Fibroblast growth factor receptor (FGFR)1 has
recently emerged as a critical regulator of bone remodeling in RA
pathogenesis (13-20). FGFR1 is widely expressed in bone,
cartilage and synovial tissue, where it regulates skeletal
development, remodeling and inflammatory responses (21-26). Dysregulation of FGFR1 signaling
is implicated in various pathological conditions, including RA,
where excessive activation contributes to synovial hyperplasia,
cartilage degradation and bone erosion (14,27-29). FGFR1, activated by FGFs, signals
through key downstream effectors including MAPK, PI3K/AKT and
phospholipase Cγ (PLCγ) (30,31). Dysregulated FGFR1 signaling has
been implicated in the aberrant bone remodeling central to RA
(32).
The present review aimed to summarize the structural
biology of FGFR1, its multifaceted roles in bone homeostasis and
its pathological contributions to RA-associated joint damage, as
well as FGFR1 as a therapeutic target and the translational
potential of FGFR1-modulating strategies for RA treatment. The
present study aimed to provide a framework for developing targeted
interventions to prevent structural progression in RA and
associated osteolytic disorders.
FGFRs are widely distributed, highly conserved
transmembrane tyrosine kinase (TK) receptors (33). FGFR adopts a bilobal kinase
architecture defined by an N- and a C-terminal lobe. These two
structural domains form a catalytic groove that accommodates both
ATP and peptide substrates. This serves an important role in the
receptor kinase activity and regulation (34). FGFR1 is one of four main members
of the FGFR family, which includes FGFR1, FGFR2, FGFR3 and FGFR4.
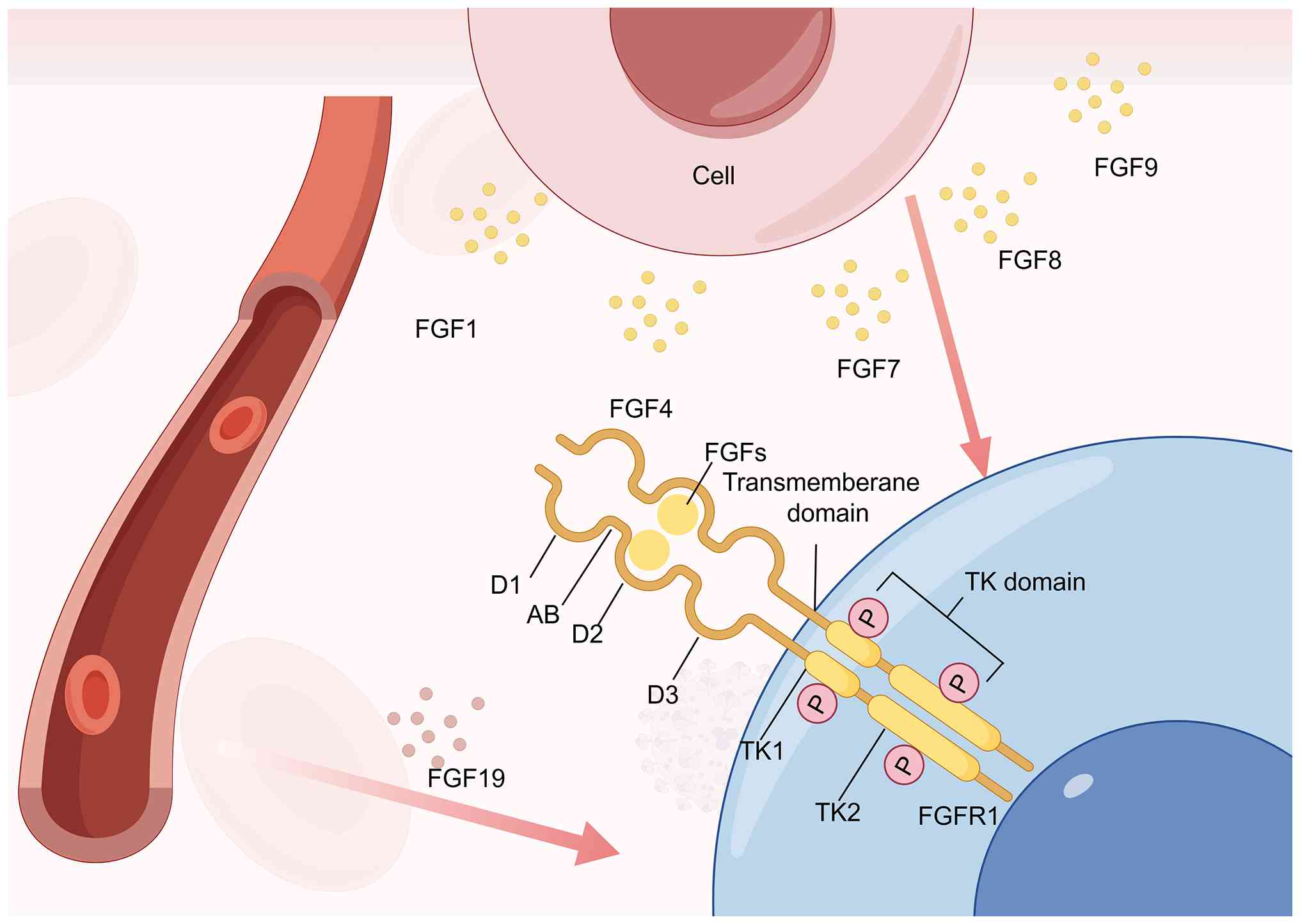
Located at 8p11.23 and comprising 68,620 bases (35). FGFR1 is a cell surface TK
receptor. FGFR1 comprises a triad of extracellular Ig-like modules
(D1-D3), followed by a transmembrane α-helix and a pair of
cytoplasmic TK units, TK1 and TK2. Numerous junctional regions are
interspersed between these domains, playing a crucial role in
maintaining the structural integrity of FGFR1 (36). Notably, the linkage between D1
and D2 is facilitated by a short sequence consisting of 7-8 amino
acids. Composed of a stretch rich in acidic and serine residues,
the acid box cooperates with Ig-like domain 1 to impose an
autoinhibitory constraint on FGFR (37,38) (Fig. 1). D2 has a heparan sulfate
(HS)-binding site, consisting of a conserved positively charged
region (39). The D2 and the D3
region are key for ligand binding and specificity (40). FGFR1-3 undergo alternative
splicing, generating b and c isoforms that modulate ligand
specificity through structural alterations in the ligand-binding
region (41-43). This regulatory mechanism is
achieved by modifying the primary amino acid sequence within the
C-terminal half of immunoglobulin-like domain D3, a key determinant
of ligand-receptor interactions. Such isoform-specific variations
in the extracellular domain enable differential recognition of
FGFs, thereby regulating signaling responses in a tissue- and
context-dependent manner (44,45). A fifth non-TK FGFR, FGFR5, is
homologous to FGFR1-4 in the extracellular ligand-binding domain,
but does not have an intracellular TK domain. FGFR5 was first
discovered in human cartilage (46). The four FGFRs exert protein TK
activity in the cell, while FGFR5 has a short non-enzymatic
intracellular element of 105 amino acids (47). FGFR5 was originally hypothesized
to function as a non-signaling decoy receptor, exerting an
inhibitory effect on the classical FGFR signaling pathway (48). This regulatory role is
hypothesized to be mediated through competitive binding to ligands,
thereby decreasing their availability for interaction with
canonical FGFRs and modulating downstream signaling responses
(49,50). FGFR5 not only serves as a
negative regulator but also actively engages in the canonical FGFR
signaling pathway (48,51-53). This dual role includes mediating
key biological functions such as the promotion of cell
differentiation, suggesting a more complex regulatory mechanism.
These findings highlight the context-dependent nature of FGFR5
activity, where its interaction with canonical FGFR components can
inhibit or enhance downstream signaling outcomes depending on cell
context and ligand availability. The expression of FGFR5 increases
with FGFR2 and FGFR1 when mesenchymal cells differentiate into
osteoblasts or adipocytes (54).
In β cells of the pancreas, FGFR5 interacts with SHP-1 and promotes
ERK1/2 signaling (55). FGFR5 is
inflammation-inducible and serves as a FGFR1 coreceptor to enhance
survival signaling (52).
Deletion of FGFR1 in osteocytes leads to an increase in trabecular
bone mass and cortical bone thickness, which is associated with
increased bone formation and impaired bone resorption (56). FGFR1 can regulate bone mass in
osteocytes by modulating Wnt/β-catenin signaling (35). This provides genetic evidence
that FGFR1 serves a crucial role in bone cells during bone
remodeling and suggests FGFR1 may be a potential therapeutic target
for preventing bone loss (35).
Activation of the FGFR1 signaling pathway is implicated in various
chronic inflammatory diseases, including RA (14,36,52,57).
FGFR1-3 use alternative splicing to generate
isoforms with distinct ligand-binding properties, exhibiting
lineage-specific expression patterns in epithelial and mesenchymal
tissues (58). This
splicing-mediated diversification enables context-dependent
modulation of receptor-ligand interactions, contributing to
tissue-specific signaling outcomes (48). FGFRb variants are predominantly
expressed in epithelial lineages and interact with
mesenchymal-derived FGFs (FGF7/10), reflecting a paracrine
signaling paradigm (59).
Conversely, FGFRc isoforms localize to mesenchymal compartments and
bind FGF ligands produced in both epithelial and mesenchymal
tissues, enabling autocrine/paracrine signaling flexibility
(60,61). This splicing-mediated receptor
diversification creates tissue-specific receptor-ligand interfaces
that modulate developmental and homeostatic processes through
differential ligand availability and binding kinetics.
The biological activity of FGF is enhanced by
interactions with HS and other sulphated glycosaminoglycans
(38,62). The protein class present on the
cell surface and extracellular matrix is primarily composed of HS
proteoglycan (HSPG), which includes HS covalently bound to a
protein core. HSPG consists of membrane-spanning proteins on the
cell surface, cell surface proteins anchored by
glycosylphosphatidylinositol and diffusible protein components of
the extracellular matrix such as basement membrane and agrin
(38). Cell surface HSPGs and
extracellular matrix regulate FGF diffusion dynamics in tissue,
acting as cofactors stabilizing FGF-FGFR heterodimer complexes
through electrostatic interactions, thereby enhancing ligand
receptor affinity and intensifying downstream receptor activation
cascades (63-65). Endocrine FGF differs from
classical FGF in that it requires protein cofactors to promote
receptor binding and activation. Members of the Klotho family
fulfil this role, as evidenced by phenotypic similarities observed
in αKlotho and FGF23 knockout (KO) mice (66-68).
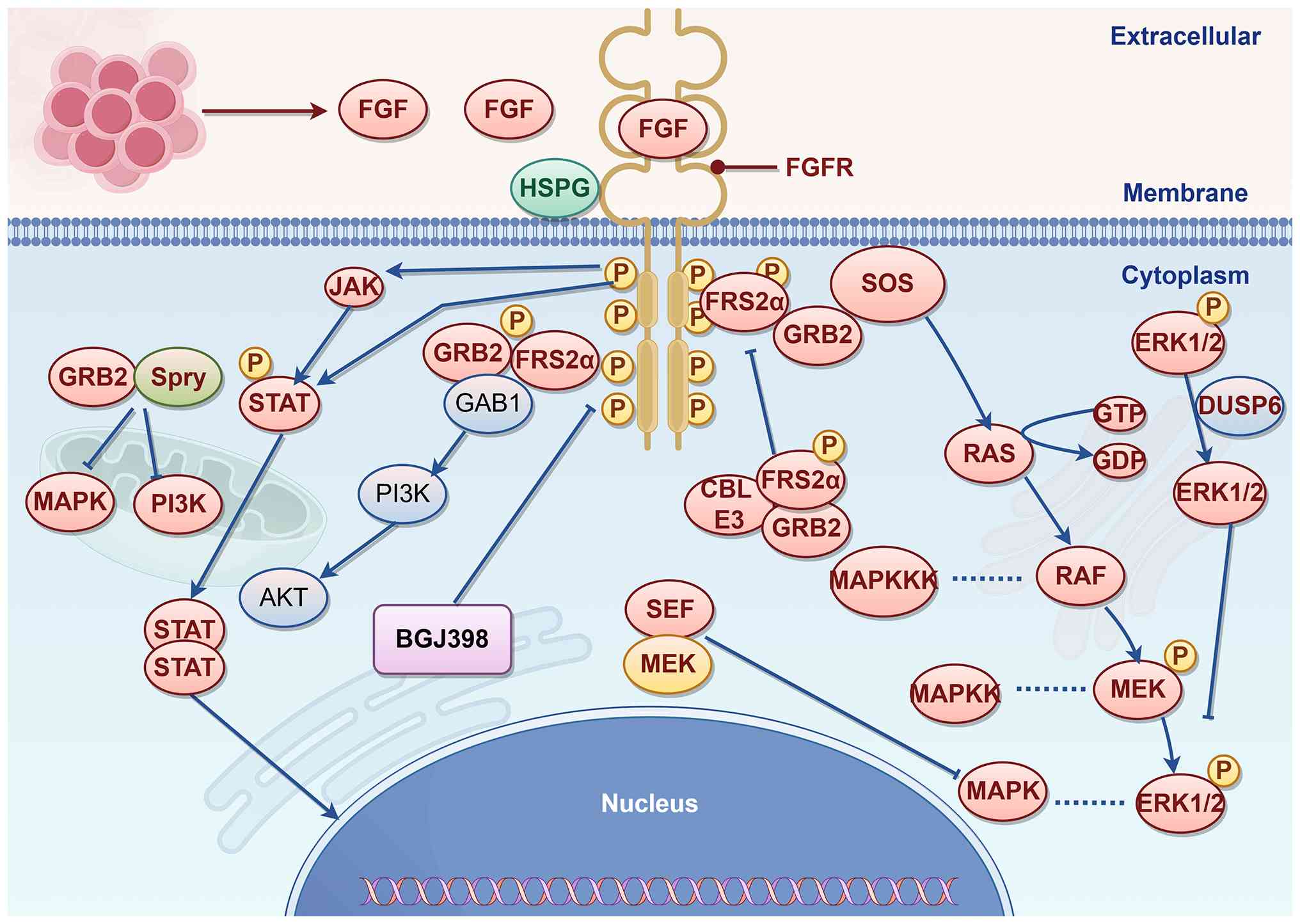
The binding of FGF ligands to FGFRs induces
dimerization and juxtaposition of the TK domains, thereby
initiating the sequential transphosphorylation of at least six
tyrosine residues (38,69,70). Activation of the FGFR TK domain
induces direct phosphorylation of FGFR substrate 2α (FRS2α) docking
binding proteins and promotes the recruitment of downstream
signaling molecules, including PLCγ, STAT1, STAT3 and STAT5,
through the formation of receptor-substrate complexes (38). In fibroblasts, FGF ligand
stimulation triggers FGFR3 to activate STAT (71,72). Phosphorylated FRS2α activates the
MAPK pathway through Son of sevenless. Additionally, phosphorylated
FRS2α binds growth factor receptor-bound protein 2 (GRB2), which
subsequently recruits GRB2-associated binder 1 to trigger the
PI3K/AKT signaling pathway (73,74). Downstream from RAS and PI3K, FGFs
activate several distinct MAPKs, including ERK1/2, JNK and p38
(75-79). The binding of v-crk sarcoma virus
CT10 oncogene homolog-like to the activated FGFR enhances the
phosphorylation of FRS2α, as well as the MAPK pathway signaling and
the activation of ERK1/2 (80).
