Introduction
Psoriasis is a chronic autoimmune skin disorder
characterized by marked keratinocyte proliferation and inflammatory
infiltration (1,2). While IL-23/IL-17-targeted biologics
(e.g., guselkumab, secukinumab) have proven beneficial in psoriasis
treatment, a notable proportion of patients exhibit partial or no
response (3). Furthermore,
long-term management of psoriasis is frequently complicated by drug
resistance and infection risks (4,5).
Thus, a deeper understanding of the molecular mechanisms governing
immune activation in the psoriatic microenvironment is
imperative.
Beyond the established immunological pathways, such
as the IL-23/Th17 axis and TNF-α signaling, increasing evidence has
indicated the crucial role of metabolic reprogramming in psoriasis
pathogenesis (6-8). Recent studies have highlighted
systemic metabolic comorbidities in patients with psoriasis,
including obesity, diabetes and cardiovascular risk (9,10), suggesting systemic 'metabolic
syndrome' of the skin. While lipid and glucose metabolism have been
extensively reviewed in this context, the role of amino acid
metabolism remains comparatively underexplored (11-13).
Perturbations of amino acid metabolism are
widespread in psoriasis, which are also positively associated with
the severity of psoriatic lesions and psoriasis relapse (14). Furthermore, treatment with
biological agents targeting TNF-α, IL-12/23 and IL-17A has been
shown to normalize these metabolic disturbances, highlighting the
potential importance of amino acids in the pathogenesis of
psoriasis (15,16). Notably, the levels of the key
amino acid phenylalanine are elevated both in psoriatic skin
lesions and the plasma of affected individuals (15,17). Although its dysregulation is
implicated in a number of metabolic disorders, including type 2
diabetes (18) and
age-associated cardiac dysfunction (19), its role in psoriasis remains
largely unexplored.
Within the complex immune landscape of psoriasis,
dendritic cells (DCs) occupy a central role, orchestrating the
transition from innate to adaptive immunity (20). Recent evidence has highlighted
that intracellular lipid accumulation resulting from impaired
autophagy, as well as altered amino acid availability that
hyperactivates nutrient-sensing pathways including mTOR, markedly
disrupts intracellular homeostasis (21,22). This overarching metabolic
rewiring forces DCs into an overmatured state and lowers their
immune activation threshold. Consequently, when these metabolically
primed DCs recognize pathogen- or damage-associated molecular
patterns through surface toll-like receptors, they exhibit
hyper-responsive behaviors. This hypersensitization leads to an
aberrantly amplified activation of the downstream NF-κB signaling
cascade, which serves as the core mechanism driving IL-23 secretion
(23,24). Consequently, DCs not only
initiate and amplify the inflammatory response through the
secretion of pro-inflammatory cytokines, including IL-6, IL-23 and
IL-1β, but also influence T-cell differentiation, particularly
toward the T helper 17 (Th17) phenotype driven by IL-23 (25). Therefore, identifying upstream
mechanisms that govern DC activation and curb their cytokine output
represents an important frontier for developing therapeutic
strategies aimed at achieving long-term remission.
Therefore, the present study was designed to bridge
the gap between systemic metabolic perturbations and local immune
dysregulation in psoriasis. Using metabolomic profiling, the study
sought to investigate phenylalanine accumulation in both the
circulation and cutaneous lesions of psoriatic mice, and to
elucidate the underlying metabolic perturbation. Furthermore, by
manipulating phenylalanine availability in vivo and in
vitro, the study aimed to delineate the mechanistic association
between phenylalanine and NF-κB signaling in DCs, and to explore
its role in driving the pathogenic IL-23/IL-17 axis, thereby
providing a mechanistic rationale for dietary intervention as an
adjunctive therapy for psoriasis.
Materials and methods
Animal studies
A total of 47 male C57BL/6 mice (8 weeks old;
average weight, 22 g) were procured from the Shanghai SLAC
Laboratory Animal Center (Shanghai, China) and housed at the Sir
Run Run Shaw Hospital, Zhejiang University School of Medicine
(Hangzhou, China). The mice were maintained in a specific
pathogen-free facility under controlled environmental conditions
(temperature, 22±2°C; relative humidity, 50±10%; and a 12-h
light/dark cycle) with ad libitum access to sterile water
and their respective diets (standard chow or customized diets as
detailed below). The animal study protocols were approved by the
Animal Research Ethical Committee of the Sir Run Run Shaw Hospital,
Zhejiang University School of Medicine (approval no.
SRRSH2025-0047). To address distinct experimental objectives, the
mice were randomly assigned to specific cohorts as follows: i) For
metabolomics profiling, mice were fed with standard chow and
randomly divided into a normal control (NC) group and an imiquimod
(IMQ)-induced model group (n=6 per group); ii) to evaluate the
efficacy of phenylalanine (Phe) diets, mice were randomly assigned
to three groups (n=5 per group) and fed diets containing either
0.25%, 1% (serving as the baseline control) or 2% Phe, which were
manufactured by Hangzhou Hangsi Biotechnology Co., Ltd. (Table SI); and iii) For the in
vivo L-type amino acid transporter 1 (LAT1) inhibition
experiment, mice were allocated into four groups (n=5 per group).
For the relevant experimental groups, the LAT1 inhibitor JPH203
(cat. no. s8667; Selleck Chemicals) was administered through
subcutaneous injection at a dose of 50 mg/kg.
To establish mouse models of psoriasis, IMQ cream
(Sichuan Med-Shine Pharmaceutical Co., Ltd.) was applied topically
for 5 consecutive days. For the back skin model, 62.5 mg IMQ cream
was applied daily to the shaved dorsal skin. For the ear
inflammation model, to ensure consistent and complete absorption of
the full dose and minimize the risk of medication loss due to
animal grooming, 25 mg IMQ cream was applied evenly across both the
dorsal and ventral surfaces of the right ear. All mice were weighed
and recorded before the induction of psoriasis. Health status and
behavioral patterns of all animals were monitored twice daily
(morning and evening) during the 6-day study period to assess
disease progression and treatment efficacy. An objective scoring
system was used to score the severity of skin inflammation based on
the clinical Psoriasis Area and Severity Index (PASI) (26). Next, erythema, thickening, and
scaling were rated on a 0-4 scale, where 0, 1, 2, 3 and 4 indicated
no symptoms, mild, moderate, severe or extremely severe symptoms,
respectively. The cumulative scores served as the severity of
inflammation index (scale 0-12). The researchers were blinded
during the experiment and data analysis.
To monitor the dynamic progression of psoriasiform
inflammation, the macroscopic clinical severity of the mouse skin
was evaluated and recorded daily. The final PASI scores analyzed
and presented in the present figures were recorded on day 6 of the
experiment, prior to euthanasia. On day 7, all terminal procedures
were performed in a strict sequence. Mice were euthanized through
CO2 asphyxiation with a controlled displacement rate of
30% of the chamber volume per min and mortality was strictly
verified by determining the absence of heartbeat and respiration.
Immediately following the confirmation of mortality, blood was
collected through retro-orbital bleeding and lesional skin tissues
and plasma were subsequently harvested for further histological and
metabolic analyses. The excised spleens were immediately weighed
and the spleen index was calculated as the ratio of spleen weight
to body weight. The acanthosis thickness was analyzed using ImageJ
software (version 1.53; National Institutes of Health) and the
thickness of the ear skin was measured with a vernier caliper.
Metabolomics profile analysis
Mouse tissues and plasma were collected and analyzed
by Wuhan Metware Biotechnology Co., Ltd. Specifically, 100 mg
tissue or 100 µl plasma was homogenized in 500 µl 70%
aqueous methanol solution. The mixture underwent a 3 min
vortex-mixing step at 2,500 rpm, followed by centrifugation (13,800
× g, 10 min, 4°C). A total of 300 µl supernatant was
isolated into fresh vials for liquid chromatography (LC)-mass
spectrometry (MS) analysis. To ensure analytical stability, quality
control samples were generated by combining equal aliquots from
every individual specimen and handled identically.
Sample extracts were analyzed using an
LC-electrospray ionization (ESI)-MS/MS system (ExionLC™ AD UPLC
coupled with a QTRAP® 6500+ System; SCIEX). The
instrument operated under both positive (5,500 V) and negative
(-4,500 V) ion spray modes utilizing an ESI Turbo Ion-Spray (SCIEX)
interface (source temperature: 550°C and curtain gas: 35.0 psi),
managed using Analyst® (version 1.6; SCIEX) software.
Multiple reaction monitoring transitions were continuously
optimized for each elution window of target amino acids.
Data analysis
Raw mass spectrometry outputs were transformed into
MzXML files utilizing ProteoWizard MSConvert (version 3.0;
http://proteowizard.sourceforge.net)
prior to XCMS integration (using the 'xcms' package on the R
platform; version 4.1.2; https://bioconductor.org/packages/xcms). Strict
thresholds [variable importance in projection (VIP) >1 and
P<0.05] were applied within the Partial Least Squares
Discriminant Analysis model to filter differential metabolites.
Pathway impact assessments were conducted using MetaboAnalyst
(version 5.0; https://www.metaboanalyst.ca), where pathways
exhibiting an impact factor >0.1 were deemed highly
significant.
Immunohistochemical and histological
analysis
Mouse skin samples were immersed in 4%
paraformaldehyde for optimal fixation at room temperature for 24 h.
Tissues were subsequently dehydrated, cast into paraffin blocks and
microtomed into 5-µm cross-sections. H&E staining was
carried out following the manufacturer's instructions (Beyotime
Biotechnology). For immunofluorescence assays, the sections were
first deparaffinized in xylene and rehydrated using a descending
ethanol gradients. Heat-mediated antigen unmasking was performed at
95°C for 15 min in a pH 6.0 citrate buffer. To prevent non-specific
binding, slides were treated with a 5% BSA solution (Beyotime
Biotechnology) enriched with 0.1% Triton X-100 for 1 h at room
temperature. Primary antibody incubation with anti-p-p65 (1:200
dilution; cat. no. 3033S; Cell Signaling Technology, Inc.) occurred
overnight at 4°C. After rigorous washing with PBS. specimens were
exposed to CoraLite® Plus 488-conjugated AffiniPure goat
anti-rabbit IgG (H+L) (1:200 dilution; cat. no. RGAR002;
Proteintech Group, Inc.) for detection of p-p65, for 1 h at room
temperature in the dark. Subsequently, specimens were incubated
with anti-CD11c (Alexa Fluor® 555 conjugate) (1:200
dilution; cat. no. 64675S; Cell Signaling Technology, Inc.)
overnight at 4°C. CD11c was visualized directly via its Alexa
Fluor® 555 conjugate without requiring a secondary
antibody. All images were eventually captured utilizing a digital
slide scanning system (Konfoong Bioinformation Tech Co., Ltd.).
