Introduction
Ischemic stroke primarily results from cerebral
artery occlusion, leading to insufficient glucose and oxygen in
brain cells. Restoring blood flow perfusion is the main treatment
strategy for ischemic stroke. Nevertheless, rapid reperfusion
inevitably leads to secondary brain tissue injury, namely, cerebral
ischemia/reperfusion injury (CIRI) (1,2).
The pathological process of CIRI is complicated and involves the
interaction of numerous factors, including the inflammatory
response, oxidative stress, energy metabolism disorders and
neuronal death (3). Pyroptosis
is a recently discovered form of programmed cell death that
mediates neuroinflammation after ischemic stroke, and the
suppression of pyroptosis may hold substantial promise for rescuing
neurological deterioration during CIRI (4,5).
Owing to irreversible neuronal death, the prognosis of CIRI is
dismal. Therefore, preventing and reducing neuronal death in CIRI
remains a medical hotspot worthy of extensive research.
Opioid receptors (ORs), including mu ORs (MORs),
delta ORs (DORs), and kappa ORs (KORs), exhibit both short- and
long-term alterations and are redistributed in brain regions after
ischemic stroke in rodents (6).
Appropriate activation of ORs at specific times and regions may be
a crucial neuroprotective strategy after stroke (7,8).
Opioid agents act by non-overlapping molecular mechanisms,
including stimulating the release of neurotrophic factors,
protecting cerebral vascular autoregulation, limiting glutamate
release, reducing oxidative injury, and maintaining brain
homeostasis to exert neuroprotective effects (9). Hydromorphone (HM), a potent opioid
analgesic widely used for pain relief (10), has been reported to protect
against ischemic oxidative stress in rat glial cells (11). HM preconditioning markedly
represses I/R-induced neuronal apoptosis in the hippocampal CA1
region and alleviates oxidative stress (12). HM restrains Nod-like receptor
protein 3 (NLRP3) inflammasome-mediated pyroptosis by activating
the Nrf2/HO-1 pathway (13).
Nevertheless, there is currently no relevant research reporting the
effect of HM on pyroptosis in CIRI. In accordance with previous
studies (14-16), human brain microvascular
endothelial cells (HBMVECs) were chosen to establish an I/R cell
model to investigate the effect of HM on cell pyroptosis.
microRNAs (miRNAs or miRs) are a class of small
non-coding RNAs (~22 nucleotides in length) that play vital roles
in CIRI (17). One such miRNA,
miR-195-5p, has been shown to mitigate CIRI via PTEN-AKT signaling
(16). A miR-195-5p inhibitor
aggravated inflammation and apoptosis in oxygen-glucose
deprivation/reperfusion (OGD/R)-induced cells (18). miR-195-5p expression is also
notably reduced in mouse I/R testis tissues, and miR-195-5p
represses testicular I/R-induced pyroptosis (19). The expression of miR-195-5p can
be upregulated by the use of propofol, another commonly used
anesthetic drug (20). However,
whether HM can affect miR-195-5p expression remains unclear.
N6-methyladenosine (m6A) is the most prevalent RNA
epigenetic modulator in eukaryotic cells and plays a pivotal role
in physiological processes during CIRI (21). m6A modification is orchestrated
by methyltransferases (writers), removed by demethylases (erasers),
and recognized by RNA-binding proteins (readers) (22). RBM15 functions as a
recruiter/adapter for the writer complex, and its expression is
upregulated in ischemic stroke samples compared with controls
(23). RNA-binding motif protein
15 (RBM15) intensifies inflammation, oxidative stress and
pyroptosis to facilitate diabetic nephropathy progression (24). A binding relationship between
miR-195-5p and RBM15 was predicted through the Starbase database
(https://rnasysu.com/encori/). Hence, it
was hypothesized that HM preconditioning affects pyroptosis in CIRI
via miR-195-5p/RBM15. The present study aimed to investigate the
underlying protective mechanism of HM preconditioning in CIRI,
providing a theoretical basis for improving the prognosis of
CIRI.
Materials and methods
Ethics statement
The study procedure was approved by the Ethics
Committee of Liaoning Cancer Hospital and Institute (approval no.
CMU20250031; Shenyang; China). All animal experimental procedures
were approved by the Animal Ethics Committee of Liaoning Cancer
Hospital and Institute and implemented on the basis of the Guide
for the Care and Use of Laboratory Animals (25).
Database prediction
The downstream target genes of miR-195-5p were
predicted using the Starbase database (https://rnasysu.com/encori/) (26), and the binding site between
miR-195-5p and RBM15 was determined. The m6A modification level of
USF2 was predicted using the SRAMP database (http://www.cuilab.cn/sramp/) (27). The binding between USF2 and NLRP3
was predicted using the JASPAR database (http://jaspar.genereg.net/) (28).
Establishment and treatment of a mouse
I/R model
Male C57BL/6J mice (7-8 weeks; 20-25 g) were
purchased from Vital River Laboratory Animal Technology Co., Ltd.
and kept under specific pathogen-free conditions at 21-24°C with a
light/dark cycle of 12/12-h. Mice had free access to water and food
(both sterilized). The water bottles and padding were regularly
replaced, and the cages were disinfected.
After they were weighed, the mice were grouped
according to random numbers, which were recorded by surgical
personnel. The mice were randomly divided into 6 groups [9 mice per
group, calculated by G * Power, Effect size: f=0.6, α err
prob=0.05, and Power (1-β err prob)=0.9]. In the sham group, mice
underwent all the same surgical procedures as the I/R group mice,
except that the arteries were not ligated; I/R group: mice with
ischemia for 30 min; I/R + saline group: mice with subcutaneous
injection of 0.9% physiological saline for 5 days, followed by 30
min of ischemia; I/R + HM group: mice with subcutaneous injection
of 2 mg/kg HM (Yichang Humanwell Pharmaceutical Co., Ltd.; NMPN:
H20120100; cat. no. TD2012-0010) for 4 consecutive days and
injection of 3 mg/kg HM 4 h before surgery (12), followed by 30 min of ischemia. In
the I/R + HM + antagomir NC group, mice were subcutaneously
injected with 2 mg/kg HM for 4 consecutive days, 100 μM
antagomir NC was injected into the mouse ventricle stereotactically
one day before surgery, and another 3 mg/kg HM was injected 4 h
before surgery, followed by 30 min of ischemia; in the I/R + HM +
antagomir-195 group, mice were subcutaneously injected with 2 mg/kg
HM for 4 consecutive days, 100 μM antagomir-195 was injected
into the mouse ventricle stereotactically one day before surgery,
and another 3 mg/kg HM was injected 4 h before surgery, followed by
30 min of ischemia. The specific experimental process is shown in
Fig. 1. The miR-195-5p antagomir
(antagomir-195) and antagomir-NC were purchased from Shanghai
GenePharma Co., Ltd. Stereotactic coordinates: Anteroposterior, 0.8
mm; mediolateral, 1.5 mm; depth, 3.5 mm. The surgery was performed
under anesthesia using a stereotaxic device (RWD Life Science). In
total, 100 μM antagomir-NC or antagomir-195 was injected
into the mouse ventricle via the stereotaxic method (16) at a rate of 0.5 μl/min. The
needle was kept in its original position for 5 min and slowly
withdrawn, after which the skin incision was sutured.
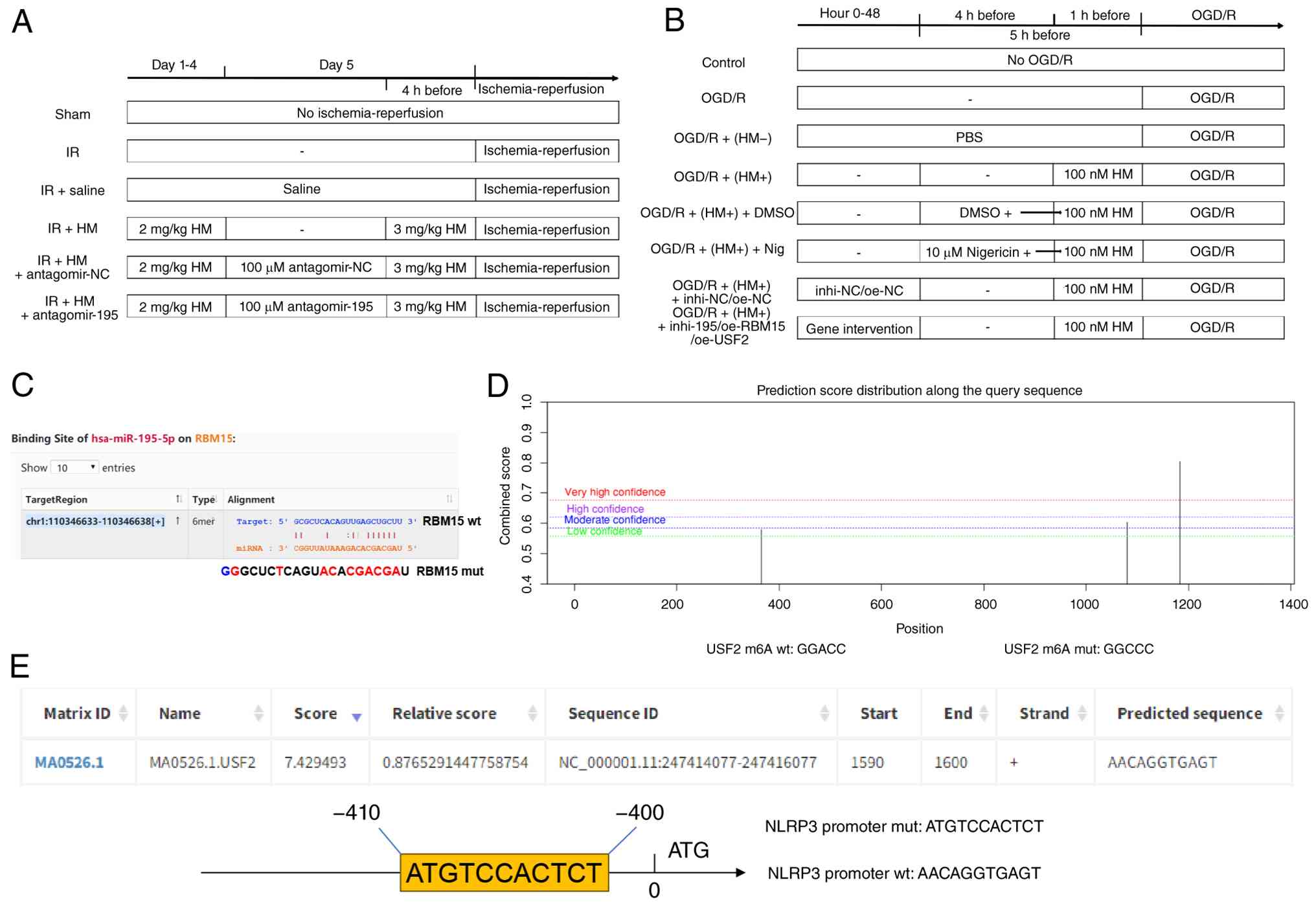 | Figure 1HM preconditioning flowchart and
database prediction results. (A) Establishment and processing flow
of the IR mouse model. (B) Establishment and processing flow of the
OGD/R cell model. (C) Prediction of the binding site between
miR-195-5p and RBM15 using the Starbase database. (D) Prediction of
the m6A modification level of USF2 using the SRAMP database. (E)
Prediction of the binding between USF2 and NLRP3 using the JASPAR
database. HM, hydromorphone; I/R, ischemia/reperfusion; OGD/R,
oxygen-glucose deprivation/reperfusion; m6A, N6-methyladenosine;
USF2, upstream stimulatory factor 2; NLRP3, Nod-like receptor
protein 3; NC, negative control; WT, wild-type; MUT, mutant; miR,
microRNA. |
The mice were anaesthetized by intraperitoneal
injection of pentobarbital sodium (30 mg/kg) 4 h after the last
administration of HM (16), and
the body temperature was maintained at 37±1.0°C on a heating pad
throughout all the experimental procedures. Afterwards, the
junction of the carotid artery and the proximal end of the internal
carotid artery was exposed through a midline incision in the neck
under a surgical microscope. A 6-0 monofilament coated with
silicone was inserted into the carotid artery at a distance of
7±0.5 mm below the carotid branch until mild resistance was felt
(29). After 30 min of ischemia,
the monofilament was removed to achieve reperfusion. Blood flow was
observed using a laser Doppler perfusion imaging device (PeriCam
PSI HR; https://www.perimed-instruments.com/). A decrease of
more than 30% in cerebral blood flow (CBF) relative to baseline was
considered sufficient induction of ischemia. After the obstructive
filament was removed, an increase in the CBF to more than 70% of
the baseline value was considered successful reperfusion. All the
animals were operated on by the same operator under the same
conditions to reduce the variability of infarction. The surgery
time for each animal did not exceed 15 min. After 24 h, the
neurological deficits of the mice were assessed. To evaluate brain
injury, 0.5 ml of blood was collected from the orbital vein of each
mouse, and serum was collected after centrifugation (2,000 × g,
4°C, 15 min). Finally, the mice were euthanized by intraperitoneal
injection of sodium pentobarbital (200 mg/kg). Death was confirmed
by observing respiratory arrest, disappearance of nerve reflex and
muscle relaxation, in combination with observation of heartbeat
(touching the apical beat). The brain tissues were collected for
subsequent experiments. During the experiment, if the mice were
unable to independently drink water, experienced severe breathing
difficulties and cyanosis, and were unable to stand, euthanasia was
immediately carried out.
Assessment of neurological deficits
In most literature on cerebral ischemia-reperfusion
(30-36), Longa score is used to evaluate
neurological deficits. In the present study, neurofunctional
deficits were assessed using the slightly modified Longa score as
previously described (16,37): 0, no neurological deficit; 1, the
contralateral front paws cannot fully extend; 2, mice have
difficulty turning to the opposite side; 3, mice cannot turn to the
opposite side; and 4, mice are unable to walk independently and
have decreased consciousness. The scoring was blindly conducted by
two independent observers.
