Introduction
Radiotherapy (RT) is a fundamental modality in the
clinical management of cancer, with more than half of all patients
receiving RT during their treatment course (1). The efficacy of RT primarily stems
from the generation of reactive oxygen species (ROS) by ionizing
radiation (IR)-induced water radiolysis, a process responsible for
the majority of cellular damage (2). Given that water constitutes ~80% of
the cellular content, this mechanism leads to significant oxidative
stress, making intracellular ROS levels a critical determinant of
tumor radiosensitivity (3). High
levels of ROS damage critical cellular components, including
proteins, nucleic acids, lipids, membranes and organelles. The
'oxidative stress' cancer biology theory highlights the critical
role of the antioxidant defense system in counteracting this
process (4). This theory
elucidates how tumor cells upregulate the activity of antioxidant
systems such as glutathione (GSH) to prevent ROS from exceeding a
lethal threshold. Bansal and Simon (5) further emphasized that this adaptive
redox homeostasis, governed largely by GSH, is a key factor in
tumor radioresistance.
A critical determinant of radioresistance is the
unregulated redox homeostasis in tumor cells, with GSH metabolism
playing a central role. GSH, a tripeptide composed of glutamate,
cysteine and glycine, is the most abundant intracellular
antioxidant and is synthesized across different cell types
(6). Its primary role is to
maintain redox balance by scavenging free radicals, thereby
protecting cells from damage induced by ROS (7). Notably, compared with normal cells,
tumor cells inherently produce more GSH to support their aberrant
proliferation (8). This notable
increase in GSH is driven by the upregulated uptake of precursors
and increased biosynthesis (9).
It is a common feature across multiple malignancies, including
breast, colon and lung cancer, as well as leukemia, and is closely
linked to tumor initiation, progression, metastasis and therapeutic
resistance (10).
Despite its success in certain malignancies, direct
tumor targeting faces inherent limitations, including intratumoral
heterogeneity, the rapid emergence of acquired resistance and a
reliance on specific molecular markers that are not universally
present across all cancer types (11). By contrast, elevated GSH
metabolism represents a shared adaptive hallmark across a wide
spectrum of tumors, driven by common stress responses to
proliferation-associated oxidative stress (12). This universality makes GSH
metabolism an attractive broad-spectrum vulnerability that
circumvents the need for individualized molecular targets.
Furthermore, elevated GSH levels are intimately linked to
radioresistance, as GSH efficiently scavenges radiation-induced ROS
(13,14). Therefore, targeting GSH
metabolism is not intended to replace radiotherapy but rather to
resensitize tumor cells to radiation by disabling their primary
antioxidant defense. Thus, targeting GSH metabolism is a
broad-spectrum, resistance-resilient adjuvant strategy that
resensitizes tumors to radiotherapy by dismantling their primary
antioxidant defense. High intracellular GSH levels in the tumor
microenvironment (TME) directly undermine RT efficacy (15). The ROS generated by IR are
effectively neutralized by GSH, thereby diminishing oxidative
damage and allowing tumor cell survival even in the presence of
radiation (16). Thus, targeting
GSH metabolism represents a promising strategy for overcoming tumor
resistance to IR-induced cell death. By promoting GSH depletion or
inhibiting its synthesis, tumor cells lose the ability to buffer
IR-induced ROS, amplifying oxidative damage (17).
This review explores the multifaceted role of GSH in
tumor radioresistance and critically evaluates the evolving
strategies for targeting GSH metabolism to achieve tumor
radiosensitization. As the field has advanced, a clear
developmental trajectory has emerged: From the first generation of
crude small-molecule inhibitors that systemically dampen GSH
synthesis to the second generation of passively targeted
nanomaterials that improve tumor accumulation via the EPR effect
and, finally, to the ongoing third generation of integrated
platforms. This review will dissect this evolutionary path,
highlighting how each generation has built upon the last generation
to overcome previous limitations and offering a perspective on the
future of innovative GSH-targeting radiosensitizers. Fig. 1 provides an abstract schematic
overview of emerging strategies that target GSH metabolism to
counteract radioresistance by amplifying cellular oxidative stress
and sensitizing tumor cells to radiotherapy.
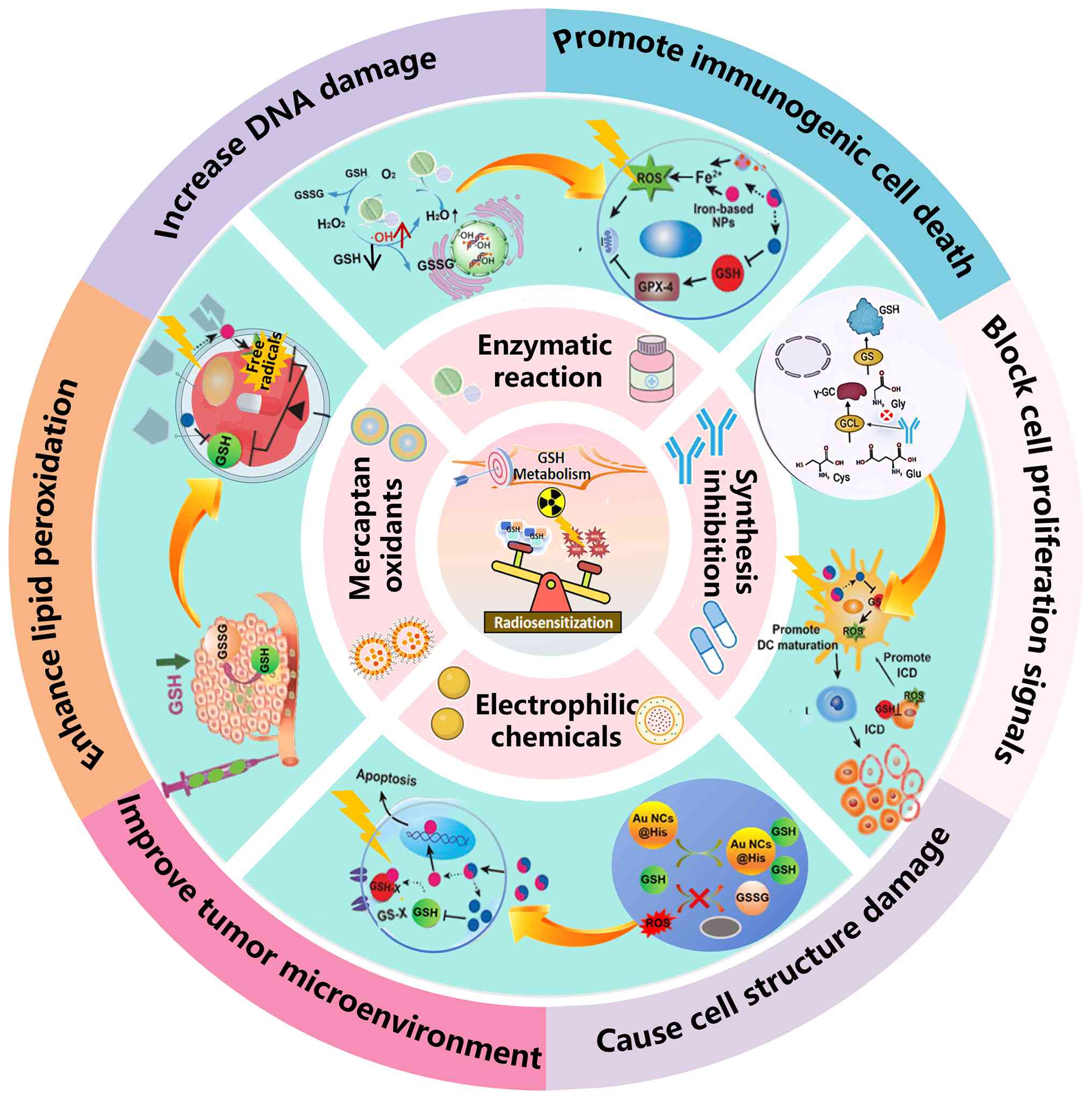 | Figure 1Abstract schematic diagram of
targeting GSH metabolism as radiosensitization strategy. The
emerging strategies are designed to counteract radioresistance by
targeting GSH metabolism. They efficiently amplify cellular
oxidative stress and sensitize tumor cells to radiotherapy. GSH,
glutathione; ROS, reactive oxygen species; NPs, nanoparticles; DNA,
deoxyribonucleic acid; GSSG, oxidized glutathione; GS-X,
glutathione polymer; GPX-4, glutathione peroxidase 4; Cys,
cysteine; Gly, glycine; Glu, glucose; GCL, glutamate cysteine
ligase; GS, glutathione synthetase; DC, dendritic cell; ICD,
immunogenic cell death. |
Tumor GSH metabolism and targeting
strategies
GSH metabolism in tumor cells
GSH is the most abundant intracellular antioxidant
and plays a critical role in maintaining cellular redox homeostasis
(18). In normal cells, GSH
helps preserve cysteine levels and detoxify xenobiotics. In tumor
cells, however, metabolic dysregulation contributes to resistance
to therapy, including radioresistance (19). A hallmark of malignant tumor
cells is elevated GSH levels. GSH levels in tumor cells are
typically 4- to 10-fold higher than those in normal cells (20). GSH is usually synthesized in two
ATP-dependent steps. The rate-limiting step, catalyzed by
glutamate-cysteine ligase (GCL), produces γ-glutamylcysteine, which
is then converted to GSH by GSH synthetase (GS) (21). Its antioxidant function is
maintained through a continuous cycle: GSH peroxidase 4 (GPX4) uses
it to neutralize lipid peroxides, generating oxidized glutathione
(GSSG), which is then reduced back to GSH by GSH reductase (GR) at
the expense of NADPH. This process safeguards tumor cells from
IR-driven oxidative damage (22).
In GSH metabolism, cystine is tightly coupled to GSH
synthesis. Cystine, a disulfide-bonded dimer of cysteine, is a
critical GSH precursor (23).
Its uptake is mediated by the cystine/glutamate antiporter (xCT).
This transporter maintains redox balance and supports GSH synthesis
by exchanging extracellular cystine for intracellular glutamate
(24). Intracellular GSH reduces
the disulfide bond of cystine and generates cysteine in tumor cells
(25). Cysteine is a substrate
for the GCL- and GS-related reactions involved in GSH synthesis.
This interdependence forms a feedback loop. In this loop, GSH
modulates the membrane redox environment and enhances cystine
uptake. The increased uptake further fuels the synthesis of GSH.
Additionally, the reduction in GSSG levels by GR sustains GSH
levels while cysteine is recycled, reinforcing cellular defenses
against oxidative stress (26,27). Elevated GSH levels in tumor cells
contribute to radioresistance by enhancing the scavenging of ROS,
thereby compromising treatment efficacy (28). Therefore, targeting GSH
metabolism represents a promising strategy to improve RT outcomes
(29).
GSH metabolism-related radioresistance
mechanisms
RT exerts its cytotoxic effects primarily through
IR-induced ROS, which disrupt redox homeostasis and cause lethal
oxidative damage to cellular components. However, the TME exhibits
dynamic adaptability in response to RT, activating multiple
signalling pathways that contribute to radioresistance (30). The elevated GSH metabolism in
tumor cells acts as a critical defense against this assault. In
addition to its fundamental role in scavenging ROS, GSH actively
orchestrates a network of pro-survival signalling pathways. This is
achieved primarily through S-glutathionylation, a key
posttranslational modification that modulates the redox state of
critical cysteine residues in signalling proteins (31).
Several intrinsic signalling pathways and their
associated proteins critically influence cellular radiosensitivity
(Fig. 2). The forkhead box
(Fox)O and p53 pathways are critically involved in maintaining
redox balance under radiation stress (32). FoxO activation enhances cystine
uptake and GSH synthesis via the upregulation of xCT, whereas
elevated GSH levels subsequently fine-tune the activity of p53 and
its apoptotic targets (33,34). This interplay highlights the role
of GSH in regulating stress response pathways. Notably, p53
activation itself is partially ROS dependent and influences RT
outcomes through caspase-mediated apoptosis (35,36).