Skin cell isolation and flow cytometric
analysis
Single cells from the ear were generated according
to the methods of a previous study (27). To prevent non-specific Fc
receptor binding, single-cell suspensions were pre-treated with the
anti-mouse CD16/32 antibody (1:500 dilution; cat. no. 553141; BD
Biosciences) on ice for 20 min prior to the surface marker
labeling. The antibodies (all used at 1:500 dilution) for flow
cytometry included allophycocyanin (APC)-A700-CD45 (56-0454081;
eBioscience; Thermo Fisher Scientific, Inc.), FITC-CD3 (cat. no.
11-0032-82; eBioscience; Thermo Fisher Scientific, Inc.),
BV650-CD11b (cat. no. 416-0112-80; eBioscience; Thermo Fisher
Scientific, Inc.) and BV421-LY6G (cat. no. 404-9668-80;
eBioscience; Thermo Fisher Scientific, Inc.). The Zombie Aqua™
Fixable Viability Kit (cat. no. 423101; BioLegend, Inc.) was also
used. After washing, the cells were assayed with a CytoFLEX LX flow
cytometer (Beckman Coulter, Inc.) and the data were analyzed using
FlowJo software (version 10; BD Biosciences).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from harvested skins or DCs
using RNAiso Plus (Takara Bio, Inc.). Reverse transcription for
cDNA synthesis was conducted utilizing the PrimeScript™ RT Reagent
Kit (cat. no. RR037A; Takara Bio, Inc.) following the provided
technical guidelines. RT-qPCR was conducted using the Hieff™ qPCR
SYBR Green Master Mix (cat. no. 11201ES08; Shanghai Yeasen
Biotechnology Co., Ltd.) on a LightCycler® 480 system
(Roche Diagnostics GmbH). The thermocycling program commenced with
a 5 min pre-denaturation at 95°C, sequentially followed by 40
cycles (95°C for 10 sec and 60°C for 30 sec). Target gene
expression levels were normalized against the housekeeping gene
β-actin leveraging the 2−ΔΔCq method (28). Primer sequences are listed in
Table SII.
Preparation of bone marrow-derived DCs
(BMDCs) and treatments
To isolate primary cells for both BMDC and naïve
CD4+ T cell experiments, healthy and untreated male
C57BL/6 donor mice (aged 6-8 weeks) obtained from the Shanghai SLAC
Laboratory Animal Center, (Shanghai, China) were utilized. A total
of 10 donor mice were euthanized using CO2 asphyxiation
(at a displacement rate of 30% of the chamber volume per min) and
mortality was verified by cervical dislocation. To obtain
sufficient cell yields, biological replicates for the following
in vitro assays were generated using pooled samples from
these donor mice.
To prepare BMDCs, femurs and tibias were flushed
with RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) to
collect bone marrow. The cell pellet was subsequently resuspended
in red cell lysis buffer (Beyotime Biotechnology) for 3 min. The
primary culture system consisted of RPMI-1640 fortified with 10%
FBS (CellMax Technologies AB), 1% penicillin-streptomycin, 50
µM β-mercaptoethanol (cat. no. abs9592; Absin Bioscience,
Inc.), 20 ng/ml GM-CSF (cat. no. 315-03; PeproTech®;
Thermo Fisher Scientific, Inc.) and 10 ng/ml recombinant mouse IL-4
(cat. no. 214-14; PeproTech®; Thermo Fisher Scientific,
Inc.). On days 3 and 5, half of the old culture medium was
carefully removed and replaced with an equal volume of fresh
medium. All cells were cultured at 37°C in a 5% CO2
incubator. On day 6, BMDCs were used for relevant experiments. All
in vitro pharmacological treatments were conducted at 37°C
in a 5% CO2 incubator. For in vitro stimulation,
IMQ powder (cat. no. R837; InvivoGen) was dissolved in sterile
water and added to the culture medium at a final concentration of
10 µg/ml. For inhibitor treatments, cells were incubated
with the NF-κB inhibitor BAY11-7082 (10 µM; MedChemExpress)
or the LAT1 inhibitor JPH203 (100 µM; Selleck Chemicals) for
2 h.
To detect maturation of BMDCs, expression of CD80,
CD86 and major histocompatibility Complex (MHC)-II was analyzed on
CD11c-positive cells using APC-labelled anti-CD80 (cat. no. 104713;
BioLegend, Inc.), primary binding-labelled anti-CD86 (cat. no.
105021; BioLegend, Inc.), phycoerythrin (PE)-labelled anti-MHCII
(cat. no. 12-5321-81, eBioscience; Thermo Fisher Scientific, Inc.)
and PE-Cy7-labelled anti-CD11c (cat. no. 117317, BioLegend, Inc.)
antibodies (all used at a 1:500 dilution). The incubation was
performed on ice for 20 min in the dark. After antibody staining,
cells were washed and analyzed with a CytoFLEX LX flow cytometer
(Beckman Coulter, Inc.).
BMDC supernatant and T cell
co-culture
On day 6, BMDCs were pre-treated with phenylalanine
(1 mM) for 48 h at 37°C in a humidified incubator containing 5%
CO2, followed by incubation with IMQ 10 µg/ml for
24 h to harvest the supernatant. Naïve CD4+ T cells were
isolated from the spleens of C57BL/6 mice using an isolation kit
(cat. no. 19765A, Stemcell Technologies, Inc.). A total of 200
µl BMDC supernatant was used to resuspend naïve
CD4+ T cells for co-culture for 72 h. Differentiated T
cells were stimulated with 750 ng/ml ionomycin (cat. no. 5608212;
PeproTech®; Thermo Fisher Scientific, Inc.), 50 ng/ml
phorbol 12-myristate 13-acetate (cat. no. S1819; Beyotime
Biotechnology) and brefeldin A (cat. no. 00-4506-51; eBioscience;
Thermo Fisher Scientific, Inc.) for another 4 h at 37°C. The cells
were then collected, stained with PE-labelled anti-CD4 (cat.
17-0041-82; eBioscience; Thermo Fisher Scientific, Inc.) and
APC-labelled anti-IL-17A (cat. 17-7177-81; eBioscience, Thermo
Fisher Scientific, Inc.) antibodies (both at a 1:500 dilution) on
ice for 20 min in the dark. The stained cells were then analyzed on
a CytoFLEX LX flow cytometer (Beckman Coulter, Inc.).
RNA sequencing (RNA-seq) data
processing
Total RNA was isolated from harvested tissues or DCs
leveraging the RNAiso Plus reagent (Takara Bio, Inc.) as directed
by the manufacturer. RNA purity and integrity were verified using a
NanoDrop 2000 spectrophotometer and an Agilent 2100 Bioanalyzer
(RIN >7.0). Transcriptomic libraries were constructed, verified
using an Agilent 2100 Bioanalyzer, and subsequently subjected to
2×150 bp paired-end sequencing. The sequencing runs were executed
on the Illumina NovaSeq™ X Plus system (Illumina, Inc.) at a final
loading concentration of 1.5 nM by LC-Bio Technologies (Hangzhou)
Co., Ltd. The 'DESeq2' package (29) was utilized within the R platform
(version 4.1.2) for transcriptomic comparisons between specific
groups, whereas the 'edgeR' algorithm (30) was employed for evaluating
individual sample variations. Differentially expressed genes (DEGs)
were defined by a false discovery rate (FDR) <0.05 and an
absolute fold change ≥2. DEGs were then subjected to enrichment
analysis of Gene Ontology (GO) functions and Kyoto Encyclopedia of
Genes and Genomes (KEGG) pathways.
Western blotting
BMDCs were lysed in standard RIPA buffer (Beyotime
Biotechnology) to liberate total cellular proteins. Lysates were
subjected to low-temperature centrifugation (10,000 × g, 4°C, 20
min) to clear cellular debris. Protein quantification was executed
using a BCA reagent kit (cat. no. FD2001; Fdbio Science).
Equivalent protein masses (30 µg per lane) were resolved
using 12% SDS-PAGE and electrically blotted onto 0.22 µm
PVDF transfer membranes. The membranes were blocked with 5% non-fat
dairy milk for 1 h at room temperature prior to overnight
incubation at 4°C with distinct primary antibodies: anti-NF-κB p65
(1:1,000 dilution; cat. no. 8242; Cell Signaling Technology, Inc.),
phosphorylated (p)-NF-κB p65 (Ser536; 1:1,000 dilution; cat. no.
3033; Cell Signaling Technology, Inc.), anti-IκBα (1:1,000
dilution; cat. no. 4814; Cell Signaling Technology, Inc.) and
anti-p-IκBα (Ser32; 1:1,000 dilution; cat. no. 2859; Cell Signaling
Technology, Inc.). An anti-β-tubulin antibody (1:1,000 dilution;
cat. no. AF2839; Beyotime Biotechnology) was used as the loading
control. The subsequent day, blots were probed with an
HRP-conjugated goat anti-rabbit secondary antibody (1:5,000
dilution; cat. no. A0208; Beyotime Biotechnology) for 1 h at room
temperature. For the detection of phosphorylated and total proteins
(such as p-p65/p65 and p-IκBα/IκBα) on the same membrane, the PVDF
membranes were first probed for the phosphorylated proteins, then
stripped using a commercial stripping buffer (cat. no. FD0050;
Fdbio Science), blocked again and reprobed for the respective total
proteins. Immunoreactive bands were optically captured using a
VILBER Fusion FX7 imaging system (Vilber Lourmat) supported by a
specialized chemiluminescent substrate (cat. no. FD8030; Fdbio
Science).
Statistical analysis
All quantitative datasets were processed and
visualized utilizing GraphPad Prism (version 9.0; Dotmatics). For
statistical comparisons involving two independent cohorts, a
two-tailed Student's t-test was executed. When evaluating datasets
containing >2 experimental groups, either one-way or two-way
ANOVAs were applied, followed by Tukey's post-hoc test for multiple
pairwise comparisons. P<0.05 was considered to indicate a
statistically significant difference. Data are presented as the
mean ± SEM.
