuates depressive-like behaviors by polarizing
microglia towards the M2 phenotype through the regulation of
Nrf2/HO-1/NLRP3 signaling pathway. Phytomedicine. 91:1536922021.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang C, Zhao M, Wang B, Su Z, Guo B, Qin
L, Zhang W and Zheng R: The Nrf2-NLRP3-caspase-1 axis mediates the
neuroprotective effects of Celastrol in Parkinson's disease. Redox
Biol. 47:1021342021. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Arioz BI, Tastan B, Tarakcioglu E, Tufekci
KU, Olcum M, Ersoy N, Bagriyanik A, Genc K and Genc S: Melatonin
attenuates LPS-induced acute depressive-like behaviors and
microglial NLRP3 inflammasome activation through the SIRT1/Nrf2
pathway. Front Immunol. 10:15112019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Song X, Fan S, Gao Y, Ma A, Zhang X, Zhou
Z, Zheng Y, Du L and Zhu X: Swietenolide inhibits the TXNIP/NLRP3
pathways via Nrf2 activation to ameliorate cognitive dysfunction in
diabetic mice. Neuropharmacology. 267:1103122025. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wang CY, Xu Y, Wang X, Guo C, Wang T and
Wang ZY: Dl-3-n-butylphthalide inhibits NLRP3 inflammasome and
mitigates Alzheimer's-like pathology via Nrf2-TXNIP-TrX axis.
Antioxid Redox Signal. 30:1411–1431. 2019. View Article : Google Scholar
|
|
70
|
Yi S, Zheng B, Zhu Y, Cai Y, Sun H and
Zhou J: Melatonin ameliorates excessive PINK1/Parkin-mediated
mitophagy by enhancing SIRT1 expression in granulosa cells of PCOS.
Am J Physiol Endocrinol Metab. 319:E91–E101. 2020. View Article : Google Scholar : PubMed/NCBI
|