Introduction
Endometrial carcinoma (EC) is the most common
gynaecological malignancy in the Western world (1). The rate is rising in developed
countries, most likely due to the rise in obesity and increased
life expectancies (2).
EC can be classified into two groups (3): type I lesions develop from excess
unopposed estrogen stimulation of the endometrium, causing
hyperplasia, which may progress to malignancy. These cancers
typically occur in younger, pre-menopausal women, are diagnosed
earlier and have a good prognosis. Type II tumours are estrogen
independent. They commonly occur in older, post-menopausal women,
are diagnosed much later than type I tumours and are usually more
aggressive histological subtypes (clear cell or serous carcinoma).
These high grade tumours have frequently already undergone
extensive invasion of surrounding structures or lymph node or
distant metastases, hence have a very poor prognosis.
Unfortunately, there is currently no appropriate
screening test available for EC. The main treatment strategy is
surgical, involving a total abdominal hysterectomy and bilateral
salpingo-oophorectomy. There is a lack of curative therapies for
patients diagnosed with late stage, recurrent or aggressive
disease. Adjuvant chemotherapy or radiotherapy is used for patients
with advanced disease (4).
Hormonal treatment with progestins are used clinically, but
response is restricted to those with hormone receptor-positive
tumours, and recurrence rate is high (5). Therefore, the identification of novel
factors involved in cancer progression is important in the
development of new pharmacological therapies to improve EC
prognosis.
Interleukin (IL) 11 is a member of the IL6 family of
cytokines. It signals via the IL11 receptor (R) α and gp130
receptor complex to activate the Janus tyrosine kinases (JAK).
This, in turn, can induce the activation of three independent
downstream signalling pathways: the signal transducer and activator
of transcription (STAT) pathway, the Ras-mitogen activated protein
kinase (MAPK) pathway; and the phosphotidylinositol-3 kinase
(PI-3K) pathway (6). In the human
endometrium, IL11 primarily acts via STAT3 (7). IL11 is a critical factor in the
establishment of pregnancy and is associated with several
pathologies of pregnancy (8–10).
Moreover, current studies have found a role for IL11 in trophoblast
migration and invasion (11,12),
processes that are similar to that of cancer progression. While
both trophoblast invasion and cancer cell invasion are similar,
trophoblast invasion is a highly controlled process unlike that of
cancer cell invasion.
Studies have reported a role for IL11 in several
epithelial cancers, including breast (13), gastric (14) and colorectal cancer (15). ECs show elevated levels of IL11 and
IL11Rα mRNA compared to normal proliferative phase endometrium
(16). Furthermore, we have
demonstrated that IL11 protein is upregulated in EC tissue and
uterine lavage fluid, compared to control post-menopausal
endometrial tissue and lavage fluid respectively (17). Whilst this evidence shows that IL11
is produced by EC, its function in EC is not yet known. In this
study, we investigated the effects of IL11 on proliferation,
adhesion and migration in endometrial cancer cell lines. We also
investigated the effects of IL11 inhibition on these actions.
Materials and methods
Materials
Human recombinant IL11 was obtained from R&D
Systems (Sydney, Australia) and diluted in 0.1% bovine serum
albumin (BSA) in phosphate-buffered saline (PBS). STAT3
phosphorylation inhibitor (STAT3i; 20 μM; Calbiochem, San
Diego, CA, USA). PEGylated IL11 inhibitor (IL11 inhibitor) was
produced by CSL Limited (Melbourne, Australia) (18,19).
Cell lines
Three endometrial cancer cell lines were used:
Ishikawa, HEC-1A and AN3CA cells, which are derived from grade I,
II and III EC, respectively (20–22).
The cell lines were cultured in DMEM, McCoy’s or MEM (Invitrogen,
Melbourne, Australia) respectively and supplemented with 10% fetal
bovine serum (FBS; SAFC Biosciences, Lenexa, KS, USA), 1%
L-glutamine (Invitrogen) and 1% penicillin-streptomycin
(Invitrogen).
SDS-PAGE and western blot analysis
Confluent cells were serum starved for 24 h before
treatment with control, IL11 (100 ng/ml, concentration chosen from
previous studies and concentration-response studies) (11) for 10, 30 or 60 min. Cell lysates
were collected using lysis buffer [50 mM Tris-Base, 150 mM NaCl, 2
mM EDTA, 2 mM EGTA, 25 mM NaF, and 25 mM B glycerolphosphate (pH
7.5)] containing protease inhibitor (2 μl/ml;
ThermoScientific, Waltham, MA, USA). Cell lysate (20 μg) was
loaded and electrophoresed on a 10% SDS-PAGE gel. The protein was
transferred to polyvinylidene fluoride (PVDF) membranes before it
was incubated with phosphorylated (p)-STAT3 (Tyr705) (no. 9131),
total STAT3 (no. 9132), phosphorylated (p)-p44/42 MAPK (Erk1/2)
(Thr202/Tyr204) (no. 4370), total p44/42 MAPK (Erk1/2) (no. 4695)
phosphorylated (p)-AKT (Ser473) (no. 4058) and total AKT (pan) (no.
