Introduction
Regular exercise is essential for maintaining
systemic metabolic homeostasis and overall health, reducing disease
susceptibility, and improving long-term health outcomes (1-5).
Reflecting these wide-ranging and sustained benefits, the World
Health Organization has identified the promotion of exercise at the
population level as a global public health priority (6). Structured exercise and lifestyle
interventions yield measurable clinical benefits, particularly in
high-risk populations. Previous studies within the LIGHT trial
framework suggest that digital health-supported programs are
associated with reduced atherosclerotic cardiovascular disease risk
estimates and improved peak VO2, underscoring the
clinical significance of exercise-induced systemic adaptation
(7,8). However, the molecular mechanisms
underlying these adaptations remain inadequately defined, including
the circulating mediators that coordinate inter-organ signaling and
the downstream pathways that support durable metabolic
reprogramming (9,10). Defining these signaling
mechanisms is essential to mechanistically linking local tissue
responses to coordinated systemic adaptations and durable metabolic
benefits.
Exercise induces profound changes in the systemic
landscape, characterized by the influx of exercise-responsive
nucleic acids, proteins, and metabolites into the circulation
(11,12). These exercise-induced signals
provide a molecular basis for the coordinated multi-organ response
to exercise and are increasingly recognized as key drivers of the
transition from short-term physiological regulation to long-term
metabolic adaptation (13-15). Numerous exercise-induced signals
are intrinsically unstable and lack canonical secretion motifs,
making them unlikely to mediate long-range effects through passive
diffusion alone. Consequently, extracellular vesicles (EVs) have
emerged as important candidates for the transmission of
exercise-related signals (16-18). EVs are lipid bilayer-enclosed
vesicles released by nucleated cells that can carry donor
cell-derived bioactive cargo, including metabolites, proteins, and
RNA, to recipient cells, mediating targeted intercellular and
inter-organ communication (19-22). However, the terminology used in
exercise-EV studies remains variable, partly because vesicles are
classified according to different criteria, including biogenesis,
size, cellular origin, and isolation or characterization strategy
(23,24). Thus, labels such as exosomes,
small EVs, and microvesicles may describe overlapping but not
necessarily identical vesicle populations across studies, with
exosomes generally referring to vesicles of endosomal origin,
microvesicles to vesicles shed from the plasma membrane, and small
EVs to size-defined vesicle fractions (25). To avoid overclassification, the
present review uses EVs as the overarching term and retains
subtype-specific terminology when supported by the original
reports. Although accumulating evidence indicates that exercise
markedly alters EV abundance and molecular features, the precise
role of these changes in mediating exercise-related biological
effects remains poorly understood (26,27).
Among the potential mechanisms, epigenetic
regulation is particularly well-suited to explain the enduring
effects of exercise adaptation as it allows environmental cues to
drive stable yet reversible changes in gene expression (28-30). Epigenetic regulation encompasses
DNA methylation, histone post-translational modifications, and
non-coding RNA (ncRNA)-mediated regulatory processes, all of which
modulate chromatin state and gene expression without altering the
underlying DNA sequence itself (31). Crucially, EVs can deliver
epigenetically relevant cargo, particularly ncRNAs, to recipient
cells, thereby providing a means for epigenetic regulation across
tissues (32). Although
EV-mediated effects may extend beyond microRNAs (miRNAs or miRs) to
other classes of ncRNAs, including long non-coding RNAs and
circular RNAs, and downstream chromatin-related regulation, direct
evidence for these mechanisms in the exercise setting remains
limited. By contrast, the strongest and most consistent evidence to
date centers on EV-associated miRNAs, which therefore constitute
the primary mechanistic focus of the present review (33,34). Substantial evidence indicates
that EVs regulate diverse physiological and pathological processes,
including metabolic homeostasis and inflammatory responses, and
influence disease progression through the delivery of ncRNAs
(35). Exercise, in turn,
dynamically modulates EV release and cargo composition, thereby
reshaping EV-mediated epigenetic regulation (36-38).
EVs have attracted broad translational interest as
minimally invasive biomarkers and potential delivery-based
therapeutic platforms, owing to their stability in biofluids and
their capacity to carry protected molecular cargo (39-41). In the exercise field, changes in
EV abundance, surface-marker profiles, and miRNA cargo have been
associated with acute exercise responses, fatigue-related
physiological stress, metabolic health, and training responsiveness
(42-44). Therapeutic translation remains
less established, with current evidence largely derived from
preclinical studies suggesting that exercise-induced EVs or
exercise-regulated cargo can modulate inflammation, tissue repair,
cardiometabolic remodeling, and organ protection (45). However, the mechanisms governing
the selection, delivery, and tissue-specific coupling of
exercise-responsive EV cargo to epigenetic remodeling and
downstream metabolic adaptation remain poorly defined. A systematic
synthesis of current evidence is therefore needed to clarify how
EV-mediated epigenetic signaling links exercise to sustained,
multisystem metabolic benefits.
EVs: An intrinsic carrier for molecular
protection and directed delivery
In 1967, Peter Wolf described platelet-derived
'platelet dust' in plasma, marking one of the earliest reports of
circulating EVs (46). Initially
considered waste disposal by-products, these vesicles are now
recognized as functional mediators of intercellular communication
(11,47). To standardize terminology, the
International Society for Extracellular Vesicles proposed in 2018
that 'extracellular vesicles' should be used as the generic term
for particles naturally released from cells (48). EVs are nanoscale, lipid
bilayer-enclosed vesicles that transfer lipids, nucleic acids,
proteins, and other biomolecules to nearby or distant recipient
cells, thereby mediating intercellular communication (49). This EV-mediated communication
pathway is evolutionarily conserved across prokaryotes, plants, and
eukaryotes, including yeast and fungi (50). Based on their biogenesis, EVs are
broadly divided into exosomes and plasma membrane-derived vesicles
(51). Exosomes, typically
within the small EV size range (30-150 nm), originate from the
endocytic pathway as intraluminal vesicles within multivesicular
bodies (MVBs) and are released when MVBs fuse with the plasma
membrane. They are often enriched in CD9, CD63, CD81,
ALG-2-interacting protein X, heat shock protein (HSP)70, and tumor
susceptibility 101 (52,53). By contrast, plasma
membrane-derived vesicles bud directly from the cell surface and
include microvesicles (MVs, typically 501,000 nm) and apoptotic
vesicles (100-5,000 nm), the latter being generated during
apoptosis (54). After release,
EVs may enter circulating biofluids, distribute locally or
systemically, and reach distant recipient cells, enabling
communication across both proximal and remote sites (39). Some EVs rapidly degrade after
release, directly releasing cytokines or growth factors, whereas
most remain stable in biofluids and can travel to specific tissues
or distant target cells (11).
Upon reaching target cells, EVs interact through three primary
mechanisms: Receptor binding at the cell surface, direct membrane
fusion, or endocytic uptake (55). The uptake of EVs varies by cell
type and is largely governed by their surface protein composition,
especially integrins, tetraspanins, and lectins (51). Additionally, EV cargo loading is
increasingly recognized as a selective process, not a purely
passive one. It involves mechanisms such as endosomal sorting
complex required for transport (ESCRT)-dependent and
ESCRT-independent pathways, lipid microdomain-associated processes,
and RNA-binding protein-mediated loading of regulatory RNAs
(56,57). This selective packaging further
contributes to the molecular composition and functional specificity
of EVs, influencing which regulatory signals are delivered to
recipient cells (58,59).
Taken together, EVs should not be viewed as passive
released cellular debris, but as active biological carriers that
protect bioactive molecules from degradation and enable their
selective delivery to recipient cells, thereby supporting
intercellular and inter-organ communication (Fig. 1).
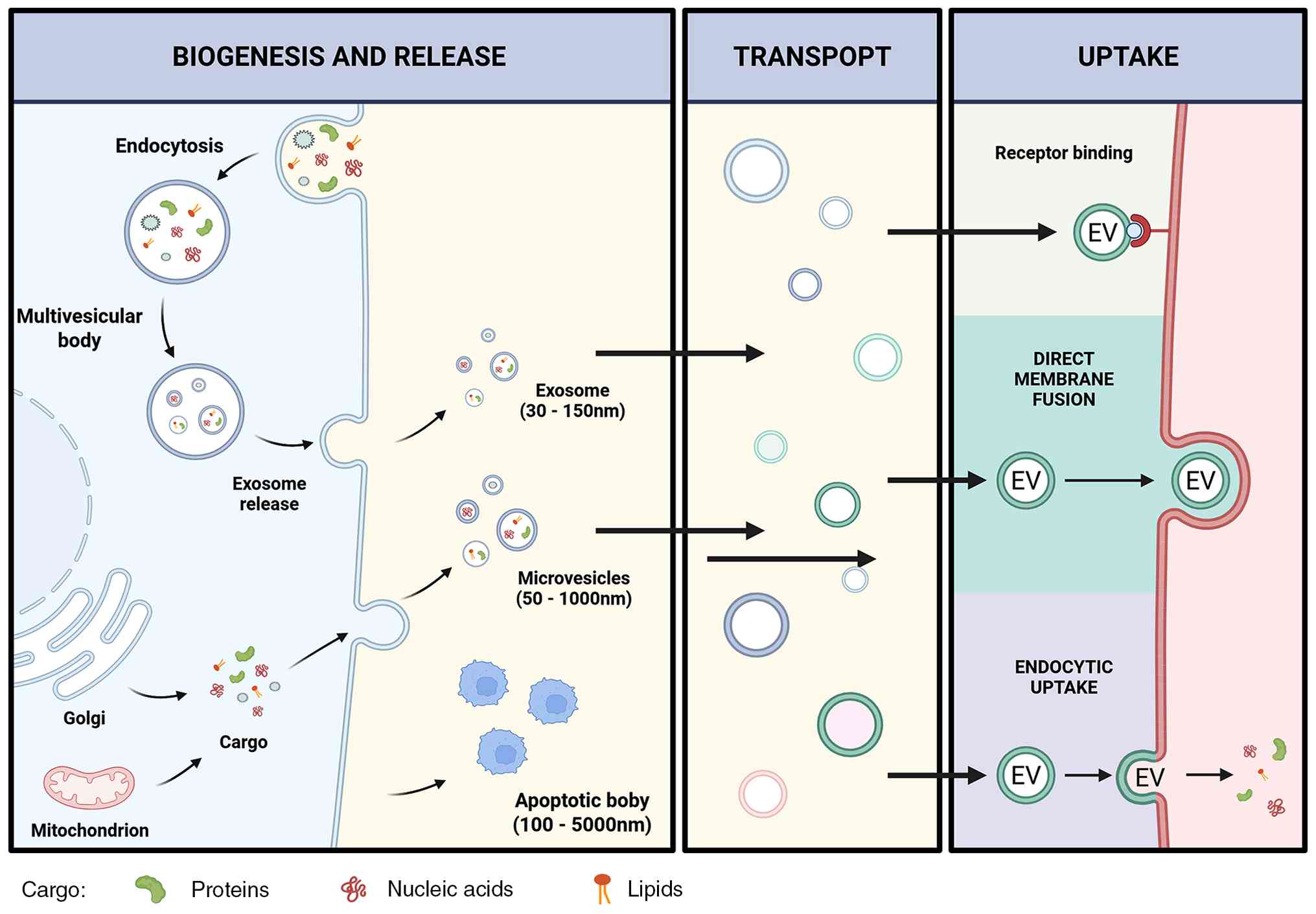 | Figure 1Biogenesis, release, transport, and
uptake of EVs. EVs are generated through distinct biogenetic routes
and carry bioactive cargo, including proteins, nucleic acids, and
lipids. Exosomes are formed through the endosomal pathway:
Endocytosis generates early endosomal compartments, which mature
into MVBs containing intraluminal vesicles; fusion of MVBs with the
plasma membrane releases these vesicles as exosomes, typically
within the small EV size range. Microvesicles are produced by
direct outward budding of the plasma membrane, whereas apoptotic
bodies are released during apoptosis. After release, EVs can travel
through extracellular fluids or the circulation and deliver donor
cell-derived signals to recipient cells. EV uptake may occur
through receptor binding at the cell surface, direct membrane
fusion, or endocytic internalization, thereby enabling transfer of
molecular cargo and contributing to intercellular and inter-organ
communication. EVs, extracellular vesicles; MVBs, multivesicular
bodies. |
Cellular origins of exercise-induced
EVs
As a systemic physiological stimulus, exercise
reshapes circulating EV profiles by altering both EV release and
cargo composition (37,60). While skeletal muscle is widely
recognized as a major source of exercise-induced EVs, current
evidence suggests that circulating EVs also originate from a
broader range of cell types, including endothelial cells,
platelets, and leukocytes, in response to both acute or chronic
exercise (61-64). Notably, despite the marked
plasticity of circulating EV signals, EVs from distinct tissue
origins often retain relatively stable source-associated features
(65) (Table I). These origin-dependent
characteristics largely determine their surface molecular identity,
cargo profile, tissue tropism, and cellular uptake efficiency,
thereby influencing downstream signaling, transcriptional programs,
and metabolic reprogramming in recipient cells, and ultimately
shaping their functional effects in target organs (66). Accordingly, a systematic
understanding of how exercise regulates EVs derived from different
cellular sources is essential not only for elucidating the systemic
adaptations to physical activity, but also for defining the
physiological functions of EVs and their potential diagnostic and
therapeutic value.
 | Table IMajor cellular sources and defining
features of exercise-related EVs. |
Table I
Major cellular sources and defining
features of exercise-related EVs.
| EV origin | Specific cargo | Exercise
response | Major biological
function | Surface
markers |
|---|
| Skeletal
muscle | miR-1, miR-133a/b,
miR-206, miR-486, miR-499 | Dynamically
remodeled by exercise mode, intensity, duration, and training
status | Mediate local
myofiber communication and long-range inter-organ signaling;
contribute to myogenesis, regeneration, metabolic adaptation,
insulin sensitivity, and systemic homeostasis | SGCA, CD81 |
| Platelets | Platelet
activation-related proteins, membrane lipids, procoagulant and
signaling molecules | Rapidly increased
after acute high-intensity exercise; transient and
intensity-dependent | Reflect platelet
activation, acute physiological stress, and vascular-hemostatic
responses; may participate in coagulation, inflammation, and
post-exercise vascular adaptation | CD41, CD42,
CD31 |
| Endothelial
cells | Endothelial
activation- or injury-related proteins, adhesion molecules, and
regulatory nucleic acids | Variable responses
depending on exercise protocol, fitness level, and study
method | Reflect endothelial
activation, vascular stress, and vascular fitness; may serve as
indicators of endothelial adaptation to exercise | CD144, CD62E,
CD105, CD146 |
| Regulatory T
cells | miR-150-5p,
miR-146a-5p, miR-21-5p | Altered with
exercise-induced immune activation | Involved in immune
cell communication, inflammatory regulation, and
exercise-associated immune adaptation | CD45, CD11b, CD14,
CD16 |
Skeletal muscle-derived EVs
Skeletal muscle, comprising 35-40% of body mass,
functions as both a primary motor effector and a critical endocrine
organ. Beyond myokine secretion, emerging evidence indicates that
skeletal muscle also releases bioactive EVs, mediating complex
intercellular and inter-organ communication (67). Guescini et al (68) were the first to show that
myoblasts release exosome-like vesicles (50-80 nm) enriched in
signaling-related proteins and mitochondrial DNA, suggesting their
role in transferring genetic and metabolic information between
muscle cells and potentially across organs. This view was further
supported by the detection of fluorescently labeled EVs in distant,
including contralateral, muscles after intramuscular injection
(69). In vitro and
animal studies further indicate that multiple skeletal muscle cell
types, including myoblasts, myotubes, and satellite cells, can
release exosomes enriched in muscle-specific miRNAs (myomiRs) such
as miR-1, miR-133a/b, miR-206, and miR-486 (70,71). These myomiRs, which are highly
enriched in muscle tissue, play important roles in myogenesis,
differentiation, regeneration, and stress responses (61,72).
