Introduction
Molecular structure of MTA2 protein
Composition of the MTA family
Metastasis and the formation of secondary malignant tumors are the primary causes of mortality in cancer patients (1-3). Recent studies have increasingly focused on the metastasis-associated (MTA) family, a chromatin-modifying group crucial for cancer invasion and metastasis regulation (1,4). This family is closely associated with Mi-2/nucleosome remodeling and deacetylase (NuRD) complexes (5). MTA1 was first identified by Xue et al (6) in 1998 as a component of the NuRD complex. Subsequently, Zhang et al (7) purified the NuRD complex and identified MTA2 instead of MTA1. Typically, distinct MTA family members are found in separate NuRD complexes and do not appear together within the same complex (8). In vertebrates, the MTA family primarily includes three highly homologous proteins, MTA1, MTA2 and MTA3, with molecular weights of ~80, 70 and 65 kDa, respectively (9). In humans, the genes encoding these proteins have well-defined chromosomal localizations: MTA1 at 14q32.3, MTA2 at 11q12-q13.1 and MTA3 at 2p21 (10). As an important member of this family, MTA2 comprises 668 amino acids and multiple functional domains and modification sites, reflecting modular structural features. It participates in transcriptional regulation and chromatin remodeling, bridges intranuclear activities with cytoskeletal dynamics and coordinates multiple signaling pathways in malignant progression (11,12). Notably, this structural uniqueness preliminarily implies MTA2 cannot be fully functionally substituted by MTA1 or MTA3 within NuRD assemblies, and the three homologs form mutually exclusive NuRD variants with divergent chromatin-targeting preference across disparate cancer types.
MTA1 is the founding homologue of the MTA family; subsequent studies indicated widespread overexpression of the MTA1 protein in most human tumors containing both solid and hematological neoplasms, in which it contributes to tumorigenesis and tumor progression (13). MTA3 (515 amino acids) and its longer isoform MTA3L (594 amino acids) are highly similar at their unique carboxyl termini (14). Alignment of the primary structure of MTA proteins shows that human MTA2 and MTA3 share 68.2 and 73.2% homology with human MTA1, respectively. The N-termini of MTA proteins are highly homologous, whereas the C-terminal regions are divergent (10). All three members of the MTA family are subunits of the NuRD complex. However, recent evidence suggests that they constitute distinct Mi-2/NuRD complexes and may exhibit specific transcriptional functions in a cell-type-specific manner (8). A detailed overview of the similarities and differences among the various MTA family members in terms of structure, expression patterns, molecular regulatory mechanisms and functions is provided in Table SI.
Although all members of the MTA family are part of the NuRD complex, each member possesses unique functions. MTA2 appears to regulate cytoskeletal remodeling by modulating genome-wide gene expression and activating the Rho signaling pathway (15,16). MTA3 also plays a role in cytoskeletal regulation by participating in the control of epithelial state through the regulation of Snail (14). Furthermore, the overall levels of MTA family members appear to be coordinately regulated at the protein level. Fujita et al (17) demonstrated that overexpression of MTA1 reduces the protein levels of MTA2 and MTA3 while increasing their mRNA expression levels. Similarly, Zhang et al (18) and Araghi et al (19) found that in breast tumors overexpressing the polyomavirus intermediate T antigen, MTA1 expression increases while MTA3 expression decreases as the tumor progresses. Thus, the overall levels of MTA may be limited by the availability of NuRD complex components capable of binding to them. This competitive binding for limited NuRD scaffold subunits is another core reason for their non-interchangeable properties: Elevated MTA1 competitively occupies the common NuRD binding pocket and displaces endogenous MTA2/MTA3 from pre-assembled complexes, which switches the global chromatin landscape from MTA2-dependent proliferation inhibition to MTA1-mediated invasive phenotype in progressive breast and liver cancers. Mutations in MTA2 are uncommon in human cancers. Furthermore, MTA2 is more frequently subject to gene amplification than deletion in cancers, suggesting that its role in cancer and metastasis is likely achieved through upregulation rather than mutational activation (20).
Collectively, multiple lines of evidence support MTA2 as the central functional element of the MTA family within the NuRD epigenomic network compared with MTA1 and MTA3: First, only MTA2 owns a complete set of four canonical conserved domains [bromo-adjacent homology (BAH)/Egl-27 and MTA1 homology 2 (ELM2)/SWI3, ADA2, N-CoR and TFIIIB (SANT)/atypical GATA zinc finger] required for both NuRD complex assembly and precise genomic targeting; partial domain truncation exists in C-terminal of MTA1 and MTA3, impairing their full-range chromatin recruitment capacity. Second, unlike MTA1 [mainly epithelial to mesenchymal transition (EMT) driver] and MTA3 (hormone-dependent tissue-specific regulator), MTA2 uniquely bridgesthe NuRD complex with multiple core oncogenic cascades including Rho, mTOR, HIF-1α and p53 signaling to coordinate epigenetic repression across proliferation, autophagy and DNA damage response simultaneously. Third, at the genetic level, MTA2 relies on gene amplification to gain oncogenic function in most cancers without frequent coding mutation, whereas MTA1/MTA3 are mainly regulated via transcriptional upregulation or isoform alternative splicing; its stable copy-number gain enables sustained central scaffolding function of NuRD during malignant evolution.
Structural domains of the MTA2 protein
MTA2 contains four highly conserved functional domains. The domains and motifs of the MTA2 protein are as follows (Fig. 1): i) The BAH domain, comprising amino acids 1-144, functions as a versatile module for chromatin-related activities. It mediates protein-protein interactions, recognizes specific histone-tail modifications and participates in nucleosome binding (21); ii) the ELM2 domain, spanning amino acids 145-256, is frequently present in transcriptional regulators and may contribute to protein dimerization and complex assembly (11); iii) the SANT domain (amino acids 263-315) serves as a fundamental unit for binding DNA and histone tails, supporting the activity of chromatin-remodeling enzymes (11); and iv) an atypical GATA zinc-finger domain (amino acids 367-394), which may be involved in specific DNA recognition or mediate interactions with specific proteins (21).
Post-translational modification sites
The biological functions of MTA2 are finely regulated by diverse post-translational modifications: i) Phosphorylation sites: Identified sites include S433 (22), S435 (22-24), T534 (24) and S548 (25). These sites may dynamically regulate MTA2 activity by sensing and integrating upstream signals, such as those activated by DNA-damage responses or growth factors. ii) Acetylation: Within the ELM2 domain, K152 carries an Nε-acetyllysine modification, suggesting dynamic and reversible regulation by acetyltransferases and deacetylases, which in turn affects the transcriptional co-repressive activity of MTA2 (26).
These modification sites collaborate with the functional domains of MTA2 to form a multilevel regulatory network that underlies its participation in nuclear processes such as chromatin remodeling and transcriptional regulation (26-29).
Mechanism of action of MTA2 protein
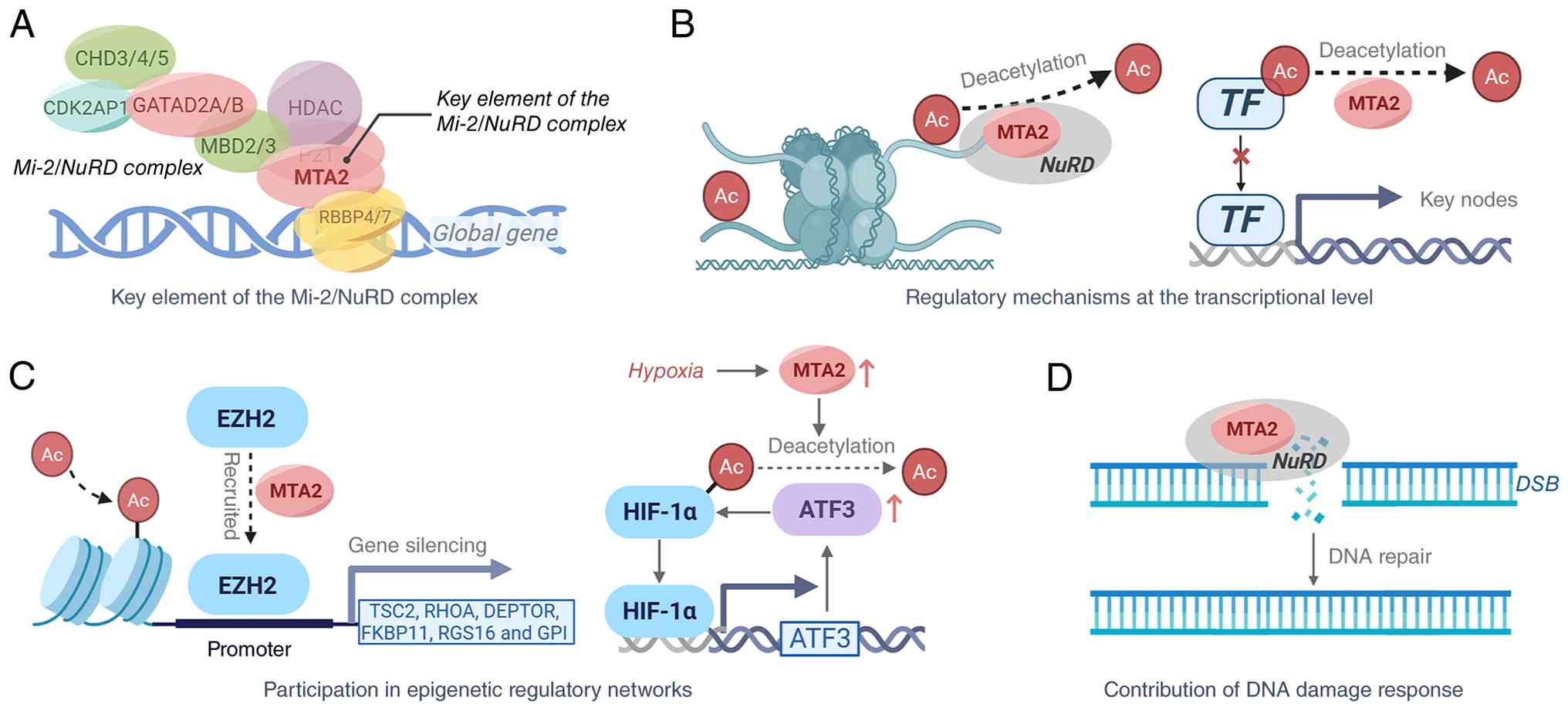
The mechanisms of action of the MTA2 protein are as follows (Fig. 2).
Key element of the Mi-2/NuRD complex
MTA family members are crucial to the Mi-2/NuRD complex (30,31), which regulates global gene expression. MTA2 is vital for maintaining the NuRD complex's structural integrity and directing histone deacetylase (HDAC) activity to specific substrates. Through its BAH, ELM2 and SANT domains, MTA2 targets the NuRD complex to defined genomic regions, mediating dynamic chromatin reorganization and transcriptional regulation (11). From the complex assembly perspective, MTA2 acts as the indispensable bridging hub for HDAC subunit and chromatin-binding module of NuRD; neither MTA1 nor MTA3 can fully restore intact HDAC enzymatic activity and genome-wide chromatin targeting efficiency when endogenous MTA2 is depleted, further verifying its irreplaceable central status in NuRD epigenetic regulation compared with other MTA paralogs.
Regulatory mechanisms at the transcriptional level
MTA2 regulates gene expression through two main mechanisms: i) Global histone deacetylation: MTA2 mediates chromatin remodeling and regulates transcriptional processes through the intranuclear NuRD complex. For example, MTA2/NuRD can specifically target the interleukin-4 (IL4) gene and repress its expression (32,33). ii) Regulation via acetylation of key nodes: MTA2 directly regulates the acetylation status of key signaling molecules [such as HIF-1α (34,35), estrogen receptor α (ERα) (28) and p53 (27)], affecting their stability and transcriptional activities. MTA2 suppresses p53 transcriptional activity by deacetylating it, thereby influencing cell-cycle arrest and apoptosis (27).
Participation in epigenetic regulatory networks
MTA2 recruits the methyltransferase enhancer of zeste homolog 2 (EZH2) to promoter regions of mechanistic target of rapamycin (mTOR) pathway repressors, including TSC complex subunit 2 (TSC2), Ras homolog family member A (RHOA), DEP domain-containing mTOR-interacting protein, FK506 binding protein 11, regulator of G protein signaling 16 and glucose-6-phosphate isomerase (1). This recruitment silences these genes, particularly TSC2, through histone methylation, thereby activating mTOR signaling and inhibiting autophagy (36).
Regulation of hypoxia-inducible factor (HIF)-1α stability: Under hypoxic conditions, transcription of the long non-coding RNA MTA2 transcriptional regulator is upregulated, increasing MTA2 expression. This stabilizes the HIF-1α protein via deacetylation and enhances its transcriptional activity. A positive feedback loop with activating transcription factor 3 promotes tumorigenesis and progression (37).
Contribution to the DNA damage response (DDR)
The MTA2/NuRD complex is a crucial regulator in the DDR (38). MTA2/NuRD collaborates at DNA double-strand break (DSB) sites to facilitate repair and regulate checkpoint activation, maintaining genomic stability.
Expression in normal human tissues
Futamura et al (39) reported MTA2 expression in all 16 adult normal tissues examined by Northern blotting, with high levels in testis, brain, heart, lung, liver and kidney. High expression in the testis and significant expression in the ovary and heart was further confirmed (40). Lung tissue expression was reported by Matsusue et al (41).