In addition to intracellular signalling, GSH
metabolism provides multilayered protection against
radiation-induced damage (37).
At the structural level, it directly safeguards membrane and
cytoskeletal integrity by maintaining thiol groups in a reduced
state, while GPX4 utilizes GSH to suppress lipid peroxidation
(38,39). Adaptively, cells can upregulate
xCT and GPX4 expression to reinforce this defense under oxidative
stress (40,41). Simultaneously, GSH plays a role
in a coordinated transcriptional response (42). It activates Nrf2 to increase
antioxidant gene expression (43-45) but conversely suppresses NF-κB
signalling through S-glutathionylation and stabilization of
inhibitor of NF-κB (IκB)α, thereby limiting its nuclear
translocation and survival output (46,47). Experimental evidence confirms
that GSH depletion disrupts IκB kinase activity and NF-κB
activation (48,49). This interconnected regulatory
network significantly influences RT efficacy.
Additionally, GSH supports the Hippo-Yes-associated
protein (YAP)1 signalling axis by maintaining S-glutathionylation
(50). YAP1 can enhance DNA
repair by upregulating the expression of factors such as X-ray
repair cross complementing 4 and promoting nonhomologous end
joining (51). In multiple
cancer types, Hippo pathway activation restrains YAP1 nuclear
translocation and transcriptional output, thereby reducing the
expression of DNA repair and antiapoptotic genes (52). Consequently, the GSH-mediated
maintenance of Hippo-YAP1 signalling reduces cellular
susceptibility to RT, highlighting another mechanism through which
GSH supports tumor cell survival under oxidative stress.
Strategies to modulate tumor GSH
metabolism
Recent advances in tumor RT research have enabled
more precise targeting of GSH metabolism to improve therapeutic
efficacy and reduce off-target effects (53). These strategies can be broadly
divided into two complementary methods. One is the inhibition of
intracellular GSH biosynthesis, which disrupts the production of
new GSH by targeting key synthesis enzymes or precursor supply
pathways. The other is the depletion of existing intracellular GSH,
which directly reduces preformed GSH reserves through oxidative or
conjugation-mediated mechanisms (Fig. 3). By making rational use of these
strategies, researchers aim to overcome radioresistance and
significantly increase oxidative stress in tumor cells (54).
 | Figure 3Targeting GSH metabolism for
radiosensitization. GSH metabolism supports direct free radical
scavenging by maintaining a reducing environment. This antioxidant
role underlies radioresistance, and targeting GSH synthesis and
utilization in radiotherapy elevates intracellular ROS by impairing
this protective function. GSH, glutathione; ROS, reactive oxygen
species; GSH, glutathione; ROS, reactive oxygen species; Cys,
cysteine; Gly, glycine; Glu, glucose; GCL, glutamate cysteine
ligase; GSS, glutathione synthetase; GGT, gamma-glutamyl
transferase; DDT, deoxynucleotidyl transferase; ER, endoplasmic
reticulum. |
Targeting key enzymes or transporters involved in
precursor supply is a common way to inhibit intracellular GSH
biosynthesis. For example, the natural compound oleanolic acid
inhibits GCL activity, thereby blocking the synthesis of
γ-glutamylcysteine and ultimately mediating radiosensitization
(55). Similarly, inhibiting GS
prevents the formation of GSH itself. The key transporter target is
xCT, as mentioned above (56).
Small molecules such as erastin impair cystine uptake through this
transporter, which depletes cysteine pools and limits GSH
production (57). Additionally,
inhibition of GSH regeneration pathways is an effective synthesis
inhibition strategy. For instance, the use of glycolysis inhibitors
can reduce NADPH availability. This strategy hinders the
GR-mediated reduction of GSSG to GSH, thereby disrupting GSH
recycling (58).
While inhibiting GSH synthesis is one viable
strategy, a complementary and more rapid approach is to directly
deplete intracellular GSH reserves. The depletion of existing
intracellular GSH can directly reduce preformed GSH reserves,
thereby amplifying tumor-cell oxidative stress. This approach
primarily employs two methods (59). One method is oxidative depletion,
in which nanoparticles (NPs) containing disulfide bonds (S-S) or
metal ions (such as copper or iron) oxidize GSH to GSSG. The
resulting GSSG is then either exported from the cell or cannot be
efficiently regenerated (60).
The other method is conjugation-mediated efflux. This process
promotes the conjugation of GSH with xenobiotics via GSH
S-transferases. The conjugated products are subsequently extruded
by transporters such as multidrug resistance-associated protein 1,
reducing intracellular GSH levels (61). However, a single approach
involving GSH depletion typically leads to only a transient
reduction in GSH levels, as tumor cells compensate by upregulating
synthesis pathways. Therefore, a rational combination of
biosynthesis inhibition and depletion strategies is essential for
achieving sustained GSH suppression, effectively overcoming this
adaptive resistance and maximizing tumor radiosensitization
(62).
Targeting GSH synthesis pathways for tumor
radiosensitization
Given that numerous tumors exhibit enhanced GSH
biosynthesis to maintain redox homeostasis, targeting its synthesis
pathways provides a direct strategy to compromise the antioxidant
defense at its source (63).
This section systematically reviews key molecular targets within
the GSH synthesis pathway and the corresponding pharmacological or
nanomaterial-based agents designed to disrupt them for effective
tumor radiosensitization (Fig.
4).
![Inhibiting GSH production for tumor
radiosensitization. Targeting key enzymes or transporters involved
in precursor supply is a common way for intracellular GSH
biosynthesis inhibition and has shown promise in radiotherapy.
[created with Biorender (biorender.com)]. GSH, glutathione; IR, ionizing
radiation; BSO, buthionine sulfoximine; xCT, cystine/glutamate
antiporter; Cys, cysteine; Gly, glycine; Glu, glucose; Gln,
glutamine; GCL, glutamate cysteine ligase; GSS, glutathione
synthetase; GR, glutathione reductase; ATP, adenosine triphosphate;
ADP, adenosine diphosphate; 2-DG, 2-deoxy-D-glucose; 2-AAPA,
2-acetylamino-3-[4-(2-acetylamino-3-carboxypropylsulfanylthiocarbonylamino)phenylthiocarbamoylsulfanyl]propionic
acid; SNAP, S-nitroso-N-acetylpenicillamine.](/article_images/ijmm/58/2/ijmm-58-02-05892-g03.jpg) | Figure 4Inhibiting GSH production for tumor
radiosensitization. Targeting key enzymes or transporters involved
in precursor supply is a common way for intracellular GSH
biosynthesis inhibition and has shown promise in radiotherapy.
[created with Biorender (biorender.com)]. GSH, glutathione; IR, ionizing
radiation; BSO, buthionine sulfoximine; xCT, cystine/glutamate
antiporter; Cys, cysteine; Gly, glycine; Glu, glucose; Gln,
glutamine; GCL, glutamate cysteine ligase; GSS, glutathione
synthetase; GR, glutathione reductase; ATP, adenosine triphosphate;
ADP, adenosine diphosphate; 2-DG, 2-deoxy-D-glucose; 2-AAPA,
2-acetylamino-3-[4-(2-acetylamino-3-carboxypropylsulfanylthiocarbonylamino)phenylthiocarbamoylsulfanyl]propionic
acid; SNAP, S-nitroso-N-acetylpenicillamine. |
Inhibiting synthetase activity
As the rate-limiting enzyme in GSH biosynthesis, GCL
is a prime target for inhibiting GSH production (64). Buthionine sulfoximine (BSO) is a
classic GCL inhibitor with excellent radiosensitizing effects
(65). At millimolar
concentrations, BSO reduces intracellular GSH levels to 40-50% of
control values. This GSH synthesis inhibition method has been shown
to achieve radiosensitization in various types of tumor cells,
including Chinese hamster V79 cells, as well as human lung, renal
and head and neck tumor cells, under both oxygenated and hypoxic
conditions (66). Notably, BSO
also synergizes with other radiosensitizers: Its combination with
the hypoxic radiosensitizer misonidazole yields enhanced antitumour
effects in multiple preclinical tumor models (67). The safety profile of BSO has been
evaluated in an early-phase clinical trial involving cancer
patients. A phase I dose-escalation study demonstrated that BSO
administered as infusions every 12 h was well tolerated (68).
In addition to classical enzyme inhibitors such as
BSO, various functional materials capable of suppressing GS have
also shown potential for depleting intracellular GSH. For instance,
Mayur et al (69)
investigated the cytotoxic mechanism of latex-capped silver NPs in
human lung carcinoma cells and reported a significant reduction in
intracellular GSH levels, attributable to the inhibition of
GSH-synthesizing enzymes. Continued efforts to develop potent and
specific GS inhibitors are therefore highly important for improving
RT efficacy. Despite its well-established preclinical efficacy, the
clinical application of BSO is limited by its modest potency,
requirement for millimolar concentrations and potential systemic
toxicity upon prolonged administration. Future efforts should focus
on developing more potent and selective GCL inhibitors, as well as
tumor-targeted delivery strategies to increase intratumoral
accumulation while minimizing off-target effects. Additionally,
combination regimens that pair GCL inhibitors with other
GSH-depleting modalities may achieve synergistic
radiosensitization.
Inhibiting synthesis substrate
uptake
An adequate amino acid supply is a fundamental
prerequisite for GSH biosynthesis. Impairment in the transport of
extracellular amino acids essential for GSH synthesis disrupts the
production of GSH, leading to reduced intracellular GSH levels
(70). Among these factors,
cysteine availability is particularly critical. Cysteine starvation
has thus emerged as a common strategy to deplete GSH, primarily
through the regulation of xCT ability. This transporter mediates
the uptake of extracellular cystine, which is subsequently reduced
to cysteine intracellularly to support both protein synthesis and
GSH synthesis (71).
Consequently, disruption of xCT function blocks cystine uptake and
reduces intracellular GSH production. Growing evidence indicates
that regulating xCT represents a promising approach for tumor
radiosensitization.
Sulfasalazine (SAS) exerts anticancer effects by
specifically inhibiting xCT function. In glioma cells, SAS-mediated
GSH depletion increases ROS accumulation, DNA damage and subsequent
cell death. When combined with radiation, SAS enhances antitumour
efficacy and improves survival in glioblastoma xenograft models
(72). However, its clinical
translation still requires balancing efficacy and safety. A
clinical trial evaluating this combination (NCT00753038) reported a
high discontinuation rate of SAS due to hematologic toxicity, with
no clear clinical benefit observed (73). Fortunately, as a safer xCT
inhibitor, sorafenib has achieved good results in clinical trials
(CTR2200066117). This combination of sorafenib and RT was superior
to RT alone (74). Further
optimization of dosing or delivery strategies may be needed to
mitigate drug toxicity and side effects. The new compound DC10
formed by scaffold jumping and structural optimization of SAS
performed better in terms of side effects and bioavailability.
Sarowar et al (75)
reported that DC10 is well tolerated and acts as a radiosensitizer
by inhibiting cystine uptake. While inhibition of xCT represents a
rational strategy for reducing the effects of GSH, its clinical
translation is limited by the narrow therapeutic window of existing
inhibitors such as SAS, which exhibit dose-limiting hematologic
toxicity. Furthermore, compensatory upregulation of alternative
cysteine acquisition pathways may limit the durability of
xCT-targeting approaches. Future directions include the development
of isoform-selective xCT inhibitors with improved safety profiles
and the exploration of combination strategies that simultaneously
target multiple cysteine uptake routes to overcome adaptive
resistance.
Inhibiting regeneration reactions
When GSH is oxidized to GSSG during antioxidant
reactions, it simultaneously catalyzes its reduction back to GSH.