2,3,5-Triphenyltetrazolium chloride (TTC)
staining
TTC staining was used to evaluate the infarct volume
in mice after reperfusion. The brain tissues were cut into 2-mm
sections and cultured in 2% TTC solution (cat. no. 17779;
Sigma-Aldrich; Merck KGaA) at 37°C for 20 min. Then, the sections
were fixed with 4% paraformaldehyde (cat. no. P0099-3L; Beyotime
Institute of Biotechnology) for 2 h and images were captured with a
digital camera. Unstained white areas were considered infarct
areas, and stained red areas were considered non-infarct areas. The
infarct volume was analyzed by Image-Pro Plus 6.0 (Media
Cybernetics, Inc.). The infarct volume was calculated by summing
the infarct areas of all the sections and multiplying them by the
section thickness. To consider possible oedema, the following
formula (38) was used to
quantify the infarct volume (%) after oedema correction:
[(contrasting hemisphere volume-non-infarct area volume of aqueous
hemisphere)/contrasting hemisphere volume] ×100%. The measurements
were conducted blindly by two separate observers.
Hematoxylin and eosin (H&E)
staining
H&E staining (cat. no. C0105M; Beyotime
Institute of Biotechnology) was used to evaluate mouse brain
histopathological injury (39).
After reperfusion, the brain tissue (cortical area) samples were
fixed overnight in 4% paraformaldehyde at 4°C and embedded in
paraffin. Afterwards, the brain tissues were cut into 5-μm
sections, deparaffinized using xylene (cat. no. 016371.M1; Thermo
Fisher Scientific, Inc.) twice, with each treatment lasting 5-10
min, and rehydrated with gradient ethanol. Afterwards, the tissues
were stained with hematoxylin for 5 min, incubated with acetic acid
for 1 min, stained with eosin for 1 min, and dehydrated with
gradient ethanol. Finally, the tissues were sealed with neutral
resin and images were captured under a light microscope.
Cell culture and establishment of OGD/R
cells
As human derived cells are more closely related to
the clinical pathological environment, HBMVECs were selected as the
main cells by referring to previous studies (14-16,40-43). HBMVECs (RRID: CVCL_4D10)
(https://www.atcc.org/products/crl-3245) were obtained
from American Type Culture Collection (cat. no. CRL-3245) and
cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% fetal bovine serum (FBS; cat. no. C0237; Beyotime
Institute of Biotechnology) at 37°C with 5% CO2. The
culture medium was changed every 3 days, and cells at the 3rd to
6th passages were used in the present study. The cells were
validated by short tandem repeats and tested negative for
mycoplasma. To simulate cerebral I/R in vivo, as previously
described (16), the cells were
subjected to OGD/R. In brief, the cells were exposed to
glucose-free DMEM and cultured at 37°C with 95% N2/5%
CO2 for 2 h. Then, the glucose-free DMEM was replaced
with DMEM containing glucose, and the cells were cultured in a
normal state for 24 h for reoxidation. The control cells were
cultured under normal conditions.
Cell treatment
The miR-195-5p inhibitor (inhi-195) and
corresponding NC (inhi-NC) were synthesized by Guangzhou RiboBio
Co., Ltd. To overexpress RBM15 (accession no. NM_001201545),
IGF2BP3 (accession no. NM_006547), and USF2 (accession no.
NM_001321150), their full length was cloned and they were inserted
into the vector pcDNA3.1 (Shanghai GeneChem Co., Ltd.) to generate
the overexpression plasmids oe-RBM15, oe-IGF2BP3 and oe-USF2, with
the empty vector pcDNA3.1 (oe-NC) as a negative control. Small
interfering (si)RNAs targeting IGF2BP3 (si-IGF2BP3) and its
NC (si-NC) were purchased from Guangzhou RiboBio Co., Ltd. Then,
transfection was performed using Lipofectamine 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.). After 48 h of transfection. For
each transfection, 50 nmol/l of RNA duplexes were used. The
transfection efficiency was validated using reverse
transcription-quantitative PCR (RT-qPCR). To determine the dosage
of HM, a preliminary experiment was conducted. The cells were
treated with three different doses (10, 100, or 1,000 nM) of HM for
1 h (11), followed by OGD/R.
For NLRP3 agonist treatment, OGD/R was performed after 5 h of
treatment with 10 μM nigericin (Nig) (44). The relevant sequences are shown
in Table SI.
Cell Counting Kit-8 (CCK-8) assay
The cells (1×104) were seeded into a
96-well plate and cultured for 24 h (16). Afterwards, 10 μl of CCK-8
solution (cat. no. 96992; MilliporeSigma) was added to each well,
and the cells were incubated at 37°C for another 2 h. The optical
density at 450 nm was measured using a microplate reader (BioTek;
Agilent Technologies, Inc.).
Caspase-1 activity assay
Caspase-1 enzyme activity was measured using a
caspase-1 fluorescence assay kit (cat. no. ab39412; Abcam)
(45). Cells were cultured in
the corresponding manner, collected, and processed according to the
assay protocol. The cell suspension was resuspended in lysis buffer
and centrifuged (3,000 × g, 4°C, 5 min). Afterwards the supernatant
was collected. The sample wells and background wells were set in
black 96-well cell culture plates with clear bottoms. Afterwards,
the reaction mixture was prepared and added to each well along with
the fluorescent label, followed by incubation at 37°C for 1-2 h.
Finally, the fluorescence output was measured by a plate
reader.
Lactate dehydrogenase (LDH) release
Serum LDH levels in mice were detected using an LDH
assay kit (cat. no. C0016; Beyotime Institute of Biotechnology)
(39). In brief, the cells were
seeded into 96-well plates (3×104 cells/well) (18) and washed once with
phosphate-buffered saline (PBS) (cat. no. C0221A; Beyotime
Institute of Biotechnology) after the culture medium was removed.
Afterwards, 400 g of cell culture plate was removed and centrifuged
for 5 min. The supernatant (120 μl) was added to each well
of a new 96-well plate supplemented with 60 μl of LDH
detection solution and incubated at 37°C for 30 min. The absorbance
at 490 nm was measured using a microplate reader.
Immunofluorescence staining of NLRP3
After they were incubated with 0.3% Triton X-100 and
10% bovine serum albumin (BSA; cat. no. ST2254; Beyotime Institute
of Biotechnology) for 1 h (46),
the brain tissue (cortical area) sections were fixed with 4%
paraformaldehyde, infiltrated with 0.2% Triton X-100 for 10 min,
and blocked with 5% BSA at room temperature for 90 min (47). Afterwards, the cells were
incubated overnight with rabbit anti-NLRP3 (1:100; RRID:
AB_2809541; cat. no. MA5-32255; Thermo Fisher Scientific, Inc.) at
4°C and the corresponding goat anti-rabbit IgG H&L (Alexa
Fluor® 488; cat. no. ab150077; Abcam) at room
temperature for 2 h. DAPI (1 μg/ml) was used to stain the
cell nucleus. The images were captured under a fluorescence
microscope (BioTek; Agilent Technologies, Inc.).
Enzyme-linked immunosorbent assay
(ELISA)
After reperfusion, the brain tissues were ice sealed
with physiological saline and frozen at −20°C for 5 min. Then, the
brain tissues were centrifuged at 800 × g for 15 min to obtain the
supernatant. In the cell experiments, cells were isolated from the
culture medium and then centrifuged at 3,000 × g for 30 min to
obtain the supernatant (42). To
determine the release of S100B (48,49), inflammatory cytokines and
pyroptosis factors, mouse serum, brain tissue homogenate and cell
culture supernatant were collected, and the concentrations of S100B
(cat. no. ab285283), TNF-α (cat. nos. ab208348/ab181421), IL-6
(cat. nos. ab222503/ab178013), IL-4 (cat. nos. ab215089/ab100710),
IL-10 (cat. nos. ab255729/ab185986), IL-1β (cat. nos.
ab197742/ab214025) and IL-18 (cat. nos. ab216165/ab215539) were
measured using ELISA kits (Abcam). The absorbance at 450 nm was
measured using a microplate reader.
RT-qPCR
Total RNA was extracted from brain tissues or cells
using TRIzol reagent (cat. no. 15596026CN; Invitrogen; Thermo
Fisher Scientific, Inc.). RT-qPCR was performed to detect gene
expression. A FastQuant RT assay kit (Tiangen Biotech Co., Ltd.)
was used to synthesize complementary DNA (cDNA) according to the
manufacturer's instructions. RT-qPCR was performed on a real-time
thermal cycler (Applied Biosystems; Thermo Fisher Scientific, Inc.)
and using a SYBR Green I fluorescence kit (Takara Biotechnology
Co., Ltd.). The amplification mixture included 20 μl: 10
μl of SYBR Green mix, 0.4 μl of forward primer, 0.4
μl of reverse primer, 3 μl of cDNA, and 6.2 μl
of RNase-free water. The reaction conditions were as follows: 95°C
for 30 sec, 95°C for 15 sec, and 60°C for 30 sec for 40 cycles. The
primer sequences are shown in Table
I. With glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6
(18) as internal reference
genes, the fold difference in gene expression was calculated via
the 2−ΔΔCq method (50).
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Mouse |
|---|
| Gene name | Primer sequence
(5'-3') | Efficiency (%) |
|---|
| miR-195-5p | F:
GCGCGATAGCAGCACAGAAAT | 101 |
| R:
GTGTCGTGGAGTCGGCAATTC | |
| RBM15 | F:
TGCCAACCGGACACTTTTCT | 107 |
| R:
GCCATAGGTACTGGTCTGGC | |
| USF2 | F:
GTAGTCCAGGTGACTGATGGT | 109 |
| R:
GGATTTTGAATTACAGCCTGGGT | |
| NLRP3 | F:
GACCGTGAGGAAAGGACCAG | 109 |
| R:
GGCCAAAGAGGAATCGGACA | |
| GAPDH | F:
AGGTTCATCAGGTAAACTCAGG | 107 |
| R:
TTGATGGCAACAATCTCCACT | |
| U6 | F:
CGCTTCGGCAGCACATATACT | 100 |
| R:
CTTCACGAATTTGCGTGTCAT | |
| Human |
| miR-195-5p | F:
GGCGTAGGTAGCAGCACAGAA | 108 |
| R:
CTCAACTGGTGTCGTGGAGTC | |
| RBM15 | F:
GCCTTCCCACCTTGTGAGTT | 107 |
| R:
TCAACCAGTTTTGCACGGAC | |
| USF2 | F:
ACAAATGGAGGACAGACAGGA | 107 |
| R:
CCTCTCATCTCGGGGTGTTC | |
| IGF2BP3 | F:
CTGCACGGGAAACCCATAGA | 103 |
| R:
TCCCACTGTAAATGAGGCGG | |
| NLRP3 | F:
AGAAGCTCTGGTTGGTCAGC | 100 |
| R:
CAAGGCATTCTCCCCCACAT | |
| NLRP3 promoter | F:
GCCTGCCACATACCAGCCATT | 96 |
| R:
GTCCTCTCACAGCAAGATGGCT | |
| GAPDH | F:
GTCAAGGCTGAGAACGGGAA | 100 |
| R:
TCGCCCCACTTGATTTTGGA | |
| U6 | F:
CTCGCTTCGGCAGCACATATA | 103 |
| R:
TGGAACGCTTCACGAATTTGC | |
Western blotting
The protein was extracted using
radioimmunoprecipitation assay (RIPA) buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology) and centrifuged at 12,000 × g
for 30 min to collect the supernatant. The protein concentration
was analyzed using a bicinchoninic acid kit (cat. no. P0010;
Beyotime Institute of Biotechnology). Afterwards, the protein (20
μg/lane) was subjected to 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred onto a
polyvinylidene fluoride membrane (39). The membrane was blocked with 5%
skim milk at room temperature for 1 h. After it was washed with
Tris-buffered saline with Tween (TBST) solution (20 mmol/l
Tris-HCl, 5% non-fat milk, 150 mmol/l NaCl, and 0.05% Tween-20, pH
7.5) (5 min, 3 times), the membranes were incubated with rabbit
anti-RBM15 (1:1,000; cat. no. ab315456; Abcam), rabbit anti-USF2
(1:2,000; cat. no. ab264330; Abcam), rabbit anti-IGF2BP3 (1:500;
RRID: AB_2808308; cat. no. PA5-96506; Thermo Fisher Scientific,
Inc.), rabbit anti-NLRP3 (1:500; cat. no. MA5-32255; Thermo Fisher
Scientific, Inc.), rabbit anti-GSDMD-N (1:2,000; RRID: AB_3663000;
cat. no. YT7991; ImmunoWay Biotechnology Company), rabbit
anti-cleaved caspase-1 (1:500; RRID: AB_2818323; cat. no.
PA5-99390; Thermo Fisher Scientific, Inc.), rabbit anti-caspase-1
(1:1,000; RRID: AB_2888675; cat. no. ab138483; Abcam) and rabbit
anti-β-actin (1:1,000; RRID: AB_2305186; cat. no. ab8227; Abcam) at
4°C overnight. After they were washed with TBST (5 min, 3 times),
the membranes were incubated with the corresponding goat
anti-rabbit secondary antibody (1:2,000; RRID: AB_2819160; cat. no.
ab205718; Abcam) at room temperature for 1 h. The membranes were
then washed with TBST (5 min, 3 times). Finally, the signal was
detected by enhanced chemiluminescence (cat. no. 32132; Thermo
Fisher Scientific Inc.), with β-actin used as a control. The film
was exposed in a dark room, fixed, scanned with an Epson scanner
(EPSON, V19ii/V39ii), and finally analyzed using Image-Pro Plus
v6.0 (Media Cybernetics, Inc.).
RNA immunoprecipitation (RIP)
The cells were lysed in RIPA lysis buffer (cat. no.