FGFR intracellular signaling cascades are subject to
inhibitory regulation mediated by molecules such as GRB2. GRB2,
which binds to FRS2α, forms a complex with the casitas B-lineage
lymphoma (CBL) E3 ligase, which transfers ubiquitin molecules to
FGFR and FRS2α, leading to their polyubiquitination (81). The ubiquitinated receptor and
adaptor proteins are targeted by lysosomes or proteasomes for
degradation, effectively shutting down ongoing signaling and
achieving negative feedback inhibition (81). Sprouty proteins, which interact
with GRB2 to suppress downstream MAPK and PI3K pathway activation
(82,83). In the downstream region, similar
expression to fibroblast growth factor antagonizes the MAPK pathway
through its interaction with MEK (84), while dual-specificity phosphatase
6 suppresses MAPK signaling through the dephosphorylation of ERK1/2
(85). CBL E3 ligase has been
shown to inhibit FGFR signaling by forming a triple complex with
phosphorylated FRS2 and GRB2, where it promotes ubiquitination and
degradation of FGFR and FRS2 (86). CBL interacts with PI3K, resulting
in its ubiquitination and degradation (87). The GRB14 adapter protein binds
phosphorylated FGFR of Tyr766, thereby interfering with
phosphorylation and activation of PLCγ (88).
As aforementioned, FGFR1 is a TK cell surface
receptor, the expression of which is primarily observed in the
extracellular matrix and smooth muscle tissue, which constitutes a
notable component of cartilage tissue. RA primarily affects
cartilage tissue, leading to bone destruction (35,56,58,89). Consequently, FGFR1 may serve a
pivotal function in the progression of RA. In vitro,
attenuation of the FGFR1 signaling pathway may lead to attenuation
of bone decay and beneficial therapeutic effects in rat models of
arthritis (14,90). The immunoglobulin-cell sII and
III domains of FGFR1, together with the interdomain hinge regions,
serve a key role in the regulation of FGF ligand binding
specificity (91). The acidic
amino acid motif AB, located within the interdomain region between
extracellular immunoglobulin-like domains I and II of FGFR1,
demonstrates an inhibitory effect on FGFR ligand-receptor
interactions when acting in concert with domain I (92). The intracellular TK domain of
FGFR1 demonstrates constitutive TK activity. In the absence of FGF,
the receptor exists as a monomeric unit with the TK domain
maintained in a phosphorylated state. Following FGF engagement,
receptor dimerization facilitates the formation of a functional
2FGF:2FGFR1 complex, leading to kinase activation and
trans-phosphorylation events. This results in full TK activation
and subsequent downstream signal transduction cascades (59,93-95).
Following exposure to FGFs and heparin/klotho, FGFR1
kinases undergo dimerization in the active
aspartate-phenylalanine-glycine motif-aspartate-in conformation
(DFG-Asp-in) conformation, a process mediated by co-receptor
interactions that stabilize the signaling complex and promote
kinase activation (96). FGF
demonstrates preferential binding to the extracellular domains of
FGFR, with a marked affinity for the D2 and D3 immunoglobulin-like
domains and their intervening interdomain linker regions. This
binding stabilizes the FGF-FGFR-HS complex and promotes receptor
dimerization (97). Upon
receptor dimerization, the FGFR kinase domain undergoes
conformational reorganization, transitioning from an Upon receptor
dimerization, the FGFR kinase domain shifts from the autoinhibitory
DFG-out conformation to the catalytically active DFG-in
conformation (97). This
structural rearrangement facilitates ATP-binding pocket alignment
and activation loop stabilization, enabling kinase activation and
downstream signaling initiation. In its active DFG-in conformation,
the FGFR kinase domain DFG motif aspartate residue orients toward
the catalytic site, enabling ATP coordination and substrate
phosphorylation (97,98). Following FGF and heparin/klotho
engagement, FGFR1 kinases assemble into active DFG-in
conformational dimers, a process stabilized by co-receptor
interactions. Trans-phosphorylation of tyrosine residues within the
activation loop amplifies kinase catalytic efficiency through
allosteric modulation of the catalytic site (99,100). The intramolecular regulatory
element within the FGFR kinase domain, composed of the conserved
never in mitosis gene A-related kinase triplet, undergoes
conformational disengagement. This structural rearrangement
disrupts the hydrogen-bonding network that stabilizes the
autoinhibited state, thereby enabling the kinase domain to adopt a
catalytically competent conformation (96). In this active conformation, the
activated kinases trigger downstream signaling cascades through the
recruitment of cytoplasmic adaptor proteins, thereby activating key
intracellular pathways including RAS/MAPK, PI3K/AKT, PLCγ and STAT
signaling. This process involves the formation of multi-protein
complexes that transduce extracellular signals to nuclear
effectors, modulating cell responses such as proliferation,
differentiation and survival (38) (Fig. 2).
FGFR1 signaling, through its downstream effectors
including RAS/MAPK and PI3K/AKT pathways, serves a crucial role in
numerous physiological processes. During embryogenesis, FGFR1 plays
a pivotal role in regulating essential cell behaviors, including
proliferation, differentiation and migration, which are fundamental
for the proper formation of tissue and organs (101-103). FGFR-mediated signaling
coordinates the morphogenesis of neural, skeletal and
cardiovascular systems, ensuring precise spatiotemporal integration
and functional maturation of developing organ structures (104-106). FGFR1 exerts key functions in
regulating developmental processes and sustaining metabolic
homeostasis during infancy and postnatal maturation. Its signaling
is key for driving tissue expansion, orchestrating organogenesis
and modulating nutrient utilization pathways to ensure proper
growth trajectories and metabolic equilibrium (107). FGFR1 modulates adipogenic
differentiation and governs glucose homeostasis alongside lipid
metabolic pathways, thereby serving as a key regulator of systemic
energy balance (108). This
multifaceted role positions FGFR1 signaling at the nexus of
metabolic regulation, integrating nutrient sensing with cell energy
utilization (109). FGFR1
serves a pivotal role in metabolic regulation, which is key for the
adaptation of the body to fluctuating nutritional and energy
demands throughout the growth and maturation process (110). FGFR1 signaling coordinates
tissue repair and regeneration in adults via stem cell activation
modulation, angiogenesis promotion and extracellular matrix
remodeling regulation (111,112). FGFR1 serves a pivotal role in
orchestrating cellular injury responses (113). It boosts the proliferation and
migration of stromal fibroblasts, vascular endothelial cells, and
other typical reparative cells including mesenchymal stem cells and
pericytes, all of which are key for tissue regeneration (114,115). This signaling axis modulates
the recruitment and activation of cells following tissue injury via
context-dependent transcriptional programs, and thus orchestrates
the full sequence of wound repair, ranging from hemostasis to
remodeling (116-118). FGFR1-mediated signaling serves
a key role in wound repair by orchestrating neovascularization,
extracellular matrix synthesis and structural reorganization of
damaged tissue. This multifaceted regulation ensures coordinated
progression through healing phases, culminating in functional
restoration and preservation of tissue architectural integrity
(119).
Genetic ablation of FGFR1 in murine models results
in increased trabecular bone mass and cortical thickness,
accompanied by impaired osteoclast function and enhanced osteoblast
activity (Table I) (35,120,121). These findings are supported by
pharmacological studies demonstrating that FGFR1 inhibition
attenuates bone erosion and synovitis in experimental arthritis
models (13,122,123). However, the cell type-specific
mechanisms through which FGFR1 coordinates bone destruction in RA
remain incompletely understood, particularly regarding its
interplay with inflammatory mediators and other signaling pathways
such as MAPK, NF-κB and PI3K/AKT pathways (32,123,124).
RA bone destruction is associated with bone tissue
remodeling and maintenance of bone homeostasis. Bone tissue
contains three types of cell: Osteocytes, osteoblasts and
osteoclasts. The most abundant type of cell in adult bones is
osteocytes, which are osteoblasts that differentiate and control
bone remodeling (89,125). Osteocytes regulate osteoblast
differentiation and anabolic action, as well as osteoclastic
activity, either by direct dendritic contact or by the secretion of
regulatory factors such as sclerostine receptor activators and
NF-κB B binders (B ligand-factor nucleic receptor activators,
RANKL) (126). Bone cells are
one of the primary sources of RANKL and FGFR1 serves an important
role in bone development and bone homeostasis. FGFR1 activation
promotes the expression of MMP 2 and RANKL by promoting ERK1/2
phosphorylation (127,128). Mice with FGFR1 and/or FGFR2
deficiency exhibit normal phenotypes at birth, yet develop a high
bone mass phenotype with age (129). These animals show enhanced
periosteal adhesion, expanded and disorganized intracortical bone
and elevated porosity, as well as improved mechanical strength and
stiffness consistent with higher bone mass (129). While KO of specific FGFRs
increases bone mass, mechanical properties such as fracture
toughness are impaired. Despite higher overall bone mass, these
bones show inferior mechanical performance compared with normal
bone. These data establish the role of FGFR1 signaling in
osteoblasts, indicating that FGFR1 is essential for osteoblast
survival and regulation of bone mass during postnatal bone growth
(129). Loss of FGFR1 in
osteocytes activates the Wnt/β-catenin signaling pathway, promotes
trabecular and cortical bone formation and increases
osteoprotegerin mRNA expression and osteoblast activity (35). In addition, the loss of FGFR1 in
osteocytes leads to decreased osteoclast formation, expression of
RANKL mRNA in cortical bone and primary osteocytes and mRNA
expression of genes associated with bone resorption, such as MMP 9
and catechsyn K, suggesting that the signals produced by FGFR1 loss
in osteocytes inhibit osteoclast formation (35). However, these structural
alterations paradoxically coincide with impaired biomechanical
properties, indicating that complete ablation of FGFR1 may disrupt
the equilibrium between bone quantity and quality. While genetic
deletion studies have elucidated the role of FGFR1 in bone
development, the ramifications of pharmacological inhibition in
established RA remain unclear (35,130). Secondly, FGFR1-mediated
regulation of both Wnt/β-catenin and RANKL pathways poses a
challenge in achieving a balanced therapeutic outcome,
necessitating dual pathway modulation strategies. Thirdly, the net
impact of FGFR1 modulation is contingent on its differential
effects on osteocytes, osteoblasts and osteoclasts, underscoring
the importance of cell type-specific responses. Collectively, these
findings position FGFR1 as a promising therapeutic target for
mitigating RA-associated bone loss, and future investigations
should ascertain whether selective, partial inhibition of FGFR1
signaling preserves its physiological functions while curbing
pathological bone destruction in RA.
Osteoclasts are large multinucleated cells whose
primary function is the absorption of bone (142). These cells originate in the
hematopoietic system and differentiate into mature osteoclasts
through the interaction between macrophage colony-stimulating
factors and RANKL. Macrophage colonystimulating factor promotes the
expansion of osteoclast precursors, whereas RANKL induces their
differentiation into functional osteoclasts (132). FGFR1 serves an important role
in osteoclast differentiation and function. Studies have shown that
activated FGFR1 and MAPK act directly on mature osteoclasts to
stimulate bone resorption (143,144). FGFR1 expression is detected on
isolated mouse osteoclasts (145). In addition, FGFR1 expression is
also detected in osteoclasts near fracture sites (146). To explore the direct effect of
FGFR1 on osteoclasts, Lu et al (147) knocked out FGFR1 in mouse bone
marrow mononuclear cells and mature osteoclasts: Mutant mice showed
abnormal bone remodeling, decreased numbers of osteoclasts,
impaired osteoclast function and decreased expression of
tartrate-resistant acid phosphatase and MMP 9. Loss of FGFR1
decreases the number and activity of osteoclasts in mice,
indicating that FGFR1 has a positive regulatory effect on
osteoclasts. FGFR1 is key for the complete differentiation and
activation of mouse osteoclasts (147). FGFR1 directly promotes the
differentiation and activation of osteoclasts by enhancing ERK 1/2
pathway signaling, thus enhancing their ability to decompose bone
(143,147). Aukes et al (148) showed that in the co-culture
environment of osteoclasts and breast cancer cells, FGFR inhibitor
BGJ398 decreases the activation of FGFR-mediated signaling and
expression of osteoclast target genes (149). FGFR1 inhibitors enhance
osteoblast differentiation in vitro and FGFR1 activation
inhibits phosphate metabolism. Overall, FGFR1 maintains bone
balance by modulating osteoblast activities, but the mechanisms and
its full therapeutic potential require further exploration
(150,151).