This recycling mechanism not only maintains intracellular GSH pools
but also facilitates cysteine regeneration. Targeting this
GR-mediated cycle has emerged as a viable therapeutic approach to
induce tumor cell death. As evidenced by
2-acetylamino-3-[4-(2-acetylamino-3-carbo
xypropylsulfanylthiocarbonylamino)phenylthiocarbamoylsulfanyl]propionic
acid, a specific GR inhibitor, suppression of GR activity induces
oxidative stress that effectively inhibits melanoma metastasis and
synergistically enhances the efficacy of chemotherapeutic agents
against human glioblastoma cells (76). Furthermore, inhibition of NADPH
is essential for maintaining cellular GSH homeostasis, as it serves
as a cofactor for the GR-mediated conversion of GSSG back to GSH.
Agents that deplete NADPH can thus indirectly inhibit GSH
regeneration and increase radiosensitivity (77). For instance, the natural
isoflavone genistein has been shown to decrease NADPH reserves in
multiple tumor cell types, resulting in significant GSH depletion
and increased susceptibility to radiation (78). This effect is partly attributed
to the interference of genistein with NADPH-producing pathways,
such as the pentose phosphate pathway, although the precise
underlying mechanisms remain unclear (79). In addition to natural compounds,
pharmacological inhibitors targeting key enzymes involved in NADPH
generation (e.g., glucose-6-phosphate dehydrogenase) also exhibit
promising radiosensitizing properties by limiting the reducing
equivalents required for GR activity (80).
Furthermore, combination strategies that
simultaneously target GR activity and the supply of NADPH may yield
synergistic effects. For example, compared with single-target
approaches, the use of GR inhibitors alongside compounds that
induce NADPH consumption could exhaust redox resilience more
comprehensively. Notably, it is also worth noting that IR itself
can alter cellular metabolism and NADPH availability, suggesting a
potential positive feedback loop when combined with metabolic
inhibitors (81).
Despite promising preclinical results, the
translation of NADPH/GR-targeting strategies into clinical practice
requires careful consideration of selectivity and toxicity, given
the role of NADPH in normal cellular functions (82). Future efforts should focus on
developing tumor-specific delivery systems to minimize off-target
effects. Additionally, identifying biomarkers of NADPH pathway
activation may help select patients most likely to benefit from
such radiosensitization approaches (83). Thus, targeting the GR pathway
represents a rational and evolving strategy to overcome
radioresistance rooted in antioxidant adaptation.
Inhibiting energy supply
The inhibition of ATP production represents a viable
strategy for suppressing the energy supply for GSH synthesis
because both steps of GSH biosynthesis are ATP-dependent (84). 2-Deoxyglucose (2-DG), a glucose
analogue, functions as a glycolytic inhibitor by targeting
hexokinase. Upon phosphorylation by hexokinase, 2-DG is converted
into 2-DG-phosphate, which cannot be further metabolized by
phosphoglucose isomerase. This results in the intracellular
accumulation of 2-DG-phosphate and the subsequent depletion of ATP
(85). In preclinical models of
cervical cancer and pancreatic tumors, 2-DG has been shown to
increase radiosensitivity through mechanisms linked to metabolic
oxidative stress and disruption of thiol metabolism (86). Importantly, the combination of
hypofractionated RT with orally administered 2-DG (200-300 mg/kg)
has demonstrated good tolerability in patients with glioblastoma
(87). Notably, 2-DG may also
mitigate radiation-induced damage to normal tissues, thereby
contributing to an improved quality of patients' life.
In addition to direct ATP inhibition, innovative
approaches have been developed to simultaneously disrupt energy
metabolism and redox balance. Recently, Qiao et al (88) constructed a liposomal
nanoplatform loaded with the nitric oxide (NO) prodrug
S-nitroso-N-acetylpenicillamine (SNAP). Following systemic
administration, the elevated GSH levels in the TME promote NO
release from SNAP. The released NO rapidly reacts with superoxide
anions generated during RT to form peroxynitrite, a highly reactive
nitrogen species that induces severe oxidative damage and cell
death. Furthermore, when combined with cold exposure, this strategy
triggered systemic thermogenesis, which exacerbated tumor energy
starvation and led to a significant reduction in both ATP and GSH
levels within the tumor.
Collectively, the strategies used to inhibit GSH
synthesis, including targeting GCL, cystine uptake or
NADPH-dependent reduction pathways, effectively decrease
intracellular GSH levels and enhance radiotherapeutic efficacy
(Table I). Although the clinical
translation of certain inhibitors, such as SAS, is limited by
systemic toxicity, other agents, such as BSO and 2-DG, demonstrate
favorable safety and tolerability profiles, underscoring their
potential as radiosensitizers (89).
 | Table IApplication of inhibiting GSH
production in tumor radiosensitization. |
Table I
Application of inhibiting GSH
production in tumor radiosensitization.
| Targeting
strategies | Materials | Mechanism of
action | Experimental
model | (Refs.) |
|---|
| GSH synthetase
inhibition | Oleanolic acid | Attenuate GCL
activity | In vitro
(C6, A549) | Qi et al
(64) |
|
L-buthioninesulphoximine | Irreversibly
inhibits GCL | In vivo
(4T1) | Zeng et al
(67) |
| Latex capped silver
nanoparticles | Inhibits GSH
synthesizing enzyme | In vitro
(A549) | Mayur et al
(69) |
| Synthesis substrate
inhibition | Quercetin | Inhibits xCT | In vivo
(CT26) | Xu et al
(71) |
| Sulfasalazine | Downregulates
xCT | In vivo
(MDA-231-MB, 4T1) | Kerkhove et
al (72) |
| Sulfasalazine | Specifically
inhibits xCT | Phase I/II clinical
trial (NCT01577966) | Takeuchi et
al (73) |
| Sorafenib | Directly inhibits
xCT | Phase II single-arm
clinical trial (CTR2200066117) | He et al
(74) |
| DC10 | Specifically
inhibits xCT | In vivo
(A375) | Sarowar et
al (75) |
| GSH reductase
inhibition | 2-AAPA | Inhibits GSSG
recycled back to GSH by GR | In vitro
(TE-13, KYSE-450, KYSE-510) | Li et al
(76) |
| Temozolomide | Inhibits GSSG
recycled back to GSH by GR | In vivo
(U251) | Zhu et al
(77) |
| ATP inhibition | 2-Deoxyglucose | Inhibits glycolysis
and ATP production | In vivo
(G361) | Dwarakanath et
al (85) |
|
3-bromopyruvate | Disrupts oxidative
phosphorylation and glycolysis | In vivo
(CT26, 4T1) | Hasse et al
(86) |
| SNAP | Inhibits ATP and
decreases GSH | In vivo
(4T1) | Qiao et al
(88) |
| Platelet membrane
biomimetic nanomedicine | Inhibits glycolysis
and ATP production | In vitro
(MDA-MB-231, PANC-1) | Chen et al
(89) |
Promoting GSH depletion for tumor
radiosensitization
Direct depletion of intracellular GSH serves as a
rapid and effective strategy to disrupt redox homeostasis and
enhance tumor radiosensitivity. By diminishing the major cellular
antioxidant reservoir, this approach significantly amplifies
radiation-induced oxidative stress, leading to increased damage to
macromolecules and potentiated cancer cell death (90,91). To achieve efficient GSH
consumption, a variety of synthetic materials and nanoplatforms
have been developed (Table II),
which operate primarily through three core mechanistic pathways: i)
The oxidation of GSH to GSSG by leveraging its strong reducibility
(92); ii) the facilitation of
GSH conjugation with electrophilic molecules, often via glutathione
S-transferase-mediated pathways, resulting in the formation and
export of GS-X conjugates (93);
and iii) the acceleration of GSH depletion through enzyme-mimetic
or catalytic reactions (94).
This section reviews the design principles and mechanisms
underlying these distinct GSH-depleting strategies and explores
their application in tumor radiosensitization (Fig. 5).
| Table IIApplication of promoting GSH
depletion in tumor radiosensitization. |
Table II
Application of promoting GSH
depletion in tumor radiosensitization.
| Targeting
strategies | Materials | Mechanism of
action | Experimental
model | (Refs.) |
|---|
| Chemical
oxidizing | GdW10@CS
nanosphere | Produces ROS and
depletes GSH | In vivo
(Hela, BEL-7402) | Yong et al
(96) |
| Cisplatin | Oxidizes the
endogenous GSH | In vivo
(Hela, 3T3) | Luo et al
(97) |
|
BiP5W30
nanoclusters | Oxidizes the
endogenous GSH | In vivo
(HeLa) | Zhou et al
(98) |
|
Bi2WO6-BP | Oxidizes the
endogenous GSH | In vivo
(MDA-231-MB, 4T1) | Ren et al
(99) |
|
Cu2-xSe | Produces ROS and
depletes GSH | In vivo
(4T1) | Chen et al
(100) |
| Disulfide
bonds | Cleaves GSH via
redox reaction | In vivo
(Hela) | Liu et al
(101) |
| Propyl gallate | Produces ROS and
depletes GSH | In vitro
(4T1) | Fu et al
(102) |
| Pt (IV)
prodrugs | Oxidizes the
endogenous GSH | In vivo
(A2780) | Ling et al
(103) |
| Covalent
conjugation | Asc-s | Binds of Asc-s with
GSH's thiols structure | In vitro
(EL4) | Mane et al
(105) |
| AuNCs@His | Conjugated with GSH
via Au-S bond | In vivo
(U14) | Zhang et al
(106) |
| AuNCs-based
biohybrid system | Conjugates with GSH
via Au-S bond | In vivo
(A549, 4T1) | Hua et al
(107) |
| PL self-assembled
polymeric NPs | Electrophilically
attacks and bonds with GSH | In vivo
(CT26) | Wang et al
(109) |
| Enzymatic
reaction | Polyaniline
inorganic-organic nanohybrids | Catalyzes GSH
oxidation through organic hydroperoxides | In vivo
(4T1) | Wang et al
(113) |
| Iron oxide
nanozymes | Catalyzes
Fenton-like reactions | In vitro
(MCF-7) | Liu et al
(114) |
| Cerium oxide
nanozymes | Increases GSH
conversion | In vitro
(A549) | Liu et al
(115) |
|
MoS2-based Nanoflowers | Enhances
self-cascade catalysis | In vivo
(Hela) | Wu et al
(117) |
| Mn-based
nanosystems | Increases GSH
conversion | In vivo
(4T1) | Pan et al
(118) |
| Au@Mn-MOF | Increases hydroxyl
radical levels | In vivo
(4T1) | Xiong et al
(119) |
| Cuhemin-Au | Reduces GSH via
multienzyme-like catalytic reactivity | In vivo
(4T1) | Qiao et al
(124) |
| Copper-based
single-atom nanozyme | Increases GSH
conversion | In vivo
(4T1) | Chen et al
(127) |
Promoting chemical oxidation
Leveraging the reducing property of GSH, a key
strategy to enhance RT efficacy is to use chemical substances that
deplete intracellular GSH via oxidation-reduction reactions,
thereby amplifying oxidative stress and limiting the ability of
tumor cells to scavenge ROS (95). For example, some high-atom-number
(high-Z) element nanomaterials are good oxidants. High-Z elements,
such as bismuth, gold and gadolinium, exhibit strong X-ray
attenuation and photoelectric effect, making them effective
radiosensitizers by enhancing local radiation energy deposition.
Zhao's group developed gadolinium-containing
polyoxometalate-conjugated chitosan (GdW10@CS) NPs via
ionotropic gelation, designed to mitigate radioresistance. Upon
X-ray IR, the GdW10@CS NPs serve as exogenous
radiosensitizers, inducing significant production of ROS (96). Notably, the oxidative tungsten
(VI) species released from these NPs reacts with intracellular GSH
in tumor cells, reducing ROS scavenging and thereby markedly
improving RT effects. Additionally, the reaction between the
released species and GSH is rapid and efficient, ensuring that the
radiosensitizing effects are maximized during the short window of
X-ray exposure (97).