Results
Psoriasis progression is associated with
abnormal phenylalanine metabolism
To investigate the metabolic landscape associated
with psoriasis progression, untargeted metabolomics profiling was
performed on skin samples from IMQ-induced psoriasis-like mouse
models and NCs (Fig. 1A). PCA
revealed distinct metabolic clustering, indicating a shift in the
metabolic profile following IMQ treatment (Fig. 1B). Specifically, out of 14,887
detected features, 3,506 significantly upregulated and 1,160
significantly downregulated metabolites were identified in the
psoriatic skin (Fig. 1C). To
interpret the biological importance of these shifts, KEGG pathway
enrichment analysis was performed. The results demonstrated that
the altered metabolites were predominantly enriched in amino acid
metabolism pathways, including 'arginine biosynthesis', 'histidine
metabolism' and notably, 'phenylalanine metabolism' (Fig. 1D). Given the high enrichment
ratio and statistical significance associated with phenylalanine
metabolism, this pathway was selected for further validation.
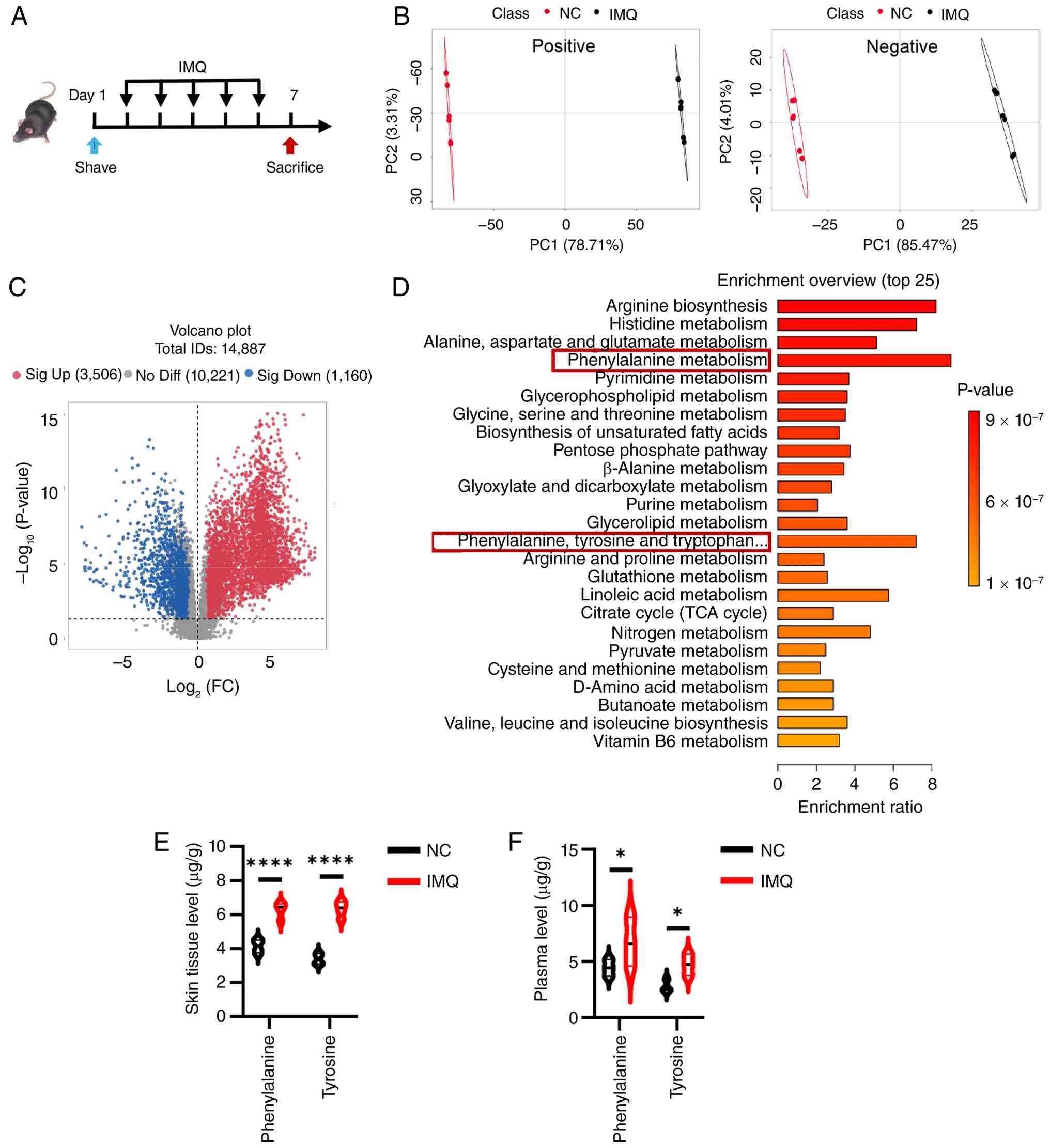 | Figure 1Psoriasis progression is associated
with aberrant phenylalanine metabolism in IMQ-induced mice. (A)
Schematic of the experimental design for the IMQ-induced psoriasis
model (n =6 mice/group). (B) PC analysis score plots exhibiting
distinct metabolic clustering and separation between the NC group
and the IMQ model group in both positive and negative ion modes.
(C) Volcano plots showing the differential abundance of total
identified metabolites (n=14,887). Red dots represent significantly
upregulated metabolites (n=3,506), blue dots represent
significantly downregulated metabolites (n=1,160), variable
importance in projection >1, P<0.05 and absolute FC >1.2.
(D) Overview of the top 25 terms in the metabolic enrichment
analysis of the differential metabolites. Absolute quantification
of phenylalanine and tyrosine levels in (E) skin tissue
(µg/g) and (F) plasma (µg/ml) using targeted
metabolomics. Data are presented as the mean ± SEM. Statistical
significance between the two groups was determined using an
unpaired two-tailed Student's t-test. *P<0.05 and
****P<0.0001. IMQ, imiquimod; NC, normal control; FC,
fold change; ns, not significant; PC, principal component. |
These untargeted findings were next corroborated
using targeted quantitative metabolomics to measure absolute
concentrations of phenylalanine and its downstream metabolite,
tyrosine. Consistent with the present initial metabolomics
screening, phenylalanine and tyrosine levels were found to be
significantly elevated in the skin lesions of IMQ-treated mice
compared with NC group (Fig.
1E). Furthermore, plasma analysis also revealed a significant
systemic increase in circulating phenylalanine and tyrosine levels
(Fig. 1F). These findings
collectively suggested that the progression of psoriasis was
closely associated with aberrant phenylalanine metabolism (Fig. S1), leading to the accumulation
of both phenylalanine and tyrosine systemically and locally in skin
lesions.
Phenylalanine exacerbates IMQ-induced
mouse psoriasis-like skin lesions
Having identified phenylalanine accumulation as a
metabolic hallmark of psoriasis, the concentration of phenylalanine
was adjusted in the diet of mice during psoriasis modeling to
determine whether altering systemic phenylalanine levels
functionally modulates disease outcome (19). C57BL/6 mice were acclimated to
diets containing varying concentrations of phenylalanine (0.25, 1
or 2%) for 1 week prior to and during the course of IMQ treatment
(Fig. 2A). Notably, these
dietary modifications did not significantly affect general health
or body weight before IMQ treatment (Fig. 2B). However, after IMQ
application, phenotypic assessment revealed a significant
dose-dependent effect of phenylalanine on skin inflammation. Mice
fed a high-phenylalanine diet (2%) exhibited exacerbated
psoriasis-like lesions characterized by severe erythema, heavy
scaling and skin thickening. Conversely, dietary restriction of
phenylalanine (0.25%) significantly alleviated these clinical
manifestations compared with the standard diet (1%) group (Fig. 2C). This observation was
quantified by PASI scores, which exhibited a clear dose-dependent
trajectory (Figs. 2D and
S2). Histological analysis of
the dorsal skin further demonstrated these findings: High dietary
phenylalanine induced pronounced epidermal hyperplasia (acanthosis)
and inflammatory cell infiltration, whereas phenylalanine
restriction effectively attenuated these pathological changes
(Fig. 2E and F). To characterize
the inflammatory milieu driven by phenylalanine, the gene
expression of key psoriasis-associated cytokines in the skin were
analyzed. It was found that elevated dietary phenylalanine
significantly promoted the gene expression levels of Il1b,
Il6 and C-X-C motif chemokine ligand (Cxcl-)-1, as
well as the key psoriasis drivers Il12b, Il23a and
Il17a (Fig. 2G).
Consistent results were observed in the ear skin, whereby high
phenylalanine aggravated ear swelling and histological severity,
accompanied by a significant increase in ear thickness (Fig. 2H-J). To further establish the
pathological severity and terminal phenotype of the disease model,
the infiltration of neutrophils was evaluated, which serve as a
well-established marker of psoriasis severity (31). Flow cytometry analysis of
single-cell suspensions from the ear skin revealed that dietary
phenylalanine significantly dose-dependently increased the
infiltration of neutrophils (Fig. 2K
and L). In summary, these results indicated that phenylalanine
acted as an aggravating factor in psoriasis, likely by amplifying
pro-inflammatory cytokine production and promoting immune cell
recruitment to skin lesions.