Exercise can markedly alter both the abundance and
molecular cargo of circulating exosomes, in a manner that depends
on exercise modality, intensity, and host-related factors (61). Different forms of exercise,
including uphill running, downhill running, endurance training, and
resistance training, induce distinct changes in exosomal miRNA
expression patterns, with dynamic fluctuations in molecules such as
miR-1, miR-133a, miR-499, and miR-486 (73). Among these, miR-486 can target
PTEN and thereby enhance insulin-like growth factor-1/protein
kinase B/mechanistic target of rapamycin (IGF-1/AKT/mTOR)
signaling, suggesting that exercise-induced exosomal miRNAs may
contribute to protein synthesis, glucose homeostasis, and improved
insulin sensitivity (74). On
the basis of these findings, skeletal muscle is now considered a
major source of circulating exosomes.
Functionally, muscle-derived EVs help maintain local
tissue homeostasis by promoting myoblast differentiation,
supporting neuromuscular junction integrity, and protecting motor
neurons (75). They can also
reach distant metabolic organs, where they influence signaling
pathways involved in insulin responsiveness, glucose handling,
lipid metabolism, and systemic metabolic homeostasis (76-79). Altered muscle exosomal cargo has
been linked to both pathological and adaptive responses in
neuromuscular and metabolic diseases, highlighting the potential of
muscle-derived EVs as biomarkers and therapeutic targets (80-84).
Platelet-derived EVs
A substantial proportion of human studies
investigating the effects of acute exercise on circulating EVs have
focused on larger vesicle populations, particularly MVs exceeding
500 nm in diameter (27). Among
these, platelet-derived microvesicles (PMVs) represent the most
abundant MV subset in circulation and have attracted significant
attention due to their robust responsiveness to exercise (85-87). Across a range of high-intensity
endurance modalities, including maximal incremental exercise
testing, heavy-intensity cycling, sustained aerobic exercise near
70-85% of maximal capacity, and high-intensity interval protocols,
PMVs consistently exhibit marked increases during or shortly after
exercise (88-91). Notably, these changes appear to
be intensity dependent, as moderate-intensity exercise does not
elicit comparable elevations (92). Moreover, the PMV surge is
transient, with concentrations peaking during exercise or early
recovery and returning toward baseline within 30-120 min (85,86,92). Together, these findings highlight
PMVs as dynamically regulated vesicles that may reflect acute
physiological stress and vascular activation during strenuous
exercise.
Endothelial cell-derived EVs
Compared with PMVs, the exercise-induced behavior of
endothelial-derived microvesicles (EMVs) is less consistent. While
several studies have demonstrated post-exercise elevations in EMVs
(91,93), others have reported no change or
even reductions, both in healthy and metabolically compromised
individuals (85,89,94). These discrepancies likely reflect
methodological variability in EV isolation and characterization, as
well as differences in exercise intensity, modality, and
participant characteristics. Notably, emerging evidence suggests
that training status may critically shape EMV dynamics. In highly
trained endurance athletes, distinct heavy-intensity exercise
formats, including prolonged submaximal workloads, repeated
high-intensity intervals, and maximal sprinting, were uniformly
associated with reductions in circulating EMVs, potentially
reflecting enhanced vascular function and accelerated vesicle
clearance in trained individuals (95,96). These observations underscore that
EMV responses are not uniform and may integrate information about
both exercise stimulus and vascular fitness.
Regulatory T cell-derived EVs
(Treg-EV)
Treg-EVs primarily carry a range of
immunosuppressive molecules and miRNAs that contribute to the
maintenance of immune tolerance and the suppression of inflammatory
responses (74). They are
enriched in molecules such as CD39, CD73, cytotoxic
T-lymphocyte-associated protein 4, IL-35 subunits, IL-10, and
TGF-β, as well as regulatory miRNAs including miR-150-5p,
miR-146a-5p, and miR-21-5p (76,97-99). Among these, miR-146a-5p has been
shown to suppress CD4+ T-cell proliferation, further
supporting the immunoregulatory function of Treg-EVs (100). During exercise, Treg-EVs may
exert immunomodulatory effects through these molecular cargos,
attenuating inflammatory responses and fostering an
anti-inflammatory milieu, particularly in chronic metabolic
disorders such as diabetes and obesity, as well as during recovery
from exercise-induced muscle injury (100). Notably, Treg cells may act in
concert with skeletal muscle-derived EVs to coordinate
post-exercise immune adaptation and tissue repair (62,101). For example, muscle-derived EVs
may enhance Treg activation and immunosuppressive function through
miRNAs such as miR-206, amplifying anti-inflammatory signals via
EV-mediated communication (102). Simultaneously, Treg-EVs may
suppress excessive immune-cell responses and reinforce immune
tolerance through the delivery of miR-146a-5p (103). In this cooperative framework,
EVs, through their carried cargo, may serve dual roles in
regulating immune responses and promoting both metabolic health and
tissue repair.
Exercise-driven changes in EV composition
and function
Exercise, including acute and chronic exercise,
refers to voluntary movements produced by skeletal muscles that
elevate energy expenditure (104,105). In response to the increased
physiological demands of exercise, the body engages coordinated
adaptations across metabolic, cardiovascular, and immune systems,
collectively underpinning the broad and sustained health benefits
of regular physical activity (106-109). In 2015, Frühbeis et al
(110) were the first to
systematically document the rapid changes in circulating EVs
following acute exercise, identifying EVs as potential mediators of
exercise-induced inter-organ communication. By transporting
proteins, lipids, and regulatory RNAs through the circulation, EVs
can relay local signals generated by contracting skeletal muscle to
distant tissues, including the vasculature, liver, adipose tissue,
and immune system, thereby linking muscle activity to systemic
adaptation (111-113). Recent studies indicate that EV
responses vary substantially across different exercise modalities.
Acute vs. chronic exercise, along with variations in intensity and
duration, each reshape EV abundance, cellular origin, and cargo
composition in distinct ways (114-116). For example, short bouts of
cycling are associated with increased exosomal miR-1, whereas
longer-duration exercise induces a broader myomiR response,
including miR-1, miR-133a/b, miR-206, miR-208a, and miR-499
(73,117,118). The present review synthesizes
recent evidence on the dynamic behavior of EVs across different
forms of exercise, particularly endurance and resistance training,
offering a clearer framework for understanding the mechanisms of
exercise adaptation (Table
II).
| Table IIExercise-induced alterations in
circulating EVs. |
Table II
Exercise-induced alterations in
circulating EVs.
| Exercise
protocol | Bio-fluid | Time of
collection | Isolation
method | Verification
method | Terminology | Exercise
effect | Year | (Refs.) |
|---|
| Acute exercise | Incremental cycling
or treadmill running to exhaustion in healthy adults | Plasma | Pre-exercise,
immediately post-exercise, and 90 min post-exercise | Differential
centrifugation, including 10,000 × g pre-clearing, filtration, and
100,000 × g ultracentrifugation | NTA and WB | Small
EVs/exosome-like vesicles | Small EVs increased
immediately after cycling and declined by 90 min; the running
response was more moderate but appeared more sustained | 2015 | (110) |
| Incremental cycling
test to exhaustion in healthy male athletes | EDTA plasma | Pre-exercise,
during exercise (RQ=0.9), and immediately post-exercise | EV Array on total
plasma; SEC; CD9/CD63/CD81 immunobead isolation | EV Array, NTA, WB,
and multiplex flow cytometry | EVs | EV levels increased
progressively during exercise and peaked at maximal load; ExerVs
showed mixed leukocyte-, platelet-, and endothelial-associated
features | 2019 | (132) |
| Single 40-min
treadmill bout (low-, moderate-, or high-intensity) in rats | Serum | Immediately after
the exercise bout | ExoQuick-based
precipitation from platelet-depleted serum | TEM/immun o-EM,
TRPS, and WB | EVs | EV concentration
increased, whereas size was not significantly changed;
CD63-positive vesicles increased and small-RNA cargo was
altered | 2018 | (131) |
| Moderate-intensity
treadmill exercise (60% VO2max) in obese and
normal-weight adults | Plasma | Pre-exercise,
immediately post-exercise, 3 h, and 24 h | Plasma EV
isolation/purification (supplementary protocol), followed by NTA
and flow-based phenotyping | NTA and flow
cytometry | Total EVs,
exosomes, and microvesicles | Total circulating
EVs decreased overall (mainly microvesicle-range EVs);
CD61+ EVs decreased,whereas SGCA+ EVs
increased; responses were sex- and BMI-dependent | 2020 | (133) |
| 60-min cycling in
healthy men (30 min at 55%, 20 min at 70%, then ~10 min at 80%
VO2max to exhaustion), with 4-h recovery | Plasma | Baseline, end of
exercise, and 4 h recovery | Rapid plasma
preparation followed by high-speed centrifugation to obtain a
20,000 × g EV fraction for proteomics | NTA, cryo-EM, and
nano-UHPLC-MS/MS proteomics | EVs, exosomes, and
small vesicles | Exercise increased
systemic release of EV-associated proteins and enriched
exosome/small-vesicle signatures; exercise-liberate | 2018 | (26) |
| Clinical exercise
stress testing in human subjects | EDTA plasma | At rest, peak
exercise, and 15 min after exercise | Plasma preparation
by sequential centrifugation; direct particle analysis by nano-flow
cytometry | Nano-flow
cytometry | EVs/plasma EVs | Acute exercise
induced a rapid increase in circulating plasma EVs | 2017 | (44) |
| Chronic
exercise | 3-week swimming in
mice | Serum | After completion of
the training period | Plasma EV isolation
followed by size and phenotyping analyses | Nano-flow
cytometry | EVs/serum EVs | Serum EVs were
increased (~1.85-fold) after training | 2017 | (44) |
| Daily moderate
treadmill exercise (20 min/day for 2 weeks) in rats | Serum | 1 h and 18 h after
the last exercise session | Precipitation-based
isolation (miRCURY Exosome Isolation Kit) | Marker assays and
biochemical profiling | EVs/exosomes | Exercise produced
delayed changes in circulating EV profile, with alterations more
evident 18 h after the last session | 2018 | (146) |
| 4-week swim
training in rats/trained human volunteers | Plasma | 24 h after the last
training session | Differential
ultracentrifugation; ExoQuick also used | Exosome
characterizati on plus functional assays | Circulating
exosomes/plasma exosomes | Total circulating
exosome level did not change significantly 24 h post-exercise, but
cargo composition/function was altered | 2019 | (45) |
| 4-week treadmill
training (low vs. moderate intensity; 60 min/day, 5 days/week) in
mice | Plasma | 24 h after the last
bout of exercise | Microbead-based
sorting of circulating EPC-derived exosomes | NTA, WB qPCR, |
Exosomes/EPC-derived exosomes | Exercise increased
circulating EPC-derived exosome release in an intensity-dependent
manner | 2018 | (150) |
Acute exercise-induced changes in
EVs
Acute exercise refers to a single bout of
short-duration physical activity, typically lasting from several
minutes to a few hours (119,120). Unlike the chronic adaptations
associated with long-term training, acute exercise primarily
represents a rapid physiological response to transient physical
stress that can quickly reconfigure multisystem homeostasis and
confer immediate health benefits (121-123). Even a single acute exercise
bout can acutely improve systemic metabolic status, enhance insulin
sensitivity, and promote energy utilization in major metabolic
organs such as skeletal muscle and the liver (124-126). In addition, acute exercise
exerts both anti-inflammatory and immune-activating effects,
providing a brief but efficient form of endogenous protection
through improved endothelial function and redistribution of immune
cell populations (127,128).
A substantial body of human evidence consistently
shows that acute exercise is a potent stimulus for the transient
elevation of circulating EVs (129). Early exploratory research used
graded cycling or treadmill tests to impose acute physiological
stress in healthy, physically active men and assessed plasma small
EV kinetics after isolation by differential centrifugation and
ultracentrifugation (100,000 × g). This research showed that acute
exercise rapidly increases circulating small EV levels: In cycling
protocols, particle concentrations rose by approximately 2- to
3-fold and exosomal marker proteins by ~5-fold, peaking immediately
after exercise and returning to baseline within 90 min of recovery
(110). Notably, EV release
occurred before the anaerobic threshold and exhibited rapid
release-and-clearance kinetics, supporting a role for EVs as
immediate physiological signaling mediators rather than passive
by-products of tissue damage (110).