To achieve a more precise RT, the same group
recently integrated bismuth heteropolytungstate
(BiP5W30) nanoclusters with reduced graphene
oxide (98). This system
provides a TME-responsive mechanism. An acidic TME can trigger the
cycle conversion of W from W5+ to W6+, and
the redox reaction between W6+ and GSH results in the
reversed activity of GSH in tumor cells and increases the
production of ROS (99).
Cu2-xSe NPs have recently garnered significant research
interest owing to their various advantages: They possess excellent
GSH oxidase-like activity and their constituent elements, copper
and selenium, are essential trace elements for the human body
(100).
In parallel, S-S provides another widely exploited
strategy to deplete GSH. These bonds can be cleaved by GSH via a
redox reaction, generating sulfhydryl groups and GSSG. This
GSH-consuming reaction is typically harnessed in redox-responsive
drug delivery systems for controlled drug release within tumor
cells with high levels of GSH (101). On the basis of this mechanism,
it is reasonable to hypothesize that a sufficient density of S-S
within a material could effectively induce intracellular GSH
depletion. A prominent strategy to incorporate substantial amounts
of S-S into nanocarriers is through framework doping, wherein S-S
is preintegrated into molecular building blocks prior to NP
assembly (102). For instance,
Ling et al (103)
constructed NPs using disulfide-rich polymers, which demonstrated
effective GSH scavenging and enhanced RT efficacy. Despite these
advances, a critical question remains unresolved: A large number of
S-S need to be present to meet the demand for depleting
intracellular GSH. The destruction of tetrasulfide bonds depletes
GSH, which is more effective than S-S. Thus, tetrasulfide bond
frameworks may be a better choice for radiosensitization
materials.
Promoting covalent conjugation
The covalent conjugation of GSH, a pivotal
antioxidant in tumor cells, has emerged as a sophisticated strategy
to disrupt redox homeostasis and potentiate anticancer therapies.
This approach exploits the nucleophilic thiol group of GSH to form
irreversible adducts with electrophilic agents, achieving dual
objectives: Permanent GSH depletion and generation of bioactive
conjugates with intrinsic cytotoxicity (104). For instance, Mane and Kamatham
(105) examined the
radiosensitizing effect of Ascorbyl stearate (Asc-s) in murine
T-cell lymphoma cells. They reported that Asc-s significantly
decreased the GSH/GSSG ratio and enhanced the RT effect through
binding with the thiol structure in GSH.
Gold nanoclusters (AuNCs) have shown considerable
promise as radiosensitizers in preclinical studies, with
evaluations at clinically relevant radiation energy levels. Zhang
et al (106) developed a
histidine-capped gold nanoclusters (AuNCs@His) to achieve
radiosensitization via intracellular GSH depletion. This is
achieved by the formation of Au-S bonds. Treatment with AuNCs@His
(at a concentration of 100 μM) reduced the intracellular GSH
concentration from 1.86 to 0.42 mM. The sensitization enhancement
ratio (SER) was ~1.54. This SER value is significantly higher than
that of most metal-based radiosensitizers. As another typical
example, Hua et al (107) designed an AuNCs-based biohybrid
system using covalent conjugation to deplete intracellular GSH.
Owing to their small size, AuNCs exhibit prolonged tumor retention
and efficient renal clearance and several AuNCs are currently under
evaluation in clinical trials for cancer treatment applications
(NCT02680535, NCT01270139) (108).
In addition to metal-based radiosensitizers, Wang
et al (109) reported
self-assembled polymeric NPs coloaded with the GSH-depleting agent
piperlongumine (PL). In vivo antitumour studies confirmed
that PL electrophilic attack-mediated GSH depletion potently
enhances the RT effect via covalent conjugation of GSH,
irreversibly exhausting antioxidant reserves. Compared with their
metal-based counterparts, polymer-based nanoradiosensitizers are
generally considered to offer superior biocompatibility and greater
potential for clinical translation, although their comparative
advantages require validation in clinical settings (110).
Promoting catalytic reaction
In addition to directly reducing GSH by oxidizing
materials, accelerating GSH oxidation by special materials with
corresponding catalytic activity is another effective approach to
indirectly promote GSH depletion (111). For example, nanozymes have
diverse catalytic properties. They have emerged as promising tools
for depleting intracellular GSH in tumors, thereby enhancing the
effects of RT by amplifying radiation-induced oxidative stress
(112). Nanozymes leverage
their catalytic activities to promote GSH depletion and disrupt
tumor cell antioxidant defenses, thereby serving as effective
radiosensitizers in cancer therapy (113). Various nanozymes have been
extensively investigated and shown to exhibit therapeutic potential
through GSH depletion and ROS modulation.
Iron oxide NPs (IONPs) have attracted significant
attention because they can catalyze Fenton-like reactions. These
reactions convert H2O2 into highly reactive
hydroxyl radicals (•OH). The generated •OH causes direct oxidative
damage to tumor cells and oxidizes GSH into GSSG (114). Complemented by their excellent
biocompatibility and surface functionalization capacity, IONPs
enable targeted delivery to tumor sites. This leads to a marked
reduction in intracellular GSH levels. Additionally, cerium oxide
NPs (CONPs) are distinguished by their unique redox switching
capabilities, arising from the reversible valence transition
between Ce3+ and Ce4+. This property endows
CONPs with dual redox reactivity: Under oxidative stress conditions
in the TME, they efficiently catalyze the oxidation of GSH to GSSG
and promote ROS accumulation. Conversely, they scavenge excess ROS
in healthy tissues. This dual functionality makes CONPs ideal for
simultaneously enhancing tumor radiosensitivity and protecting
healthy tissues from radiation exposure (115).
Manganese-based nanozymes have recently emerged as
potent catalytic agents for depleting intracellular GSH (116). Through rational structural
design, these nanozymes can efficiently promote the oxidation of
GSH to GSSG. For instance, Wu et al (117) demonstrated that MnS2
functions as an effective catalyst to accelerate GSH oxidation. In
another study, Pan et al (118) developed a targeted
radiosensitizer by hybridizing Mn3O4 with the
metal-organic framework ZIF-8. The high surface area and porosity
of ZIF-8 enhanced reactant adsorption and transport, whereas its
integration with Mn3O4 strengthened the
interaction between manganese species and GSH in the TME, leading
to improved GSH depletion efficiency. Furthermore, Xiong et
al (119) synthesized a
heterojunction nanozyme named Au@Mn-MOF, where efficient electron
transfer within the heterostructure facilitated energy delivery to
catalytic active sites, thereby promoting the
Mn2+-catalyzed Fenton reaction.
Emerging strategies for tumor-targeted
and ROS-responsive GSH-depleting radiosensitizers
To facilitate clinical translation, next-generation
GSH-depleting radiosensitizers must achieve tumor-specific delivery
and spatiotemporally controlled activity to minimize systemic
toxicity. Recent advances in stimuli-responsive nanoplatforms have
opened promising avenues to address these challenges (120,121).
TME-responsive delivery systems exploit unique TME
features, such as acidic pH, elevated GSH levels or overexpressed
enzymes. Among these, GSH-responsive nanocarriers incorporating S-S
or tetrasulfide bonds are particularly attractive, as these
linkages are specifically cleaved by high intracellular GSH
concentrations in tumor cells, triggering payload release while
simultaneously consuming GSH (122). Notably, compared with
conventional disulfide bonds, tetrasulfide bonds exhibit higher GSH
depletion efficiency, making them superior candidates for
radiosensitization.
ROS-responsive prodrugs and nanozymes offer another
powerful tool, given that RT itself generates a burst of ROS.
ROS-responsive chemical moieties include arylboronic esters,
thioketals and proline oligomers (123). When integrated into
nanocarriers, these groups undergo structural cleavage upon ROS
exposure, enabling radiation-triggered drug release and creating a
positive feedback loop: Radiation generates ROS → ROS cleaves the
carrier → released GSH-depleting agent amplifies oxidative stress →
further ROS production → enhanced radiosensitization.
Dual-responsive systems (e.g., GSH/ROS or GSH/pH)
provide even greater precision. For example, heterojunction
nanozymes such as Cuhemin-Au nanosheets have been engineered to
simultaneously respond to TME cues and exogenous radiation,
resulting in both GSH depletion and catalytic ROS generation
(124). These platforms not
only consume GSH but also convert H2O2 into
highly toxic •OH, further exacerbating oxidative damage.
In summary, integrating tumor-targeting ligands,
GSH/ROS-responsive linkages and catalytic nanozymes into a single
platform represents a frontier in radiosensitizer design. Future
efforts should focus on simplifying material synthesis, validating
efficacy in large-animal models and establishing scalable
manufacturing processes to accelerate clinical adoption.
Conclusion
Tumor cells often exhibit elevated levels of GSH,
which is closely associated with the maintenance of redox
homeostasis, thereby promoting cell proliferation and protecting
against free radical-induced damage in numerous instances. On the
basis of this understanding, targeting the characteristically
elevated GSH levels in tumor cells for depletion represents a
viable therapeutic strategy. This approach works by disrupting
intracellular redox homeostasis, thereby inhibiting tumor cell
proliferation and reversing established radioresistance. The
efficacy of GSH-depleting interventions has been empirically
validated, with numerous studies demonstrating their ability to
suppress tumor cell growth and induce apoptosis. These collective
findings provide a solid rationale for the use of GSH elimination
as a method to improve therapeutic outcomes in RT (Fig. 6). In this review, the central
role of GSH in tumor radioresistance is elucidated and contemporary
strategies targeting GSH metabolism to improve RT efficacy are
explored. Elevated GSH levels in tumor cells scavenge
radiation-induced ROS, thereby mitigating oxidative damage and
sustaining survival. To counteract this adaptive mechanism,
interventions focusing on both the inhibition of GSH synthesis and
the depletion of existing pools have shown considerable promise.
Pharmacological agents, such as BSO, SAS and 2-DG, disrupt key
synthetic and regenerative pathways, while various
nanotechnology-mediated approaches (e.g., catalytic oxidation,
covalent conjugation and enzyme-mimetic reactions) enable spatially
controlled and efficient GSH consumption within the TME.