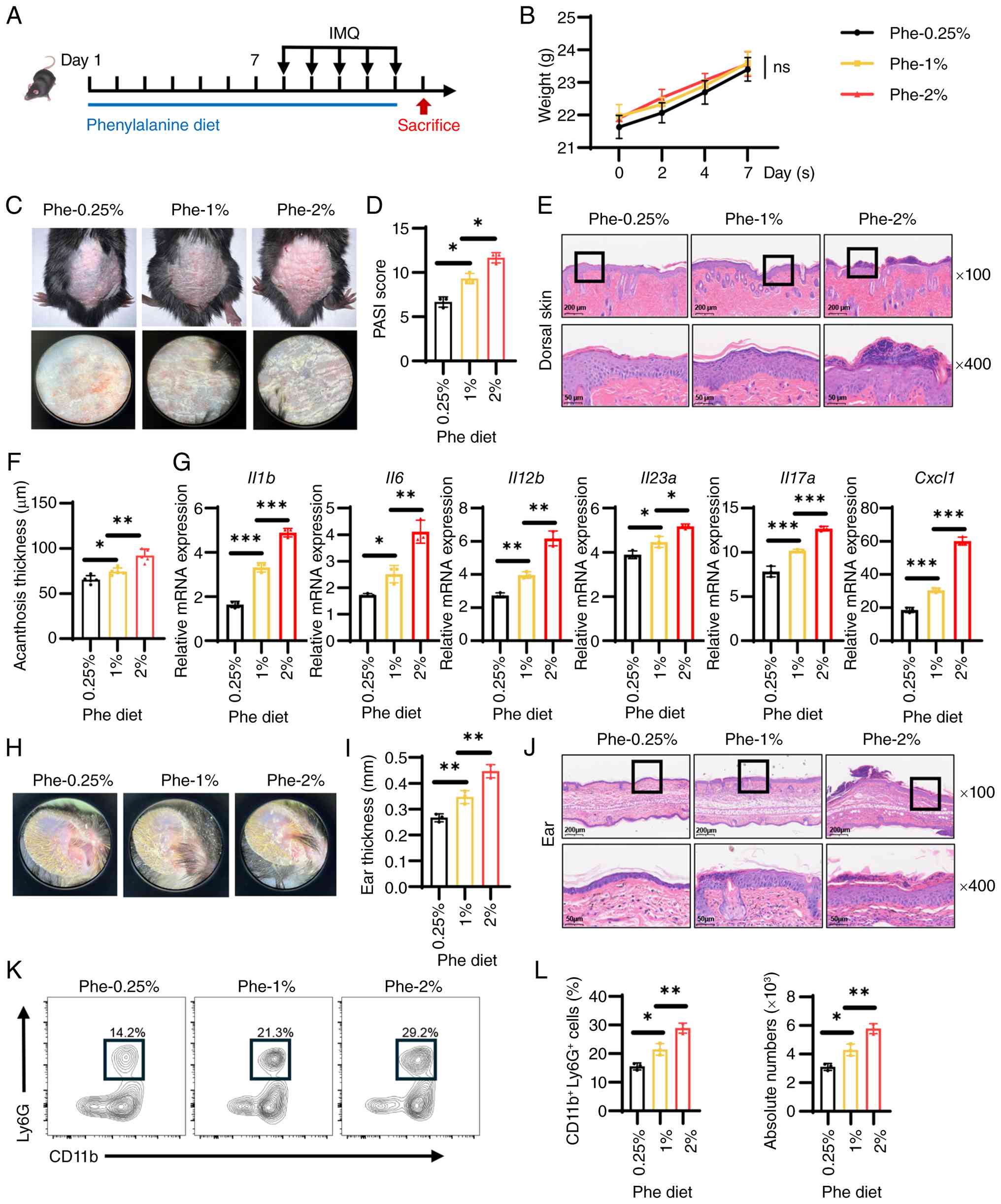 | Figure 2Dietary phenylalanine exacerbates
IMQ-induced psoriasis-like dermatitis in a dose-dependent manner.
(A) Schematic representation of the dietary intervention: Mice were
fed diets containing 0.25% (low), 1% (normal) or 2% (high) Phe for
7 days prior to and during IMQ application (n=5/group). (B)
Recorded body weights of mice fed with Phe diet prior to IMQ
application. (C) Representative phenotypic images of dorsal skin
lesions across dietary groups after IMQ treatment. (D) PASI scores
of the mice in each group evaluated on day 6. (E) Representative
H&E-stained sections of the dorsal skin (scale bars, 50 and 200
µm; the black rectangles indicate the regions of interest
magnified in the 400× views below). (F) Histological quantification
of epidermal thickness (acanthosis) in dorsal skin lesions. (G)
Reverse transcription-quantitative PCR analysis of pro-inflammatory
cytokines (Il1b, Il6, Il12b, Il23a, and
Il17a) and chemokine Cxcl1 gene expression in skin
tissues. (H) Representative images of ears skin (ventral side). (I)
Ear thickness measurement. (J) Representative H&E-stained
sections of the ear skin (scale bars, 50 and 200 µm). Flow
cytometry analysis showing (K) representative relative percentage
and (L) absolute quantification of infiltrating neutrophils
(CD11b+ Ly6G+) in ear tissues. Data are
presented as the mean ± SEM. Statistical significance was
determined using two-way ANOVA (for B and D) or one-way ANOVA (for
all others), followed by Tukey's multiple comparisons test.
*P<0.05, **P<0.01 and
***P<0.001. IMQ, imiquimod; ns, not significant;
Cxcl1, C-X-C motif chemokine ligand 1; Phe, phenylalanine. |
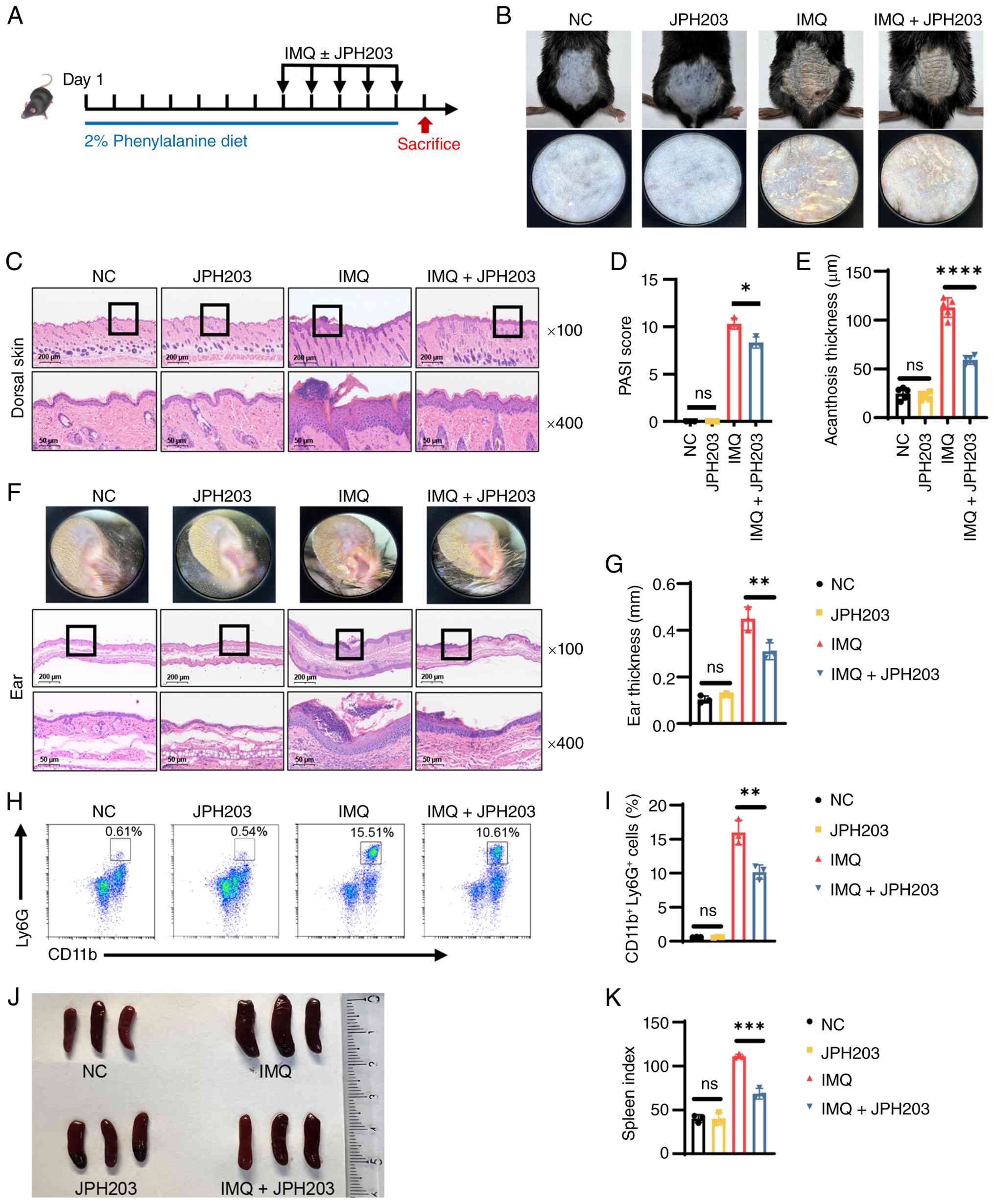
LAT1 inhibitor JPH203 mitigates
phenylalanine-driven psoriatic inflammation
To validate the therapeutic potential of restricting
cellular phenylalanine availability, LAT1, the primary transporter
for large neutral amino acids, was targeted. JPH203, a selective
high-affinity LAT1 inhibitor, was utilized in mice subjected to the
high-phenylalanine diet and IMQ challenge (Fig. 3A). Notably, administration of
JPH203 alone to healthy mice did not induce any cutaneous
abnormalities or toxicity. Aligning with the present hypothesis,
JPH203 exerted a potent protective effect. Treatment with this
inhibitor markedly abrogated the exacerbating impact of dietary
phenylalanine. Clinically, this was evidenced by a marked reduction
in erythema and scaling compared with the vehicle-treated IMQ group
(Fig. 3B). Consistent with these
macroscopic observations, histological assessment of the dorsal
skin revealed that JPH203 effectively attenuated pathological
epidermal thickening (Fig. 3C).
Quantitative analyses further demonstrated these protective
effects, demonstrating significant reductions in PASI scores
(Figs. 3D and S3) and epidermal acanthosis thickness
(Fig. 3E). Similarly, in the ear
skin model, JPH203 treatment alleviated tissue swelling and
restored epidermal architecture, characterized by reduced
hyperkeratosis and fewer pustules (Fig. 3F), as well as a significant
reduction in ear thickness (Fig.
3G). At the cellular level, flow cytometry analysis revealed
that JPH203 treatment effectively suppressed the immune response,
leading to a significant reduction in the frequency of infiltrating
neutrophils in skin lesions (Fig. 3H
and I). Furthermore, systemic inflammation was assessed by
examining the spleen. While IMQ treatment induced significant
splenomegaly, JPH203 co-administration normalized both spleen size
and the spleen index (Fig. 3J and
K). Collectively, these findings demonstrated that blocking
phenylalanine transport through JPH203 effectively disrupted the
phenylalanine-dependent inflammatory loop, providing therapeutic
benefits both locally and systemically.