Pathogenic mechanisms of MTA2
MTA2 is notably overexpressed in various malignant tumor types, including cervical cancer (CC) (7), hepatocellular carcinoma (HCC) (42), osteosarcoma (43), non-small cell lung cancer (NSCLC) (44), thymoma (45), esophageal squamous cell carcinoma (ESCC) (46), pancreatic ductal adenocarcinoma (PDAC) (47), gastric cancer (GC) (48), breast cancer (BC) (16) and glioma (49). This overexpression correlates with more aggressive tumor biology and increased metastatic potential. In cancer, MTA2 is more often amplified than deleted, indicating that its role in carcinogenesis is mainly due to increased expression rather than mutations that alter protein function (11). Functionally, MTA2 activates multiple signaling pathways, including the Rho pathway, and regulates global gene-expression networks. This induces cytoskeletal reorganization and enhances the distant migration and invasive capabilities of tumor cells (11). The pathogenic mechanisms of MTA2 in human diseases have now been largely elucidated (Fig. 3 and Table I).
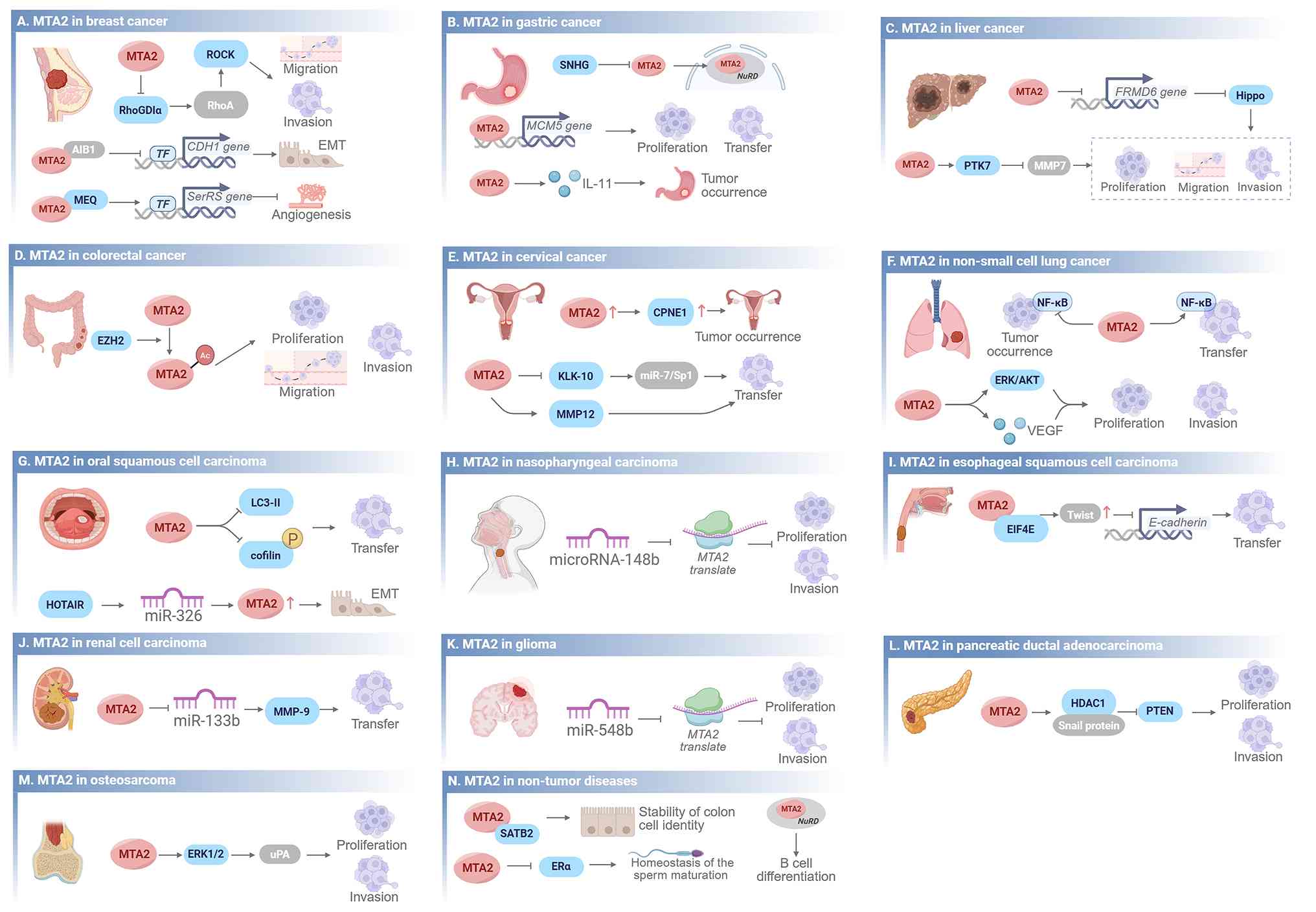 |
Figure 3
Pathogenic mechanisms of MTA2; this figure provides a detailed overview of the pathogenic mechanisms of MTA2 in 14 diseases. Arrows indicate a promoting effect on downstream signaling pathways and blunt-ended arrows indicate an inhibitory effect. (A) Breast cancer: MTA2 promotes migration, invasion and EMT via the RhoGDIα/ROCK and CDH1 pathways, and inhibits angiogenesis via the MEQ/SerRS axis. (B) Gastric cancer: MTA2 promotes proliferation, transfer and tumor occurrence through SNHG, MCM5 and IL-11 signaling. (C) Liver cancer: MTA2 drives proliferation, migration and invasion by inhibiting the Hippo pathway via FRMD6, and through the PTK7/MMP7 axis. (D) Colorectal cancer: MTA2 promotes proliferation, migration and invasion via EZH2-related epigenetic regulation. (E) Cervical cancer: MTA2 contributes to tumor occurrence via CPNE1, and promotes transfer via the KLK-10/miR-7/Sp1 and MMP12 pathways. (F) Non-small cell lung cancer: MTA2 promotes tumor occurrence, transfer, proliferation and invasion through NF-κB, ERK/AKT and VEGF signaling. (G) Oral squamous cell carcinoma: MTA2 regulates transfer via the LC3-II/cofilin pathway, and the HOTAIR/miR-326 axis upregulates MTA2 to drive EMT. (H) Nasopharyngeal carcinoma: miR-148b inhibits MTA2 translation and MTA2 subsequently promotes proliferation and invasion. (I) Esophageal squamous cell carcinoma: MTA2 drives transfer via the EIF4E/Twist/E-cadherin pathway. (J) Renal cell carcinoma: MTA2 promotes transfer via the miR-133b/MMP-9 axis. (K) Glioma: miR-548b inhibits MTA2 translation, and MTA2 subsequently promotes proliferation and invasion. (L) Pancreatic ductal adenocarcinoma: MTA2 regulates proliferation and invasion via the HDAC1/Snail/PTEN pathway. (M) Osteosarcoma: MTA2 promotes proliferation and invasion through the ERK1/2/uPA pathway. (N) Non-tumor diseases: MTA2 maintains colon cell identity stability, sperm maturation homeostasis and B-cell differentiation. AIB1, amplified in breast cancer 1; CDH1, cadherin 1; CPNE1, copine 1; EIF4E, eukaryotic initiation factor 4E; EMT, epithelial-mesenchymal transition; ERK, extracellular signal-regulated kinase; EZH2, enhancer of zeste homolog 2; FRMD6, FERM domain containing 6; IL-11, interleukin-11; KLK10, kallikrein-related peptidase 10; MCM5, minichromosome maintenance complex component 5; MMP, matrix metalloproteinase; MTA, metastasis-associated; NuRD, nucleosome remodeling and deacetylase; PTK7, protein tyrosine kinase 7; PTZN, phosphatase and tensin homolog deleted on chromosome ten; RhoGDIα, Rho GTPase dissociation inhibitor α; ROCK, Rho associated coiled-coil containing protein kinase; TF, transcription factor; uPA, urokinase-type plasminogen activator; VEGF, vascular endothelial growth factor.
|
 |
Table I
Pathogenic mechanisms of MTA2.
|
Table I
Pathogenic mechanisms of MTA2.
| Cancer type |
Authors, year |
Study population |
Mechanism |
(Refs.) |
| Breast cancer |
Zhang et al, 2019 |
Human BC cell line (ZR-75-30, MDA-MB-231) |
MTA1 induces MTA2 C-terminal cleavage, leading to its degradation and establishing an antagonistic relationship |
(55) |
| Gastric cancer |
Shi et al, 2021 |
Human GC cell line (MKN45, AGS, KATO III) |
Combined with replication-initiation sequences, intensifies olaparib-induced replication stress and enhances PARP inhibitor sensitivity |
(61) |
| Xie et al, 2024 |
Human GC cell line (MKN45, AGS, SGC7901) |
circMTA2 binds UCHL3, inhibits MTA2 ubiquitination and degradation, stabilizes its expression and promotes tumor progression |
(62) |
| Rectal cancer |
Zhou et al, 2014 |
Human RC cell line, mouse model (HCT116, Rat1) |
p300 mediates K152 acetylation, enhancing MTA2 oncogenic activity and promoting proliferation, migration and invasion |
(73) |
| Uraki et al, 2014 |
Human RC cell line (COLO-320, SW480, SW620) |
hBD-3 downregulates MTA2 mRNA via paracrine signaling, inhibiting colorectal cancer cell migration |
(74) |
| Non-small cell lung cancer |
Zhang et al, 2015 |
Human NSCLC cell line, human lung fibroblast cell line (A549, H460, WI-38) |
Combines with epithelial markers and their suppressor promoters to trigger EMT and tumor metastasis |
(84) |
| Renal cell carcinoma |
Lee et al, 2022 |
Human RCC cell line (Caki-1, ACHN) |
Co-suppression of MTA2 expression reduces renal cell carcinoma migration and invasion capabilities |
(112) |
| Glioma |
Cheng et al, 2014 |
Human glioma cell line (GBM8401, M059J, M059K) |
MTA2 downregulation inhibits cell growth and migration, induces G0/G1 arrest and correlates with tumor grading |
(49) |
| Pancreatic ductal adenocarcinoma |
Zhu et al, 2018 |
Human pancreatic carcinoma cell line (PANC-1, BxPC-3, SW1990) |
Under hypoxia, HIF-1α upregulates MTA2, which enhances HIF-1α stability, jointly driving EMT |
(106) |
| Spermatogenesis |
Zhu et al, 2014 |
mouse model (Primary murine spermatogenic cells) |
Upregulates MTA2 in supporting cells, transcriptionally suppresses Fshr, and maintains phagocytic function and spermatogenesis |
(125) |
Across diverse malignancies, MTA2 displays three conserved downstream regulatory patterns: First, it generally represses epithelial markers to trigger EMT; second, it commonly upregulates multiple matrix metalloproteinase (MMP) family proteins to boost tumor invasiveness; third, frequent PI3K/AKT overactivation occurs after MTA2-mediated phosphatase and tensin homolog deleted on chromosome ten (PTEN) suppression. Meanwhile, various non-coding RNAs form conserved competitive endogenous RNA (ceRNA) loops to modulate MTA2 abundance. Despite these shared mechanisms, distinct upstream regulators and downstream effector molecules are employed in different tumor types, as elaborated below for each disease.
MTA2 in BC
BC remains a major global health challenge due to its complex pathogenesis and varied clinical presentations, which hinder effective treatment and prevention (50-53). In BC, MTA2 primarily exerts its functions by regulating cytoskeletal reorganization and the Rho/Rho associated coiled-coil containing protein kinase (ROCK) signaling pathway. MTA2 inhibits Rho GTPase dissociation inhibitor α (RhoGDIα), thereby activating Rho GTPases such as RhoA and enhancing downstream ROCK activity (16). This activation promotes actin restructuring and lamellipodia formation, conferring amoeboid-like motility to breast cancer cells and significantly increasing migration and invasion. Reintroducing RhoGDIα or administering ROCK-specific inhibitors suppresses the MTA2-induced migration phenotype (11). In ERα-negative breast cancers, MTA2 expression undergoes reciprocal regulation by the Rho pathway: RhoGDIα overexpression reduces MTA2 protein levels, while MTA2 upregulation inhibits RhoGDIα, forming a positive feedback loop (16). In ERα-positive breast cancers, MTA2 overexpression is frequently accompanied by RhoGDIα downregulation. This is linked to tamoxifen resistance and increased anchorage-independent growth (15).
The antitumor small molecule MEQ directly binds MTA2, disrupting its interaction with the Mi-2/NuRD complex. This lifts repression of the seryl-tRNA synthetase (SerRS) gene and inhibits vascular endothelial growth factor A (VEGFA)-driven angiogenesis (54).
In BC, MTA1 overexpression reduces MTA2 protein levels without affecting mRNA. MTA1 induces activation of neutrophil elastase, which cleaves the C-terminal region of MTA2 at residues 486, 497, 542, 583 and 621, leading to MTA2 degradation. This antagonism partially explains why MTA1 can inhibit metastasis, whereas MTA2 promotes it (55).
MTA2 in GC
GC is a significant global health concern (56). It ranks as the fifth most prevalent cancer and the third leading cause of cancer-related mortality worldwide, with >1 million new cases annually (57,58). MTA2 contributes to GC progression via several mechanisms. As a transcriptional regulator, MTA2 binds the promoter of minichromosome maintenance complex component 5, enhancing GC cell proliferation and metastatic potential (59). Overexpression of MTA2 promotes clonogenic formation and tumorigenesis in vivo, partly through activation of the downstream interleukin-11 (IL-11) pathway (60).
