Introduction
Hepatocellular carcinoma (HCC) is a common
malignancy worldwide. Of the numerous therapies available, hepatic
resection is currently the first choice, although it is often
accompanied by a poor prognosis. Another therapeutic option, liver
transplantation, is not widely used as there is a shortage of
livers available, and many patients succumb to the disease while
waiting for a transplant (1).
Several experts have focused on biotherapy for decades; however,
the mechanism underlying HCC development remains unclear, making it
difficult to develop effective therapies.
Following injury, non-parenchymal cells, especially
hepatic stellate cells (HSCs), are activated and change from a
quiescent state to myofibroblast-like cells. Activated HSCs can
secrete cytokines and produce collagen (2,3), and
they can potentially contribute to the repair of liver damage.
However, the excessive activation of HSCs can induce liver fibrosis
(4–6), which can progress to hepatocirrhosis
or even HCC. As a result of these studies, it has been suggested
that there may be a relationship between HSCs and HCC.
Recent studies have confirmed that HSCs have
immunomodulatory activities and can inhibit T-cell-mediated tissue
destruction and, notably, can prolong the survival of allografts
(7–11). Furthermore, activated HSCs can
promote the occurrence and development of HCC (12,13).
In our previous study, we found that activated HSCs induce an
immunosuppressive microenvironment that promotes HCC development
in vivo(14). However, how
HSCs affect T cells has yet to be elucidated. Additionally, whether
T cells can still attack cancer cells when their functions are
inhibited by HSCs also remains unknown.
In this study, we investigated whether HSCs could
block the cancer-fighting ability of T cells and therefore promote
the development of HCC in vitro. We found that activated
HSCs induced T cell hyporesponsiveness by promoting T-cell
apoptosis and expanding regulatory T cells (Treg cells), thereby
changing the balance of the different T-cell populations in mixed
leukocyte reactions (MLRs). In addition, HSCs inhibited the
cytotoxic T cell-mediated lysis of tumour cells. Moreover,
activated HSCs altered the cytokines that were released during the
MLRs, and this alteration could promote the development of HCC.
Materials and methods
Animals
C57BL/6 (B6; H2b) and BALB/c (H2d) mice were
purchased from the National Rodent Laboratory Animal Resources,
Shanghai branch. All animals were maintained under specific
pathogen-free conditions in the Laboratory Animal Centre of Xiamen
University Medical Department and provided with rodent chow and tap
water. All mice were 8–12 weeks old when used, and all experiments
were performed following the Laboratory Animal Centre care
guidelines.
Isolation, culture, and identification of
HSCs
HSCs were isolated and cultured as previously
described (14,15). Cell viability was determined using
trypan blue exclusion, and the appearance of the HSCs was verified
using light microscopy. The purity of the HSCs was examined using
desmin immunofluorescence. HSCs were obtained after passage for 2–3
generations, and the cells were plated on glass slides, fixed at
70–80% confluence, and stained with antibodies specific for desmin
(NeoMarker) or α-smooth muscle actin (α-SMA, Abcam). The stained
HSCs were examined using confocal microscopy.
Isolation and culture of dendritic cells
(DCs)
Bone marrow cells were isolated from C57BL/6 mouse
femurs and tibias. After lysing the red blood cells with lysis
buffer (Beyotime), the remaining cells were cultured in 6-well
plates (Corning) in RPMI-1640 medium (Hyclone) supplemented with
10% fetal bovine serum (FBS) (complete RPMI-1640 medium) in the
presence of mouse recombinant granulocyte-macrophage
colony-stimulating factor (GM-CSF, 10 ng/ml, Hangzhou LongGene) and
interleukin-4 (IL-4,10 ng/ml, Peprotech), as previously described
(16). The media was replaced
every 2 days. Non-adherent cells were spontaneously released from
the proliferating cell clusters, and these floating cells were
harvested on Day 7.