Angiogenesis serves a key role in maintaining and
promoting the progression of RA (199). Studies have shown that
angiopoietin, hypoxia-inducible factor, VEGF and FGF serve an
important role in inhibiting synovial angiogenesis in RA,
suggesting that targeting angiogenesis may be an effective
treatment for RA (200,201). Binding of these GFs to their
receptors leads to VE cell deformation, division, migration and
proliferation, inducing neovascularization; in addition,
angiogenesis in synovial tissue stimulates intravascular
extravasation, exacerbating synovial inflammation and vessel
formation (202). Targeting
FGFR1 to inhibit angiogenesis has been demonstrated in tumor
therapy (203-205). In RA neovascularization, many
proangiogenic factors, including VEGF and sphingosine-1-phosphate,
induce abnormal activation of VE cells and promote angiogenesis
(206). Studies have shown that
activated FGFR1 upregulates VEGF A expression (207,208). Promoting VEGF A expression and
secretion mediates FGFR1 activation to induce angiogenesis
(209). Lowering FGFR1 protein
levels impairs endothelial cell function, particularly during
angiogenesis (210). FGFR1
activation promotes endothelial cell proliferation and migration by
activating MAPK signaling pathways (211). To the best of our knowledge, no
studies have linked FGFR1 with angiogenesis and bone destruction in
RA. Therefore, investigating the role of FGFR1 in inhibiting
synovial vascularization in RA may facilitate treatment of RA
vascularization and improvement of RA bone destruction. In RA,
FGFR1 is associated with VEGFA production and key endothelial
activities including proliferation and migration. Nevertheless, its
specific roles in synovial angiogenesis and secondary bone damage
remain unclear (Fig. 3).
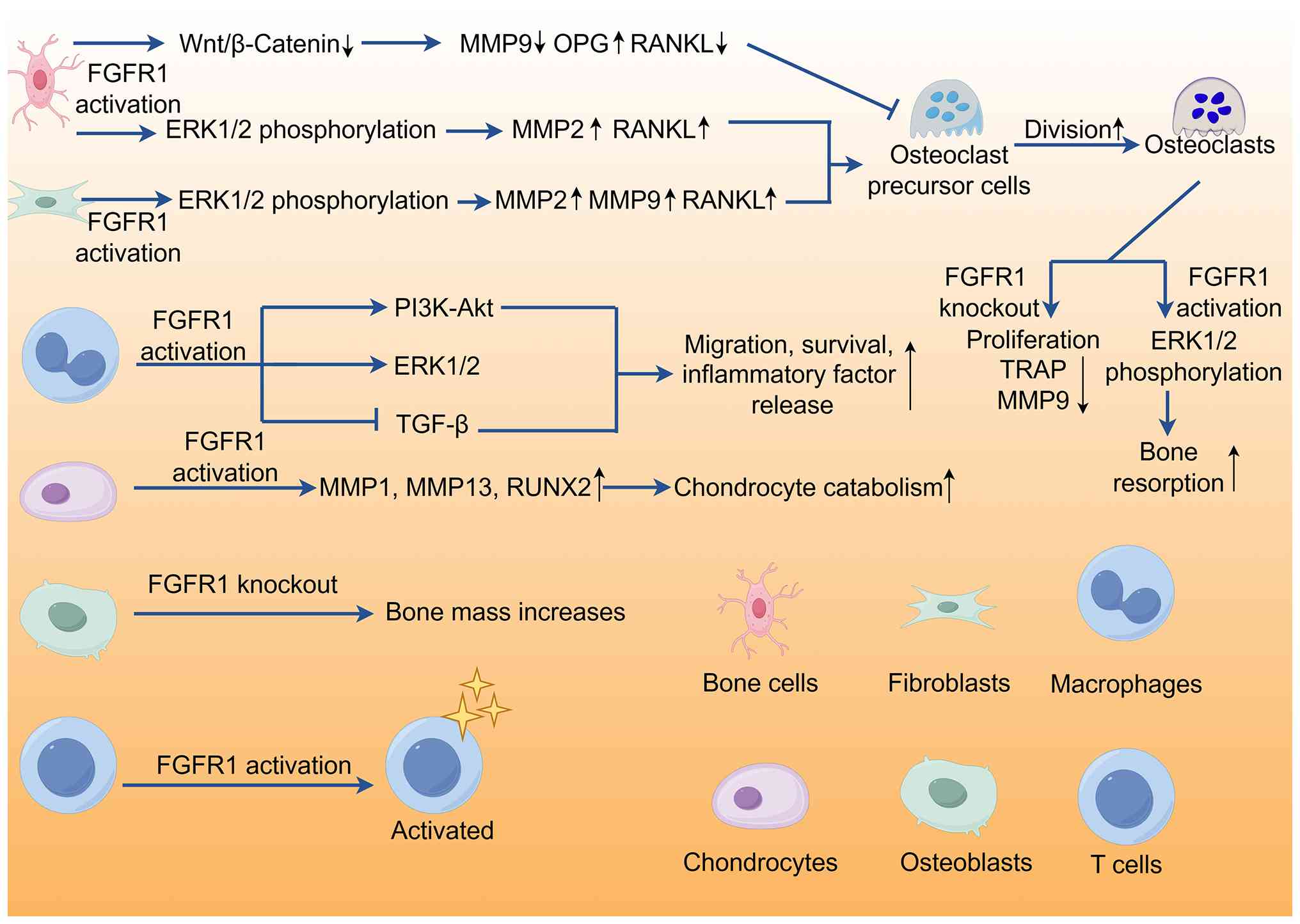
The contradictory roles of FGFR1 in different cell
types primarily stem from its complex signaling network and cell
type-specific responses (106,212). Environmental factors, cell
type-specific characteristics and stages of disease influence the
ultimate effects of FGFR1. Therefore, when studying FGFR1
functions, it is necessary to consider these factors to understand
its mechanisms of action under different physiological and
pathological conditions (106).
Moreover, targeted therapeutic strategies against FGFR1 need to be
adjusted according to the specific disease and cell type to achieve
the best therapeutic outcome (213). In bone cells, FGFR1 activation
promotes the formation of osteoclasts and bone resorption by
upregulating the expression of MMP 2 and RANKL, leading to a
decrease in bone mass and enhanced bone resorption; FGFR1 also
serves a role in promoting bone formation in bone cells (127,128). The loss of FGFR1 in bone cells
activates the Wnt/β-catenin signaling pathway, promotes the
formation of trabecular and cortical bone and increases the
expression of osteoprotegerin mRNA, thereby enhancing bone
formation (214). The
microenvironment of bone cells, including extracellular matrix
components and local concentrations of GF, may affect FGFR1
signaling pathways, causing it to promote bone resorption or
enhance bone formation. In different stages of RA, bone cell
responses may differ, with early stages dominated by inflammation
and bone resorption, while later stages may involve reparative bone
formation (215). In
osteoblasts, FGFR1 expression is key for bone mineralization and
formation (17). However,
excessive activation of FGFR1 may inhibit osteoblast maturation and
mineralization, leading to decreased bone formation. During
osteoblast development, FGFR1 promotes the proliferation of
progenitor cells and differentiation into pre-osteoblasts in the
early phase. In mature osteoblasts, by contrast, FGFR1 restrains
excessive mineralization to sustain bone homeostasis. Notably,
these biological processes are tightly modulated by multiple
downstream signaling pathways. In articular chondrocytes, FGFR1
activation upregulates the expression of MMP1 and MMP13 in the
matrix, promoting matrix degradation and cartilage damage (216). However, in gene KO models
(128,217), the loss of FGFR1 protects
cartilage from degeneration and decreases cartilage destruction. In
inflammatory diseases such as RA, inflammatory factors may alter
the response of chondrocytes to FGFR1 signaling, causing its role
to shift from protective to destructive (128,154).
As FGFR1 is key for bone resorption and bone
reconstruction in the induction and development of RA, blockade of
FGFR1 might represent a novel therapeutic strategy for the
treatment of RA. There is evidence that mice lacking FGFR1 in
osteocytes have increased trabecular bone mass and cortical bone
thickness, which is attributed to increased bone formation and
impaired bone resorption (35,129). Consequently, FGFR1 is
hypothesized to be a good molecular target for decreasing bone loss
in RA. Tang et al (35)
used osteocyte-specific FGFR1 KO mice to demonstrate that FGFR1
deficiency enhances trabecular bone mass and cortical bone
thickness through augmented bone formation and attenuated bone
resorption. In vitro, Meng et al (14) showed that the FGF pathway is a
critical signaling pathway in relapsed RA, and targeted
tissue-specific inhibition of FGF10/FGFR1 may provide new
opportunities to treat patients with relapse RA. FGFR1 inhibitors
could effectively improve the effectiveness of RA treatment.
Additionally, treating rats with PD173074 (FGFR1 inhibitor)
significantly decreases bone erosion (14).
Drugs used in clinical practice primarily include
non-steroidal anti-inflammatory drugs (NSAIDs), glucocorticoids and
DMARDs (223,224). Clinical trials have
demonstrated that these drugs effectively improve the condition of
patients with RA (225,226). NSAIDs and glucocorticoids serve
a role in relieving pain and reducing inflammation (227,228). DMARDs are effective first-line
drugs for treating RA in clinical practice (229). FGFR1 inhibitors may inhibit
inflammatory responses, counteract bone erosion, and reduce bone
loss by suppressing the bone-resorbing activity of osteoclasts,
inhibiting chondrocyte degradation, suppressing macrophage
activity, and protecting degenerated chondrocytes. The advantages
of FGFR1 inhibitors in the treatment of RA as well as potential
risks are summarized in Table
III.
FGFR1 is a key regulator in RA-associated bone
destruction, orchestrating crosstalk between synovial angiogenesis,
osteoclast activation and inflammatory signaling. However, the
specific regulatory mechanisms remain unclear, particularly the
signal transduction mechanisms of FGFR1 in the pathogenesis of RA
and across different cell types, which require further validation.
FGFR1 inhibition attenuates bone erosion and synovitis in
preclinical models (230-232), suggesting its therapeutic
potential. However, challenges persist, including the paradoxical
roles of FGFR1 in different cell types and the need for targeted
delivery strategies to minimize off-target effects. Advanced
single-cell sequencing and spatial transcriptomics may delineate
cell-type-specific FGFR1 signaling networks in RA joints. The
therapeutic potential of selective FGFR1 inhibitors, particularly
their capacity to simultaneously target bone erosion and synovitis,
warrants validation in human clinical trials. By bridging these
gaps, FGFR1-targeted therapies may redefine RA management,
particularly for refractory cases with aggressive joint
destruction.
All authors have read and approved the final
manuscript. CQ, ZZ, XW, XL and FLY conceived the study. CQ, HT, JW,
ZY and ZZ constructed figures. CQ and FLY wrote the manuscript.
Data authentication is not applicable.
Not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by National Natural Science
Foundation of China (grant nos. 82372412 and 82572767), the Social
Development Project of Jiangsu Province (grant no. BE2022701),
Jiangsu Provincial Traditional Chinese Medicine Science and
Technology Development Program (grant no. MS2023069), Jiangsu
Province Leading Talents Cultivation Project for Traditional
Chinese Medicine (grant no. SLJ0322) and Wuxi Municipal Health
Commission Research Project (grant no. Z202405).
|
1
|
Shen Y, Zou L, Zeng Y, Xia T, Liu Z, Hu K,
Wang L and Feng J: Potential of Alkaloids from Zanthoxylum nitidum
var. tomentosum in treating rat rheumatoid arthritis model and
validation of molecular mechanisms. Curr Issues Mol Biol.
47:6612025. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu L, Wang J, Chen Y, Fang Y, Wang Z, Liu
X, Hong L, Tao Q and Xu Y: Efficacy and safety of the bushen quhan
zhiwang decoction to achieve clinical deep remission in rheumatoid
arthritis: Protocol for a double-blind, randomized,
placebo-controlled trial. J Pain Res. 18:4237–4249. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Z, Dong Q, Zhang L, Wang X, Jiao Y,
Geng Q, Cai W, Zhang Q, Xiao C, Tao Q and Xu Y: Fibroblast
activation protein-α drives rheumatoid arthritis inflammation
through the AKT/mTOR signaling pathway and its therapeutic effect
by Wangbi granules. Phytomedicine. 147:1571792025. View Article : Google Scholar
|
|
4
|
Peng Z, Huang W, Tang M, Chen B, Yang R,
Liu Q, Liu C and Long P: Investigating the shared genetic
architecture between hypothyroidism and rheumatoid arthritis. Front
Immunol. 14:12864912024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y, Deng Y, Zheng H, Li B, Yang Y,
Huang J, Yuan H, Wang M, Wang W and Yu H: Xuetongsu attenuates
synovial inflammation in rheumatoid arthritis by inhibiting the
IL-23/IL-17/NF-κB inflammatory axis. Front Pharmacol.