MTA2 preferentially binds DNA replication origins. This exacerbates olaparib-induced replication stress and renders BRCA-wild-type GC cells more sensitive to poly(ADP ribose) polymerase (PARP) inhibition, supporting targeted clinical use of PARP inhibitors in selected GC groups (61).
Circular RNA (circ)MTA2 binds the deubiquitinating enzyme ubiquitin C-terminal hydrolase L3 (UCHL3), preventing ubiquitination and degradation of the MTA2 protein. This stabilizes MTA2 and promotes tumor progression; the axis can be transmitted via exosomes (62). Conversely, long non-coding (lnc)RNAs in the small nucleolar RNA host gene family (SNHGs) retain MTA2 in the cytoplasm, preventing NuRD complex formation. This elevates acetylation of histone H3 and p53 and suppresses GC-cell proliferation and metastasis (63).
MTA2 in HCC
Liver cancer poses a significant global health challenge, with projections indicating it will impact >1 million individuals each year by 2025. HCC accounts for 90% of primary liver cancers and ranks as the fourth leading cause of cancer-related mortality globally (64-66). MTA2, an integral part of the NuRD complex, suppresses the transcription of target genes by facilitating histone deacetylation and independently regulates gene expression without relying on transcription factors. Immunohistochemical analysis reveals significantly elevated MTA2 expression in HCC tissues, primarily localized to the nucleus. Analysis of The Cancer Genome Atlas database further confirms markedly elevated MTA2 mRNA levels in HCC. Furthermore, MTA2 expression correlates with increased α-fetoprotein (ATP) levels in patient cohorts, suggesting a potential association with HCC recurrence (67).
Research indicates that MTA2 accelerates the growth, proliferation and metastasis of HCC cells by inhibiting the Hippo signaling pathway through suppression of genes such as FERM Domain Containing 6 (FRMD6). The MTA2/FRMD6/Hippo signaling axis plays a crucial role in HCC development and progression (68).
MTA2 in rectal cancer (RC)
RC ranks as the third most prevalent cancer type worldwide and is the second leading cause of cancer-related mortality, comprising one-third of these cases (69). Despite a general decline in colorectal cancer incidence and mortality, RC rates among individuals under 50 years of age are rapidly increasing, with projections indicating a 124.2% rise for those aged 20 to 34 by 2030 in the United States (70-72). In RC, the biological function of MTA2 is precisely regulated through post-translational modifications. Among these, acetylation at lysine 152 mediated by the histone acetyltransferase p300 significantly enhances the oncogenic activity of MTA2, promoting proliferation, migration and invasion in RC cells. Conversely, mutations at this site inhibit RC cell growth and attenuate the migration and invasion capabilities of Rat-1 fibroblasts (73).
At the level of the immune microenvironment, RC progression is also regulated by MTA2. Tumor-infiltrating monocytes secrete human beta-defensin 3 (hBD-3) and inhibit cancer cell migration through a paracrine mechanism. This inhibitory effect is closely associated with downregulation of MTA2 mRNA. Importantly, hBD-3 does not directly inhibit cancer cell proliferation (74).
MTA2 in CC
CC, primarily caused by persistent high-risk human papillomavirus (HR-HPV) infection, is a major cause of mortality among women globally and in the United States (75). CC remains a significant public health issue, with ~600,000 new cases and 300,000 deaths annually, making it the fourth most prevalent cancer type among women (76). MTA2 promotes tumor metastasis by negatively regulating kallikrein-related peptidase 10 (KLK10) expression and activating the microRNA (miR)-7/specificity protein 1 (Sp1) signaling axis. MTA2 is highly expressed in CC tissues, while KLK10, a known inhibitor of tumor migration, shows a significant negative correlation with MTA2 levels. Knockout of MTA2 leads to marked upregulation of KLK10. Concurrently, downregulation of MTA2 activates miR-7, which suppresses the activity of transcription factor SP1, forming a multilevel cascade regulatory network. Within this network, SP1 undergoes both direct regulation by MTA2 and indirect suppression by miR-7. Together, these factors act on downstream targets to influence the migratory capacity of CC cells (77).
Additionally, MTA2 exhibits synergistic carcinogenic effects with the calcium-calcium-phospholipid-binding protein copine 1 (CPNE1) in CC. MTA2 and CPNE1 expression levels show a significant positive correlation (r=0.668, P<0.01), suggesting they may act through shared signaling pathways to promote tumorigenesis and progression (78).
MTA2 in NSCLC
Lung cancer is the most common cause of cancer-associated death worldwide, with an estimated 1.6 million deaths each year (79,80). NSCLC accounts for ~85% of lung cancer cases, with lung adenocarcinoma and lung squamous cell carcinoma being the predominant subtypes (81). NSCLC is characterized by significantly elevated MTA2 expression. Research confirms that a specific antibody targeting MTA2 effectively blocks proliferation, migration and invasion in NSCLC cells. Further findings reveal that MTA2 activation is closely associated with enhanced activity in the extracellular signal-regulated kinase (ERK)/AKT signaling pathway and the VEGF pathway, while inhibition of MTA2 expression markedly reduces the phosphorylation levels and signaling efficiency of key molecules within these pathways. Silencing MTA2 reduces phosphorylation of ERK and AKT, significantly inhibiting NSCLC cell growth and invasiveness (82).
MTA2, a crucial part of the NuRD complex, plays dual roles in tumor progression. During the primary lung tumor stage, MTA2 negatively regulates NF-κB signaling independently of inhibitor of NF-κB kinase subunit beta (IKK2), thereby inhibiting tumor growth (83). However, when tumors progress to the metastatic stage, MTA2 dissociates from NF-κB target gene promoters and shifts to positively regulating NF-κB in an IKK2-dependent manner, thereby promoting EMT and cancer metastasis (83).
MTA2 is essential for maintaining the malignant phenotype of lung cancer and is highly expressed in invasive lung cancer cells. At the molecular level, MTA2 directly binds to the promoter regions of epithelial markers such as epithelial cell adhesion molecule (EPCAM) and cadherin 1 (CDH1), thereby suppressing the transcription of these genes and inducing EMT and tumor metastasis. Functional experiments demonstrate that MTA2 overexpression suppresses the promoter activity of CDH1 or EPCAM, whereas its deletion enhances the activity of these promoters (84). This stage-dependent dual role (tumor-suppressive in primary lesions, oncogenic in metastatic stages) indicates MTA2 function is highly context-dependent, regulated by NF-κB signaling and IKK2 dependency. Such stage-dependent dual functions of MTA2 in NSCLC are further fine-tuned by competitive dynamics between MTA2 and other MTA paralogs, where altered MTA1 abundance can displace MTA2 from NuRD complexes and reshape NF-κB-dependent tumor phenotypes in a context-specific manner.
MTA2 in head and neck malignant tumors
Head and neck cancer, which arises in the lining or mucosa of the upper aerodigestive tract, specifically the oral cavity, oropharynx, larynx, hypopharynx and paranasal sinuses, is the seventh most common cancer type worldwide (85).
Oral squamous cell carcinoma (OSCC) significantly contributes to the morbidity and mortality associated with head and neck squamous cell carcinoma (86,87). Although there have been recent advancements in surgical methods, adjuvant therapy, molecular targeted therapy and immunotherapy, the outcomes remain inadequate (88). Studies on OSCC reveal that MTA2 enhances oral cancer cell migration and invasion by reducing the expression of autophagy markers light chain 3-II and phosphorylated cofilin. Suppression of MTA2 expression via gene knockdown markedly reverses these malignant phenotypes (89). At the regulatory level, lncRNA HOTAIR functions as a competitive endogenous RNA. By binding to miR-326, it relieves miR-326-mediated translational repression of MTA2 mRNA, thereby indirectly upregulating MTA2 expression. This HOTAIR/miR-326/MTA2 axis shows a significant positive correlation with EMT in OSCC, suggesting a pivotal role in tumor progression (90). Nasopharyngeal carcinoma (NPC) is an epithelial cancer originating from the mucosal lining of the nasopharynx (91). Tumors in the nasopharynx are frequently located at the pharyngeal recess. NPC exhibits a unique geographic distribution, being notably common in East and Southeast Asia (92). In NPC pathogenesis, MTA2 is directly targeted and regulated by miR-148b. miR-148b is notably downregulated in NPC and suppresses MTA2 expression by targeting the 3'-UTR of MTA2 mRNA to inhibit its translation. Restoration of miR-148b expression significantly inhibits NPC cell proliferation, migration, and invasion. Reintroduction of MTA2 partially counteracts the tumor-suppressive effects mediated by miR-148b, indicating that miR-148b exerts its antitumor activity in part by inhibiting MTA2 (93). Furthermore, analysis across ethnic groups reveals that in Han Chinese patients with NPC, MTA2 expression negatively correlates with patient age, with no significant association with clinical stage or lymph node metastasis. In Zhuang patients, MTA2 expression shows no correlation with clinical indicators (94).
MTA2 in other solid tumors of the digestive system
Globally, esophageal cancer ranks as the ninth most prevalent cancer and the sixth leading cause of cancer-related mortality (95). The two primary histological subtypes, ESCC and esophageal adenocarcinoma (EAC), exhibit notable geographical distribution differences (96,97). ESCC is the most prevalent type, commonly found in developing countries, with its incidence declining in developed nations (98). Research on ESCC has identified a regulatory positive feedback loop between eukaryotic initiation factor 4E (EIF4E) and Twist in controlling EMT. MTA2 binds to EIF4E, which subsequently increases the expression of Twist, a crucial EMT regulator. Twist recruits MTA2 to the E-cadherin gene promoter, suppressing its transcription by decreasing histone acetylation. This positive feedback loop significantly promotes the metastatic process in ESCC (99).
Conversely, the lncRNA SNHG5 binds directly to the MTA2 protein, preventing its nuclear translocation and facilitating its degradation through ubiquitination. Furthermore, overexpression of SNHG5 can downregulate MTA2 expression at the transcriptional level. In esophageal cancer, downregulation of SNHG5 leads to abnormal accumulation of MTA2 protein, thereby accelerating the EMT process (100).
PDAC ranks as the fourth most common cause of cancer-related mortality in developed nations (101). By 2030, PDAC is projected to become the second leading cause of cancer-related mortality in the United States (102,103). Research on PDAC indicates that MTA2 recruits Snail protein and interacts with HDAC1 at the PTEN promoter, forming a transcriptional repression complex. The complex suppresses PTEN expression via histone deacetylation, diminishing negative regulation of the PI3K/Akt pathway and thereby enhancing PDAC cell proliferation, migration and invasion (104). Experimental evidence shows that benzyl isothiocyanate (BITC) treatment of MIA Paca-2 and PANC-1 cell lines leads to a dose- and time-dependent reduction in MTA2 expression, along with increased PTEN expression and reduced phosphorylated PI3K and AKT levels (105).
Research shows that MTA2 and HIF-1α create a positive feedback loop under hypoxic conditions. HIF-1α binds to the hypoxia response element (HRE) in the MTA2 promoter, boosting MTA2 transcription. Concurrently, the MTA2-HDAC1 complex deacetylates HIF-1α, increasing its protein stability. Activated HIF-1α further recruits MTA2 to the E-cadherin promoter region, suppressing the expression of this epithelial marker and driving the EMT process (106).
MTA2 in renal cell carcinoma (RCC)
RCC accounts for 2-3% of all malignant diseases in adults. It ranks as the seventh most prevalent cancer in men and the ninth in women. Globally, there are ~209,000 new cases and 102,000 deaths annually (107). The rising incidence of all stages of this cancer over the years has led to a continuous increase in population-based mortality rates (108-110). Previous studies have confirmed that MTA2 negatively regulates miR-133b expression, thereby weakening the latter's binding to and inhibition of the 3' untranslated region of MMP9 mRNA. This ultimately promotes MMP9 expression, further enhancing the migration and invasive capacity of RCC. Research indicates that reducing MTA2 expression significantly inhibits RCC migration, invasion and metastatic potential in vivo, without appreciably affecting cell proliferation. In RCC cells with low MTA2 expression, the enzymatic activity, protein levels and mRNA transcription levels of MMP9 are all reduced (111). Another study indicated that melatonin combined with sorafenib synergistically reduces MTA2 expression, effectively inhibiting cancer cell migration and invasion in RCC cells (112).
MTA2 in glioma
Gliomas, comprising 81% of central nervous system malignancies, are the most prevalent primary brain tumors. Gliomas arise from neuroglial progenitor cells (113), evolving into astrocytoma, oligodendroglioma, ependymoma or oligoastrocytoma (114,115). The World Health Organization (WHO) classifies gliomas into four grades: Grades 1 and 2 are low-grade, while grades 3 and 4 are high-grade gliomas (HGG) (116). During the development of malignant gliomas, MTA2 expression is directly negatively regulated by miR-548b. This miRNA targets the 3'-UTR of MTA2 mRNA, leading to its degradation or translational inhibition, thereby decreasing MTA2 expression. Experimental evidence shows that miR-548b overexpression decreases MTA2 protein levels, thereby inhibiting glioma cell proliferation and invasive capabilities. Further rescue experiments demonstrate that restoring MTA2 expression can counteract the tumor-suppressive effects mediated by miR-548b (117).
Additionally, MTA2 participates in the progression of glioma by regulating cell-cycle processes. Research shows that significantly lowering MTA2 expression hinders the proliferation, migration and invasion of human glioblastoma cell lines GBM8401 and Hs683 and induces cell-cycle arrest at the G0/G1 phase. MTA2 expression was notably elevated in GBM8401 and Hs683 cells compared to other glioma cell lines, including M059J, M059K and U-87MG, at both the mRNA and protein levels. MTA2 expression in glioma tissues significantly correlates with tumor grade (P<0.001), indicating its potential as a biomarker for disease progression (49).