4691) (Cell Signaling Technology, Danvers, MA, USA) primary
antibodies overnight at 4°C. Membranes were incubated with
horseradish peroxidase (HRP)-conjugated secondary antibody (50
μg/l; DakoCytomation) and HRP activity visualized using the
enhanced chemiluminescence system (ThermoScientific). Membranes
were exposed to film and developed, and the relative band intensity
determined using densitometric analysis (ImageJ v1.43, National
Institutes of Health, Bethesda, MD, USA).
Enzyme-linked immunoassays (ELISAs)
Phosphorylated (p)-STAT3 and total STAT3 were
measured using an established sandwich-ELISA (Cell Signaling
Technology) according to manufacturer’s protocol. Prior to assay,
confluent cells were serum starved for 24 h before treatment with
control, IL11 (100 ng/ml) + PEGylated IL11 inhibitor (IL11
inhibitor; 10 μg/ml) or IL11 inhibitor alone (10 g/ml). Cell
lysates were collected using lysis buffer [50 mM Tris-Base, 150 mM
NaCl, 2 mM EDTA, 2 mM EGTA, 25 mM NaF, and 25 mM B
glycerolphosphate (pH 7.5)] containing protease inhibitors (2
μl/ml; ThermoScientific).
Briefly, a 96-well plate was incubated overnight at
4°C with capture antibody. The plate was washed and blocked with
blocking buffer (1X PBS, 0.05% Tween-20, 1% BSA) for 1 h at 37°C.
The plate was washed four times and then incubated with 30
μg of protein lysates per well for 2 h at 37°C. A further
wash was required followed by incubation with detection antibody
for 1 h at 37°C. Following this, the plate was washed and secondary
antibody was applied for 30 min at 37°C before measuring the
absorbance at 450 nm with a Wallac Envision 2103 plate reader
(Perkin Elmer).
Bromodeoxyuridine (BrdU) assay
BrdU ELISA kit (Roche Applied Science, Indianapolis,
IN, USA) was used to assess cell proliferation as per the
manufacturer’s instructions and as previously described (12). Briefly, cells were seeded into
96-well plates (5,000 cells/well) and allowed to adhere for 8 h.
They were then serum starved for 24 h before treatment with
control, IL11 [100 ng/ml + IL11 inhibitor (10 μg/ml)] for 48
h at 37°C. BrdU (10 mM) was added to each well and incubated for 2
h at 37°C. BrdU incorporation was measured at 450 nm with a Wallac
Envision 2103 plate reader (Perkin Elmer).
WST-1 assay
A Wst-1 assay was used to determine the effect of
IL11 on cell viability as previously described (12). Cells were seeded into 96-well
plates (10,000 cells/well) and allowed to adhere for 8 h. Cells
were then serum starved for 24 h before treatment with control,
IL11 [100 + IL11 inhibitor (10 μg/ml) ng/ml] for 48 h at
37°C. Wst-1 dye (1:10; Roche Applied Science) was then added to
each well and incubated for 4 h at 37°C. The viable cell number was
determined by measuring the absorbance at 450 nm with a Wallac
Envision 2103 plate reader (Perkin Elmer).
Adhesion assay
To determine the effect of IL11 on cell adhesion,
CytoMatrix cell adhesion strips (Millipore, Sydney, Australia)
coated with extracellular matrix (ECM) proteins (fibronectin,
laminin, collagen I, collagen IV and vitronectin) were used
according to the manufacturer’s instructions. Briefly, cells were
serum starved for 24 h, then treated with IL11 [100 or 100 + IL11
inhibitor (10 μg/ml)]. Cells were seeded [50,000 (Ishikawa
and HEC-1A) or 60,000 (AN3CA)] into the wells and incubated for 2 h
at 37°C. Non-adherent cells were washed off and adherent cells
stained with 0.2% crystal violet in 50% ethanol solution. The stain
was eluted with solubilisation buffer (1:1 solution of
NaH2PO4 and 50% ethanol) and absorbance was
read at 560 nm with a Wallac Envision 2103 plate reader (Perkin
Elmer).
Migration assay
Chemotaxis cell (96-well) migration kits (Millipore)
were used according to the manufacturer’s instructions to assess
chemotactic cell migration. Briefly, 50,000 cells in 1% FBS were
seeded into each of the upper wells. Cells were pre-treated with
the STAT-3 inhibitor (10 μM) for 18 h prior to the migration
assay (11,23). Control, IL11 [(100 ng/ml) or IL11
(100 ng/ml) + IL11 inhibitor (10 μg/ml)] was added to the
lower wells and cells were incubated for 24 h at 37°C. A positive
control consisting of 10% FCS was added to the lower chamber as
previously described (12).