20-188; Sigma-Aldrich; Merck KGaA) containing an RNase inhibitor
and a protein inhibitor for 30 min in an ice bath and then
centrifuged at 16,000 × g for 10 min at 4°C to obtain the
supernatant. The preequilibrated protein A/G Sepharose beads were
mixed with an appropriate amount of mouse anti-m6A (1:100; RRID:
AB_3674612; cat. no. MABE1006; Sigma-Aldrich; Merck KGaA), rabbit
anti-IGF2BP3 (1:100; cat. no. PA5-96506; Thermo Fisher Scientific,
Inc.) or rabbit anti-USF2 (1:100; cat. no. ab264330; Abcam) and
rotated at 4°C for ~6 h. The supernatant was added to the
bead-antibody mixture and rotated overnight at 4°C, with IgG
(1:100; RRID: AB_2687931; cat. no. ab172730; Abcam) used as the
control. The mixed solution was centrifuged at 4°C and 100 × g for
10 sec to remove the supernatant, after which it was washed twice
with cold low-salt rinse solution in an ice bath (5 min each time)
and then washed twice with high-salt rinse solution (5 min each
time). RNA was extracted and subjected to RT-qPCR. The lysate was
incubated overnight at 4°C with a specific anti-Ago2 antibody
(1:100; RRID: AB_2687492; cat. no. ab156870; Abcam) or control IgG
(1:100; cat. no. ab172730; Abcam). After it was washed, the
immunoprecipitated RNA was isolated by treatment with proteinase K
and extraction with phenol/chloroform. The purified RNA was reverse
transcribed, and the target RNAs (miR-195-5p and RBM15) were
evaluated by RT-qPCR analysis (39).
m6A quantitative analysis
Total RNA was extracted using TRIzol reagent, and
the m6A RNA level of total RNA was measured using a m6A RNA
methylation quantification kit (cat. no. ab185912; Abcam) (51). The detection well was coated with
RNA, followed by the addition of a capture antibody, detection
antibody, and enhancer solution separately. Finally, the
colorimetric solution was added, and the absorbance was measured at
450 nm. The m6A level was quantitatively analyzed by measuring the
absorbance of each well.
Actinomycin D treatment
Actinomycin D (5 μg/ml; cat. no. A4262;
Sigma-Aldrich; Merck KGaA) was added to the culture medium to block
cell transcription (52), and
the USF2 mRNA level was analyzed by RT-qPCR.
Chromatin immunoprecipitation (ChIP)
At room temperature, the cells were crosslinked with
1% formaldehyde (cat. no. 47608; Sigma-Aldrich; Merck KGaA) for 10
min. After they were washed 4 times with 20 ml of PBS in a 50 ml
conical tube, the cells were scraped and incubated on ice for 10
min. After they were centrifuged at 2,000 × g for 5 min, the cells
were ultrasonicated by a Branson 150 Ultrasonic Processor. For
ChIP, anti-USF2 antibody (1:100; cat. no. ab264330; Abcam) and
anti-IgG antibody (1:100; cat. no. ab172730; Abcam) were used as
negative controls. Specific primers (Table I) were used for PCR amplification
of the USF2 binding region of the NLRP3 promoter (29).
Dual-luciferase assay
To investigate the interaction between miR-195-5p
and RBM15, wild-type (WT) and mutant (MUT) RBM15 3'-untranslated
regions (UTRs) containing putative binding sites for miR-195-5p
were constructed. miR-195-5p mimics or NC mimics and reporter
vectors (pmirGLO vector; Promega Corporation) containing WT or MUT
RBM15 3'-UTRs were co-transfected into cells using Lipofectamine
3000. Similarly, to investigate the regulation of the NLRP3
promoter by USF2, WT and MUT NLRP3 promoter sequences containing
the putative binding site of USF2 were constructed based on
the psiCHECK-2 vector (Promega Corporation) and co-transfected with
oe-USF2 or oe-NC using Lipofectamine 3000. Similarly, to
investigate the regulation of the USF2 m6A site by
RBM15 and IGF2BP3, WT and MUT sequences containing
predicted USF2 m6A sites were constructed based on the psiCHECK-2
vector and co-transfected with oe-RBM15, oe-IGF2BP3, or oe-NC using
Lipofectamine 3000. After 24 h of cultivation, the luciferase
activity was measured using the luciferase assay system (Ambion;
Thermo Fisher Scientific, Inc.). The ratio of the luminescence of
firefly luciferase to that of Renilla luciferase was
calculated (46).
Statistical analysis
Data analysis and map plotting were performed using
SPSS 21.0 (IBM Corp.) and GraphPad Prism 8.0 (GraphPad Software
Inc.; Dotmatics). All the data were tested for normality using the
Shapiro-Wilk test. If the data between multiple groups did not
follow a normal distribution, a non-parametric test (Kruskal-Wallis
test) was performed, followed by Dunn's multiple comparisons test.
If the data between multiple groups followed a normal distribution,
the Brown-Forsythe test was used for the homogeneity of variance
test. To ensure homogeneity of variance, one-way analysis of
variance (ANOVA) or two-way ANOVA was performed, with Tukey's
multiple comparisons test or Sidak's multiple comparisons test for
post hoc testing. If the data did not follow a normal distribution,
a non-parametric test (Mann-Whitney test) was performed. If the
data between two groups followed a normal distribution, a t-test
was performed. The homogeneity of variance was determined using an
F test. If equal variances were not assumed, the Welch t-test was
performed again. P<0.05 was considered to indicate a
statistically significant difference.
Results
HM preconditioning alleviates cerebral
I/R inflammatory injury in mice and represses NLRP3-mediated
pyroptosis
HM can protect against CIRI (12), but its mechanism is unclear.
Middle cerebral artery occlusion/reperfusion (MCAO/R) mice were
established and treated with HM (Fig. 1A). Compared with those in the
sham group, the serum LDH and S100B levels in the IR group
increased, whereas compared with those in the I/R + saline group,
the serum LDH and S100B levels in the I/R + HM group decreased
(P<0.01; Fig. 2A). After
MCAO/R, the infarct volume of the mice increased, but HM
preconditioning significantly reduced the infarct volume of the
ischemic mice (P<0.01; Fig.
2B). As shown in Fig. 2C, HM
preconditioning exerted neuroprotective effects (P<0.01).
H&E staining revealed no significant abnormalities in the brain
tissues of the sham-operated mice, whereas infarction accompanied
by inflammatory cell infiltration was detected in the brain tissues
of the MCAO/R-treated mice, but HM preconditioning improved these
symptoms (Fig. 2D). After
MCAO/R, the concentrations of the inflammatory factors TNF-α and
IL-6 in brain tissues increased, whereas the concentrations of IL-4
and IL-10 decreased. However, HM preconditioning reversed these
trends (P<0.01; Fig. 2E). HM
inhibited pyroptosis (13), and
the present results demonstrated that HM preconditioning
significantly decreased the expression levels of NLRP3, GSDMD-N,
caspase-1 and cleaved caspase-1 in brain tissues (P<0.01;
Fig. 2F and G). Moreover, HM
preconditioning also led to significant decreases in the
concentrations of the pyroptosis factors IL-1β and IL-18
(P<0.01; Fig. 2H). These
results indicate that HM preconditioning alleviates cerebral I/R
injury in mice and represses NLRP3-mediated pyroptosis.
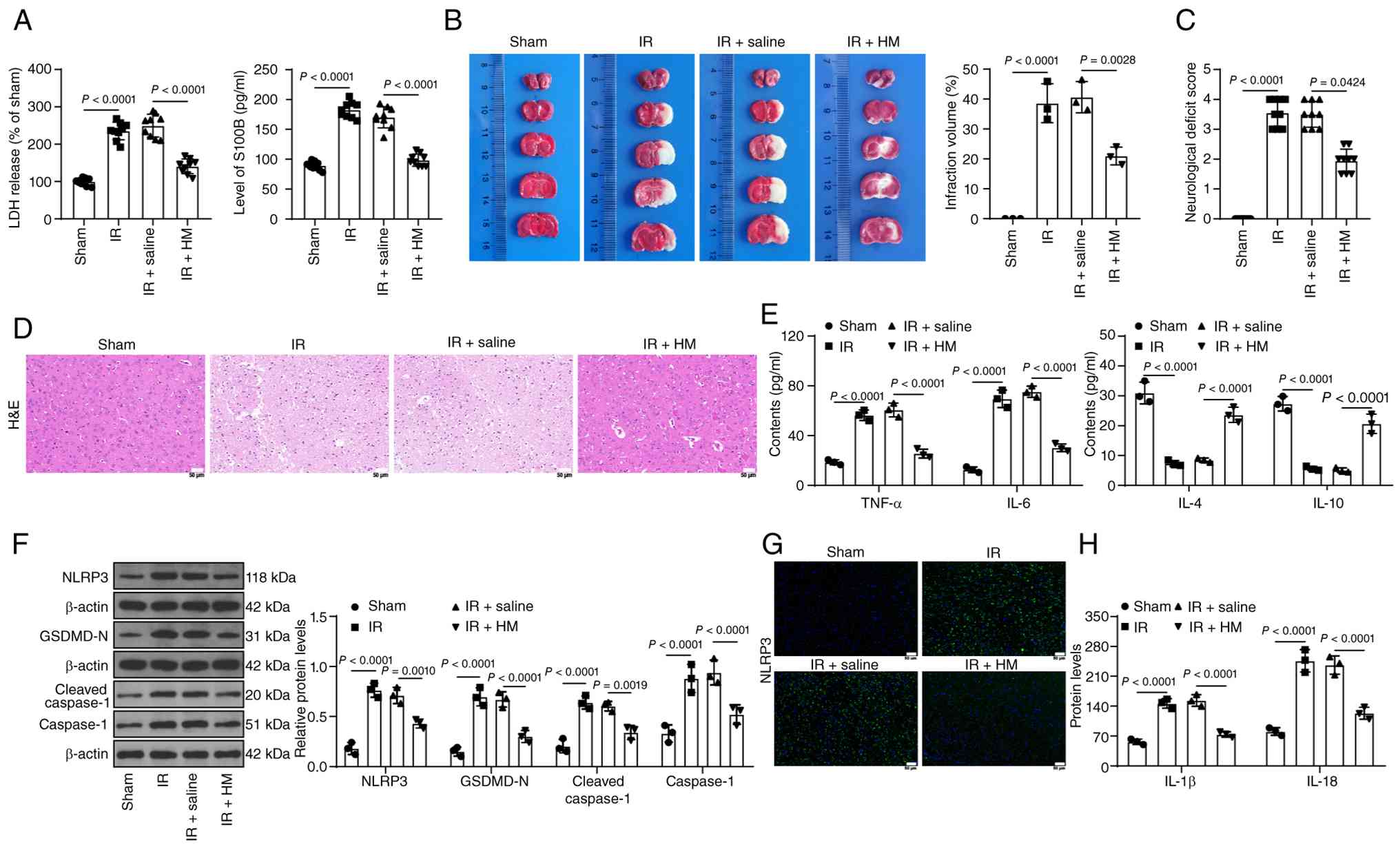 | Figure 2HM preconditioning alleviates
cerebral I/R inflammatory injury in mice and represses
NLRP3-mediated pyroptosis. The mice were treated according to the
process shown in Fig. 1A. (A)
LDH and S100B levels in the serum of each group of mice. (B)
2,3,5-triphenyltetrazolium chloride staining for detecting infarct
size in brain tissue. (C) Neurological function scores of the mice
in each group. (D) H&E staining for detecting brain tissue
injury; magnification, ×200. (E) ELISA detection of inflammatory
factors in brain tissues. (F) Western blot analysis of the protein
expression levels of NLRP3, GSDMD-N, caspase-1 and cleaved
caspase-1 in brain tissues. (G) Immunofluorescence staining for
detecting the percentage of NLRP3-positive brain tissues;
magnification, ×200. (H) ELISA detection of pyroptosis factors in
brain tissues. Animal experiments in A and C: n=9; others: n=3. The
data are expressed as the mean ± standard deviation. Comparisons
between multiple groups in panel C were conducted using the
Kruskal-Wallis test, and Dunn's multiple comparisons test was used
for post hoc testing. The data in panels A and B were analyzed via
one-way ANOVA, and the data in panels E, F and H were analyzed via
two-way ANOVA, followed by Tukey's multiple comparisons test. HM,
hydromorphone; I/R, ischemia/reperfusion; NLRP3, Nod-like receptor
protein 3; LDH, lactate dehydrogenase; GSDMD, gasdermin D; IL,
interleukin; TNF-α, tumor necrosis factor alpha. |
HM preconditioning alleviates
OGD/R-induced inflammatory injury by repressing NLRP3-mediated
pyroptosis
Next, HBMVECs were induced with OGD/R (Fig. 1B) and a significant reduction in
cell viability and an increase in LDH levels were observed, whereas
HM preconditioning alleviated cell injury (P<0.01; Fig. 3A and B). HBMVECs were
subsequently treated with 100 nM HM for further experiments. HM
preconditioning reduced TNF-α and IL-6 levels induced by OGD/R but
increased IL-4 and IL-10 levels inhibited by OGD/R (P<0.01;
Fig. 3C). OGD/R upregulated the
expression levels of NLRP3, GSDMD-N, caspase-1 and cleaved
caspase-1, as well as the concentrations of IL-1β and IL-18 and the
activity of caspase-1, but HM preconditioning decreased the
expression levels of these factors (P<0.01; Fig. 3D-G). Furthermore, the NLRP3
agonist Nig was used in combination with HM. Compared with HM
preconditioning alone, the combined treatment significantly
increased pyroptosis but decreased cell viability, elevated LDH
levels, and increased cell inflammation (P<0.01; Fig. 3A-G). Briefly, HM preconditioning
alleviates OGD/R-induced inflammatory injury by repressing
NLRP3-mediated pyroptosis.
