Introduction
Osteoarthritis (OA) has historically been
categorized within orthopedics and sports medicine as a localized
condition resulting from 'wear and tear' or mechanical overload.
However, this hemodynamic and biomechanical perspective fails to
explain the prevalence of joint degeneration in non-weight-bearing
joints or the accelerated progression observed in patients with
metabolic comorbidities. Contemporary epidemiological data indicate
that OA is a leading cause of global disability, with its incidence
rising in parallel with the obesity epidemic (1,2).
Consequently, the paradigm has shifted toward defining OA as a
whole-organ disease characterized by low-grade systemic
inflammation, often termed 'metabolic OA', a clinical phenotype
defined by the co-occurrence of OA with components of metabolic
syndrome (obesity, dyslipidemia, hypertension and/or insulin
resistance) in the absence of overt mechanical trauma. This
phenotype is distinct from posttraumatic etiologies and is
intricately linked with metabolic syndrome components such as
dyslipidemia, hypertension and insulin resistance (3). Recent evidence suggests that the
chronic low-grade inflammation associated with aging, known as
'inflammaging', which is as the age-related, low-grade, sterile
systemic inflammation that drives cellular senescence and tissue
dysfunction, acts as a critical driver of chondrocyte senescence
and synovial dysfunction (4,5).
Therefore, understanding the systemic origins of this inflammation
is paramount for developing non-surgical interventions that go
beyond symptomatic pain relief.
The gastrointestinal tract has emerged as a primary
candidate for the origin of this systemic inflammation. The
'gut-joint axis' hypothesis posits that the intestinal ecosystem
communicates with articular tissues through complex immunological
and metabolic pathways. Under physiological conditions, the gut
barrier prevents the translocation of luminal pathogens; however,
dysbiosis (an imbalance in microbial composition) can compromise
this barrier, leading to a 'leaky gut' phenomenon (6,7).
Systematic reviews have consistently identified distinct microbial
signatures in patients with OA compared to healthy controls, often
characterized by a reduction in diversity and an alteration in the
Firmicutes to Bacteroidetes ratio (8,9).
Furthermore, recent Mendelian randomization studies have provided
causal evidence suggesting that gut microbiota dysbiosis is not
merely a consequence of OA or analgesic use but a potential
initiating factor in the disease pathogenesis (10,11). This bidirectional relationship
suggests that the joint is susceptible to inflammatory signals
originating from the gut, challenging the traditional dogma that
the articular cavity is a sterile and isolated environment.
While the presence of bacterial DNA in synovial
fluid has been documented, the primary mediators of the gut-joint
crosstalk are small-molecule metabolites produced by the
fermentation of dietary substrates. These metabolites enter the
systemic circulation and act as signaling molecules that regulate
bone extracellular matrix homeostasis and immune responses in
distal tissues (12,13). This 'biological highway' explains
how dietary inputs directly influence joint health. Recent
multi-omics analyses have revealed that specific alterations in the
serum metabolome, driven by gut microbial shifts, correlate
strongly with the severity of synovitis and cartilage degradation
(14,15). Unlike short-lived cytokines,
these metabolites can induce epigenetic changes in chondrocytes and
osteoblasts, thereby modulating the gene expression profiles
responsible for tissue repair or degradation. Thus, the gut
microbiota functions as a 'forgotten organ' with endocrine
capabilities, secreting bioactive molecules that dictate the
metabolic fate of the musculoskeletal system (14,15).
Among the myriad of microbial byproducts, three
classes of metabolites have garnered significant attention due to
their opposing roles in joint homeostasis: Short-chain fatty acids
(SCFAs), trimethylamine N-oxide (TMAO) and bile acids (BAs). SCFAs,
such as butyrate and propionate, are generally regarded as
'protective guardians' that suppress inflammation and promote
regulatory T-cell differentiation (16). Conversely, TMAO, a product of
dietary choline metabolism, has been identified as a
'pro-inflammatory aggressor' linked to oxidative stress and the
newly described gut-microbiota-ferroptosis axis in OA (17). BAs occupy a complex middle
ground; while they are essential for nutrient absorption,
dysregulated BA metabolism has been associated with symptomatic
hand and knee OA through the modulation of the farnesoid X receptor
(FXR) and inflammatory pathways (18,19). The delicate balance between these
metabolite classes may determine whether the joint microenvironment
remains in a state of homeostasis or progresses toward
catabolism.
As sports medicine evolves toward precision health
and lifestyle medicine, unraveling these metabolic networks offers
novel therapeutic avenues. Current management strategies largely
rely on analgesics and eventual arthroplasty, but targeting the
gut-joint axis presents an opportunity for disease modification
through diet, exercise and microbiome modulation (20,21). Exercise, a cornerstone of OA
rehabilitation, has been shown to independently improve gut
microbial diversity and serum metabolomics, potentially mitigating
the effects of a high-fat diet (20,22). Furthermore, the integration of
multi-omics data allows for the identification of personalized
metabolic phenotypes, paving the way for targeted interventions
such as prebiotics, probiotics or postbiotics (21,23). Thus, a significant unmet
cognitive need remains: No existing framework systematically
integrates the opposing yet complementary roles of SCFAs, TMAO and
BAs to guide the transition from generalized microbiome modulation
toward stratified, mechanism-driven interventions.
This review aims to synthesize the current state of
knowledge regarding the crosstalk between gut microbiota-derived
metabolites and OA, specifically focusing on the mechanistic roles
of SCFAs, TMAO and BAs, to provide a theoretical basis for novel
clinical strategies in the preservation of joint health.
Furthermore, this review deconstructs the conventional 'biological
highway' metaphor into a more precise three-tier mechanistic
schema. This hierarchical schema provides a structured framework
for identifying therapeutic targets at each level of the gut-joint
axis and directly addresses the identified cognitive gap by
translating the conceptual model of metabolic equilibrium into
testable mechanistic hypotheses. A dedicated conceptual schematic
(Fig. 1) is placed below this
paragraph to systematically visualize the proposed 'metabolite
homeostasis imbalance' theoretical framework and three-tier
gut-joint axis mechanistic hierarchy. This diagram hierarchically
divides gut-joint communication into intestinal barrier integrity,
systemic trafficking and local joint effector tiers, and
graphically demonstrates how disrupted balance among protective
SCFAs, detrimental TMAO and bidirectionally regulated BAs initiates
metabolic OA lesions (10,18).
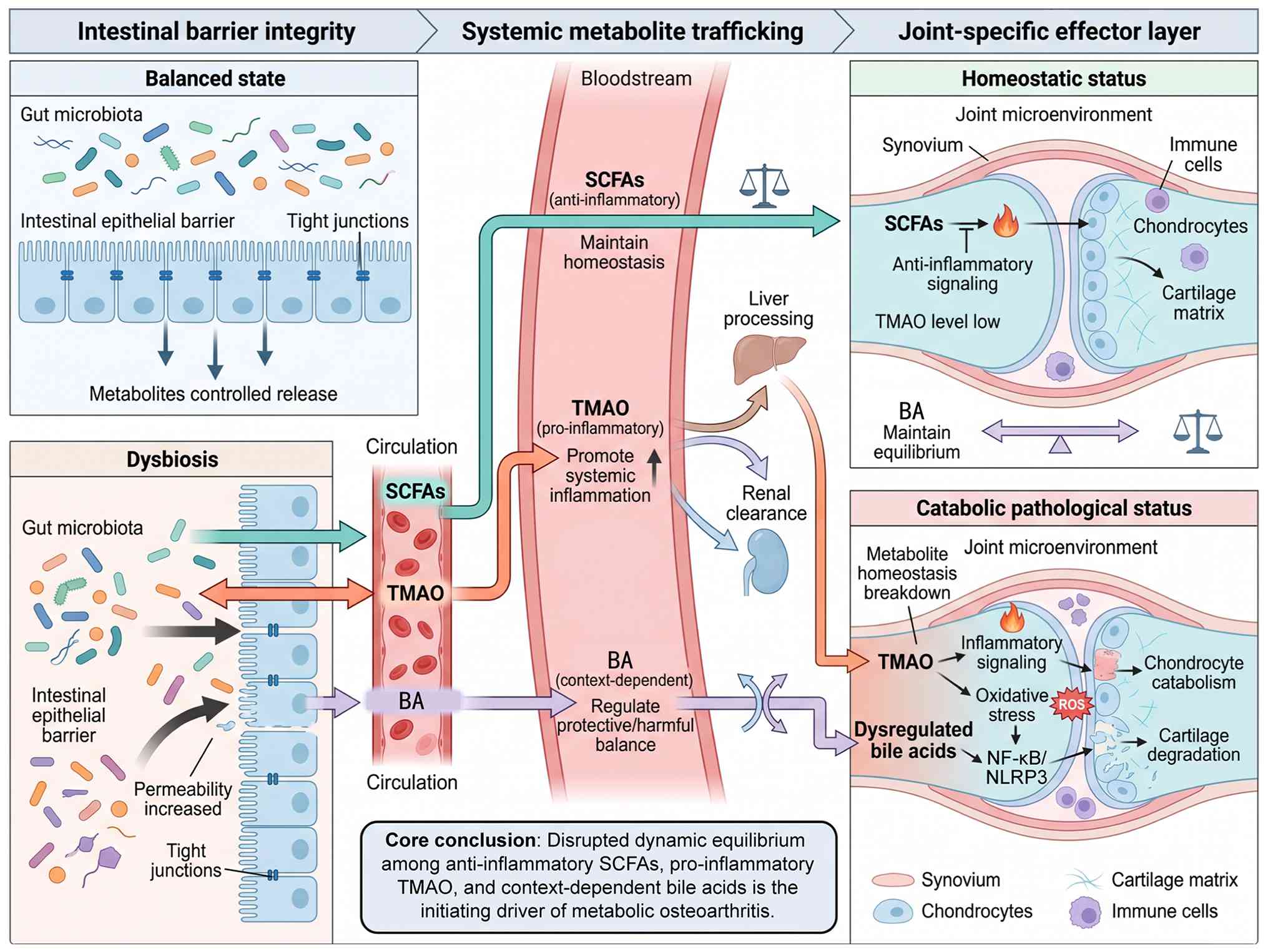 | Figure 1Schematic illustration of the
proposed 'metabolite homeostasis imbalance' theoretical framework
and three-tier mechanistic gut-joint axis schema. The figure
stratifies the whole pathological cascade into intestinal barrier
integrity, systemic metabolite trafficking and joint-specific
effector three tiers, and demonstrates that disrupted dynamic
equilibrium between anti-inflammatory SCFAs, pro-inflammatory TMAO
and context-dependent bile acids is the core initiating driver of
metabolic osteoarthritis. SCFAs, short-chain fatty acids; TMAO,
trimethylamine N-oxide; BA, bile acid; OA, osteoarthritis; FXR,
farnesoid X receptor; TGR5, Takeda G-protein-coupled receptor 5;
NLRP3, NLR family pyrin domain containing 3; NF-κB, nuclear factor
κB. |
The biological highway: The gut-joint
axis
The concept of the 'gut-joint axis' represents a
paradigm shift in understanding OA pathogenesis, transforming it
from a localized disorder of wear and tear into a complex systemic
condition driven by metabolic and inflammatory mediators. This
bidirectional communication channel facilitates the transit of
gut-derived molecules to distal musculoskeletal tissues via the
systemic circulation. Consistent with the three-tier mechanistic
schema proposed in the introduction, the gut-joint axis operates
through a sequential cascade of molecular events that can be
precisely delineated at the barrier, trafficking and effector
levels. Recent evidence suggests that the integrity of the
intestinal barrier and the subsequent translocation of microbial
products or metabolites constitute the 'biological highway' through
which intestinal dysbiosis exacerbates articular degeneration.