Migrated cells were detached, lysed and quantified with a dye by
measuring the fluorescence at 485/535 with a Wallac Envision 2103
plate reader (Perkin Elmer).
Statistical analysis
All data are expressed as mean ± standard error of
the mean (SEM) fold change in comparison to control for graphical
representation. Raw data were analysed using GraphPad Prism 5
(GraphPad Software, San Diego, CA, USA) for Windows. The raw data
were tested for normal distribution and the appropriate statistical
test utilized as described below. All functional assays had a
minimum of 3 independent experiments in triplicate wells, unless
otherwise stated. Normally distributed data were analysed
statistically by one-way analysis of variance for three or more
groups or a paired t-test was used to analyse differences between
two groups. Not normally distributed data were analysed by
Mann-Whitney (comparison of two groups) or Kruskal-Wallis
(comparison of three of more groups) tests. A p-value of <0.05
was considered statistically significantly different between
groups.
Results
IL11 signals via STAT3 in endometrial
cancer cell lines
We examined the effect of IL11 on three downstream
intracellular pathways. IL11 (100 ng/ml) phosphorylated (p)-STAT3
in all three cell lines after 10 min (Fig. 1A) and had no effect on STAT3
protein abundance. Moreover, a sandwich-ELISA, used for
quantitative STAT3 phosphorylation analysis in the AN3CA cells,
demonstrated that IL11 (100 ng/ml) significantly upregulated
(p)-STAT3 compared to control and this was reversed by co-treatment
with the IL11 inhibitor (Fig. 1B).
Treatment of ANC3A cells with IL11 inhibitor alone or IL11 at a
concentration of 1 ng/ml did not alter (p)-STAT3 compared to
control. As IL11 is known to activate a number of other signalling
pathways in cancer cells we investigated whether IL11 altered the
MAPK or PI3K pathways. We demonstrated that IL11 did not
activate/enhance the MAPK or PI3K signalling pathways in Ishikawa,
HEC-IA and ANC3A cells [Fig. 1C (i)
and (ii)].
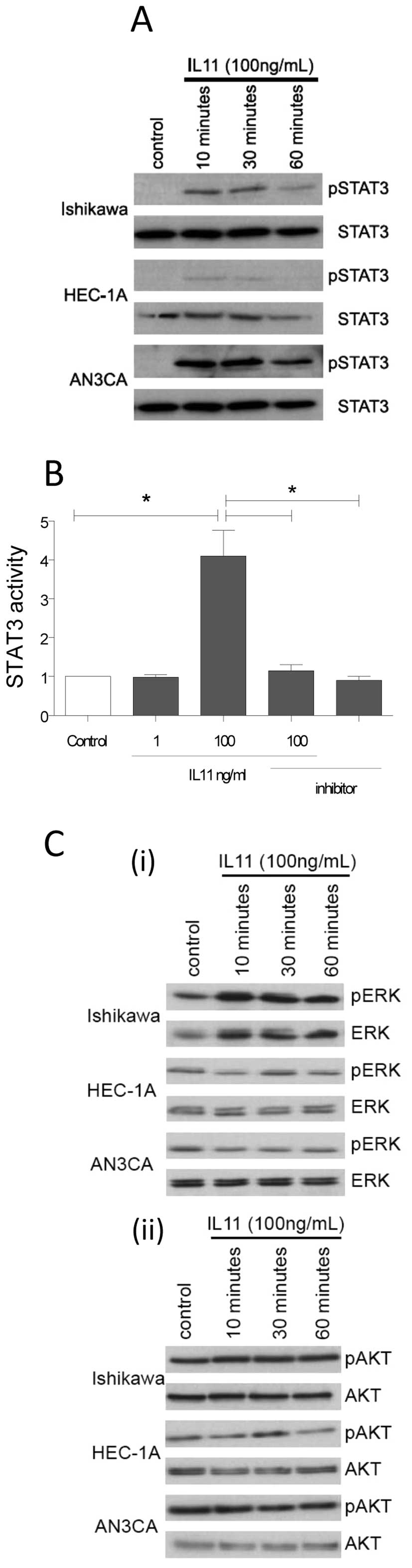 | Figure 1IL11 signals via phosphorylated STAT3
in AN3CA, Ishikawa and HEC-1A cells. (A) Cells were cultured with
IL11 (100 ng/ml) and the abundance of phosphorylated (p) STAT3 and
STAT3 was determined after 10, 30 and 60 min. Data are
representative of n=3 experiments. (B) pSTAT3 ELISA assay: ANC3A
cells were treated with control, IL11 [1, 100, 100 ng/ml +
PEGylated IL11 inhibitor (IL11 inhibitor; 10 μg/ml) and IL11
inhibitor (10 μg/ml) alone] for 10 min. Data are expressed
as mean (expressed as fold change from control) ± SEM and
represents triplicate treatments from n=3 individual experiments.