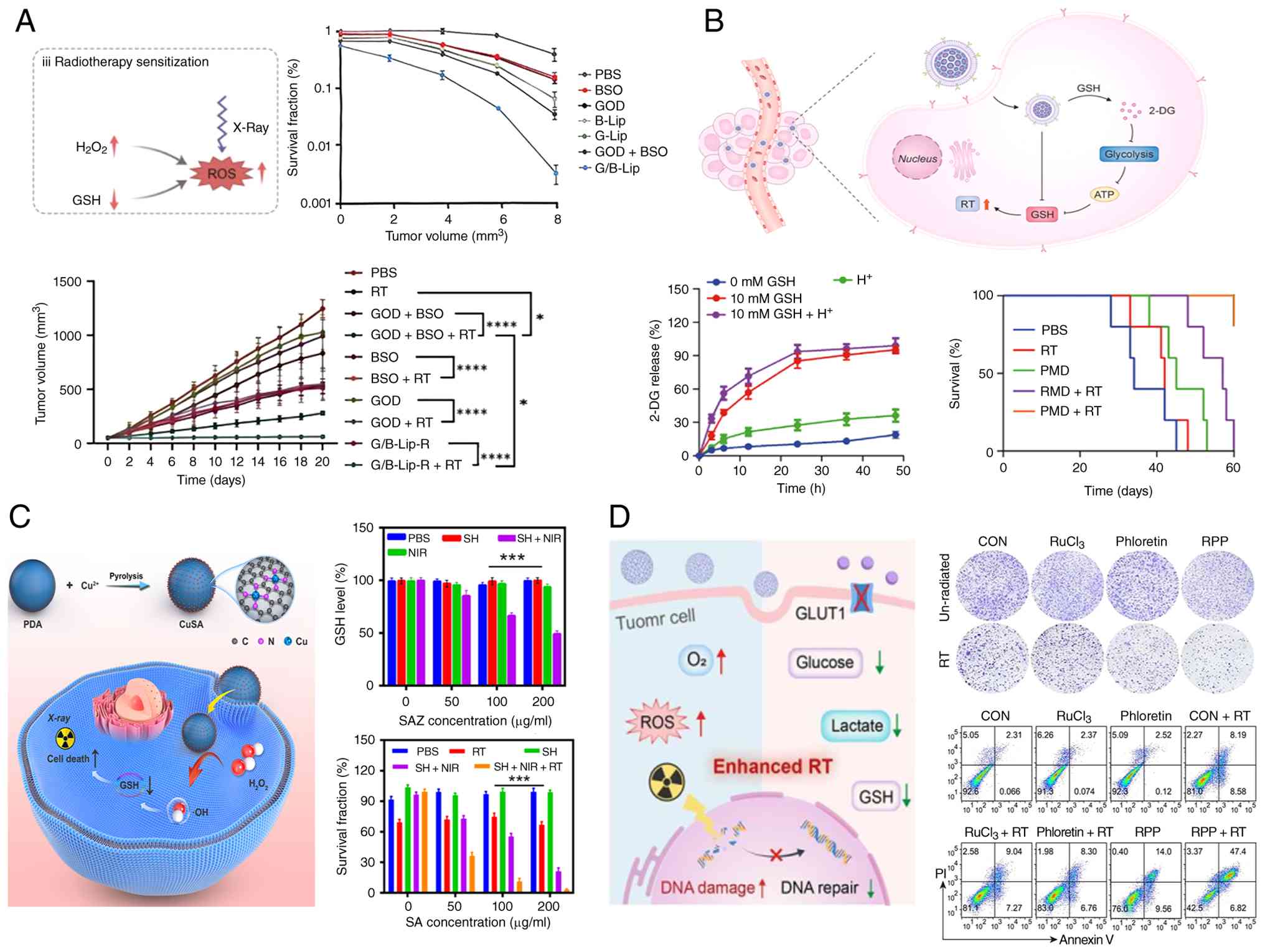 | Figure 6(A) Glucose and GSH synergistic
dual-starvation strategy for RT sensitization enhancement.
Copyright 2025 Elsevier. (B) Platelet membrane biomimetic
nanomedicine induces dual GSH consumption for enhancing cancer
radioimmunotherapy. Copyright 2024 Elsevier. (C) Copper single atom
nanozyme for GSH-depleted enhanced RT. Copyright 2024 IOP
Publishing. (D) RPP nanozyme with multienzyme activities and
glucose metabolism inhibition for radiosensitization. Copyright
2025 Elsevier. *P<0.05, ***P<0.001,
****P<0.0001. RT, radiotherapy; GSH, glutathione;
2-DG, 2-deoxy-D-glucose; BSO, buthionine sulfoximine; GOD, glucose
oxidase; ROS, reactive oxygen species; NIR, near infrared; PDA,
polydopamine; SA, single atom; SAZ, single atom nanozyme; ATP,
adenosine triphosphate; PMD, platelet membrane biomimetic
nanomedicine; CON, copper single atom nanozyme; GLUT1, glucose
transporter 1. |
Notably, concurrent chemoradiotherapy represents a
standard of care for many locally advanced cancers. The rationale
is that GSH metabolism plays an equally critical role in
chemoresistance, particularly against platinum-based agents (e.g.,
cisplatin and carboplatin) and alkylating drugs, through direct
drug conjugation and enhanced DNA repair (125). Preclinical studies have
demonstrated that GSH depletion using inhibitors such as BSO or
xCT-targeting agents can resensitize tumors to both chemotherapy
and radiotherapy. Therefore, an emerging and highly promising
direction is the triple combination of GSH-depleting strategies
with concurrent chemoradiotherapy, which could simultaneously
overcome chemo- and radioresistance. Although clinical validation
is still lacking, the mechanistic framework established in this
review provides a solid foundation for future investigations into
GSH-targeted chemoradiosensitization, ultimately broadening the
therapeutic scope beyond radiotherapy alone.
Despite significant advances in the development of
bioactive molecules and inhibitors for GSH depletion, several
challenges impede their clinical translation. First, the precise
mechanisms underlying the effects of GSH depletion-based therapies
remain incompletely elucidated. A deeper understanding of these
processes, facilitated by advanced biochemical technologies, is
essential to inform the rational design of more effective and
specific GSH-targeting strategies. Second, given the indispensable
role of GSH in maintaining cellular metabolism, off-target
depletion poses a substantial risk of damaging normal tissues and
potentially exacerbating conditions such as liver disease or cystic
fibrosis. It is therefore imperative to rigorously evaluate the
long-term biosafety of molecular and nanoscale GSH-depleting agents
before clinical application. Concurrently, developing delivery
systems with functional tumor-derived exosome modifications is
crucial for minimizing systemic toxicity (126). Third, many current
GSH-depleting systems rely on intricately designed molecules or NPs
with complex syntheses, which hinders their scalable production and
broad clinical adoption. Future efforts should therefore focus on
innovative and practical material designs. By simplifying the
material design and preparation process, copper-based single-atom
nanozymes can also achieve excellent sensitization effects
(127).
To accelerate clinical translation, several
promising directions merit particular attention. First, the
integration of predictive biomarkers, such as baseline intratumoral
GSH levels or GCL/xCT expression, could enable patient
stratification to identify those most likely to benefit from
GSH-targeting radiosensitization. Second, combination regimens that
pair GSH modulators with established immunotherapies (e.g., immune
checkpoint inhibitors) warrant systematic evaluation, given that
GSH-mediated redox regulation profoundly influences antitumour
immunity. Third, the development of stimuli-responsive
nanoplatforms that enable spatiotemporally controlled GSH depletion
within tumors while sparing normal tissues represents a critical
frontier. Advances in these areas, coupled with rigorous toxicity
assessment and scalable manufacturing processes, will be essential
to transition GSH-targeting radiosensitization from bench to
bedside as a clinically viable and personalized therapeutic
modality. Such improvements will be vital to advancing
GSH-targeting radiosensitization into clinically viable and
personalized anticancer therapeutics (Table III).
| Table IIILimitations and development prospects
of various GSH targeting strategies. |
Table III
Limitations and development prospects
of various GSH targeting strategies.
| Targeting
strategies | Categories | Limitations | Development
prospects |
|---|
| Promotion of GSH
depletion | Oxidizing
agents | Temporary
reduction, uncertain biosafety | Design of
sustained-release or cascade-reaction nanoplatforms to prolong GSH
depletion; incorporation of biodegradable or renal-clearable
materials to improve long-term biosafety. |
| Metal-based
nanomaterials | Quick elimination,
unavoidable damage, low cellular uptake | Surface engineering
with tumor-targeting ligands or biomimetic membranes to enhance
tumor accumulation and cellular internalization while minimizing
off-target damage. |
| Quinone
methide | Unstable structure,
short half-life | Develop prodrug
strategies or encapsulation within stimuli-responsive nanocarriers
to improve stability and extend half-life. |
| Nanozyme | Off-target
consequences | Engineering tumor
microenvironment-activated nanozymes that exert catalytic
GSH-depleting activity only within tumor tissues; integration with
imaging guidance for precise localization. |
| Electrophilic
reagents | Short
half-life | Conjugation to
self-assembling nanoplatforms or use of slow-release depots;
combination with radiotherapy-induced reactive oxygen species to
create positive-feedback activation loops. |
| Inhibition of GSH
biosynthesis | GCL inhibitors | Short
half-life | Encapsulating in
long-circulating nanoparticles to protect from rapid metabolism;
development of more selective GCL inhibitors via structure-based
drug design. |
| NADPH
inhibition | Unpredictable
effect | Integrating with
metabolomic profiling to identify predictive biomarkers for patient
stratification; combining NADPH-targeting agents with real-time
metabolic monitoring. |
| Cysteine
inhibition | Slow process,
uncertain efficiency | Combination with
rapid GSH-depleting agents to achieve synergistic and accelerated
effect; use of high-efficiency cysteine/glutamate antiporter
inhibitors with better pharmacokinetics. |
| ATP inhibition | Off-target
consequences | Exploit
tumor-specific metabolic vulnerabilities to design ATP-uncoupling
strategies that spare normal cells; design local delivery via
intratumoral injection or radiation-responsive prodrugs to restrict
systemic exposure. |
In summary, accumulating evidence suggests that a
rational combination of GSH metabolism-related radiosensitizers
with IR is an attractive approach to improve the tumor treatment
response. Research on GSH-depleting agents is advancing in several
areas, including their mechanisms of action, improved specificity
and biosafety, and simplified preparation. The discovery of other
methods to modulate cellular GSH further supports this progress.
Consequently, targeting GSH metabolism is an emerging
radiosensitization strategy with great potential for future
clinical applications.
Availability of data and materials
Not applicable.
Authors' contributions
All authors contributed to the study conception and
revision. The first draft of the manuscript was written by HX.
Literature search and data analysis were performed by MD and XL. ZC
and XX provided resources and critically revised the work. Data
authentication is not applicable. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
GSH
|
glutathione
|
|
ROS
|
reactive oxygen species
|
|
RT
|
radiotherapy
|
|
GCL
|
glutamate-cysteine ligase
|
|
GS
|
GSH synthetase
|
|
TME
|
tumor microenvironment
|
|
SS
|
disulfide bonds
|
|
Nrf2
|
nuclear factor-erythroid related
factor 2
|
|
Cys
|
cysteine
|
|
IR
|
ionizing radiation
|
|
2-DG
|
2-deoxyglucose
|
|
SER
|
sensitization enhancement ratio
|
|
AuNCs
|
gold nanoclusters
|
|
SAS
|
sulfasalazine
|
|
GSTs
|
GSH S-transferases
|
|
PL
|
piperlongumine
|
|
Asc-s
|
ascorbyl stearate
|
|
SNAP
|
S-nitroso-N-acetylpenicillamine
|
|
NO
|
nitric oxide
|
|
ATP
|
adenosine triphosphate
|
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant nos. 82572257 and 82272028), the Health
Research Project of Hunan Provincial Health Commission (grant no.
W20241010) and the Hunan Provincial Health High-Level Talent
Scientific Research Project (grant no. R2023010).
References
|
1
|
Citrin DE and Timmerman RD: Effects of
radiotherapy in normal tissue. N Engl J Med. 394:996–1009. 2026.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang RX and Zhou PK: DNA damage response
signaling pathways and targets for radiotherapy sensitization in
cancer. Signal Transduct Target Ther. 5:602020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jia S, Ge S, Fan X, Leong KW and Ruan J:
Promoting reactive oxygen species generation: A key strategy in
nanosensitizer-mediated radiotherapy. Nanomedicine (Lond).
16:759–778. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Trachootham D, Alexandre J and Huang P:
Targeting tumor cells by ROS-mediated mechanisms: A radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bansal A and Simon MC: Glutathione
metabolism in cancer progression and treatment resistance. J Cell
Biol. 217:2291–2298. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gasmi A, Nasreen A, Lenchyk L, Lysiuk R,
Peana M, Shapovalova N, Piscopo S, Komisarenko M, Shanaida M,
Smetanina K, et al: An update on glutathione's biosynthesis,
metabolism, functions, and medicinal purposes. Curr Med Chem.
31:4579–4601. 2024. View Article : Google Scholar
|
|
7
|
Liu R, Bian Y, Liu L, Liu L, Liu X and Ma
S: Molecular pathways associated with oxidative stress and their
potential applications in radiotherapy (Review). Int J Mol Med.
49:652022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mirhadi E, Mashreghi M, Faal Maleki M,
Alavizadeh SH, Arabi L, Badiee A and Jaafari MR: Redox-sensitive
nanoscale drug delivery systems for cancer treatment. Int J Pharm.
589:1198822020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang G, Ji J and Liu Z: Multifunctional
MnO2 nanoparticles for tumor microenvironment modulation
and cancer therapy. Wiley Interdiscip Rev Nanomed Nanobiotechnol.