MTA2 in osteosarcoma
Osteosarcoma, the most prevalent primary malignant bone tumor, is characterized by malignant mesenchymal cells that produce osteoid or immature bone. It has a global incidence of about one to three cases per million annually (118,119). It mainly occurs in children and teenagers, with another increase in cases seen in individuals aged >50 years (120). Research on osteosarcoma has identified that MTA2 regulates urokinase-type plasminogen activator (uPA) expression through the ERK1/2 signaling pathway, significantly enhancing the migration and invasion of osteosarcoma cells. Experimental results indicate that knocking down MTA2 downregulates uPA expression and inhibits the formation of pulmonary metastatic lesions. Notably, treatment of MTA2-knockdown osteosarcoma cells with recombinant human (rh)-uPA leads to a marked recovery in migration and invasion capabilities (121).
Mechanistic roles of MTA2 in non-neoplastic disease-related biological processes
In non-neoplastic diseases, earlier findings indicate that MTA2 regulates the plasticity of intestinal cell lineages. It acts synergistically with SATB homeobox 2, a protein that binds specifically to AT-rich sequences, to maintain the stability of colonic cell identity. Further studies indicate that MTA2 and hepatocyte nuclear factor 4α (HNF4A) co-occupy DNA on colonic chromatin. In the absence of MTA2, HNF4A detaches from colonic chromatin and binds to small intestinal chromatin, leading to the activation of genes involved in lipid absorption. This promotes functional lipid uptake processes and ultimately drives the transformation of colonic cells toward a small intestinal phenotype. This mechanism highlights the central role of MTA2 in tissue-specific enhancer regulatory networks (122).
In the epididymal epithelial cells of the male reproductive system, MTA2 was found to colocalize with ERα in the nucleus. By inhibiting the transactivation function of ERα, it participates in regulating the luminal microenvironment homeostasis essential for sperm maturation (123). MTA2 expression is notably increased in testicular Sertoli cells during the phagocytosis of apoptotic spermatogenic cells, leading to the transcriptional suppression of the follicle-stimulating hormone (FSH) receptor (FSHR) gene (124). This FSH/MTA2/FSHR negative feedback loop is crucial for maintaining the integrity of spermatogenesis by sustaining normal phagocytic function in supporting cells through the reduction of circulating FSH levels (125).
The MTA2/NuRD complex modulates the expression of crucial genes, including immunoglobulin lambda-like polypeptide 1 and V-set pre-B cell surrogate light chain 1, in pre-B cells during B-lymphocyte development by interacting with transcription factors IKAROS family zinc finger 3 (AIOLOS, IKZF3) and IKAROS family zinc finger 1 (IKAROS, IKZF1). The absence of MTA2 increases histone H3 lysine 27 acetylation (H3K27ac) at gene promoter regions, disrupting B-cell differentiation. This results in developmental defects in pro-B cells, immature B cells and marginal zone B cells, as well as abnormal differentiation of germinal center B cells (126).
MTA2 and MTA3 collaboratively maintain pluripotency in human embryonic stem cells by repressing genes linked to mesodermal differentiation. Studies show that ID8, an inhibitor targeting dual-specificity tyrosine phosphorylation-regulated kinases (DYRKs), partially mitigates the differentiation phenotype resulting from MTA2 or MTA3 knockdown. This effect is primarily mediated by inhibiting DYRK4 activity, suggesting that DYRK4 may function downstream of MTA2 or MTA3 in this pathway (127).
Collectively, MTA2 functions as a conserved epigenetic and metastatic driver across nearly all human cancers examined. All tumor types share core MTA2-mediated mechanisms, including NuRD complex-dependent transcriptional repression, induction of EMT, enhancement of MMP activity and non-coding RNA-mediated regulation. However, tissue-specific pathogenic mechanisms are evident: BC relies on Rho/ROCK cytoskeletal signaling; digestive system malignancies converge on epigenetic silencing, hypoxia/HIF-1α loops and mTOR pathway dysregulation; NSCLC uniquely exhibits stage-specific biphasic NF-κB regulation; gynecologic and urologic tumors act through distinct miRNA-MMP axes; head and neck cancers feature ceRNA crosstalk and ethnicity-associated expression patterns; and glioma and osteosarcoma are characterized by aberrant cell-cycle control and ERK/uPA-driven invasion. By contrast, MTA2 contributes to non-neoplastic homeostasis by maintaining lineage stability, reproductive function, immune development and stem cell pluripotency, representing a fundamentally distinct role from its oncogenic activity. This combination of shared core machinery and tissue-specific effector pathways defines the multifaceted landscape of MTA2-mediated pathogenesis.
Clinical implications and therapeutic frontiers across disease landscapes of MTA2
MTA2, a protein with a key mechanistic role in tumorigenesis and progression, is significantly overexpressed in multiple malignant tumor types. Consistently across most malignancies, elevated MTA2 expression serves as a universal poor prognostic marker linked to advanced tumor stage, lymphatic metastasis and shortened survival. A total of four shared therapeutic directions have been validated preclinically: Disrupting the MTA2-NuRD complex, restoring tumor-suppressive miRNAs, small-molecule intervention against MTA2-associated axes and inhibiting downstream EMT/MMP/PI3K cascades. Despite these universal rules, each tumor displays distinctive prognostic characteristics and exclusive targeted regimens are detailed below with cross-cancer comparison (Fig. 4, Table II).
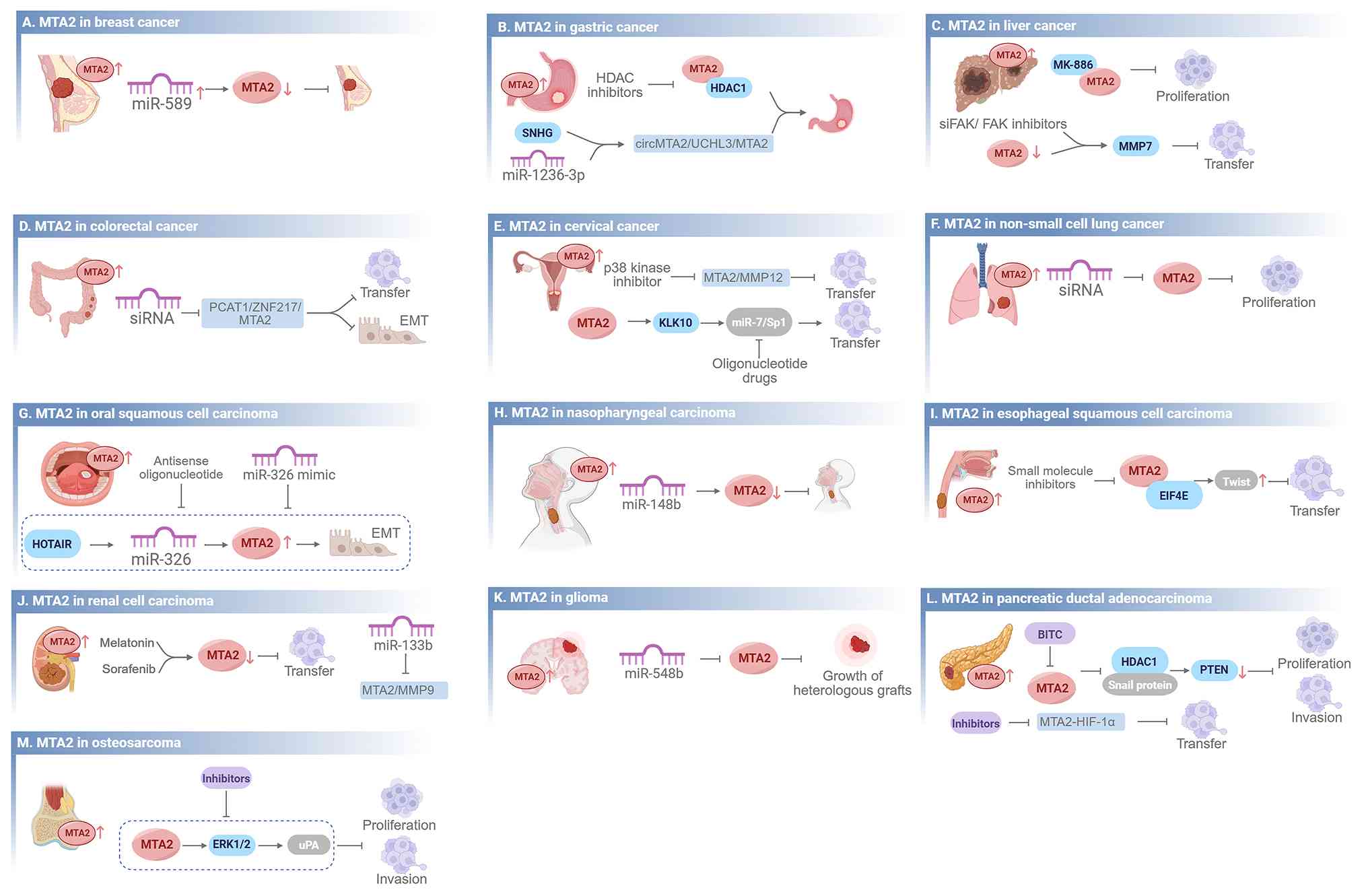 |
Figure 4
Clinical implications and therapeutic frontiers across disease landscapes of MTA2. This figure highlights the regulatory effects of microRNAs, targeted inhibitors, and therapeutic interventions on MTA2 expression and function. Arrows indicate a promoting effect on downstream signaling pathways and blunt-ended arrows indicate an inhibitory effect. (A) Breast cancer: miR-589 inhibits MTA2 expression, thereby suppressing tumor progression. (B) Gastric cancer: HDAC inhibitors suppress MTA2 activity; SNHG and miR-1236-3p regulate the circMTA2/UCHL3/MTA2 axis. (C) Liver cancer: MK-886 inhibits proliferation; siFAK/FAK inhibitors and MTA2-mediated MMP7 suppression reduce transfer. (D) Colorectal cancer: MTA2 siRNA and targeting the PCAT1/ZNF217/MTA2 axis inhibit transfer and EMT. (E) Cervical cancer: p38 kinase inhibitors and oligonucleotide drugs targeting the miR-7/Sp1 pathway inhibit MTA2-mediated transfer. (F) Non-small cell lung cancer: MTA2 siRNA inhibits proliferation. (G) Oral squamous cell carcinoma: Antisense oligonucleotides and miR-326 mimics target the HOTAIR/miR-326/MTA2 axis to inhibit EMT. (H) Nasopharyngeal carcinoma: miR-148b inhibits MTA2 expression and suppresses tumor progression. (I) Esophageal squamous cell carcinoma: Small molecule inhibitors targeting the MTA2-EIF4E-Twist axis inhibit transfer. (J) Renal cell carcinoma: Melatonin and sorafenib downregulate MTA2 to inhibit transfer; miR-133b suppresses the MTA2/MMP9 axis. (K) Glioma: miR-548b inhibits MTA2 and suppresses the growth of heterologous grafts. (L) Pancreatic ductal adenocarcinoma: BITC and MTA2-HIF-1α inhibitors suppress proliferation, invasion and transfer via the HDAC1/Snail/PTEN pathway. (M) Osteosarcoma: Inhibitors targeting the MTA2-ERK1/2-uPA axis suppress proliferation and invasion. BITC, benzyl isothiocyanate; EIF4E, eukaryotic initiation factor 4E; EMT, epithelial-mesenchymal transition; ERK, extracellular signal-regulated kinase; FAK, focal adhesion kinase; HDAC, histone deacetylase; HIF-1α, hypoxia-inducible factor 1α; KLK10, kallikrein-related peptidase 10; MMP, matrix metalloproteinase; MTA, metastasis-associated; PTZN, phosphatase and tensin homolog deleted on chromosome ten; SNHGs, small nucleolar RNA host gene family; UCHL3, ubiquitin C-terminal hydrolase L3; uPA, urokinase-type plasminogen activator.
|
|
Table II
Clinical implications and therapeutic frontiers across disease landscapes of MTA2.
|
Table II
Clinical implications and therapeutic frontiers across disease landscapes of MTA2.