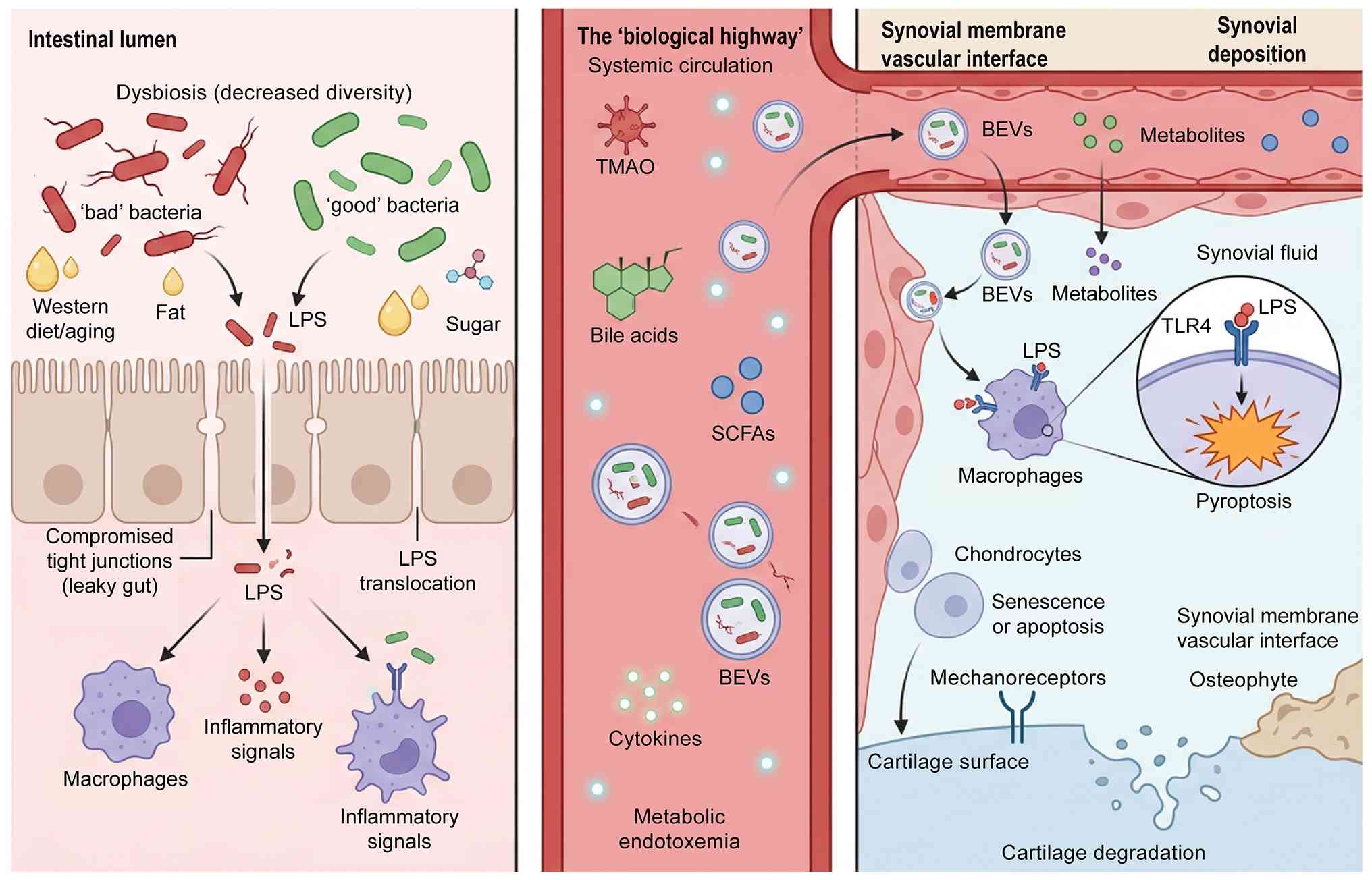
Fig. 2 centrally illustrates the
complete mechanistic cascade of the gut-joint axis in OA
pathogenesis. As shown in the schematic, gut dysbiosis disrupts
intestinal barrier integrity and increases gut permeability,
allowing microbial metabolites and bacterial extracellular vesicles
(BEVs) to enter systemic circulation (24). These gut-derived bioactive
substances translocate to joint tissues, activate TLR4-mediated
inflammatory signaling and chondrocyte pyroptosis and ultimately
drive cartilage degeneration and OA progression (25). This graphical model intuitively
validates the three-tier gut-joint axis mechanism proposed in this
review.
The 'leaky gut' hypothesis
The primary initiation point of this pathological
axis lies in the disruption of the intestinal epithelial barrier, a
phenomenon clinically termed 'leaky gut'. Under physiological
conditions, the intestinal epithelium serves as a selective
barrier, regulated by complex apical junctional complexes including
tight junctions (zona occludens1, occludin) and adherens junctions.
However, dysbiosis induced by Western diets, aging or obesity can
impair these junctional proteins, leading to increased intestinal
permeability. Guido et al (26) conducted a systematic review
highlighting that increased intestinal permeability is
significantly associated with the severity of OA symptoms and
radiographic progression, suggesting that barrier dysfunction is a
prerequisite for gut-derived inflammation to reach the joint.
Furthermore, Escalante et al (7) recently elucidated that age-related
deterioration of the gastrointestinal tract exacerbates this
permeability, thereby permitting the paracellular leakage of
luminal antigens into the lamina propria.
Once the physical barrier is compromised, the immune
tolerance of the gut is breached. Jiang et al (27) identified that the
mechanosensitive channel Piezo1 plays a critical role in regulating
intestinal inflammation and barrier function. This offers a novel
molecular explanation for how mechanical signals in the gut might
influence systemic immunity (27). The breakdown of these defense
mechanisms allows for the influx of pathogen-associated molecular
patterns, such as lipopolysaccharides (LPS), into the portal
circulation. This phenomenon, known as metabolic endotoxemia,
triggers a low-grade systemic inflammatory state. Such a state is
highly characteristic of metabolic OA phenotypes (21).
Translocation to the joint
The second segment of this biological highway
involves the physical translocation of microbial components and
metabolites from the circulation into the synovial fluid and
cartilage matrix. Historically, the joint was considered a sterile
environment; however, the application of next-generation sequencing
has challenged this dogma. Bardi et al (28) performed a systematic review of
synovial fluid microbiota characterization, confirming the presence
of bacterial DNA signatures in OA patients that closely resemble
intestinal microbial profiles. This suggests that bacterial genetic
material or dormant bacteria can traverse the endothelial barrier
of the synovial microvasculature. He et al (29) further proposed the concept of
'joint dysbiosis', where the local accumulation of translocated
microbial products directly alters the homeostasis of the joint
microenvironment.
A critical mechanism facilitating this transport
involves BEVs. Niu et al (24) demonstrated that BEVs act as a
'Trojan horse', encapsulating bioactive bacterial components and
protecting them from degradation in the bloodstream until they
reach the target joint tissue. Upon arrival, these vesicles can
fuse with synoviocytes or chondrocytes, delivering their cargo.
Consequently, the presence of LPS and other bacterial metabolites
in the joint activates Toll-like receptors (TLRs), particularly
TLR4, on the surface of macrophages and chondrocytes. Yang et
al (25) detailed how this
activation triggers pyroptosis, a highly inflammatory form of
programmed cell death, thereby accelerating cartilage matrix
degradation and osteophyte formation.
Furthermore, the translocation process is not
limited to pro-inflammatory agents but also includes metabolic
regulators. Li et al (30) discussed how gut
microbiota-derived metabolites, once translocated, directly
regulate bone extracellular matrix homeostasis through specific
signaling pathways. This implies that the 'biological highway' is a
dual-use route, transporting both destructive endotoxins and
potentially therapeutic metabolites depending on the state of the
gut microbiome. Zhang et al (31) provided recent evidence that this
microbial regulation extends to the Wnt/β-catenin signaling
pathway, which is pivotal for joint remodeling and cartilage
repair. Therefore, the gut-joint axis functions as a continuous
feedback loop where intestinal permeability determines the load of
systemic trafficking, ultimately dictating the metabolic and
inflammatory fate of the OA joint.
The anti-inflammatory guardians
The integrity of the gut-joint axis, as previously
described, determines the systemic dissemination of microbial
metabolites, with their biological impact contingent upon their
specific chemical nature (16).
In contrast to pro-inflammatory agents like TMAO, SCFAs, primarily
acetate, propionate and butyrate, emerge as a crucial class of
gut-derived metabolites with demonstrated anti-inflammatory and
tissue-protective properties (16). Synthesized from dietary fiber
fermentation, SCFAs are now recognized as essential signaling
molecules that modulate joint homeostasis and combat OA progression
through multifaceted mechanisms (32,33). The multifaceted protective
mechanisms of SCFAs, spanning receptor signaling, epigenetic
regulation and immunomodulation, are supported by a growing body of
experimental and clinical evidence, as summarized in Table I, which is organized to highlight
the receptor target, molecular mechanism and biological outcome in
joint health for each key study. Most of the detailed mechanistic
insights into SCFA actions, such as G-protein coupled receptor
(GPCR) engagement, histone deacetylase (HDAC) inhibition and
autophagy regulation, derive from preclinical models (cell cultures
and animal studies) (34,35).
Where available, human translational evidence is explicitly noted.
The following subsections first summarize the preclinical
mechanistic framework and then highlight clinically validated
findings.
 | Table ISummary of key preclinical and
clinical evidence elucidating the protective roles of SCFAs in OA
and associated musculoskeletal disorders. |
Table I
Summary of key preclinical and
clinical evidence elucidating the protective roles of SCFAs in OA
and associated musculoskeletal disorders.
| Authors, year | Study
model/subjects | Specific
metabolite/intervention | Target
receptor/signaling node | Molecular
mechanism | Biological outcome
involved in joint health | (Refs.) |
|---|
| Pirozzi et
al, 2018 | Human OA
chondrocytes | Butyrate | GPR43 (FFAR2) | Inhibition of NF-κB
nuclear translocation and reduction of inflammatory cytokines | Attenuation of
IL-1β-induced inflammation and cartilage matrix degradation | (34) |
| Zhou et al,
2021 | Rat OA model
(ACLT)/chondrocytes | Sodium
butyrate |
HDAC/PI3K/Akt/mTOR | Restoration of
impaired autophagic flux (LC3II/p62 modulation) | Inhibition of
chondrocyte apoptosis and attenuation of cartilage destruction | (35) |
| Tang et al,
2020 | Ovariectomized
rats/osteoblasts | Sodium
butyrate | GSK-3β/Nrf2 | Enhancement of
mitochondrial function and antioxidant defense | Prevention of
subchondral bone loss and promotion of osteoblastic activity | (42) |
| Duscha et
al, 2022 | Human cohort
(n=290)/in vitro | Propionic acid | Tregs | Enhancement of Treg
functionality and suppression of Th1/Th17 axis | Improvement of
systemic bone health biomarkers and immunomodulation | (43) |
| Bo et al,
2018 | Human OA
chondrocytes | Sodium
butyrate | MMPs/ADAMTSs | Downregulation of
catabolic enzymes expression (MMP-13, ADAMTS-5) | Prevention of type
II collagen degradation and preservation of chondrocyte
phenotype | (44) |
| Cho et al,
2022 | Rat OA model
(MIA)/chondrocytes |
Lactobacillus (SCFA
producer)/butyrate | AMPK/mTOR | Suppression of
NLRP3 inflammasome-mediated pyroptosis | Reduction of
inflammatory cell death and relief of OA pain behaviors | (45) |
| Wu et al,
2022 | Murine macrophages
(RAW264.7) |
Propionate/butyrate | GPR43/NLRP3 | Inhibition of ROS
production and caspase-1 activation | Attenuation of wear
particle-induced macrophage pyroptosis and osteoclastogenesis | (48) |
| Korsten et
al, 2024 | Human OA patients
(RCT) | Sustained-release
butyrate tablet | T helper cells | Systemic
immunomodulation of effector T-cell responses | Suppression of
ex vivo T-cell activation, demonstrating translational
potential | (51) |
Biosynthesis and systemic distribution of
SCFAs
SCFAs are primarily generated in the cecum and colon
through the anaerobic fermentation of indigestible carbohydrates by
commensal bacteria such as Faecalibacterium prausnitzii,
Roseburia spp. and Eubacterium rectale (32). The levels and ratios of
individual SCFAs are dynamically influenced by host diet, with
high-fiber intake robustly promoting their production, whereas
Western-style diets are linked to reduced SCFA levels and gut
dysbiosis (33). Following
production, SCFAs are absorbed through the colonic epithelium.
Butyrate is preferentially metabolized by colonocytes, while
acetate and propionate enter the portal circulation. A significant
proportion evades hepatic clearance, achieving systemic
bioavailability and enabling these metabolites to reach peripheral
tissues, including synovial joints and bone marrow (36,37). This distribution establishes a
direct biochemical link between colonic microbial ecology and
musculoskeletal tissue metabolism, positioning SCFAs as systemic
mediators of the gut-joint axis.
SCFAs as regulators of inflammatory
signaling pathways
A primary mechanism of SCFA action involves
activation of specific G protein-coupled receptors, namely GPR41
(FFAR3), GPR43 (FFAR2) and GPR109A, expressed on immune cells,
chondrocytes and synoviocytes (38). Ligand binding to these receptors
initiates intracellular signaling that suppresses key inflammatory
cascades. Li et al (38)
demonstrated that in endothelial cells, SCFAs inhibit LPS or
TNF-α-induced inflammation via G protein-coupled receptor 41/43
activation and concurrent HDAC inhibition. In chondrocytes, this
receptor-mediated action is particularly relevant. Pirozzi et
al (34) provided pivotal
evidence that butyrate mitigates IL-1β-induced inflammation and
matrix degradation predominantly through GPR43. This effect
frequently involves the suppression of the nuclear factor-kappa B
(NF-κB) pathway, a central driver of OA pathology. For instance,
sodium butyrate reduces inflammatory responses in bovine mammary
epithelial cells by inactivating NF-κB signaling (39). Sun et al (40) further showed that butyrate
ameliorates high-fat-diet-induced inflammation partly by activating
peroxisome proliferator-activated receptor α. Thus, through GPCR
engagement, SCFAs directly interfere with pro-inflammatory
signaling hubs within joint tissues.
Epigenetic modulation and autophagy
regulation by SCFAs
Beyond receptor activation, SCFAs function as potent
endogenous inhibitors of HDAC, leading to histone hyperacetylation
and altered gene transcription (41). This epigenetic regulation
promotes an anti-inflammatory and pro-reparative cellular state. In
the context of OA, this mechanism is critical for maintaining
cellular homeostasis. Zhou et al (35) elucidated that sodium butyrate
attenuates cartilage degradation by restoring impaired autophagy
and autophagic flux in chondrocytes. Autophagy is a vital cellular
clearance process and its dysregulation contributes to OA
progression. The HDAC inhibitory activity of butyrate is central to
this effect, influencing the expression of autophagy-related genes.