*P<0.05; IL11 (100 ng/ml) compared to all other
groups. (C) Effect of IL11 on ERK and AKT phosphorylation in
Ishikawa, HEC-1A and AN3CA cells. Cells were cultured with control,
IL11 (100 ng/ml) for 10, 30 and 60 min. Following western blot
analysis protein bands for pERK (42, 44 kDa), total ERK (42, 44
kDa), p-Ser473-AKT (60 kDa) and total AKT (60 kDa) were detected.
Data are representative of two independent experiments assayed in
triplicate. |
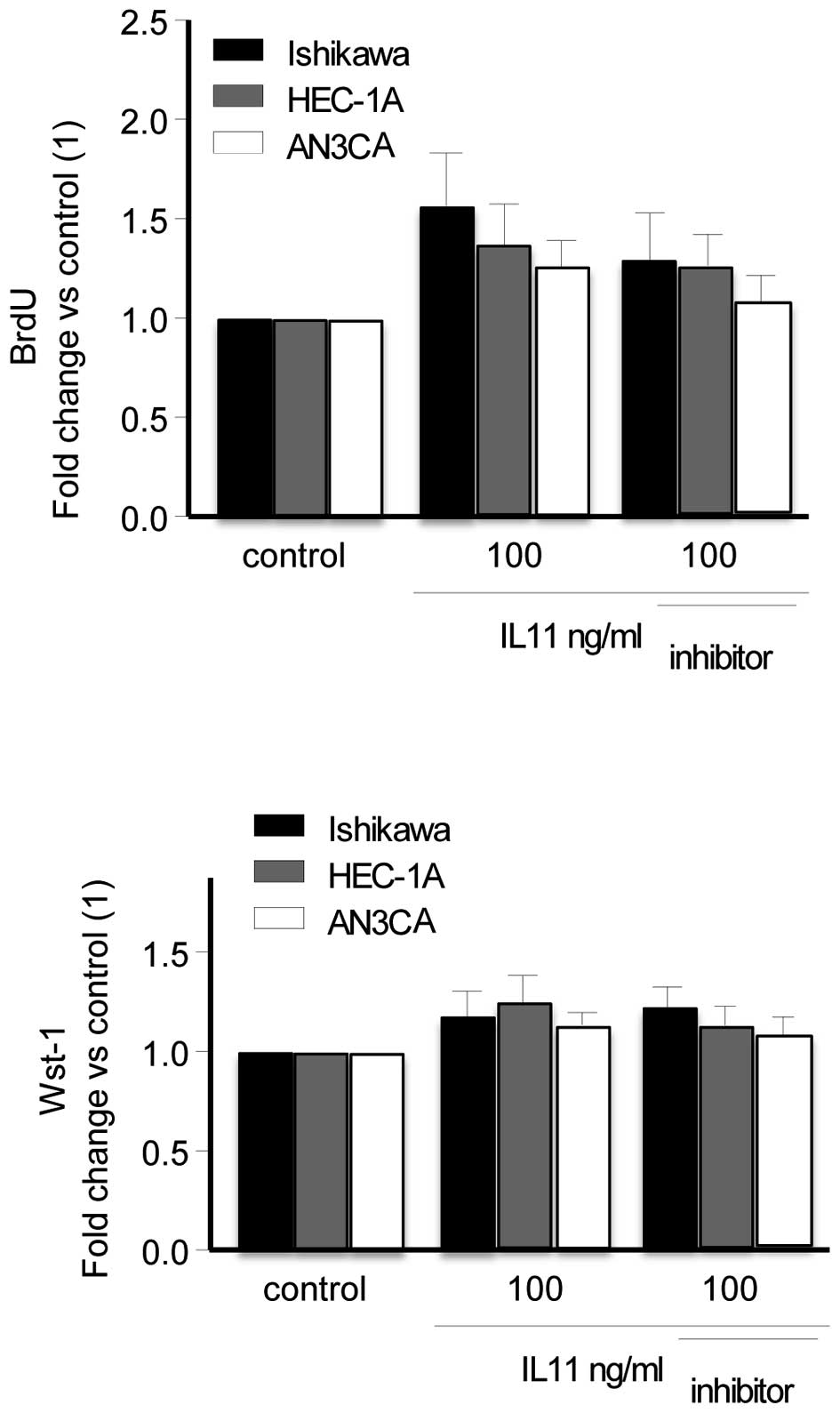
IL11 had no effect on cell proliferation
or viability
To determine the effect of IL11 on cell
proliferation and cell viability, BrdU and Wst-1 assays were used,
respectively. IL11 (100 ng/ml) or IL11 (100 ng/ml) + IL11 inhibitor
(10 μg/ml) had no significant effect on cell proliferation
(BrdU; Fig. 2A) or cell viability
(Wst-1; Fig. 2B) in the three cell
lines. IL11 inhibitor alone did not affect cell proliferation (data
not shown).