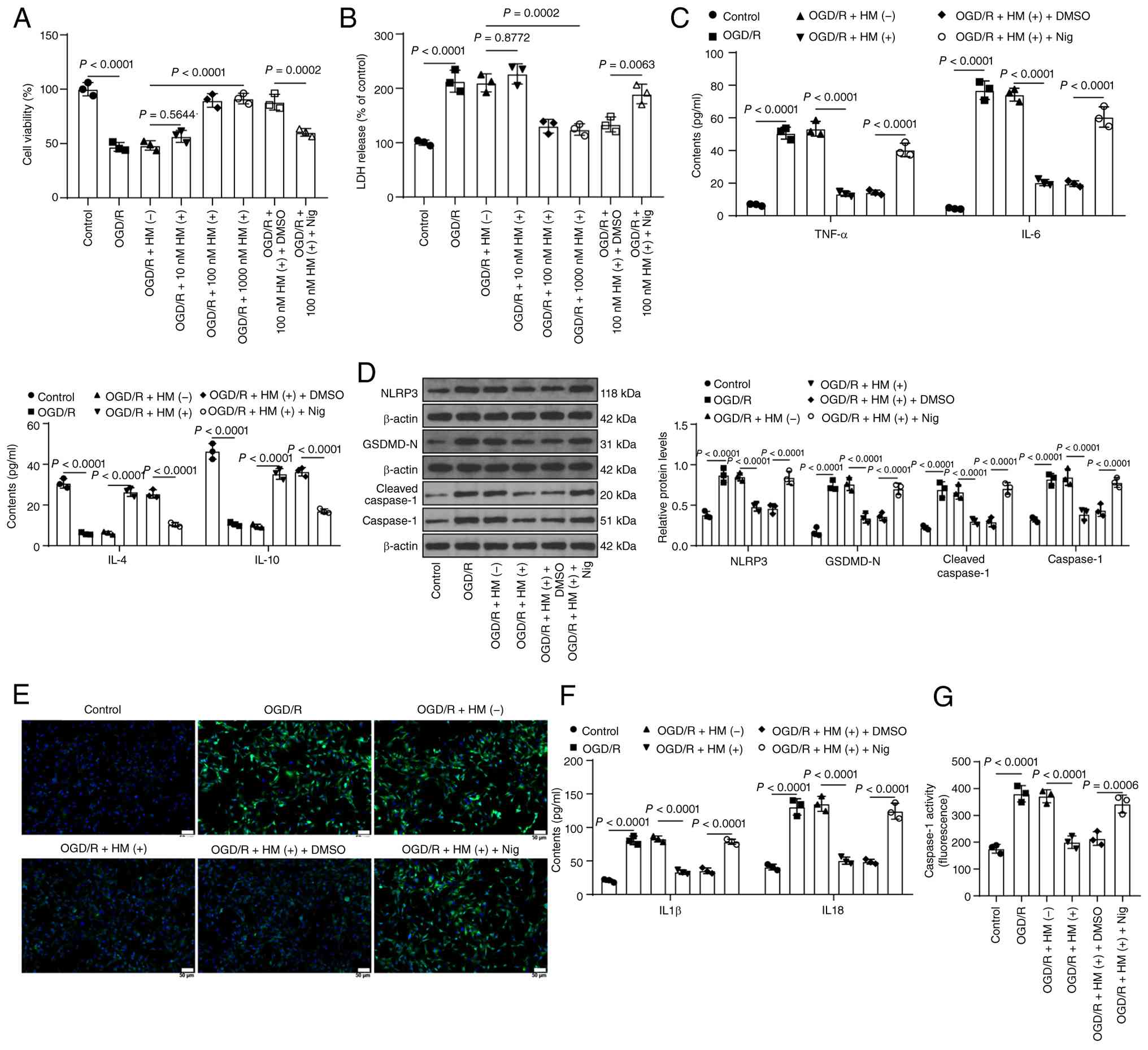 | Figure 3HM preconditioning alleviates
OGD/R-induced inflammatory injury by repressing NLRP3-mediated
pyroptosis. Cells were treated according to the process shown in
Fig. 1B. (A) Cell Counting Kit-8
assay detection of cell viability. (B) LDH levels in each group of
cells. (C) ELISA detection of inflammatory factors in cells. (D)
Western blot analysis of the protein expression levels of NLRP3,
GSDMD-N, caspase-1 and cleaved caspase-1 in cells. (E)
Immunofluorescence staining for detecting the percentage of
NLRP3-positive cells; magnification, ×200. (F) ELISA detection of
pyroptosis factors in cells. (G) Detection of caspase 1 enzyme
activity using a caspase 1 fluorescence assay kit. A total of three
technical replicates of the cell experiments were performed. The
data are expressed as the mean ± standard deviation. The data in
panels A, B and G were analyzed by one-way ANOVA, and the data in
panels C, D and F were analyzed by two-way ANOVA, followed by
Tukey's multiple comparisons test. HM, hydromorphone; OGD/R,
oxygen-glucose deprivation/reperfusion; NLRP3, Nod-like receptor
protein 3; GSDMD, gasdermin D; Nig, nigericin; IL, interleukin;
TNF-α, tumor necrosis factor alpha. |
HM preconditioning alleviates
OGD/R-induced inflammatory injury by upregulating the expression of
miR-195-5p to suppress NLRP3-mediated pyroptosis
HM can regulate miRNA expression (53). miR-195-5p expression is reduced
during CIRI (16,18), and miR-195-5p is associated with
pyroptosis (19). It was
hypothesized that miR-195-5p may be a downstream miRNA target of
HM. The expression of miR-195-5p decreased in the model group but
increased in the HM preconditioning group (P<0.01; Fig. 4A). Considering that the
pretreatment effect of HM has been verified in Fig. 3 (the negative control of HM is
demonstrated in Fig. 3), the
combined experiment was set up on the basis of Fig. 3. miR-195-5p expression was
inhibited in HBMVECs, followed by OGD/R treatment in combination
with HM (P<0.01; Fig. 4A and
B). After inhibition of miR-195-5p expression, cell viability
decreased, and LDH levels increased (P<0.01; Fig. 4C and D). Moreover, inhibition of
miR-195-5p led to increases in the concentrations of TNF-α and IL-6
in cells, decreases in the concentrations of IL-4 and IL-10
(P<0.01; Fig. 4E), and
significant increases in pyroptosis (P<0.01; Fig. 4F-I). These results suggest that
HM preconditioning alleviates OGD/R-induced inflammatory injury by
upregulating the expression of miR-195-5p to repress NLRP3-mediated
pyroptosis.
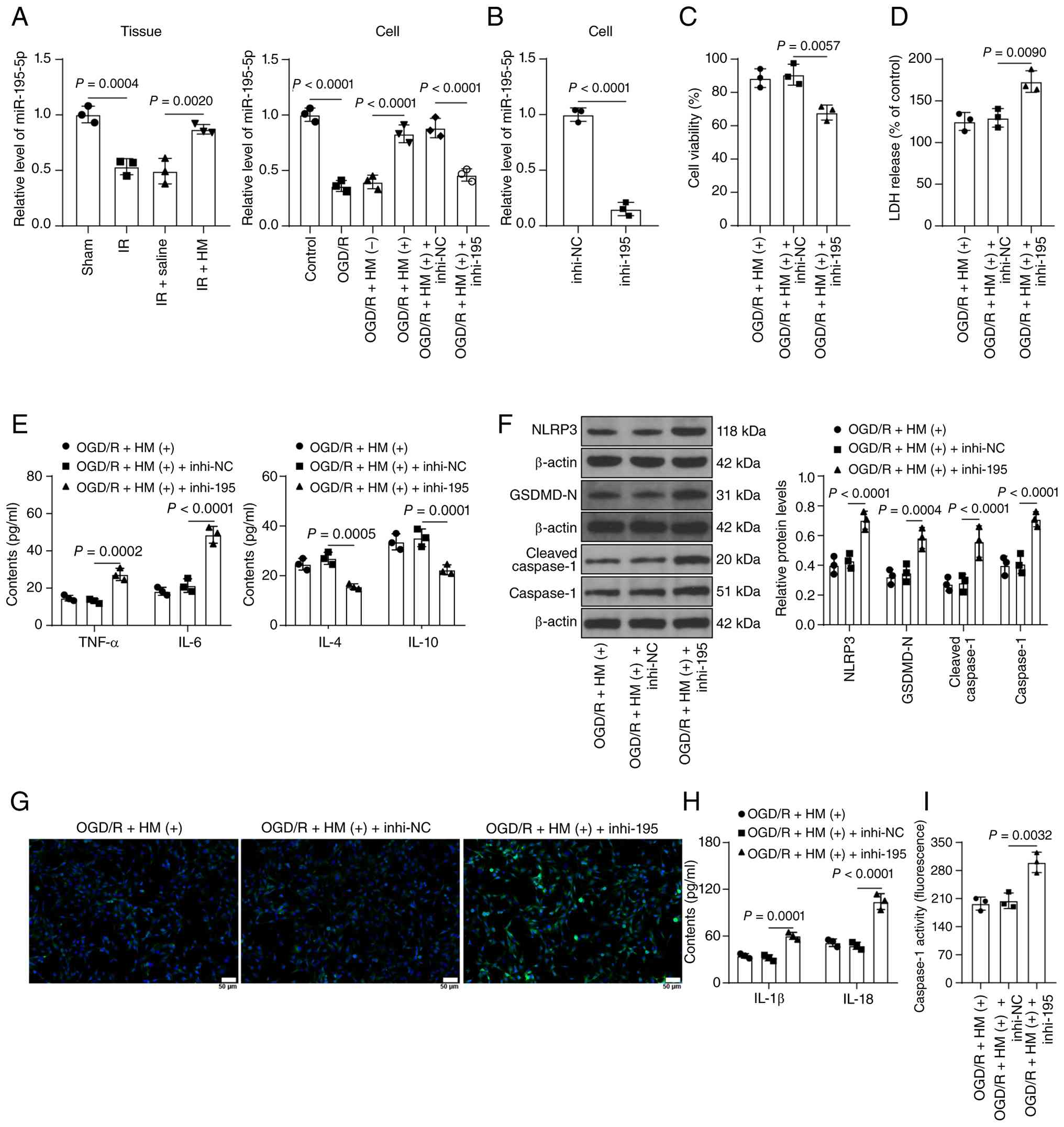 | Figure 4HM preconditioning alleviates
OGD/R-induced inflammatory injury by upregulating the expression of
miR-195-5p to suppress NLRP3-mediated pyroptosis. (A) RT-qPCR
detection of the expression of miR-195-5p in brain tissues and
cells; a miR-195-5p inhibitor (inhi-195) was transfected into human
brain microvascular endothelial cells, with NC (inhi-NC) using as
the negative control. (B) RT-qPCR detection of the transfection
efficiency of miR-195-5p in cells. (C) Cell Counting Kit-8 assay
detection of cell viability in each group. (D) LDH levels in each
group of cells. (E) ELISA detection of inflammatory factors in
cells. (F) Western blot analysis of the protein expression levels
of NLRP3, GSDMD-N, caspase-1 and cleaved caspase-1 in cells. (G)
Immunofluorescence staining for detecting the percentage of
NLRP3-positive cells; magnification, ×200. (H) ELISA detection of
pyroptosis factors in cells. (I) Detection of caspase 1 enzyme
activity using a caspase 1 fluorescence assay kit. Animal
experiments: n=3. Three technical replicates of the cell
experiments were performed. The data are expressed as the mean ±
standard deviation. The data in panel B were analyzed via a t-test.
The data in panels A, C, D and I were analyzed via one-way ANOVA,
and the data in panels E, F and H were analyzed via two-way ANOVA,
followed by Tukey's multiple comparisons test. HM, hydromorphone;
OGD/R, oxygen-glucose deprivation/reperfusion; miR, microRNA;
NLRP3, Nod-like receptor protein 3; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; LDH, lactate
dehydrogenase; GSDMD, gasdermin D; I/R, ischemia/reperfusion; IL,
interleukin; TNF-α, tumor necrosis factor alpha. |
miR-195-5p targets RBM15 and reduces USF2
expression, thus transcriptionally inhibiting NLRP3 expression
The downstream target genes of miR-195-5p were
predicted (Fig. 1C) and it was
reported that RBM15 was upregulated in cerebral I/R (21), but its role remains unclear.
Therefore, RBM15 was chosen as the downstream target gene of
miR-195-5p. RIP and dual-luciferase assays confirmed the target
binding of miR-195-5p and the RBM15 3'-UTR (P<0.01; Fig. 5A-C). RBM15 expression increased
in the model group but decreased in the HM preconditioning group
(P<0.01; Fig. 5D-G), and
after inhibition of miR-195-5p, RBM15 expression increased again
(P<0.01; Fig. 5E and G).
RBM15 functions as a m6A methyltransferase, and USF2 has m6A
modification levels (Fig. 1D).
USF2 expression is increased in cerebral I/R (29). Therefore, USF2 was chosen as a
downstream factor of RBM15. It was found that the change in m6A
content was consistent with the trend in RBM15 expression
(P<0.01; Fig. 5H) and was
accompanied by the enrichment of m6A in USF2 (P<0.01; Fig. 6A). Furthermore, a dual-luciferase
assay was performed to determine the binding of RBM15 to USF2 m6A.
The results revealed that the luciferase activity of the oe-RBM15
group was significantly greater than that of the oe-NC group
(P<0.01; Fig. 6B). Moreover,
overexpression of RBM15 significantly increased the mRNA stability
and expression of USF2 (P<0.01; Fig. 6C-G). As a m6A-reading protein,
IGF2BP3 often participates in the regulation of m6A modification by
RBM15 (52). Similarly, the
luciferase activity of the oe-IGF2BP3 group was significantly
greater than that of the oe-NC group (P<0.01; Fig. 6B-D). After silencing of IGF2BP3,
the mRNA stability and expression of USF2 decreased (P<0.01;
Fig. 6C-G). USF2 expression
increased in the model group but decreased in the HM
preconditioning group. However, after inhibition of miR-195-5p,
USF2 expression increased again (P<0.01; Fig. 5D-G). USF2, a transcription
factor, is predicted by the JASPAR database to bind to the NLRP3
promoter (Fig. 1E). The ChIP
results revealed that the enrichment trend of USF2 in the NLRP3
promoter was consistent with the expression trends of USF2 and
NLRP3 (P<0.01; Fig. 6H).
Dual-luciferase assays confirmed the binding of USF2 to the NLRP3
promoter (P<0.01; Fig. 5B),
and the expression trend of NLRP3 was consistent with that of USF2
(P<0.01; Fig. 5D and E). In
short, miR-195-5p targets RBM15 and reduces USF2 expression through
IGF2BP3-mediated m6A modification, ultimately leading to the
transcriptional inhibition of NLRP3 expression.