This epigenetic action extends to bone metabolism, where SCFAs
promote osteoblast function and inhibit osteoclastogenesis. Tang
et al (42) demonstrated
that sodium butyrate prevents osteoporosis in rats by promoting the
osteal glycogen synthase kinase-3β/nuclear factor erythroid
2-related factor 2 signaling axis and improving mitochondrial
function. Furthermore, Duscha et al (43) reported that propionic acid
beneficially modulates osteoporosis biomarkers in patients with
multiple sclerosis, indicating a translatable systemic
bone-protective effect via similar mechanisms.
Direct protective effects on chondrocytes
and cartilage matrix
SCFAs exert direct cytoprotective effects on
articular chondrocytes, preserving their phenotype and inhibiting
catabolism. Bo et al (44) showed that sodium butyrate
abolishes the degradation of type II collagen in human
chondrocytes, a hallmark of OA cartilage destruction. This
protection involves the downregulation of matrix-degrading enzymes
such as Matrix metalloproteinases and a disintegrin and
metalloproteinase with thrombospondin motifs. Furthermore, SCFAs
can counteract cellular senescence and apoptosis in chondrocytes
exposed to inflammatory stress. Cho et al (45) demonstrated that
Lactobacillus and butyrate inhibit OA by controlling
autophagy and inflammatory cell death (pyroptosis) of chondrocytes.
This direct cellular protection is synergistic with their
anti-inflammatory actions, creating a comprehensive defense for
cartilage matrix integrity against the degradative OA
microenvironment.
Immunomodulatory effects in the synovial
microenvironment
The synovial inflammation in OA is characterized by
immune cell infiltration and cytokine production. SCFAs play a
decisive role in modulating this synovial immune landscape. They
promote the polarization of macrophages from a pro-inflammatory M1
phenotype towards an anti-inflammatory, tissue-reparative M2
phenotype (46,47). Wu et al (48) confirmed that propionate and
butyrate attenuate macrophage pyroptosis and osteoclastogenesis
induced by wear particles. This modulation is crucial for reducing
synovitis and subchondral bone resorption. SCFAs also regulate
other immune cells; for example, butyrate has been shown to
ameliorate allergic inflammation by limiting eosinophil trafficking
and survival (49), and to
constrain neutrophil functions in inflammatory settings (50). Korsten et al (51) provided clinical translational
evidence, showing that a sustained-release butyrate tablet
suppresses ex vivo T helper cell activation in patients with
OA. By reshaping the immune response within the joint towards a
more regulatory state, SCFAs effectively dampen the chronic
low-grade inflammation that fuels OA progression.
In conclusion, SCFAs serve as fundamental
anti-inflammatory guardians within the gut-joint axis. Their
pleiotropic mechanisms, which span GPCR signaling, epigenetic
regulation, direct chondroprotection and immunomodulation,
establish them as critical metabolites for maintaining joint
homeostasis. Strategies aimed at augmenting SCFA levels, such as
dietary fiber supplementation, prebiotics, probiotics or even
postbiotic administration, therefore represent promising,
multi-targeted approaches for the prevention and management of OA.
This perspective resonates strongly with the holistic principles of
contemporary sports and regenerative medicine.
The pro-inflammatory aggressor
In stark contrast to the protective and reparative
nature of SCFAs, TMAO has emerged as a potent pro-inflammatory
metabolite that exacerbates musculoskeletal degeneration. While
originally identified as a cardiovascular risk factor, accumulating
evidence suggests that TMAO acts as a systemic 'aggressor'
affecting cartilage viability, subchondral bone architecture and
muscle function. Elevated serum TMAO levels are increasingly
recognized as a metabolic link connecting dietary habits, gut
dysbiosis and the low-grade systemic inflammation (inflammaging)
observed in metabolic OA phenotypes. As depicted in the
accompanying conceptual framework, TMAO operates through a
multi-hit mechanism involving oxidative stress induction,
inflammasome activation and the disruption of the gut-kidney-joint
axis. The molecular mechanisms linking TMAO to joint and bone
pathology, including NLRP3 inflammasome activation, NF-κB signaling
and Piezo1 upregulation, have been primarily established in
cellular and rodent models. However, corroborating clinical
evidence exists for associations between circulating TMAO levels
and adverse musculoskeletal outcomes, as detailed in this chapter.
The following discussion distinguishes preclinical mechanistic
findings from human observational data.
The diet-microbe-host axis
The biosynthesis of TMAO is a classic example of
co-metabolism between the host and the gut microbiome. It begins
with the dietary intake of quaternary amine precursors such as
choline, L-carnitine and phosphatidylcholine, which are abundant in
red meat, eggs and dairy products. Alisson-Silva et al
(52) reviewed the human risk
associated with high red meat intake, noting that unabsorbed
precursors reach the cecum and colon. Here, specific bacterial
clostridia possess the cutC/D gene cluster (choline utilization) or
cntA/B genes (carnitine monooxygenase), converting these substrates
into trimethylamine (TMA).
TMA is a volatile gas that traverses the intestinal
barrier and enters the portal circulation to reach the liver. In
hepatocytes, flavin-containing monooxygenases (FMOs), specifically
the FMO3 isoform, oxidize TMA into TMAO. Thomas and Fernandez
(53) highlighted that this
conversion is highly variable and influenced by host genetics,
hormonal status and the extent of gut dysbiosis. Crucially, this
axis is bidirectional; studies indicate that TMAO itself can
further alter the gut microbiota composition. For instance, sports
nutrition reviews such as that by Sawicka et al (54) have scrutinized L-carnitine
supplementation, noting that while it aids performance, chronic
high-dose ingestion in the presence of a dysbiotic gut may
inadvertently fuel TMAO production. Furthermore, since TMAO is
primarily cleared by the kidneys, renal function plays a
gatekeeping role. Lau et al (55) and Lei et al (56) described a
'gut-kidney-vascular-bone axis', suggesting that even mild renal
insufficiency, which is common in elderly OA populations, can lead
to the accumulation of uremic toxins like TMAO, thereby amplifying
systemic osteotoxicity.
Molecular mechanisms of joint and tissue
damage
Once in the systemic circulation, TMAO translocates
to synovial joints and skeletal muscles, where it exerts
deleterious effects through several converging molecular pathways.
The most prominent mechanism involves the induction of
mitochondrial dysfunction and oxidative stress.
Oxidative stress and senescence
TMAO has been shown to impair mitochondrial electron
transport, leading to an excessive production of reactive oxygen
species (ROS). In the context of bone and cartilage, Li et
al (57) demonstrated that
TMAO promotes oxidative stress, which in turn suppresses Sirtuin 6
(Sirt6), a longevity gene essential for DNA repair and genomic
stability. The downregulation of Sirt6 accelerates cellular
senescence in chondrocytes and osteoblasts, impairing their ability
to maintain the extracellular matrix. Similarly, Zou et al
(58) utilized metabolomic
analysis on C2C12 myoblasts to show that TMAO treatment exacerbates
oxidative damage in muscle cells, providing a metabolic basis for
the sarcopenia often comorbid with OA.
Inflammasome activation and NF-κB
signaling
Beyond direct oxidative damage, TMAO is a potent
activator of the innate immune system. Zhang et al (59) provided pivotal evidence that TMAO
triggers the activation of the NLR family pyrin domain containing 3
(NLRP3) inflammasome. This multiprotein complex is responsible for
the cleavage and secretion of the pro-inflammatory cytokines IL-1β
and IL-18. In the OA joint, IL-1β is the 'master cytokine' driving
cartilage catabolism. Notably, this activation often occurs in
parallel with the NF-κB pathway. Wang et al (60) and Zhao et al (61) independently confirmed that TMAO
enhances osteoclast differentiation and bone resorption via the
ROS-dependent NF-κB signaling cascade. This leads to an uncoupling
of bone remodeling, characterized by increased osteoclast activity
and suppressed osteoblast function, ultimately resulting in
subchondral bone loss and fragility.
Mechanosensitivity and the
'mechanical-metabolic' interface
Perhaps the most intriguing mechanism for sports
medicine is the interaction between TMAO and mechanotransduction.
Zhuang et al (62)
recently reported that TMAO sensitizes chondrocytes to mechanical
loading through the upregulation of Piezo1, a mechanosensitive ion
channel. Under physiological loading, Piezo1 maintains homeostasis;
however, in a TMAO-rich environment, the channel becomes
hypersensitive, converting normal mechanical signals into
pro-inflammatory responses and calcium influx overload. This
finding suggests that metabolic dysregulation (high TMAO) makes the
joint more susceptible to mechanical injury, fundamentally linking
the 'wear and tear' theory with metabolic pathology.
Clinical correlations: Bone, muscle and
cartilage
The mechanistic toxicity of TMAO translates into
observable clinical outcomes across the musculoskeletal spectrum.
High circulating levels of TMAO are no longer viewed solely as
cardiovascular markers but are now implicated in 'osteosarcopenic
obesity'.
Osteoporosis and bone quality
Several clinical studies have established a negative
correlation between serum TMAO and bone mineral density. Elam et
al (63), in a large-scale
analysis of the Cardiovascular Health Study, found that elevated
TMAO levels were associated with a higher risk of hip fractures in
older adults, independent of traditional risk factors. This is
supported by Lin et al (64), who observed that TMAO impairs the
functional capacity of bone marrow mesenchymal stem cells,
diverting their differentiation away from osteogenesis and towards
adipogenesis. This 'fatty marrow' conversion compromises bone
quality and structural support for the overlying cartilage.
Sarcopenia and muscle function
Given the close functional unit of the muscle and
joint, muscle wasting (sarcopenia) significantly accelerates OA
progression by reducing joint stability. Mo et al (65) elucidated a 'gut-TMAO-muscle'
axis, demonstrating that high-fat diet-induced TMAO accumulation
drives sarcopenic obesity by impairing muscle protein synthesis and
promoting proteolysis. Furthermore, Lin et al (66) found an association between uremic
toxins (including TMAO precursors) and reduced skeletal muscle mass
in patients with compromised renal function. This suggests that
TMAO-mediated myopathy may precede or exacerbate articular
pathology.
Synovial inflammation
In the specific context of joint disease,
Murillo-Saich et al (67)
performed metabolomic profiling of synovial tissue, identifying
distinct metabolic signatures associated with inflammation. While
direct synovial TMAO quantification is an emerging field, the
presence of its downstream effectors and the strong association
between gut dysbiosis and hand OA severity, as noted by Silvestre
et al (68), supports the
hypothesis that this metabolite is a key driver of synovitis.
Additionally, Huang et al (69) showed in animal models that
periodontal inflammation (another source of systemic bacterial
burden) aggravates TMAO metabolism, further linking oral-gut
dysbiosis to systemic inflammatory loads that burden the joint.
Collectively, these data position TMAO as a critical therapeutic
target, where reducing its generation could simultaneously benefit
cardiovascular, renal and musculoskeletal health. Key experimental
and clinical findings linking TMAO to musculoskeletal dysfunction
are consolidated in Table II,
which categorizes evidence by tissue/cell type, molecular pathway
and translational outcome to help readers trace the mechanistic
chain from TMAO elevation to bone, muscle or synovial pathology.
Before translating these findings into clinical strategies, it is
essential to examine how well the mechanistic evidence from model
systems aligns with human OA pathophysiology.
| Table IISummary of experimental and clinical
evidence linking TMAO to OA pathogenesis and musculoskeletal
dysfunction. |
Table II
Summary of experimental and clinical
evidence linking TMAO to OA pathogenesis and musculoskeletal
dysfunction.