A parallel study of healthy men highlights a
distinct kinetic divergence between exercise paradigms: While
cycling triggered a sharp, transient peak in EV abundance, an
incremental treadmill test resulted in a more moderate (~1.5-fold)
elevation in circulating small EVs, which remained elevated for 90
min and required 6 h to return to baseline (130). Animal models further
corroborate these dynamics. In Wistar rats, acute treadmill running
significantly enhanced serum EV concentrations and protein content
without a concomitant surge in total small-RNA yield (131). These findings suggest that the
exercise-induced signal resides not in the sheer abundance of RNA,
but in the selective remodeling of the molecular cargo. Deep
sequencing has identified 12 differentially expressed miRNAs
following exercise, whose targets predominantly converge on the
MAPK signaling pathway (131).
These data indicate that acute aerobic exercise rapidly reshapes
both the physical abundance and the molecular signature of
circulating EVs, positioning them as precise mediators of immediate
inter-organ signaling.
The cellular origins of acute exercise-induced EVs
are highly context-dependent. In healthy men performing a fasting,
graded cycling-to-exhaustion test, EV-associated signals
progressively increased during exercise peaking at exhaustion.
Under these acute exhaustive conditions, exercise-induced EVs
predominantly originated from lymphocytes, monocytes, endothelial
cells, and platelets, with no significant contribution from
skeletal muscle-derived EVs (132). By contrast, research using
moderate-intensity exercise revealed a different pattern. In
sex-stratified cohorts of both obese and normal-weight individuals,
acute treadmill exercise (~60% VO2max) rapidly reshaped
the circulating EV landscape. Skeletal muscle-derived α-sarcoglycan
(SGCA)+ EVs increased immediately after exercise,
whereas platelet-derived CD61+ EVs declined during
recovery, particularly at 24 h (133). These responses also varied by
sex and metabolic status: Women showed higher MV but lower exosome
levels, and normal-weight individuals exhibited a stronger MV
response than obese participants (133). Consistent with this, additional
research has demonstrated marked sex differences in
exercise-induced microparticle responses, with men showing
post-exercise increases in CD62E+ endothelial
microparticles, while women showed a more pronounced rise in
CD34+ microparticles (91). Together, these findings indicate
that the source composition of exercise-induced EVs is shaped not
only by exercise intensity and modality, but also by sex and
metabolic state.
Beyond changes in abundance and cellular origin,
acute exercise also reshapes EV function. In healthy men undergoing
incremental cycling, Whitham et al (26) showed that acute exercise induces
a marked but transient remodeling of the circulating EV proteome,
with 322 proteins significantly altered immediately after exercise
and most changes largely resolving within 4 h. Upregulated proteins
were enriched in pathways related to vesicle biogenesis, membrane
trafficking, adhesion, cytoskeletal remodeling, and signal
transduction, highlighting a rapid activation of EV-associated
signaling machinery (26).
Notably, exercise-derived EVs showed greater hepatic accumulation
than resting EVs and were capable of transferring protein cargo to
hepatocytes, supporting their role as active mediators linking
contracting tissues to metabolic organs rather than passive
by-products of exercise (26).
The functional relevance of exercise-induced EVs extends to
vascular biology. In healthy men, vigorous acute exercise rapidly
increased circulating endothelial-derived microparticles, and
phenotypic analysis indicated that these vesicles primarily
reflected endothelial activation rather than overt endothelial
apoptosis (94). This suggests
that acute exercise-induced EV release is more closely associated
with adaptive endothelial stress signaling than with widespread
vascular injury. Accordingly, circulating EVs may serve not only as
sensitive indicators of exercise stress, but also as early
effectors of vascular repair, endothelial regulation, and
exercise-induced cardiovascular protection.
Although still limited, current in vivo
evidence strongly supports a functional role for EVs in mediating
the cross-organ protective effects of acute physical activity.
Acute exercise rapidly increases circulating EVs, many of which are
predominantly derived from activated platelets and endothelial
cells, while simultaneously remodeling their cargo, including
proteins, miRNAs, and metabolites. By carrying these
exercise-responsive signals through the circulation, EVs may
coordinate multiorgan metabolic adaptation and functional
integration through epigenetic regulation and the modulation of key
signaling pathways.
Chronic exercise-induced changes in
EVs
Chronic exercise refers to regular physical activity
performed over an extended period (134,135). Unlike the transient
physiological stress triggered by a single bout of acute exercise,
chronic exercise is characterized by repeated exercise stimuli that
drive durable functional adaptation and structural remodeling,
resulting in broad, long-lasting, and systemic health benefits
(136-138). Long-term exercise training
substantially lowers the risk of multiple non-communicable
diseases, including metabolic disorders (such as obesity and type 2
diabetes), cardiovascular disease, cancer, and neurodegenerative
conditions (134,136,139,140). Through long-term physiological
regulation, chronic exercise reshapes systemic metabolic
homeostasis, supports more efficient energy utilization and insulin
sensitivity, delays biological aging, and ultimately extends
healthspan (141-143). In addition, by improving the
overall immune microenvironment, chronic exercise enhances
long-term host defense against pathogen exposure (144,145).
At the molecular level, in contrast to the rapid
post-exercise rise in circulating small EVs observed after acute
exercise, the effects of chronic exercise training on circulating
EVs appear to be more consistent with a delayed, homeostatic form
of remodeling (27). In a study
of Wistar rats across different age groups, Bertoldi et al
(146) applied a 2-week
moderate-intensity treadmill training protocol (20 min/day, ~60%
VO2max) and collected blood at 1 h and 18 h after the
final session. Training-related effects were observed mainly at the
18-h time point, suggesting that regular aerobic exercise may
promote exosome release or increase circulating exosome-associated
components in a delayed manner (146). In aged animals, elevated
EV-associated acetylcholinesterase activity was reduced after
training, indicating that chronic exercise may also partially
correct aging-associated EV abnormalities. Notably, this study also
identified time-of-day differences in CD63 measurements (afternoon
vs. the following morning), underscoring the importance of strict
control of sampling time and circadian factors in studies of
chronic exercise and EV biology (146). Overall, these findings suggest
that chronic exercise contributes to sustained systemic protection,
at least in part, by remodeling the quantitative profile of
circulating EVs.
Further evidence for the organ-protective effects of
exercise-derived EVs comes from chronic training models. In a
3-week swimming-trained mouse model, Bei et al (44) showed that chronic endurance
exercise increased circulating EV abundance and, more importantly,
that the transfer of EVs from exercised mice reduced ischemic
myocardial injury, suppressed cardiomyocyte apoptosis, and improved
cardiac function in recipient animals (44). Similar protective effects were
observed in vitro, where exercise-derived EVs enhanced
cardiomyocyte survival and attenuated apoptotic signaling. These
actions were closely linked to activation of ERK1/2-HSP27
signaling, supporting the view that exercise-induced EVs are not
merely by-products of training, but active effectors of remote
organ protection and post-stress tissue recovery. When sampling
time is carefully controlled, however, the effects of chronic
training on circulating exosome levels appear more nuanced. In
trained young male rowers and swim-trained rats, plasma exosome
size and concentration were unchanged 24 h after the final exercise
bout, yet exosomes from trained subjects displayed enhanced
cardioprotective activity (137). Specifically, these exosomes
were more readily taken up by cardiomyocytes, reduced
hypoxia/reoxygenation injury in vitro, and attenuated
ischemia-reperfusion damage in vivo. This functional
remodeling was linked to enrichment of miR-342-5p, which inhibits
caspase-9/c-Jun N-terminal kinase 2(Caspase9/JNK2)-dependent
apoptosis and enhances AKT-mediated survival signaling (137). More broadly, chronic exercise
appears to promote long-term adaptation and disease protection by
selectively remodeling the cargo of EVs from different tissues. In
db/db mice, 8 weeks of treadmill training increased exosome
release and elevated miR-455 and miR-29b, thereby attenuating
diabetic myocardial remodeling through suppression of matrix
metalloproteinase 9 (MMP9) (147). A recent human study further
extends these observations to metabolic disease settings. In
individuals with insulin resistance or type 2 diabetes, 12 weeks of
high-intensity interval training followed by detraining was
associated with partially sustained improvements in energy
metabolism and hepatic insulin sensitivity, raising the possibility
that small EV remodeling contributes to the persistence of some
exercise-induced metabolic benefits (148). Similarly, a study in sedentary
adults with obesity showed that endurance training significantly
remodeled the circulating EV proteome and reduced systemic
inflammation, even without marked changes in EV abundance (149). Together, these findings suggest
that, under metabolically compromised conditions, exercise may
reprogram EV function not only by altering vesicle levels, but also
by reshaping EV cargo and signaling properties (149). Chronic exercise has also been
shown to enhance vascular repair through endothelial progenitor
cell (EPC)-derived EVs: 4 weeks of treadmill training increased
circulating EPCs, boosted EPC exosome release and miR-126 content,
and promoted adaptive vascular remodeling via the sprouty-related
EVH1 domain-containing 1/vasular endothelial growth factor axis
(150). In addition to RNA
cargo, chronic exercise may also remodel EV-associated proteins, as
suggested by increased skeletal muscle HSP60 after endurance
training, raising the possibility that EV-associated protein
signals contribute to systemic adaptation (151).
Taken together, current evidence supports a central
role for EVs in mediating the long-term benefits of chronic
exercise. Rather than simply increasing circulating EV levels,
chronic exercise appears to induce a more stable functional
reprogramming of EV cargo, enabling sustained miRNA- and
protein-mediated signaling that may reinforce epigenetic
remodeling, metabolic adaptation, and systemic homeostasis.
Exercise-induced EVs as mediators of
epigenetic remodeling
Exercise-induced changes in EVs are likely to be
functionally important because they may reshape the regulatory
information delivered to recipient cells (152). By altering EV abundance,
subtype distribution, and cargo composition in a context-dependent
manner, exercise can modify the intercellular transfer of miRNAs,
proteins, metabolites, and other bioactive molecules (153). Once delivered to recipient
cells, these cargos can influence signaling networks and gene
programs involved in metabolism, inflammation, stress adaptation,
and tissue repair (154-156).
Among them, ncRNAs are especially notable, as they provide a
plausible route through which exercise-responsive EVs may
participate in epigenetic regulation across tissues (157). EV remodeling may therefore
represent a mechanistic link between transient exercise stimuli and
longer-term molecular adaptation (158). This perspective provides an
important framework for considering how exercise-induced EV
signaling intersects with epigenetic remodeling.
Exercise-induced epigenetic
remodeling
Epigenetics refers to the dynamic regulation of gene
expression through a series of plastic and reversible mechanisms
that occur without altering the underlying DNA sequence (159-161). First introduced by Waddington
in 1942, the concept describes how organisms establish structurally
dynamic regulatory states within chromatin to precisely control
transcriptional programs and adapt to environmental cues (162-164). Broadly, epigenetic regulation
encompasses DNA methylation, histone post-translational
modifications, chromatin remodeling, and regulation mediated by
ncRNAs (165-167). These mechanisms allow external
stimuli to be translated into changes in gene expression, thereby
influencing metabolic homeostasis, inflammatory responses, stress
adaptation, tissue repair, and disease progression (168-170). Notably, exercise can induce
epigenetic reprogramming through changes in energy metabolism,
redox status, and fluctuations in key metabolites such as
acetyl-CoA, NAD+, and lactate (171-173). These effects include modulation
of DNA methylation at metabolism-related genes, shifts in histone
acetylation and methylation, and remodeling of miRNA expression
profiles, ultimately influencing mitochondrial biogenesis, fatty
acid oxidation, glucose metabolism, insulin sensitivity, and
inflammatory-oxidative stress responses (174-178). Because these epigenetic
responses are dynamic, context-dependent, and partially persistent
after training, they provide a plausible mechanistic basis for the
sustained metabolic and systemic benefits of exercise (5,179-181).
Within this framework, ncRNAs, especially miRNAs,
have emerged as key regulatory effectors (182-184). Exercise markedly alters miRNA
profiles in both tissues and in the circulation, with these changes
being increasingly linked to pathways governing metabolism,
inflammation, angiogenesis, mitochondrial function, and tissue
repair (185-188). Notably, a subset of miRNAs can
be selectively packaged into EVs and transferred between tissues
(37,189,190). This suggests that
exercise-induced EV-associated miRNAs may serve as mobile
epigenetic signals that connect local muscle activity to systemic
adaptive remodeling (38,191,192).