| Cancer type |
Role of MTA2 in disease diagnosis or prognosis assessment |
Role of MTA2 in disease treatment |
(Refs.) |
| Breast cancer |
Overexpression correlates with lymph node metastasis, advanced stage and poor prognosis; it predicts endocrine resistance in ER+ patients. |
Isoflavone derivative MEQ inhibits triple-negative breast cancer progression by targeting MTA2. |
(54,128-131) |
| Gastric cancer |
High expression is an independent poor-prognosis indicator, associated with stage, metastasis and short survival; combined with HDAC1 testing improves accuracy. |
Gastric cancer cells with high MTA2 expression are sensitive to Olaparib and AZD6738, targeted inhibition of MTA2 can enhance the therapeutic effect of gastric cancer when combined with drugs. |
(61-63,133,134) |
| HCC |
High expression correlates with short survival, TNM staging, vascular invasion and immune infiltration characteristics. |
MK-886, SB203580 and PF-573228 can target MTA2 to inhibit the proliferation and induce the death of hepatocellular carcinoma cells. |
(67,135,136) |
| Rectal cancer |
High expression is positively correlated with tumor grade and associated with short progression-free survival. |
GSK126 can inhibit colorectal cancer progression by targeting MTA2-related pathways, and human β-defensin-3 can downregulate MTA2 expression to suppress colon cancer cell migration. |
(36,73,74,137) |
| Cervical cancer |
High expression correlates with malignant progression, FIGO staging and metastasis, shortening disease-free survival as a prognostic biomarker. |
SB203580 can inhibit cervical cancer cell metastasis by targeting MTA2-related pathways. |
(77,78,138,140) |
| Non-small cell lung cancer |
High expression correlates with low differentiation, metastasis and poor prognosis as an independent prognostic indicator. |
HDAC inhibitors can intervene in tumor growth and metastasis by regulating the MTA2/NuRD and NF-κB pathways. |
(82-84) |
| Oral squamous cell carcinoma |
High expression correlates with increased tumor grade and low survival rates in stage III patients. |
- |
(90) |
| Nasopharyngeal carcinoma |
Expression varies by ethnicity; low expression in Zhuang patients is associated with higher survival rates under specific chemotherapy regimens. |
miR-148b can target and inhibit MTA2 to suppress the proliferation, migration and invasion of nasopharyngeal carcinoma cells. |
(93,94) |
| Esophageal squamous cell carcinoma |
- |
SNHG5 can inhibit cancer cell proliferation, metastasis and epithelial-mesenchymal transition by ubiquitinating and degrading MTA2. |
(99,100) |
| Renal cell carcinoma |
High expression correlates with tumor grade, stage and short survival as an independent prognostic indicator. |
miR-133b can target and regulate the MTA2 pathway to inhibit renal cancer cell metastasis, and melatonin combined with sorafenib can synergistically suppress MTA2 expression and block the invasion and metastasis of renal cancer cells. |
(111,112) |
| Glioma |
High expression correlates positively with tumor grade and can assist in assessing malignancy. |
miR-548b can target and inhibit MTA2 expression to suppress glioma cell proliferation, invasion and tumorigenesis in vivo. |
(49,117) |
| Pancreatic ductal adenocarcinoma |
High expression correlates positively with tumor grade, serving as an independent adverse prognostic indicator associated with short survival duration. |
BITC and miR-146a inhibit MTA2 to suppress cancer cell proliferation. |
(105,106) |
| Osteosarcoma |
High expression correlates with advanced stage and short survival, enabling prognosis assessment. |
- |
(121) |
MTA2 in BC
In BC targeted therapy, small-molecule inhibitors such as MEQ, which target the MTA2-NuRD complex, can restore the expression of tumor suppressor genes such as SerRS, thereby inhibiting tumor angiogenesis (54). Furthermore, disrupting the interaction between MTA2 and amplified in breast cancer 1 (AIB1) can upregulate E-cadherin, partially reversing EMT (128). Enhancing miR-589 expression or altering the TTC39A-AS1/miR-483-3p/MTA2 signaling pathway markedly decreases MTA2 levels, suggesting a promising therapeutic approach for triple-negative BC (129,130). In ER+ BC, the synergistic inhibitory effect mediated by MTA2 and AIB1 indicates that therapy should be combined with endocrine strategies (128). Additional evidence indicates that knocking out the MTA2 gene increases tumor-cell sensitivity to chemotherapeutic drugs and induces apoptosis (131). In ER+ BC, the presence of the MTA2-AIB1 complex may predict resistance to endocrine therapy (128).
MTA2 in GC
In targeted strategies for GC, epigenetic modulators targeting the MTA2-HDAC1 complex (e.g., HDAC inhibitors) may reverse the malignant phenotype of GC. MTA2 expression may also serve as a biomarker for predicting the efficacy of PARP-inhibitor therapy, such as Olaparib; when used with ataxia-telangiectasia and Rad3-related (ATR) inhibitors, this approach increases tumor-cell sensitivity and treatment effectiveness (61). Furthermore, disrupting the circMTA2/UCHL3/MTA2 signaling axis may offer novel strategies for curbing GC progression, particularly by targeting exosome-mediated intercellular communication. This approach also lays the groundwork for developing targeted inhibitors and designing combination regimens (62). The natural inhibitory molecules SNHG5 (132) and miR-1236-3p exert antitumor effects by regulating MTA2; their analogues or overexpression techniques may represent novel avenues for GC gene therapy (63,133). Combined detection with HDAC1 may further enhance prognostic accuracy (134).
MTA2 in HCC
In HCC targeted-therapy research, intervention strategies targeting MTA2 show potential value. MK-886 interacts with MTA2 through α-alkyl and alkyl interactions, leading to significant inhibition of HepG2 cell growth and dose-dependent apoptosis (135). Additionally, small interfering (si)RNA targeted at focal adhesion kinase (FAK) or FAK inhibitors (PF-573228) exhibit synergistic effects in the context of low MTA2 expression, jointly inhibiting MMP7-mediated invasion and metastasis (136).
For prognosis in HCC, MTA2 shows significantly higher expression in tumor cells, and its upregulation is closely linked to poor clinical outcomes, with both MTA2 and the proliferation marker Ki-67 markedly higher than in adjacent normal tissue. Nuclear MTA2 expression is positively correlated with Ki-67 (P=0.019). Furthermore, patients with HCC with elevated MTA2 display immune-microenvironment features marked by higher levels of activated CD4+ T cells and M0-type macrophages, indicating that immune regulation may contribute to tumor development (135).
MTA2 in RC
In targeted therapy for RC, acetylation-focused interventions have gained attention. Inhibiting acetylation at K152 of MTA2 (e.g., with p300-specific inhibitors) may block its tumor-promoting functions, effectively suppressing tumor growth and metastasis (73). Preclinical models indicate that targeting lncRNAs in exosomes with antisense oligonucleotides or siRNA directed at the prostate cancer-associated transcript 1 (PCAT1)/ZNF217 (zinc finger protein 217)/MTA2 pathway may inhibit EMT and metastasis (137). In immune regulation, analogues or inducers of hBD-3 may suppress tumor cell migration by downregulating MTA2, offering a potential approach for metastatic disease in advanced RC (74). Small-molecule inhibitors that disrupt the MTA2-EZH2 interaction may restore TSC2 expression and suppress mTOR pathway activation, showing therapeutic potential in RC (36).
MTA2 in CC
Studies of therapeutic targets for CC show that small-molecule inhibitors, such as p38 kinase inhibitors, can hinder the MTA2/MMP12 pathway. This disruption affects activator protein-1 (c-Fos/c-Jun) binding to the MMP12 promoter by interfering with phosphorylation of Y-box binding protein 1 (YB1), ultimately suppressing MMP12 expression and reducing metastatic potential (138). Additionally, oligonucleotide agents targeting the miR-7/SP1 pathway have shown therapeutic potential; these may reverse MTA2-mediated metastasis by promoting KLK10 expression, informing combination strategies (77). In molecular subtyping and personalized therapy, a positive correlation between MTA2 and CPNE1 expression in patients with HR-HPV (types 16 and 18) may serve as an additional marker for individualized treatment (78,139).
For prognosis, high MTA2 expression is associated with malignant progression. Positive expression rates are 0% in normal cervical tissue, ~30% in cervical intraepithelial neoplasia (CIN) and up to 73.4% in cervical squamous cell carcinoma. Expression correlates with International Federation of Obstetrics and Gynecology stage and lymph node metastasis (140).
MTA2 in NSCLC
Targeting MTA2 shows promise for NSCLC. Small-molecule inhibitors aimed at the MTA2-ERK/AKT/VEGF axis (e.g., AbMTA2) in vivo effectively suppress tumor growth, and siRNA-mediated MTA2 knockdown significantly inhibits proliferation (82). HDAC inhibitors targeting the MTA2-NuRD complex may relieve repression of tumor suppressors such as E-cadherin, facilitating re-epithelialization (84). Strategies may be stage-specific: During the primary phase, enhance the inhibitory effect of MTA2-NuRD on NF-κB signaling; at the metastatic stage, block the IKK2-MTA2 interaction (83). Meanwhile, MTA1 antagonizes MTA2 by promoting its degradation, and competitive dynamics within the NuRD complex further determine MTA2's pro- or anti-tumor activity. This inter-MTA competitive balance is highly disease- and stage-specific, making NuRD functional outputs and therapeutic responses distinct in NSCLC compared with other cancer types.
MTA2 in head and neck malignant tumors
Targeting ceRNA networks shows promise. Antisense oligonucleotides or synthetic miR-326 mimics that suppress HOTAIR and restore miR-326 can disrupt the HOTAIR/miR-326/MTA2 axis, markedly reducing MTA2 and inhibiting EMT (90). Clinically, MTA2 levels in OSCC tissues are higher than in adjacent normal mucosa (P<0.01). Overexpression is associated with higher histological grade (P<0.01), underscoring MTA2's value for assessing OSCC progression and prognosis (90).
In NPC, miRNA-targeted therapy restoring or mimicking miR-148b suppresses MTA2 pathway activity and exerts antitumor effects (93). Given ethnic differences in MTA2 expression, treatment can be optimized. For patients of Zhuang ethnicity with lower MTA2 expression, carboplatin plus fluorouracil often yields more favorable outcomes. MTA2 expression also has prognostic value. In Zhuang patients with NPC and low MTA2 expression, the specified chemotherapy regimen notably enhances overall survival (OS) (hazard ratio=0.42, P<0.05) (94).
MTA2 in other solid tumors of the digestive system
Targeting the EIF4E/TWIST signaling pathway is a promising strategy for ESCC. Designing small-molecule inhibitors to disrupt the MTA2-EIF4E interaction could block TWIST-mediated upstream EMT events and reduce metastatic foci (99). Epigenetic regulation is another route: Applying SNHG5 analogs or overexpression vectors promotes MTA2 ubiquitination and degradation, significantly inhibiting proliferation, migration and invasion in ESCC cells. This approach reverses the EMT phenotype and restores epithelial characteristics (100).
In PDAC treatment strategies, intervening through epigenetic regulation by using small-molecule inhibitors (such as structural analogues of BITC) to target the interaction between MTA2 and HDAC1 can restore PTEN expression. In preclinical experiments, this intervention significantly reduces tumor volume (105). MTA2 is a central component and potential therapeutic target in the regulatory network of the hypoxic tumor microenvironment, crucial for inhibiting PDAC progression and metastasis. Blocking the MTA2-HIF-1α signaling axis using antisense oligonucleotides or specific inhibitors effectively suppresses metastasis formation, offering a therapeutic option for patients with advanced PDAC (106).
Prognostic evaluations indicate that MTA2 expression is significantly elevated in PDAC tissues compared to adjacent normal tissues (P<0.01), with a positive correlation between its expression intensity and tumor grading (105).
MTA2 in RCC
In RCC, combined targeted strategies show potential. Melatonin plus sorafenib synergistically downregulates MTA2 expression and attenuates migration and invasion of RCC cells (112). miRNA-based interventions are also feasible. Restoring miR-133b or developing its analogs can block the MTA2-MMP9 axis and inhibit RCC metastasis (111). From a prognostic perspective, MTA2 expression is significantly higher in RCC tissues than in adjacent normal tissues (P<0.01). MTA2 overexpression is associated with higher tumor grade (P=0.002) and advanced TNM stage (P<0.001), indicating its potential as a prognostic biomarker (112).
MTA2 in glioma
In miRNA-targeted therapy for glioma, restoring or mimicking miR-548b inhibits the MTA2 pathway and exerts antitumor effects. miR-548b overexpression markedly suppresses growth of U-87MG xenografts (117). In clinical samples, MTA2 expression is higher in HGG (e.g., GBM8401 and Hs683) than in low-grade lines (e.g., M059J, M059K and U-87MG). MTA2 levels positively correlate with WHO grade (P<0.001), supporting its potential as an auxiliary marker of malignancy (49).
MTA2 in osteosarcoma
In clinical treatment strategies for osteosarcoma, interventions targeting the uPA signaling pathway demonstrate potential value. It has been indicated that rh-uPA treatment of MTA2-low osteosarcoma cells significantly reverses the suppression of their migration and invasion. Correspondingly, the application of small-molecule inhibitors targeting the MTA2/uPA axis markedly reduces the number of lung metastases. Furthermore, combining these inhibitors with ERK pathway inhibitors holds promise for further enhancing the suppression of tumor metastasis (121). In the prognostic analysis of patients with osteosarcoma, elevated MTA2 expression correlated significantly with advanced tumor stages (P<0.01). In vivo experiments also confirmed that reducing MTA2 expression markedly suppressed tumor metastatic activity and the formation of pulmonary nodules (121).
Clinical implications of MTA2 in non-neoplastic diseases
Research indicates that the MTA2/HNF4A signaling axis may serve as a therapeutic target for inflammatory bowel disease and short bowel syndrome. Modulating this pathway may enable colonic epithelial cells to acquire small-intestine-like absorptive properties, thereby improving nutritional deficiencies caused by malabsorption (122). Research on male infertility has revealed that abnormalities in the FSH/MTA2/FSHR negative-feedback regulatory pathway are associated with impaired spermatogenesis. Developing drugs targeting this pathway may correct defects in the phagocytic clearance function of supporting cells, offering a therapeutic strategy for such infertility conditions (125). Additionally, the study revealed that MTA2 interacts with ERα in the epididymis through a transactivation mechanism, offering insights for developing selective ER modulators to help maintain male reproductive function (123).
It should be noted that the current understanding of MTA2 in non-neoplastic diseases remains relatively limited compared with its well-established roles in various malignant tumors. Most existing studies have concentrated on the oncogenic functions of MTA2 in cancer initiation, progression and metastasis, whereas investigations into its physiological and pathological roles in non-neoplastic conditions are still in the preliminary stage with relatively scarce clinical and translational evidence. Therefore, the coverage of non-neoplastic diseases in this review is appropriately concise, while future studies are warranted to further explore the clinical and translational potential of MTA2 in these non-oncological contexts.