Preparation of T lymphocytes
Enriched T cells were prepared as previously
described (17) with minor
modifications. Spleens were isolated from BALB/c mice under sterile
conditions and pushed through a 75-μm steel mesh. After the
red blood cells were lysed, the remaining cells were placed in a
nylon wool column and incubated for 45 min at 37°C in 5%
CO2 in air. The non-adherent cells were then collected
for experiments. The purity of the T cells was >90%.
T-cell proliferation assay
One-way MLRs assays were performed in triplicate in
96-well microculture plates (Corning). First, HSCs were treated
with mitomycin C (40 μg/ml) for 25 min and washed twice
before use. Nylon wool-purified splenic T cells from BALB/c mice
were used as responders (2×105) and were co-cultured
with mitomycin C (20 μg/ml)-treated DCs at a ratio of 10:1.
To evaluate the ability of HSCs to inhibit T-cell proliferation,
mitomycin C-treated HSCs were added at the beginning of each MLRs
at different ratios (T:HSCs = 10:1, 20:1, 40:1, or 80:1), while the
control had no HSCs added. All of the cells were cultured in
complete RPMI-1640 medium for 72 h before proliferation was
measured using the BrdU Cell Proliferation Assay (Roche) and a
microtitre plate reader at 450 nm. The supernatant was collected
separately.
Hepa1-6-cell proliferation assay
The supernatant (referred to as ‘conditioned
medium’) from the MLRs was collected. To determine the effect of
the conditioned medium on tumour cell proliferation, Hepa1-6 cells
(purchased from the Shanghai Cell Bank, Chinese Academy of
Sciences) were plated in triplicate at 4×103 cells/well
in 96-well plates and allowed to adhere overnight. The culture
medium was then replaced with the conditioned medium. After 72 h,
cell proliferation was measured using the BrdU Cell Proliferation
Assay. The absorbance values are directly correlated with the
amount of DNA synthesis and, thus, with the number of proliferating
cells in culture.
In vitro T-cell apoptosis assay
Purified T cells and mitomycin C-treated DCs with or
without mitomycin C-treated HSCs were co-cultured T:DC:HSCs at a
ratio of 20:2:1 in 24-well plates (Corning). After 72 h, the cells
were collected, stained with fluorescein isothiocyanate
(FITC)-conjugated Annexin V and propidium iodide (PI) using the
Apoptosis Analysis Kit (Keygen, Nanjing), according to the
manufacturer’s protocol, and analysed using flow cytometry. In
addition, the supernatant was collected.
Detection of Treg cells
The 3 types of cells (T cells, DCs, and HSC) were
co-cultured as described for the apoptosis assay. After 72 h of
incubation, the cells were collected and examined using the Mouse
Regulatory T Cell Staining Kit (eBioscience) according to the
manufacturer’s instructions. The supernatant was also
collected.
In vitro cytotoxic T-lymphocyte
assay
Hepa1–6 cells were collected and subjected to 4
cycles of snap freeze-thaw to obtain tumour lysates that could be
used to pulse DCs. DCs were stimulated using the tumour lysate on
day 4 of culture, incubated for another 48 h, and collected as
stimulators. As targets, Hepa1–6 cells were labelled with CFSE (10
μM). The effectors, stimulators, and targets were plated at
a ratio of 50:5:1 (T:DC:Hepa1–6, T cells = 2×106).
Mitomycin C-treated HSCs were added to the experimental groups at
the start of the experiment. After co-culturing the cells for 72 h,
the cells were harvested, stained using PI (1 mg/ml) in a total
volume of 500 μl, and examined using flow cytometry. The
CFSE/PI double-positive cells were considered as dead Hepa1–6
cells; therefore, the percentage of CFSE/PI double-positive cells
was as the percentage of the mortality rate of Hepa1–6.
Transwell assay
Transwell migration chambers (Corning) were used to
observe the migration of HCC cells. Hepa1–6 cells
(5×104) in 200 μl serum-free medium were added to
the upper chamber, and the supernatant (800 μl) collected
from the MLRs experiments with or without HSCs was added to the
lower chamber. The cells were allowed to migrate for 18 h at 37°C
in a 5% CO2 atmosphere. The non-migrating cells on the
upper surface of the membrane were removed using a cotton swab. The
remaining cells were fixed in methanol, stained using crystal
violet, and air-dried. The number of migrating cells on each
membrane was counted using a microscope.