16:16155192025. View Article : Google Scholar
|
|
6
|
Zhang Z, Liu L, Ti H, Chen M, Chen Y, Du
D, Zhan W, Wang T, Wu X, Wu J, et al: Synovial fibroblast derived
small extracellular vesicles miRNA15-29148 promotes articular
chondrocyte apoptosis in rheumatoid arthritis. Bone Res. 13:612025.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan FL, Li X, Lu WG, Li CW, Xu RS and
Dong J: IL-33: A promising therapeutic target for rheumatoid
arthritis? Expert Opin Ther Targets. 15:529–534. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan FL, Hu W, Lu WG, Li X, Li JP, Xu RS,
Li CW, Chen FH and Jin C: Targeting interleukin-21 in rheumatoid
arthritis. Mol Biol Rep. 38:1717–1721. 2011. View Article : Google Scholar
|
|
9
|
Firestein GS: Evolving concepts of
rheumatoid arthritis. Nature. 423:356–361. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ngo ST, Steyn FJ and McCombe PA: Gender
differences in autoimmune disease. Front Neuroendocrinol.
35:347–369. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hajar H and Lahcen A: insufficiency
fracture in a patient with rheumatoid arthritis in remission
receiving long-term methotrexate without conventional risk factors:
A case report and literature review. Cureus.
18:e1078062026.PubMed/NCBI
|
|
12
|
Hirose H, Higuchi T, Ichimura Y, Katsumata
Y, Tanaka E, Yamamoto T, Koenuma N, Yano K, Shinohara A, Seo S and
Harigai M: Periarticular other iatrogenic
immunodeficiency-associated lymphoproliferative disorder mimicking
knee synovitis in a patient with rheumatoid arthritis. Intern Med.
65:905–910. 2026. View Article : Google Scholar :
|
|
13
|
Ji L, Kong R, Yu Y, Wan W, Zhao D and Gao
J: Klotho promotes bone formation in rheumatoid
arthritis-associated osteoporosis by modulating the
Fgf23/Fgfr1/NF-κB pathway and inhibiting ferroptosis. Mol Immunol.
184:64–75. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meng X, Chen Z, Li T, Nie Z, Han H, Zhong
S, Yin Z, Sun S, Xie J, Shen J, et al: Role and therapeutic
potential for targeting fibroblast growth factor 10/FGFR1 in
relapsed rheumatoid arthritis. Arthritis Rheumatol. 76:32–47. 2024.
View Article : Google Scholar
|
|
15
|
Etori K, Tanaka S, Tamura J, Hattori K,
Kagami SI, Nakamura J, Ohtori S and Nakajima H: Fibroblast growth
factor receptor 1 as a potential marker of terminal effector
peripheral T helper cells in rheumatoid arthritis patients.
Rheumatology (Oxford). 62:3763–3769. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Plotnikov AN, Schlessinger J, Hubbard SR
and Mohammadi M: Structural basis for FGF receptor dimerization and
activation. Cell. 98:641–650. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Ellman MB, Kroin JS, Chen D, Yan D,
Mikecz K, Ranjan KC, Xiao G, Stein GS, Kim SG, et al:
Species-specific biological effects of FGF-2 in articular
cartilage: Implication for distinct roles within the FGF receptor
family. J Cell Biochem. 113:2532–2542. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu L, Leijten J, van Blitterswijk CA and
Karperien M: Fibroblast growth factor-1 is a mesenchymal stromal
cell-secreted factor stimulating proliferation of osteoarthritic
chondrocytes in co-culture. Stem Cells Dev. 22:2356–2367. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao S, Wang Y, Hou L, Wang Y, Xu N and
Zhang N: Pentraxin 3 inhibits fibroblast growth factor 2 induced
osteoclastogenesis in rheumatoid arthritis. Biomed Pharmacother.
131:1106282020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang ZZ, Liu F, Gong YF, Huang TY, Zhang
XM and Huang XY: Antiarthritic effects of sorafenib in rats with
adjuvant-induced arthritis. Anat Rec (Hoboken). 301:1519–1526.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Demuynck B, Shah BP, Mayeux F, Vasseur L,
Barbault F, Ding J, Paull M, Reddi T, Muslimova E and Legeai-Mallet
L: Infigratinib low dose therapy is an effective strategy to treat
hypochondroplasia. J Bone Miner Res. 40:1255–1264. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Othman AA, Babcock HE, Gill CS, Fraser JL,
Regier DS, Kaur R, Simpson KL and Ferreira CR: New Phenotypic
features in FGFR1-Related osteoglophonic dysplasia. Am J Med Genet
A. 197:e640922025. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Savarirayan R, De Bergua JM, Arundel P,
Salles JP, Saraff V, Delgado B, Leiva-Gea A, McDevitt H, Nicolino
M, Rossi M, et al: Oral infigratinib therapy in children with
achondroplasia. N Engl J Med. 392:865–874. 2025. View Article : Google Scholar
|
|
24
|
Rikimaru S, Nakao-Kuroishi K,
Kometani-Gunjigake K, Mizuhara M, Nakatomi C, Toyono T, Ono K and
Kawamoto T: Fibroblast growth factor 2 stimulates differentiation
of mechanically-stressed human periodontal ligament fibroblasts
into cementoblasts. Am J Orthod Dentofacial Orthop. 168:477–488.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yi X, Zimath PL, Martin-Vazquez E,
Oliveira JG, Jawurek S, Title AC, Yesildag B, Mourad NI, Buemi A,
Pattou F, et al: Transcriptomics of autoimmune diseases identifies
FGFR1 as a target for pancreatic β-cell protection. J Autoimmun.
156:1034692025. View Article : Google Scholar
|
|
26
|
Zhou H, Pan H, Li X, Huang L, Zhang R, Yan
X and Xu J: Ginsenoside reprogramming microglia through the
FGF/FGFR1 inhibits post traumatic stress disorder. Int
Immunopharmacol. 145:1137632025. View Article : Google Scholar
|
|
27
|
Li HZ, Zhang JL, Yuan DL, Xie WQ, Ladel
CH, Mobasheri A and Li YS: Role of signaling pathways in
age-related orthopedic diseases: Focus on the fibroblast growth
factor family. Mil Med Res. 11:402024.PubMed/NCBI
|
|
28
|
Demuynck B, Flipo J, Kaci N, Dambkowski C,
Paull M, Muslimova E, Shah BP and Legeai-Mallet L: Low-dose
infigratinib increases bone growth and corrects growth plate
abnormalities in an achondroplasia mouse model. J Bone Miner Res.
39:765–774. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dutra EH, Chen PJ, Kalajzic Z, Wadhwa S,
Hurley M and Yadav S: FGF ligands and receptors in osteochondral
tissues of the temporomandibular joint in young and aging mice.
Cartilage. 15:195–199. 2024. View Article : Google Scholar :
|
|
30
|
Wang Q, Li H, Mao Y, Garg A, Park ES, Wu
Y, Chow A, Peregrin J and Zhang X: Shc1 cooperates with Frs2 and
Shp2 to recruit Grb2 in FGF-induced lens development. Elife.
13:RP1036152025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Q, Tao C, Wu Y, Anderson KE, Makrides
N, Hannan A, Peregrin J, Lin CS, Ding Z, Li W, et al: FGF-induced
phospholipase Cγ signaling regulates lacrimal gland branching by
competing with PI3K in phosphoinositide metabolism. Cell Rep.
44:1160462025. View Article : Google Scholar
|
|
32
|
Nakano K, Okada Y, Saito K and Tanaka Y:
Induction of RANKL expression and osteoclast maturation by the
binding of fibroblast growth factor 2 to heparan sulfate
proteoglycan on rheumatoid synovial fibroblasts. Arthritis Rheum.
50:2450–2458. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Soutto M, Zhang X, Bhat N, Chen Z, Zhu S,
Maacha S, Genoula M, El-Gazzaz O, Peng D, Lu H, et al: Fibroblast
growth factor receptor-4 mediates activation of nuclear factor
erythroid 2-Related Factor-2 in gastric tumorigenesis. Redox Biol.
69:1029982024. View Article : Google Scholar :
|
|
34
|
Farrell B and Breeze AL: Structure,
activation and dysregulation of fibroblast growth factor receptor
kinases: Perspectives for clinical targeting. Biochem Soc Trans.
46:1753–1770. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tang Y, Yang P, Jin M, Huang S, Chen H,
Chen L, Yang J and Su N: Fgfr1 deficiency in osteocytes leads to
increased bone mass by enhancing Wnt/β-catenin signaling. Bone.
174:1168172023. View Article : Google Scholar
|
|
36
|
Wang L, Luo W, Zhang S, Zhang J, He L, Shi
Y, Gao L, Wu B, Nie X, Hu C, et al: Macrophage-derived FGFR1 drives
atherosclerosis through PLCγ-mediated activation of NF-κB
inflammatory signalling pathway. Cardiovasc Res. 120:1385–1399.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kalinina J, Dutta K, Ilghari D, Beenken A,
Goetz R, Eliseenkova AV, Cowburn D and Mohammadi M: The
alternatively spliced acid box region plays a key role in FGF
receptor autoinhibition. Structure. 20:77–88. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ornitz DM and Itoh N: The fibroblast
growth factor signaling pathway. Wiley Interdiscip Rev Dev Biol.
4:215–266. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schlessinger J, Plotnikov AN, Ibrahimi OA,
Eliseenkova AV, Yeh BK, Yayon A, Linhardt RJ and Mohammadi M:
Crystal structure of a ternary FGF-FGFR-heparin complex reveals a
dual role for heparin in FGFR binding and dimerization. Mol Cell.
6:743–750. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Farooq M, Khan AW, Kim MS and Choi S: The
role of fibroblast growth factor (FGF) signaling in tissue repair
and regeneration. Cells. 10:32422021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Orr-Urtreger A, Bedford MT, Burakova T,
Arman E, Zimmer Y, Yayon A, Givol D and Lonai P: Developmental
localization of the splicing alternatives of fibroblast growth
factor receptor-2 (FGFR2). Dev Biol. 158:475–486. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu X, Weinstein M, Li C, Naski M, Cohen
RI, Ornitz DM, Leder P and Deng C: Fibroblast growth factor
receptor 2 (FGFR2)-mediated reciprocal regulation loop between FGF8
and FGF10 is essential for limb induction. Development.
125:753–765. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jin C, Wang F, Wu X, Yu C, Luo Y and
McKeehan WL: Directionally specific paracrine communication
mediated by epithelial FGF9 to stromal FGFR3 in two-compartment
premalignant prostate tumors. Cancer Res. 64:4555–4562. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yeh BK, Igarashi M, Eliseenkova AV,
Plotnikov AN, Sher I, Ron D, Aaronson SA and Mohammadi M:
Structural basis by which alternative splicing confers specificity
in fibroblast growth factor receptors. Proc Natl Acad Sci USA.
100:2266–2271. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Olsen SK, Li JY, Bromleigh C, Eliseenkova
AV, Ibrahimi OA, Lao Z, Zhang F, Linhardt RJ, Joyner AL and
Mohammadi M: Structural basis by which alternative splicing
modulates the organizer activity of FGF8 in the brain. Genes Dev.
20:185–198. 2006. View Article : Google Scholar :
|
|
46
|
Wiedemann M and Trueb B: Characterization
of a novel protein (FGFRL1) from human cartilage related to FGF
receptors. Genomics. 69:275–279. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Goida J and Pilmane M: The evaluation of
FGFR1, FGFR2 and FOXO1 in orofacial cleft tissue. Children (Basel).
9:5162022.PubMed/NCBI
|
|
48
|
Liu Q, Huang J, Yan W, Liu Z, Liu S and
Fang W: FGFR families: Biological functions and therapeutic
interventions in tumors. MedComm (2020). 4:e3672023. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Trueb B, Amann R and Gerber SD: Role of
FGFRL1 and other FGF signaling proteins in early kidney
development. Cell Mol Life Sci. 70:2505–2518. 2013. View Article : Google Scholar
|
|
50
|
Steinberg F, Zhuang L, Beyeler M, Kälin
RE, Mullis PE, Brändli AW and Trueb B: The FGFRL1 receptor is shed
from cell membranes, binds fibroblast growth factors (FGFs), and
antagonizes FGF signaling in Xenopus embryos. J Biol Chem.
285:2193–2202. 2010. View Article : Google Scholar
|
|
51
|
Salmaso N, Stevens HE, McNeill J, ElSayed
M, Ren Q, Maragnoli ME, Schwartz ML, Tomasi S, Sapolsky RM, Duman R
and Vaccarino FM: Fibroblast growth factor 2 modulates hypothalamic
pituitary axis activity and anxiety behavior through glucocorticoid
receptors. Biol Psychiatry. 80:479–489. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Regeenes R, Silva PN, Chang HH, Arany EJ,
Shukalyuk AI, Audet J, Kilkenny DM and Rocheleau JV: Fibroblast
growth factor receptor 5 (FGFR5) is a co-receptor for FGFR1 that is
up-regulated in beta-cells by cytokine-induced inflammation. J Biol
Chem. 293:17218–17228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Pal V, Wang Y, Regeenes R, Kilkenny DM and
Rocheleau JV: Laminin matrix regulates beta-cell FGFR5 expression
to enhance glucose-stimulated metabolism. Sci Rep. 12:61102022.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kähkönen TE, Ivaska KK, Jiang M, Büki KG,
Väänänen HK and Härkönen PL: Role of fibroblast growth factor
receptors (FGFR) and FGFR like-1 (FGFRL1) in mesenchymal stromal
cell differentiation to osteoblasts and adipocytes. Mol Cell
Endocrinol. 461:194–204. 2018. View Article : Google Scholar
|
|
55
|
Silva PN, Altamentova SM, Kilkenny DM and
Rocheleau JV: Fibroblast growth factor receptor like-1 (FGFRL1)
interacts with SHP-1 phosphatase at insulin secretory granules and
induces beta-cell ERK1/2 protein activation. J Biol Chem.