| Authors, year | Study
model/subjects | TMAO-related
intervention/exposure | Target tissue/cell
type | Molecular
mechanism/signaling pathway | Key biological
outcome | (Refs.) |
|---|
| Li et al,
2019 | Aging mice and
osteoblast culture | High TMAO diet | Bone
tissue/osteoblasts | Oxidative stress
and Sirt6 suppression | Accelerated bone
aging, reduced bone quality | (57) |
| Zou et al,
2022 | C2C12 myoblasts
under oxidative stress | TMAO treatment | Skeletal muscle
cells | Mitochondrial
dysfunction and ROS overproduction | Exacerbated
oxidative damage in muscle cells | (58) |
| Zhang et al,
2020 | Vascular smooth
muscle cells and calcification models | TMAO treatment | Vascular
cells/osteoclast precursors | NLRP3 inflammasome
activation and NF-κB signaling | Promotion of
vascular calcification and osteogenic differentiation | (59) |
| Wang et al,
2022 | Osteoclast
precursors (RAW264.7) and ovariectomized mice | TMAO
administration | Bone marrow-derived
macrophages | ROS-dependent NF-κB
activation | Enhanced osteoclast
differentiation and bone loss | (60) |
| Zhao et al,
2024 | Osteoclast
precursors and mouse calvarial osteolysis model | TMAO exposure | Osteoclasts/bone
tissue | NF-κB/MAPK pathway
activation | Increased
osteoclastogenesis and bone resorption | (61) |
| Zhuang et
al, 2023 | Human chondrocytes
and mechanical loading model | TMAO
pre-treatment | Articular
chondrocytes | Upregulation of
Piezo1 mechanosensitive channel | Sensitization to
mechanical stress, promoting inflammatory response | (62) |
| Elam et al,
2022 | Older adults
(cardiovascular health study) | Serum TMAO
measurement | Systemic/BMD | Not specified
(observational) | Association between
high TMAO levels and increased hip fracture risk | (63) |
| Lin et al,
2020 | Osteoporotic
patients and BMSC culture | TMAO exposure | BMSCs | Impaired osteogenic
differentiation | Reduced bone
formation and increased adipogenesis in BMSCs | (64) |
| Mo et al,
2025 | Aging rats on
high-fat diet | Diet-induced TMAO
elevation | Skeletal
muscle | Gut-muscle axis
dysregulation | Promotion of
sarcopenic obesity and muscle atrophy | (65) |
| Murillo-Saich et
al, 2022 | OA patients
(synovial tissue metabolomics) | Metabolomic
profiling | Synovial
tissue | Inflammatory
metabolite signature (incl. TMAO-related pathways) | Correlation with
synovitis severity and metabolic dysregulation | (67) |
| Huang et al,
2025 | ApoE(−/−) mice with
periodontitis | Periodontal
treatment | Systemic/gut-kidney
axis | Modulation of TMAO
metabolism | Reduced systemic
TMAO and inflammatory burden | (69) |
Translational considerations for
TMAO-mediated pathogenesis
The translational relevance of TMAO-induced
pathogenic mechanisms in human OA is supported by several lines of
clinical evidence. Large-scale epidemiological studies have
established that elevated serum TMAO levels are associated with
increased risk of hip fractures 63 and reduced bone mineral density
(57), consistent with the
osteoclastogenic effects observed in murine models. Synovial tissue
metabolomic profiling of patients with OA has identified
TMAO-related metabolic signatures that correlate with synovitis
severity (67), suggesting that
TMAO accumulates in human joint tissues and contributes to local
inflammation. Furthermore, the observation that periodontal
treatment reduces systemic TMAO levels and inflammatory burden
provides indirect evidence that modulating TMAO metabolism can
alter systemic inflammation in humans (69). However, direct evidence that TMAO
activates the NLRP3 inflammasome or induces chondrocyte senescence
in human OA joints remains limited, and it is unclear whether the
concentrations of TMAO achieved in the human circulation are
sufficient to replicate the robust effects observed in cell culture
studies. Future human studies should measure TMAO levels in
synovial fluid and correlate them with local expression of NLRP3
inflammasome components and chondrocyte senescence markers to
validate these mechanistic pathways in vivo.
The complex regulators
BAs are historically viewed merely as digestive
surfactants required for lipid absorption; however, contemporary
research has redefined them as potent signaling steroid hormones
that regulate systemic metabolic homeostasis. Following synthesis
in the liver as primary BAs, these molecules undergo significant
modification by gut microbiota into secondary BAs, such as
deoxycholic acid (DCA) and lithocholic acid (LCA). These
metabolites re-enter the systemic circulation and reach distant
organs, including the synovial joints, where they interact with
specific nuclear and membrane receptors. In the context of OA, BAs
exhibit a dual nature, functioning as either protective agents or
destructive proinflammatory mediators depending on their
hydrophobicity and concentration, and the specific receptor
activated. Preclinical studies [e.g., FXR agonist studies in murine
OA models, Takeda G-protein-coupled receptor 5 (TGR5) activation in
chondrocyte cultures and DCA-induced pyroptosis in hepatocytes]
have defined the core BA signaling pathways. Recent human
metabolomic profiling and interventional trials have started to
validate these pathways in patients with OA (18,19). The following subsections clearly
separate preclinical mechanistic evidence from clinical
translational data.
Microbiota-driven alterations in the BA
pool
The composition of the systemic BA pool is dictated
by the metabolic activity of the intestinal microbiome. Dysbiosis,
characterized by a reduction in beneficial commensals and an
increase in pathogenic strains, fundamentally alters the ratio of
primary to secondary BAs. Chen et al (70) demonstrated that maintaining
gut-liver axis homeostasis through microbiota-mediated secondary BA
pathways is essential for systemic health. When this balance is
disrupted, there is an accumulation of hydrophobic secondary BAs,
which are often cytotoxic at high concentrations. This perturbation
is not merely local to the gut; rather, it leads to increased
intestinal permeability and the translocation of these metabolites
into the blood, eventually accumulating within the synovial fluid
of joints. Yu et al (71)
identified that BA insufficiency and specific metabolic alterations
are mechanically linked to oxidative stress-mediated pathology,
providing a bridge between metabolic liver diseases and the
development of OA. Consequently, the specific profile of BAs
circulating in the host serves as a critical determinant of whether
the joint microenvironment remains homeostatic or shifts toward a
catabolic state.
FXR: The protective guardian
The FXR acts as a nuclear transcription factor and
is widely regarded as a protective sensor in joint tissues. Under
physiological conditions, FXR activation suppresses inflammatory
signaling and maintains metabolic quiescence in chondrocytes. Hu
et al (72) provided
compelling evidence regarding the therapeutic potential of this
pathway; they found that an FXR agonist significantly attenuated
osteochondral pathologies in an OA model. Their study revealed that
FXR activation suppressed the c-Jun N-terminal kinase 1/2/nuclear
factor of activated T-cells 1 pathway, thereby inhibiting
osteoclast fusion in the subchondral bone (72). This is a crucial finding because
it suggests that BAs affect not only the cartilage surface but also
the underlying bone remodeling, which is a key driver of OA
progression. Furthermore, the anti-inflammatory capacity of FXR is
linked to the inhibition of the NLRP3 inflammasome. Sun et
al (73) reported that
regulation of the FXR-NLRP3 signaling pathway could alleviate
inflammatory conditions, suggesting a similar mechanism may protect
synovial tissues from cytokine-induced degradation. Therefore,
strategies that preserve FXR expression or enhance its activation
by specific BA ligands represent a promising avenue for halting the
structural deterioration of the joint.
The TGR5 and NLRP3 axis: A
context-dependent interaction
While FXR generally exerts protective effects, the
role of the TGR5 and its interaction with specific secondary BAs is
more complex and context-dependent. TGR5 is expressed on
chondrocytes and immune cells within the synovium and its
activation has been shown to inhibit catabolic enzymes. Huang et
al (74) demonstrated that
activation of TGR5 ameliorates IL-1β induced chondrocyte
senescence, indicating a direct anti-aging effect on the cartilage
matrix. Similarly, Zhuo et al (75) observed that TGR5 activation
inhibits the degradation of type II collagen and aggrecan in human
chondrocytes, further supporting a chondroprotective role for this
receptor.
However, the specific ligands available to bind
these receptors can drastically alter the outcome. High levels of
hydrophobic secondary BAs, particularly DCA, have been implicated
in triggering inflammation rather than resolving it. Mai et
al (76) highlighted that
DCA promotes pyroptosis, a highly inflammatory form of programmed
cell death, by inhibiting mitophagy and activating the NLRP3
inflammasome. This aligns with findings by Holtmann et al
(77), who reported that certain
BAs specifically activate the NLRP3 inflammasome to promote
inflammation in a cell-type-specific manner.
However, not all secondary BAs are detrimental;
Zhong et al (78) found
that LCA, another secondary BA, could alleviate inflammatory
conditions via inhibition of the NLRP3 inflammasome, contrasting
with the effects of high-dose DCA. Additionally, Liu et al
(79) reported that LCA
ameliorates inflammation via the pregnane X receptor
(PXR)/TLR4/NF-κB/NLRP3 signaling pathway, further illustrating the
nuance that different metabolites from the same class can have
opposing biological effects. Thus, the 'complex regulator'
designation arises from this delicate balance: TGR5 activation is
beneficial, but an excess of cytotoxic secondary BAs like DCA can
bypass these protective checks to trigger the NLRP3 inflammasome
and induce chondrocyte death.
Clinical evidence and translational
perspectives
The transition from basic science to clinical
reality is supported by recent metabolomic profiling of patients
with OA. Li et al (18)
conducted a comprehensive analysis of BA metabolism in patients
with symptomatic hand OA, providing direct clinical evidence of
altered BA profiles in human OA populations. Their work validates
the hypothesis that systemic metabolic perturbations are reflected
in the clinical phenotype of joint disease. Furthermore, the
connection between metabolic health and joint integrity is
reinforced by broader comorbidity studies. Zemedikun et al
(80) and Sharafi et al
(81) utilized latent class
analysis to identify comorbidity phenotypes, consistently finding
that metabolic clusters (often involving dyslipidemia and liver
dysfunction) are high-risk groups for severe OA and mortality.
Collectively, these studies suggest that the
'gut-joint axis' is not a passive system but an active signaling
highway mediated by BAs. The presence of specific BA transporters
and receptors on joint tissues renders the cartilage and synovium
sensitive to microbiome-derived metabolic shifts. In summary, BAs
regulate OA pathology through a tripartite mechanism: Protecting
subchondral bone via FXR, modulating chondrocyte senescence via
TGR5 and potentially driving inflammation via NLRP3 activation when
hydrophobic secondary BAs accumulate. Future therapeutic
interventions may focus on manipulating the gut microbiota to
optimize the BA pool or developing selective receptor modulators
that uncouple the anti-inflammatory benefits from the cytotoxic
risks. The dual and context-dependent roles of BAs, as elucidated
by pivotal preclinical and clinical studies, are detailed in
Table III, which separates
protective (FXR, TGR5) from detrimental (DCA, NLRP3) pathways and
includes recent human validation studies for easier comparison.
| Table IIIComplex and dual roles of bile acids
in OA pathophysiology: A summary of mechanistic insights from
preclinical and clinical studies. |
Table III
Complex and dual roles of bile acids
in OA pathophysiology: A summary of mechanistic insights from
preclinical and clinical studies.
| Authors, year | Study
model/subjects | Specific bile
acid/intervention | Target
receptor/signaling node | Molecular
mechanism | Biological outcome
in joint health | (Refs.) |
|---|
| Chen et al,
2025 | Weaned
piglets/gut-liver axis | Metasilicate water
(modulates BA pool) | Microbiota-mediated
secondary BA pathway | Maintenance of
gut-liver axis homeostasis | Systemic health
promotion, implying stabilized joint microenvironment | (70) |
| Yu et al,
2025 | PFOS-induced mouse
model | Bile acid
insufficiency | Oxidative stress
pathways | Linking BA
insufficiency to oxidative stress | Proposed mechanism
connecting metabolic liver disease to OA development | (71) |
| Hu et al,
2022 | Mouse OA model | FXR agonist |
FXR/JNK1/2/NFATc1 | Suppression of
JNK1/2/NFATc1 pathway | Attenuation of
subchondral bone osteoclast fusion and osteochondral pathology | (72) |
| Sun et al,
2023 | DSS-induced colitis
mice | Bacteroides
dorei BDX-01 | FXR-NLRP3 axis | Regulation of
intestinal BSH activity and FXR-NLRP3 signaling | Alleviation of
systemic inflammation, suggesting potential joint protection | (73) |
| Huang et al,
2018 | IL-1β-treated human
chondrocytes | TGR5 agonist | TGR5 (GPBAR1) | Activation of TGR5
signaling | Amelioration of
IL-1β-induced chondrocyte senescence | (74) |
| Zhuo et al,
2019 | IL-1β-treated human
chondrocytes | TGR5 agonist | TGR5 (GPBAR1) | Activation of TGR5
signaling | Inhibition of type
II collagen and aggrecan degradation | (75) |
| Mai et al,
2023 | Steatotic HepG2
cells | DCA | NLRP3
inflammasome | Inhibition of
mitophagy, activation of NLRP3 | Induction of
pyroptosis (pro-inflammatory cell death) | (76) |
| Holtmann et
al, 2021 | Murine liver
inflammation models | Specific bile
acids | NLRP3
inflammasome | Cell-type-specific
activation of NLRP3 | Promotion of
inflammation or fibrosis | (77) |
| Zhong et al,
2026 | Colitis mouse
model | LCA/EGCG | NLRP3
inflammasome | Gut
microbiota-derived LCA inhibits NLRP3 | Alleviation of
colitis inflammation, suggesting anti-inflammatory potential | (78) |
| Liu et al,
2025 | Ulcerative colitis
mouse model | LCA |
PXR/TLR4/NF-κB/NLRP3 | Modulation of
PXR/TLR4/NF-κB/NLRP3 axis and gut microbiota | Amelioration of
inflammation, highlighting metabolite-specific effects | (79) |
| Li et al,
2025 | Human patients with
symptomatic hand OA | Bile acid
metabolomics profile | Systemic metabolic
perturbation | Altered bile acid
metabolism | Direct clinical
correlation of BA dysregulation with human OA phenotype | (18) |
Translational validation of BA signaling
in human OA
Recent clinical studies have significantly advanced
the current understanding of BA signaling in human OA
pathophysiology. Li et al (18) conducted the first comprehensive
metabolomic analysis of BA profiles in patients with symptomatic
hand OA, identifying specific alterations in primary and secondary
BA levels that correlate with disease severity. Most notably, Yang
et al (19) demonstrated
in a landmark study that glucagon like peptide 1 receptor agonists
improve OA outcomes by targeting intestinal FXR signaling,
providing direct clinical validation of the therapeutic potential
of modulating BA receptors in human OA. This study confirmed that
the FXR-NLRP3 signaling axis, previously characterized in
preclinical models, plays a critical role in human disease
pathogenesis. However, the dual nature of BA signaling presents
unique translational challenges. While FXR activation is
consistently protective, the effects of TGR5 activation and
individual secondary BAs in human OA remain incompletely
understood. For example, while DCA induces pyroptosis in
vitro (76), it is unclear
whether the concentrations of DCA present in human synovial fluid
are sufficient to trigger this response. Further clinical studies
are needed to define the optimal BA profile for joint health and to
develop selective receptor modulators that maximize therapeutic
benefits while minimizing potential adverse effects.