Phenylalanine promotes expression of
psoriasis-associated cytokines and chemokines in DCs
DCs serve as key orchestrators in the initiation of
psoriasis, primarily by driving the IL-23/IL-17 axis (32). The present in vivo
findings demonstrated that systemic phenylalanine markedly
aggravated IMQ-induced psoriasis-like dermatitis. To elucidate the
molecular basis of this effect, RNA-seq was performed on BMDCs
exposed to phenylalanine in the presence or absence of IMQ
stimulation. Transcriptomics analysis revealed that phenylalanine
exposure alone was sufficient to notably alter the DC landscape
(Fig. 4A). Compared with vehicle
controls, phenylalanine treatment significantly upregulated 116
genes (Fig. 4B). KEGG enrichment
analysis showed phenylalanine significantly regulated genes
associated with the 'cytokine-cytokine receptor interaction',
'NOD-like receptor signaling pathway' and 'NF-κB signaling pathway'
(Fig. 4C). Notably, in the
context of inflammatory challenge (IMQ stimulation), phenylalanine
acted as a potent amplifier. Co-treatment with phenylalanine and
IMQ induced a distinct transcriptional profile compared with IMQ
alone. Differential expression analysis highlighted further
upregulation of 245 genes, including key pro-inflammatory mediators
such as Il1a, Il1b, Il6 and Cxcl1-3.
Functional enrichment analysis determined that phenylalanine
potentiated pathways key in psoriasis pathogenesis, including the
'IL-17 signaling pathway' and 'cytokine-cytokine receptor
interaction' (Fig. 4D).
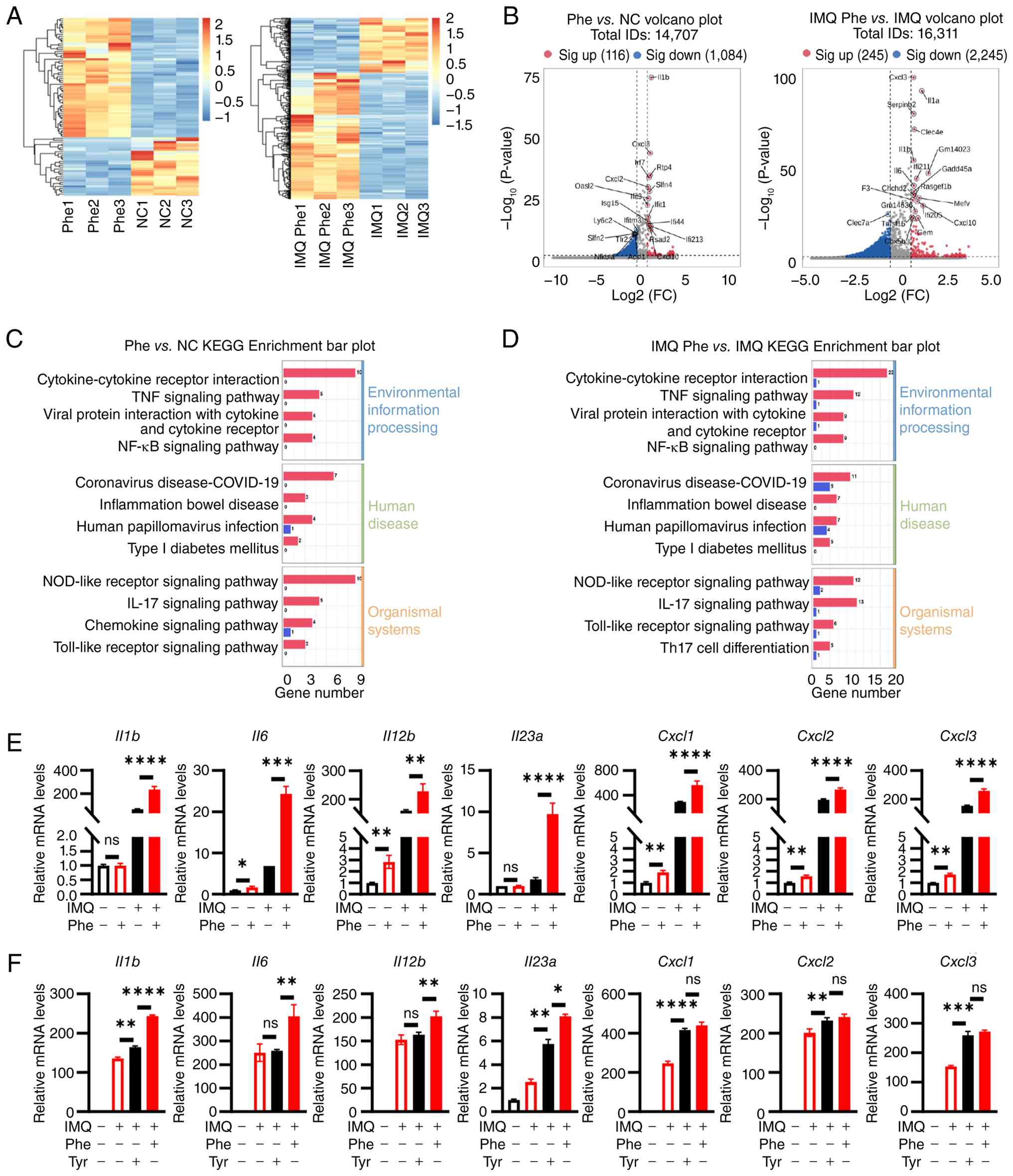 | Figure 4Phe potentiates the inflammatory
response in BMDCs. BMDCs were treated with Phe or the vehicle for
48 h, with or without IMQ stimulation, followed by RNA sequencing
and qPCR analysis. (A) Hierarchical clustering heatmap of DEGs
comparing Phe alone vs. NC and IMQ + Phe vs. IMQ alone. (B) Volcano
plots depicting DEGs in Phe vs. NC and IMQ + Phe vs. IMQ
comparisons. KEGG pathway enrichment analysis showing top enriched
pathways for the upregulated genes in (C) Phe vs. NC and (D) IMQ +
Phe vs. IMQ comparisons. (E) RT-qPCR validation of key
psoriasis-associated cytokines (Il1b, Il6,
Il12b and Il23a) and chemokines (Cxcl1,
Cxcl2 and Cxcl3) across four experimental groups (NC,
Phe, IMQ and IMQ + Phe). (F) RT-qPCR analysis comparing the effects
of Phe and tyrosine supplementation on cytokine expression under
IMQ stimulation. Data are representative of three independent
experiments and presented as the mean ± SEM. Statistical
significance was determined by two-way ANOVA (for E) and one-way
ANOVA (for F), followed by Tukey's multiple comparisons test.
*P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001. IMQ,
imiquimod; ns, not significant; NC, normal control; DEGs,
differentially expressed genes; Cxcl, C-X-C motif chemokine ligand;
Phe, phenylalanine; qPCR, quantitative PCR; RT-qPCR, reverse
transcription-qPCR; KEGG, Kyoto Encyclopedia of Genes and Genomes;
BMDCs, bone marrow-derived dendritic cells; FC, fold change. |
To validate these transcriptomic signatures, qPCR
was performed on independent samples. Consistent with the RNA-seq
data, phenylalanine treatment alone significantly increased the
basal expression of Il6 and Il12b, while only
slightly elevating Il1b and Il23a levels without
statistical significance. Notably, it significantly and
synergistically enhanced the IMQ-induced expression of these
cytokines, as well as the neutrophil-attracting chemokines
Cxcl1, Cxcl2 and Cxcl3 (Fig. 4E). To determine whether this
effect was specific to phenylalanine or shared by its downstream
metabolites, the impact of tyrosine supplementation was evaluated.
While tyrosine also exacerbated the IMQ-induced expression of
Il1b, Il6 and Il12b, its effect was notably
less potent compared with that of phenylalanine in combination with
IMQ (Fig. 4F). This suggested
that while tyrosine contributes to the phenotype, phenylalanine
itself acted as the primary driver of the hyper-inflammatory state
in DCs. Collectively, these data suggest that phenylalanine
reprogramed the DC transcriptome, lowering the threshold for
activation and amplifying the magnitude of the inflammatory
response.
Phenylalanine enhances NF-κB activation
in dendritic cells to promote cytokine production
To mechanistically validate the present
transcriptomic signatures, the NF-κB signaling axis, a central
regulator of inflammatory cytokine production (33), was investigated. The activation
status of DCs was first examined in vivo. Immunofluorescence
staining of skin lesions from mice fed different diets revealed
that while the total density of infiltrating DCs
(CD11c+) remained comparable across dietary groups
(Fig. 5A and B), the activation
state was markedly altered. Specifically, the fluorescence
intensity of p-p65+ within DCs exhibited an upward
trend, though a statistically significant increase was only
observed between the 1 and 2% phenylalanine groups (Fig. 5A and C). The signaling kinetics
were next dissected in vitro using BMDCs. Western blotting
analysis demonstrated that phenylalanine treatment independently
elevated the basal phosphorylation levels of IκBα and p65.
Following the addition of IMQ, the expression of phosphorylated
IκBα and p65 were further upregulated (Fig. 5D). Quantification showed that the
phosphorylation levels of IκBα and p65 were significantly amplified
at numerous time points compared with the controls (Fig. 5E and F). To prove causality,
cells were treated with BAY11-7082, a specific NF-κB inhibitor.
While phenylalanine strongly potentiated the IMQ-induced expression
of Il1b, Il6, Il12b, Il23a and
Cxcl1, pharmacological blockade of NF-κB abrogated this
amplification effect (Fig. 5G),
determining that the pro-inflammatory action of phenylalanine was
dependent on the NF-κB axis. Finally, to determine an association
between intracellular signaling to amino acid transport, the LAT1
inhibitor JPH203 was utilized. Western blotting revealed that
blocking phenylalanine uptake effectively suppressed
hyperactivation of the pathway, evidenced by reduced
phosphorylation of IκBα and p65 in JPH203-treated cells (Fig. 5H-J). Consistent with this, JPH203
treatment significantly attenuated gene expression of downstream
cytokines and chemokines in phenylalanine-exposed DCs (Fig. 5K). In summary, these data
indicated that LAT1-mediated phenylalanine uptake fueled the
hyperactivation of NF-κB signaling, thereby drove the excessive
production of psoriasis-associated cytokines in DCs.