IL11 alters endometrial cancer cell
adhesion
Using the three different cell lines Ishikawa,
HEC-1A and AN3CA (as models of grade I, II and III, respectively)
we investigated the effects of IL11 on EC cell adhesion to ECM
proteins. AN3CA cells showed maximal binding to fibronectin,
laminin and vitronectin but no binding to both collagen I and IV
(Fig. 3A). AN3CA cells treated
with IL11 (100 ng/ml) increased binding to fibronectin (P<0.05);
Fig. 3B) compared to control.
Co-culture of AN3CA cells with IL11 (100 ng/ml) and IL11 inhibitor
or STAT3 inhibitor significantly decreased adhesion to fibronectin
compared to cells treated with IL11 (100 ng/ml) (Fig. 3B). Treatment of Ishikawa and HEC-1A
cells with IL11 (100 ng/ml) did not significantly alter cell
adhesion to the ECM (data not shown).
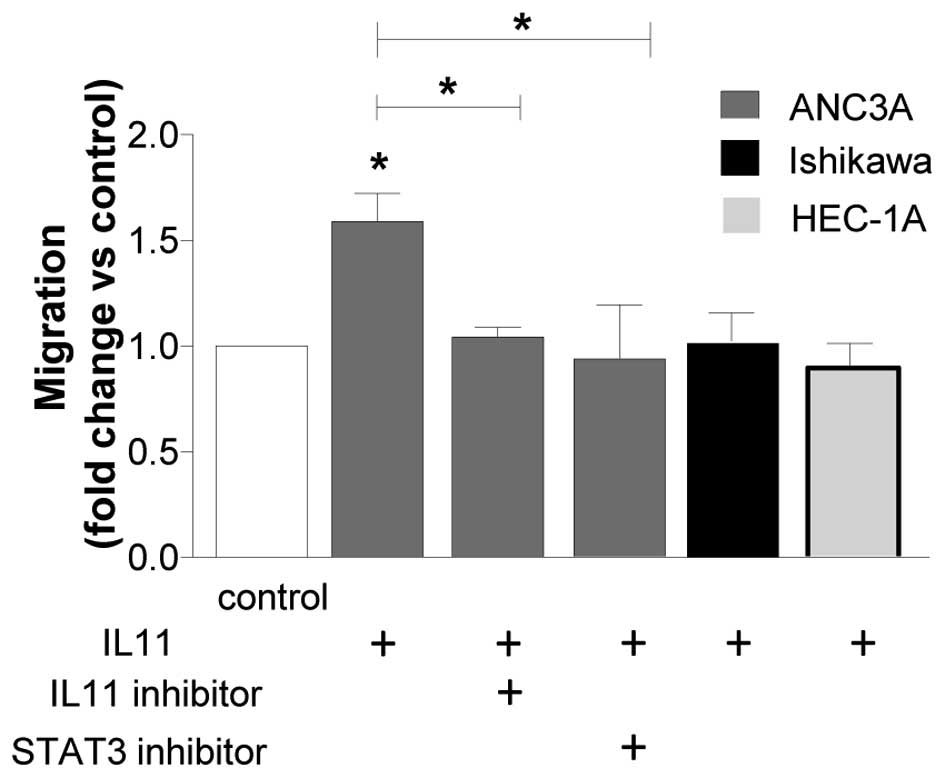
IL11 regulates ANC3A cell migration via
STAT3
The effect of IL11 on cell migration was examined in
ANC3A, Ishikawa and HEC-1A cells (Fig.
4). AN3CA cells treated with IL11 (100 ng/ml) resulted in a 50%
increase in cell migration compared to control (P<0.05; Fig. 4). This effect was reversed by
co-incubation with IL11 (100 ng/ml) and the IL11 specific inhibitor
(10 μg/ml) (Fig. 4).
Similarly, ANC3A cells co-incubated with IL11 and the STAT3
inhibitor reduced cell migration compared to cells treated with
IL11 alone. By contrast, IL11 had no effect on Ishikawa and HEC-1A
cell migration (Fig. 4).
Discussion
This study investigated the effect of IL11 in
endometrial cancer cell function – proliferation, viability,
adhesion and migration using three human endometrial carcinoma cell
lines (Ishikawa, HEC-1A and AN3CA). We demonstrated that IL11
increased cell adhesion of ANC3A cells to fibronectin but did not
affect Ishikawa and HEC-1A cell adhesive properties. IL11 had no
effect on cell proliferation or viability. By contrast, IL11
increased AN3CA cell migration while it did not significantly alter
Ishikawa or HEC-1A cell migration. A unique IL11 inhibitor reversed
the observed effects on both cell adhesion and migration.