288:17859–17870. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Han X, Yang J, Li L, Huang J, King G and
Quarles LD: Conditional deletion of Fgfr1 in the proximal and
distal tubule identifies distinct roles in phosphate and calcium
transport. PLoS One. 11:e01478452016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu Z, Luo W, Chen L, Zhuang Z, Yang D,
Qian J, Khan ZA, Guan X, Wang Y, Li X and Liang G: Ang II
(Angiotensin II)-Induced FGFR1 (Fibroblast Growth Factor Receptor
1) activation in tubular epithelial cells promotes hypertensive
kidney fibrosis and injury. Hypertension. 79:2028–2041. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pepe F, Russo G, Scimone C, Palumbo L,
Tommasi S, Pinto R, De Biase D, Maloberti T, Busico A, Santoro A,
et al: Harmonization trial of FGFR1-3 testing strategies in
cholangiocarcinoma patients: An Italian multicenter experience.
Pathologica. 117:496–507. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Cheng YM, Chou CY, Hsu YC and Chen MJ:
Influence of HPV16 E6/7 on the expression of FGF2 and FGFR type B
in cervical carcinogenesis. Reprod Sci. 19:580–586. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Muh SJ, Hovhannisyan RH and Carstens RP: A
Non-sequence-specific double-stranded RNA structural element
regulates splicing of two mutually exclusive exons of fibroblast
growth factor receptor 2 (FGFR2). J Biol Chem. 277:50143–50154.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shin JW, Min M, Larrieu-Lahargue F, Canron
X, Kunstfeld R, Nguyen L, Henderson JE, Bikfalvi A, Detmar M and
Hong YK: Prox1 promotes lineage-specific expression of fibroblast
growth factor (FGF) receptor-3 in lymphatic endothelium: A role for
FGF signaling in lymphangiogenesis. Mol Biol Cell. 17:576–584.
2006. View Article : Google Scholar :
|
|
62
|
Shimokawa K, Kimura-Yoshida C, Nagai N,
Mukai K, Matsubara K, Watanabe H, Matsuda Y, Mochida K and Matsuo
I: Cell surface heparan sulfate chains regulate local reception of
FGF signaling in the mouse embryo. Dev Cell. 21:257–272. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Xu D and Esko JD: Demystifying heparan
sulfate-protein interactions. Annu Rev Biochem. 83:129–157. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Belov AA and Mohammadi M: Molecular
mechanisms of fibroblast growth factor signaling in physiology and
pathology. Cold Spring Harb Perspect Biol. 5:a0159582013.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wu ZL, Zhang L, Yabe T, Kuberan B, Beeler
DL, Love A and Rosenberg RD: The involvement of heparan sulfate
(HS) in FGF1/HS/FGFR1 signaling complex. J Biol Chem.
278:17121–17129. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kuro-o M: Endocrine FGFs and Klothos:
Emerging concepts. Trends Endocrinol Metab. 19:239–245. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Urakawa I, Yamazaki Y, Shimada T, Iijima
K, Hasegawa H, Okawa K, Fujita T, Fukumoto S and Yamashita T:
Klotho converts canonical FGF receptor into a specific receptor for
FGF23. Nature. 444:770–774. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kurosu H, Ogawa Y, Miyoshi M, Yamamoto M,
Nandi A, Rosenblatt KP, Baum MG, Schiavi S, Hu MC, Moe OW and
Kuro-o M: Regulation of fibroblast growth factor-23 signaling by
klotho. J Biol Chem. 281:6120–6123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Goetz R and Mohammadi M: Exploring
mechanisms of FGF signalling through the lens of structural
biology. Nat Rev Mol Cell Biol. 14:166–180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Furdui CM, Lew ED, Schlessinger J and
Anderson KS: Autophosphorylation of FGFR1 kinase is mediated by a
sequential and precisely ordered reaction. Mol Cell. 21:711–717.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Sahni M, Ambrosetti DC, Mansukhani A,
Gertner R, Levy D and Basilico C: FGF signaling inhibits
chondrocyte proliferation and regulates bone development through
the STAT-1 pathway. Genes Dev. 13:1361–1366. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Su WC, Kitagawa M, Xue N, Xie B, Garofalo
S, Cho J, Deng C, Horton WA and Fu XY: Activation of Stat1 by
mutant fibroblast growth-factor receptor in thanatophoric dysplasia
type II dwarfism. Nature. 386:288–292. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lamothe B, Yamada M, Schaeper U,
Birchmeier W, Lax I and Schlessinger J: The docking protein Gab1 is
an essential component of an indirect mechanism for fibroblast
growth factor stimulation of the phosphatidylinositol 3-kinase/Akt
antiapoptotic pathway. Mol Cell Biol. 24:5657–5666. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Kouhara H, Hadari YR, Spivak-Kroizman T,
Schilling J, Bar-Sagi D, Lax I and Schlessinger J: A lipid-anchored
Grb2-binding protein that links FGF-receptor activation to the
Ras/MAPK signaling pathway. Cell. 89:693–702. 1997. View Article : Google Scholar
|
|
75
|
Kanazawa S, Fujiwara T, Matsuzaki S,
Shingaki K, Taniguchi M, Miyata S, Tohyama M, Sakai Y, Yano K,
Hosokawa K and Kubo T: bFGF regulates PI3-kinase-Rac1-JNK pathway
and promotes fibroblast migration in wound healing. PLoS One.
5:e122282010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Liao S, Porter D, Scott A, Newman G,
Doetschman T and Schultz Jel J: The cardioprotective effect of the
low molecular weight isoform of fibroblast growth factor-2: the
role of JNK signaling. J Mol Cell Cardiol. 42:106–120. 2007.
View Article : Google Scholar
|
|
77
|
House SL, Branch K, Newman G, Doetschman T
and Schultz Jel J: Cardioprotection induced by cardiac-specific
overexpression of fibroblast growth factor-2 is mediated by the
MAPK cascade. Am J Physiol Heart Circ Physiol. 289:H2167–H2175.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Tsang M and Dawid IB: Promotion and
attenuation of FGF signaling through the Ras-MAPK pathway. Sci
STKE. 2004:pe172004. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Tan Y, Rouse J, Zhang A, Cariati S, Cohen
P and Comb MJ: FGF and stress regulate CREB and ATF-1 via a pathway
involving p38 MAP kinase and MAPKAP kinase-2. EMBO J. 15:4629–4642.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Moon AM, Guris DL, Seo JH, Li L, Hammond
J, Talbot A and Imamoto A: Crkl deficiency disrupts Fgf8 signaling
in a mouse model of 22q11 deletion syndromes. Dev Cell. 10:71–80.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Bogale DE: The roles of FGFR3 and c-MYC in
urothelial bladder cancer. Discov Oncol. 15:2952024. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Timsah Z, Ahmed Z, Lin CC, Melo FA, Stagg
LJ, Leonard PG, Jeyabal P, Berrout J, O'Neil RG, Bogdanov M and
Ladbury JE: Competition between Grb2 and Plcγ1 for FGFR2 regulates
basal phospholipase activity and invasion. Nat Struct Mol Biol.
21:180–188. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hanafusa H, Torii S, Yasunaga T and
Nishida E: Sprouty1 and Sprouty2 provide a control mechanism for
the Ras/MAPK signalling pathway. Nat Cell Biol. 4:850–858. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Torii S, Kusakabe M, Yamamoto T, Maekawa M
and Nishida E: Sef is a spatial regulator for Ras/MAP kinase
signaling. Dev Cell. 7:33–44. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Camps M, Nichols A, Gillieron C, Antonsson
B, Muda M, Chabert C, Boschert U and Arkinstall S: Catalytic
activation of the phosphatase MKP-3 by ERK2 mitogen-activated
protein kinase. Science. 280:1262–1265. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wong A, Lamothe B, Lee A, Schlessinger J
and Lax I: FRS2 alpha attenuates FGF receptor signaling by
Grb2-mediated recruitment of the ubiquitin ligase Cbl. Proc Natl
Acad Sci USA. 99:6684–6689. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Dufour C, Guenou H, Kaabeche K, Bouvard D,
Sanjay A and Marie PJ: FGFR2-Cbl interaction in lipid rafts
triggers attenuation of PI3K/Akt signaling and osteoblast survival.
Bone. 42:1032–1039. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Browaeys-Poly E, Blanquart C, Perdereau D,
Antoine AF, Goenaga D, Luzy JP, Chen H, Garbay C, Issad T, Cailliau
K and Burnol AF: Grb14 inhibits FGF receptor signaling through the
regulation of PLCγ recruitment and activation. FEBS Lett.
584:4383–4388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Huang SW, Yu HC, Huang HB, Chen PC and Lu
MC: Inhibiting pyruvate carboxylase as a potential therapeutic
strategy for alleviating rheumatoid arthritis.
Inflammopharmacology. 34:4239–4252. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Morice A, de La Seiglière A, Kany A,
Khonsari RH, Bensidhoum M, Puig-Lombardi ME and Legeai Mallet L:
FGFR antagonists restore defective mandibular bone repair in a
mouse model of osteochondrodysplasia. Bone Res. 13:122025.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Liao S, Li J, Gao S, Han Y, Han X, Wu Y,
Bi J, Xu M and Bi W: Sulfatinib, a novel multi-targeted tyrosine
kinase inhibitor of FGFR1, CSF1R, and VEGFR1-3, suppresses
osteosarcoma proliferation and invasion via dual role in tumor
cells and tumor microenvironment. Front Oncol. 13:11588572023.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wehrli M, Oppliger Leibundgut E, Gattiker
HH, Manz MG, Müller AM and Goede JS: Response to tyrosine kinase
inhibitors in myeloproliferative neoplasia with 8p11 translocation
and CEP110-FGFR1 rearrangement. Oncologist. 22:480–483. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Peng H, Myers J, Fang X, Stachowiak EK,
Maher PA, Martins GG, Popescu G, Berezney R and Stachowiak MK:
Integrative nuclear FGFR1 signaling (INFS) pathway mediates
activation of the tyrosine hydroxylase gene by angiotensin II,
depolarization and protein kinase C. J Neurochem. 81:506–524. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Sait SF, Bale T and Karajannis MA: FGFR
alterations and potential role of FGFR inhibitors in pediatric
brain tumors. Neurooncol Adv. 8(Suppl 2): ii31–ii40. 2025.
|
|
95
|
Pichler R, van Creij NCH, Mertens LS, Del
Giudice F, Koll F, Soria F, Subiela JD, Plage H, Tymoszuk P, Mayr
R, et al: FGFR1/3 signaling as an achilles' heel of phenotypic
diversity in urothelial carcinoma. Eur Urol Oncol. 9:220–226. 2026.
View Article : Google Scholar
|
|
96
|
Lu X, Chen H, Patterson AV, Smaill JB and
Ding K: Fibroblast growth factor receptor 4 (FGFR4) selective
inhibitors as hepatocellular carcinoma therapy: Advances and
prospects. J Med Chem. 62:2905–2915. 2019. View Article : Google Scholar
|
|
97
|
Katoh M: FGFR inhibitors: Effects on
cancer cells, tumor microenvironment and whole-body homeostasis
(Review). Int J Mol Med. 38:3–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Suzuki Y, Shimizu H, Ishizuka N, Kubota N,
Kubota T, Senoo A, Kageyama H, Osaka T, Hirako S, Kim HJ, et al:
Vagal hyperactivity due to ventromedial hypothalamic lesions
increases adiponectin production and release. Diabetes.
63:1637–1648. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Abdel-Mohsen HT, Ibrahim MA, Nageeb AM and
El Kerdawy AM: Receptor-based pharmacophore modeling, molecular
docking, synthesis and biological evaluation of novel VEGFR-2,
FGFR-1, and BRAF multi-kinase inhibitors. BMC Chem. 18:422024.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Roskoski R Jr: The role of fibroblast
growth factor receptor (FGFR) protein-tyrosine kinase inhibitors in
the treatment of cancers including those of the urinary bladder.
Pharmacol Res. 151:1045672020. View Article : Google Scholar
|
|
101
|
Yoon GH, Park DS, Kim MS and Choi SC:
Perillic acid disrupts the specification of germ layers by
upregulating the FGF/MAPK pathway. Genes Genomics. 47:637–649.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Orsmond A, Krishnan G, Palmer LJ, De Sousa
SMC and McCormack A: FGFR1 variation in the divergent settings of
congenital hypopituitarism and pituitary tumours. Pituitary.