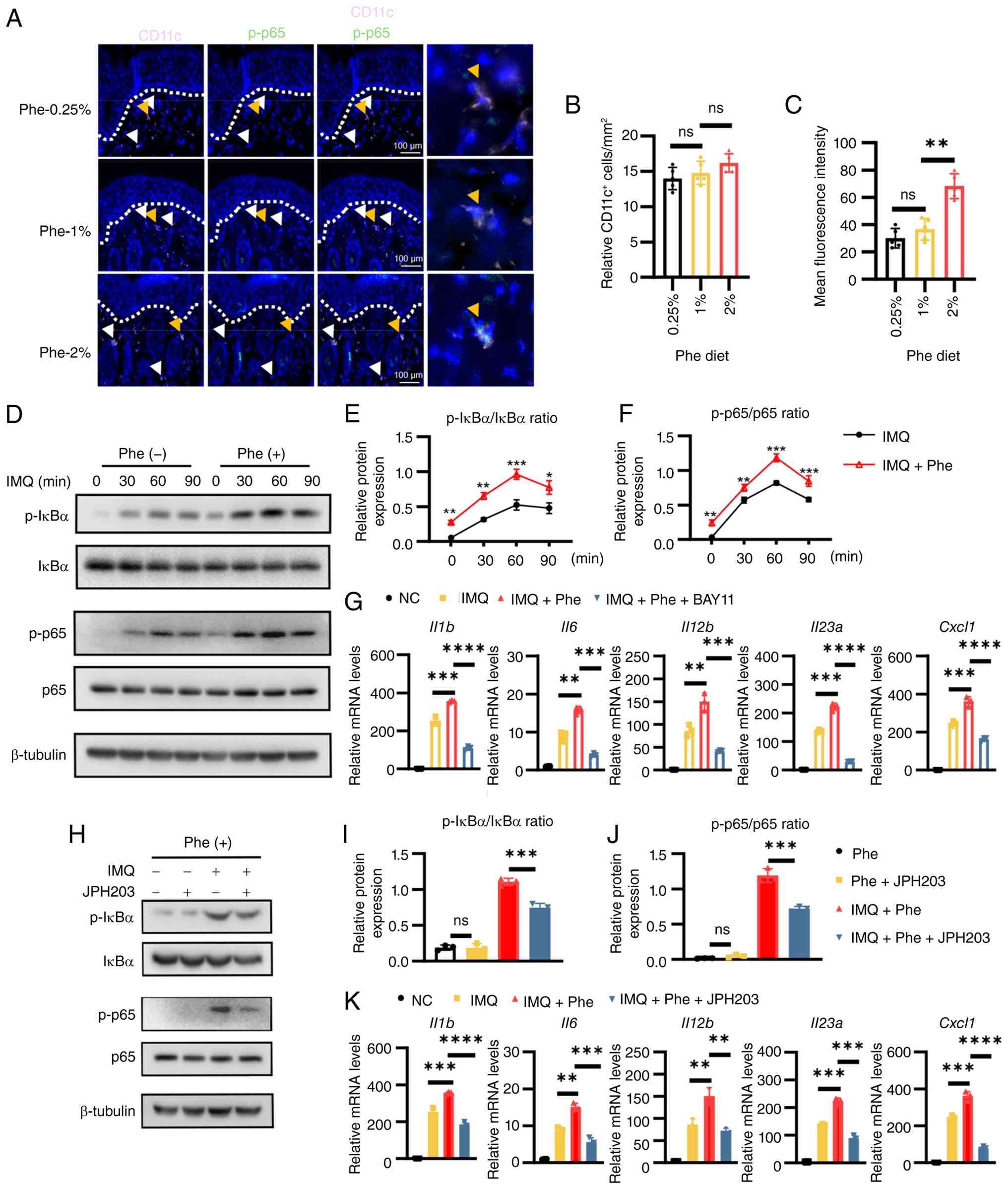 | Figure 5Phe enhances NF-κB signaling
activation in dendritic cells to promote pro-inflammatory cytokine
production. (A) Representative immunofluorescence images of dorsal
skin sections from mice fed different phenylalanine diets (0.25, 1
and 2%). Sections were stained for CD11c (pink), p-p65 (green) and
DAPI (blue; scale bars, 100 µm). (B) Quantification of the
relative density of CD11c+ cells in the dermis. (C)
Quantification of the relative fluorescence intensity of
p-p65+cells within the CD11c+ population. (D)
Western blotting analysis showing protein expression of the NF-κB
pathway in BMDCs. (Expected molecular weights are as follows:
p-IκBα, ~39 kDa; IκBα, ~37 kDa, p-p65, ~65 kDa; p65, ~65 kDa;
β-tubulin, ~55 kDa). (E,F) Densitometry quantification of (E)
p-IκBα/IκBα and (F) p-p65/p65 ratios from the blots in (D). (G)
RT-qPCR analysis of pro-inflammatory genes in BMDCs treated with
IMQ + Phe, in the presence or absence of the NF-κB inhibitor
BAY11-7082. (H) Western blotting analysis of NF-κB activation in
BMDCs treated with IMQ + Phe, in the presence or absence of the
L-type amino acid transporter 1 inhibitor JPH203. (Densitometry
quantification of (I) p-IκBα/IκBα and (J) p-p65/p65 ratios from the
blots in (H). (K) RT-qPCR analysis of pro-inflammatory genes in
BMDCs treated with IMQ + Phe, with or without JPH203. Data are
representative of three independent experiments and presented as
the mean ± SEM. Statistical significance was determined by two-way
ANOVA (for E and F) and one-way ANOVA (for B, C, G, I, J and K),
followed by Tukey's multiple comparisons test.
*P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001. Phe,
phenylalanine; ns, not significant; BMDCs, bone marrow-derived
dendritic cells; p-, phosphorylated; IMQ, imiquimod; RT-qPCR,
reverse transcription-quantitative PCR. |
Phenylalanine exacerbates psoriasis by
promoting Th17 cell differentiation and IL-17 cytokine
production
DCs serve an important role in immature T cell
differentiation and the present data showed that phenylalanine
amplified the production of Th17-polarizing cytokines (IL-6, IL-23
and IL-1β) by DCs. The present study hypothesized that this
metabolic cue would ultimately exacerbate psoriasis by fueling a
Th17-dominant response. To test this in vivo, the systemic
immune profile of mice fed diets with varying phenylalanine
concentrations were analyzed. Flow cytometry analysis of drained
lymph nodes revealed a notable dose-dependent expansion of Th17
cells. Mice on the 2% phenylalanine diet exhibited the highest
frequency and absolute numbers of Th17 cells among all diet groups
(Fig. 6A-C). To determine that
this Th17 expansion was mechanistically driven by DC-derived
signals rather than a direct effect on T cells, an in vitro
differentiation assay was performed using DC-conditioned medium.
Naïve CD4+ T cells were cultured in supernatants
collected from BMDCs that had been subjected to different
treatments. Results indicated that the conditioned medium from
phenylalanine-primed, IMQ-stimulated DCs possessed a potent
capacity to drive Th17 polarization. T cells cultured in this
medium exhibited significantly upregulated gene expression of the
master transcription factor RAR-related orphan receptor C
(Rorc) and the effector cytokines Il17a, Il17f
and Il22 compared with controls (Fig. 6D). This was further corroborated
by flow cytometry, which exhibited a robust increase in the
population of CD4+ IL-17A+ T cells (Fig. 6E-G).
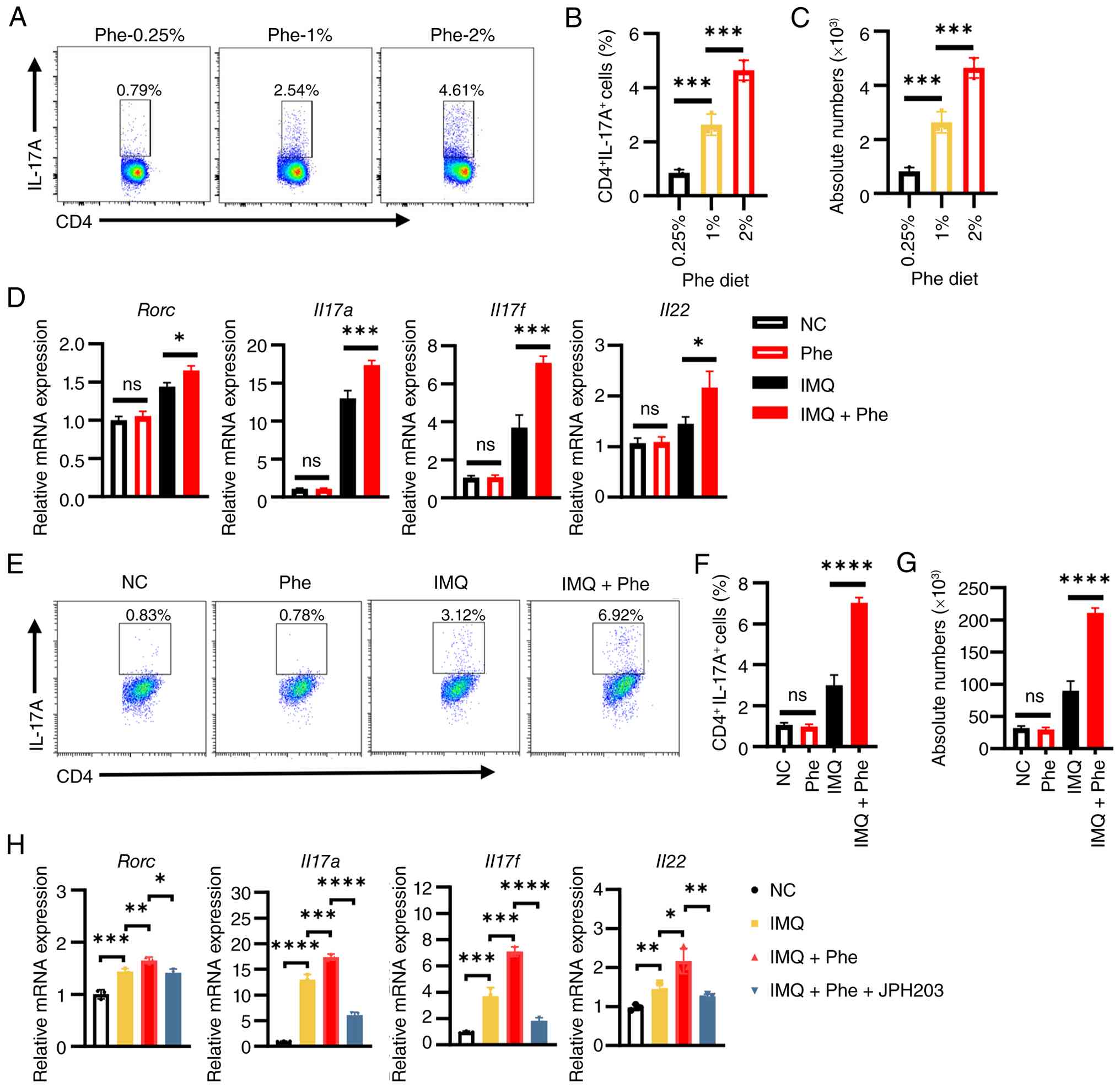 | Figure 6Phe amplifies the Th17 immune
response through dendritic cell modulation. (A) Representative flow
cytometry plots showing intracellular IL-17A staining in
CD4+ T cells isolated from the draining lymph nodes of
mice fed diets containing 0.25, 1 or 2% Phe. Quantification of the
frequency (B) and absolute number (C) of CD4+
IL-17A+ Th17 cells. (D) RT-qPCR analysis of
Th17-associated genes (Rorc, Il17a, Il17f and
Il22) expression in naïve CD4+ T cells cultured
for 3 days in conditioned medium derived from BMDCs treated as
indicated. (E) Representative flow cytometry plots of
CD4+ IL-17A+ T cells after in vitro
differentiation in DC-conditioned medium. (F,G) Quantification of
the (F) frequency and (G) absolute number of CD4
+IL-17A+ Th17 cells from the in vitro
assay. (H) RT-qPCR analysis of Th17 markers in T cells cultured in
conditioned medium from BMDCs treated with IMQ + Phe, in the
presence or absence of the L-type amino acid transporter 1
inhibitor JPH203. Data are representative of three independent
experiments and presented as the mean ± SEM. Statistical
significance was determined by one-way ANOVA followed by Tukey's
multiple comparisons test. *P<0.05,
**P<0.01, ***P<0.001 and
****P<0.0001. Phe, Phenylalanine; Th17, T helper cell
17; ns, not significant; RT-qPCR, reverse
transcription-quantitative PCR; DC, dendritic cell; BMDCs, bone
marrow-derived DCs; NC, normal control; RORC, RAR-related orphan
receptor C. |
Finally, to demonstrate that blocking phenylalanine
transport in DCs could reverse this pathogenic axis, LAT1 inhibitor
JPH203 was utilized. Notably, T cells cultured in supernatants from
DCs co-treated with JPH203 failed to promote a strong Th17
response, as evidenced by the normalized expression of Rorc,
Il17a, Il17f and Il22 (Fig. 6H). In conclusion, these findings
demonstrated that phenylalanine acted as a metabolic checkpoint
that instructed DCs to create a pro-Th17 microenvironment, thereby
sustaining the chronic inflammation characteristic of
psoriasis.