Cytokine analysis
To measure cytokine production in the MLRs, the
collected supernatant was lyophilised to a total volume of 1 ml and
analysed using the Mouse Cytokine Array Panel A (R&D), as
recommended by the manufacturer. The data were analysed using image
analysis software and the expression of cytokines with fold change
>±1.5 in the experimental group are shown.
Animal model
In vivo tumour growth was measured using a
previously described animal model (18). Briefly, C57BL/6 mice were
subcutaneously injected in their backs with a 0.1 ml cell
suspension containing either 1×106 Hepa1–6 cells or a
mixture of 1×106 Hepa1–6 cells and 4×105
activated HSCs. Each experimental group consisted of 6 animals. The
tumour growth kinetics were monitored by measuring the length and
width of the tumour mass at the inoculation site. At the end of the
experiment, the mice were euthanised, and the tumours were
collected and stored for subsequent analysis.
Histochemistry and
immunohistochemistry
Paraffin-embedded tissue samples were serially
sectioned and immunohistochemically examined using antibodies
against PCNA (Cell Signalling) or stained using an FITC-conjugated
anti-Foxp3 antibody. Slides were visualised and photographed using
a Leica DM2500 light and fluorescence microscope.
Statistical analysis
The data were analysed using SPSS software (version
13.0). The results are expressed as the mean ± SEM. Statistical
analyses were performed using a one-way ANOVA and a Student’s
t-test. The statistical significance level was set at 0.05.
Results
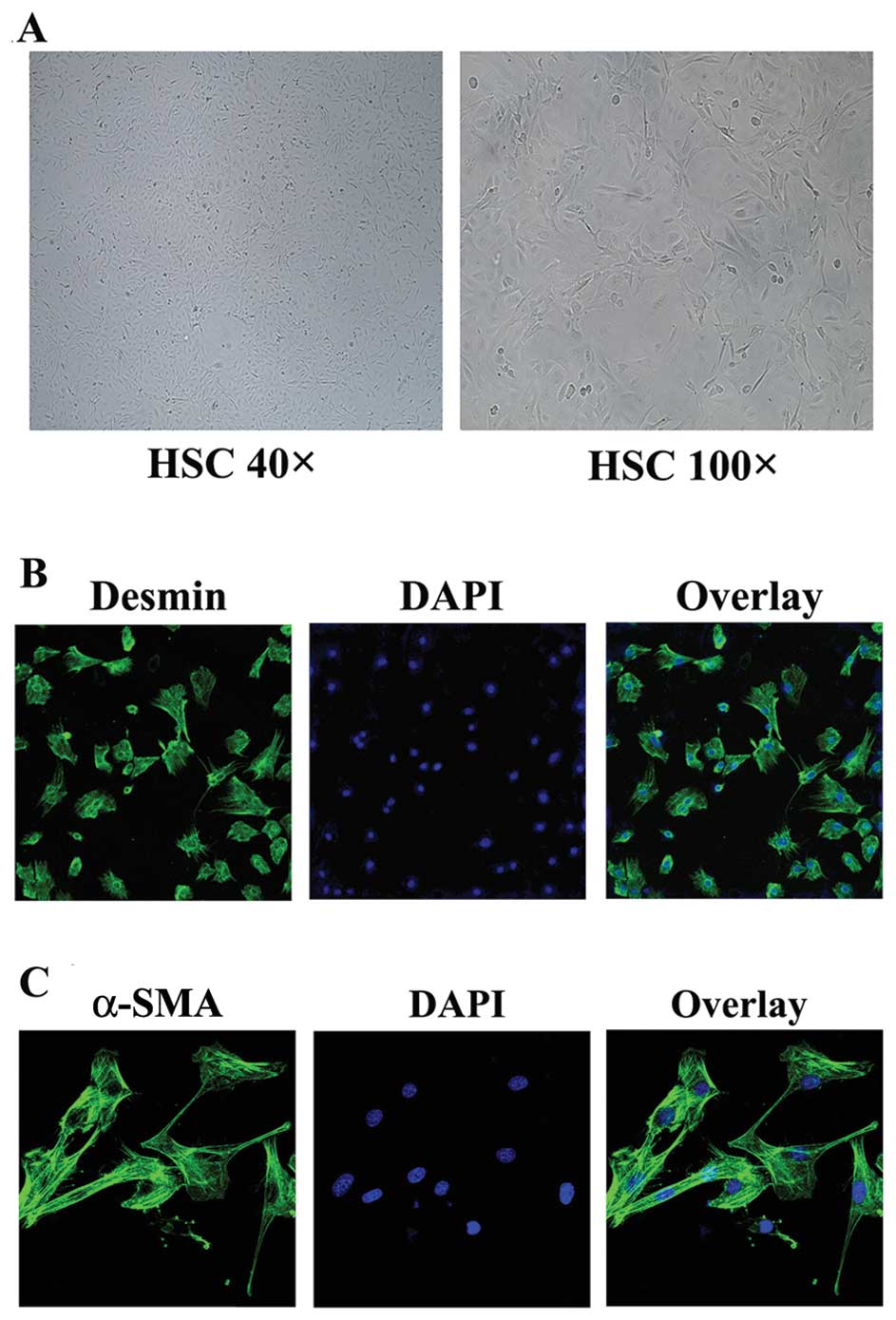
Culture and identification of HSCs
Previous studies have demonstrated that desmin, the
gold standard for identifying HSCs, is expressed in both quiescent
and activated HSCs (18); however,
α-SMA has only been detected in activated HSCs (6). Following isolation and culture, the
HSCs gradually displayed a myofibroblast-like shape (Fig. 1A) and became mature. The purity of
the HSCs was >90% based on desmin staining (Fig. 1B). After in vitro culture
for 14 days, the HSCs were activated and strongly expressed α-SMA
(Fig. 1C).
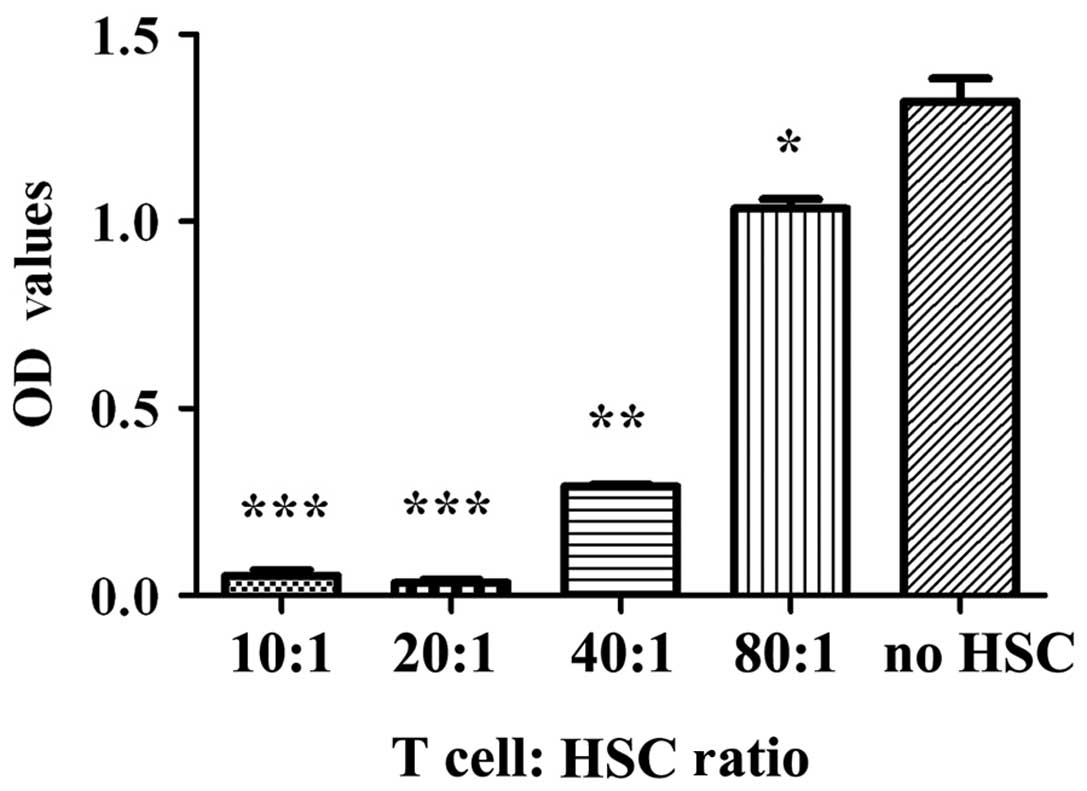
HSCs inhibit T-cell responses
To examine the effect of HSCs on the proliferation
of T cells, we used one-way MLRs. As shown in Fig. 2, the inhibition of T-cell
proliferation could be correlated with the T:HSC cell ratio in the
culture. The highest level of HSC-mediated inhibition of T-cell
proliferation was observed at a ratio of 20:1 (p<0.001).