Similarly, a STAT3 inhibitor blocked the IL11 mediated effects on
ANC3A cell adhesion suggesting IL11 regulated these processes via
(p)-STAT3. In agreement, IL11 phosphorylated (p)-STAT3 and did not
affect the MAPK or PI3K pathways in the Ishikawa, HEC-1A and AN3CA
cells.
IL11 signalling is initiated via the IL11Rα and the
gp130 receptor complex (24). In
the endometrium, IL11 primarily signals via the JAK/STAT
intracellular pathway (25). This
is also the case in endometrial cancer cells as demonstrated in the
present study. JAK/STAT signalling has been well studied in other
cancers (26), including breast
cancer, pancreatic, colorectal, prostate and head and neck cancer
as well as leukaemia and lymphoma (27). Evidence shows that inappropriate
STAT activation results in an environment that is conducive to
tumour development, such as inhibition of apoptosis, promotion of
angiogenesis and proliferation (28). For instance, IL11 signals via STAT3
to increase tumourigenesis in gastric cancer, as demonstrated with
a gp130 Y757 knock-in mutant mouse model (14). In the present study we demonstrated
that blocking the IL11 pathway inhibited adhesion and migration of
the ANC3A cells that were derived from grade III (undifferentiated)
tumours via the activation of STAT3.
IL11 had no effect on cell proliferation or
viability in all three cancer cell lines tested. This finding is
consistent with the lack of effect of IL11 on proliferation of
human trophoblast cells (12),
lung and colon cancers (29). It
is well known that IL11 has an anti-apoptotic effect under
stressful conditions. In the gastrointestinal system, IL11 protects
against mucosal injury in diseases such as inflammatory bowel
disease, intestinal ischemia, necrotising enterocolitis and
irradiation. However, from our data, it is unlikely that IL11
prevents apoptosis due to the lack of effect on cell proliferation
and cell viability.
We demonstrated that IL11 significantly altered
ANC3A cell adhesion to fibronectin, while no significant effect was
seen with the other extracellular matrix proteins tested and
furthermore no effect on Ishikawa and HEC-1A cell adhesion was
detected. By comparison we previously demonstrated that IL11
stimulated cell adhesion of human endometrial epithelial cells to
fibronectin and collagen IV, as well as primary trophoblast cells
(30). Integrins are likely
candidates by which cells regulate adhesion (22). Whether IL11 regulates integrins in
endometrial cancer cells, however, remains to be determined.
Tumours of higher grades are more likely to be less adhesive,
lending themselves to a more migratory phenotype. This has been
found in pancreatic (31) and
prostate cancer (32).
Paradoxically, the present study demonstrated IL11 increased ANC3A
cell adhesion to fibronectin and migration. Cell migration involves
the adhesion and de-adhesion of cells and our data suggest there
may be mechanisms additional to cell adhesion by which IL11
facilitated ANC3A cell migration. A recent study reported, that
fibronectin gene expression is reduced in endometrial cancer tissue
compared to normal endometrium and suggest that fibronectin may be
useful as a marker of disease progression (33).
IL11 increased migration of ANC3A cells via
(p)-STAT3 while no effect was seen with IL11 on Ishikawa and HEC-1A
cell migration. IL11 maximally activated (p)-STAT3 in the ANC3A
cells compared to the Ishikawa and HEC-1A cells suggesting that
IL11 was less responsive to the latter two cell lines.
Interestingly, Ishikawa and HEC-1A cells are derived from grade I
and II endometrial cancers, while ANC3A cell are derived from grade
III cancers suggesting that IL11 has a role primarily in the
migratory higher grade tumours. Similarly, IL11 has previously been
demonstrated to stimulate the migration of EVT-hybridoma cells
(12) and outgrowth of primary
extravillous trophoblast cells (23). In other cancers such as colorectal
cancer, IL11 increases cell migration via the PI3K pathway and
inhibiting this pathway reverses the effect (15). Similarly IL11 stimulates migration
in other cells, including breast cancer cell lines (34) and endothelial progenitor cells
(35).
A number of studies demonstrate that IL11 increases
the invasive capacity of many other cancers. IL11 increases
invasion of JEG3 choriocarcinoma cells (36), squamous cell carcinomas (29) and breast cancer cells (37). In gastric cancer, IL11 promotes
cellular invasion via the STAT3 pathway (14). Similarly, in colorectal cancer,
IL11 levels are associated with the extent of tumour invasion and
infiltration into vessels (15).
In conclusion, these results demonstrate a role for
IL11 in endometrial carcinoma cells likely acting via (p)-STAT3.