Exercise-derived EVs as epigenetic
messengers
Given the central roles of EVs and epigenetic
regulation in exercise adaptation and metabolic health, increasing
attention has recently focused on whether exercise-induced EV
remodeling may function as a cross-tissue information carrier,
mediating ncRNA-driven epigenetic regulation and thereby
contributing to systemic metabolic benefits (193). Although this field remains in
its early stages, available evidence increasingly suggests that
physical activity can alter the abundance, subtype composition, and
cargo profile of circulating EVs, particularly their ncRNA content,
including miRNAs, and may thereby influence transcriptional
programs, signaling states, and long-term adaptive remodeling in
target tissues (61). To
systematically evaluate the evidence on exercise-induced EV
remodeling and its potential epigenetic relevance, a PubMed search
was conducted using the following terms:
('exercise'[Title/Abstract] OR 'physical activity'[Title/Abstract])
AND ('extracellular vesicles'[Title/Abstract] OR
'exosomes'[Title/Abstract] OR 'EVs'[Title/Abstract]). After merging
results, removing duplicates, and applying stringent screening
criteria, studies were included in which exercise was the primary
intervention and EVs were evaluated as mediators, biomarkers, or
functional effectors, with particular emphasis on studies
explicitly examining EV cargo remodeling and its roles in
epigenetic regulation, inter-organ communication, or downstream
transcriptional/metabolic reprogramming (Table III).
| Table IIIEpigenetic effects associated with
exercise-induced EVs. |
Table III
Epigenetic effects associated with
exercise-induced EVs.
| Exercise
protocol | EV
source/sample | Epigenetic change
(cargo remodeling) | Epigenetic
effects | Year | (Refs.) |
|---|
| Single
flywheel-based iso-inertial resistance session in trained men | Circulating EVs
from blood | Acute exercise
increased EV-encapsulated miR-206 and miR-146a | Suggests rapid
EV-mediated transfer of regulatory miRNAs during early
post-exercise adaptation, potentially coupling inflammation to
muscle remodeling | 2019 | (194) |
| Acute
exercise/fitness-related comparison in young men | Plasma EVs,
including SGCA+ muscle-enriched EVs | Muscle-derived EVs
were enriched in miR-206; EV miR-133b and miR-181a-5p increased
after acute exercise | Supports skeletal
muscle as a source of circulating EV-miRNAs that may contribute to
muscle remodeling, homeostasis, and inter-organ communication | 2015 | (68) |
| Two consecutive
bouts of muscle-damaging exercise (plyometrics + downhill
running) | Plasma EVs | No major change in
EV number or canonical myomiRs; EV miR-31 decreased at 24 h | Indicates selective
EV cargo repackaging after muscle-damaging exercise, with miR-31
emerging as a potential regulator of early recovery/remodeling | 2018 | (195) |
| Acute uphill vs.
downhill running in rats | Plasma, exosomal,
and exosome-free fractions | After downhill
exercise, exosomal miR-1, miR-133a, miR-133b, miR-206, miR-208a,
miR-499 increased during early recovery; uphill exercise produced
little exosomal change | Suggests exercise
mode-specific EV-miRNA remodeling, with eccentric loading
preferentially engaging exosomal miRNA signaling during
recovery | 2019 | (73) |
| Acute 40-min
cycling in sedentary vs. endurance-trained older men; comparison of
chronic training status | Plasma
exosomes | Baseline exomiR
profiles differed by training status; acute exercise-induced
distinct exomiR signatures in trained vs. sedentary men | Suggests regular
training reshapes EV-mediated epigenetic signaling and may
counteract age-related anabolic resistance | 2020 | (196) |
| Long-term swim
training in rats; trained vs. untrained human volunteers | Circulating plasma
exosomes | Long-term exercise
enriched exosomal miR-342-5p without changing total exosome
abundance at 24 h | Exercise-derived
exosomes conveyed sustained cardioprotection, reduced cardiomyocyte
apoptosis, and protected against myocardial ischemia/reperfusion
injury | 2019 | (45) |
| Treadmill exercise
in db/db diabetic mice | Serum and
heart-associated exosomes ('cardiosomes') | Exercise enhanced
exosomal release and increased miR-29b and miR-455 | Reduced MMP9
activity, attenuated extracellular matrix remodeling, and
potentially limited fibrosis and diabetic cardiac injury | 2015 | (147) |
The earliest evidence for exercise-induced EV-miRNA
remodeling came from studies showing that exercise alters not only
the abundance of circulating EVs, but also the loading of specific
miRNAs into these vesicles. Guescini et al (68) first identified a circulating EV
subpopulation with myogenic features
(SGCA+/CD81+) enriched in muscle-related
miRNAs such as miR-206. They also showed that several EV-associated
miRNAs were positively correlated with VO2max at rest.
After acute aerobic exercise, EV-miR-181a-5p levels significantly
increased, while EV-miR-133b showed an upward trend, supporting the
notion that exercise-related changes in circulating miRNAs may
arise, at least in part, from active EV-mediated secretion and
cargo loading rather than passive leakage from damaged tissues
(68). Subsequent research
reinforced this concept by showing that exercise modality
influences EV-miRNA sorting. In rats, eccentric-biased downhill
running, but not concentric-biased uphill running, increased
multiple myomiRs in exosome-enriched fractions during early
recovery. This suggests that higher mechanical strain more readily
drives myomiRs into EV-associated transport pathways (73). This pattern is consistent with
observations from high mechanical-load resistance exercise. After a
single bout of flywheel-based inertial squats, circulating EV
levels increased and EV-encapsulated miR-206 and miR-146a were
selectively elevated, whereas several other miRNAs remained
unchanged (194). These changes
occurred alongside markers of muscle inflammation, immune
activation, and recovery signaling, suggesting that EV-mediated
ncRNA output may be integrated into a broader program of
post-exercise stress, inflammation, and tissue remodeling. Notably,
some research indicates that exercise-induced EV remodeling may be
driven more by selective cargo changes than by changes in vesicle
number. In untrained men subjected to a muscle-damaging exercise
protocol, EV size and particle number remained unchanged, whereas
EV cargo was selectively remodeled, with a significant decline in
miR-31 at 24 h. Together with concurrent increases in creatine
kinase and delayed-onset muscle soreness, this finding suggests
that EV-mediated miRNA remodeling may contribute to post-exercise
repair, potentially through satellite cell-related pathways
(195). In chronic training,
EV-miRNA responses appear further shaped by training status. In
older men, long-term endurance training produced a stable resting
exosomal miRNA signature distinct from that of sedentary controls
(196). Following an acute bout
of cycling, both trained and sedentary individuals exhibited exomiR
remodeling, but the composition and temporal dynamics of these
changes differed markedly between groups. Pathway analysis
suggested that numerous exercise-responsive exosomal miRNAs target
key nodes within the IGF-1/AKT/mTOR/Forkhead box O axis, with
trained individuals showing a profile more consistent with IGF-1
pathway activation (196).
These findings indicate that long-term training can reprogram the
EV-miRNA response to acute exercise, supporting the view that
EV-miRNAs may function both as molecular fingerprints of training
status and as mediators of adaptive signaling.
Previous research has advanced from descriptive
profiling to functional and mechanistic validation of
exercise-induced EV-miRNA remodeling. In a diabetic heart model,
regular exercise increased cardiac-associated EV release and
remodeled their miRNA cargo, notably upregulating miR-29b and
miR-455. Mechanistically, these exercise-induced EV-miRNAs,
particularly miR-455, were linked to reduced MMP9 expression and
activity, thereby attenuating extracellular matrix remodeling,
fibrosis, and myocardial dysfunction (146). In parallel animal and human
studies, Hou et al (45)
showed that chronic exercise did not necessarily increase
circulating exosome abundance 24 h after training, but
substantially altered exosomal miRNA composition and function. They
identified miR-342-5p as a key exercise-induced exosomal miRNA that
protects against ischemia-reperfusion injury by suppressing
apoptosis-related targets, including Caspase9, Jnk2,
and Ppm1f, while enhancing AKT-dependent survival signaling.
Notably, laminar shear stress also increased miR-342-5p expression
in endothelial cells and their exosomes, suggesting that
exercise-induced hemodynamic signals may contribute to systemic
protection through an endothelial EV-miRNA pathway (45). In addition to systemic
cross-organ signaling, EV-miRNA communication also shapes local
tissue remodeling. Fry et al (197) demonstrated that during
mechanical overload-induced skeletal muscle hypertrophy, myogenic
progenitor cells (MPCs) regulate the stromal microenvironment
through exosome-mediated transfer of miR-206. MPC-derived exosomes
delivered miR-206 to fibrogenic cells, suppressed its target gene
ribosome binding protein 1 (Rrbp1), reduced collagen and
other extracellular matrix-related transcripts, and thereby limited
excessive fibrosis while supporting appropriate muscle remodeling.
By contrast, loss of MPCs or disruption of miRNA
processing/exosomal miR-206 signaling exacerbated extracellular
matrix deposition and impaired remodeling quality. These findings
establish a direct mechanistic axis linking mechanical loading,
EV-mediated miRNA transfer, recipient-cell gene reprogramming, and
tissue adaptation (197).
Although much of the available literature remains
associative, several recent studies have begun to provide stronger
functional support that exercise-responsive EV-miRNA cargoes can
regulate recipient-cell gene programs and tissue adaptation
(34,198). In particular, some reports now
extend beyond descriptive profiling by linking exercise-induced
changes in EV-miRNA cargo to downstream target repression and
measurable phenotypic effects in recipient tissues. In a diabetic
heart model, exercise-induced enrichment of EV-associated miR-455
was linked to reduced MMP9 expression and attenuation of
extracellular matrix remodeling and myocardial dysfunction
(146). Similarly,
exercise-responsive exosomal miR-342-5p has been functionally
linked with suppression of apoptosis-related targets and enhanced
cardioprotection (45). Among
the currently available examples, the MPC-derived exosomal
miR-206/Rrbp1 axis provides especially direct support that
EV-mediated miRNA transfer can reshape recipient-cell gene
expression and local tissue remodeling (197). Nevertheless, direct evidence
that specific exercise-responsive EV-miRNA cargoes induce canonical
epigenetic remodeling in recipient tissues, rather than broader
post-transcriptional or signaling changes, remains limited and
warrants further investigation.
Unresolved challenges in EV research
Current studies suggest that exercise-induced EV
remodeling may contribute to cross-organ metabolic adaptation,
inflammatory regulation, and tissue protection, but the field
remains largely at a transitional stage between descriptive
observation and mechanistic validation (34,37,118,199). A major limitation is
methodological heterogeneity, much of which arises from differences
in EV isolation and characterization strategies. Commonly used EV
isolation techniques include differential ultracentrifugation, size
exclusion chromatography, and immunoaffinity-based isolation, each
with distinct strengths and limitations in terms of recovery,
purity, and subtype selectivity (200). In general, ultracentrifugation
is widely used but may co-isolate contaminants, size exclusion
chromatography often improves purity, and immunoaffinity-based
approaches provide higher specificity for selected EV populations
(201). Terminology remains
inconsistent across studies, with EVs, exosomes, microvesicles, and
small EVs often used interchangeably, even though isolation
procedures alone are usually insufficient to define vesicle subtype
with precision (202). As
emphasized in the Minimal Information for Studies of Extracellular
Vesicles (MISEV)2018 recommendations, no single method is
universally optimal, and isolation procedures alone are often
insufficient to define EV subtypes with precision (48). MISEV2018 therefore advocates the
use of 'extracellular vesicle' as a generic term when subtype
assignment remains uncertain, together with transparent reporting
of isolation workflows, complementary characterization methods,
inclusion of both positive and negative markers, and careful
assessment of co-isolated non-vesicular contaminants. These
considerations are particularly important in exercise-EV studies,
where methodological variation can substantially affect the
apparent abundance, composition, and inferred function of
circulating EV preparations (48). In addition, plasma and serum
contain lipoproteins, albumin, and protein-RNA complexes that
overlap extensively with small EVs in size and physicochemical
properties; when sample purity is inadequate, apparent changes in
EV number or EV-miRNA cargo may be misinterpreted (203). Another critical source of
confounding, particularly in training studies, is sampling time.
Because circulating small EV dynamics can be rapid, samples
collected too close to the final exercise bout may reflect acute
release and clearance rather than true training-induced homeostatic
remodeling, thereby obscuring the distinction between acute
responses and chronic adaptation (66). At the same time, although
numerous studies report exercise-associated changes in circulating
miRNA profiles, direct evidence remains limited regarding their
precise tissue origins, dominant carrier subtypes, destination
organs, and functional uptake (37,65,204). Moving the field forward will
therefore require more rigorous experimental design under the MISEV
framework. This includes improved EV isolation and characterization
using multi-step workflows, more systematic reporting of positive
and negative markers as well as contamination indicators, and
clearer separation of acute stress responses from chronic training
adaptations through appropriate sampling windows (such as ≥24-48 h
after the last exercise bout). Recent advances in multi-omics are
also helping to refine EV characterization beyond particle counting
and bulk marker detection (205). Integrative analyses combining
proteomics, lipidomics, and surfaceome profiling can improve
discrimination between circulating EVs and non-vesicular
contaminants, while providing a more detailed molecular map of EV
composition in human plasma (206). For the exercise-EV field, these
approaches may be especially useful for resolving vesicle
heterogeneity, identifying tissue-informative cargoes, and
connecting EV remodeling with downstream metabolic and epigenetic
effects. More importantly, tissue-specific tracing, single-particle
profiling, and causal intervention studies will be needed to define
the contribution of distinct EV subtypes to ncRNA-mediated
inter-organ communication and to clarify how these signals
intersect with DNA methylation, histone modification, chromatin
accessibility, and transcriptional reprogramming (203).
An additional translational limitation is that much
of the current mechanistic framework has been established in animal
models or cell-based systems, whereas direct human evidence remains
comparatively limited. In preclinical studies, exercise-derived EVs
have been linked to specific miRNA cargoes, downstream target
regulation, and functional effects in recipient tissues. By
contrast, most human studies still rely on measurements of
circulating EV abundance, surface markers, or cargo associations
before and after exercise, without directly establishing tissue
origin, target-organ delivery, cellular uptake, or causal
contribution to physiological adaptation. As a result, although the
available data strongly support the biological plausibility of
EV-mediated exercise signaling, the extent to which these
mechanisms operate in humans remains incompletely resolved.
Bridging this gap will require better integration of mechanistic
preclinical studies with rigorously designed human investigations
that incorporate refined EV characterization, longitudinal
sampling, and functional validation.
Overall, as methodological standardization, refined
EV subtyping, and integrative multi-omics approaches continue to
advance, research on the exercise-EV-miRNA-epigenetic axis is
likely to move from predominantly descriptive association toward
more reproducible and causally interpretable mechanistic
investigation. Such progress will not only deepen our understanding
of how exercise promotes systemic health and metabolic adaptation,
but may also support the development of biomarkers of exercise
responsiveness, more precise exercise prescriptions, and future
exercise-mimetic strategies.
Conclusions
EVs have emerged as key mediators of the systemic
benefits of exercise. Far from being passive by-products of
cellular stress, they act as dynamic carriers of bioactive cargo,
including proteins, metabolites, and regulatory ncRNAs, that
transmit exercise-induced signals to recipient cells and reshape
downstream signaling and gene expression. Both acute and chronic
exercise induce context-dependent remodeling of EV abundance,
origin, and cargo, influenced by factors such as exercise modality,
intensity, timing, training status, and host characteristics. Among
the various cargoes, miRNAs play a particularly pivotal role as
mobile epigenetic regulators that link exercise to sustained
changes in metabolism, inflammation, tissue repair, and organ
protection (Fig. 2). Current
evidence supports the exercise-EV-epigenetic axis as a meaningful
framework for understanding long-term physiological adaptation.