In summary, MTA2 serves as a pan-cancer prognostic biomarker and a broadly actionable therapeutic target, with conserved clinical value in predicting poor differentiation, lymph node metastasis and shortened survival across most malignancies. Common therapeutic strategies include disrupting NuRD assembly, inhibiting post-translational modifications, restoring regulatory miRNAs and blocking upstream regulatory axes. At the same time, tissue-specific clinical implications are critical: Digestive system tumors show unique sensitivities to PARP or HDAC inhibition; BC exhibits endocrine therapy resistance linked to MTA2; NSCLC requires stage-specific intervention; and head and neck, bone and brain tumors benefit from miRNA-based or ceRNA-targeted strategies. These conserved and distinct features highlight that MTA2-targeted diagnosis and therapy should be tumor-type-specific and mechanism-guided to maximize clinical efficacy.
Future applications, prospects and challenges of MTA2
In conclusion, MTA2 is a central element of the NuRD complex, featuring several highly conserved functional domains, including BAH, ELM2, SANT and an atypical GATA zinc finger. It primarily achieves dual regulation of gene expression through histone deacetylation and modulation of transcription factors. Distinct from MTA1 and MTA3 with incomplete C-terminal domains, the full-set intact conserved domains equip MTA2 with the exclusive capability to assemble intact functional NuRD complex (10,141), laying the structural basis for its unique central regulatory role unavailable for other MTA homologs.
Although MTA2 exhibits consistent prognostic value across most cancer types, its clinical translation is currently hindered by substantial methodological heterogeneity. Variable detection platforms, lack of unified cut-off thresholds, inconsistent immunohistochemical scoring systems, and ambiguous subcellular localization criteria collectively limit the standardization of MTA2 as a routine clinical biomarker (142). Standardized quantitative protocols and prospective cohort validation are therefore urgently required to achieve stable clinical application.
Objectively speaking, clinical application still faces significant challenges. First, structural homology within the MTA family poses a major obstacle. MTA2 shares high sequence and domain similarity with MTA1 and MTA3, especially in the conserved BAH, ELM2 and SANT domains, making it difficult to design small molecules that selectively target MTA2 without cross-reacting with other MTA family members (10,13). Second, the lack of a well-defined druggable domain within the NuRD complex further hinders inhibitor development. From the perspective of structural biology and druggability, MTA2 possesses no intrinsic enzymatic catalytic activity and functions predominantly as a core scaffold protein. It utilizes intact BAH, ELM2, SANT and atypical GATA zinc-finger domains to mediate extensive protein-protein interactions among various subunits inside the NuRD complex. The deficiency of explicit independent ligand-binding pocket across its full-length sequence greatly restricts rational design and screening of conventional small-molecule inhibitors targeting MTA2 specifically (20). This structural feature also differentiates MTA2 from MTA1 and MTA3 with incomplete domain composition, partially explaining why selective inhibitor development against single MTA paralog remains challenging. MTA2 functions as a core scaffold subunit of the NuRD complex rather than an enzyme with a distinct active pocket; it lacks a clear ligand-binding pocket or catalytic site that can be readily targeted by conventional small-molecule inhibitors, complicating the rational design of highly selective MTA2-targeted agents. As of now, no targeted drugs for MTA2 have entered the clinical trial phase, indicating that uncertainties remain regarding the development of MTA2-specific inhibitors and rapid MTA2 detection technologies for real-time intraoperative margin assessment. It is important to note that some inconsistent or neutral findings regarding MTA2 expression and clinical outcomes have been reported in the literature. In certain tumor types and ethnic subgroups, MTA2 overexpression does not uniformly correlate with poor prognosis. For instance, in NPC, MTA2 expression shows no significant association with clinical stage or lymph node metastasis in Zhuang patients, in contrast to its clear prognostic value in Han Chinese patients. Furthermore, while MTA2 is widely linked to aggressive phenotypes and unfavorable prognosis in advanced-stage tumors across multiple cancer types, limited evidence supports a consistent prognostic role in early-stage lesions, indicating a stage-dependent functional pattern rather than a universal oncogenic effect. Importantly, such context-dependent inconsistent phenotypes are largely attributed to the competitive and dynamic substitution among MTA family members in assembling NuRD complexes. MTA1, MTA2 and MTA3 compete for limited NuRD core scaffolds to form mutually exclusive complexes, and the dominant MTA paralog determines the overall functional output of NuRD. In early-stage tumors and specific ethnic subgroups, elevated MTA1 or MTA3 can competitively replace MTA2 within NuRD, rewiring chromatin regulatory programs and offsetting MTA2-mediated oncogenic effects; in advanced malignancies, MTA2 gains dominant occupancy of NuRD complexes, driving persistent malignant progression (8). These discrepancies may arise from ethnic heterogeneity, tumor stage differences, tissue-specific regulatory contexts or variations in detection methods, highlighting the variability and inherent limitations of current evidence regarding the prognostic value of MTA2.
Therefore, while the understanding of MTA2 is gradually deepening, its clinical application will require the active participation of more high-level researchers and sustained investment from multiple biopharmaceutical companies in the future. Future research should prioritize the analysis of the competitive and context-dependent substitution patterns of MTA1, MTA2 and MTA3 across different disease contexts to precisely define the subtype-specific functions of NuRD, overcome current bottlenecks in MTA2-based clinical diagnosis and targeted therapy, and accelerate the translation of MTA2 from basic research into mature clinical biomarkers and targeted drugs.
Supplementary Data
Availability of data and materials
Not applicable.
Authors' contributions
HX contributed to writing - original draft, investigation, writing - review and editing and funding acquisition. LC was involved in investigation, methodology and writing - review and editing. JG was responsible for writing - original draft and resources. XL contributed to writing - original draft and investigation. YY was involved in writing - original draft and methodology. HH performed investigation and methodology. PW was responsible for writing - original draft and resources. QY contributed to writing - review and editing and supervision. YW was involved in writing - review and editing, supervision and conceptualization.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Abbreviations:
|
AIB1
|
amplified in breast cancer 1
|
|
BAH
|
bromo-adjacent homology
|
|
BITC
|
benzyl isothiocyanate
|
|
ceRNA
|
competing endogenous RNA
|
|
DDR
|
DNA damage response
|
|
EMT
|
epithelial-mesenchymal transition
|
|
ESCC
|
esophageal squamous cell carcinoma
|
|
EZH2
|
enhancer of zeste homolog 2
|
|
FAK
|
focal adhesion kinase
|
|
FSHR
|
follicle-stimulating hormone receptor
|
|
MMP
|
matrix metalloproteinase
|
|
MTA
|
metastasis-associated
|
|
NuRD
|
nucleosome remodeling and deacetylase
|
|
PTZN
|
phosphatase and tensin homolog deleted on chromosome ten
|
|
ROCK
|
Rho associated coiled-coil containing protein kinase
|
|
uPA
|
urokinase-type plasminogen activator
|
Acknowledgements
All authors would like to express our sincere gratitude to Dr Zhaoyun Liu from Tianjin Medical University General Hospital for his invaluable guidance and support throughout the writing of this review.
Funding
This work was supported by the Research Projects of the Jintang County Medical Association (grant no. 2024028) and the Medical Research Projects of the Chengdu Municipal Health Commission (grant no. 2025486).
References
|
1
|
Liu J, Wang H, Huang C and Qian H: Subcellular localization of MTA proteins in normal and cancer cells. Cancer Metastasis Rev. 33:843–856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qiong L, Shuyao X, Shan X, Qian F, Jiaying T, Yao X and Hui L: Recent advances in the glycolytic processes linked to tumor metastasis. Curr Mol Pharmacol. 17:e187614293083612024. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu B, Yang C, Liu J, Peng M, Mao J, Tang S and Huang W: Targeting of lysosomes as a therapeutic target in cancer. Curr Mol Pharmacol. 17:e187614293546592024. View Article : Google Scholar
|
|
4
|
Ning Z, Gan J, Chen C, Zhang D and Zhang H: Molecular functions and significance of the MTA family in hormone-independent cancer. Cancer Metastasis Rev. 33:901–919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li DQ, Pakala SB, Nair SS, Eswaran J and Kumar R: Metastasis-associated protein 1/nucleosome remodeling and histone deacetylase complex in cancer. Cancer Res. 72:387–394. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xue Y, Wong J, Moreno GT, Young MK, Côté J and Wang W: NURD, a novel complex with both ATP-dependent chromatin-remodeling and histone deacetylase activities. Mol Cell. 2:851–861. 1998. View Article : Google Scholar
|
|
7
|
Zhang Y, Ng HH, Erdjument-Bromage H, Tempst P, Bird A and Reinberg D: Analysis of the NuRD subunits reveals a histone deacetylase core complex and a connection with DNA methylation. Genes Dev. 13:1924–1935. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yao YL and Yang WM: The metastasis-associated proteins 1 and 2 form distinct protein complexes with histone deacetylase activity. J Biol Chem. 278:42560–42568. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chrysovergis K, Headley K, Harper KM, Moy SS, Grimm SA, Jefferson WN, Sifre MI, Mahapatra D, Rodriguez Y, Williams CJ and Wade PA: A novel mutant allele of Mta3 in the mouse: genetic analysis of roles in immunity and androgen biology. G3 (Bethesda). 15:jkaf1712025. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Manavathi B and Kumar R: Metastasis tumor antigens, an emerging family of multifaceted master coregulators. J Biol Chem. 282:1529–1533. 2007. View Article : Google Scholar
|
|
11
|
Covington KR and Fuqua SA: Role of MTA2 in human cancer. Cancer Metastasis Rev. 33:921–928. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qin Z and Xu Y: Dexmedetomidine alleviates brain ischemia/reperfusion injury by regulating metastasis-associated lung adenocarcinoma transcript 1/MicroRNA-140-5p/nuclear factor erythroid-derived 2-like 2 axis. Protein Pept Lett. 31:116–127. 2024. View Article : Google Scholar
|
|
13
|
Toh Y and Nicolson GL: The role of the MTA family and their encoded proteins in human cancers: Molecular functions and clinical implications. Clin Exp Metastasis. 26:215–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fujita N, Jaye DL, Kajita M, Geigerman C, Moreno CS and Wade PA: MTA3, a Mi-2/NuRD complex subunit, regulates an invasive growth pathway in breast cancer. Cell. 113:207–219. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Barone I, Brusco L, Gu G, Selever J, Beyer A, Covington KR, Tsimelzon A, Wang T, Hilsenbeck SG, Chamness GC, et al: Loss of Rho GDIα and resistance to tamoxifen via effects on estrogen receptor α. J Natl Cancer Inst. 103:538–552. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Covington KR, Brusco L, Barone I, Tsimelzon A, Selever J, Corona-Rodriguez A, Brown P, Kumar R, Hilsenbeck SG and Fuqua SA: Metastasis tumor-associated protein 2 enhances metastatic behavior and is associated with poor outcomes in estrogen receptor-negative breast cancer. Breast Cancer Res Treat. 141:375–384. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fujita N, Kajita M, Taysavang P and Wade PA: Hormonal regulation of metastasis-associated protein 3 transcription in breast cancer cells. Mol Endocrinol. 18:2937–2949. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang H, Stephens LC and Kumar R: Metastasis tumor antigen family proteins during breast cancer progression and metastasis in a reliable mouse model for human breast cancer. Clin Cancer Res. 12:1479–1486. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Araghi M, Gharebakhshi F, Faramarzi F, Mafi A, Mousavi T, Alimohammadi M and Soleimantabar H: Efficacy and safety of pembrolizumab monotherapy or combined therapy in patients with metastatic triple-negative breast cancer: A systematic review and meta-analysis of randomized controlled trials. Curr Gene Ther. 25:72–88. 2025. View Article : Google Scholar
|
|
20
|
Liu X, Jiang Y, Hou Y, Li X, Li H and Si W: The expression of metastasis associated protein 2 in normal development and cancers: Mechanism and clinical significance. PeerJ. 13:e201072025. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
UniProt Consortium: Update on activities at the Universal Protein Resource (UniProt) in 2013. Nucleic Acids Res. 41(Database Issue): D43–D47. 2013.