Therapeutic implications: A sports medicine
perspective
The delineation of the gut-joint axis, mediated by
microbial metabolites such as SCFAs, TMAO and BAs, reframes OA
management. It advocates for a systemic strategy targeting
underlying metabolic and inflammatory dysregulation, moving beyond
local symptom control. This paradigm aligns with the holistic
principles of sports medicine, which emphasizes optimizing the
whole-body environment for tissue resilience and long-term
musculoskeletal health. Consequently, strategic modulation of the
gut microbiota and its metabolic output emerge as a compelling
therapeutic frontier. To translate these concepts into actionable
clinical practice, the following mechanism-guided strategies are
proposed. Fig. 3 serves as the
core therapeutic schematic of this review, summarizing the
multi-modal sports medicine interventions for precision OA
management via targeting the gut-joint axis. As illustrated,
exercise, dietary regulation, probiotic supplementation and
targeted pharmacotherapy collectively remodel gut microbial
metabolite profiles, including upregulating protective SCFAs,
inhibiting pro-inflammatory TMAO production and balancing BA
receptor signaling (20,51,82). This integrated graphical
framework systematizes mechanism-based interventions to restore
joint metabolic homeostasis and delay OA progression.
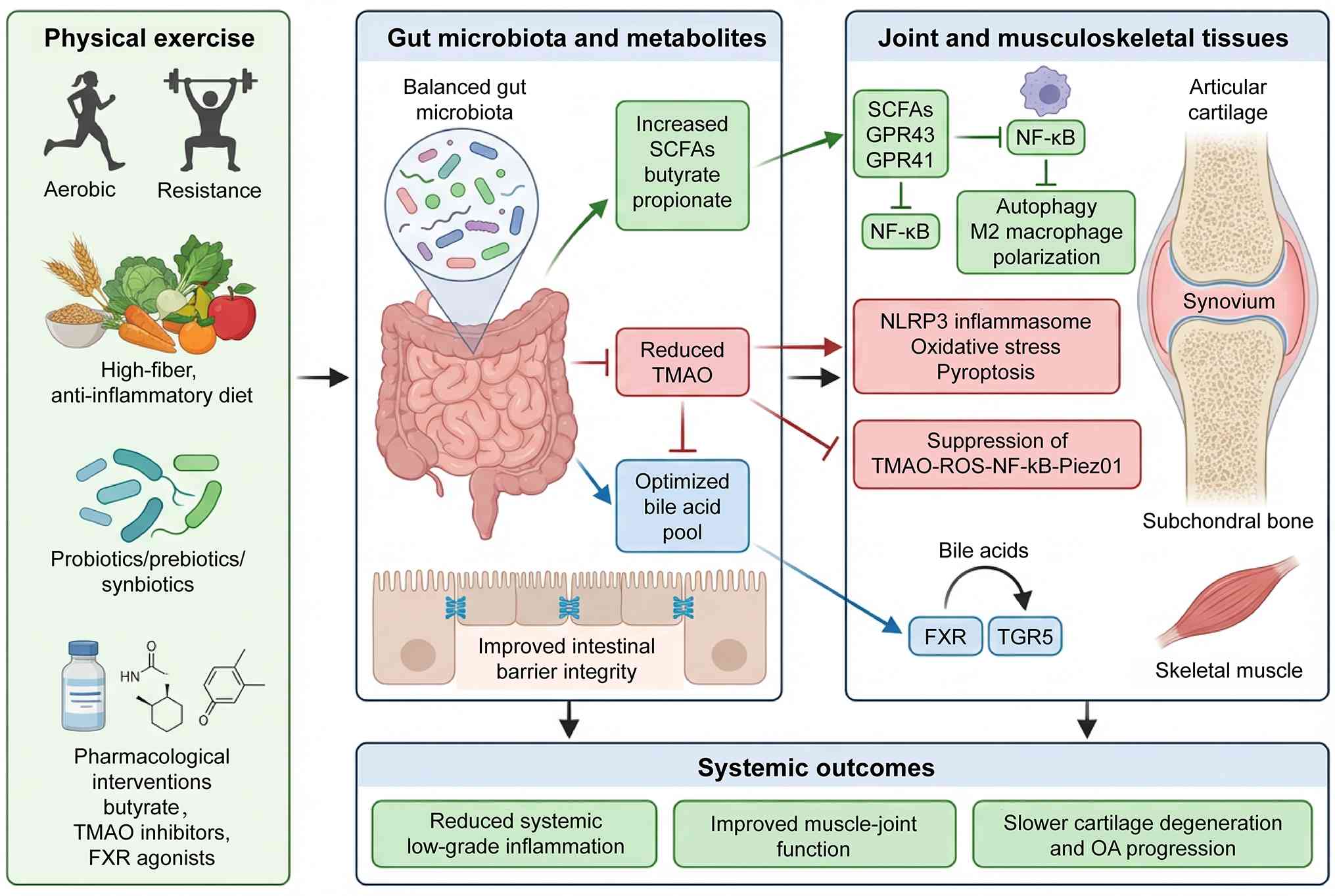 | Figure 3Therapeutic modulation of the
gut-joint axis in osteoarthritis: A sports medicine perspective.
Scientific diagram illustrating how exercise, diet, probiotics and
targeted metabolite-based therapies modulate the gut
microbiota-derived metabolites (SCFAs, TMAO, bile acids) to
influence the gut-joint axis in osteoarthritis, highlighting
anti-inflammatory SCFA signaling, inhibition of TMAO-driven
oxidative stress, and bile acid receptor regulation (FXR/TGR5) in
cartilage, synovium, bone and muscle. SCFAs, short-chain fatty
acids; TMAO, trimethylamine N-oxide; BAs, bile acids; FXR,
farnesoid X receptor; TGR5, Takeda G-protein-coupled receptor 5;
NLRP3, NLR family pyrin domain containing 3; NF-κB, nuclear factor
κB; ROS, reactive oxygen species; TLR4, Toll-like receptor 4. |
Exercise as a microbiome modulator:
Towards prescription
Physical activity is a cornerstone of sports
medicine and OA management, renowned for its direct benefits on
muscle strength and systemic inflammation. Compelling evidence now
positions regular exercise as a potent non-pharmacological
modulator of the gut ecosystem. Both aerobic and resistance
training are associated with increased gut microbial diversity, a
hallmark of a stable and functional microbiome often observed in
athletes.
This 'athlete's microbiome' is frequently enriched
with taxa capable of producing beneficial metabolites, particularly
SCFAs. Li et al (83)
demonstrated that moderate exercise ameliorated OA progression in
mice, an effect linked to reduced systemic LPS and a favorable
microbial shift. Further reinforcing this link, Hao et al
(21,84) showed that exercise intervention
altered gut microbiome profiles and serum metabolomics in rat
models of post-traumatic OA, promoting a less inflammatory
state.
Exercise also appears to counteract the detrimental
TMAO pathway. Brunt et al (85) reported that voluntary wheel
running mitigated vascular dysfunction and exercise intolerance
induced by a Western diet in mice, with suppression of circulating
TMAO implicated as a key mechanism. Furthermore, Pedersini et
al (86) reviewed the
evidence linking physical activity to gut microbiota composition,
supporting its role in maintaining gut barrier integrity and
microbial homeostasis. Therefore, tailored exercise prescriptions
serve a dual purpose: Directly enhancing musculoskeletal function
and indirectly cultivating a gut environment that favors protective
metabolites while suppressing harmful ones.
From a prescription standpoint, existing evidence
supports the use of moderate-intensity continuous training (MICT)
performed for 30-45 min per session, at least 3-5 times per week,
as a feasible regimen to increase gut microbial diversity and
SCFA-producing taxa (20). For
patients in whom MICT is limited by joint symptoms, accumulating
evidence suggests that resistance training and high-intensity
interval training may offer comparable or even superior benefits
for specific metabolites. However, data in OA populations remain
preliminary (83). Importantly,
the gut microbiome responds to exercise in a dose-dependent manner,
with responders showing increased Bifidobacterium and
Lactobacillus abundance, whereas non-responders exhibit a
resilient microbial profile that may require longer training
durations or adjunctive dietary modulation.
Nutritional interventions: Moving from
general advice to quantified targets
Dietary patterns are primary determinants of gut
microbiota composition and the systemic metabolite profile. From a
sports medicine perspective, nutritional strategies can be
optimized to foster a joint-protective gut environment. The
dichotomy between a Western diet and diets rich in fiber and
polyphenols is critical. The former promotes dysbiosis and elevates
TMAO production, while high intake of fermentable fibers stimulates
SCFA production. Fortuna et al (82) conducted a randomized controlled
trial demonstrating that prebiotic fiber supplementation in adults
with knee OA and obesity improved physical function and shifted gut
bacterial taxa beneficially.
Probiotic supplementation represents a direct
strategy to introduce beneficial microbes. Substantial evidence
supports specific Lactobacillus strains such as L.
casei Shirota (87), L.
acidophilus (88) and L.
rhamnosus (89), all of
which have been shown to improve knee symptoms, attenuate pain and
cartilage damage, or ameliorate OA progression. The efficacy of
Bifidobacterium species is also well-documented for OA and bone
health (90-92). Henrotin et al (91) demonstrated that
Bifidobacterium longum CBi0703 protected against spontaneous
OA in guinea pigs. Synbiotic combinations may offer synergistic
effects, as suggested by studies on bone metabolism (93-95). Ishizu et al (96) further highlighted the potential
in athletes, finding that prebiotic food intake may improve bone
resorption markers in Japanese female athletes. Thus, nutritional
counseling should prioritize anti-inflammatory, fiber-rich dietary
patterns, potentially augmented with evidence-based prebiotic or
probiotic supplements.
To operationalize these findings, a daily
fermentable fiber intake target of 25-35 g/day is recommended based
on data showing that this amount increases circulating acetate and
butyrate concentrations and reduces inflammatory markers in
patients with OA (82).
Furthermore, dietary interventions to lower TMAO should prioritize
reducing red meat intake to ≤2 servings per week, given that
L-carnitine is a major TMAO precursor (52). In athletic populations requiring
higher protein intake for muscle maintenance, alternative protein
sources such as plant-based proteins or egg white (which are lower
in TMA precursors than red meat) should be emphasized while
maintaining total protein goals.
Targeted pharmacotherapy and
personalization
Beyond lifestyle modifications, the precise
targeting of specific microbial metabolic pathways represents a
novel frontier in pharmacotherapy. The concept of 'postbiotics',
which involves administering beneficial bacterial metabolites,
holds significant promise. Butyrate, a principal SCFA, has
demonstrated chondroprotective effects in OA models (45,97). Translating this to humans,
Korsten et al (51)
conducted a double-blind trial showing that a sustained-release
butyrate tablet suppressed ex vivo T helper cell activation
in patients with OA, suggesting direct metabolite supplementation
could bypass variable microbial fermentation.
A major therapeutic target is the TMAO-generating
pathway. Inhibition of microbial enzymes like trimethylamine (TMA)
lyase can block TMA formation. Wang et al (98) and Fechtner et al (99,100) demonstrated that the inhibitor
3,3-dimethyl-1-butanol and its metabolite attenuated pathology in
models of heart failure and arthritis by reducing TMAO levels.
Natural compounds also show potential; Baptista et al
(101) investigated
resveratrol's effects on TMAO changes post-exercise in older
adults, while Huang et al (102) evaluated resveratrol butyrate
esters for interrupting TMA metabolism in vitro.
Furthermore, strategies to modulate BA receptors, such as FXR
agonists, have shown promise in preclinical OA models by protecting
subchondral bone (103). These
approaches underscore the potential for developing interventions
that selectively inhibit detrimental metabolites or supplement
beneficial ones.
A critical step toward personalization is the
integration of multi-omics data to identify patients who are most
likely to respond to specific interventions. Machine learning
algorithms have recently been employed to associate gut microbiota
compositions with physical functioning in patients with OA,
enabling the identification of distinct 'metabolic phenotypes'
(104). For example, patients
with high baseline TMAO may benefit more from TMA lyase inhibitors
or dietary red meat restriction, whereas those with low SCFA levels
may respond preferentially to prebiotic fiber or butyrate
supplementation. Furthermore, genetic factors such as polymorphisms
in the flavin-containing FMO3 gene, which encodes the hepatic
enzyme converting TMA to TMAO, can influence individual TMAO
responses to dietary choline (53). Screening for such variants could
identify patients with genetically determined high TMAO production
who would derive the greatest benefit from microbial TMA lyase
inhibition rather than dietary modification alone.
In summary, the sports medicine perspective is
uniquely equipped to implement a holistic strategy informed by the
gut-joint axis. Exercise establishes a foundational pro-homeostatic
shift in the gut microbiome. Nutrition provides the necessary
substrates to sustain this beneficial environment. Emerging
targeted therapies offer precision tools to correct specific
metabolic imbalances. The proposed algorithm provides a
mechanism-guided, implementable pathway for translating gut-joint
axis biology into clinical practice. This integrative, multi-modal
framework aligns with the preventive, systems-oriented ethos of
sports medicine, addressing the root systemic dysregulation in OA
and paving the way for personalized, mechanism-driven preservation
of joint health.