Discussion
Within the present study, the results highlighted
the key role of phenylalanine in psoriasis, which was associated
with amplified inflammatory responses. Elevated phenylalanine was
shown to exacerbate skin inflammation through enhanced NF-κB
signaling in DCs, ultimately promoting Th17 cell
differentiation.
The association between diet and psoriasis has
attracted marked attention. However, current dietary guidelines
predominantly stem from survey-based studies and lack robust direct
evidence (34). To the best of
our knowledge, the present study offers the first evidence that
phenylalanine, a central dietary component, serves a role in
exacerbating psoriasis symptoms. This amino acid is typically
present in high levels in red meat and serves as a primary
ingredient in aspartame, a commonly utilized artificial sweetener
(18). Consequently, a number of
patients may unknowingly consume excessive levels of phenylalanine,
unaware of its potential hazards. Understanding the effects of
phenylalanine on psoriasis is important, especially since
individuals with phenylketonuria often experience symptom
alleviation through dietary restriction of phenylalanine (35). These findings therefore highlight
the importance of dietary considerations in managing psoriasis,
potentially empowering patients to make informed dietary choices
that could positively affect their condition.
In the present study, a significant increase in
phenylalanine concentrations within the skin lesions and peripheral
blood of the psoriasis-like mouse model was observed. This finding
prompted the hypothesis that inhibition of the transporters
responsible for moving phenylalanine into cells may alleviate
psoriasis-associated inflammation. JPH203, known as a specific
inhibitor of LAT, has been shown in a previous study to ameliorate
cutaneous symptoms in a psoriasis-like mouse model when applied
topically (36). The present
experiments, both in vivo and in vitro, supported
this notion, highlighting the effectiveness of JPH203 in
attenuating the psoriasis-associated inflammatory response. JPH203
has been studied for its application in inhibiting tumor growth
(37) and modulating immune
responses by suppressing IL-17-producing T-cell expansion via the
PI3K/AKT/mTOR pathway (38), and
is currently undergoing clinical trials (39,40). However, the specific molecular
mechanisms underlying the anti-inflammatory effects of JPH203 in
the context of psoriasis remain to be fully elucidated.
DCs serve a central role in orchestrating immune
responses and inflammation in both healthy and diseased states.
Studies have indicated that tryptophan and branched-chain amino
acids alter the immunogenicity and tolerance of DCs (41,42). In addition, previous studies have
also shown that metal-ion-chelating L-phenylalanine nanostructures
activate DCs through the NOD-like receptor family pyrin domain
containing 3 inflammasome and the calcium-mediated NF-κB pathway
(43). In the present study, the
levels of p-p65 were found to be significantly elevated in a
phenylalanine dose-dependent manner. Through transcriptomics and
subsequent validation, it was determined that the presence of
phenylalanine amplified the inflammatory responses associated with
psoriasis in DCs. Furthermore, the inhibition of phenylalanine
transport led to suppression of the NF-κB pathway, suggesting that
phenylalanine partially exerted its effects by enhancing NF-κB
signaling.
Naive T cells can differentiate into Th17 cells when
induced by IL-6 and sustain their disease-associated phenotype in
the presence of IL-23 (44).
Therefore, the present study revealed that exposure to
phenylalanine enhanced the production of IL-1β, IL-6, IL-12 and
IL-23 in DCs, thus promoting a Th17-polarizing environment
(45). Furthermore, it was
observed that phenylalanine positively regulated the gene
expression of numerous chemokines, such as CXCL1, CXCL2 and CXCL3,
in DCs. These chemokines are likely key in recruiting neutrophils
and exacerbating tissue inflammation (31). Notably, this phenomenon can be
effectively suppressed by JPH203, reinforcing the notion that
phenylalanine serves a key role in Th17 differentiation within the
context of psoriasis.
A number of limitations in the present study warrant
future exploration. Although the present data and recent
metabolomics profiling demonstrated elevated phenylalanine levels
in the psoriatic lesional skin, the precise mechanisms driving this
local accumulation remain unclear. Recent studies have implicated
specific transporters, such as SLC16A10 (46) and the ultraviolet B-responsive
SLC6A15 (47) in cutaneous
phenylalanine uptake. Whether the dysregulation of these
transporters contributes to psoriasis lesions requires further
investigation. Consequently, while the present study observed
marked immune activation, the direct impact of accumulated
phenylalanine on keratinocyte dynamics remains to be fully
characterized. Given that phenylalanine hydroxylase is
transcriptionally upregulated during terminal differentiation
(48), future studies should aim
to experimentally validate whether exogenous phenylalanine directly
modulates keratinocyte proliferation rates. On a molecular level,
the direct intermediary mechanisms associating phenylalanine
exposure with NF-κB signaling activation, as well as the
comprehensive pharmacological pathways through which JPH203
operates in the psoriatic context, have yet to be fully elucidated.
Finally, the translational relevance of the present preclinical
findings necessitates future validation in human clinical
settings.
In conclusion, the present study demonstrated that
phenylalanine accumulation elevated the baseline expression of key
pro-inflammatory cytokines through the NF-κB signaling pathway,
thereby driving the pathogenic IL-23/IL-17 axis. Consequently, the
present study revealed a marked association between diet-derived
amino acids and immune regulation, offering a viable adjunctive
therapeutic strategy for psoriasis.
Supplementary Data
Availability of data and materials
The data generated in the present study may be found
in the China National Center for Bioinformation of the Genome
Sequence Archive under accession number PRJCA055303 or at the
following URL: (https://ngdc.cncb.ac.cn/bioproject/browse/PRJCA055303)
and in the Open Archive for Miscellaneous Data under accession
number OMIX014354 or at the following URL: (https://ngdc.cncb.ac.cn/omix/release/OMIX014354).
Authors' contributions
YX and YS conceived the present study. SV and HC
designed the present study. YX, ZM, JC and YZ performed the
experiments. SC, YW, JP and KL and MC analyzed the data and curated
the figures. YX and TY interpreted the data and wrote the
manuscript. HC, SV and MC revised the manuscript content. HC, YS
and YX confirm the authenticity of all the raw data. All authors
read and approved the final version of the manuscript.
Ethics approval and consent to
participate
All experimental procedures involving mice were
approved by the Animal Research Ethical Committee of the Sir Run
Run Shaw Hospital, Zhejiang University School of Medicine
(Hangzhou, China; approval no. SRRSH2025-0047).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 82373491) and the Zhejiang Province
Traditional Chinese Medicine Science and Technology Project (grant
no. 2024ZL587).
References
|
1
|
Griffiths CEM, Armstrong AW, Gudjonsson JE
and Barker J: Psoriasis. Lancet. 397:1301–1315. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Armstrong AW and Read C: Pathophysiology,
clinical presentation, and treatment of psoriasis: A Review. JAMA.
323:1945–1960. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ghoreschi K, Balato A, Enerbäck C and
Sabat R: Therapeutics targeting the IL-23 and IL-17 pathway in
psoriasis. Lancet. 397:754–766. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Masson Regnault M, Shourick J, Jendoubi F,
Tauber M and Paul C: Time to relapse after discontinuing systemic
treatment for psoriasis: A systematic review. Am J Clin Dermatol.
23:433–447. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Torres T, Chiricozzi A, Puig L, Lé AM,
Marzano AV, Dapavo P, Dauden E, Carrascosa JM, Lazaridou E, Duarte
G, et al: Treatment of psoriasis patients with latent tuberculosis
using IL-17 and IL-23 inhibitors: A retrospective, multinational,
multicentre study. Am J Clin Dermatol. 25:333–342. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yan L, Wang W, Dong M, Wang R and Li C:
Skin metabolic signatures of psoriasis and psoriasis concurrent
with metabolic syndrome. J Inflamm Res. 18:505–517. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yan L, Wang W, Qiu Y, Yu C, Wang R and Li
C: Role of glucose metabolism reprogramming in keratinocytes in the
link between psoriasis and metabolic syndrome. Int Immunopharmacol.