|
|
22
|
Dephoure N, Zhou C, Villén J, Beausoleil SA, Bakalarski CE, Elledge SJ and Gygi SP: A quantitative atlas of mitotic phosphorylation. Proc Natl Acad Sci USA. 105:10762–10767. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mayya V, Lundgren DH, Hwang SI, Rezaul K, Wu L, Eng JK, Rodionov V and Han DK: Quantitative phosphoproteomic analysis of T cell receptor signaling reveals system-wide modulation of protein-protein interactions. Sci Signal. 2:ra462009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Olsen JV, Vermeulen M, Santamaria A, Kumar C, Miller ML, Jensen LJ, Gnad F, Cox J, Jensen TS, Nigg EA, et al: Quantitative phosphoproteomics reveals widespread full phosphorylation site occupancy during mitosis. Sci Signal. 3:ra32010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rigbolt KT, Prokhorova TA, Akimov V, Henningsen J, Johansen PT, Kratchmarova I, Kassem M, Mann M, Olsen JV and Blagoev B: System-wide temporal characterization of the proteome and phosphoproteome of human embryonic stem cell differentiation. Sci Signal. 4:rs32011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choudhary C, Kumar C, Gnad F, Nielsen ML, Rehman M, Walther TC, Olsen JV and Mann M: Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science. 325:834–840. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo J, Su F, Chen D, Shiloh A and Gu W: Deacetylation of p53 modulates its effect on cell growth and apoptosis. Nature. 408:377–381. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cui Y, Niu A, Pestell R, Kumar R, Curran EM, Liu Y and Fuqua SA: Metastasis-associated protein 2 is a repressor of estrogen receptor alpha whose overexpression leads to estrogen-independent growth of human breast cancer cells. Mol Endocrinol. 20:2020–2035. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar R, Wang RA and Bagheri-Yarmand R: Emerging roles of MTA family members in human cancers. Semin Oncol. 30(5 Suppl 16): S30–S37. 2003. View Article : Google Scholar
|
|
30
|
Bowen NJ, Fujita N, Kajita M and Wade PA: Mi-2/NuRD: Multiple complexes for many purposes. Biochim Biophys Acta. 1677:52–57. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lai AY and Wade PA: Cancer biology and NuRD: A multifaceted chromatin remodelling complex. Nat Rev Cancer. 11:588–596. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu X, Kovalev GI, Chang H, Kallin E, Knudsen G, Xia L, Mishra N, Ruiz P, Li E, Su L and Zhang Y: Inactivation of NuRD component Mta2 causes abnormal T cell activation and lupus-like autoimmune disease in mice. J Biol Chem. 283:13825–13833. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hosokawa H, Tanaka T, Suzuki Y, Iwamura C, Ohkubo S, Endoh K, Kato M, Endo Y, Onodera A, Tumes DJ, et al: Functionally distinct Gata3/Chd4 complexes coordinately establish T helper 2 (Th2) cell identity. Proc Natl Acad Sci USA. 110:4691–4696. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moon HE, Cheon H, Chun KH, Lee SK, Kim YS, Jung BK, Park JA, Kim SH, Jeong JW and Lee MS: Metastasis-associated protein 1 enhances angiogenesis by stabilization of HIF-1alpha. Oncol Rep. 16:929–935. 2006.PubMed/NCBI
|
|
35
|
Singh RR and Kumar R: MTA family of transcriptional meta-regulators in mammary gland morphogenesis and breast cancer. J Mammary Gland Biol Neoplasia. 12:115–125. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei FZ, Cao Z, Wang X, Wang H, Cai MY, Li T, Hattori N, Wang D, Du Y, Song B, et al: Epigenetic regulation of autophagy by the methyltransferase EZH2 through an MTOR-dependent pathway. Autophagy. 11:2309–2322. 2015. View Article : Google Scholar
|
|
37
|
Zeng Z, Xu FY, Zheng H, Cheng P, Chen QY, Ye Z, Zhong JX, Deng SJ, Liu ML, Huang K, et al: LncRNA-MTA2TR functions as a promoter in pancreatic cancer via driving deacetylation-dependent accumulation of HIF-1α. Theranostics. 9:5298–5314. 2019. View Article : Google Scholar
|
|
38
|
Smeenk G, Wiegant WW, Vrolijk H, Solari AP, Pastink A and van Attikum H: The NuRD chromatin-remodeling complex regulates signaling and repair of DNA damage. J Cell Biol. 190:741–749. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Futamura M, Nishimori H, Shiratsuchi T, Saji S, Nakamura Y and Tokino T: Molecular cloning, mapping, and characterization of a novel human gene, MTA1-L1, showing homology to a metastasis-associated gene, MTA1. J Hum Genet. 44:52–56. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhou LH, Gu LF, Chen YX, Jing P, Yang TT, He Y, Bao YL, Zhang XZ, Du C, Wang SB, et al: HMGB2 promotes cardiomyocyte proliferation and heart regeneration through MTA2-Driven metabolic reprogramming. Adv Sci (Weinh). 13:e058202026. View Article : Google Scholar :
|
|
41
|
Matsusue K, Takiguchi S, Toh Y and Kono A: Characterization of mouse metastasis-associated gene 2: Genomic structure, nuclear localization signal, and alternative potentials as transcriptional activator and repressor. DNA Cell Biol. 20:603–611. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee H, Ryu SH, Hong SS, Seo DD, Min HJ, Jang MK, Kwon HJ, Yu E, Chung YH and Kim KW: Overexpression of metastasis-associated protein 2 is associated with hepatocellular carcinoma size and differentiation. J Gastroenterol Hepatol. 24:1445–1450. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Park HR, Jung WW, Kim HS, Bacchini P, Bertoni F and Park YK: Overexpression of metastatic tumor antigen in osteosarcoma: Comparison between conventional high-grade and central low-grade osteosarcoma. Cancer Res Treat. 37:360–364. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu SL, Han Y, Zhang Y, Xie CY, Wang EH, Miao Y, Li HY, Xu HT and Dai SD: Expression of metastasis-associated protein 2 (MTA2) might predict proliferation in non-small cell lung cancer. Target Oncol. 7:135–143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang Y, Li L, Li Q, Xie C and Wang E and Wang E: Expression of P120 catenin, Kaiso, and metastasis tumor antigen-2 in thymomas. Tumour Biol. 33:1871–1879. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu YP, Shan BE, Wang XL and Ma L: Correlation between MTA2 overexpression and tumour progression in esophageal squamous cell carcinoma. Exp Ther Med. 3:745–749. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen DW, Fan YF, Li J and Jiang XX: MTA2 expression is a novel prognostic marker for pancreatic ductal adenocarcinoma. Tumour Biol. 34:1553–1557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhou C, Ji J, Cai Q, Shi M, Chen X, Yu Y, Liu B, Zhu Z and Zhang J: MTA2 promotes gastric cancer cells invasion and is transcriptionally regulated by Sp1. Mol Cancer. 12:1022013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cheng CY, Chou YE, Ko CP, Yang SF, Hsieh SC, Lin CL, Hsieh YH and Chen KC: Metastasis tumor-associated protein-2 knockdown suppresses the proliferation and invasion of human glioma cells in vitro and in vivo. J Neurooncol. 120:273–281. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 74:229–263. 2024.PubMed/NCBI
|
|
51
|
Giaquinto AN, Sung H, Miller KD, Kramer JL, Newman LA, Minihan A, Jemal A and Siegel RL: Breast cancer statistics, 2022. CA Cancer J Clin. 72:524–541. 2022.PubMed/NCBI
|
|
52
|
Loibl S, Poortmans P, Morrow M, Denkert C and Curigliano G: Breast cancer. Lancet. 397:1750–1769. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Veronesi U, Boyle P, Goldhirsch A, Orecchia R and Viale G: Breast cancer. Lancet. 365:1727–1741. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang X, Zou G, Li X, Wang L, Xie T, Zhao J, Wang L, Jiao S, Xiang R, Ye H and Shi Y: An isoflavone derivative potently inhibits the angiogenesis and progression of triple-negative breast cancer by targeting the MTA2/SerRS/VEGFA pathway. Cancer Biol Med. 17:693–706. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhang L, Wang Q, Zhou Y, Ouyang Q, Dai W, Chen J, Ding P, Li L, Zhang X, Zhang W, et al: Overexpression of MTA1 inhibits the metastatic ability of ZR-75-30 cells in vitro by promoting MTA2 degradation. Cell Commun Signal. 17:42019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Duan Y, Xu Y, Dou Y and Xu D: Helicobacter pylori and gastric cancer: Mechanisms and new perspectives. J Hematol Oncol. 18:102025. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI
|
|
58
|
Smyth EC, Nilsson M, Grabsch HI, van Grieken NC and Lordick F: Gastric cancer. Lancet. 396:635–648. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li A, Guo Y, Yin Z, Liu X and Xie G: MTA2 is one of 14 Transcription factors predicting recurrence free survival in gastric cancer and promotes cancer progression by targeting MCM5. J Cancer. 14:262–274. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhou C, Ji J, Cai Q, Shi M, Chen X, Yu Y, Zhu Z and Zhang J: MTA2 enhances colony formation and tumor growth of gastric cancer cells through IL-11. BMC Cancer. 15:3432015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shi J, Zhang X, Li J, Huang W, Wang Y, Wang Y and Qin J: MTA2 sensitizes gastric cancer cells to PARP inhibition by induction of DNA replication stress. Transl Oncol. 14:1011672021. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xie G, Lei B, Yin Z, Xu F and Liu X: CircMTA2 drives gastric cancer progression through suppressing MTA2 degradation via interacting with UCHL3. Int J Mol Sci. 25:28172024. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhao L, Guo H, Zhou B, Feng J, Li Y, Han T, Liu L, Li L, Zhang S, Liu Y, et al: Long non-coding RNA SNHG5 suppresses gastric cancer progression by trapping MTA2 in the cytosol. Oncogene. 35:5770–5780. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
No authors listed. Hepatocellular carcinoma. Nat Rev Dis Primers. 7:72021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Nagaraju GP, Dariya B, Kasa P, Peela S and El-Rayes BF: Epigenetics in hepatocellular carcinoma. Semin Cancer Biol. 86:622–632. 2022. View Article : Google Scholar
|
|
66
|
Foglia B, Turato C and Cannito S: Hepatocellular Carcinoma: Latest research in pathogenesis, detection and treatment. Int J Mol Sci. 24:122242023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang H, Chang Z, Qin LN, Liang B, Han JX, Qiao KL, Yang C, Liu YR, Zhou HG and Sun T: MTA2 triggered R-loop trans-regulates BDH1-mediated β-hydroxybutyrylation and potentiates propagation of hepatocellular carcinoma stem cells. Signal Transduct Target Ther. 6:1352021. View Article : Google Scholar
|
|
68
|
Guan C, Chang Z, Gu X and Liu R: MTA2 promotes HCC progression through repressing FRMD6, a key upstream component of hippo signaling pathway. Biochem Biophys Res Commun. 515:112–118. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Siegel RL, Miller KD and Jemal A: Cancer statistics, 2018. CA Cancer J Clin. 68:7–30. 2018.PubMed/NCBI
|
|
70
|
Siegel RL, Miller KD and Jemal A: Cancer Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017.PubMed/NCBI
|
|
71
|
Henley SJ, Singh SD, King J, Wilson R, O'Neil ME and Ryerson AB; Centers for Disease Control and Prevention (CDC): Invasive cancer incidence and survival-United States, 2011. MMWR Morb Mortal Wkly Rep. 64:237–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ferlay J, Colombet M, Soerjomataram I, Mathers C, Parkin DM, Piñeros M, Znaor A and Bray F: Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int J Cancer. 144:1941–1953. 2019. View Article : Google Scholar
|
|
73
|
Zhou J, Zhan S, Tan W, Cheng R, Gong H and Zhu Q: P300 binds to and acetylates MTA2 to promote colorectal cancer cells growth. Biochem Biophys Res Commun. 444:387–390. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Uraki S, Sugimoto K, Shiraki K, Tameda M, Inagaki Y, Ogura S, Kasai C, Nojiri K, Yoneda M, Yamamoto N, et al: Human β-defensin-3 inhibits migration of colon cancer cells via downregulation of metastasis-associated 1 family, member 2 expression. Int J Oncol. 45:1059–1064. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Olusola P, Banerjee HN, Philley JV and Dasgupta S: Human papilloma virus-associated cervical cancer and health disparities. Cells. 8:6222019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Arbyn M, Weiderpass E, Bruni L, de Sanjosé S, Saraiya M, Ferlay J and Bray F: Estimates of incidence and mortality of cervical cancer in 2018: A worldwide analysis. Lancet Glob Health. 8:e191–e203. 2020. View Article : Google Scholar :
|
|
77
|
Lin CL, Ying TH, Yang SF, Wang SW, Cheng SP, Lee JJ and Hsieh YH: Transcriptional suppression of miR-7 by MTA2 Induces Sp1-Mediated KLK10 expression and metastasis of cervical cancer. Mol Ther Nucleic Acids. 20:699–710. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wang L, Chen G, Zhou C, Wu C and Jiang J: Expression and significance of MTA2 and CPNE1 in cervical squamous cell carcinoma. Appl Immunohistochem Mol Morphol. 31:569–573. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA Cancer J Clin. 65:87–108. 2015.PubMed/NCBI
|
|
80
|
Nasim F, Sabath BF and Eapen GA: Lung cancer. Med Clin North Am. 103:463–473. 2019. View Article : Google Scholar
|
|
81
|
Molina JR, Yang P, Cassivi SD, Schild SE and Adjei AA: Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594. 2008. View Article : Google Scholar
|
|
82
|
Zhang B, Tao F and Zhang H: Metastasis-associated protein 2 promotes the metastasis of non-small cell lung carcinoma by regulating the ERK/AKT and VEGF signaling pathways. Mol Med Rep. 17:4899–4908. 2018.PubMed/NCBI
|
|
83
|
El-Nikhely N, Karger A, Sarode P, Singh I, Weigert A, Wietelmann A, Stiewe T, Dammann R, Fink L, Grimminger F, et al: Metastasis-associated protein 2 represses NF-κB to reduce lung tumor growth and inflammation. Cancer Res. 80:4199–4211. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang B, Zhang H and Shen G: Metastasis-associated protein 2 (MTA2) promotes the metastasis of non-small-cell lung cancer through the inhibition of the cell adhesion molecule Ep-CAM and E-cadherin. Jpn J Clin Oncol. 45:755–766. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dunn LA, Ho AL and Pfister DG: Head and neck cancer: A review. JAMA. 335:531–541. 2026. View Article : Google Scholar