13:e17202021. View Article : Google Scholar
|
|
10
|
Xue X, Wang M, Cui J, Yang M, Ma L, Kang
R, Tang D and Wang J: Glutathione metabolism in ferroptosis and
cancer therapy. Cancer Lett. 621:2176972025. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cabrales P, Izquierdo-García D, V Onecha
V, Pérez-Liva M, Fraile LM, Udías JM and L Herraiz J: Mapping
intratumoral heterogeneity through PET-derived washout and deep
learning after proton therapy. Comput Biol Med. 208:1116462026.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuo CC, Chuang YC, Wu PH, Chang WM, Lo LW,
Chen HY, Lee KH, Lee HL, Chiou JF and Shen YA: Targeting glutamine
metabolism combined with a nanoradioenhancer in radioresistant
hepatocellular carcinoma. Nanoscale. Apr 14–2026.Epub ahead of
print. View Article : Google Scholar
|
|
13
|
Kennedy L, Sandhu JK, Harper ME and
Cuperlovic-Culf M: Role of glutathione in cancer: From mechanisms
to therapies. Biomolecules. 10:14292020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao W, Zhao L, Sun Y, Yang X and Fu Q:
Stimuli-responsive nanoradiosensitizers for enhanced cancer
radiotherapy. Small Methods. 8:e23011312024. View Article : Google Scholar
|
|
15
|
Xiong Y, Xiao C, Li Z and Yang X:
Engineering nanomedicine for glutathione depletion-augmented cancer
therapy. Chem Soc Rev. 50:6013–6041. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu P, Hao L, Liu M and Hu S:
Glutathione-responsive and -exhausting metal nanomedicines for
robust synergistic cancer therapy. Front Bioeng Biotechnol.
11:11614722023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li B, Ming H, Qin S, Nice EC, Dong J, Du Z
and Huang C: Redox regulation: Mechanisms, biology and therapeutic
targets in diseases. Signal Transduct Target Ther. 10:722025.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fan Y, Tan X, Zhao H, Tu X, Liu X and Wang
Y: Cysteine metabolism in tumor redox homeostasis. Curr Med Chem.
30:1813–1823. 2023. View Article : Google Scholar
|
|
19
|
Yang B, Wang H, Xiao J and Chen W and Chen
W: ELK1/KIFC1 axis promotes breast cancer cell proliferation by
regulating glutathione metabolism. J Obstet Gynaecol Res.
49:2175–2184. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fu X, Wu H, Li C, Deng G and Chen C: YAP1
inhibits RSL3-induced castration-resistant prostate cancer cell
ferroptosis by driving glutamine uptake and metabolism to GSH. Mol
Cell Biochem. 479:2415–2427. 2024. View Article : Google Scholar :
|
|
21
|
Lv H, Zhen C, Liu J, Yang P, Hu L and
Shang P: Unraveling the potential role of glutathione in multiple
forms of cell death in cancer therapy. Oxid Med Cell Longev.
2019:31501452019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Niu B, Liao K, Zhou Y, Wen T, Quan G, Pan
X and Wu C: Application of glutathione depletion in cancer therapy:
Enhanced ROS-based therapy, ferroptosis, and chemotherapy.
Biomaterials. 277:1211102021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jones DP: Redefining oxidative stress.
Antioxid Redox Signal. 8:1865–1879. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin Z, Yang S, Qiu Q, Cui G, Zhang Y, Yao
M, Li X, Chen C, Gu J, Wang T, et al: Hypoxia-induced cysteine
metabolism reprogramming is crucial for the tumorigenesis of
colorectal cancer. Redox Biol. 75:1032862024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo W, Li K, Sun B, Xu D, Tong L, Yin H,
Liao Y, Song H, Wang T, Jing B, et al: Dysregulated glutamate
transporter SLC1A1 propels cystine uptake via Xc- for
glutathione synthesis in lung cancer. Cancer Res. 81:552–566. 2021.
View Article : Google Scholar
|
|
26
|
Zhang W, Trachootham D, Liu J, Chen G,
Pelicano H, Garcia-Prieto C, Lu W, Burger JA, Croce CM, Plunkett W,
et al: Stromal control of cystine metabolism promotes cancer cell
survival in chronic lymphocytic leukaemia. Nat Cell Biol.
14:276–286. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Venè R, Castellani P, Delfino L, Lucibello
M, Ciriolo MR and Rubartelli A: The cystine/cysteine cycle and GSH
are independent and crucial antioxidant systems in malignant
melanoma cells and represent druggable targets. Antioxid Redox
Signal. 15:2439–2453. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Q, Jia X, Li X, He M, Hao JN, Guan M,
Mao Y, Cao Y, Dai B and Li Y: One-pot fabrication of a
polydopamine-based nanoplatform for GSH triggered trimodal
ROS-amplification for cancer therapy. Biomater Sci. 10:4208–4217.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang G and Pan ST: ROS-mediated
therapeutic strategy in chemo-/radiotherapy of head and neck
cancer. Oxid Med Cell Longev. 2020:50479872020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Narayanan D, Ma S and Özcelik D: Targeting
the redox landscape in cancer therapy. Cancers (Basel).
12:17062020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li X and Li D: Natural compounds targeting
cellular redox homeostasis in malignancies. Cell Mol Biol
(Noisy-le-grand). 69:255–265. 2023. View Article : Google Scholar
|
|
32
|
Zhang Y and Martin SG: Redox proteins and
radiotherapy. Clin Oncol (R Coll Radiol). 26:289–300. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Milković L, Mlinarić M, Lučić I and Čipak
Gašparović A: The involvement of peroxiporins and antioxidant
transcription factors in breast cancer therapy resistance. Cancers
(Basel). 15:57472023. View Article : Google Scholar
|
|
34
|
Zhang J, Shen L and Sun LQ: The regulation
of radiosensitivity by p53 and its acetylation. Cancer Lett.
363:108–118. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Suzuki N, Johmura Y, Wang TW, Migita T, Wu
W, Noguchi R, Yamaguchi K, Furukawa Y, Nakamura S, Miyoshi I, et
al: TP53/p53-FBXO22-TFEB controls basal autophagy to govern
hormesis. Autophagy. 17:3776–3793. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Song B, Yang P and Zhang S: Cell fate
regulation governed by p53: Friends or reversible foes in cancer
therapy. Cancer Commun (Lond). 44:297–360. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mao C, Lei G, Horbath A and Gan B:
Assessment of lipid peroxidation in irradiated cells. Methods Cell
Biol. 172:37–50. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ursini F and Maiorino M: Lipid
peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic
Biol Med. 152:175–185. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hussain S, Gupta G, Shahwan M, Bansal P,
Kaur H, Deorari M, Pant K, Ali H, Singh SK, Rama Raju Allam VS, et
al: Non-coding RNA: A key regulator in the glutathione-GPX4 pathway
of ferroptosis. Noncoding RNA Res. 9:1222–1234. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li Q, Wu Z, Chen S, Liang Y, Zhu K, Su N,
Liu T and Zhao B: Enhancing ferroptosis-mediated radiosensitization
via synergistic disulfidptosis induction. ACS Nano. 19:1757–1770.
2025. View Article : Google Scholar
|
|
41
|
Feng L, Zhao K, Sun L, Yin X, Zhang J, Liu
C and Li B: SLC7A11 regulated by NRF2 modulates esophageal squamous
cell carcinoma radiosensitivity by inhibiting ferroptosis. J Transl
Med. 19:3672021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang Q, Zhang M, Huang X, Liu X and Li W:
Inhibition of cytoskeletal protein carbonylation may protect
against oxidative damage in traumatic brain injury. Exp Ther Med.
12:4107–4112. 2016. View Article : Google Scholar
|
|
43
|
Zepeta-Flores N, Valverde M,
Lopez-Saavedra A and Rojas E: Glutathione depletion triggers actin
cytoskeleton changes via actin-binding proteins. Genet Mol Biol.
41:475–487. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Patel M, Horgan PG, McMillan DC and
Edwards J: NF-κB pathways in the development and progression of
colorectal cancer. Transl Res. 197:43–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fu H, Zhang Y, Duan Y, Zhang X, Yao J,
Yang D, Wei Z, Zhu Z, Xu J, Hu Z, et al: Superoxide dismutase
promotes gastric tumorigenesis mediated by Helicobacter pylori and
enhances resistance to 5-fluorouracil in gastric cancer. iScience.
28:1115532024. View Article : Google Scholar
|
|
46
|
Filomeni G, Aquilano K, Rotilio G and
Ciriolo MR: Antiapoptotic response to induced GSH depletion:
Involvement of heat shock proteins and NF-kappaB activation.
Antioxid Redox Signal. 7:446–455. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Seidel P, Roth M, Ge Q, Merfort I, S'Ng CT
and Ammit AJ: IκBα glutathionylation and reduced histone H3
phosphorylation inhibit eotaxin and RANTES. Eur Respir J.
38:1444–1452. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lou H and Kaplowitz N: Glutathione
depletion down-regulates tumor necrosis factor alpha-induced
NF-kappaB activity via IkappaB kinase-dependent and -independent
mechanisms. J Biol Chem. 282:29470–29481. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang Y, Zhang Y, Xue J, Gao L, Li X, Zhao
M, Zhao D and Zhou X: Ferroptosis mediates decabromodiphenyl
ether-induced liver damage and inflammation. Ecotoxicol Environ
Saf. 255:1147712023. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhong Z, Jiao Z and Yu FX: The Hippo
signaling pathway in development and regeneration. Cell Rep.
43:1139262024. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hatem E, Berthonaud V, Dardalhon M,
Lagniel G, Baudouin-Cornu P, Huang ME, Labarre J and Chdin S:
Glutathione is essential to preserve nuclear function and cell
survival under oxidative stress. Free Radic Biol Med. 75(Suppl 1):
S25–S26. 2014. View Article : Google Scholar
|
|
52
|
Ciamporcero E, Daga M, Pizzimenti S,
Roetto A, Dianzani C, Compagnone A, Palmieri A, Ullio C, Cangemi L,
Pili R and Barrera G: Crosstalk between Nrf2 and YAP contributes to
maintaining the antioxidant potential and chemoresistance in
bladder cancer. Free Radic Biol Med. 115:447–457. 2018. View Article : Google Scholar
|
|
53
|
Banjac A, Perisic T, Sato H, Seiler A,
Bannai S, Weiss N, Kölle P, Tschoep K, Issels RD, Daniel PT, et al:
The cystine/cysteine cycle: A redox cycle regulating susceptibility
versus resistance to cell death. Oncogene. 27:1618–1628. 2008.
View Article : Google Scholar
|
|
54
|
Cheng X, Xu HD, Ran HH, Liang G and Wu FG:
Glutathione-depleting nanomedicines for synergistic cancer therapy.
ACS Nano. 15:8039–8068. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhang H, Shan G, Liu M, Sun Q, Yang T,
Peng R, Li X, Mei Y, He X and Qiao L: Harnessing ROS amplification
and GSH depletion using a carrier-free nanodrug to enhance
ferroptosis-based cancer therapy. Small. 21:e24092502025.
View Article : Google Scholar
|
|
56
|
Cao S, Li X, Gao Y, Li F, Li K, Cao X, Dai
Y, Mao L, Wang S and Tai X: A simultaneously GSH-depleted
bimetallic Cu(ii) complex for enhanced chemodynamic cancer therapy.