However, progress in the field is still constrained by
methodological heterogeneity, inconsistent terminology, and limited
mechanistic insight into EV biogenesis, targeting, and functional
uptake. Addressing these challenges through rigorous
standardization, refined EV characterization, and in vivo
causal studies will be essential. A clearer understanding of
exercise-derived EVs may not only deepen our mechanistic
understanding of exercise adaptation, but also advance the
development of biomarkers, precision exercise strategies, and
EV-based therapeutic approaches. This overall framing is consistent
with the synthesis in the present review, in which exercise
remodels EV abundance, subtype distribution, and cargo, with miRNAs
emerging as a functionally important component, while the field
remains limited by heterogeneity and incomplete mechanistic
validation.
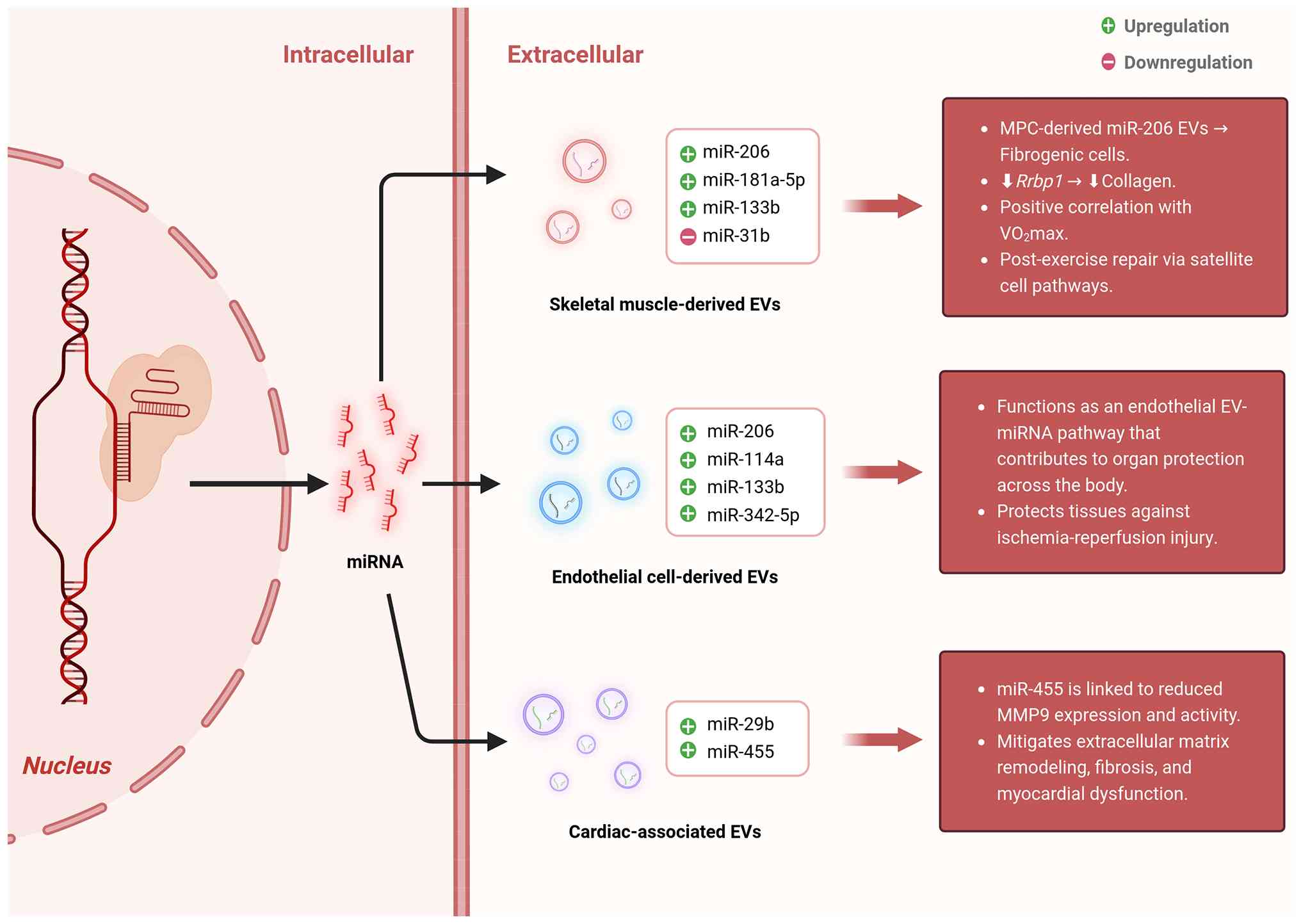 | Figure 2Representative exercise-responsive
miRNAs enriched in EVs and their potential biological functions.
The schematic summarizes representative miRNAs carried by
exercise-responsive EVs from different cellular or
tissue-associated sources. Exercise may alter the abundance of
EV-associated miRNAs released from skeletal muscle-related cells,
endothelial cells, and cardiac-associated compartments. Skeletal
muscle-derived EV miRNAs, including miR-206, miR-181a-5p, miR-133b,
and miR-31, are implicated in muscle remodeling, satellite
cell-related repair, fibrogenic cell regulation, and exercise
capacity. Endothelial cell-derived EV miRNAs, such as miR-206,
miR-114a, miR-133b, and miR-342-5p, may contribute to vascular and
organ-protective responses, including protection against
ischemia-reperfusion injury. Cardiac-associated EV miRNAs,
including miR-29b and miR-455, have been linked to regulation of
extracellular matrix remodeling, fibrosis, and myocardial
dysfunction. Green and red symbols indicate reported upregulation
and downregulation, respectively. Together, these examples
illustrate how exercise-induced EV-miRNA remodeling may connect
local cellular responses with systemic tissue protection and
adaptive remodeling. EVs, extracellular vesicles; miR, microRNA;
MPC, myogenic progenitor cell; Rrbp1, ribosome binding protein 1;
MMP9, matrix metalloproteinase 9. |
Availability of data and materials
Not applicable.
Authors' contributions
XZ and BZ contributed to the study conception and
design, data collection, figure drawing, and manuscript drafting.
Both authors read and approved the final manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
JNK2
|
c-Jun N-terminal kinase 2
|
|
EMVs
|
endothelial-derived microvesicles
|
|
EPC
|
endothelial progenitor cell
|
|
EVs
|
extracellular vesicles
|
|
HSP
|
heat shock protein
|
|
IGF-1
|
insulin-like growth factor-1
|
|
AKT
|
protein kinase B
|
|
mTOR
|
mechanistic target of rapamycin
|
|
miRNA or miR
|
microRNA
|
|
MISEV
|
Minimal Information for Studies of
Extracellular Vesicles
|
|
MMP9
|
matrix metalloproteinase 9
|
|
MPCs
|
myogenic progenitor cells
|
|
MVBs
|
multivesicular bodies
|
|
MVs
|
microvesicles
|
|
myomiRs
|
muscle-specific microRNAs
|
|
ncRNA
|
non-coding RNA
|
|
PMVs
|
platelet-derived microvesicles
|
|
Treg
|
regulatory T cell
|
Acknowledgements
Not applicable.
Funding
This work was supported by the Youth Project of Education of
National Social Science Foundation of China: A study on the
accurate model of traditional exercises for the intervention of
juvenile myopia in the new era (grant no. CLA210279).
References
|
1
|
Myers J, Prakash M, Froelicher V, Do D,
Partington S and Atwood JE: Exercise capacity and mortality among
men referred for exercise testing. N Engl J Med. 346:793–801. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vina J, Sanchis-Gomar F, Martinez-Bello V
and Gomez-Cabrera MC: Exercise acts as a drug; the pharmacological
benefits of exercise. Br J Pharmacol. 167:1–12. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wen CP, Wai JP, Tsai MK, Yang YC, Cheng
TY, Lee MC, Chan HT, Tsao CK, Tsai SP and Wu X: Minimum amount of
physical activity for reduced mortality and extended life
expectancy: A prospective cohort study. Lancet. 378:1244–1253.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ling C and Rönn T: Epigenetics in human
obesity and type 2 diabetes. Cell Metab. 29:1028–1044. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
McGee SL and Hargreaves M: Epigenetics and
exercise. Trends Endocrinol Metab. 30:636–645. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mendis S, Davis S and Norrving B:
Organizational update: The world health organization global status
report on noncommunicable diseases 2014; one more landmark step in
the combat against stroke and vascular disease. Stroke.
46:e121–e122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hayıroğlu M, Çınar T, Çinier G, Karakaya
A, Yıldırım M, Güney BÇ, Öz A, Gündoğmuş PD, Ösken A, Özkan A, et
al: The effect of 1-year mean step count on the change in the
atherosclerotic cardiovascular disease risk calculation in patients
with high cardiovascular risk: A sub-study of the LIGHT randomized
clinical trial. Kardiol Pol. 79:1140–1142. 2021. View Article : Google Scholar
|
|
8
|
Hayıroğlu M, Çınar T, Cilli Hayıroğlu S,
Şaylık F, Uzun M and Tekkeşin AI: The role of smart devices and
mobile application on the change in peak VO(2) in patients with
high cardiovascular risk: A sub-study of the LIGHT randomised
clinical trial. Acta Cardiol. 78:1000–1005. 2023. View Article : Google Scholar
|
|
9
|
Yi J, Chen J, Yao X, Zhao Z, Niu X, Li X,
Sun J, Ji Y, Shang T, Gong L, et al: Myokine-mediated muscle-organ
interactions: Molecular mechanisms and clinical significance.
Biochem Pharmacol. 242:1173262025. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Smith JAB, Murach KA, Dyar KA and Zierath
JR: Exercise metabolism and adaptation in skeletal muscle. Nat Rev
Mol Cell Biol. 24:607–632. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mathieu M, Martin-Jaular L, Lavieu G and
Théry C: Specificities of secretion and uptake of exosomes and
other extracellular vesicles for cell-to-cell communication. Nat
Cell Biol. 21:9–17. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wortzel I, Dror S, Kenific CM and Lyden D:
Exosome-mediated metastasis: Communication from a distance. Dev
Cell. 49:347–360. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Walzik D, Wences Chirino TY, Zimmer P and
Joisten N: Molecular insights of exercise therapy in disease
prevention and treatment. Signal Transduct Target Ther. 9:1382024.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Egan B and Zierath JR: Exercise metabolism
and the molecular regulation of skeletal muscle adaptation. Cell
Metab. 17:162–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Furrer R and Handschin C: Molecular
aspects of the exercise response and training adaptation in
skeletal muscle. Free Radic Biol Med. 223:53–68. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo Z, Sun Y, Qi B, Lin J, Chen Y, Xu Y
and Chen J: Human bone marrow mesenchymal stem cell-derived
extracellular vesicles inhibit shoulder stiffness via let-7a/Tgfbr1
axis. Bioact Mater. 17:344–359. 2022.PubMed/NCBI
|
|
17
|
Luo Z, Qi B, Sun Y, Chen Y, Lin J, Qin H,
Wang N, Shi R, Shang X, Chen S and Chen J: Engineering Bioactive M2
macrophage-polarized, anti-inflammatory, miRNA-Based liposomes for
functional muscle repair: From exosomal mechanisms to biomaterials.
Small. 18:e22019572022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun Y, Luo Z, Chen Y, Lin J, Zhang Y, Qi B
and Chen J: si-Tgfbr1-loading liposomes inhibit shoulder capsule
fibrosis via mimicking the protective function of exosomes from
patients with adhesive capsulitis. Biomater Res. 26:392022.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Raposo G and Stoorvogel W: Extracellular
vesicles: Exosomes, microvesicles, and friends. J Cell Biol.
200:373–383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hugel B, Martínez MC, Kunzelmann C and
Freyssinet JM: Membrane microparticles: Two sides of the coin.
Physiology (Bethesda). 20:22–27. 2005.PubMed/NCBI
|
|
21
|
Smalley DM, Sheman NE, Nelson K and
Theodorescu D: Isolation and identification of potential urinary
microparticle biomarkers of bladder cancer. J Proteome Res.
7:2088–2096. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Erdbrügger U and Lannigan J: Analytical
challenges of extracellular vesicle detection: A comparison of
different techniques. Cytometry A. 89:123–134. 2016. View Article : Google Scholar
|
|
23
|
Zhang Y, Lan M and Chen Y: Minimal
information for studies of extracellular vesicles (MISEV): Ten-year
evolution (2014-2023). Pharmaceutics. 16:13942024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vaiaki EM and Falasca M: Comparative
analysis of the minimal information for studies of extracellular
vesicles guidelines: Advancements and implications for
extracellular vesicle research. Semin Cancer Biol. 101:12–24. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Silvestri M, Grazioli E, Duranti G, Sgrò P
and Dimauro I: Exploring the impact of exercise-derived
extracellular vesicles in cancer biology. Biology (Basel).
13:7012024.PubMed/NCBI
|
|
26
|
Whitham M, Parker BL, Friedrichsen M,
Hingst JR, Hjorth M, Hughes WE, Egan CL, Cron L, Watt KI, Kuchel
RP, et al: Extracellular vesicles provide a means for tissue
crosstalk during exercise. Cell Metab. 27:237–251.e4. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nederveen JP, Warnier G, Di Carlo A,
Nilsson MI and Tarnopolsky MA: Extracellular vesicles and exosomes:
insights from exercise science. Front Physiol. 11:6042742021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen Y, Hong T, Wang S, Mo J, Tian T and
Zhou X: Epigenetic modification of nucleic acids: From basic
studies to medical applications. Chem Soc Rev. 46:2844–2872. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu YL, Lin ZJ, Li CC, Lin X, Shan SK, Guo
B, Zheng MH, Li F, Yuan LQ and Li ZH: Epigenetic regulation in
metabolic diseases: Mechanisms and advances in clinical study.
Signal Transduct Target Ther. 8:982023. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dimauro I, Paronetto MP and Caporossi D:
Exercise, redox homeostasis and the epigenetic landscape. Redox
Biol. 35:1014772020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Oh ES and Petronis A: Origins of human
disease: The chrono-epigenetic perspective. Nat Rev Genet.
22:533–546. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou H, Hu S and Yan W: Extracellular
vesicles as modifiers of epigenomic profiles. Trends Genet.