Challenges and future directions
The proposed 'metabolite homeostasis imbalance'
model has important implications for addressing the current
challenges in gut-joint axis research. This integrative framework
suggests that future studies should move beyond investigating
individual metabolites in isolation and instead focus on
characterizing the global metabolic profile of patients to
determine their specific 'metabolic imbalance phenotype'. Despite
the exponential growth in delineating the gut-joint axis, a
significant dichotomy remains between the robust causal evidence
established in animal models and the largely associative nature of
human clinical studies. Most current research relies on
cross-sectional sequencing of the microbiome, which captures a
static snapshot of dysbiosis rather than the dynamic temporal
changes preceding joint degeneration. While recent investigations
using fecal microbiota transplantation in germ-free mice have
successfully demonstrated that OA susceptibility is transferable
and immunologically mediated (105), translating these findings to
human cohorts remains complex due to confounding lifestyle
variables. Longitudinal studies are scarce but essential to
determine whether gut dysbiosis is a precursor to OA or a
consequence of disease-related lifestyle changes such as immobility
and analgesic use. Recent efforts analyzing gut microbiomics in
relation to sustained knee pain represent a positive step toward
establishing temporality (106), yet large-scale prospective
cohorts integrating multi-omics are required to definitively map
the transition from gut dysbiosis to the onset of articular
cartilage degradation.
A critical challenge in gut-joint axis research is
the translation of mechanistic findings from cellular and animal
models to human OA pathophysiology. While reductionist models are
essential for defining molecular pathways, such as TMAO activating
NLRP3 inflammasomes or SCFAs inhibiting NF-κB signaling, the human
joint microenvironment is complex and validation in clinical
settings remains limited. Nevertheless, convergent human evidence
already supports these mechanisms, including Mendelian
randomization studies establishing causal links between gut
microbiota and OA risk (10,11), human synovial fluid metabolomics
identifying TMAO-related signatures associated with synovitis
(67), clinical trials
demonstrating butyrate-mediated immunomodulation in patients with
OA (51) and large-scale
metabolomic and interventional studies validating BA-FXR signaling
in human OA (18,19). Future efforts should prioritize
direct quantification of these metabolites in human synovial fluid
and single-cell analyses of patient-derived joint tissues to
further bridge the translational gap.
Furthermore, the influence of sexual dimorphism on
the gut-joint axis represents a critical knowledge gap that current
literature often overlooks. OA prevalence exhibits a distinct
female predominance particularly after menopause, yet many
preclinical mechanistic studies utilize male animals to avoid
hormonal variability, thereby obscuring potential sex-specific
microbial interactions. Recent evidence indicates that sex
differences significantly alter how external factors like alcohol
or high-fat diets impact the intestinal flora and subsequent bone
resorption (107,108). The interplay between estrogen
depletion and the microbiome is pivotal, as estrogens regulate both
gut barrier integrity and microbial composition (109). Consequently, future
investigative frameworks must rigorously stratify data by sex to
determine if the 'estrogen-gut-bone axis' requires distinct
therapeutic approaches for male vs. female patients, potentially
explaining the variable efficacy of metabolic interventions
observed in clinical trials.
A second major challenge lies in deciphering the
precise molecular mechanisms by which specific metabolites
influence joint tissue homeostasis beyond general anti-inflammatory
effects. While the protective roles of SCFAs and the detrimental
impact of TMAO are broadly categorized, the specific signaling
cascades within the chondrocyte and osteoblast microenvironment
require further elucidation. Emerging research has begun to map
these interactions, such as the regulation of bone extracellular
matrix homeostasis by gut-derived metabolites and the modulation of
the Wnt/β-catenin signaling pathway in joint remodeling (30,31). However, the interplay is often
non-linear and involves complex feedback loops including the
'gut-microbiota-ferroptosis axis', which has been recently proposed
as a critical pathogenic pathway (17). Future research must focus on
validating specific receptor targets on synovial cells for these
metabolites, moving from general observations of 'metabolic shifts'
to identifying druggable molecular targets that can halt cartilage
senescence and subchondral bone loss. A specific knowledge gap
regarding SCFAs concerns the absence of direct quantification in
human OA synovial fluid and the unknown relationship between oral
supplementation doses and clinically relevant concentration ranges
within the joint microenvironment. While serum SCFA levels have
been associated with OA severity, direct measurements in synovial
fluid remain scarce and existing metabolomic profiling of OA
synovial tissue has focused on other metabolite classes rather than
systematically quantifying SCFAs (67). Future studies should prioritize
paired serum-synovial fluid SCFA quantification in patients
receiving SCFA-based interventions to determine whether the
butyrate doses shown to improve clinical outcomes (300-600 mg/day)
achieve sufficient synovial fluid concentrations to directly
modulate chondrocyte and synovial cell functions.
The heterogeneity of OA phenotypes necessitates a
shift from a 'one-size-fits-all' probiotic strategy toward
personalized precision medicine based on distinct metabolomic
signatures. Current clinical applications are hindered by the lack
of reliable biomarkers to predict which patients will respond to
microbiome modulation. Advanced analytical techniques are beginning
to bridge this gap, with machine learning algorithms now being
employed to associate specific gut microbiota compositions with
physical functioning in patients with OA (104). Furthermore, distinct metabolic
profiles have been identified for specific disease subtypes, such
as erosive hand OA (110),
suggesting that the gut-joint axis may operate differently
depending on the affected joint and systemic metabolic status.
Future directions should prioritize the development of 'companion
diagnostics' that utilize fecal or serum metabolomics to phenotype
patients, thereby enabling clinicians to prescribe targeted
prebiotic or postbiotic regimens tailored to the individual's
specific dysbiotic profile.
Finally, the clinical translation of
microbiome-based therapies faces significant hurdles regarding
standardization, delivery and safety. While probiotic supplements
are popular, their colonization efficiency is transient and highly
variable among individuals. The field is progressively pivoting
toward 'postbiotics' or metabolite-based therapies, which offer
more predictable pharmacokinetics. For instance, the use of
sustained-release butyrate tablets has shown promise in suppressing
immune cell activation in patients with OA (51), providing a proof-of-concept for
direct metabolite supplementation. However, rigorous Mendelian
randomization studies are needed to confirm the causal impact of
the microbiota on arthritis outcomes to prevent premature clinical
application (111). The
ultimate goal for sports medicine and rheumatology is to develop
standardized, evidence-based protocols that integrate exercise,
which independently modulates the microbiome (21), with targeted metabolic
interventions to restore joint homeostasis through the gut-joint
axis.
Conclusion and future perspectives
The 'gut-joint axis' redefines OA from simple
mechanical wear to a systemic metabolic pathology driven by
intestinal dysbiosis. This review highlights how microbial
metabolites, including protective SCFAs, destructive TMAO and
complex BAs, act as critical molecular switches regulating joint
homeostasis. Future clinical innovation lies in precision sports
medicine where integrating exercise, nutrition and targeted
microbiome therapies will transition OA management from symptomatic
relief to proactive and mechanism-driven disease modification.
Availability of data and materials
Not applicable.
Authors' contributions
RZ, LZ, YW, XK and JZ made substantial
contributions to the conception and design of the article. RZ and
JZ performed acquisition, analysis and interpretation of data. RZ,
LZ, YW and XK drafted and wrote the manuscript. Data authentication
is not applicable. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
OA
|
osteoarthritis
|
|
SCFAs
|
short-chain fatty acids
|
|
TMAO
|
trimethylamine N-oxide
|
|
BAs
|
bile acids
|
|
FXR
|
farnesoid X receptor
|
|
TGR5
|
Takeda G-protein-coupled receptor
5
|
|
PXR
|
pregnane X receptor
|
|
TLR4
|
Toll-like receptor 4
|
|
NF-κB
|
nuclear factor κB
|
|
NLRP3
|
NLR family pyrin domain containing
3
|
|
HDAC
|
histone deacetylase
|
|
ROS
|
reactive oxygen species
|
|
LPS
|
lipopolysaccharides
|
|
BEVs
|
bacterial extracellular vesicles
|
|
ADAMTS
|
A disintegrin and metalloproteinase
with thrombospondin motifs
|
|
DCA
|
deoxycholic acid
|
|
LCA
|
lithocholic acid
|
|
Sirt6
|
sirtuin 6
|
|
MICT
|
moderate-intensity continuous
training
|
Acknowledgements
Not applicable.
Funding
No funding was received.
References
|
1
|
Weng Q, Chen Q, Jiang T, Zhang Y, Zhang W,
Doherty M, Xie J, Liu K, Li J, Yang T, et al: Global burden of
early-onset osteoarthritis, 1990-2019: Results from the global
burden of disease study 2019. Ann Rheum Dis. 83:915–925. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kloppenburg M, Namane M and Cicuttini F:
Osteoarthritis. Lancet. 405:71–85. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Batushansky A, Zhu S, Komaravolu RK, South
S, Mehta-D'souza P and Griffin TM: Fundamentals of OA. An
initiative of osteoarthritis and cartilage. Obesity and metabolic
factors in OA. Osteoarthritis Cartilage. 30:501–515. 2022.
View Article : Google Scholar
|
|
4
|
Motta F, Barone E, Sica A and Selmi C:
Inflammaging and osteoarthritis. Clin Rev Allergy Immunol.
64:222–238. 2023. View Article : Google Scholar
|
|
5
|
Gaspar MG, Núñez-Carro C, Blanco-Blanco M,
Blanco FJ and de Andrés MC: Inflammaging contributes to
osteoarthritis development and human microbiota variations and vice
versa: A systematic review. Osteoarthritis Cartilage. 33:218–230.
2025. View Article : Google Scholar
|
|
6
|
Bhardwaj A, Sapra L, Tiwari A, Mishra PK,
Sharma S and Srivastava RK: 'Osteomicrobiology': The nexus between
bone and bugs. Front Microbiol. 12:8124662022. View Article : Google Scholar
|
|
7
|
Escalante J, Artaiz O, Diwakarla S and
McQuade RM: Leaky gut in systemic inflammation: Exploring the link
between gastrointestinal disorders and age-related diseases.
Geroscience. 47:1–22. 2025. View Article : Google Scholar :
|
|
8
|
Bonato A, Zenobi-Wong M, Barreto G and
Huang Z: A systematic review of microbiome composition in
osteoarthritis subjects. Osteoarthritis Cartilage. 30:786–801.
2022. View Article : Google Scholar
|
|
9
|
Gilat R, Yazdi AA, Weissman AC, Joyce KM,
Bouftas FA, Muth SA, Chisari E, Shohat N and Cole BJ: The gut
microbiome and joint microbiome show alterations in patients with
knee osteoarthritis versus controls: A systematic review.
Arthroscopy. 41:1226–1238. 2025. View Article : Google Scholar
|
|
10
|
Li J, Liang J, Liu Y and Sun W and Sun W:
Basal metabolic rate mediates the causal relationship between gut
microbiota and osteoarthritis: A two-step bidirectional Mendelian
randomization study. Front Microbiol. 15:13716792024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang J and Qiu X: A Mendelian
randomization study of the gut microbiota and risk of knee
osteoarthritis and the mediating role of immune cells. Sci Rep.
15:284552025. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu L, Tian F, Li GY, Xu W and Xia R: The
effects and significance of gut microbiota and its metabolites on
the regulation of osteoarthritis: Close coordination of gut-bone
axis. Front Nutr. 9:10120872022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Femia M, Valenti Pittino C, Fumarola EM,
Tramarin M, Papa M, Giurazza F, Francioso AP, Fior D, Moramarco LP,
Antonini G, et al: Genicular artery embolization: A new tool for
the management of refractory osteoarthritis-related knee pain. J
Pers Med. 14:6862024. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang W, Liu X, Nan H, Li H and Yan L:
Specific gut microbiota and serum metabolite changes in patients
with osteoarthritis. Front Cell Dev Biol. 13:15435102025.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang X, Liu Y, Sun Z, Li J, Lu Z, Huang J,
Hu S, Cao P, Cao X, Li S, et al: Multi-omics reveal the
dysregulated gut-joint axis in knee synovitis: Data from two
osteoarthritis studies in China. Adv Sci (Weinh). 13:e120202026.
View Article : Google Scholar :
|
|
16
|
Han J, Meng X, Kong H, Li X, Chen P and
Zhang XA: Links between short-chain fatty acids and osteoarthritis
from pathology to clinic via gut-joint axis. Stem Cell Res Ther.
16:2512025. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang T, Xu Y, Zha H and Wu L: The 'gut
microbiota-ferroptosis axis': A hypothesis for osteoarthritis
pathogenesis and therapeutic implications. Front Microbiol.
16:16859622025. View Article : Google Scholar
|
|
18
|
Li J, Ye J, Yang T, Hunter DJ, Zhang W,
Doherty M, Zhang Y, Yang Z, Li H, Wang Y, et al: Bile acids
metabolism in symptomatic hand osteoarthritis. Arthritis Rheumatol.