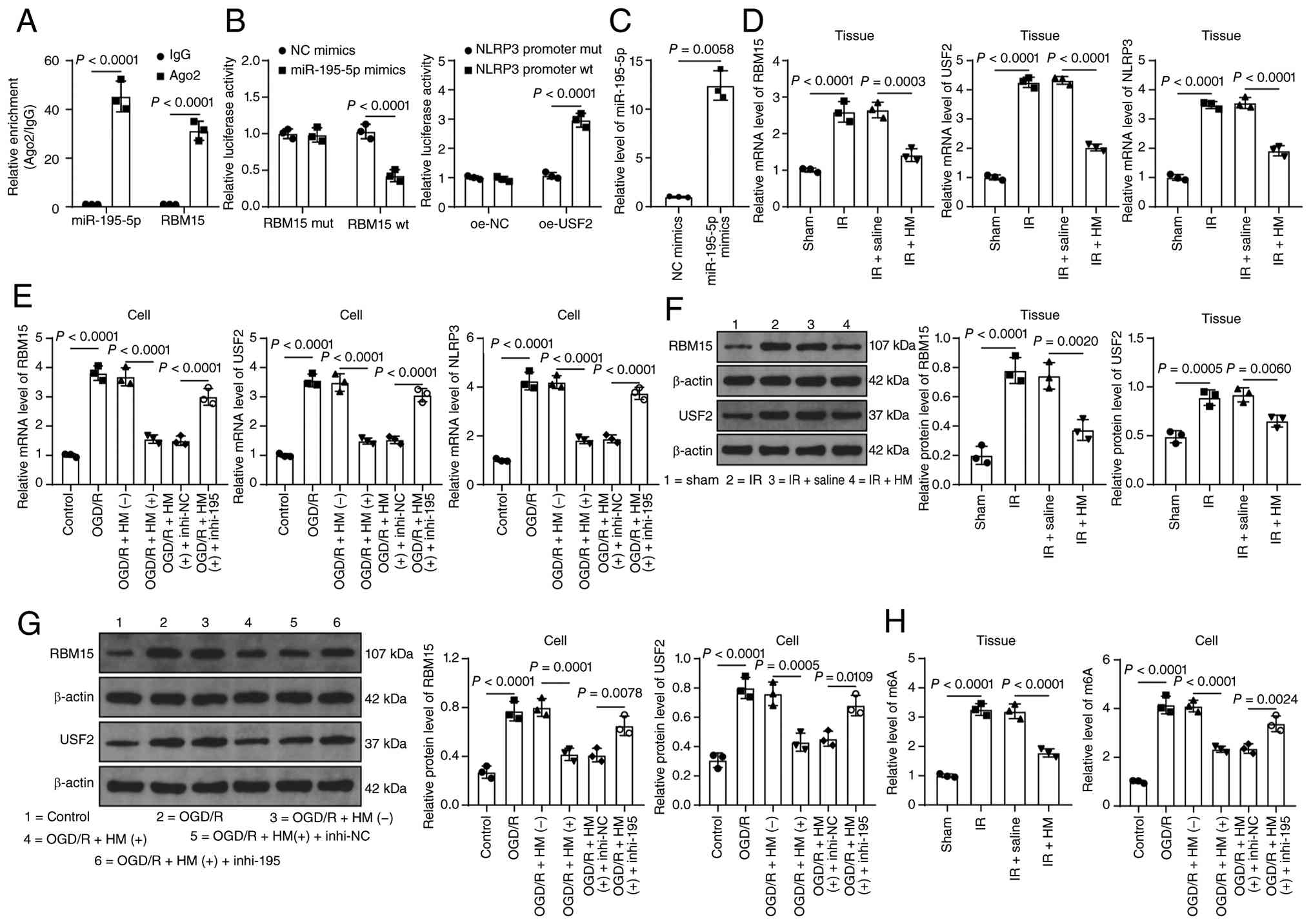 | Figure 5miR-195-5p targets RBM15 and reduces
USF2 expression. (A) RNA immunoprecipitation detection of the
binding relationship between miR-195-5p and RBM15. (B) Verification
of the binding relationship by dual-luciferase assay. (C) RT-qPCR
detection of miR-195-5p expression. (D and E) RT-qPCR detection of
the mRNA expression levels of RBM15, USF2 and NLRP3 in brain
tissues and cells. (F and G) Western blot detection of the protein
expression levels of RBM15 and USF2 in brain tissues and cells. (H)
Quantitative analysis of m6A content in brain tissues and cells.
Animal experiments: n=3. A total of three technical replicates of
the cell experiments were performed. The data are expressed as the
mean ± standard deviation. The data comparisons between two groups
in panel C were conducted using the Welch t-test. Comparisons
between multiple groups in panels A and B were conducted using
two-way ANOVA, and Sidak's multiple comparisons test was used for
post hoc testing. The data in panels D-H were analyzed via one-way
ANOVA, followed by Tukey's multiple comparisons test. miR,
microRNA; RBM15, RNA-binding motif protein 15; USF2, upstream
stimulatory factor 2; RT-qPCR, reverse transcription-quantitative
PCR; NLRP3, Nod-like receptor protein 3; m6A, N6-methyladenosine;
HM, hydromorphone; OGD/R, oxygen-glucose deprivation/reperfusion;
I/R, ischemia/reperfusion; NC, negative control; oe-,
overexpression; WT, wild-type; MUT, mutant. |
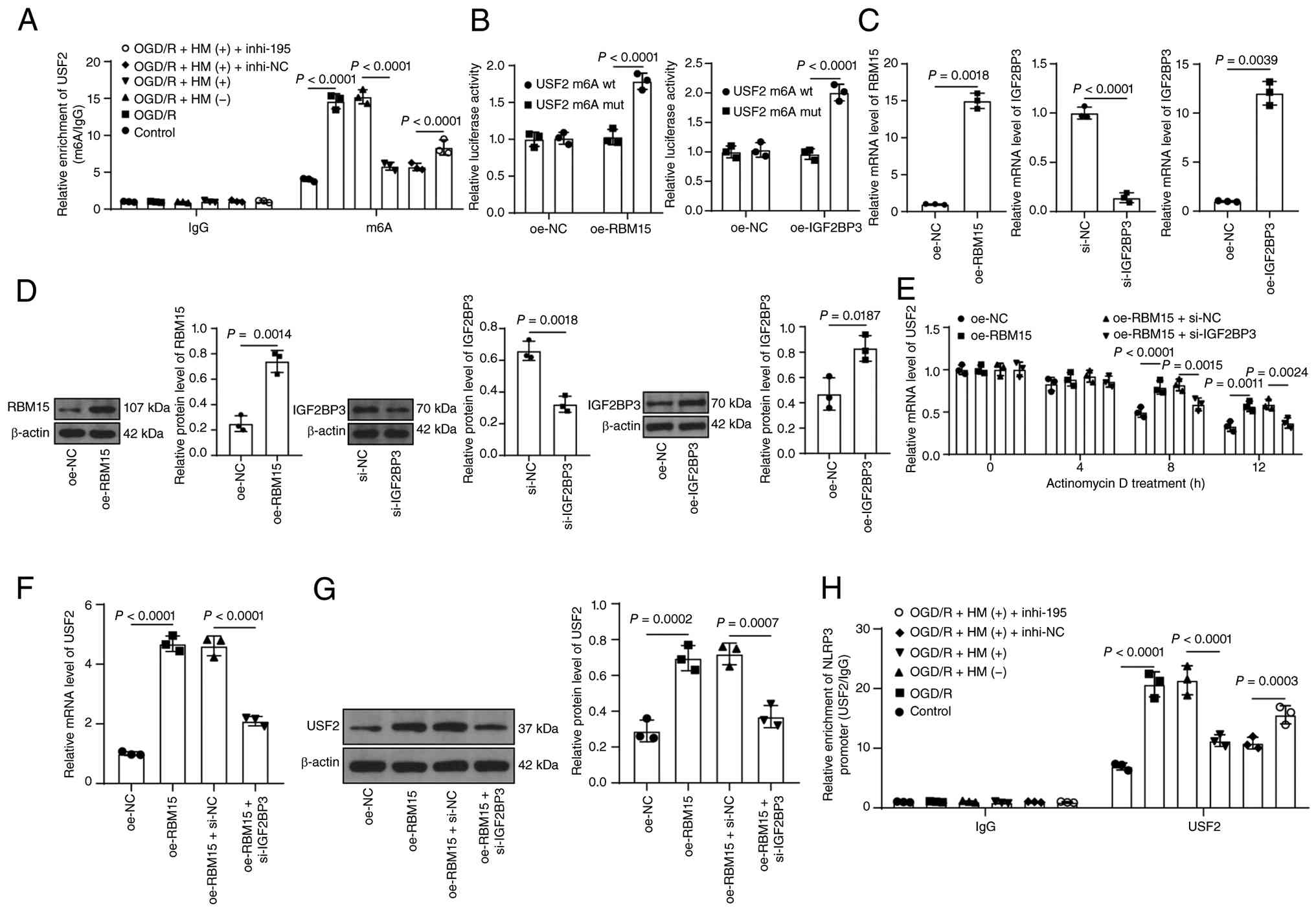 | Figure 6miR-195-5p inhibits NLRP3 expression
via the RBM15/USF2 axis. (A) RNA immunoprecipitation analysis of
USF2 m6A modification in each group of cells. (B) Dual-luciferase
assay for detecting the binding of RBM15 and IGF2BP3 to USF2 m6A.
(C and D) RT-qPCR and western blot analysis of the transfection
efficiency of RBM15 and IGF2BP3 in cells. (E) RT-qPCR detection of
the mRNA stability of USF2 in cells. (F and G) The expression of
USF2 in cells was detected by RT-qPCR and western blotting. (H)
Chromatin immunoprecipitation analysis of the enrichment of USF2 on
the NLRP3 promoter in each group of cells. Animal experiments: n=3.
Three technical replicates of the cell experiments were performed.
The data are expressed as the mean ± standard deviation. The data
comparisons between two groups in panel C (left, right) were
conducted using the Welch t-test, while the data comparisons
between two groups in panels C (middle) and D were conducted using
the t-test. Comparisons between multiple groups in panel B were
conducted using two-way ANOVA, and Sidak's multiple comparisons
test was used for post hoc testing. The data in panels F and G were
analyzed via one-way ANOVA, and the data in panels A, E and H were
analyzed via two-way ANOVA, followed by Tukey's multiple
comparisons test. miR, microRNA; NLRP3, Nod-like receptor protein
3; RBM15, RNA-binding motif protein 15; USF2, upstream stimulatory
factor 2; RT-qPCR, reverse transcription-quantitative PCR; IGF2BP3,
insulin-like growth factor 2 mRNA-binding protein 3; m6A,
N6-methyladenosine; HM, hydromorphone; OGD/R, oxygen-glucose
deprivation/reperfusion; si-, small interfering; NC, negative
control; oe-, overexpression. |
Overexpression of RBM15/USF2 abolishes
the protective effect of HM preconditioning on OGD/R-induced
pyroptosis
To verify the role of RBM15 in the ability of HM to
protect against OGD/R-induced cell injury, the combined experiment
was set up on the basis of Fig.
3 as the pretreatment effect of HM has been verified in
Fig. 3 (the negative control of
HM is shown in Fig. 3). RBM15
expression was upregulated in cells for a combined treatment with
HM (P<0.05; Fig. 7A and C).
As a result, the m6A content increased, and the m6A enrichment of
USF2 in cells increased (P<0.05; Fig. 7E and F). After overexpression of
RBM15, USF2 expression increased, and enrichment of USF2 in the
NLRP3 promoter increased (P<0.05; Fig. 7A, D and G). NLRP3 expression also
increased accordingly (P<0.05; Fig. 7A, D and H). In addition,
overexpression of RBM15 led to a decrease in cell viability, an
increase in LDH levels, and a significant increase in inflammatory
and pyroptosis levels (P<0.05; Figs. 7D and H; 8A-E), alleviating the protective effect
of HM preconditioning on OGD/R-induced pyroptosis.