40:797–809. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang S, Kang S, Kwon O, Seo W, Jin EJ, Ha
H and Huh JY: Small extracellular vesicles derived from damaged
muscle aggravate kidney injury progression. J Cachexia Sarcopenia
Muscle. 16:e138612025. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pinto-Hernandez P, Fernandez-Sanjurjo M,
Paget D, Caravia XM, Roiz-Valle D, Castilla-Silgado J, Diez-Robles
S, Coto-Vilcapoma A, Fernandez-Vivero D, Gama-Perez P, et al:
Training-induced plasma miR-29a-3p is secreted by skeletal muscle
and contributes to metabolic adaptations to resistance exercise in
mice. Mol Metab. 98:1021732025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Wei Y, Liu H and Guo Y:
Protective effect of exercise on metabolic dysfunction-associated
fatty liver disease: Potential epigenetic mechanisms (Review). Int
J Mol Med. 56:1462025. View Article : Google Scholar
|
|
36
|
Safdar A, Saleem A and Tarnopolsky MA: The
potential of endurance exercise-derived exosomes to treat metabolic
diseases. Nat Rev Endocrinol. 12:504–517. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li H, Liu G, Wang B and Momeni MR:
Exosomes and microRNAs as mediators of the exercise. Eur J Med Res.
30:382025. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Joaquim VHA, Pereira NP, Improta-Caria AC,
de Avila Pelozin BR, Fernandes T and Oliveira EM: Exosomes and
microRNAs: Molecular mediators linking obesity, exercise, and
metabolic regulation. Curr Diab Rep. 25:552025. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cheng L and Hill AF: Therapeutically
harnessing extracellular vesicles. Nat Rev Drug Discov. 21:379–399.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yu D, Li Y, Wang M, Gu J, Xu W, Cai H,
Fang X and Zhang X: Exosomes as a new frontier of cancer liquid
biopsy. Mol Cancer. 21:562022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Welsh JA, Goberdhan DCI, O'Driscoll L,
Buzas EI, Blenkiron C, Bussolati B, Cai H, Di Vizio D, Driedonks
TAP, Erdbrügger U, et al: Minimal information for studies of
extracellular vesicles (MISEV2023): From basic to advanced
approaches. J Extracell Vesicles. 13:e124042024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park S and Moon HY: Urinary extracellular
vesicle as a potential biomarker of exercise-induced fatigue in
young adult males. Eur J Appl Physiol. 122:2175–2188. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang X, Kraus WE, Houmard JA, Johnson JL
and Kraus VB: Plasma extracellular vesicle signatures of metabolic
health and exercise response in a pilot study of older adults. Am J
Physiol Cell Physiol. 330:C379–C389. 2026. View Article : Google Scholar :
|
|
44
|
Bei Y, Xu T, Lv D, Yu P, Xu J, Che L, Das
A, Tigges J, Toxavidis V, Ghiran I, et al: Exercise-induced
circulating extracellular vesicles protect against cardiac
ischemia-reperfusion injury. Basic Res Cardiol. 112:382017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hou Z, Qin X, Hu Y, Zhang X, Li G, Wu J,
Li J, Sha J, Chen J, Xia J, et al: Longterm exercise-derived
exosomal miR-342-5p: A novel exerkine for cardioprotection. Circ
Res. 124:1386–1400. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wolf P: The nature and significance of
platelet products in human plasma. Br J Haematol. 13:269–288. 1967.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Johnstone RM, Mathew A, Mason AB and Teng
K: Exosome formation during maturation of mammalian and avian
reticulocytes: Evidence that exosome release is a major route for
externalization of obsolete membrane proteins. J Cell Physiol.
147:27–36. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Théry C, Witwer KW, Aikawa E, Alcaraz MJ,
Anderson JD, Andriantsitohaina R, Antoniou A, Arab T, Archer F,
Atkin-Smith GK, et al: Minimal information for studies of
extracellular vesicles 2018 (MISEV2018): A position statement of
the International Society for Extracellular Vesicles and update of
the MISEV2014 guidelines. J Extracell Vesicles. 7:15357502018.
View Article : Google Scholar
|
|
49
|
Johnstone RM, Adam M, Hammond JR, Orr L
and Turbide C: Vesicle formation during reticulocyte maturation.
Association of plasma membrane activities with released vesicles
(exosomes). J Biol Chem. 262:9412–9420. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lötvall J, Hill AF, Hochberg F, Buzás EI,
Di Vizio D, Gardiner C, Gho YS, Kurochkin IV, Mathivanan S,
Quesenberry P, et al: Minimal experimental requirements for
definition of extracellular vesicles and their functions: A
position statement from the International Society for extracellular
vesicles. J Extracell Vesicles. 3:269132014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xiang H, Bao C, Chen Q, Gao Q, Wang N, Gao
Q and Mao L: Extracellular vesicles (EVs)' journey in recipient
cells: From recognition to cargo release. J Zhejiang Univ Sci B.
25:633–655. 2024.In English, Chinese. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rilla K, Pasonen-Seppänen S, Deen AJ,
Koistinen VVT, Wojciechowski S, Oikari S, Kärnä R, Bart G, Törrönen
K, Tammi RH and Tammi MI: Hyaluronan production enhances shedding
of plasma membrane-derived microvesicles. Exp Cell Res.
319:2006–2018. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
György B, Szabó TG, Pásztói M, Pál Z,
Misják P, Aradi B, László V, Pállinger E, Pap E, Kittel A, et al:
Membrane vesicles, current state-of-the-art: Emerging role of
extracellular vesicles. Cell Mol Life Sci. 68:2667–2688. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
McCullough J, Fisher RD, Whitby FG,
Sundquist WI and Hill CP: ALIX-CHMP4 interactions in the human
ESCRT pathway. Proc Natl Acad Sci USA. 105:7687–7691. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gurung S, Perocheau D, Touramanidou L and
Baruteau J: The exosome journey: From biogenesis to uptake and
intracellular signalling. Cell Commun Signal. 19:472021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lee YJ, Shin KJ and Chae YC: Regulation of
cargo selection in exosome biogenesis and its biomedical
applications in cancer. Exp Mol Med. 56:877–889. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hou S, Wang L, Yang C, Li Y and Liu H:
MACC1 drives metastasis in colorectal cancer by coordinating
YKT6-dependent exosome biogenesis and c-Met cargo selection. Cell
Signal. 135:1120732025. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Arya SB, Collie SP and Parent CA: The
ins-and-outs of exosome biogenesis, secretion, and internalization.
Trends Cell Biol. 34:90–108. 2024. View Article : Google Scholar
|
|
59
|
Wei H, Chen Q, Lin L, Sha C, Li T, Liu Y,
Yin X, Xu Y, Chen L, Gao W, et al: Regulation of exosome production
and cargo sorting. Int J Biol Sci. 17:163–177. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Navasiolava NM, Dignat-George F, Sabatier
F, Larina IM, Demiot C, Fortrat JO, Gauquelin-Koch G, Kozlovskaya
IB and Custaud MA: Enforced physical inactivity increases
endothelial microparticle levels in healthy volunteers. Am J
Physiol Heart Circ Physiol. 299:H248–H256. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ni P, Yang L and Li F: Exercise-derived
skeletal myogenic exosomes as mediators of intercellular crosstalk:
A major player in health, disease, and exercise. J Physiol Biochem.
79:501–510. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Blank M, Israeli D and Shoenfeld Y:
Exercise, autoimmune diseases and T-regulatory cells. J Autoimmun.
149:1033172024. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sukhnanan K, Ross JR, Chao NJ and Chen BJ:
Endothelial cell derived extracellular vesicles and hematopoiesis.
Radiat Res. 202:215–226. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tao SC, Guo SC and Zhang CQ:
Platelet-derived extracellular vesicles: An emerging therapeutic
approach. Int J Biol Sci. 13:828–834. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Eichner NZM, Erdbrügger U and Malin SK:
Extracellular vesicles: A novel target for exercise-mediated
reductions in type 2 diabetes and cardiovascular disease risk. J
Diabetes Res. 2018:78072452018. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Caporossi D, Jackson MJ and
Henriquez-Olguin C: Intracellular and extracellular redox signals
during exercise and aging. Free Radic Biol Med. 242:521–534. 2026.
View Article : Google Scholar
|
|
67
|
Pedersen BK and Febbraio MA: Muscles,
exercise and obesity: Skeletal muscle as a secretory organ. Nat Rev
Endocrinol. 8:457–465. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Guescini M, Canonico B, Lucertini F,
Maggio S, Annibalini G, Barbieri E, Luchetti F, Papa S and Stocchi
V: Muscle releases alpha-sarcoglycan positive extracellular
vesicles carrying miRNAs in the Bloodstream. PLoS One.
10:e01250942015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Jalabert A, Vial G, Guay C, Wiklander OP,
Nordin JZ, Aswad H, Forterre A, Meugnier E, Pesenti S, Regazzi R,
et al: Exosome-like vesicles released from lipid-induced
insulin-resistant muscles modulate gene expression and
proliferation of beta recipient cells in mice. Diabetologia.
59:1049–1058. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Forterre A, Jalabert A, Chikh K, Pesenti
S, Euthine V, Granjon A, Errazuriz E, Lefai E, Vidal H and Rome S:
Myotube-derived exosomal miRNAs downregulate Sirtuin1 in myoblasts
during muscle cell differentiation. Cell Cycle. 13:78–89. 2014.
View Article : Google Scholar
|
|
71
|
Liu HC, Han DS, Hsu CC and Wang JS:
Circulating MicroRNA-486 and MicroRNA-146a serve as potential
biomarkers of sarcopenia in the older adults. BMC Geriatr.
21:862021. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Sempere LF, Freemantle S, Pitha-Rowe I,
Moss E, Dmitrovsky E and Ambros V: Expression profiling of
mammalian microRNAs uncovers a subset of brain-expressed microRNAs
with possible roles in murine and human neuronal differentiation.
Genome Biol. 5:R132004. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yin X, Zhao Y, Zheng YL, Wang JZ, Li W, Lu
QJ, Huang QN, Zhang CY, Chen X and Ma JZ: Time-course responses of
muscle-specific MicroRNAs following acute uphill or downhill
exercise in sprague-dawley rats. Front Physiol. 10:12752019.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Aoi W and Tanimura Y: Roles of skeletal
muscle-derived exosomes in organ metabolic and immunological
communication. Front Endocrinol (Lausanne). 12:6972042021.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Madison RD, McGee C, Rawson R and Robinson
GA: Extracellular vesicles from a muscle cell line (C2C12) enhance
cell survival and neurite outgrowth of a motor neuron cell line
(NSC-34). J Extracell Vesicles. 3:2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wu Z, Gao Y, Chen Q and Yuan L:
Inter-Organ crosstalk and regulatory mechanisms of skeletal
muscle-derived extracellular vesicles in systemic metabolic
homeostasis. J Cell Mol Med. 29:e708962025. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Matsuzaka Y, Tanihata J, Komaki H,
Ishiyama A, Oya Y, Rüegg U, Takeda SI and Hashido K:
Characterization and functional analysis of extracellular vesicles
and muscle-abundant miRNAs (miR-1, miR-133a, and miR-206) in C2C12
Myocytes and mdx Mice. PLoS One. 11:e01678112016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang A, Li M, Wang B, Klein JD, Price SR
and Wang XH: miRNA-23a/27a attenuates muscle atrophy and renal
fibrosis through muscle-kidney crosstalk. J Cachexia Sarcopenia
Muscle. 9:755–770. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Noboa-Velástegui JA, Valdez-Vega RI,
Castro-Albarran J, Madrigal-Ruiz PM, Fletes-Rayas AL, Ruiz-Quezada
SL, Ramos-Márquez ME, López-Jiménez JJ, Álvarez I and
Navarro-Hernández RE: Expression of serum and exosomal microRNA-34a
in subjects with increased fat mass. Int J Mol Sci. 27:2702025.
View Article : Google Scholar
|
|
80
|
Roberts TC, Godfrey C, McClorey G, Vader
P, Briggs D, Gardiner C, Aoki Y, Sargent I, Morgan JE and Wood MJ:
Extracellular microRNAs are dynamic non-vesicular biomarkers of
muscle turnover. Nucleic Acids Res. 41:9500–9513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Cacchiarelli D, Legnini I, Martone J,
Cazzella V, D'Amico A, Bertini E and Bozzoni I: miRNAs as serum
biomarkers for Duchenne muscular dystrophy. EMBO Mol Med.
3:258–265. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Karolina DS, Armugam A, Tavintharan S,
Wong MT, Lim SC, Sum CF and Jeyaseelan K: MicroRNA 144 impairs
insulin signaling by inhibiting the expression of insulin receptor
substrate 1 in type 2 diabetes mellitus. PLoS One. 6:e228392011.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Donaldson A, Natanek SA, Lewis A, Man WD,
Hopkinson NS, Polkey M and Kemp PR: Increased skeletal
muscle-specific microRNA in the blood of patients with COPD.
Thorax. 68:1140–1149. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Miyachi M, Tsuchiya K, Yoshida H, Yagyu S,
Kikuchi K, Misawa A, Iehara T and Hosoi H: Circulating
muscle-specific microRNA, miR-206, as a potential diagnostic marker
for rhabdomyosarcoma. Biochem Biophys Res Commun. 400:89–93. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Sossdorf M, Otto GP, Claus RA, Gabriel HH
and Lösche W: Release of pro-coagulant microparticles after
moderate endurance exercise. Platelets. 21:389–391. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sossdorf M, Otto GP, Claus RA, Gabriel HH
and Lösche W: Cell-derived microparticles promote coagulation after
moderate exercise. Med Sci Sports Exerc. 43:1169–1176. 2011.
View Article : Google Scholar
|
|
87
|
Headland SE, Jones HR, D'Sa AS, Perretti M
and Norling LV: Cutting-edge analysis of extracellular
microparticles using ImageStream(X) imaging flow cytometry. Sci
Rep. 4:52372014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Keir DA, Fontana FY, Robertson TC, Murias
JM, Paterson DH, Kowalchuk JM and Pogliaghi S: Exercise intensity
thresholds: Identifying the boundaries of sustainable performance.
Med Sci Sports Exerc. 47:1932–1940. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Guiraud T, Gayda M, Juneau M, Bosquet L,
Meyer P, Théberge-Julien G, Galinier M, Nozza A, Lambert J, Rhéaume
E, et al: A single bout of high-intensity interval exercise does
not increase endothelial or platelet microparticles in stable,
physically fit men with coronary heart disease. Can J Cardiol.