Dec 26–2025.Epub ahead of print. PubMed/NCBI
|
|
19
|
Yang Y, Hao C, Jiao T, Yang Z, Li H, Zhang
Y, Zhang W, Doherty M, Sun C, Yang T, et al: Osteoarthritis
treatment via the GLP-1-mediated gut-joint axis targets intestinal
FXR signaling. Science. 388:eadt05482025. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hao X, Shang X, Zhang Y, Hou W, Chi R, Pan
C, Liu J, Deng X, Zhang J and Xu T: Effects of exercise on gut
microbiome and serum metabolomics in post-traumatic osteoarthritis
rats. Metabolites. 15:3412025. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xi Y, Wang Z, Wei Y, Xiao N, Duan L, Zhao
T, Zhang X, Zhang L, Wang J, Li Z and Qin D: Gut microbiota and
osteoarthritis: From pathogenesis to novel therapeutic
opportunities. Am J Chin Med. 53:43–66. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bridges SL Jr, Sun D, Graham ZA, McAdam
JS, Mayo ED and Bamman MM: Muscle inflammation susceptibility: A
potential phenotype for guiding precision rehabilitation after
total hip arthroplasty in end-stage osteoarthritis. HSS J.
19:453–458. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Zeng T, Tang D, Cui H, Wan Y and
Tang H: Integrated multi-omics analyses reveal lipid metabolic
signature in osteoarthritis. J Mol Biol. 437:1688882025. View Article : Google Scholar
|
|
24
|
Niu L, Chen W, Yin Z, Tan H, Cui J and Su
J: Bacterial extracellular vesicles in osteoarthritis: A new bridge
of the gut-joint axis. Gut Microbes. 17:24890692025. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang F, Li D, Long W, Li E and Wei B: Role
of pyroptosis in the pathogenesis of osteoarthritis: An updated
review. J Inflamm Res. 18:15065–15079. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guido G, Ausenda G, Iascone V and Chisari
E: Gut permeability and osteoarthritis, towards a mechanistic
understanding of the pathogenesis: A systematic review. Ann Med.
53:2380–2390. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang Q, Li Z, Dang D, Wei J and Wu H:
Role of mechanosensitive channel Piezo1 protein in intestinal
inflammation regulation: A potential target. FASEB J.
38:e701222024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bardi E, D'Arrigo D, Pozzi C, Gatti A,
Bertolino L, Favaro A, Rescigno M and Bonanzinga T: Current methods
in synovial fluid microbiota characterization: A systematic review.
Int J Mol Sci. 26:46902025. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He M, Kolhoff F, Mont MA and Parvizi J: Is
osteoarthritis a state of joint dysbiosis? Antibiotics (Basel).
14:6092025. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Liu K, Li Y, Tang Z, Li Y, Li A and
Zhang H: The gut microbiota-derived metabolites regulate bone
extracellular matrix homeostasis: Mechanisms and therapeutic
implications. Cell Signal. 138:1122452026. View Article : Google Scholar
|
|
31
|
Zhang R, Zhang L, Tian B, Wang Y, Kang X
and Zheng J: The gut-bone-cartilage triad: Microbial regulation of
the Wnt/β-catenin signaling pathway in osteoarthritis joint
remodeling (review). Mol Med Rep. 33:232026. View Article : Google Scholar
|
|
32
|
Tominaga K, Tsuchiya A, Mizusawa T,
Matsumoto A, Minemura A, Oka K, Takahashi M, Yosida T, Kawata Y,
Takahashi K, et al: Evaluation of intestinal microbiota,
short-chain fatty acids, and immunoglobulin a in diversion colitis.
Biochem Biophys Rep. 25:1008922020.
|
|
33
|
Kumar A, Toghyani M, Kheravii SK, Pineda
L, Han Y, Swick RA and Wu SB: Organic acid blends improve
intestinal integrity, modulate short-chain fatty acids profiles and
alter microbiota of broilers under necrotic enteritis challenge.
Anim Nutr. 8:82–90. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pirozzi C, Francisco V, Guida FD, Gómez R,
Lago F, Pino J, Meli R and Gualillo O: Butyrate modulates
inflammation in chondrocytes via GPR43 receptor. Cell Physiol
Biochem. 51:228–243. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou H, Li G, Wang Y, Jiang R, Li Y, Wang
H, Wang F, Ma H and Cao L: Microbial metabolite sodium butyrate
attenuates cartilage degradation by restoring impaired autophagy
and autophagic flux in osteoarthritis development. Front Pharmacol.
12:6595972021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Balderas C, de Ancos B and Sánchez-Moreno
C: Bile acids and short-chain fatty acids are modulated after onion
and apple consumption in obese zucker rats. Nutrients. 15:30352023.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kirschner SK, Engelen MP, Haas P, Bischoff
SC and Deutz NE: Short-chain fatty acid kinetics and concentrations
are higher after inulin supplementation in young and older adults:
A randomized trial. Am J Clin Nutr. 121:1224–1235. 2025. View Article : Google Scholar
|
|
38
|
Li M, van Esch BCAM, Henricks PAJ,
Folkerts G and Garssen J: The anti-inflammatory effects of short
chain fatty acids on lipopolysaccharide- or tumor necrosis factor
α-stimulated endothelial cells via activation of GPR41/43 and
inhibition of HDACs. Front Pharmacol. 9:5332018. View Article : Google Scholar
|
|
39
|
Sun X, Luo S, Jiang C, Tang Y, Cao Z, Jia
H, Xu Q, Zhao C, Loor JJ and Xu C: Sodium butyrate reduces bovine
mammary epithelial cell inflammatory responses induced by exogenous
lipopolysaccharide, by inactivating NF-κB signaling. J Dairy Sci.
103:8388–8397. 2020. View Article : Google Scholar
|
|
40
|
Sun B, Jia Y, Hong J, Sun Q, Gao S, Hu Y,
Zhao N and Zhao R: Sodium butyrate ameliorates
high-fat-diet-induced non-alcoholic fatty liver disease through
peroxisome proliferator-activated receptor α-mediated activation of
β oxidation and suppression of inflammation. J Agric Food Chem.
66:7633–7642. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ziabska K, Gargas J, Sypecka J and
Ziemka-Nalecz M: The impact of the histone deacetylase inhibitor
sodium butyrate on microglial polarization after oxygen and glucose
deprivation. Pharmacol Rep. 74:909–919. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tang X, Ma S, Li Y, Sun Y, Zhang K, Zhou Q
and Yu R: Evaluating the activity of sodium butyrate to prevent
osteoporosis in rats by promoting osteal GSK-3β/Nrf2 signaling and
mitochondrial function. J Agric Food Chem. 68:6588–6603. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Duscha A, Hegelmaier T, Dürholz K, Desel
C, Gold R, Zaiss MM and Haghikia A: Propionic acid beneficially
modifies osteoporosis biomarkers in patients with multiple
sclerosis. Ther Adv Neurol Disord. 15:175628642211039352022.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bo W, Zhou J and Wang K: Sodium butyrate
abolishes the degradation of type II collagen in human
chondrocytes. Biomed Pharmacother. 102:1099–1104. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cho KH, Na HS, Jhun J, Woo JS, Lee AR, Lee
SY, Lee JS, Um IG, Kim SJ, Park SH and Cho ML: Lactobacillus (LA-1)
and butyrate inhibit osteoarthritis by controlling autophagy and
inflammatory cell death of chondrocytes. Front Immunol.
13:9305112022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Huang C, Du W, Ni Y, Lan G and Shi G: The
effect of short-chain fatty acids on M2 macrophages polarization in
vitro and in vivo. Clin Exp Immunol. 207:53–64. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu M, Xie WJ, Zhang X, Wu W, Li G and
Wang L: Sodium butyrate regulates macrophage polarization by
TGR5/β-arrestin2 in vitro. Mol Med. 31:312025. View Article : Google Scholar
|
|
48
|
Wu YL, Zhang CH, Teng Y, Pan Y, Liu NC,
Liu PX, Zhu X, Su XL and Lin J: Propionate and butyrate attenuate
macrophage pyroptosis and osteoclastogenesis induced by CoCrMo
alloy particles. Mil Med Res. 9:462022.PubMed/NCBI
|
|
49
|
Theiler A, Bärnthaler T, Platzer W,
Richtig G, Peinhaupt M, Rittchen S, Kargl J, Ulven T, Marsh LM,
Marsche G, et al: Butyrate ameliorates allergic airway inflammation
by limiting eosinophil trafficking and survival. J Allergy Clin
Immunol. 144:764–776. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li G, Lin J, Zhang C, Gao H, Lu H, Gao X,
Zhu R, Li Z, Li M and Liu Z: Microbiota metabolite butyrate
constrains neutrophil functions and ameliorates mucosal
inflammation in inflammatory bowel disease. Gut Microbes.
13:19682572021. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Korsten SGPJ, Hartog M, Berends AJ,
Koenders MI, Popa CD, Vromans H, Garssen J, van de Ende CHM,
Vermeiden JPW and Willemsen LEM: A sustained-release butyrate
tablet suppresses ex vivo T helper cell activation of
osteoarthritis patients in a double-blind placebo-controlled
randomized trial. Nutrients. 16:33842024. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Alisson-Silva F, Kawanishi K and Varki A:
Human risk of diseases associated with red meat intake: Analysis of
current theories and proposed role for metabolic incorporation of a
non-human sialic acid. Mol Aspects Med. 51:16–30. 2016. View Article : Google Scholar
|
|
53
|
Thomas MS and Fernandez ML: Trimethylamine
N-oxide (TMAO), diet and cardiovascular disease. Curr Atheroscler
Rep. 23:122021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sawicka AK, Renzi G and Olek RA: The
bright and the dark sides of L-carnitine supplementation: A
systematic review. J Int Soc Sports Nutr. 17:492020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lau WL, Savoj J, Nakata MB and Vaziri ND:
Altered microbiome in chronic kidney disease: Systemic effects of
gut-derived uremic toxins. Clin Sci (Lond). 132:509–522. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lei Y, Xiong G, Lei D, Wu H, Peng X, Chen
L, Fang Q, Wu Y, Wu Y, Li X and Li Y: Gut microbiota-derived TMAO
drives the kidney-bone-vascular axis in chronic kidney disease
complications. Ren Fail. 47:25754342025. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li L, Chen B, Zhu R, Li R, Tian Y, Liu C,
Jia Q, Wang L, Tang J, Zhao D, et al: Fructus Ligustri Lucidi
preserves bone quality through the regulation of gut microbiota
diversity, oxidative stress, TMAO and Sirt6 levels in aging mice.
Aging (Albany NY). 11:9348–9368. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zou H, Huang C, Zhou L, Lu R, Zhang Y and
Lin D: NMR-based metabolomic analysis for the effects of
trimethylamine N-oxide treatment on C2C12 myoblasts under oxidative
stress. Biomolecules. 12:12882022. View Article : Google Scholar :
|
|
59
|
Zhang X, Li Y, Yang P, Liu X, Lu L, Chen
Y, Zhong X, Li Z, Liu H, Ou C, et al: Trimethylamine-N-oxide
promotes vascular calcification through activation of NLRP3
(nucleotide-binding domain, leucine-rich-containing family, pyrin
domain-containing-3) inflammasome and NF-κB (nuclear factor κB)
signals. Arterioscler Thromb Vasc Biol. 40:751–765. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang N, Hao Y and Fu L:
Trimethylamine-N-oxide promotes osteoclast differentiation and bone
loss via activating ROS-dependent NF-κB signaling pathway.
Nutrients. 14:39552022. View Article : Google Scholar
|
|
61
|
Zhao Y, Wang C, Qiu F, Liu J, Xie Y, Lin
Z, He J and Chen J: Trimethylamine-N-oxide promotes osteoclast
differentiation and oxidative stress by activating NF-κB pathway.
Aging (Albany NY). 16:9251–9263. 2024. View Article : Google Scholar :
|
|
62
|
Zhuang H, Ren X, Zhang Y, Jiang F and Zhou
P: Trimethylamine-N-oxide sensitizes chondrocytes to mechanical
loading through the upregulation of Piezo1. Food Chem Toxicol.
175:1137262023. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Elam RE, Bůžková P, Barzilay JI, Wang Z,
Nemet I, Budoff MJ, Cauley JA, Fink HA, Lee Y, Robbins JA, et al:
Trimethylamine N-oxide and hip fracture and bone mineral density in
older adults: The cardiovascular health study. Bone.
161:1164312022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lin H, Liu T, Li X, Gao X, Wu T and Li P:
The role of gut microbiota metabolite trimethylamine N-oxide in
functional impairment of bone marrow mesenchymal stem cells in
osteoporosis disease. Ann Transl Med. 8:10092020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mo X, Cheng R, Shen L, Sun Y, Wang P,
Jiang G, Wen L, Li X, Peng X, Liao Y, et al: High-fat diet induces
sarcopenic obesity in natural aging rats through the
gut-trimethylamine N-oxide-muscle axis. J Adv Res. 70:405–422.
2025. View Article : Google Scholar :
|
|
66
|
Lin YL, Liu CH, Lai YH, Wang CH, Kuo CH,
Liou HH and Hsu BG: Association of serum indoxyl sulfate levels
with skeletal muscle mass and strength in chronic hemodialysis
patients: A 2-year longitudinal analysis. Calcif Tissue Int.
107:257–265. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Murillo-Saich JD, Coras R, Meyer R,
Llorente C, Lane NE and Guma M: Synovial tissue metabolomic
profiling reveal biomarkers of synovial inflammation in patients
with osteoarthritis. Osteoarthr Cartil Open. 4:1002952022.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Silvestre MP, Rodrigues AM, Canhão H,
Marques C, Teixeira D, Calhau C and Branco J: Cross-talk between
diet-associated dysbiosis and hand osteoarthritis. Nutrients.