28:392025. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Clark JF and Soriano P: Diverse Fgfr1
signaling pathways and endocytic trafficking regulate mesoderm
development. Genes Dev. 38:393–414. 2024.PubMed/NCBI
|
|
104
|
Thisse B and Thisse C: Functions and
regulations of fibroblast growth factor signaling during embryonic
development. Dev Biol. 287:390–402. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Lu W, Jiang M, Zhuang J, Song J, Zhou C,
Zhou Y, Zhu Z, Wu A, Sheng S, Zhu S and Wang Z: FGF4 activates
FGFR1 - PI3K/AKT signaling to enhance Clec10a-mediated
intracellular myelin debris processing and promote spinal cord
repair. J Neuroinflammation. 23:1072026. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Yang X, Li X, Zhang X, Meng L, Wang J and
Zheng N: Comparative evaluation of the short-chain fatty acids
formate, propionate, and valerate on intestinal barrier maturation
and bone development in neonatal mice. Food Funct. 17:3372–3385.
2026. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Sawano K, Nagasaki K, Suzuki E, Ogiwara Y,
Kageyama I, Fukami M and Kuroki Y: De novo retrotransposon
insertion into the FGFR1 gene in a boy with congenital
hypogonadotropic hypogonadism: A case report. Front Endocrinol
(Lausanne). 16:15653162025. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Tuokkola JE and Schwertfeger KL: Breast
cancer progression by the FGF/FGFR axis: A metabolic perspective. J
Mammary Gland Biol Neoplasia. 31:22025. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wu AL, Kolumam G, Stawicki S, Chen Y, Li
J, Zavala-Solorio J, Phamluong K, Feng B, Li L, Marsters S, et al:
Amelioration of type 2 diabetes by antibody-mediated activation of
fibroblast growth factor receptor 1. Sci Transl Med. 3:113ra262011.
View Article : Google Scholar
|
|
110
|
Stamou MI, Chiu CJ, Jadhav SV, Lopes VF,
Salnikov KB, Plummer L, Lippincott MF, Lee H, Seminara SB and
Balasubramanian R: Defective FGFR1 signaling disrupts glucose
regulation: Evidence from humans with FGFR1 mutations. J Endocr
Soc. 8:bvae1182024. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Seki D and Honda M: FGF-2-Overexpressing
adipose-derived stem cells as a paracrine platform for
angiogenesis-driven tissue regeneration. Cell Mol Bioeng. 19:73–87.
2026. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Feng Y, Chen B, Xu Y, Xu X, Hu B, Li T,
Zhang D, Bai B, Liu C, Xu W, et al: Multifunctional bioactive
scaffold facilitating BMSCs-Driven osteogenesis and vascularization
in critical-sized bone defect repair. Adv Sci (Weinh).
13:e226922026. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Dong Q, Tao J, Yuan J, Tian J, Cheng Y,
Wang J, Xiao C, Chen X, Wang Y, Li W and Shen F: FGF20 activates
FGFR1-PI3K-AKT signaling to coordinate barrier integrity and
alveolar coagulation in sepsis-induced lung injury. Cell Signal.
145:1125742026. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Seitz T and Hellerbrand C: Role of
fibroblast growth factor signalling in hepatic fibrosis. Liver Int.
41:1201–1215. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Kim S, An S, Lee J, Jeong Y, You CL, Kim
H, Bae JH, Yun CE, Ryu D, Bae GU and Kang JS: Cdon ablation in
motor neurons causes age-related motor neuron degeneration and
impaired sciatic nerve repair. J Cachexia Sarcopenia Muscle.
14:2239–2252. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Meng J, Lv Z, Chen X, Sun C, Jin C, Ding K
and Chen C: LBP1C-2 from Lycium barbarum maintains skeletal muscle
satellite cell pool by interaction with FGFR1. iScience.
26:1065732023. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Farooq M, Hwang M, Khan AW, Batool M,
Ahmad B, Kim W, Kim MS and Choi S: Identification of a novel
fibroblast growth factor receptor-agonistic peptide and its effect
on diabetic wound healing. Life Sci. 364:1234322025. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Lee SB, Abdal Dayem A, Kmiecik S, Lim KM,
Seo DS, Kim HT, Kumar Biswas P, Do M, Kim DH and Cho SG: Efficient
improvement of the proliferation, differentiation, and
anti-arthritic capacity of mesenchymal stem cells by simply
culturing on the immobilized FGF2 derived peptide,
44-ERGVVSIKGV-53. J Adv Res. 62:119–141. 2024. View Article : Google Scholar :
|
|
119
|
Eng L, Azad AK, Habbous S, Pang V, Xu W,
Maitlandvan der Zee AH, Savas S, Mackay HJ, Amir E and Liu G:
Vascular endothelial growth factor pathway polymorphisms as
prognostic and pharmacogenetic factors in cancer: A systematic
review and meta-analysis. Clin Cancer Res. 18:4526–4537. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Michel ZD, Aitken SF, Glover OD, Alejandro
LO, Randazzo D, Dambkowski C, Martin D, Collins MT, Somerman MJ and
Chu EY: Infigratinib, a selective FGFR1-3 tyrosine kinase
inhibitor, alters dentoalveolar development at high doses. Dev Dyn.
252:1428–1448. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Grinspon RP, Castro S, Brunello FG, Sansó
G, Ropelato MG and Rey RA: Diagnosis of male central hypogonadism
during childhood. J Endocr Soc. 5:bvab1452021. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Lu Y, Li B, Zheng X, Xu L, Zeng L, Zhang C
and Zhang J: Discovery and characterization of novel FGFR1 V561M
inhibitors via virtual screening and molecular dynamics
simulations. ACS Med Chem Lett. 16:1575–1584. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Lv M, Shangguan W, Zhao Q, Li Y, Zhao Q
and Li F: Protein kinase B inhibitors enhance the sensitivity of
translocated promoter region-fibroblast growth factor receptor 1
cells to fibroblast growth factor receptor 1 inhibitor-induced
apoptosis. J Int Med Res. 53:30006052513629682025. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Qin S, Sun D, Li H, Li X, Pan W, Yan C,
Tang R and Liu X: The Effect of SHH-Gli signaling pathway on the
synovial fibroblast proliferation in rheumatoid arthritis.
Inflammation. 39:503–512. 2016. View Article : Google Scholar
|
|
125
|
Kschonsak YT, Liang WC, Chan J, Kan D,
Nguyen T, Ruiz K, Kee YS, Tsai WK, Xi H, Moskalenko M, et al:
Tumor-targeted bispecific antibodies effectively inhibit oncogenic
pathways while minimizing toxicity. Sci Adv. 12:eadx39592026.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Compton JT and Lee FY: A review of
osteocyte function and the emerging importance of sclerostin. J
Bone Joint Surg Am. 96:1659–1668. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Chen LC, Shibu MA, Liu CJ, Han CK, Ju DT,
Chen PY, Viswanadha VP, Lai CH, Kuo WW and Huang CY: ERK1/2
mediates the lipopolysaccharide-induced upregulation of FGF-2, uPA,
MMP-2, MMP-9 and cellular migration in cardiac fibroblasts. Chem
Biol Interact. 306:62–69. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Wang K, Ji W, Yu Y, Li Z, Niu X, Xia W and
Lu S: FGFR1-ERK1/2-SOX2 axis promotes cell proliferation,
epithelial-mesenchymal transition, and metastasis in
FGFR1-amplified lung cancer. Oncogene. 37:5340–5354. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
McKenzie J, Smith C, Karuppaiah K,
Langberg J, Silva MJ and Ornitz DM: Osteocyte death and bone
overgrowth in mice lacking fibroblast growth factor receptors 1 and
2 in mature osteoblasts and osteocytes. J Bone Miner Res.
34:1660–1675. 2019. View Article : Google Scholar
|
|
130
|
O'Shea PJ, Guigon CJ, Williams GR and
Cheng SY: Regulation of fibroblast growth factor receptor-1 (FGFR1)
by thyroid hormone: identification of a thyroid hormone response
element in the murine Fgfr1 promoter. Endocrinology. 148:5966–5976.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Florez-Martin M, Radu AM, Shakib K and
Jell G: Osteoblastic cell responses to zoledronate treatment
in-vitro: A systematic review. Bone. 205:1177962026. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Salhotra A, Shah HN, Levi B and Longaker
MT: Mechanisms of bone development and repair. Nat Rev Mol Cell
Biol. 21:696–711. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
White KE, Cabral JM, Davis SI, Fishburn T,
Evans WE, Ichikawa S, Fields J, Yu X, Shaw NJ, McLellan NJ, et al:
Mutations that cause osteoglophonic dysplasia define novel roles
for FGFR1 in bone elongation. Am J Hum Genet. 76:361–367. 2005.
View Article : Google Scholar :
|
|
134
|
Wilkie AO: Bad bones, absent smell,
selfish testes: The pleiotropic consequences of human FGF receptor
mutations. Cytokine Growth Factor Rev. 16:187–203. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Jacob AL, Smith C, Partanen J and Ornitz
DM: Fibroblast growth factor receptor 1 signaling in the
osteo-chondrogenic cell lineage regulates sequential steps of
osteoblast maturation. Dev Biol. 296:315–328. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Maruyama T, Mirando AJ, Deng CX and Hsu W:
The balance of WNT and FGF signaling influences mesenchymal stem
cell fate during skeletal development. Sci Signal. 3:ra402010.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Rice DP, Rice R and Thesleff I: Fgfr mRNA
isoforms in craniofacial bone development. Bone. 33:14–27. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Zhang Y, Su N, Luo F, Wen X, Tang Y, Yang
J, Chen S, Jiang W, Du X and Chen L: Deletion of Fgfr1 in
osteoblasts enhances mobilization of EPCs into peripheral blood in
a mouse endotoxemia model. Int J Biol Sci. 10:1064–1071. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Xiao Z, Huang J, Cao L, Liang Y, Han X and
Quarles LD: Osteocyte-specific deletion of Fgfr1 suppresses FGF23.
PLoS One. 9:e1041542014. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Lee Y, Bae KJ, Chon HJ, Kim SH, Kim SA and
Kim J: A receptor tyrosine kinase inhibitor, dovitinib (TKI-258),
Enhances BMP-2-Induced osteoblast differentiation in vitro. Mol
Cells. 39:389–394. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Wu AL, Feng B, Chen MZ, Kolumam G,
Zavala-Solorio J, Wyatt SK, Gandham VD, Carano RA and Sonoda J:
Antibody-mediated activation of FGFR1 induces FGF23 production and
hypophosphatemia. PLoS One. 8:e573222013. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Chen ZH, Wu JJ, Guo DY, Li YY, Chen MN,
Zhang ZY, Yuan ZD, Zhang KW, Chen WW, Tian F, et al: Physiological
functions of podosomes: From structure and function to therapy
implications in osteoclast biology of bone resorption. Ageing Res
Rev. 85:1018422023. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Chen Z, Encarnacion AM, Rajan RPS, Yao H,
Lee S, Kim E and Lee TH: Discovery of a novel homoisoflavonoid
derivative 5g for anti-osteoclastic bone loss via targeting FGFR1.
Eur J Med Chem. 270:1163352024. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Kawaguchi H, Chikazu D, Nakamura K,
Kumegawa M and Hakeda Y: Direct and indirect actions of fibroblast
growth factor 2 on osteoclastic bone resorption in cultures. J Bone
Miner Res. 15:466–473. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Chikazu D, Hakeda Y, Ogata N, Nemoto K,
Itabashi A, Takato T, Kumegawa M, Nakamura K and Kawaguchi H:
Fibroblast growth factor (FGF)-2 directly stimulates mature
osteoclast function through activation of FGF receptor 1 and
p42/p44 MAP kinase. J Biol Chem. 275:31444–31450. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Nakajima A, Nakajima F, Shimizu S,
Ogasawara A, Wanaka A, Moriya H, Einhorn TA and Yamazaki M: Spatial
and temporal gene expression for fibroblast growth factor type I
receptor (FGFR1) during fracture healing in the rat. Bone.
29:458–466. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Lu X, Su N, Yang J, Huang W, Li C, Zhao L,
He Q, Du X, Shen Y, Chen B and Chen L: Fibroblast growth factor
receptor 1 regulates the differentiation and activation of
osteoclasts through Erk1/2 pathway. Biochem Biophys Res Commun.
390:494–499. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Aukes K, Forsman C, Brady NJ, Astleford K,
Blixt N, Sachdev D, Jensen ED, Mansky KC and Schwertfeger KL:
Breast cancer cell-derived fibroblast growth factors enhance
osteoclast activity and contribute to the formation of metastatic
lesions. PLoS One. 12:e01857362017. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
O'Hagan A, Siefker-Radtke A, Loriot Y,
Deprince K, Crow L, Laron M, Adelman R, Sweiti H and Triantos S:
Incidence and outcomes of FGFR inhibitor-associated retinopathy of
patients treated with oral erdafitinib across the clinical trial
program. Oncologist. 31:oyag1742026. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Zhang Y, Ling L, Ajay D/OAA, Eio YM, van
Wijnen AJ, Nurcombe V and Cool SM: FGFR2 accommodates osteogenic
cell fate determination in human mesenchymal stem cells. Gene.
818:1461992022. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Michigami T: Advances in understanding of
phosphate homeostasis and related disorders. Endocr J. 69:881–896.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Karolak MR, Yang X and Elefteriou F: FGFR1
signaling in hypertrophic chondrocytes is attenuated by the Ras-GAP
neurofibromin during endochondral bone formation. Hum Mol Genet.
24:2552–2564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Tuzon CT, Rigueur D and Merrill AE:
Nuclear fibroblast growth factor receptor signaling in skeletal
development and disease. Curr Osteoporos Rep. 17:138–146. 2019.
View Article : Google Scholar
|
|
154
|
Wang Z, Huang Y, He Y, Khor S, Zhong X,
Xiao J, Ye Q and Li X: Myocardial protection by heparin-based
coacervate of FGF10. Bioact Mater. 6:1867–1877. 2020.PubMed/NCBI
|
|
155
|
Song H, Du H, Li J, Wang M, Wang J, Ju X
and Mu W: Effect of fibroblast growth factor 2 on degenerative
endplate chondrocyte: From anabolism to catabolism. Exp Mol Pathol.