Dalton Trans. 49:11851–11858. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kang N, Son S, Min S, Hong H, Kim C, An J,
Kim JS and Kang H: Stimuli-responsive ferroptosis for cancer
therapy. Chem Soc Rev. 52:3955–3972. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Liu X, Li Y, Wang K, Chen Y, Shi M, Zhang
X, Pan W, Li N and Tang B: GSH-responsive nanoprodrug to inhibit
glycolysis and alleviate immunosuppression for cancer therapy. Nano
Lett. 21:7862–7869. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kou L, Sun R, Xiao S, Zheng Y, Chen Z, Cai
A, Zheng H, Yao Q, Ganapathy V and Chen R: Ambidextrous approach to
disrupt redox balance in tumor cells with increased ROS production
and decreased GSH synthesis for cancer therapy. ACS Appl Mater
Interfaces. 11:26722–26730. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen M, Liu D, Liu F, Wu Y, Peng X and
Song F: Recent advances of redox-responsive nanoplatforms for tumor
theranostics. J Control Release. 332:269–284. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang C, Chen L, Bai Q, Wang L, Li S, Sui
N, Yang D and Zhu Z: Nonmetal graphdiyne nanozyme-based
ferroptosis-apoptosis strategy for colon cancer therapy. ACS Appl
Mater Interfaces. 14:27720–27732. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mittal A, Yadav M, Biswas L, Verma AK and
Roy I: Nanoparticles for combined photo- and chemo-dynamic therapy
of cancer cells involving endogenous glutathione depletion.
Nanomedicine (Lond). 18:679–694. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Dong Z, Feng L, Chao Y, Hao Y, Chen M,
Gong F, Han X, Zhang R, Cheng L and Liu Z: Amplification of tumor
oxidative stresses with liposomal fenton catalyst and glutathione
inhibitor for enhanced cancer chemotherapy and radiotherapy. Nano
Lett. 19:805–815. 2019. View Article : Google Scholar
|
|
64
|
Qi R and Jin W, Wang J, Yi Q, Yu M, Xu S
and Jin W: Oleanolic acid enhances the radiosensitivity of tumor
cells under mimetic hypoxia through the reduction in intracellular
GSH content and HIF-1α expression. Oncol Rep. 31:2399–2406. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cardinali CAEF, Fabiano de Freitas C,
Sonchini Gonçalves R, Amanda Pedroso de Morais F, Nunes de Lima
Martins J, Martins YA, Fernando Comar J, de Souza Bonfim-Mendonça
P, Tessaro AL, Kimura E, et al: 'Effects of redox status on
immediate hypericin-mediated photodynamic therapy in human
glioblastoma T98G cell line'. ACS Omega. 10:1100–1109. 2024.
View Article : Google Scholar
|
|
66
|
Quindoza GM III, Nakagawa Y, Anraku Y and
Ikoma T: Adsorption of l-buthionine sulfoximine on Bi(III) and
Eu(III) co-substituted hydroxyapatite nanocrystals for enhancing
radiosensitization effects. Colloids Surf B Biointerfaces.
228:1134032023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zeng L, Ding S, Cao Y, Li C, Zhao B, Ma Z,
Zhou J, Hu Y, Zhang X, Yang Y, et al: A MOF-based potent
ferroptosis inducer for enhanced radiotherapy of triple negative
breast cancer. ACS Nano. 17:13195–13210. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kong A, Johnson N, Kitchener HC and Lawrie
TA: Adjuvant radiotherapy for stage I endometrial cancer. Cochrane
Database Syst Rev. CD0039162012.PubMed/NCBI
|
|
69
|
Valodkar M, Jadeja RN, Thounaojam MC,
Devkar RV and Thakore S: In vitro toxicity study of plant latex
capped silver nanoparticles in human lung carcinoma cells. Mater
Sci Eng. 31:1723–1728. 2011. View Article : Google Scholar
|
|
70
|
Muraoka M, Yoshida S, Ohno M, Matsuura H,
Nagano K, Hirata Y, Arai M and Hirata K: Reactivity of
γ-glutamyl-cysteine with intracellular and extracellular
glutathione metabolic enzymes. FEBS Lett. 596:180–188. 2022.
View Article : Google Scholar
|
|
71
|
Xu S, Gao W, Lin S, Kang B, Shi X, Zheng
D, Xiao L, Zeng L, Zhang Q, Tian W and Song H: Self-assembly
albumin nanoparticles for improved anticancer effect and reinforced
ferroptosis in metastatic colorectal cancer treatment. ACS Appl
Mater Interfaces. 17:66475–66491. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Kerkhove L, Geirnaert F, Rifi AL, Law KL,
Gutiérrez A, Oudaert I, Corbet C, Gevaert T, Dufait I and De Ridder
M: Repurposing sulfasalazine as a radiosensitizer in hypoxic human
colorectal cancer. Cancers (Basel). 15:23632023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Takeuchi S, Wada K, Nagatani K, Otani N,
Osada H and Nawashiro H: Sulfasalazine and temozolomide with
radiation therapy for newly diagnosed glioblastoma. Neurol India.
62:42–47. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
He J, Zhang Y, Luo S, Zhao Z, Mo T, Guan
H, Li H, Bian Z, Zhang X, Qiu S, et al: Targeting SLC7A11 with
sorafenib sensitizes stereotactic body radiotherapy in colorectal
cancer liver metastasis. Drug Resist Updat. 81:1012502025.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Sarowar S, Cirillo D, Játiva P, Nilsen MH,
Otragane SMA, Heggdal J, Selheim F, Ceña V, Bjørsvik HR and Enger
PØ: The Styryl benzoic acid derivative DC10 potentiates
radiotherapy by targeting the xCT-glutathione axis. Front Oncol.
12:7867392022. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Li X, Jiang Z, Feng J, Zhang X, Wu J and
Chen W: 2-Acetylamino-3-[4-(2-acetylamino-2-carboxyethylsulfanylca
rbon ylamino) phenyl carbamoylsulfanyl] propionic acid, a
glutathione reductase inhibitor, induces G2/M cell cycle
arrest through generation of thiol oxidative stress in human
esophageal cancer cells. Oncotarget. 8:61846–61860. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhu Z, Du S, Du Y, Ren J, Ying G and Yan
Z: Glutathione reductase mediates drug resistance in glioblastoma
cells by regulating redox homeostasis. J Neurochem. 144:93–104.
2018. View Article : Google Scholar
|
|
78
|
Liu X, Zheng T, Bao Y, Li P, Zhao T, Liu
Y, Wang H and Sun C: Genistein implications in radiotherapy: Kill
two birds with one stone. Molecules. 30:1882025. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Tang Q, Ma J, Sun J, Yang L, Yang F, Zhang
W, Li R, Wang L, Wang Y and Wang H: Genistein and AG1024
synergistically increase the radiosensitivity of prostate cancer
cells. Oncol Rep. 40:579–588. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
González-Blanco A, Allo A, Barcia R and
Ramos-Martínez JI: Inhibition of glutathione reductase uncovers the
activation of NADPH-inhibited glucose-6-phosphate dehydrogenase.
Biotechnol Appl Biochem. 69:1690–1695. 2022. View Article : Google Scholar
|
|
81
|
Rashmi R, Huang X, Floberg JM, Elhammali
AE, McCormick ML, Patti GJ, Spitz DR and Schwarz JK: Radioresistant
cervical cancers are sensitive to inhibition of glycolysis and
redox metabolism. Cancer Res. 78:1392–1403. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Narayanankutty A, Job JT and
Narayanankutty V: Glutathione, an antioxidant tripeptide: Dual
roles in carcinogenesis and chemoprevention. Curr Protein Pept Sci.
20:907–917. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Shi X, Zhang W, Gu C, Ren H, Wang C, Yin
N, Wang Z, Yu J, Liu F and Zhang H: NAD+ depletion radiosensitizes
2-DG-treated glioma cells by abolishing metabolic adaptation. Free
Radic Biol Med. 162:514–522. 2021. View Article : Google Scholar
|
|
84
|
Bianchi ME and Mezzapelle R: The chemokine
receptor CXCR4 in cell proliferation and tissue regeneration. Front
Immunol. 11:21092020. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dwarakanath BS: Cytotoxicity,
radiosensitization, and chemosensitization of tumor cells by
2-deoxy-D-glucose in vitro. J Cancer Res Ther. 5(Suppl 1): S27–S31.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hasse FC, Koerber SA, Prigge ES, Liermann
J, von Knebel Doeberitz M, Debus J and Sterzing F: Overcoming
radioresistance in WiDr cells with heavy ion irradiation and
radiosensitization by 2-deoxyglucose with photon irradiation. Clin
Transl Radiat Oncol. 19:52–58. 2019.PubMed/NCBI
|
|
87
|
Varshney R, Dwarakanath B and Jain V:
Radiosensitization by 6-aminonicotinamide and 2-deoxy-D-glucose in
human cancer cells. Int J Radiat Biol. 81:397–408. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Qiao K, Pan Y, Zhang S, Shi G, Yang J,
Zhang Z, Wang K, Chen X and Ning S: Cold exposure therapy
sensitizes nanodrug-mediated radioimmunotherapy of breast cancer.
ACS Nano. 18:29689–29703. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Chen J, Feng C, Lan Y, Chen X, Peng Z,
Huang Z, Wang R, Zhang W, Ye Y, Mao Z, et al: Bidirectional
regulation of reactive oxygen species for radiosensitization in
nasopharyngeal carcinoma. J Nanobiotechnology. 23:962025.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kim W, Lee S, Seo D, Kim D, Kim K, Kim E,
Kang J, Seong KM, Youn H and Youn B: Cellular stress responses in
radiotherapy. Cells. 8:11052019. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Dangi K, Kumar V, Mittal D, Yadav P, Malik
M and Verma AK: Nanotherapeutics induced redox resetting of
oxidative and nitrosative stress: Targeting glutathione-depletion
in cancer. Nanomedicine (Lond). 20:955–965. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Jiang W, Huang G, Pan S, Chen X, Liu T,
Yang Z, Chen T and Zhu X: TRAIL-driven targeting and reversing
cervical cancer radioresistance by seleno-nanotherapeutics through
regulating cell metabolism. Drug Resist Updat. 72:1010332024.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ding Y, Sun Z, Tong Z, Zhang S, Min J, Xu
Q, Zhou L, Mao Z, Xia H and Wang W: Tumor
microenvironment-responsive multifunctional peptide coated
ultrasmall gold nanoparticles and their application in cancer
radiotherapy. Theranostics. 10:5195–5208. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wang X, Wei N, Zhang Y, Fang Y, Li Y, Li
S, Wang Z and Sun C: Nanozyme-mediated glutathione depletion for
enhanced ROS-based cancer therapies: A comprehensive review.
Nanomedicine (Lond). 20:279–290. 2025. View Article : Google Scholar :
|
|
95
|
Zhang Q, Sun Z, Sun W, Yu B, Liu J, Jiang
C and Lu L: Engineering a synergistic antioxidant inhibition
nanoplatform to enhance oxidative damage in tumor treatment. Acta
Biomater. 158:625–636. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yong Y, Zhang C, Gu Z, Du J, Guo Z, Dong
X, Xie J, Zhang G, Liu X and Zhao Y: Polyoxometalate-based
radiosensitization platform for treating hypoxic tumors by
attenuating radioresistance and enhancing radiation response. ACS
Nano. 11:7164–7176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Luo K, Guo W, Yu Y, Xu S, Zhou M, Xiang K,
Niu K, Zhu X, Zhu G, An Z, et al: Reduction-sensitive platinum
(IV)-prodrug nano-sensitizer with an ultra-high drug loading for
efficient chemo-radiotherapy of Pt-resistant cervical cancer in
vivo. J Control Release. 326:25–37. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Zhou R, Wang H, Yang Y, Zhang C, Dong X,
Du J, Yan L, Zhang G, Gu Z and Zhao Y: Tumor
microenvironment-manipulated radiocatalytic sensitizer based on
bismuth heteropolytungstate for radiotherapy enhancement.
Biomaterials. 189:11–22. 2019. View Article : Google Scholar
|
|
99
|
Ren S, Zhang Q, Fu H, Cheng J, Xie Y,
Liang Q and Xiao X: A reactive oxygen species amplifier based on a
Bi WO2/BP6 heterojunction for high efficiency
radiotherapy enhancement. J Mater Chem B. 13:3128–3137. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Chen X, Qiu M, Liu L, Ji Q, Xu Z, Xiong Z
and Yang S: Intelligent
Bi2Se3@Cu2-xSe heterostructures
with enhanced photoabsorption and photoconversion efficiency for
tri-modal imaging guided combinatorial cancer therapy by
near-infrared II light. J Colloid Interface Sci. 625:614–627. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Liu HJ, Chen W, Wu G, Zhou J, Liu C, Tang
Z, Huang X, Gao J, Xiao Y, Kong N, et al: Glutathione-scavenging
nanoparticle-mediated PROTACs delivery for targeted protein
degradation and amplified antitumor effects. Adv Sci (Weinh).