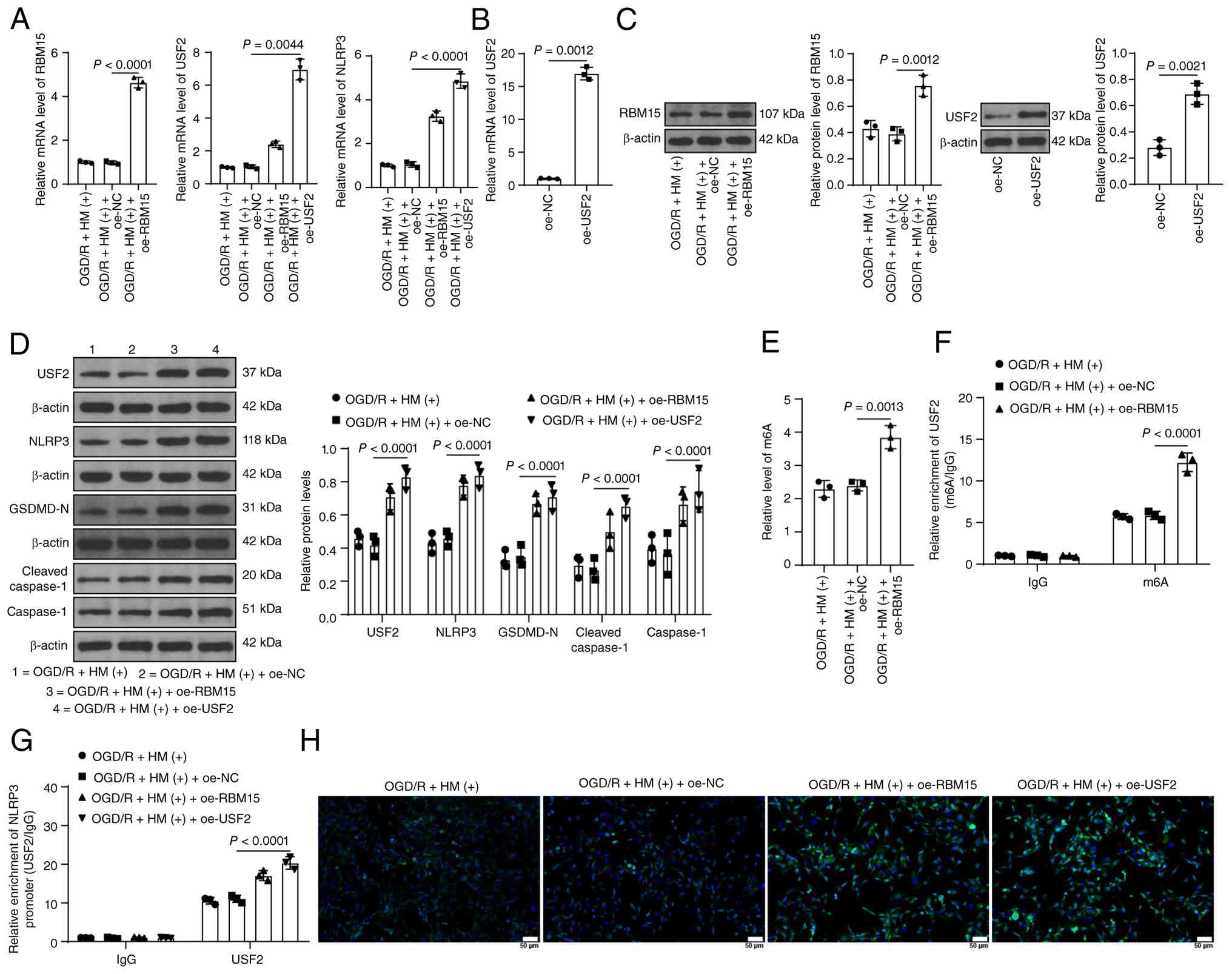 | Figure 7RBM15 promotes NLRP3 expression by
upregulating USF2. The overexpression plasmids oe-RBM15 and oe-USF2
were transfected into human brain microvascular endothelial cells,
with the empty vector pcDNA3.1 (oe-NC) as a negative control. (A)
RT-qPCR detection of the mRNA expression levels of RBM15, USF2 and
NLRP3 in cells. (B) RT-qPCR detection of the transfection
efficiency of USF2 in cells. (C and D) Western blot detection of
protein expression. (E) Quantitative analysis of m6A content in
cells. (F) RNA immunoprecipitation analysis of USF2 m6A
modification in each group of cells. (G) Chromatin
immunoprecipitation analysis of the enrichment of USF2 on the NLRP3
promoter in each group of cells. (H) Immunofluorescence staining
for detecting the percentage of NLRP3-positive cells;
magnification, ×200. A total of three technical replicates of the
cell experiments were performed. The data are expressed as the mean
± standard deviation. The data in panel B were analyzed via the
Welch t-test, while the data comparisons between two groups in
panel C (right) were conducted using the t-test. The data in panels
A, C (left) and E were analyzed via one-way ANOVA, and the data in
panels D, F and G were analyzed via two-way ANOVA, followed by
Tukey's multiple comparisons test. RBM15, RNA-binding motif protein
15; NLRP3, Nod-like receptor protein 3; USF2, upstream stimulatory
factor 2; oe-, overexpression; NC, negative control; RT-qPCR,
reverse transcription-quantitative PCR; m6A, N6-methyladenosine;
HM, hydromorphone; OGD/R, oxygen-glucose deprivation/reperfusion;
GSDMD, gasdermin D. |
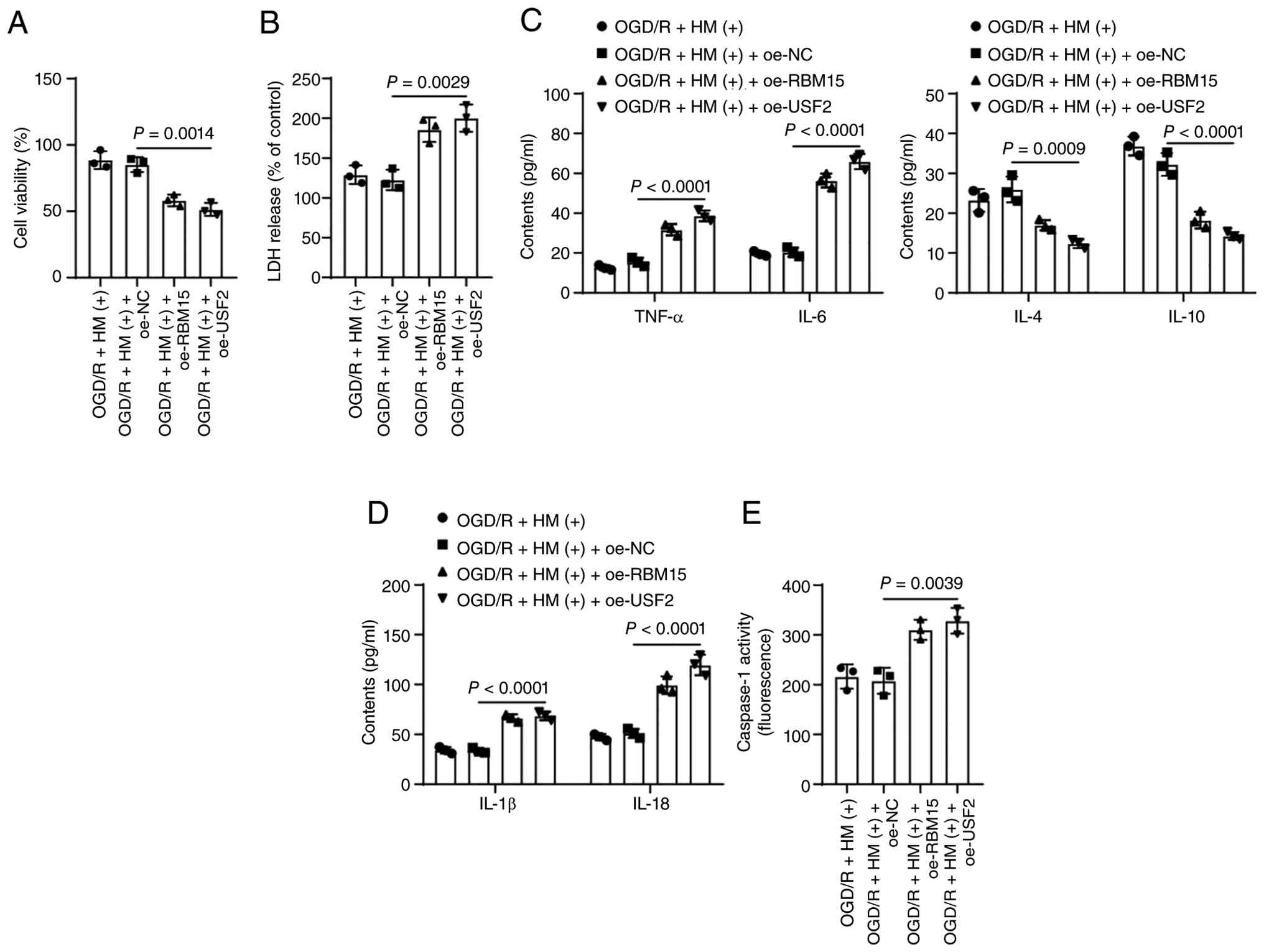 | Figure 8Overexpression of RBM15/USF2
abolishes the protective effect of HM preconditioning on
OGD/R-induced pyroptosis. (A) Cell Counting Kit-8 assay detection
of cell viability in each group. (B) LDH levels in each group of
cells. (C and D) ELISA detection of inflammatory factors and
pyroptosis factors in cells. (E) Detection of caspase 1 enzyme
activity using a caspase 1 fluorescence assay kit. A total of three
technical replicates of the cell experiments were performed. The
data are expressed as the mean ± standard deviation. The data in
panels A, B and E were analyzed via one-way ANOVA, and the data in
panels C and D were analyzed via two-way ANOVA, followed by Tukey's
multiple comparisons test. RBM15, RNA-binding motif protein 15;
USF2, upstream stimulatory factor 2; OGD/R, oxygen-glucose
deprivation/reperfusion; LDH, lactate dehydrogenase; HM,
hydromorphone; oe-, overexpression; NC, negative control; IL,
interleukin; TNF-α, tumor necrosis factor alpha. |
To verify the role of USF2 in the protective effect
of HM against OGD/R-induced cell injury, the combined experiment
was set up on the basis of Fig.
3 as the pretreatment effect of HM has been verified in
Fig. 3 (the negative control of
HM is shown in Fig. 3). USF2
expression was upregulated in cells for a combined treatment with
HM (P<0.05; Fig. 7A-D), which
resulted in an increase in USF2 enrichment in the NLRP3 promoter
(P<0.05; Fig. 7G) and an
increase in NLRP3 expression (P<0.05; Fig. 7A, D and H). Overexpression of
USF2 also counteracted the protective effect of HM preconditioning
on OGD/R-induced cell pyroptosis (P<0.05; Figs. 7D and H; 8A-E).
HM preconditioning alleviates cerebral
I/R injury in mice by upregulating the expression of miR-195-5p to
repress NLRP3-mediated pyroptosis
Finally, to validate in vitro results, the
combined experiment was set up on the basis described in Fig. 2, as the pretreatment effect of HM
has been verified in (the negative control of HM is shown in
Fig. 2). miR-195-5p expression
was reduced in the mouse brain (P<0.01; Fig. 9A and B), which resulted in
increases in m6A content and RBM15, USF2 and NLRP3 expression
levels (P<0.01; Fig. 9B-D).
After inhibition of miR-195-5p expression, the serum LDH and S100B
levels in mice increased, and the infarct volume increased,
indicating a decrease in the neuroprotective effect of HM
preconditioning (P<0.05; Fig.
9E-G). H&E staining revealed that the brain tissues of mice
in which miR-195-5p was inhibited exhibited infarction accompanied
by inflammatory cell infiltration (Fig. 9I). The inhibition of miR-195-5p
exacerbated inflammatory injury in brain tissues (P<0.01;
Fig. 9J) and increased the
expression levels of NLRP3, GSDMD-N, caspase-1 and cleaved
caspase-1 (P<0.01; Fig. 9C and
I), as well as the concentrations of IL-1β and IL-18
(P<0.01; Fig. 9H). Briefly,
inhibition of miR-195-5p promotes NLRP3-mediated pyroptosis by
upregulating the RBM15/USF2 axis and alleviating the protective
effect of HM preconditioning on inflammatory injury in mice with
cerebral I/R.
 | Figure 9HM preconditioning alleviates mouse
cerebral I/R injury by upregulating the expression of miR-195-5p to
repress NLRP3-mediated pyroptosis. The mice were treated according
to the process shown in Fig. 1A.
(A) RT-qPCR detection of the miR-195-5p transfection efficiency in
brain tissues. (B) RT-qPCR detection of the RNA expression of
various genes in brain tissues. (C) Western blot detection of the
protein expression levels of various proteins in brain tissues. (D)
Quantitative analysis of m6A content in brain tissues. (E) LDH and
S100B levels in the serum of mice. (F) 2,3,5-Triphenyltetrazolium
chloride staining for detecting infarct size in brain tissues. (G)
Neurological function scores of the mice in each group. (H) ELISA
detection of pyroptosis factors in brain tissues. (I) Tissue
staining for detecting brain tissue injury and the positive rate of
NLRP3 expression. (J) ELISA detection of inflammatory factors in
brain tissues. Animal experiments in panels E and G: n=9; others:
n=3. The data are expressed as the mean ± standard deviation.
Comparisons between two groups in panel A were conducted using the
t-test; comparisons between multiple groups in panels E (right) and
G were conducted using the Kruskal-Wallis test, and Dunn's multiple
comparisons test was used for post hoc testing. The data in panels
D, E (left) and F were analyzed via one-way ANOVA, and the data in
panels B, C, H and J were analyzed via two-way ANOVA, followed by
Tukey's multiple comparisons test. HM, hydromorphone; I/R,
ischemia/reperfusion; miR, microRNA; NLRP3, Nod-like receptor
protein 3; RT-qPCR, reverse transcription-quantitative PCR; m6A,
N6-methyladenosine; LDH, lactate dehydrogenase; RBM15, RNA-binding
motif protein 15; USF2, upstream stimulatory factor 2; NC, negative
control; IL, interleukin; TNF-α, tumor necrosis factor alpha. |
Discussion
NLRP3 inflammasome-mediated pyroptosis is a vital
pathological link in the inflammatory response to CIRI (1). HM is a potent opioid analgesic
commonly administered in acute care settings (54). The present study revealed that HM
preconditioning downregulates the RBM15/USF2 axis by upregulating
the expression of miR-195-5p, thus repressing NLRP3-mediated
pyroptosis and ultimately alleviating CIRI (Fig. 10).