139:1127042024. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hong D, Xiong H, Lu S, Ma J and Shi Z:
Metabolic regulation of the immune cell in psoriasis: Mechanisms
and interventions. Curr Opin Immunol. 96:1026142025. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wan MT, Shin DB, Hubbard RA, Noe MH, Mehta
NN and Gelfand JM: Psoriasis and the risk of diabetes: A
prospective population-based cohort study. J Am Acad Dermatol.
78:315–322.e311. 2018. View Article : Google Scholar
|
|
10
|
Garshick MS, Ward NL, Krueger JG and
Berger JS: Cardiovascular risk in patients with psoriasis: JACC
review topic of the week. J Am Coll Cardiol. 77:1670–1680. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Herbert D, Franz S, Popkova Y, Anderegg U,
Schiller J, Schwede K, Lorz A, Simon JC and Saalbach A: High-fat
diet exacerbates early psoriatic skin inflammation independent of
obesity: Saturated fatty acids as key players. J Invest Dermatol.
138:1999–2009. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cappello A, Mancini M, Madonna S, Rinaldo
S, Paone A, Scarponi C, Belardo A, Zolla L, Zuccotti A, Panatta E,
et al: Extracellular serine empowers epidermal proliferation and
psoriasis-like symptoms. Sci Adv. 8:eabm79022022. View Article : Google Scholar
|
|
13
|
He X, Mo Y, Shi P, Xu Y, Zhou M and Zhang
T: Tryptophan metabolism in psoriasis and its complications: Future
opportunities. J Adv Res. 84:883–894. 2026. View Article : Google Scholar :
|
|
14
|
Chen C, Hou G, Zeng C, Ren Y, Chen X and
Peng C: Metabolomic profiling reveals amino acid and carnitine
alterations as metabolic signatures in psoriasis. Theranostics.
11:754–767. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pohla L, Ottas A, Kaldvee B, Abram K,
Soomets U, Zilmer M, Reemann P, Jaks V and Kingo K:
Hyperproliferation is the main driver of metabolomic changes in
psoriasis lesional skin. Sci Rep. 10:30812020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miao H, Bai Y, Shen S, Chu M, Miao C, Yang
J, Li X, Li L, Shao S, Wang G and Dang E: Biological agent exerts
therapeutic effects by reversing abnormalities in amino acid
metabolic pathways in psoriasis. Exp Dermatol. 33:e150592024.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kamleh MA, Snowden SG, Grapov D, Blackburn
GJ, Watson DG, Xu N, Ståhle M and Wheelock CE: LC-MS metabolomics
of psoriasis patients reveals disease severity-dependent increases
in circulating amino acids that are ameliorated by anti-TNFα
treatment. J Proteome Res. 14:557–566. 2015. View Article : Google Scholar :
|
|
18
|
Zhou Q, Sun WW, Chen JC, Zhang HL, Liu J,
Lin Y, Lin PC, Wu BX, An YP, Huang L, et al: Phenylalanine impairs
insulin signaling and inhibits glucose uptake through modification
of IRβ. Nat Commun. 13:42912022. View Article : Google Scholar
|
|
19
|
Czibik G, Mezdari Z, Altintas DM, Bréhat
J, Pini M, d'Humières T, Delmont T, Radu C, Breau M, Liang H, et
al: Dysregulated phenylalanine catabolism plays a key role in the
trajectory of cardiac aging. Circulation. 144:559–574. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Puig L, Costanzo A, Muñoz-Elías EJ, Jazra
M, Wegner S, Paul CF and Conrad C: The biological basis of disease
recurrence in psoriasis: A historical perspective and current
models. Br J Dermatol. 186:773–781. 2022. View Article : Google Scholar :
|
|
21
|
Zhang X, Li X, Wang Y, Chen Y, Hu Y, Guo
C, Yu Z, Xu P, Ding Y, Mi QS, et al: Abnormal lipid metabolism in
epidermal Langerhans cells mediates psoriasis-like dermatitis. JCI
Insight. 7:e1502232022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jaswal A, Kumar A, Patel P and Kurmi BD:
Immunometabolic dysregulation in psoriasis: Mechanisms driving
inflammation and emerging therapeutic targets. Immunol Invest.
55:948–974. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chang J, Voorhees TJ, Liu Y, Zhao Y and
Chang CH: Interleukin-23 production in dendritic cells is
negatively regulated by protein phosphatase 2A. Proc Natl Acad Sci
USA. 107:8340–8345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo Q, Jin Y, Chen X, Ye X, Shen X, Lin M,
Zeng C, Zhou T and Zhang J: NF-κB in biology and targeted therapy:
New insights and translational implications. Signal Transduct
Target Ther. 9:532024. View Article : Google Scholar
|
|
25
|
McGeachy MJ, Chen Y, Tato CM, Laurence A,
Joyce-Shaikh B, Blumenschein WM, McClanahan TK, O'Shea JJ and Cua
DJ: The interleukin 23 receptor is essential for the terminal
differentiation of interleukin 17-producing effector T helper cells
in vivo. Nat Immunol. 10:314–324. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Langley RG and Ellis CN: Evaluating
psoriasis with Psoriasis Area and Severity Index, Psoriasis Global
Assessment, and Lattice System Physician's Global Assessment. J Am
Acad Dermatol. 51:563–569. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Broggi A, Cigni C, Zanoni I and Granucci
F: Preparation of single-cell suspensions for cytofluorimetric
analysis from different mouse skin regions. J Vis Exp.
e525892016.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Love MI, Huber W and Anders S: Moderated
estimation of fold change and dispersion for RNA-seq data with
DESeq2. Genome Biol. 15:5502014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Robinson MD, McCarthy DJ and Smyth GK:
edgeR: A Bioconductor package for differential expression analysis
of digital gene expression data. Bioinformatics. 26:139–140. 2010.
View Article : Google Scholar
|
|
31
|
Chiang CC, Cheng WJ, Korinek M, Lin CY and
Hwang TL: Neutrophils in psoriasis. Front Immunol. 10:23762019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wohn C, Ober-Blöbaum JL, Haak S,
Pantelyushin S, Cheong C, Zahner SP, Onderwater S, Kant M,
Weighardt H, Holzmann B, et al: Langerin(neg) conventional
dendritic cells produce IL-23 to drive psoriatic plaque formation
in mice. Proc Natl Acad Sci USA. 110:10723–10728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu T, Zhang L, Joo D and Sun SC: NF-κB
signaling in inflammation. Signal Transduct Target Ther.
2:170232017. View Article : Google Scholar
|
|
34
|
Ford AR, Siegel M, Bagel J, Cordoro KM,
Garg A, Gottlieb A, Green LJ, Gudjonsson JE, Koo J, Lebwohl M, et
al: Dietary recommendations for adults with psoriasis or psoriatic
arthritis from the medical board of the national psoriasis
foundation: A Systematic Review. JAMA Dermatol. 154:934–950. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Deon M, Sitta A, Faverzani JL, Guerreiro
GB, Donida B, Marchetti DP, Mescka CP, Ribas GS, Coitinho AS,
Wajner M and Vargas CR: Urinary biomarkers of oxidative stress and
plasmatic inflammatory profile in phenylketonuric treated patients.
Int J Dev Neurosci. 47:259–265. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang X, Jiang Z, Huang S, Mao P, Zhang L,
Wang M, Ye J, Sun L, Sun M, Lu R, et al: Ultraviolet B
radiation-induced JPH203-loaded keratinocyte extracellular vesicles
exert etiological interventions for psoriasis therapy. J Control
Release. 362:468–478. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hayashi K and Anzai N: Novel therapeutic
approaches targeting L-type amino acid transporters for cancer
treatment. World J Gastrointest Oncol. 9:21–29. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cibrian D, Castillo-González R,
Fernández-Gallego N, la Fuente HD, Jorge I, Saiz ML, Punzón C,
Ramírez-Huesca M, Vicente-Manzanares M, Fresno M, et al: Targeting
L-type amino acid transporter 1 in innate and adaptive T cells
efficiently controls skin inflammation. J Allergy Clin Immunol.
145:199–214.e111. 2020. View Article : Google Scholar
|
|
39
|
Lemberg KM, Gori SS, Tsukamoto T, Rais R
and Slusher BS: Clinical development of metabolic inhibitors for
oncology. J Clin Invest. 132:e1485502022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nishikubo K, Ohgaki R, Okanishi H, Okuda
S, Xu M, Endou H and Kanai Y: Pharmacologic inhibition of LAT1
predominantly suppresses transport of large neutral amino acids and
downregulates global translation in cancer cells. J Cell Mol Med.
26:5246–5256. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen J, Liu J and Cao X: Functional and
metabolic heterogeneity of dendritic cells in self-tolerance and
autoimmunity. Immunol Rev. 336:e700682025. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kakazu E, Ueno Y, Kondo Y, Fukushima K,
Shiina M, Inoue J, Tamai K, Ninomiya M and Shimosegawa T: Branched
chain amino acids enhance the maturation and function of myeloid
dendritic cells ex vivo in patients with advanced cirrhosis.
Hepatology. 50:1936–1945. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tan M, Cao G, Wang R, Cheng L, Huang W,
Yin Y, Ma H, Ho SH, Wang Z, Zhu M, et al: Metal-ion-chelating
phenylalanine nanostructures reverse immune dysfunction and
sensitize breast tumour to immune checkpoint blockade. Nat
Nanotechnol. 19:1903–1913. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dong C: TH17 cells in development: An
updated view of their molecular identity and genetic programming.
Nat Rev Immunol. 8:337–348. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hawkes JE, Yan BY, Chan TC and Krueger JG:
Discovery of the IL-23/IL-17 signaling pathway and the treatment of
psoriasis. J Immunol. 201:1605–1613. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Luo L, Zeng H, Hu Y, Jiang L, Fu C, Huang
J, Chen J and Zeng Q: The amino acid transporter SLC16A10 promotes
melanogenesis by facilitating the transportation of phenylalanine.
Exp Dermatol. 33:e151652024. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou S, Ouyang Y, Hu Y, Dai X, Jiang L, Fu
C, Wen Y, Huang J, Zhang K, Chen J and Zeng Q: UVB enhances
SLC6A15-mediated phenylalanine transport to promote melanogenesis.
J Photochem Photobiol B. 274:1133292026. View Article : Google Scholar
|
|
48
|
Dörner C, Steinbinder J, Sachslehner AP,
Sukseree S and Eckhart L: Amino acid metabolism of the skin:
Control by specific enzymes and contribution to protective
functions. Metabolites. 15:2025. View Article : Google Scholar : PubMed/NCBI
|