|
|
86
|
Johnson DE, Burtness B, Leemans CR, Lui VWY, Bauman JE and Grandis JR: Head and neck squamous cell carcinoma. Nat Rev Dis Primers. 6:922020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhou J, Hu Z, Wang L, Hu Q, Chen Z, Lin T, Zhou R, Cai Y, Wu Z, Zhang Z, et al: Tumor-colonized Streptococcus mutans metabolically reprograms tumor microenvironment and promotes oral squamous cell carcinoma. Microbiome. 12:1932024. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Chinn SB and Myers JN: Oral cavity carcinoma: Current management, controversies, and future directions. J Clin Oncol. 33:3269–3276. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Tseng TY, Chiou HL, Lin CW, Chen YS, Hsu LS, Lee CH and Hsieh YH: Repression of metastasis-associated protein 2 for inhibiting metastasis of human oral cancer cells by promoting the p-cofilin-1/LC3-II expression. J Oral Pathol Med. 48:959–966. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Tao D, Zhang Z, Liu X, Zhang Z, Fu Y, Zhang P, Yuan H, Liu L, Cheng J and Jiang H: LncRNA HOTAIR promotes the invasion and metastasis of oral squamous cell carcinoma through metastasis-associated gene 2. Mol Carcinog. 59:353–364. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Wang Z, Fang M, Zhang J, Tang L, Zhong L, Li H, Cao R, Zhao X, Liu S, Zhang R, et al: Radiomics and deep learning in nasopharyngeal carcinoma: A review. IEEE Rev Biomed Eng. 17:118–135. 2024. View Article : Google Scholar
|
|
92
|
Chen YP, Chan ATC, Le QT, Blanchard P, Sun Y and Ma J: Nasopharyngeal carcinoma. Lancet. 394:64–80. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wu M, Ye X, Wang S, Li Q, Lai Y and Yi Y: MicroRNA-148b suppresses proliferation, migration, and invasion of nasopharyngeal carcinoma cells by targeting metastasis-associated gene 2. Onco Targets Ther. 10:2815–2822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang BJ, Lan GP, Si JY, Li YL, Huang B, Deng ZX, Si YF, Chen MM, Shen XY and Wang Y: Correlation of metastasis-associated protein expression with prognosis and chemotherapy in nasopharyngeal carcinoma. Int J Clin Exp Pathol. 11:2537–2549. 2018.PubMed/NCBI
|
|
95
|
Global Burden of Disease Cancer Collaboration; Fitzmaurice C, Dicker D, Pain A, Hamavid H, Moradi-Lakeh M, MacIntyre MF, Allen C, Hansen G, Woodbrook R, et al: The global burden of cancer 2013. JAMA Oncol. 1:505–527. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Ferlay J, Shin HR, Bray F, Forman D, Mathers C and Parkin DM: Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
97
|
Day NE: Some aspects of the epidemiology of esophageal cancer. Cancer Res. 35(11 Pt. 2): 3304–3307. 1975.PubMed/NCBI
|
|
98
|
Codipilly DC and Wang KK: Squamous cell carcinoma of the esophagus. Gastroenterol Clin North Am. 51:457–484. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Dai SL, Wei SS, Zhang C, Li XY, Liu YP, Ma M, Lv HL, Zhang Z, Zhao LM and Shan BE: MTA2 promotes the metastasis of esophageal squamous cell carcinoma via EIF4E-Twist feedback loop. Cancer Sci. 112:1060–1074. 2021. View Article : Google Scholar :
|
|
100
|
Wei S, Sun S, Zhou X, Zhang C, Li X, Dai S, Wang Y, Zhao L and Shan B: SNHG5 inhibits the progression of EMT through the ubiquitin-degradation of MTA2 in oesophageal cancer. Carcinogenesis. 42:315–326. 2021. View Article : Google Scholar
|
|
101
|
Thiel V, Renders S, Panten J, Dross N, Bauer K, Azorin D, Henriques V, Vogel V, Klein C, Leppä AM, et al: Characterization of single neurons reprogrammed by pancreatic cancer. Nature. 640:1042–1051. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ferlay J, Partensky C and Bray F: More deaths from pancreatic cancer than breast cancer in the EU by 2017. Acta Oncol. 55:1158–1160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and deaths to 2030: the unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 74:2913–2921. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Kwak M, Hua TC, Jin H, Lee J and Kim DE: AS1411 aptamer-conjugated liposomal siRNA Targeting MTA2 Suppresses PI3K/AKT signaling in pancreatic cancer cells. Int J Mol Sci. 26:84672025. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Si W, Liu X, Wei R, Zhang Y, Zhao Y, Cui L and Hong T: MTA2-mediated inhibition of PTEN leads to pancreatic ductal adenocarcinoma carcinogenicity. Cell Death Dis. 10:2062019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhu S, Deng S, He C, Liu M, Chen H, Zeng Z, Zhong J, Ye Z, Deng S, Wu H, et al: Reciprocal loop of hypoxia-inducible factor-1α (HIF-1α) and metastasis-associated protein 2 (MTA2) contributes to the progression of pancreatic carcinoma by suppressing E-cadherin transcription. J Pathol. 245:349–360. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Gupta K, Miller JD, Li JZ, Russell MW and Charbonneau C: Epidemiologic and socioeconomic burden of metastatic renal cell carcinoma (mRCC): A literature review. Cancer Treat Rev. 34:193–205. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Chow WH, Devesa SS, Warren JL and Fraumeni JF Jr: Rising incidence of renal cell cancer in the United States. JAMA. 281:1628–1631. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hollingsworth JM, Miller DC, Daignault S and Hollenbeck BK: Rising incidence of small renal masses: A need to reassess treatment effect. J Natl Cancer Inst. 98:1331–1334. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Hollingsworth JM, Miller DC, Daignault S and Hollenbeck BK: Five-year survival after surgical treatment for kidney cancer: A population-based competing risk analysis. Cancer. 109:1763–1768. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chen YS, Hung TW, Su SC, Lin CL, Yang SF, Lee CC, Yeh CF, Hsieh YH and Tsai JP: MTA2 as a potential biomarker and its involvement in metastatic progression of human renal cancer by miR-133b targeting MMP-9. Cancers (Basel). 11:18512019. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Lee CC, Huang PY, Hsieh YH, Chen YS and Tsai JP: Melatonin combined with sorafenib synergistically inhibit the invasive ability through targeting metastasis-associated protein 2 expression in human renal cancer cells. Tzu Chi Med J. 34:192–199. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Xu S, Tang L, Li X, Fan F and Liu Z: Immunotherapy for glioma: Current management and future application. Cancer Lett. 476:1–12. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Ostrom QT, Gittleman H, Truitt G, Boscia A, Kruchko C and Barnholtz-Sloan JS: CBTRUS statistical report: Primary brain and other central nervous system tumors diagnosed in the United States in 2011-2015. Neuro Oncol. 20(Suppl 4): iv1–iv86. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhang N, Zhang L, Qiu B, Meng L, Wang X and Hou BL: Correlation of volume transfer coefficient Ktrans with histopathologic grades of gliomas. J Magn Reson Imaging 2012. 36:355–363. 2012. View Article : Google Scholar
|
|
116
|
Louis DN, Perry A, Reifenberger G, von Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD, Kleihues P and Ellison DW: The 2016 World health organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Pan Y, Liang W, Zhao X, Liu L, Qing Y and Li Y: miR-548b inhibits the proliferation and invasion of malignant gliomas by targeting metastasis tumor-associated protein-2. Neuroreport. 27:1266–1273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Arndt CA and Crist WM: Common musculoskeletal tumors of childhood and adolescence. N Engl J Med. 341:342–352. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Ritter J and Bielack SS: Osteosarcoma. Ann Oncol. 21(Suppl 7): vii320–vii325. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Kansara M, Teng MW, Smyth MJ and Thomas DM: Translational biology of osteosarcoma. Nat Rev Cancer. 14:722–735. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Tseng C, Chen CM, Hsieh YH, Lin CY, Chen JW, Hsiao PH, Fong YC, Wang PH, Chen PN and Lin RC: MTA2 knockdown suppresses human osteosarcoma metastasis by inhibiting uPA expression. Aging (Albany NY). 16:12239–12251. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Gu W, Huang X, Singh PNP, Li S, Lan Y, Deng M, Lacko LA, Gomez-Salinero JM, Rafii S, Verzi MP, et al: A MTA2-SATB2 chromatin complex restrains colonic plasticity toward small intestine by retaining HNF4A at colonic chromatin. Nat Commun. 15:35952024. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Cheng P, Zhao J, Qiao H, Ma H, Ma B, Wei J, Li Y and Li Z: Colocalization of metastasis-associated proteins 1/2 and estrogen receptor alpha in rat epididymis. Tissue Cell. 49:582–588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Zhang S, Li W, Zhu C, Wang X, Li Z, Zhang J, Zhao J, Hu J, Li T and Zhang Y: Sertoli cell-specific expression of metastasis-associated protein 2 (MTA2) is required for transcriptional regulation of the follicle-stimulating hormone receptor (FSHR) gene during spermatogenesis. J Biol Chem. 287:40471–40483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhu CJ, Zhang S, Liang Y and Li W: Elicitation of metastasis associated protein 2 expression in the phagocytosis by murine testicular Sertoli cells. Biochem Biophys Res Commun. 445:667–672. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Lu X, Chu CS, Fang T, Rayon-Estrada V, Fang F, Patke A, Qian Y, Clarke SH, Melnick AM, Zhang Y, et al: MTA2/NuRD Regulates B cell development and cooperates with OCA-B in controlling the Pre-B to immature B cell transition. Cell Rep. 28:472–485.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
You Y, Cui Y, Li Y, Zhang M and Wang X, Ji J, Zhang X, Zhou M, Zhang Z, Ye SD and Wang X: Inhibition of MTA2 and MTA3 induces mesendoderm specification of human embryonic stem cells. Biochem Biophys Res Commun. 552:142–149. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Varešlija D, Ward E, Purcell SP, Cosgrove NS, Cocchiglia S, O'Halloran PJ, Charmsaz S, Bane FT, Brett FM, Farrell M, et al: Comparative analysis of the AIB1 interactome in breast cancer reveals MTA2 as a repressive partner which silences E-Cadherin to promote EMT and associates with a pro-metastatic phenotype. Oncogene. 40:1318–1331. 2021. View Article : Google Scholar
|
|
129
|
Zhou Z, Yang P, Zhang B, Yao M, Jia Y, Li N, Liu H, Bai H and Gong X: Long Noncoding RNA TTC39A-AS1 promotes breast cancer tumorigenicity by sponging MicroRNA-483-3p and thereby upregulating MTA2. Pharmacology. 106:573–587. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Chu J: MicroRNA-589 serves as a tumor suppressor microRNA through directly targeting metastasis-associated protein 2 in breast cancer. Oncol Lett. 18:2232–2239. 2019.PubMed/NCBI
|
|
131
|
Lu J and Jin ML: Short-hairpin RNA-mediated MTA2 silencing inhibits human breast cancer cell line MDA-MB231 proliferation and metastasis. Asian Pac J Cancer Prev. 15:5577–5582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Zhao L, Guo H, Zhou B, Feng J, Li Y, Han T, Liu L, Li L, Zhang S, Liu Y, et al: Retraction note: Long non-coding RNA SNHG5 suppresses gastric cancer progression by trapping MTA2 in the cytosol. Oncogene. 45:14002026. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
An JX, Ma MH, Zhang CD, Shao S, Zhou NM and Dai DQ: miR-1236-3p inhibits invasion and metastasis in gastric cancer by targeting MTA2. Cancer Cell Int. 18:662018. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Jiang Z, Sun X, Zhang Q, Ji X, Yu Q, Huang T, Chen D, Chen H, Mei X, Wang L, et al: Identification of candidate biomarkers that involved in the epigenetic transcriptional regulation for detection gastric cancer by iTRAQ based quantitative proteomic analysis. Clin Chim Acta. 471:29–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Huang X, Tan J, Chen M, Zheng W, Zou S, Ye X, Li Y and Wu M: Prognostic, immunological, and mutational analysis of MTA2 in pan-cancer and drug screening for hepatocellular carcinoma. Biomolecules. 13:8832023. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Hu HM, Lee HL, Liu CJ, Hsieh YH, Chen YS and Hsueh KC: Loss of MTA2-mediated downregulation of PTK7 inhibits hepatocellular carcinoma metastasis progression by modulating the FAK-MMP7 axis. Environ Toxicol. 39:1897–1908. 2024. View Article : Google Scholar
|
|
137
|
Zhang S, Zhu K, Han Q, Wang Q and Yang B: lncRNA PCAT1 might coordinate ZNF217 to promote CRC adhesion and invasion through regulating MTA2/MTA3/Snai1/E-cadherin signaling. Cell Mol Biol (Noisy-le-grand). 67:1–9. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Lin CL, Ying TH, Yang SF, Chiou HL, Chen YS, Kao SH and Hsieh YH: MTA2 silencing attenuates the metastatic potential of cervical cancer cells by inhibiting AP1-mediated MMP12 expression via the ASK1/MEK3/p38/YB1 axis. Cell Death Dis. 12:4512021. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Burd EM: Human papillomavirus and cervical cancer. Clin Microbiol Rev. 16:1–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Xiao L, Huang S, Cheng H and Chen J: Expression of metastasis tumor-associated protein 2 in cervical squamous carcinoma and its relationship with prognosis. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 41:1031–1038. 2016.In Chinese. PubMed/NCBI
|
|
141
|
Torchy MP, Hamiche A and Klaholz BP: Structure and function insights into the NuRD chromatin remodeling complex. Cell Mol Life Sci. 72:2491–2507. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Ludwig JA and Weinstein JN: Biomarkers in cancer staging, prognosis and treatment selection. Nat Rev Cancer. 5:845–856. 2005. View Article : Google Scholar : PubMed/NCBI
|