118:1045902021. View Article : Google Scholar
|
|
156
|
Tang J, Su N, Zhou S, Xie Y, Huang J, Wen
X, Wang Z, Wang Q, Xu W, Du X, et al: Fibroblast growth factor
receptor 3 inhibits osteoarthritis progression in the knee joints
of adult mice. Arthritis Rheumatol. 68:2432–2443. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Weng T, Yi L, Huang J, Luo F, Wen X, Du X,
Chen Q, Deng C, Chen D and Chen L: Genetic inhibition of fibroblast
growth factor receptor 1 in knee cartilage attenuates the
degeneration of articular cartilage in adult mice. Arthritis Rheum.
64:3982–3992. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Shen J, Li J, Wang B, Jin H, Wang M, Zhang
Y, Yang Y, Im HJ, O'Keefe R and Chen D: Deletion of the
transforming growth factor β receptor type II gene in articular
chondrocytes leads to a progressive osteoarthritis-like phenotype
in mice. Arthritis Rheum. 65:3107–3119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Zhou YX, Xu X, Chen L, Li C, Brodie SG and
Deng CX: A Pro250Arg substitution in mouse Fgfr1 causes increased
expression of Cbfa1 and premature fusion of calvarial sutures. Hum
Mol Genet. 9:2001–2008. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Wang Z, Huang J, Zhou S, Luo F, Tan Q, Sun
X, Ni Z, Chen H, Du X, Xie Y and Chen L: Loss of Fgfr1 in
chondrocytes inhibits osteoarthritis by promoting autophagic
activity in temporomandibular joint. J Biol Chem. 293:8761–8774.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Gao Z, Hou Y, Kong F and Hou S:
Downregulation of microRNA-574-3p expression in human lumbar disc
degeneration and its improvement of degenerative changes in nucleus
pulposus cells via targeting FGFR3. Arch Biochem Biophys.
782:1108462026. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Yao X, Zhang J, Jing X, Ye Y, Guo J, Sun K
and Guo F: Fibroblast growth factor 18 exerts anti-osteoarthritic
effects through PI3K-AKT signaling and mitochondrial fusion and
fission. Pharmacol Res. 139:314–324. 2019. View Article : Google Scholar
|
|
163
|
Delucchi Á, Toro L, Alzamora R, Barrientos
V, González M, Andaur R, León P, Villanueva F, Galindo M, Las Heras
F, et al: Glucocorticoids decrease longitudinal bone growth in
pediatric kidney transplant recipients by stimulating the
FGF23/FGFR3 signaling pathway. J Bone Miner Res. 34:1851–1861.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Davidson D, Blanc A, Filion D, Wang H,
Plut P, Pfeffer G, Buschmann MD and Henderson JE: Fibroblast growth
factor (FGF) 18 signals through FGF receptor 3 to promote
chondrogenesis. J Biol Chem. 280:20509–20515. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Valverde-Franco G, Binette JS, Li W, Wang
H, Chai S, Laflamme F, Tran-Khanh N, Quenneville E, Meijers T,
Poole AR, et al: Defects in articular cartilage metabolism and
early arthritis in fibroblast growth factor receptor 3 deficient
mice. Hum Mol Genet. 15:1783–1792. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Peng Z, Gao L, Zhang L, Yao R, Li X, Deng
L, Fan J, Ying L and Wang Y: Development of FGF21 mutant with
potent cardioprotective effects in T2D Mice via FGFR1-AMPK-Mediated
inhibition of oxidative stress. Int J Mol Sci. 26:65772025.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Reed JR, Stone MD, Beadnell TC, Ryu Y,
Griffin TJ and Schwertfeger KL: Fibroblast growth factor receptor 1
activation in mammary tumor cells promotes macrophage recruitment
in a CX3CL1-dependent manner. PLoS One. 7:e458772012. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Zhao YN, Liu ZD, Yan T, Xu TX, Jin TY,
Jiang YS, Zuo W, Lee KY, Huang LJ and Wang Y: Macrophage-specific
FGFR1 deletion alleviates high-fat-diet-induced liver inflammation
by inhibiting the MAPKs/TNF pathways. Acta Pharmacol Sin.
45:988–1001. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Bohrer LR and Schwertfeger KL: Macrophages
promote fibroblast growth factor receptor-driven tumor cell
migration and invasion in a CXCR2-dependent manner. Mol Cancer Res.
10:1294–1305. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Yan M, Yang H, He W, Lei H, Feng N, Guan
W, Feng W and Han L: PZP deficiency impairs M2 macrophage
polarization via TGF-β/Smad3 signaling, contributing to immune
dysregulation at the maternal-fetal interface in preeclampsia. Mol
Immunol. 194:1–12. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Haolan, Wang Y, Gao Y, Liu Y, Deng B, Li
P, Guo J and Sun Z: Esketamine attenuated sepsis-induced organ
injury in mice via the TGF-β-Smad3 signaling pathway. Cytokine.
203:1571632026. View Article : Google Scholar
|
|
172
|
Wu JJ, Sun ZL, Liu SY, Chen ZH, Yuan ZD,
Zou ML, Teng YY, Li YY, Guo DY and Yuan FL: The ASIC3-M-CSF-M2
macrophage-positive feedback loop modulates
fibroblast-to-myofibroblast differentiation in skin fibrosis
pathogenesis. Cell Death Dis. 13:5272022. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Davidson S, Coles M, Thomas T, Kollias G,
Ludewig B, Turley S, Brenner M and Buckley CD: Fibroblasts as
immune regulators in infection, inflammation and cancer. Nat Rev
Immunol. 21:704–717. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Marsh LJ, Kemble S, Reis Nisa P, Singh R
and Croft AP: Fibroblast pathology in inflammatory joint disease.
Immunol Rev. 302:163–183. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Komatsu N and Takayanagi H: Mechanisms of
joint destruction in rheumatoid arthritis - immune
cell-fibroblast-bone interactions. Nat Rev Rheumatol. 18:415–429.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Buckley CD, Ospelt C, Gay S and Midwood
KS: Author correction: Location, location, location: how the tissue
microenvironment affects inflammation in RA. Nat Rev Rheumatol.
17:2462021. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Bartok B and Firestein GS: Fibroblast-like
synoviocytes: key effector cells in rheumatoid arthritis. Immunol
Rev. 233:233–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Danks L, Komatsu N, Guerrini MM, Sawa S,
Armaka M, Kollias G, Nakashima T and Takayanagi H: RANKL expressed
on synovial fibroblasts is primarily responsible for bone erosions
during joint inflammation. Ann Rheum Dis. 75:1187–1195. 2016.
View Article : Google Scholar
|
|
179
|
Lang J, Li L, Quan Y, Tan R, Zhao J, Li M,
Zeng J, Chen S, Wang T, Li Y, et al: LC-MS-based metabolomics
reveals the mechanism of anti-gouty arthritis effect of Wuwei
Shexiang pill. Front Pharmacol. 14:12136022023. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Gao X, Zhu X, Lin Y, Liu C, Wang Q and Lin
X: Targeting 5-methylcytosine RNA methylation-mediated
CX26/IκBα/NF-κB signaling by salvianolic acid B attenuates
rheumatoid arthritis progression. Phytomedicine. 157:1583002026.
View Article : Google Scholar
|
|
181
|
Li S, Li F, Xu M, Cheng N, Wan L and Zhang
X: m6A-associated GAS6 expression is associated with
pathogenic activation of fibroblast-like synoviocytes in rheumatoid
arthritis. Sci Rep. May 21–2026.Epub ahead of print.
|
|
182
|
Tang C, Wang Z, Xie Y, Fei Y, Luo J, Wang
C, Ying Y, He P, Yan R, Chen Y, et al: Classification of distinct
tendinopathy subtypes for precision therapeutics. Nat Commun.
15:94602024. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Nygaard G and Firestein GS: Restoring
synovial homeostasis in rheumatoid arthritis by targeting
fibroblast-like synoviocytes. Nat Rev Rheumatol. 16:316–333. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Qiao Z, Meng Q, Xiao B, Long Y, Liang Q
and Su T: Brevilin A, a novel LRRC15 inhibitor, exerts potent
anti-rheumatoid arthritis effects by inhibiting the LRRC15/STAT3
signaling pathway. Arthritis Res Ther. 27:1672025. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Chang CJ, Chang TM, Sun YS, Lin JF, Lee
CW, Fang ML and Liu JF: Lutein attenuates rheumatoid arthritis
progression by suppressing MAPK/NF-κB signaling and MMP3 and MMP13
expression in fibroblast-like Synoviocytes. Int Immunopharmacol.
164:1153602025. View Article : Google Scholar
|
|
186
|
Liu J, Zhang C, Ma R, Yin C, Ren J, Yang
S, Zhao Y, Tang Y, Wei J and Li X: Knockdown of MiD49 and MiD51
alleviates collagen-induced arthritis and suppresses mitophagy and
fatty acid oxidation (FAO) in rheumatoid arthritis fibroblast-like
synoviocytes. Free Radic Biol Med. 240:514–531. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Gao X, Lyu S, Lin T, He J, Pan W, He J,
Wang X, Lin X, Chen J and Wang Q: Oxyimperatorin suppresses the
pathologies of rheumatoid arthritis and the growth of synovial
organoids through targeting the PI3K/AKT Pathway. Phytother Res.
39:3526–3544. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Wang Q, Peng X, Xu H, Zhao Y, Lu X, Lu C,
Wang Q, Lu W, Sheng Q, Lu X, et al: Narirutin mitigates
inflammatory arthritis and osteoporosis through modulating
macrophage phenotype and osteoclastogenesis. J Orthop Translat.
54:115–130. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Wang W, Wang Z, Ling A, Zhang C, Lv M,
Huang L and Niu Y: Research progress in treatment of rheumatoid
arthritis with Sinomenine and related formulations based on
different administration routes. Front Pharmacol. 16:16136792025.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Yue S, Fan J, Xie D, Cao C, Wang Z, Huang
J, Qiu F, Yang X, He D, Lu A and Liang C: Unveiling the therapeutic
potential: Targeting fibroblast-like synoviocytes in rheumatoid
arthritis. Expert Rev Mol Med. 27:e182025. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
DeSalvo J, Ban Y, Li L, Sun X, Jiang Z,
Kerr DA, Khanlari M, Boulina M, Capecchi MR, Partanen JM, et al:
ETV4 and ETV5 drive synovial sarcoma through cell cycle and DUX4
embryonic pathway control. J Clin Invest. 131:e1419082021.
View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Shoda J, Tanaka S, Etori K, Hattori K,
Kasuya T, Ikeda K, Maezawa Y, Suto A, Suzuki K, Nakamura J, et al:
Semaphorin 3G exacerbates joint inflammation through the
accumulation and proliferation of macrophages in the synovium.
Arthritis Res Ther. 24:1342022. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Zhao XM, Byrd VM, McKeehan WL, Reich MB,
Miller GG and Thomas JW: Costimulation of human CD4+ T cells by
fibroblast growth factor-1 (acidic fibroblast growth factor). J
Immunol. 155:3904–3911. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Gould PW, Zemel BS, Taratuta EG and Baker
JF: Circulating fibroblast growth factor-21 levels in rheumatoid
arthritis: Associations with disease characteristics, body
composition, and physical functioning. J Rheumatol. 48:504–512.
2021. View Article : Google Scholar
|
|
195
|
Sato H, Kazama JJ, Murasawa A, Otani H,
Abe A, Ito S, Ishikawa H, Nakazono K, Kuroda T, Nakano M and Narita
I: Serum fibroblast growth factor 23 (FGF23) in patients with
rheumatoid arthritis. Intern Med. 55:121–126. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Manabe N, Oda H, Nakamura K, Kuga Y,
Uchida S and Kawaguchi H: Involvement of fibroblast growth factor-2
in joint destruction of rheumatoid arthritis patients. Rheumatology
(Oxford). 38:714–720. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Byrd V, Zhao XM, McKeehan WL, Miller GG
and Thomas JW: Expression and functional expansion of fibroblast
growth factor receptor T cells in rheumatoid synovium and
peripheral blood of patients with rheumatoid arthritis. Arthritis
Rheum. 39:914–922. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Li L, Sun Y, Luo J and Liu M: Circulating
immune cells and risk of osteosarcoma: A Mendelian randomization
analysis. Front Immunol. 15:13812122024. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Balogh E, Biniecka M, Fearon U, Veale DJ
and Szekanecz Z: Angiogenesis in inflammatory arthritis. Isr Med
Assoc J. 21:345–352. 2019.PubMed/NCBI
|
|
200
|
Wang Y, Wu H and Deng R: Angiogenesis as a
potential treatment strategy for rheumatoid arthritis. Eur J
Pharmacol. 910:1745002021. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Leblond A, Allanore Y and Avouac J:
Targeting synovial neoangiogenesis in rheumatoid arthritis.