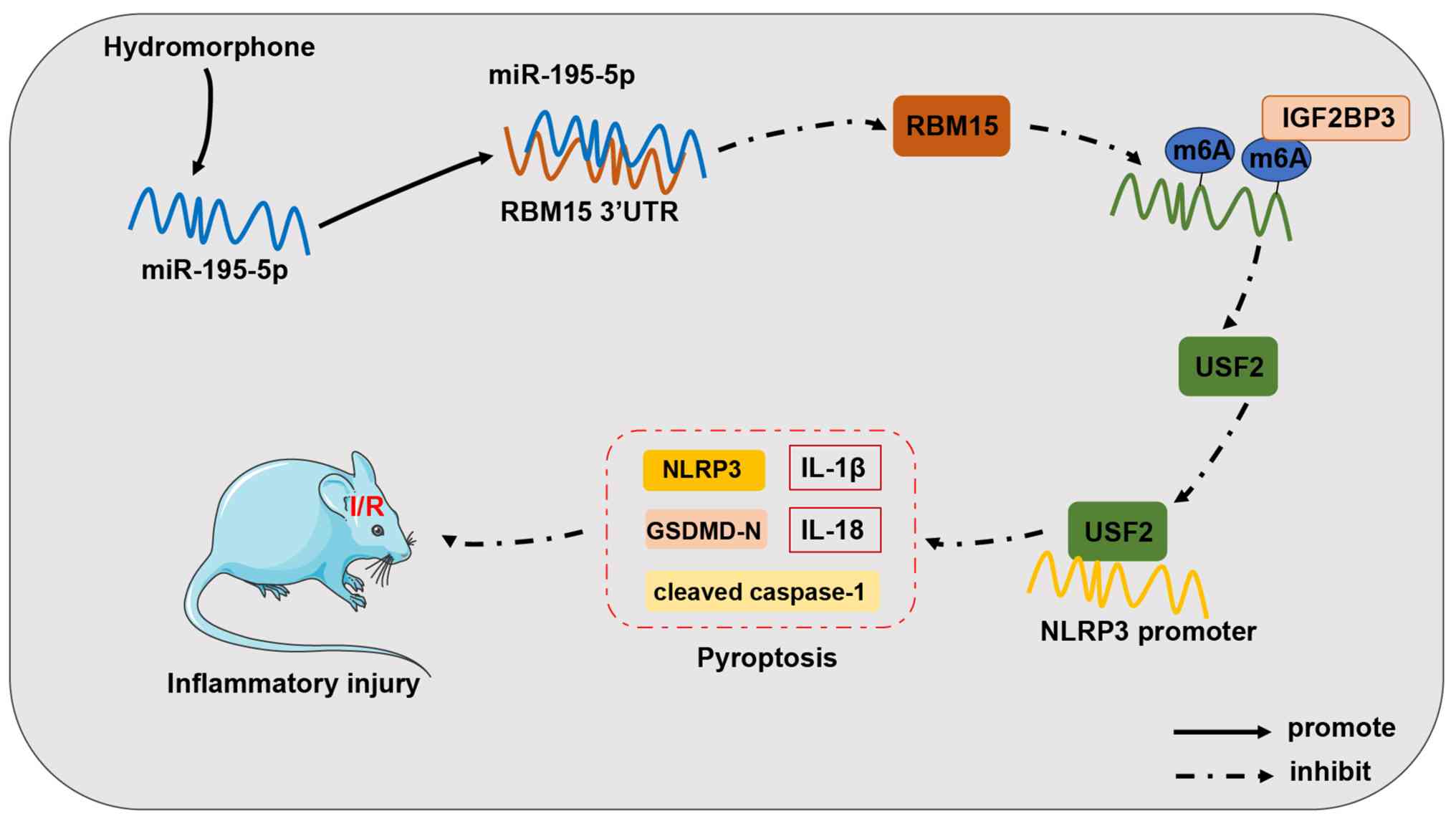 | Figure 10Mechanism through which HM
preconditioning alleviates inflammatory injury caused by cerebral
I/R injury. HM preconditioning can upregulate miR-195-5p
expression, target RBM15 expression, reduce IGF2BP3-mediated m6A
modification, and decrease USF2 expression, thus reducing the
enrichment of USF2 on the NLRP3 promoter, inhibiting NLRP3-mediated
pyroptosis, and alleviating cerebral I/R inflammatory injury. I/R,
ischemia/reperfusion; HM, hydromorphone; miR, microRNA; RBM15,
RNA-binding motif protein 15; m6A, N6-methyladenosine; USF2,
upstream stimulatory factor 2; NLRP3, Nod-like receptor protein 3;
UTR, untranslated region; IGF2BP3, insulin-like growth factor 2
mRNA-binding protein 3. |
HM is an opioid analgesic with a strong analgesic
effect, long duration, and few adverse events (55). In terms of pharmacodynamics, the
analgesic effect of HM is 5-10-fold greater than that of the
equivalent dose of morphine (56). A double-blind, randomized and
controlled trial revealed that preoperative 2 mg of HM in patients
undergoing laparoscopic radical gastrectomy can reduce
intraoperative changes in blood pressure and heart rate and relieve
postoperative pain intensity (57). In the present study, it was
revealed that HM preconditioning notably reduced the infarct size
of MCAO/R mice, alleviated inflammatory cell infiltration in brain
tissues, and diminished inflammatory cytokines in brain tissues.
Additionally, HM has been demonstrated to protect hippocampal
neurons from I/R injury by activating mTOR signaling (12). Pyroptosis is a type of
proinflammatory cell death that occurs in an NLRP3-dependent manner
after CIRI (4,58). HM preconditioning significantly
reduced the expression levels of NLRP3, GSDMD-N and cleaved
caspase-1 in the brain tissues of MCAO/R mice, as well as the
concentrations of the pyroptosis factors IL-1β and IL-18,
indicating that HM preconditioning alleviated CIRI and restrained
NLRP3-mediated pyroptosis in MCAO/R mice. Consistently, it has been
reported that HM represses NLRP3 inflammasome-mediated pyroptosis
in alveolar macrophages by upregulating Nrf2/HO-1 expression
(13). In vitro, it was
found that HM preconditioning reduced OGD/R-induced inflammatory
factor expression and suppressed pyroptosis in HBMVECs. After
combined treatment with the NLRP3 agonist Nig and HM, cell
pyroptosis significantly increased, but cell viability decreased
and the inflammatory response increased, indicating that HM
preconditioning alleviated OGD/R-induced inflammatory injury by
suppressing NLRP3-mediated pyroptosis.
Thereafter, it was attempted to determine the
underlying mechanism through which HM preconditioning alleviates
CIRI. miRNAs positively modulate inflammation and cell
survival/death in response to CIRI (59). miRNA features can be utilized as
clinical biomarkers for predicting the analgesic effect of HM
(53). The overexpression of
miR-195-5p efficiently increased cell viability while reducing LDH
release and apoptosis in OGD-induced HBMVECs in vitro and
ameliorating cerebral injury in MCAO mice in vivo (16). Silencing of miR-195-5p
exacerbates inflammation and apoptosis in OGD/R-induced HT22 cells
(18). miR-195-5p attenuates
pyroptosis in OGD/R-induced GC-1 cells by repressing PELP1
expression (19). Similarly, it
was found that miR-195-5p expression decreased in the model group
but increased in the HM preconditioning group. The inhibition of
miR-195-5p expression decreased cell viability, elevated cellular
inflammation, and significantly enhanced pyroptosis. These results
indicated that HM preconditioning alleviated OGD/R-induced
inflammatory injury by upregulating the expression of miR-195-5p to
inhibit NLRP3-mediated pyroptosis.
Furthermore, the target genes downstream of
miR-195-5p were predicted. The binding between miR-195-5p and RBM15
was confirmed by a dual-luciferase assay. RBM15 expression is
upregulated after cerebral ischemia (21). The silencing of RBM15 diminishes
inflammatory factors and decreases pyroptosis in high
glucose-induced HK-2 cells (24). The present results demonstrated
that RBM15 expression increased in the model group but decreased in
the HM preconditioning group, but RBM15 expression increased again
after inhibition of miR-195-5p. USF2 belongs to the bHLH-LZ
transcription factor family and interacts with the E-box of the
DNA-core sequence in its target genes (60). USF2 is crucial for controlling
growth and developmental processes, as it affects embryonic
development, fertility and brain function (61). m6A modification was detected on
USF2, and the changes in m6A content in USF2 were consistent with
the expression trends of RBM15. Overexpression of RBM15
significantly promoted the mRNA stability and expression of USF2.
USF2 is highly expressed in MCAO mice, and knockdown of USF2
markedly reduces autophagy and alleviates CIRI (29). USF2 increases NLRP3 expression at
the transcriptional level, and inhibition of USF2 represses the
pyroptosis of podocytes and relieves kidney injury in patients with
lupus nephritis (62). It was
also revealed that HM preconditioning limited the upregulation of
USF2 expression in MCAO mice and OGD/R-induced HBMVECs. Functional
rescue experiments confirmed that overexpression of RBM15/USF2
abolished the protective effect of HM preconditioning on
OGD/R-induced pyroptosis. Finally, to validate the in vitro
results, miR-195-5p expression was reduced in mice and it was found
that inhibition of miR-195-5p exacerbated inflammatory injury to
the brain tissue and increased pyroptosis, indicating that
inhibition of miR-195-5p facilitated NLRP3-mediated pyroptosis by
upregulating the RBM15/USF2 axis and reversing the protective
effect of HM preconditioning on CIRI in mice.
In conclusion, HM preconditioning upregulates
miR-195-5p expression to suppress RBM15 expression, reduces
IGF2BP3-mediated m6A modification, and decreases USF2 expression,
thus reducing USF2 enrichment on the NLRP3 promoter, repressing
NLRP3-mediated pyroptosis, and alleviating CIRI. The present study
provides new strategies and targets for the treatment of CIRI.
Multicenter preclinical studies are needed to further clarify the
neuroprotective effects of HM, which is expected to fill the gap in
existing neuroprotective drugs.
However, the present study has certain limitations.
First, long-term functional assessments are lacking. Owing to the
limited funding and experimental technology in the authors'
research group, the functional outcomes of the mice by adhesive
removal, rotarod and corner tests were not assessed. In addition,
additional experiments on the combination of MCC950 or
Nlrp3−/− mice were not conducted. MCC950 or
Nlrp3−/− mice will be used for deeper validation in
future studies. Second, only endothelial cells were used in
vitro. Considering that human-derived cells are more closely
related to the clinical pathological environment, HBMVECs were
chosen as the objective cells but did not replicate key nodes of
the mechanism in primary murine brain microvascular endothelial
cells. Hence, cross-species limitations such as differences in
inflammatory response and culture conditions may affect the
results. In the combined experiments, the data of OGD/R+
(HM−) and IR+ saline groups in Figs. 4, 6 and 7 were not shown, which were the
negative controls of HM in the in vitro and in vivo
experiments. Third, the absence of antagonist experiments (for
example, naloxone) render unclear whether the protective effects of
HM are mediated by opioid receptors. Fourth, although a combined
experiment involving HM preconditioning and genetic intervention
was conducted, the causal relationship using caspase-1 inhibitors
to enhance the rigor of the manuscript has not yet been validated.
Fifth, a seed-mutant RBM15 3'UTR (insensitive to miR-195-5p) in
AGO2-RIP assay was not used. This experiment may be included in
future studies to further confirm the target binding of miR-195-5p
to RBM15. Moreover, ChIP and RIP assays merely included IgG as a
negative control, but did not include positive control loci. Last
but not least, only miR-195-5p was selected as the downstream
target of HM, and whether other potential off-target miRNAs are
involved remains unclear. For the downstream target of miR-195-5p,
only RBM15 was selected as the target gene from the database. It is
unknown whether RBM15 affects USF2 through other m6A readers, and
additional genes downstream of RBM15 remain to be clarified.
As the present study remains in the exploratory
stage, more experimental evidence is needed to apply the
conclusions to clinical testing. In the future, other downstream
mechanisms of HM preconditioning will be explored through
differential analysis to further elucidate the impact of HM
preconditioning on CIRI and provide new theoretical knowledge for
the treatment of CIRI.
Supplementary Data
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
JY conceptualized the study, visualized, curated and
validated data, conducted formal analysis, developed methodology,
wrote the original draft, and wrote, reviewed and edited the
manuscript. ZH conceptualized the study, curated and visualized
data, and conducted investigation. SL supervised the study, curated
and validated data, and developed methodology. ML conceptualized
the study, conducted formal analysis and investigation, and
developed methodology. JF supervised the study, conducted formal
analysis and investigation, validated data, and wrote, reviewed and
edited the manuscript. JY and YF confirm the authenticity of all
the raw data. All authors read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
The study procedure was approved by the Ethics
Committee of Liaoning Cancer Hospital and Institute (approval no.
CMU20250031; Shenyang, China). All animal experiment schemes were
approved by the Animal Ethics Committee of Liaoning Cancer Hospital
and Institute and implemented based on the Guide for the Care and
Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
HM
|
hydromorphone
|
|
I/R
|
ischemia/reperfusion
|
|
CIRI
|
cerebral ischemia/reperfusion
injury
|
|
RBM15
|
RNA-binding motif protein 15
|
|
USF2
|
upstream stimulatory factor 2
|
|
IGF2BP3
|
insulin-like growth factor 2
mRNA-binding protein 3
|
|
miRNA or miR
|
microRNA
|
|
m6A
|
N6-methyladenosine
|
|
MCAO/R
|
middle cerebral artery
occlusion/reperfusion
|
|
OGD/R
|
oxygen-glucose
deprivation/reperfusion
|
|
CBF
|
cerebral blood flow
|
|
ORs
|
opioid receptors
|
|
Nig
|
nigericin
|
|
HBMVECs
|
human brain microvascular endothelial
cells
|
|
H&E
|
hematoxylin and eosin
|
|
TTC
|
2,3,5-triphenyltetrazolium
chloride
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
IL
|
interleukin
|
|
TNF-α
|
tumor necrosis factor alpha
|
|
DMEM
|
Dulbecco's modified Eagle's
medium
|
|
NLRP3
|
nod-like receptor protein 3
|
|
FBS
|
fetal bovine serum
|
|
PBS
|
phosphate-buffered saline
|
|
CCK-8
|
Cell Counting Kit-8
|
|
LDH
|
lactate dehydrogenase
|
|
CPK
|
creatine phosphokinase
|
|
RIP
|
RNA immunoprecipitation
|
|
ChIP
|
chromatin immunoprecipitation
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
ANOVA
|
analysis of variance
|
Acknowledgements
Not applicable.
Funding
No funding was received.
References
|
1
|
Mao R, Zong N, Hu Y, Chen Y and Xu Y:
Neuronal death mechanisms and therapeutic strategy in ischemic
stroke. Neurosci Bull. 38:1229–1247. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sommer CJ: Ischemic stroke: Experimental
models and reality. Acta Neuropathol. 133:245–261. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang L, Ren W, Wu Q, Liu T, Wei Y, Ding J,
Zhou C, Xu H and Yang S: NLRP3 inflammasome activation: A
therapeutic target for cerebral ischemia-reperfusion injury. Front
Mol Neurosci. 15:8474402022. View Article : Google Scholar :
|
|
4
|
Gou X, Xu D, Li F, Hou K, Fang W and Li Y:
Pyroptosis in stroke-new insights into disease mechanisms and
therapeutic strategies. J Physiol Biochem. 77:511–529. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Duan WL, Wang XJ, Ma YP, Sheng ZM, Dong H,
Zhang LY, Zhang BG and He MT: Therapeutic strategies targeting the
NLRP3-mediated inflammatory response and pyroptosis in cerebral
ischemia/reperfusion injury (Review). Mol Med Rep. 29:462024.