29:1285–1291. 2013.PubMed/NCBI
|
|
90
|
Maruyama K, Kadono T and Morishita E:
Plasma levels of platelet-derived microparticles are increased
after anaerobic exercise in healthy subjects. J Atheroscler Thromb.
19:585–587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Lansford KA, Shill DD, Dicks AB, Marshburn
MP, Southern WM and Jenkins NT: Effect of acute exercise on
circulating angiogenic cell and microparticle populations. Exp
Physiol. 101:155–167. 2016. View Article : Google Scholar
|
|
92
|
Wilhelm EN, González-Alonso J, Parris C
and Rakobowchuk M: Exercise intensity modulates the appearance of
circulating microvesicles with proangiogenic potential upon
endothelial cells. Am J Physiol Heart Circ Physiol.
311:H1297–H1310. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kirk RJ, Peart DJ, Madden LA and Vince RV:
Repeated supra-maximal sprint cycling with and without sodium
bicarbonate supplementation induces endothelial microparticle
release. Eur J Sport Sci. 14:345–352. 2014. View Article : Google Scholar
|
|
94
|
Möbius-Winkler S, Hilberg T, Menzel K,
Golla E, Burman A, Schuler G and Adams V: Time-dependent
mobilization of circulating progenitor cells during strenuous
exercise in healthy individuals. J Appl Physiol (1985).
107:1943–1950. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wahl P, Jansen F, Achtzehn S, Schmitz T,
Bloch W, Mester J and Werner N: Effects of high intensity training
and high volume training on endothelial microparticles and
angiogenic growth factors. PLoS One. 9:e960242014. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wilhelm EN, Mourot L and Rakobowchuk M:
Exercise-Derived microvesicles: A review of the literature. Sports
Med. 48:2025–2039. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Lin C, Guo J and Jia R: Roles of
regulatory T cell-derived extracellular vesicles in human diseases.
Int J Mol Sci. 23:112062022. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Azimi M, Ghabaee M, Moghadasi AN,
Noorbakhsh F and Izad M: Immunomodulatory function of Treg-derived
exosomes is impaired in patients with relapsing-remitting multiple
sclerosis. Immunol Res. 66:513–520. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Punga AR, Andersson M, Alimohammadi M and
Punga T: Disease specific signature of circulating miR-150-5p and
miR-21-5p in myasthenia gravis patients. J Neurol Sci. 356:90–96.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Lovinsky-Desir S, Jung KH, Jezioro JR,
Torrone DZ, de Planell-Saguer M, Yan B, Perera FP, Rundle AG,
Perzanowski MS, Chillrud SN and Miller RL: Physical activity, black
carbon exposure, and DNA methylation in the FOXP3 promoter. Clin
Epigenetics. 9:652017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Catitti G, De Bellis D, Vespa S, Simeone
P, Canonico B and Lanuti P: Extracellular vesicles as players in
the anti-inflammatory inter-cellular crosstalk induced by exercise
training. Int J Mol Sci. 23:140982022. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Darkwah S, Park EJ, Myint PK, Ito A,
Appiah MG, Obeng G, Kawamoto E and Shimaoka M: Potential roles of
muscle-derived extracellular vesicles in remodeling cellular
microenvironment: proposed implications of the exercise-induced
myokine, irisin. Front Cell Dev Biol. 9:6348532021. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Jiang T, Huang Y, Liu H, Xu Q, Gong Y,
Chen Y, Hu X, Han Z and Gao M: Reduced miR-146a promotes REG3A
expression and macrophage migration in polymyositis and
dermatomyositis. Front Immunol. 11:372020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Petriz BA, Gomes CP, Almeida JA, de
Oliveira GP Jr, Ribeiro FM, Pereira RW and Franco OL: The effects
of acute and chronic exercise on skeletal muscle proteome. J Cell
Physiol. 232:257–269. 2017. View Article : Google Scholar
|
|
105
|
Fine J: Exercise and fitness. JAMA.
188:433–436. 1964. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Thyfault JP and Bergouignan A: Exercise
and metabolic health: Beyond skeletal muscle. Diabetologia.
63:1464–1474. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Papaioannou F, Karatzanos E, Chatziandreou
I, Philippou A, Nanas S and Dimopoulos S: Epigenetic effects
following acute and chronic exercise in cardiovascular disease: A
systematic review. Int J Cardiol. 341:88–95. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Mora JC and Valencia WM: Exercise and
older adults. Clin Geriatr Med. 34:145–162. 2018. View Article : Google Scholar
|
|
109
|
Andrade C: Physical exercise and health,
2: Benefits associated with different levels and patterns of
activity. J Clin Psychiatry. 84:23f151102023. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Frühbeis C, Helmig S, Tug S, Simon P and
Krämer-Albers EM: Physical exercise induces rapid release of small
extracellular vesicles into the circulation. J Extracell Vesicles.
4:282392015. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chow LS, Gerszten RE, Taylor JM, Pedersen
BK, van Praag H, Trappe S, Febbraio MA, Galis ZS, Gao Y, Haus JM,
et al: Exerkines in health, resilience and disease. Nat Rev
Endocrinol. 18:273–289. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Kong H, Zheng J, Han J, Meng X, Li X, Wang
X and Zhang XA: The crosstalk among macrophages, chondrocytes and
mesenchymal stem cells in osteoarthritis: The role of extracellular
vesicles. Stem Cell Res Ther. 16:5532025. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wu Y, Hou M, Deng Y, Xia X, Liu Y, Yu J,
Yu C, Yang H, Zhang Y and Zhu X: Swimming exercise induces
redox-lipid crosstalk to ameliorate osteoarthritis progression.
Redox Biol. 81:1035352025. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Vechetti IJ Jr, Valentino T, Mobley CB and
McCarthy JJ: The role of extracellular vesicles in skeletal muscle
and systematic adaptation to exercise. J Physiol. 599:845–861.
2021. View Article : Google Scholar :
|
|
115
|
Safdar A and Tarnopolsky MA: Exosomes as
mediators of the systemic adaptations to endurance exercise. Cold
Spring Harb Perspect Med. 8:a0298272018. View Article : Google Scholar
|
|
116
|
Lombardo M, Aiello G, Fratantonio D, Karav
S and Baldelli S: Functional role of extracellular vesicles in
skeletal muscle physiology and sarcopenia: the importance of
physical exercise and nutrition. Nutrients. 16:30972024. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
D'Souza RF, Woodhead JST, Zeng N,
Blenkiron C, Merry TL, Cameron-Smith D and Mitchell CJ: Circulatory
exosomal miRNA following intense exercise is unrelated to muscle
and plasma miRNA abundances. Am J Physiol Endocrinol Metab.
315:E723–E733. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Estébanez B, Jiménez-Pavón D, Huang CJ,
Cuevas MJ and González-Gallego J: Effects of exercise on exosome
release and cargo in in vivo and ex vivo models: A systematic
review. J Cell Physiol. 236:3336–3353. 2021. View Article : Google Scholar
|
|
119
|
Egan B and Sharples AP: Molecular
responses to acute exercise and their relevance for adaptations in
skeletal muscle to exercise training. Physiol Rev. 103:2057–2170.
2023. View Article : Google Scholar
|
|
120
|
Karpovich PV: Exercise. Annu Rev Physiol.
9:149–162. 1947. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Thomas HJ, Ang T, Morrison DJ, Keske MA
and Parker L: Acute exercise and high-glucose ingestion elicit
dynamic and individualized responses in systemic markers of redox
homeostasis. Front Immunol. 14:11270882023. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Morgan WP: Affective beneficence of
vigorous physical activity. Med Sci Sports Exerc. 17:94–100. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Hammerman H, Schoen FJ and Kloner RA:
Short-term exercise has a prolonged effect on scar formation after
experimental acute myocardial infarction. J Am Coll Cardiol.
2:979–982. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Díaz-Castro F, Tuñón-Suárez M, Rivera P,
Botella J, Cancino J, Figueroa AM, Gutiérrez J, Cantin C, Deldicque
L, Zbinden-Foncea H, et al: A single bout of resistance exercise
triggers mitophagy, potentially involving the ejection of
mitochondria in human skeletal muscle. Acta Physiol (Oxf).
240:e142032024. View Article : Google Scholar
|
|
125
|
Brown EC, Franklin BA, Regensteiner JG and
Stewart KJ: Effects of single bout resistance exercise on glucose
levels, insulin action, and cardiovascular risk in type 2 diabetes:
A narrative review. J Diabetes Complications. 34:1076102020.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Bittel AJ, Bittel DC, Mittendorfer B,
Patterson BW, Okunade AL, Yoshino J, Porter LC, Abumrad NA, Reeds
DN and Cade WT: A single bout of resistance exercise improves
postprandial lipid metabolism in overweight/obese men with
prediabetes. Diabetologia. 63:611–623. 2020. View Article : Google Scholar
|
|
127
|
Feng W, Yu WL, Zhao Z, Guo YD, Zhou XJ,
Zhai KQ, Liu X and Yan ZW: Acute effects of high-intensity interval
exercise and moderate-intensity continuous training on arterial
stiffness and endothelial function in hypertension: A crossover
trial. Sci Rep. 15:370862025. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Pedersen BK and Hoffman-Goetz L: Exercise
and the immune system: Regulation, integration, and adaptation.
Physiol Rev. 80:1055–1081. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Siqueira IR, Palazzo RP and Cechinel LR:
Circulating extracellular vesicles delivering beneficial cargo as
key players in exercise effects. Free Radic Biol Med. 172:273–285.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Helmig S, Frühbeis C, Krämer-Albers EM,
Simon P and Tug S: Release of bulk cell free DNA during physical
exercise occurs independent of extracellular vesicles. Eur J Appl
Physiol. 115:2271–2280. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Oliveira GP Jr, Porto WF, Palu CC, Pereira
LM, Petriz B, Almeida JA, Viana J, Filho NNA, Franco OL and Pereira
RW: Effects of acute aerobic exercise on rats serum extracellular
vesicles diameter, concentration and small RNAs content. Front
Physiol. 9:5322018. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Brahmer A, Neuberger E, Esch-Heisser L,
Haller N, Jorgensen MM, Baek R, Möbius W, Simon P and Krämer-Albers
EM: Platelets, endothelial cells and leukocytes contribute to the
exercise-triggered release of extracellular vesicles into the
circulation. J Extracell Vesicles. 8:16158202019. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Rigamonti AE, Bollati V, Pergoli L, Iodice
S, De Col A, Tamini S, Cicolini S, Tringali G, De Micheli R, Cella
SG and Sartorio A: Effects of an acute bout of exercise on
circulating extracellular vesicles: tissue-, sex-, and BMI-related
differences. Int J Obes (Lond). 44:1108–1118. 2020. View Article : Google Scholar
|
|
134
|
Sapp RM, Shill DD, Roth SM and Hagberg JM:
Circulating microRNAs in acute and chronic exercise: more than mere
biomarkers. J Appl Physiol (1985). 122:702–717. 2017. View Article : Google Scholar
|
|
135
|
Bass H, Whitcomb JF and Forman R: Exercise
training: Therapy for patients with chronic obstructive pulmonary
disease. Chest. 57:116–121. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Balietti M, Fattoretti P, Skalicky M,
Viidik A, Giorgetti B, Grossi Y and Bertoni-Freddari C: The effect
of chronic physical exercise on succinic dehydrogenase activity in
the heart muscle of old rats. Biogerontology. 6:95–100. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Liu J, Xiao F, Choubey A, Kumar SU, Wang
Y, Hong S, Yang T, Otlu HG, Oturmaz ES, Loro E, et al: Muscle
Rev-erb controls time-dependent adaptations to chronic exercise in
mice. Nat Commun. 16:57082025. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Mackinnon LT: Chronic exercise training
effects on immune function. Med Sci Sports Exerc. 32(Suppl 7):
S369–S376. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Audiffren M and André N: The
exercise-cognition relationship: A virtuous circle. J Sport Health
Sci. 8:339–347. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Ploeger HE, Takken T, de Greef MH and
Timmons BW: The effects of acute and chronic exercise on
inflammatory markers in children and adults with a chronic
inflammatory disease: A systematic review. Exerc Immunol Rev.
15:6–41. 2009.PubMed/NCBI
|
|
141
|
Fedewa MV, Hathaway ED, Ward-Ritacco CL,
Williams TD and Dobbs WC: The effect of chronic exercise training
on leptin: A systematic review and meta-analysis of Randomized
controlled trials. Sports Med. 48:1437–1450. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Brennan AM, Benson M, Morningstar J,
Herzig M, Robbins J, Gerszten RE and Ross R: Plasma metabolite
profiles in response to chronic exercise. Med Sci Sports Exerc.
50:1480–1486. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
de Las Heras N, Klett-Mingo M, Ballesteros
S, Martín-Fernández B, Escribano Ó, Blanco-Rivero J, Balfagón G,
Hribal ML, Benito M, Lahera V and Gómez-Hernández A: Chronic
exercise improves mitochondrial function and insulin sensitivity in
brown adipose tissue. Front Physiol. 9:11222018. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Joisten N, Walzik D, Metcalfe AJ, Bloch W
and Zimmer P: Physical exercise as kynurenine pathway modulator in
chronic diseases: Implications for immune and energy homeostasis.
Int J Tryptophan Res. 13:11786469209386882020. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Gleeson M, Bishop NC, Stensel DJ, Lindley
MR, Mastana SS and Nimmo MA: The anti-inflammatory effects of
exercise: Mechanisms and implications for the prevention and
treatment of disease. Nat Rev Immunol. 11:607–615. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Bertoldi K, Cechinel LR, Schallenberger B,
Corssac GB, Davies S, Guerreiro ICK, Belló-Klein A, Araujo ASR and
Siqueira IR: Circulating extracellular vesicles in the aging
process: Impact of aerobic exercise. Mol Cell Biochem. 440:115–125.