10:e22074392023. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Fu S, Rempson CM, Puche V, Zhao B and
Zhang F: Construction of disulfide containing redox-responsive
polymeric nanomedicine. Methods. 199:67–79. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Ling X, Chen X, Riddell IA, Tao W, Wang J,
Hollett G, Lippard SJ, Farokhzad OC, Shi J and Wu J:
Glutathione-scavenging poly(disulfide amide) nanoparticles for the
effective delivery of Pt(IV) prodrugs and reversal of cisplatin
resistance. Nano Lett. 18:4618–4625. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Xie J, Gong L, Zhu S, Yong Y, Gu Z and
Zhao Y: Emerging strategies of nanomaterial-mediated tumor
radiosensitization. Adv Mater. 31:e18022442019. View Article : Google Scholar
|
|
105
|
Mane SD and Kamatham AN: Ascorbyl stearate
and ionizing radiation potentiate apoptosis through intracellular
thiols and oxidative stress in murine T lymphoma cells. Chem Biol
Interact. 281:37–50. 2018. View Article : Google Scholar
|
|
106
|
Zhang X, Chen X, Jiang YW, Ma N, Xia LY,
Cheng X, Jia HR, Liu P, Gu N, Chen Z and Wu FG:
Glutathione-depleting gold nanoclusters for enhanced cancer
radiotherapy through synergistic external and internal regulations.
ACS Appl Mater Interfaces. 10:10601–10606. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Hua S, Zhao J, Li L, Liu C, Zhou L, Li K,
Huang Q, Zhou M and Wang K: Photosynthetic bacteria-based
whole-cell inorganic-biohybrid system for multimodal enhanced tumor
radiotherapy. J Nanobiotechnology. 22:3792024. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Yao L, Bojic D and Liu M: Applications and
safety of gold nanoparticles as therapeutic devices in clinical
trials. J Pharm Anal. 13:960–967. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wang H, Jiang H, Corbet C, de Mey S, Law
K, Gevaert T, Feron O and De Ridder M: Piperlongumine increases
sensitivity of colorectal cancer cells to radiation: Involvement of
ROS production via dual inhibition of glutathione and thioredoxin
systems. Cancer Lett. 450:42–52. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Liu F, Zhou Q, Jiang HF, Zhang TT, Miao C,
Xu XH, Wu JX, Yin SL, Xu SJ, Peng JY, et al: Piperlongumine
conquers temozolomide chemoradiotherapy resistance to achieve
immune cure in refractory glioblastoma via boosting oxidative
stress-inflamation-CD8+-T cell immunity. J Exp Clin
Cancer Res. 42:1182023. View Article : Google Scholar
|
|
111
|
Sun H, Bai Y, Zhao D, Wang J and Qiu L:
Transition-metal-oxide-based nanozymes for antitumor applications.
Materials (Basel). 17:28962024. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Shao L, Wang X, Du X, Yin S, Qian Y, Yao Y
and Yang L: Application of multifunctional nanozymes in tumor
therapy. ACS Omega. 9:15753–15767. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wang J, Tan X, Pang X, Liu L, Tan F and Li
N: MoS2 quantum Dot@Polyaniline inorganic-organic nanohybrids for
in vivo dual-modal imaging guided synergistic
photothermal/radiation therapy. ACS Appl Mater Interfaces.
8:24331–24338. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Liu H, Zhen Z, Chen F, Chen J and Chen W:
Advancements in iron oxide nanoparticles for multimodal imaging and
tumor theranostics. Curr Med Chem. 32:301–321. 2025. View Article : Google Scholar
|
|
115
|
Liu J, Liu C, Tang J, Chen Q, Yu Y, Dong
Y, Hao J and Wu W: Synergistic cerium oxide nanozymes: Targeting
DNA damage and alleviating tumor hypoxia for improved NSCLC
radiotherapy efficiency. J Nanobiotechnology. 22:252024. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Pan S, Sun Z, Zhao B, Miao L, Zhou Q, Chen
T and Zhu X: Therapeutic application of manganese-based nanosystems
in cancer radiotherapy. Biomaterials. 302:1223212023. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wu S, Liu X, Ren J and Qu X: Glutathione
depletion in a benign manner by MoS2-based nanoflowers
for enhanced hypoxia-irrelevant free-radical-based cancer therapy.
Small. 15:e19048702019. View Article : Google Scholar
|
|
118
|
Pan S, Huang G, Sun Z, Chen X, Xiang X,
Jiang W, Xu Y, Chen T and Zhu X: X-ray-responsive zeolitic
imidazolate framework-capped nanotherapeutics for cervical
cancer-targeting radiosensitization (Adv. Funct. Mater. 13/2023).
Adv Funct Mater. 33:22133362023.
|
|
119
|
Xiong Z, Yang M, Liu P, Tang Z, Yang Y,
Zhan M, Chen T, Li X and Lu L: Designing bimetallic metal-organic
framework-based heterojunction radiosensitizer for enhanced
radiodynamic therapy and immunotherapy. Adv Funct Mater.
34:23129192024. View Article : Google Scholar
|
|
120
|
Vo QNQ and Park CH: Interfacial
engineering of hollow reduced graphene oxide-molybdenum disulfide
nanosphere for surface-regulated targeted drug release and
photothermal conversion. J Colloid Interface Sci. 717:1404612026.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Ruiz-Hitzky E, Martín-Morales C, Barra A,
Nunes C and Ferreira P: Clay-polysaccharides as engineered
nanoplatforms for cancer treatment. Carbohydr Polym.
381:1251692026. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Xu J, Qiu W, Liang M, Ye M, Hu J, Ma X,
Shi X, Xue P, Kang Y, Xiao B and Xu Z: Dual-stimulus
phototherapeutic nanogel for triggering pyroptosis to promote
cancer immunotherapy. J Control Release. 358:219–231. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Chen D, Zhang P, Li M, Li C, Lu X, Sun Y
and Sun K: Hyaluronic acid-modified redox-sensitive hybrid
nanocomplex loading with siRNA for non-small-cell lung carcinoma
therapy. Drug Deliv. 29:574–587. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Qiao K, Mo J, Pan Y, Zhang S, Jiang C, Lyu
M, Cui Y, Huang Y and Ning S: Gold nanoparticle doped Cuhemin
nanosheets with a remodeling tumor microenvironment for multiple
radiotherapy sensitization. J Mater Chem B. 11:4095–4101. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Li Y, Wang Q, Xia H, Wang F, Jiao P, Song
M and Jiang G: Silver nanoparticles specially amplify the
anticancer effects of platinum-based drugs by disrupting cellular
nucleotide and sulfur homeostasis. Nano Lett. 26:4353–4361. 2026.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Chauhan AS, Chand P and Parashar T:
Lipid-based nanoparticles: Strategy for targeted cancer therapy.
BIO Integr. 6:9942025. View Article : Google Scholar
|
|
127
|
Chen C, Chen N, Qi Y, Lyu M, Wu C, Xie C
and Yu H: Copper-based single-atom nanozyme system mimicking
platelet cells for enhancing the outcome of radioimmunotherapy. Int
J Nanomedicine. 19:403–414. 2024. View Article : Google Scholar : PubMed/NCBI
|















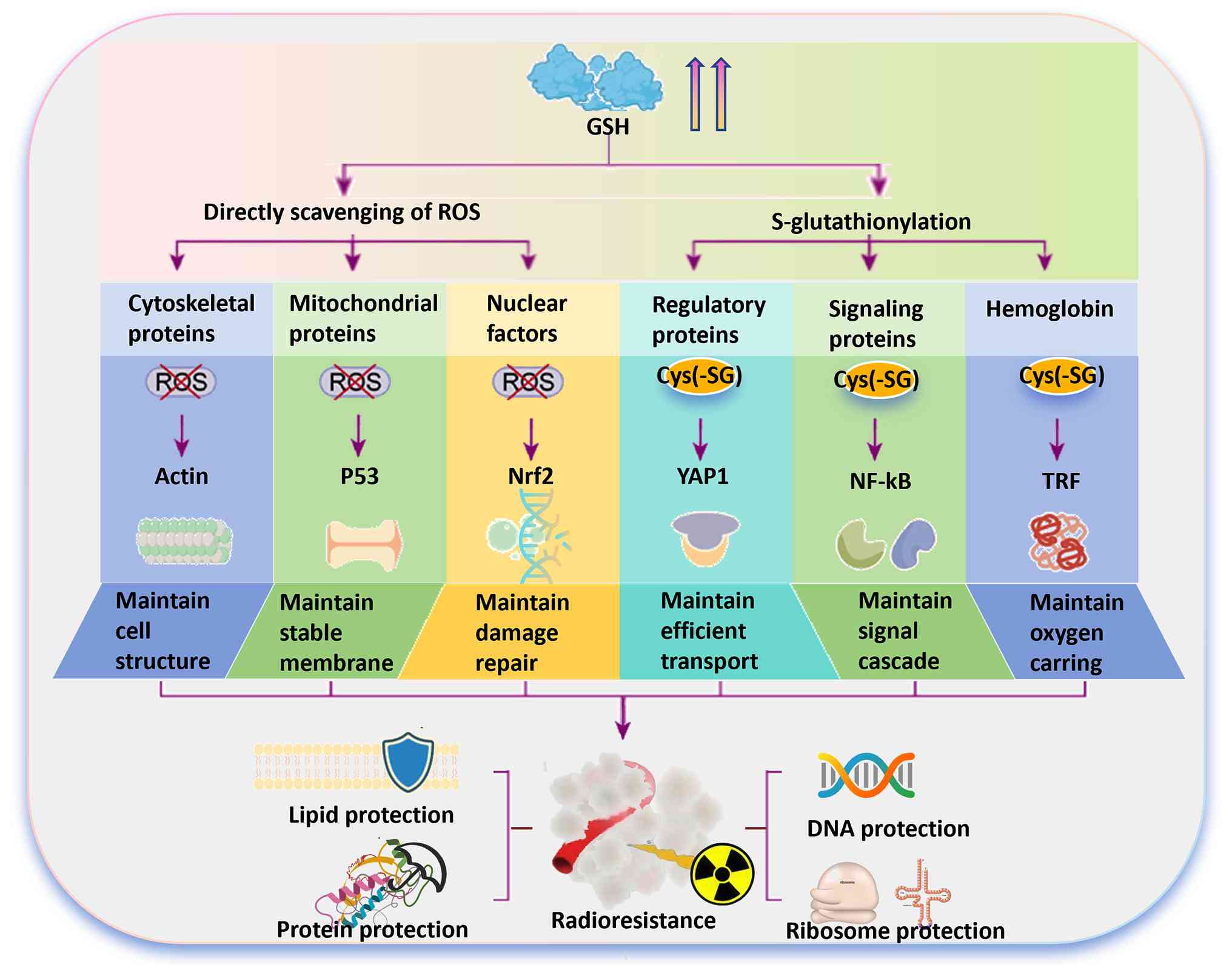
![Promoting GSH depletion for tumor
radiosensitization. Various targeting strategies of promoting GSH
depletion (oxidation reaction, covalent linkage and enzymatic
reaction) reduce GSH levels and enhance radiotherapy efficiency
[created with Biorender (biorender.com)]. GSH, glutathione; ROS, reactive
oxygen species; TME, tumor microenvironment; GSSG, oxidized
glutathione; GS-X, glutathione polymer.](/article_images/ijmm/58/2/ijmm-58-02-05892-g04.jpg)