12:34692020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Huang L, Huang L, Wang L, Wang Y, Xie W,
Zhou Y, Mei Y, Yang J, Yan F and Xie S: Nonsurgical periodontal
treatment improved the abnormal trimethylamine n-oxide metabolism
in Apoe-/- mice with periodontitis. Biochim Biophys Acta
Mol Basis Dis. 1871:1677522025. View Article : Google Scholar
|
|
70
|
Chen J, Malhi KK, Li X, Xu X, Kang J, Zhao
B, Xu Y, Li X and Li J: Metasilicate-based alkaline mineral water
improves the growth performance of weaned piglets by maintaining
gut-liver axis homeostasis through microbiota-mediated secondary
bile acid pathway. Anim Nutr. 20:95–109. 2024. View Article : Google Scholar
|
|
71
|
Yu J, Huang S, Zhang J, Su Z, Chang Y, Liu
K, Yang L, Guo H, Ye J, Liang X, et al: Bile acids insufficiency
links perfluorooctane sulfonate-induced oxidative stress-mediated
fatty liver with osteoarthritis. J Hazard Mater. 501:1407502026.
View Article : Google Scholar
|
|
72
|
Hu W, Cai C, Li Y, Kang F, Chu T and Dong
S: Farnesoid X receptor agonist attenuates subchondral bone
osteoclast fusion and osteochondral pathologies of osteoarthritis
via suppressing JNK1/2/NFATc1 pathway. FASEB J. 36:e222432022.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sun X, Chen Z, Yu L, Zeng W, Sun B, Fan H
and Bai Y: Bacteroides dorei BDX-01 alleviates DSS-induced
experimental colitis in mice by regulating intestinal bile salt
hydrolase activity and the FXR-NLRP3 signaling pathway. Front
Pharmacol. 14:12053232023. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Huang H, Lei H, Yang F, Fan X, Dang Q and
Li Y: Activation of the bile acid receptor GPBAR1 (TGR5)
ameliorates interleukin-1β (IL-1β)-induced chondrocytes senescence.
Biomed Pharmacother. 106:1713–1719. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhuo W, Li B and Zhang D: Activation of
G-protein-coupled bile acid receptor Gpbar1 (TGR5) inhibits
degradation of type II collagen and aggrecan in human chondrocytes.
Eur J Pharmacol. 856:1723872019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Mai M, Wang Y, Luo M, Li Z, Wang D, Ruan Y
and Guo H: Silibinin ameliorates deoxycholic acid-induced
pyroptosis in steatotic HepG2 cells by inhibiting NLRP3
inflammasome activation. Biochem Biophys Rep.
35:1015452023.PubMed/NCBI
|
|
77
|
Holtmann TM, Inzaugarat ME, Knorr J,
Geisler L, Schulz M, Bieghs V, Frissen M, Feldstein AE, Tacke F,
Trautwein C and Wree A: Bile acids activate NLRP3 inflammasome,
promoting murine liver inflammation or fibrosis in a cell
type-specific manner. Cells. 10:26182021. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhong K, Yao Y, Shi B, Li S, Zhao Y, Wang
X, Zhou C, Wang B, Shao X and Zhu X: EGCG alleviates colitis in
inflammatory bowel disease via gut microbiota-derived LCA and
inhibition of NLRP3 inflammasome. Int Immunopharmacol.
168:1159622026. View Article : Google Scholar
|
|
79
|
Liu Y, Gao J, Chen L, Chen Y, Jiang J,
Chen H and Ma L: Lithocholic acid ameliorates ulcerative colitis
via the PXR/TLR4/NF-κB/NLRP3 signaling pathway and gut microbiota
modulation. Cell Mol Life Sci. 82:3362025. View Article : Google Scholar
|
|
80
|
Zemedikun DT, Lee H, Nirantharakumar K,
Raza K, Chandan JS, Lord JM and Jackson TA: Comorbidity phenotypes
and risk of mortality in patients with osteoarthritis in the UK: A
latent class analysis. Arthritis Res Ther. 24:2312022. View Article : Google Scholar :
|
|
81
|
Sharafi M, Baeradeh N, Mohsenpour MA,
Afrashteh S, Bagheri P, Keshavarzian O, Annabi MA, Nekouei F and
Farjam M: Investigating multimorbidity patterns and associated risk
factors in the fasa adults cohort study (FACS): A latent class
analysis. PLoS One. 20:e03351772025. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Fortuna R, Wang W, Mayengbam S, Tuplin
EWN, Sampsell K, Sharkey KA, Hart DA and Reimer RA: Effect of
prebiotic fiber on physical function and gut microbiota in adults,
mostly women, with knee osteoarthritis and obesity: A randomized
controlled trial. Eur J Nutr. 63:2149–2161. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Li K, Liu A, Zong W, Dai L, Liu Y, Luo R,
Ge S and Dong G: Moderate exercise ameliorates osteoarthritis by
reducing lipopolysaccharides from gut microbiota in mice. Saudi J
Biol Sci. 28:40–49. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hao X, Zhang J, Shang X, Sun K, Zhou J,
Liu J, Chi R and Xu T: Exercise modifies the disease-relevant gut
microbial shifts in post-traumatic osteoarthritis rats. Bone Joint
Res. 11:214–225. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Brunt VE, Greenberg NT, Sapinsley ZJ,
Casso AG, Richey JJ, VanDongen NS, Gioscia-Ryan RA, Ziemba BP,
Neilson AP, Davy KP and Seals DR: Suppression of trimethylamine
N-oxide with DMB mitigates vascular dysfunction, exercise
intolerance, and frailty associated with a western-style diet in
mice. J Appl Physiol (1985). 133:798–813. 2022. View Article : Google Scholar
|
|
86
|
Pedersini P, Turroni S and Villafañe JH:
Gut microbiota and physical activity: Is there an evidence-based
link? Sci Total Environ. 727:1386482020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Lei M, Guo C, Wang D, Zhang C and Hua L:
The effect of probiotic Lactobacillus casei Shirota on knee
osteoarthritis: A randomised double-blind, placebo-controlled
clinical trial. Benef Microbes. 8:697–703. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Lee SH, Kwon JY, Jhun J, Jung K, Park SH,
Yang CW, Cho Y, Kim SJ and Cho ML: Lactobacillus acidophilus
ameliorates pain and cartilage degradation in experimental
osteoarthritis. Immunol Lett. 203:6–14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Jhun J, Cho KH, Lee DH, Kwon JY, Woo JS,
Kim J, Na HS, Park SH, Kim SJ and Cho ML: Oral Administration of
Lactobacillus rhamnosus ameliorates the progression of
osteoarthritis by inhibiting joint pain and inflammation. Cells.
10:10572021. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Fernández-Murga ML, Olivares M and Sanz Y:
Bifidobacterium pseudocatenulatum CECT 7765 reverses the adverse
effects of diet-induced obesity through the gut-bone axis. Bone.
141:1155802020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Henrotin Y, Patrier S, Pralus A, Roche M
and Nivoliez A: Protective actions of oral administration of
Bifidobacterium longum CBi0703 in spontaneous osteoarthritis in
dunkin hartley guinea pig model. Cartilage. 13(Suppl 2):
1204S–1213S. 2021. View Article : Google Scholar
|
|
92
|
Sapra L, Shokeen N, Porwal K, Saini C,
Bhardwaj A, Mathew M, Mishra PK, Chattopadhyay N, Dar HY, Verma B
and Srivastava RK: Bifidobacterium longum ameliorates
ovariectomy-induced bone loss via enhancing anti-osteoclastogenic
and immunomodulatory potential of regulatory B cells (Bregs). Front
Immunol. 13:8757882022. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Drabińska N, Jarocka-Cyrta E, Złotkowska
D, Abramowicz P and Krupa-Kozak U: Daily oligofructose-enriched
inulin intake impacts bone turnover markers but not the cytokine
profile in pediatric patients with celiac disease on a gluten-free
diet: Results of a randomised, placebo-controlled pilot study.
Bone. 122:184–192. 2019. View Article : Google Scholar
|
|
94
|
Seijo M, Bryk G, Zeni Coronel M, Bonanno
M, Río ME, Pita Martín de Portela ML and Zeni SN: Effect of adding
a galacto-oligosaccharides/fructo-oligosaccharides (GOS/FOS®)
mixture to a normal and low calcium diet, on calcium absorption and
bone health in ovariectomy-induced osteopenic rats. Calcif Tissue
Int. 104:301–312. 2019. View Article : Google Scholar
|
|
95
|
Turbić A, Vandenput L, Gandham A and
Lorentzon M: Effects of synbiotic supplementation on bone and
metabolic health in caucasian postmenopausal women: Rationale and
design of the OsteoPreP trial. Nutrients. 16:42192024. View Article : Google Scholar
|
|
96
|
Ishizu T, Takai E, Torii S and Taguchi M:
Prebiotic food intake may improve bone resorption in Japanese
female athletes: A pilot study. Sports (Basel). 9:822021.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Hsu CN, Chang-Chien GP, Lin S, Hou CY and
Tain YL: Targeting on gut microbial metabolite
trimethylamine-N-oxide and short-chain fatty acid to prevent
maternal high-fructose-diet-induced developmental programming of
hypertension in adult male offspring. Mol Nutr Food Res.
63:e19000732019. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Wang G, Kong B, Shuai W, Fu H, Jiang X and
Huang H: 3,3-Dimethyl-1-butanol attenuates cardiac remodeling in
pressure-overload-induced heart failure mice. J Nutr Biochem.
78:1083412020. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Fechtner S, Allen BE, Chriswell ME, Jubair
WK, Robertson CE, Kofonow JN, Frank DN, Holers VM and Kuhn KA:
3,3-Dimethyl-1-butanol and its metabolite 3,3-dimethylbutyrate
ameliorate collagen-induced arthritis independent of choline
trimethylamine lyase activity. Res Sq [Preprint].
rs.3.rs-32970182023.PubMed/NCBI
|
|
100
|
Fechtner S, Allen BE, Chriswell ME, Jubair
WK, Robertson CE, Kofonow JN, Frank DN, Holers VM and Kuhn KA:
3,3-Dimethyl-1-butanol and its metabolite 3,3-dimethylbutyrate
ameliorate collagen-induced arthritis independent of choline
trimethylamine lyase activity. Inflammation. 48:1350–1365. 2025.
View Article : Google Scholar
|
|
101
|
Baptista LC, Wilson L, Barnes S, Anton SD
and Buford TW: Effects of resveratrol on changes in
trimethylamine-N-oxide and circulating cardiovascular factors
following exercise training among older adults. Exp Gerontol.
194:1124792024. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Huang PH, Chen DQ, Chen YW, Shih MK, Lee
BH, Tain YL, Hsieh CW and Hou CY: Evaluation of the feasibility of
in vitro metabolic interruption of trimethylamine with resveratrol
butyrate esters and its purified monomers. Molecules. 29:4292024.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Roberts JL, Liu G, Darby TM, Fernandes LM,
Diaz-Hernandez ME, Jones RM and Drissi H: Bifidobacterium
adolescentis supplementation attenuates fracture-induced systemic
sequelae. Biomed Pharmacother. 132:1108312020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
de Sire A, Mancuso E, Marotta N, Massimino
M, Zito R, Averta C, Bartalotta I, Palummo A, Cerantonio A,
Citrigno L, et al: Association between gut microbiota composition
and physical functioning in patients with knee osteoarthritis: A
machine learning study. Sci Rep. 15:408262025. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Prinz E, Schlupp L, Dyson G, Barrett M,
Szymczak A, Velasco C, Izda V, Dunn CM and Jeffries MA: OA
susceptibility in mice is partially mediated by the gut microbiome,
is transferrable via microbiome transplantation and is associated
with immunophenotype changes. Ann Rheum Dis. 83:382–393. 2024.
View Article : Google Scholar
|
|
106
|
Huang J, Liu M, Furey A, Rahman P and Zhai
G: Gut microbiomics of sustained knee pain in patients with knee
osteoarthritis. J Rheumatol. 51:1218–1225. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Griffin TM, Lopes EBP, Cortassa D,
Batushansky A, Jeffries MA, Makosa D, Jopkiewicz A, Mehta-D'souza
P, Komaravolu RK and Kinter MT: Sexually dimorphic metabolic
effects of a high fat diet on knee osteoarthritis in mice. Biol Sex
Differ. 15:1032024. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Cheng M, Lu H, Wu Y, Jia L, Xiang T, Deng
LI, Zhao G and Feng J: Sex differences in alcohol inhibits bone
formation and promotes bone resorption in young male and female
rats by altering intestinal flora, metabolites, and bone
microenvironment. PLoS One. 20:e03232222025. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Kverka M and Stepan JJ: Associations among
estrogens, the gut microbiome and osteoporosis. Curr Osteoporos
Rep. 23:22024. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Xie D, Wang Y, Li J, Yang T, Zhang Y,
Zhang W, Doherty M, Zhu Y, Yang Z, Wang Y, et al: Correlation
between gut dysbiosis, metabolite alterations and erosive hand
osteoarthritis-An observational study within the community-based
Xiangya Osteoarthritis (XO) cohort. Osteoarthritis Cartilage.
33:1246–1257. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Pan B, Guo Q, Cai J, Chen L, Zhao Z, Shen
P and Wang Y: Investigating the causal impact of gut microbiota on
arthritis via inflammatory proteins using mendelian randomization.
Sci Rep. 14:274332024. View Article : Google Scholar : PubMed/NCBI
|