2018. View Article : Google Scholar
|
|
147
|
Chaturvedi P, Kalani A, Medina I,
Familtseva A and Tyagi SC: Cardiosome mediated regulation of MMP9
in diabetic heart: role of mir29b and mir455 in exercise. J Cell
Mol Med. 19:2153–2161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Mastrototaro L, Apostolopoulou M, Hartwig
S, Strassburger K, Lipaeva P, Trinks N, Karusheva Y, Gancheva S,
Trenkamp S, Lehr S, et al: The role of exosomes for sustained
specific cardiorespiratory and metabolic improvements in males with
type 2 diabetes after detraining. EBioMedicine. 110:1054712024.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Warnier G, van Doorslaer de Ten Ryen S,
Lannoy C, Mahy T, Antoine N, Boyer E, Kienlen-Campard P, Verboven
K, Copine S, Francaux M and Deldicque L: Effect of a 12-Week
endurance training program on circulating extracellular vesicle
proteome in sedentary adults with obesity. J Extracell Biol.
4:e700872025. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Ma C, Wang J, Liu H, Chen Y, Ma X, Chen S,
Chen Y, Bihl JI and Yang YI: Moderate exercise enhances endothelial
progenitor cell exosomes release and function. Med Sci Sports
Exerc. 50:2024–2032. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Barone R, Macaluso F, Sangiorgi C,
Campanella C, Marino Gammazza A, Moresi V, Coletti D, Conway de
Macario E, Macario AJ, Cappello F, et al: Skeletal muscle Heat
shock protein 60 increases after endurance training and induces
peroxisome proliferator-activated receptor gamma coactivator 1 α1
expression. Sci Rep. 6:197812016. View Article : Google Scholar
|
|
152
|
Kalluri R: The biology and function of
extracellular vesicles in immune response and immunity. Immunity.
57:1752–1768. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Joshi BS, de Beer MA, Giepmans BNG and
Zuhorn IS: Endocytosis of extracellular vesicles and release of
their cargo from endosomes. ACS Nano. 14:4444–4455. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Guo Q, Chen J, Wu J, Mo Z, Ye L, Zhong W,
Zhang Y, Lai H, Zhang Y, Qiu J, et al: Bioprinted mesenchymal stem
cell microfiber-derived extracellular vesicles alleviate unilateral
renal ischemia-reperfusion injury and fibrosis by inhibiting
tubular epithelial cells ferroptosis. Bioact Mater. 40:649–664.
2024.PubMed/NCBI
|
|
155
|
Gao H, Zeng Y, Huang X, A L, Liang Q, Xie
J, Lin X, Gong J, Fan X, Zou T and Xu H: Extracellular vesicles
from organoid-derived human retinal progenitor cells prevent lipid
overload-induced retinal pigment epithelium injury by regulating
fatty acid metabolism. J Extracell Vesicles. 13:e124012024.
View Article : Google Scholar
|
|
156
|
Zhao Q, Li B, Zhang X, Zhao H, Xue W, Yuan
Z, Xu S and Duan G: M2 macrophage-derived lncRNA NORAD in EVs
promotes NSCLC progression via miR-520g-3p/SMIM22/GALE axis. NPJ
Precis Oncol. 8:1852024. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Muraca M and Cappariello A: The role of
extracellular vesicles (EVs) in the epigenetic regulation of bone
metabolism and osteoporosis. Int J Mol Sci. 21:86822020. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Man K, Brunet MY, Fernandez-Rhodes M,
Williams S, Heaney LM, Gethings LA, Federici A, Davies OG, Hoey D
and Cox SC: Epigenetic reprogramming enhances the therapeutic
efficacy of osteoblast-derived extracellular vesicles to promote
human bone marrow stem cell osteogenic differentiation. J Extracell
Vesicles. 10:e121182021. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Cavalli G and Heard E: Advances in
epigenetics link genetics to the environment and disease. Nature.
571:489–499. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Pal S and Tyler JK: Epigenetics and aging.
Sci Adv. 2:e16005842016. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Wu Q, Gao ZJ, Yu X and Wang P: Dietary
regulation in health and disease. Signal Transduct Target Ther.
7:2522022. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Peixoto P, Cartron PF, Serandour AA and
Hervouet E: From 1957 to nowadays: A brief history of epigenetics.
Int J Mol Sci. 21:75712020. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Sapienza C and Issa JP: Diet, nutrition,
and cancer epigenetics. Annu Rev Nutr. 36:665–681. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Qu P, Li L, Jin Q, Liu D, Qiao Y, Zhang Y,
Sun Q, Ran S, Li Z, Liu T and Peng L: Histone methylation
modification and diabetic kidney disease: Potential molecular
mechanisms and therapeutic approaches (Review). Int J Mol Med.
54:1042024. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Cutfield WS, Hofman PL, Mitchell M and
Morison IM: Could epigenetics play a role in the developmental
origins of health and disease? Pediatr Res. 61:68R–75R. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Goldberg AD, Allis CD and Bernstein E:
Epigenetics: A landscape takes shape. Cell. 128:635–638. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Stefanska B, Karlic H, Varga F,
Fabianowska-Majewska K and Haslberger A: Epigenetic mechanisms in
anti-cancer actions of bioactive food components-the implications
in cancer prevention. Br J Pharmacol. 167:279–297. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Stover PJ, James WPT, Krook A and Garza C:
Emerging concepts on the role of epigenetics in the relationships
between nutrition and health. J Intern Med. 284:37–49. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Dehan P, Kustermans G, Guenin S, Horion J,
Boniver J and Delvenne P: DNA methylation and cancer diagnosis: new
methods and applications. Expert Rev Mol Diagn. 9:651–657. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Zhao Q, Liu J, Deng H, Ma R, Liao JY,
Liang H, Hu J, Li J, Guo Z, Cai J, et al: Targeting
mitochondria-located circRNA SCAR alleviates NASH via reducing mROS
output. Cell. 183:76–93.e22. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Lisi V, Senesi G and Balbi C: Converging
protective pathways: Exploring the linkage between physical
exercise, extracellular vesicles and oxidative stress. Free Radic
Biol Med. 208:718–727. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Goodpaster BH and Sparks LM: Metabolic
flexibility in health and disease. Cell Metab. 25:1027–1036. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Harris RT and Dudley GA: Exercise alters
the distribution of ammonia and lactate in blood. J Appl Physiol
(1985). 66:313–317. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
King SE and Skinner MK: Epigenetic
transgenerational inheritance of obesity susceptibility. Trends
Endocrinol Metab. 31:478–494. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Gibson F, Hanly A, Grbic N, Grunberg N, Wu
M, Collard M and Alani RM: Epigenetic dysregulation in autoimmune
and inflammatory skin diseases. Clin Rev Allergy Immunol.
63:447–471. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Yang Y, Cai J, Yang X, Wang K, Sun K, Yang
Z, Zhang L, Yang L, Gu C, Huang X, et al: Dysregulated m6A
modification promotes lipogenesis and development of non-alcoholic
fatty liver disease and hepatocellular carcinoma. Mol Ther.
30:2342–2353. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Ge T, Gu X, Jia R, Ge S, Chai P, Zhuang A
and Fan X: Crosstalk between metabolic reprogramming and
epigenetics in cancer: Updates on mechanisms and therapeutic
opportunities. Cancer Commun (Lond). 42:1049–1082. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Wątroba M, Dudek I, Skoda M, Stangret A,
Rzodkiewicz P and Szukiewicz D: Sirtuins, epigenetics and
longevity. Ageing Res Rev. 40:11–19. 2017. View Article : Google Scholar
|
|
179
|
Avelar RA, Palmer D, Kulaga AY and Fuellen
G: Conserved biological processes in partial cellular
reprogramming: Relevance to aging and rejuvenation. Ageing Res Rev.
108:1027372025. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Wu G, Zhang X and Gao F: The epigenetic
landscape of exercise in cardiac health and disease. J Sport Health
Sci. 10:648–659. 2021. View Article : Google Scholar
|
|
181
|
Widmann M, Nieß AM and Munz B: Physical
exercise and epigenetic modifications in skeletal muscle. Sports
Med. 49:509–523. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Polakovičová M, Musil P, Laczo E, Hamar D
and Kyselovič J: Circulating MicroRNAs as Potential Biomarkers of
Exercise Response. Int J Mol Sci. 17:15532016. View Article : Google Scholar
|
|
183
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Nemeth K, Bayraktar R, Ferracin M and
Calin GA: Non-coding RNAs in disease: From mechanisms to
therapeutics. Nat Rev Genet. 25:211–232. 2024. View Article : Google Scholar
|
|
185
|
Xu T, Liu Q, Yao J, Dai Y, Wang H and Xiao
J: Circulating microRNAs in response to exercise. Scand J Med Sci
Sports. 25:e149–e154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Statello L, Guo CJ, Chen LL and Huarte M:
Gene regulation by long non-coding RNAs and its biological
functions. Nat Rev Mol Cell Biol. 22:96–118. 2021. View Article : Google Scholar
|
|
187
|
Zhao H, Chen X, Hu G, Li C, Guo L, Zhang
L, Sun F, Xia Y, Yan W, Cui Z, et al: Small extracellular vesicles
from brown adipose tissue mediate exercise cardioprotection. Circ
Res. 130:1490–1506. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Dos Santos JAC, Veras ASC, Batista VRG,
Tavares MEA, Correia RR, Suggett CB and Teixeira GR: Physical
exercise and the functions of microRNAs. Life Sci. 304:1207232022.
View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Wang J, Li L, Zhang Z, Zhang X, Zhu Y,
Zhang C and Bi Y: Extracellular vesicles mediate the communication
of adipose tissue with brain and promote cognitive impairment
associated with insulin resistance. Cell Metab. 34:1264–1279.e8.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Katayama M, Caria E, Van Simaeys D, Sanz
AY, Barrès R, Caidahl K, Wiklander OPB, El Andaloussi S, Berggren
PO, Zierath JR and Krook A: Exercise training-induced extracellular
miR-136-3p modulates glucose uptake and myogenesis through
targeting of NRDC in human skeletal muscle. J Sport Health Sci.
15:1010912025. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Ionescu A, Ankol L, Ganapathy Subramaniam
A, Altman T, Magen I, Cohen Y, Danino Y, Gradus-Pery T, Niv Y, Bar
Avi O, et al: Muscle-derived miR-126 regulates TDP-43 axonal local
synthesis and NMJ integrity in ALS models. Nat Neurosci.
28:2201–2216. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Chen M, Li Y, Zhang M, Ge S, Feng T, Chen
R, Shen J, Li R, Wang Z, Xie Y, et al: Histone deacetylase
inhibition enhances extracellular vesicles from muscle to promote
osteogenesis via miR-873-3p. Signal Transduct Target Ther.
9:2562024. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Zhao Y, Gao L, Chen J, Wei J, Lin G, Hu K,
Zhao W, Wei W, Huang W, Gao L, et al: Remote limb ischemic
conditioning alleviates steatohepatitis via extracellular
vesicle-mediated muscle-liver crosstalk. Cell Metab. 37:886–902.e7.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Annibalini G, Contarelli S, Lucertini F,
Guescini M, Maggio S, Ceccaroli P, Gervasi M, Ferri Marini C,
Fardetti F, Grassi E, et al: Muscle and systemic molecular
responses to a single flywheel based iso-inertial training session
in resistance-trained men. Front Physiol. 10:5542019. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Lovett JAC, Durcan PJ and Myburgh KH:
Investigation of circulating extracellular vesicle MicroRNA
following two consecutive bouts of muscle-damaging exercise. Front
Physiol. 9:11492018. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Nair VD, Ge Y, Li S, Pincas H, Jain N,
Seenarine N, Amper MAS, Goodpaster BH, Walsh MJ, Coen PM and
Sealfon SC: Sedentary and trained older men have distinct
circulating exosomal microRNA profiles at baseline and in response
to acute exercise. Front Physiol. 11:6052020. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Fry CS, Kirby TJ, Kosmac K, McCarthy JJ
and Peterson CA: Myogenic progenitor cells control extracellular
matrix production by fibroblasts during skeletal muscle
hypertrophy. Cell Stem Cell. 20:56–69. 2017. View Article : Google Scholar
|
|
198
|
Ruan S, Du B, Dai C, Wang L, Sun W, Jiang
J, Tang Y, Zhang Y, Wang K and Wei Q: Cognitive impairment in
patients with middle cerebral artery severe stenosis or occlusion
is associated with hemodynamic abnormalities. BMC Neurol.
26:3662026. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Perera N, De Blasio MJ and Febbraio MA:
Harnessing the therapeutic potential of exercise in extracellular
vesicle-based therapy in metabolic disease associated
cardiovascular complications. Free Radic Biol Med. 226:230–236.
2025. View Article : Google Scholar
|
|
200
|
Tiwari S, Kumar V, Randhawa S and Verma
SK: Preparation and characterization of extracellular vesicles. Am
J Reprod Immunol. 85:e133672021. View Article : Google Scholar
|
|
201
|
Coumans FAW, Brisson AR, Buzas EI,
Dignat-George F, Drees EEE, El-Andaloussi S, Emanueli C, Gasecka A,
Hendrix A, Hill AF, et al: Methodological guidelines to study
extracellular vesicles. Circ Res. 120:1632–1648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Brahmer A, Neuberger EWI, Simon P and
Krämer-Albers EM: Considerations for the analysis of small
extracellular vesicles in physical exercise. Front Physiol.
11:5761502020. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Darragh IAJ, O'Driscoll L and Egan B:
Exercise training and circulating small extracellular vesicles:
Appraisal of methodological approaches and current knowledge. Front
Physiol. 12:7383332021. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Karvinen S, Korhonen TM, Sievänen T,
Karppinen JE, Juppi HK, Jakoaho V, Kujala UM, Laukkanen JA, Lehti M
and Laakkonen EK: Extracellular vesicles and high-density
lipoproteins: Exercise and oestrogen-responsive small RNA carriers.
J Extracell Vesicles. 12:e123082023. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Greening DW, Rai A and Simpson RJ:
Extracellular vesicles-An omics view. Proteomics. 24:e24001282024.
View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Wijerathna-Yapa A, Aubert D and Kulasinghe
A: Integrating mass spectrometry-based multi-omic signatures of
extracellular vesicles: from discovery to clinical translation.
Clin Transl Immunology. 15:e700782026. View Article : Google Scholar : PubMed/NCBI
|