IL11 is upregulated in grade I tumours and in uterine lavage fluid
(17) suggesting IL11 may be
useful as a marker for endometrial cancer. This study supports a
role for IL11 in endometrial cancer progression and in vivo
studies are required to determine whether targeting IL11 may be
useful as a treatment option for endometrial cancer
progression.
Acknowledgements
We thank Dr Ellen Menkhorst for her
contribution in editing the manuscript and technical advice. We
thank Kirsten Edwards, Brent McKenzie, Andrew Nash and Pierre
Scotney from CSL Biotherapies for provision of the IL11 antagonist.
This study was funded by the NHMRC of Australia: ED is the funding
recipient of Career Development Fellowship no. 550905 and V.L.,
J.Y. and S.S. are funded from project grant no. 388901. The study
was also supported by the Victorian Government’s Operational
Infrastructure Support Program. PHI Data Audit 11–16.
References
|
1
|
Australian Institute of Health and Welfare
(AIHW): Australasian Association of Cancer Registries (AACR):
Cancer in Australia: an overview, 2008. Cancer series no. 46. Cat.
no. CAN 42. AIHW; Canberra: 2008
|
|
2
|
Amant F, Moerman P, Neven P, Timmerman D,
Van Limbergen E and Vergote I: Endometrial cancer. Lancet.
366:491–505. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bokhman JV: Two pathogenetic types of
endometrial carcinoma. Gynecol Oncol. 15:10–17. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tangjitgamol S, Anderson BO, See HT, et
al: Management of endometrial cancer in Asia: consensus statement
from the Asian Oncology Summit 2009. Lancet Oncol. 10:1119–1127.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Niwa K, Tagami K, Lian Z, Onogi K, Mori H
and Tamaya T: Outcome of fertility-preserving treatment in young
women with endometrial carcinomas. BJOG. 112:317–320. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Heinrich PC, Behrmann I, Haan S, Hermanns
HM, Muller-Newen G and Schaper F: Principles of interleukin
(IL)-6-type cytokine signalling and its regulation. Biochem J.
374:1–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dimitriadis E, Stoikos C, Tan YL and
Salamonsen LA: Interleukin 11 signaling components signal
transducer and activator of transcription 3 (STAT3) and suppressor
of cytokine signaling 3 (SOCS3) regulate human endometrial stromal
cell differentiation. Endocrinology. 147:3809–3817. 2006.
View Article : Google Scholar
|
|
8
|
Chen HF, Lin CY, Chao KH, Wu MY, Yang YS
and Ho HN: Defective production of interleukin-11 by decidua and
chorionic villi in human anembryonic pregnancy. J Clin Endocrinol
Metab. 87:2320–2328. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Karpovich N, Chobotova K, Carver J, Heath
JK, Barlow DH and Mardon HJ: Expression and function of
interleukin-11 and its receptor alpha in the human endometrium. Mol
Hum Reprod. 9:75–80. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
von Rango U, Alfer J, Kertschanska S, et
al: Interleukin-11 expression: its significance in eutopic and
ectopic human implantation. Mol Hum Reprod. 10:783–792.
2004.PubMed/NCBI
|
|
11
|
Paiva P, Salamonsen LA, Manuelpillai U and
Dimitriadis E: Interleukin 11 inhibits human trophoblast invasion
indicating a likely role in the decidual restraint of trophoblast
invasion during placentation. Biol Reprod. 80:302–310. 2009.
View Article : Google Scholar
|
|
12
|
Paiva P, Salamonsen LA, Manuelpillai U, et
al: Interleukin-11 promotes migration, but not proliferation, of
human trophoblast cells, implying a role in placentation.
Endocrinology. 148:5566–5572. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hanavadi S, Martin TA, Watkins G, Mansel
RE and Jiang WG: Expression of interleukin 11 and its receptor and
their prognostic value in human breast cancer. Ann Surg Oncol.
13:802–808. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ernst M, Najdovska M, Grail D, et al:
STAT3 and STAT1 mediate IL-11-dependent and inflammation-associated
gastric tumorigenesis in gp130 receptor mutant mice. J Clin Invest.
118:1727–1738. 2008.PubMed/NCBI
|
|
15
|
Yamazumi K, Nakayama T, Kusaba T, et al:
Expression of interleukin-11 and interleukin-11 receptor alpha in
human colorectal adenocarcinoma; immunohistochemical analyses and
correlation with clinicopathological factors. World J
Gastroenterol. 12:317–321. 2006.
|
|
16
|
Sales KJ, Grant V, Cook IH, et al:
Interleukin-11 in endometrial adenocarcinoma is regulated by
prostaglandin F2alpha-F-prostanoid receptor interaction via the
calcium-calcineurin-nuclear factor of activated T cells pathway and
negatively regulated by the regulator of calcineurin-1. Am J
Pathol. 176:435–445. 2010. View Article : Google Scholar
|
|
17
|
Yap J, Salamonsen LA, Jobling T, Nicholls
PK and Dimitriadis E: Interleukin 11 is upregulated in uterine
lavage and endometrial cancer cells in women with endometrial
carcinoma. Reprod Biol Endocrinol. 8:632010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee CG, Hartl D, Matsuura H, et al:
Endogenous IL-11 signaling is essential in Th2- and IL-13-induced
inflammation and mucus production. Am J Resp Cell Mol Biol.