Autoimmun Rev. 16:594–601. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Zhang YF, Gao SS, Li JL, Zuo WS, Qiu YW
and Xiao YC: Comparison and correlation study of synovial
ultrasound indices and serum VEGF in rheumatoid wrist arthritis
before and after treatment. Clin Rheumatol. 41:2677–2683. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Tan Q, Wang Z, Wang Q, Wang Y, Huang Z, Su
N, Jin M, Kuang L, Qi H, Ni Z, et al: A novel FGFR1-binding peptide
exhibits anti-tumor effect on lung cancer by inhibiting
proliferation and angiogenesis. Int J Biol Sci. 14:1389–1398. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Zhu X, Qiu C, Wang Y, Jiang Y, Chen Y, Fan
L, Ren R, Wang Y, Chen Y, Feng Y, et al: FGFR1 SUMOylation
coordinates endothelial angiogenic signaling in angiogenesis. Proc
Natl Acad Sci USA. 119:e22026311192022. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Lee C, Chen R, Sun G, Liu X, Lin X, He C,
Xing L, Liu L, Jensen LD, Kumar A, et al: VEGF-B prevents excessive
angiogenesis by inhibiting FGF2/FGFR1 pathway. Signal Transduct
Target Ther. 8:3052023. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Wang Y, Wu H, Gui BJ, Liu J, Rong GX, Deng
R, Bu YH and Zhang H: Geniposide alleviates VEGF-induced
angiogenesis by inhibiting VEGFR2/PKC/ERK1/2-mediated SphK1
translocation. Phytomedicine. 100:1540682022. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Luo J, Liang Y, Kong F, Qiu J, Liu X, Chen
A, Luxon BA, Wu HW and Wang Y: Vascular endothelial growth factor
promotes the activation of hepatic stellate cells in chronic
schistosomiasis. Immunol Cell Biol. 95:399–407. 2017. View Article : Google Scholar
|
|
208
|
Wu Q, Han L, Gui W, Wang F, Yan W and
Jiang H: MiR-503 suppresses fibroblast activation and myofibroblast
differentiation by targeting VEGFA and FGFR1 in silica-induced
pulmonary fibrosis. J Cell Mol Med. 24:14339–14348. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Gao M, Li X, Yang M, Feng W, Lin Y and He
T: TNFAIP3 mediates FGFR1 activation-induced breast cancer
angiogenesis by promoting VEGFA expression and secretion. Clin
Transl Oncol. 24:2453–2465. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Oberkersch RE, Pontarin G, Astone M,
Spizzotin M, Arslanbaeva L, Tosi G, Panieri E, Ricciardi S, Allega
MF, Brossa A, et al: Aspartate metabolism in endothelial cells
activates the mTORC1 pathway to initiate translation during
angiogenesis. Dev Cell. 57:1241–1256.e8. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Nourse MB, Rolle MW, Pabon LM and Murry
CE: Selective control of endothelial cell proliferation with a
synthetic dimerizer of FGF receptor-1. Lab Invest. 87:828–835.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Oliva-Ferrusola E, Baus-Domínguez M,
Torres-Lagares D and Serrera-Figallo MA: Genetic aspects of dental
impaction: A scoping review. Genes (Basel). 17:2652026. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Čemažar L, Šlajpah K, Gredelj Šimec N and
Podgornik H: A case report and a literature review of
myeloid/lymphoid neoplasm with eosinophilia and PCM1::JAK2
rearrangement representing as B-cell acute lymphoblastic leukemia
B-ALL. Ann Hematol. 105:52026. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Li D, Yan M, Yang M, Yu Y, Bai X, He Y,
Liu K and Fan H: 20(S)-PPD-HSA NPs enhance the recovery of
intramedullary and extramedullary hematopoiesis in
cyclophosphamide-treated mice through activation of the FGFR1/ERK
pathway. Int Immunopharmacol. 170:1160732026. View Article : Google Scholar
|
|
215
|
Huang Y, Liao J, Shen P, He Y, Sun F,
Zhang Q, Zheng C, Zhang X, Li H and Chen G: Nf2/FGFR1/AKT axis
directs cranial neural crest-derived skull morphogenesis via
collagen synthesis and trafficking. JCI Insight. 10:e1911122025.
View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Zhang X and Xu J: Fibroblast growth factor
23-mediated regulation of osteoporosis: Assessed via Mendelian
randomization and in vitro study. J Cell Mol Med. 28:e185512024.
View Article : Google Scholar
|
|
217
|
Hagan AS, Boylan M, Smith C,
Perez-Santamarina E, Kowalska K, Hung IH, Lewis RM, Hajihosseini
MK, Lewandoski M and Ornitz DM: Generation and validation of novel
conditional flox and inducible Cre alleles targeting fibroblast
growth factor 18 (Fgf18). Dev Dyn. 248:882–893. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Freeman KW, Gangula RD, Welm BE, Ozen M,
Foster BA, Rosen JM, Ittmann M, Greenberg NM and Spencer DM:
Conditional activation of fibroblast growth factor receptor (FGFR)
1, but not FGFR2, in prostate cancer cells leads to increased
osteopontin induction, extracellular signal-regulated kinase
activation, and in vivo proliferation. Cancer Res. 63:6237–6243.
2003.PubMed/NCBI
|
|
219
|
Pendurthi UR, Rao LV, Williams JT and
Idell S: Regulation of tissue factor pathway inhibitor expression
in smooth muscle cells. Blood. 94:579–586. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Pond AC, Herschkowitz JI, Schwertfeger KL,
Welm B, Zhang Y, York B, Cardiff RD, Hilsenbeck S, Perou CM,
Creighton CJ, et al: Fibroblast growth factor receptor signaling
dramatically accelerates tumorigenesis and enhances oncoprotein
translation in the mouse mammary tumor virus-Wnt-1 mouse model of
breast cancer. Cancer Res. 70:4868–4879. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Stubbs JR, He N, Idiculla A, Gillihan R,
Liu S, David V, Hong Y and Quarles LD: Longitudinal evaluation of
FGF23 changes and mineral metabolism abnormalities in a mouse model
of chronic kidney disease. J Bone Miner Res. 27:38–46. 2012.
View Article : Google Scholar
|
|
222
|
Yuan B, Feng JQ, Bowman S, Liu Y, Blank
RD, Lindberg I and Drezner MK: Hexa-D-arginine treatment increases
7B2•PC2 activity in hyp-mouse osteoblasts and rescues the HYP
phenotype. J Bone Miner Res. 28:56–72. 2013. View Article : Google Scholar
|
|
223
|
Smolen JS, Landewé RBM, Bergstra SA,
Kerschbaumer A, Sepriano A, Aletaha D, Caporali R, Edwards CJ,
Hyrich KL, Pope JE, et al: EULAR recommendations for the management
of rheumatoid arthritis with synthetic and biological
disease-modifying antirheumatic drugs: 2022 update. Ann Rheum Dis.
82:3–18. 2023. View Article : Google Scholar
|
|
224
|
Firestein GS and McInnes IB:
Immunopathogenesis of rheumatoid arthritis. Immunity. 46:183–196.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Karabhari A, Rathee S, Soni S and Patil
UK: Rheumatoid arthritis: A comprehensive review of etiology,
pathophysiology, and innovative therapeutic advancements. Curr
Pharm Des. May 8–2026.Epub ahead of print. View Article : Google Scholar
|
|
226
|
Cruciani G, Cafaro G, Perricone C, Dal
Pozzolo R, Bruno L, Tromby F, Colangelo A, Gerli R and Bartoloni E:
Sequential treatment strategies following methotrexate inadequate
response in rheumatoid arthritis: A real-world retrospective cohort
study. Clin Exp Rheumatol. May 13–2026.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Chen Y, Zhou YF, Long ZY, Peng YM, Wu SX,
Peng F, Yuan MH, Zhou YZ, Wang L and Yuan H: Therapeutic potential
of spermidine in rheumatoid arthritis: Mechanisms and future
directions. Int J Rheum Dis. 29:e706102026. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Leiva-Escobar I, Chennas AP, Alayash Z,
Reckelkamm SL, Holtfreter B, Kocher T, Baumeister SE and Nolde M:
Nonsteroidal anti-inflammatory drugs and the risk of periodontitis
among adults with osteoarthritis: A target trial emulation. J Clin
Periodontol. May 23–2026.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
229
|
Konzett V, Laskou F, Smolen JS, Edwards
CJ, Aletaha D, van der Heijde D, Winthrop KL, Takeuchi T, Caporali
R, Verschueren P, et al: Efficacy of synthetic and biological
DMARDs: A systematic literature review informing the 2025 update of
the EULAR recommendations for the management of rheumatoid
arthritis. Ann Rheum Dis. 85:1039–1054. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Ellis H, Balasooriya ER, Varkaris A,
Hajian B, Wan J, Shekhar M, Gritti I, Vijay V, Albertelli L, Piot
S, et al: Mechanisms of clinical resistance to selective FGFR2
inhibition by lirafugratinib. Ann Oncol. 37:798–812. 2026.
View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Suzuki R, Watanabe S, Shono K, Masuda T,
Yanai K, Yamazaki R, Ando Y, Wakabayashi T, Tanaka S, Sekiya T, et
al: Nintedanib enhances the antitumor efficacy of pd-1 blockade,
potentially through inhibition of myeloid-derived suppressor cells
and cancer-associated fibroblasts. Cancer Immunol Immunother.
75:942026. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Boggavarapu RK and Chimakurthy J:
Preclinical pharmacokinetics of erdafitinib, an FGFR tyrosine
kinase inhibitor, via LC-MS/MS in sprague dawley rats. Rapid Commun
Mass Spectrom. 40:e701162026. View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Pirvola U, Ylikoski J, Trokovic R, Hébert
JM, McConnell SK and Partanen J: FGFR1 is required for the
development of the auditory sensory epithelium. Neuron. 35:671–680.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
234
|
Wang C, Chang JY, Yang C, Huang Y, Liu J,
You P, McKeehan WL, Wang F and Li X: Type 1 fibroblast growth
factor receptor in cranial neural crest cell-derived mesenchyme is
required for palatogenesis. J Biol Chem. 288:22174–22183. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Kawai M, Herrmann D, Fuchs A, Cheng S,
Ferrer-Vaquer A, Götz R, Driller K, Neubüser A and Ohura K: Fgfr1
conditional-knockout in neural crest cells induces heterotopic
chondrogenesis and osteogenesis in mouse frontal bones. Med Mol
Morphol. 52:156–163. 2019. View Article : Google Scholar
|
|
236
|
Takamori K, Hosokawa R, Xu X, Deng X,
Bringas P Jr and Chai Y: Epithelial fibroblast growth factor
receptor 1 regulates enamel formation. J Dent Res. 87:238–243.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Pei F, Ma L, Guo T, Zhang M, Jing J, Wen
Q, Feng J, Lei J, He J, Janečková E, et al: Sensory nerve regulates
progenitor cells via FGF-SHH axis in tooth root morphogenesis.
Development. 151:dev2020432024. View Article : Google Scholar
|
|
238
|
Maniou E, Farah F, Marshall AR,
Crane-Smith Z, Krstevski A, Stathopoulou A, Greene NDE, Copp AJ and
Galea GL: Caudal Fgfr1 disruption produces localised spinal
mis-patterning and a terminal myelocystocele-like phenotype in
mice. Development. 150:dev2021392023. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Siefker-Radtke AO, Necchi A, Park SH,
García-Donas J, Huddart RA, Burgess EF, Fleming MT, Rezazadeh
Kalebasty A, Mellado B, Varlamov S, et al: Efficacy and safety of
erdafitinib in patients with locally advanced or metastatic
urothelial carcinoma: Long-term follow-up of a phase 2 study.
Lancet Oncol. 23:248–258. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
240
|
CADTH Reimbursement Reviews and
Recommendations: Pemigatinib (Pemazyre): Indication: For the
treatment of adults with previously treated, unresectable locally
advanced or metastatic Cholangiocarcinoma with a FGFR2 fusion or
other rearrangement: Reimbursement Recommendation. Canadian Agency
for Drugs and Technologies in Health; Ottawa, ON: 2025
|
|
241
|
Li P, Wu L and Song Z: Data mining and
safety analysis of FGFR tyrosine kinase inhibitors based on the
FAERS database. Sci Rep. 15:261502025. View Article : Google Scholar : PubMed/NCBI
|
|
242
|
Sasanuma H and Yamaguchi H: Sequential
fibroblast growth factor receptor inhibition in intrahepatic
cholangiocarcinoma: Navigating an evolving landscape of resistance
and opportunity-A case report and current opinion. Oncol Ther.
13:1243–1253. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Adashek JJ, Kato S, Sicklick JK, Lippman
SM and Kurzrock R: If it's a target, it's a pan-cancer target:
Tissue is not the issue. Cancer Treat Rev. 125:1027212024.
View Article : Google Scholar : PubMed/NCBI
|
|
244
|
Thein KZ, Myat YM, Park BS, Panigrahi K
and Kummar S: Target-driven tissue-agnostic drug approvals-A new
path of drug development. Cancers (Basel). 16:25292024. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Newbold K and Cheng L: Targeting RET in
medullary thyroid cancer. Endocr Relat Cancer. 32:e2402912025.
View Article : Google Scholar : PubMed/NCBI
|
|
246
|
Meertens M, de Jong LD, de Vries N, Rosing
H, Beijnen JH and Huitema ADR: LC-MS/MS method development and
validation for novel targeted anticancer therapies adagrasib,
capmatinib, ensartinib, entrectinib, larotrectinib, lorlatinib,
pralsetinib, selpercatinib and sotorasib. J Pharm Biomed Anal.
266:1170782025. View Article : Google Scholar : PubMed/NCBI
|
|
247
|
Seguchi K, Enokida T, Oba T, Yoshikawa AL,
Nakatake N and Yu O: A case of micro-medullary thyroid carcinoma
presenting as cancer of unknown primary. Cureus.
17:e870852025.PubMed/NCBI
|