View Article : Google Scholar
|
|
6
|
Kao TK, Ou YC, Liao SL, Chen WY, Wang CC,
Chen SY, Chiang AN and Chen CJ: Opioids modulate post-ischemic
progression in a rat model of stroke. Neurochem Int. 52:1256–1265.
2008. View Article : Google Scholar
|
|
7
|
Crowley MG, Liska MG, Lippert T, Corey S
and Borlongan CV: Utilizing delta opioid receptors and peptides for
cytoprotection: implications in stroke and other neurological
disorders. CNS Neurol Disord Drug Targets. 16:414–424. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grant Liska M, Crowley MG, Lippert T,
Corey S and Borlongan CV: Delta opioid receptor and peptide: A
dynamic therapy for stroke and other neurological disorders. Handb
Exp Pharmacol. 247:277–299. 2018. View Article : Google Scholar
|
|
9
|
Vaidya B, Sifat AE, Karamyan VT and
Abbruscato TJ: The neuroprotective role of the brain opioid system
in stroke injury. Drug Discov Today. 23:1385–1395. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Murray A and Hagen NA: Hydromorphone. J
Pain Symptom Manage. 29(5 Suppl): S57–S66. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim YS, Kim WY, Kim YH, Yoo JW and Min TJ:
The protective effect of hydromorphone to ischemia in rat glial
cells. Springerplus. 5:6102016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xie W, Xie W, Kang Z, Jiang C and Liu N:
Hydromorphone protects CA1 neurons by activating mTOR pathway.
Neurosci Lett. 687:49–54. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Li J, An Z and Qi J:
Hydromorphone mitigates cardiopulmonary bypass-induced acute lung
injury by repressing pyroptosis of alveolar macrophages. Shock.
60:92–99. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Li X, Qiao S, Yang D, Li Z, Xu J,
Li W, Su L and Liu W: Occludin degradation makes brain
microvascular endothelial cells more vulnerable to reperfusion
injury in vitro. J Neurochem. 156:352–366. 2021. View Article : Google Scholar
|
|
15
|
Long J, Sun Y, Liu S, Yang S, Chen C,
Zhang Z, Chu S, Yang Y, Pei G, Lin M, et al: Targeting pyroptosis
as a preventive and therapeutic approach for stroke. Cell Death
Discov. 9:1552023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren X, Wang Z and Guo C: MiR-195-5p
ameliorates cerebral ischemia-reperfusion injury by regulating the
PTEN-AKT signaling pathway. Neuropsychiatr Dis Treat. 17:1231–1242.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Forouzanfar F, Shojapour M, Asgharzade S
and Amini E: Causes and consequences of MicroRNA dysregulation
following cerebral ischemia-reperfusion injury. CNS Neurol Disord
Drug Targets. 18:212–221. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu F, Luo E, Yi F, Xiong J, Huang C and
Li R: LncRNA ITSN1-2 knockdown inhibits OGD/R-induced inflammation
and apoptosis in mouse hippocampal neurons via sponging miR-195-5p.
Neuroreport. 32:1325–1334. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He KX, Xu L, Ning JZ and Cheng F:
MiR-195-5p is involved in testicular ischemia/reperfusion injury by
directly targeting PELP1 and regulating spermatogonia pyroptosis.
Int Immunopharmacol. 121:1104272023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao L and Zhang X: Propofol enhances the
lethality of cisplatin on liver cancer cells by up-regulating
miR-195-5p. Tissue Cell. 74:1016802022. View Article : Google Scholar
|
|
21
|
Liang E, Xiao S, Zhao C, Zhang Y and Fu G:
M6A modification promotes blood-brain barrier breakdown during
cerebral ischemia/reperfusion injury through increasing matrix
metalloproteinase 3 expression. Heliyon. 9:e169052023. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang X, Liu B, Nie Z, Duan L, Xiong Q,
Jin Z, Yang C and Chen Y: The role of m6A modification in the
biological functions and diseases. Signal Transduct Target Ther.
6:742021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia K, Xia W, Su Q, Yang S, Zhang Y, Ni X,
Su Z and Meng D: RNA methylation pattern and immune
microenvironment characteristics mediated by m6A regulator in
ischemic stroke. Front Genet. 14:11485102023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qin Y, Wu S, Zhang F, Zhou X, You C and
Tan F: N6-methyladenosine methylation regulator RBM15 promotes the
progression of diabetic nephropathy by regulating cell
proliferation, inflammation, oxidative stress, and pyroptosis
through activating the AGE-RAGE pathway. Environ Toxicol.
38:2772–2782. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jones-Bolin S: Guidelines for the care and
use of laboratory animals in biomedical research. Curr Protoc
Pharmacol Appendix. 4:Appendix 4B. 2012.
|
|
26
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42(Database Issue): D92–D97. 2014. View Article : Google Scholar :
|
|
27
|
Zhou Y, Zeng P, Li YH, Zhang Z and Cui Q:
SRAMP: Prediction of mammalian N6-methyladenosine (m6A) sites based
on sequence-derived features. Nucleic Acids Res. 44:e912016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rauluseviciute I, Riudavets-Puig R,
Blanc-Mathieu R, Castro-Mondragon JA, Ferenc K, Kumar V, Lemma RB,
Lucas J, Chèneby J, Baranasic D, et al: JASPAR 2024: 20th
anniversary of the open-access database of transcription factor
binding profiles. Nucleic Acids Res. 52(D1): D174–D182. 2024.
View Article : Google Scholar :
|
|
29
|
Liu C, Gao Q, Dong J and Cai H: Usf2
deficiency promotes autophagy to alleviate cerebral
ischemia-reperfusion injury through suppressing YTHDF1-m6A-mediated
Cdc25A translation. Mol Neurobiol. 61:2556–2568. 2024. View Article : Google Scholar
|
|
30
|
Zhong Y, Gu L, Ye Y, Zhu H, Pu B, Wang J,
Li Y, Qiu S, Xiong X and Jian Z: JAK2/STAT3 axis intermediates
microglia/macrophage polarization during cerebral
ischemia/reperfusion injury. Neuroscience. 496:119–128. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo P, Jin Z, Wu H, Li X, Ke J, Zhang Z
and Zhao Q: Effects of irisin on the dysfunction of blood-brain
barrier in rats after focal cerebral ischemia/reperfusion. Brain
Behav. 9:e014252019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang Y, Hu F, Yang G and Meng Q: Lack of
sphingomyelin synthase 2 reduces cerebral ischemia/reperfusion
injury by inhibiting microglial inflammation in mice. Exp Ther Med.
20:2412020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cai L, Yao ZY, Yang L, Xu XH, Luo M, Dong
MM and Zhou GP: Mechanism of electroacupuncture against cerebral
ischemia-reperfusion injury: reducing inflammatory response and
cell pyroptosis by inhibiting NLRP3 and caspase-1. Front Mol
Neurosci. 15:8220882022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Diaz-Ruiz A, Vacio-Adame P, Monroy-Noyola
A, Mendez-Armenta M, Ortiz-Plata A, Montes S and Rios C:
Metallothionein-II inhibits lipid peroxidation and improves
functional recovery after transient brain ischemia and reperfusion
in rats. Oxid Med Cell Longev. 2014:4364292014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou J, Wu JS, Yan Y, Li J, Ni T, Shao W,
Mei JH, Xiong WZ and Wu H: MiR-199a modulates autophagy and
inflammation in rats with cerebral infarction via regulating mTOR
expression. Eur Rev Med Pharmacol Sci. 24:6338–6345.
2020.PubMed/NCBI
|
|
36
|
Rong W, Yang L, Li CY, Wu XT, Zhou ZD, Zhu
WL and Yan Y: MiR-29 inhibits neuronal apoptosis in rats with
cerebral infarction through regulating Akt signaling pathway. Eur
Rev Med Pharmacol Sci. 24:843–850. 2020.PubMed/NCBI
|
|
37
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang P, Cui Y, Ren Q, Yan B, Zhao Y, Yu P,
Gao G, Shi H, Chang S and Chang YZ: Mitochondrial ferritin
attenuates cerebral ischaemia/reperfusion injury by inhibiting
ferroptosis. Cell Death Dis. 12:4472021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hu Y, Ye C, Cheng S and Chen J: Propofol
downregulates lncRNA MALAT1 to alleviate cerebral
ischemia-reperfusion injury. Inflammation. 44:2580–2591. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Guo Y, Yang JH, He Y, Zhou HF, Wang Y,
Ding ZS, Jin B and Wan HT: Protocatechuic aldehyde prevents
ischemic injury by attenuating brain microvascular endothelial cell
pyroptosis via lncRNA Xist. Phytomedicine. 94:1538492022.
View Article : Google Scholar
|
|
41
|
Kong LY, Zhu SY, Si MY, Xu XH, Yu JJ,
Zhong WX, Sang CP, Rao DY, Xie FC, Liu ZY and Tang ZX: TAK-242
protects against oxygen-glucose deprivation and
reoxygenation-induced injury in brain microvascular endothelial
cells and alters the expression pattern of lncRNAs. J Thorac Dis.
15:2571–2584. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Han C, Zhai L, Shen H, Wang J and Guan Q:
Advanced glycation end-products (AGEs) promote endothelial cell
pyroptosis under cerebral ischemia and hypoxia via
HIF-1α-RAGE-NLRP3. Mol Neurobiol. 60:2355–2366. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kong LY, Liang MY, Liu JP, Lai P, Ye JS,
Zhang ZX, Du ZM, Yu JJ, Gu L, Xie FC, et al: Mesenchymal stem
cell-derived exosomes rescue oxygen-glucose deprivation-induced
injury in endothelial cells. Curr Neurovasc Res. 17:155–163. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao B, Fei Y, Zhu J, Yin Q, Fang W and Li
Y: PAF receptor inhibition attenuates neuronal pyroptosis in
cerebral ischemia/reperfusion injury. Mol Neurobiol. 58:6520–6539.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Waithe OY, Anderson A, Muthusamy S,
Seplovich GM and Tharakan B: Homocysteine induces brain and retinal
microvascular endothelial cell barrier damage and hyperpermeability
via NLRP3 inflammasome pathway differentially. Microcirculation.
32:e700192025. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang J, Xu Z, Chen X, Li Y, Chen C, Wang
C, Zhu J, Wang Z, Chen W, Xiao Z and Xu R: MicroRNA-182-5p
attenuates cerebral ischemia-reperfusion injury by targeting
Toll-like receptor 4. Biochem Biophys Res Commun. 505:677–684.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu C, Xu X, Huang C, Zhang L, Shang D,
Cai W and Wang Y: Circ_002664/miR-182-5p/Herpud1 pathway
importantly contributes to OGD/R-induced neuronal cell apoptosis.
Mol Cell Probes. 53:1015852020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu C, Sui H, Li Z, Sun Z, Li C, Chen G,
Ma Z, Cao H and Xi H: THBS1 in macrophage-derived exosomes
exacerbates cerebral ischemia-reperfusion injury by inducing
ferroptosis in endothelial cells. J Neuroinflammation. 22:482025.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dimopoulos C, Damaskos C, Papadakis M,
Garmpis N, Kontzoglou K, Perrea D, Moraitis S, Daskalopoulou A,
Papaspirou I, Georgopoulos S and Nikiteas N: Expression of S100B
protein in ischemia/reperfusion-induced brain injury after
cyclosporine therapy: A biochemical serum marker with prognostic
value? Med Sci Monit. 25:1637–1644. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
51
|
Jiang Z, Shi L, Huang H, Lei D, Lou L, Jin
Y, Sun J and Wang L: Downregulated FTO promotes
MicroRNA-155-mediated inflammatory response in cerebral
ischemia/reperfusion injury. Neuroscience. 526:305–313. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang X, Tian L, Li Y, Wang J, Yan B, Yang
L, Li Q, Zhao R, Liu M, Wang P and Sun Y: RBM15 facilitates
laryngeal squamous cell carcinoma progression by regulating TMBIM6
stability through IGF2BP3 dependent. J Exp Clin Cancer Res.
40:802021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kiyosawa N, Watanabe K, Toyama K and
Ishizuka H: Circulating miRNA signature as a potential biomarker
for the prediction of analgesic efficacy of hydromorphone. Int J
Mol Sci. 20:16652019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mazer-Amirshahi M, Motov S and Nelson LS:
Hydromorphone use for acute pain: Misconceptions, controversies,
and risks. J Opioid Manag. 14:61–71. 2018. View Article : Google Scholar
|
|
55
|
Pickering G, Kotlinska-Lemieszek A,
Krcevski Skvarc N, O'Mahony D, Monacelli F, Knaggs R, Morel V and
Kocot-Kepska M: Pharmacological pain treatment in older persons.
Drugs Aging. 41:959–976. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu L, Xu M, Wang J, Hu Y and Huang Z:
Research progress of hydromorphone in clinical application. Physiol
Res. 74:41–48. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang J, Wang N, Zhou H and Wang Y: Effect
of preoperative hydromorphone in patients undergoing laparoscopic
radical gastrectomy. A double-blind, randomized and controlled
trial. Saudi Med J. 39:1023–1027. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gao L, Dong Q, Song Z, Shen F, Shi J and
Li Y: NLRP3 inflammasome: A promising target in ischemic stroke.
Inflamm Res. 66:17–24. 2017. View Article : Google Scholar
|
|
59
|
Ghafouri-Fard S, Shoorei H and Taheri M:
Non-coding RNAs participate in the ischemia-reperfusion injury.
Biomed Pharmacother. 129:1104192020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Corre S and Galibert MD: Upstream
stimulating factors: Highly versatile stress-responsive
transcription factors. Pigment Cell Res. 18:337–348. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fujimi TJ and Aruga J: Upstream
stimulatory factors, USF1 and USF2 are differentially expressed
during Xenopus embryonic development. Gene Expr Patterns.
8:376–381. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xie Y, Li X, Deng W, Nan N, Zou H, Gong L,
Chen M, Yu J, Chen P, Cui D and Zhang F: Knockdown of USF2 inhibits
pyroptosis of podocytes and attenuates kidney injury in lupus
nephritis. J Mol Histoln. 54:313–327. 2023. View Article : Google Scholar
|