39:739–746. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Menkhorst E, Salamonsen L, Robb L and
Dimitriadis E: IL11 antagonist inhibits uterine stromal
differentiation, causing pregnancy failure in mice. Biol Reprod.
80:920–927. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dimitriadis E, Stoikos C, Baca M, Fairlie
WD, McCoubrie JE and Salamonsen LA: Relaxin and prostaglandin E(2)
regulate interleukin 11 during human endometrial stromal cell
decidualization. J Clin Endocrinol Metab. 90:3458–3465. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hilton DJ, Hilton AA, Raicevic A, et al:
Cloning of a murine IL-11 receptor alpha-chain; requirement for
gp130 for high affinity binding and signal transduction. EMBO J.
13:4765–4775. 1994.PubMed/NCBI
|
|
22
|
Marwood M, Visser K, Salamonsen LA and
Dimitriadis E: Interleukin-11 and leukemia inhibitory factor
regulate the adhesion of endometrial epithelial cells: implications
in fertility regulation. Endocrinology. 150:2915–2923. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sonderegger S, Yap J, Menkhorst E, Weston
G, Stanton PG and Dimitriadis E: Interleukin (IL)11 mediates
protein secretion and modification in human extravillous
trophoblasts. Hum Reprod. 26:2841–2849. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Painter KJ, Armstrong NJ and Sherratt JA:
The impact of adhesion on cellular invasion processes in cancer and
development. J Theor Biol. 264:1057–1067. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Arao S, Masumoto A and Otsuki M: Beta1
integrins play an essential role in adhesion and invasion of
pancreatic carcinoma cells. Pancreas. 20:129–137. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Torimura T, Ueno T, Kin M, et al: Integrin
alpha6beta1 plays a significant role in the attachment of hepatoma
cells to laminin. J Hepatol. 31:734–740. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakahara H, Nomizu M, Akiyama SK, Yamada
Y, Yeh Y and Chen WT: A mechanism for regulation of melanoma
invasion. Ligation of alpha6beta1 integrin by laminin G peptides. J
Biol Chem. 271:27221–27224. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lessey BA: Integrins and the endometrium:
new markers of uterine receptivity. Ann NY Acad Sci. 828:111–122.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang J, Homer RJ, Hong L, et al: IL-11
selectively inhibits aeroallergen-induced pulmonary eosinophilia
and Th2 cytokine production. J Immunol. 165:2222–2231. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luo X, Ding L and Chegini N:
Gonadotropin-releasing hormone and TGF-beta activate MAP kinase and
differentially regulate fibronectin expression in endometrial
epithelial and stromal cells. Am J Physiol Endocrinol Metab.
287:E991–E1001. 2004. View Article : Google Scholar
|
|
31
|
Jarnicki A, Putoczki T and Ernst M: Stat3:
linking inflammation to epithelial cancer - more than a ‘gut’
feeling? Cell Div. 5:142010.PubMed/NCBI
|
|
32
|
Seval Y, Cakmak H, Kayisli UA and Arici A:
Estrogen-mediated regulation of p38 mitogen-activated protein
kinase in human endometrium. J Clin Endocrinol Metab. 91:2349–2357.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Futyma K, Kubiatowski T, Rozynska K, et
al: Decreased osteonectin and fibronectin gene expression in
endometrial cancer cancer as a prognostic marker. Ginekol Pol.
80:907–913. 2009.(In Polish).
|
|
34
|
Soda H, Raymond E, Sharma S, et al:
Recombinant human interleukin-11 is unlikely to stimulate the
growth of the most common solid tumors. Anticancer Drugs.
10:97–101. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pignatelli M, Ansari TW, Gunter P, et al:
Loss of membranous E-cadherin expression in pancreatic cancer:
correlation with lymph node metastasis, high grade, and advanced
stage. J Pathol. 174:243–248. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suman P, Poehlmann TG, Prakash GJ, Markert
UR and Gupta SK: Interleukin-11 increases invasiveness of JEG-3
choriocarcinoma cells by modulating STAT3 expression. J Reprod
Immunol. 82:1–11. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gentilini D, Busacca M, Di Francesco S,
Vignali M, Vigano P and Di Blasio AM: PI3K/Akt and ERK1/2
signalling pathways are involved in endometrial cell migration
induced by 17beta-estradiol and growth factors. Mol Hum Reprod.
13:317–322. 2007. View Article : Google Scholar : PubMed/NCBI
|