However, this inhibition did not increase when more HSCs were added
to the culture (compare the proliferation in the 10:1 and 20:1
cultures, p>0.05).
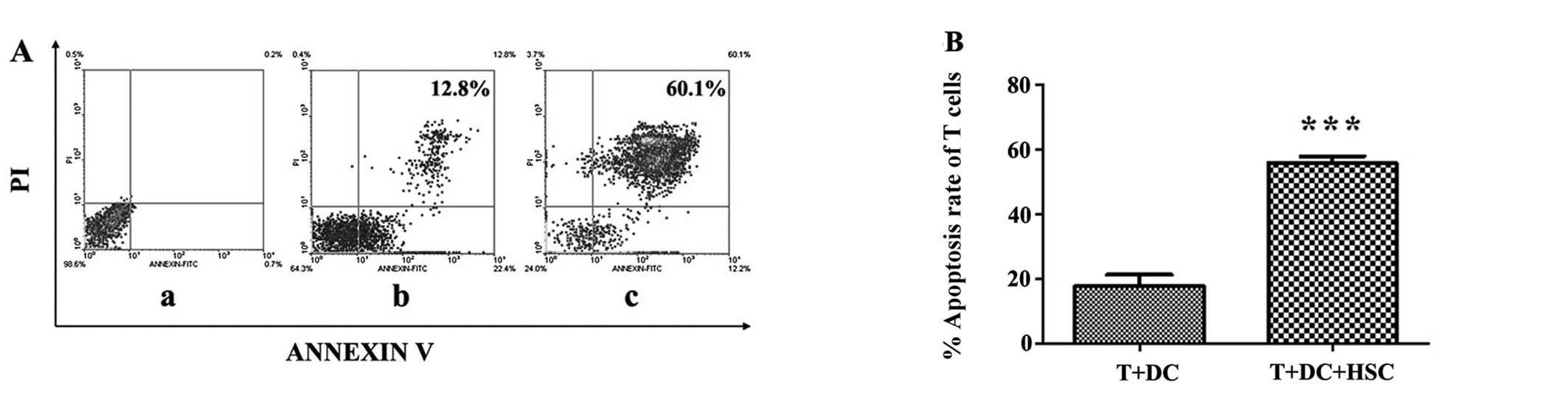
HSCs enhance T-cell apoptosis
We speculated that the HSC-induced T-cell
hyporesponsiveness may result from the apoptosis of activated T
cells. To determine the effect of HSCs on T-cell apoptosis, we
seeded T cells, DCs, and mitomycin C-treated HSCs in a 24-well
plate, and after 3 days of culture, the cells were stained using
Annexin V and PI. As shown in Fig.
3A, the proportion of cells that were double-positive for
Annexin V and PI staining increased significantly from 12.8 to
60.1% (Fig. 3B, p<0.001). These
results confirm that HSCs greatly enhance T-cell apoptosis.
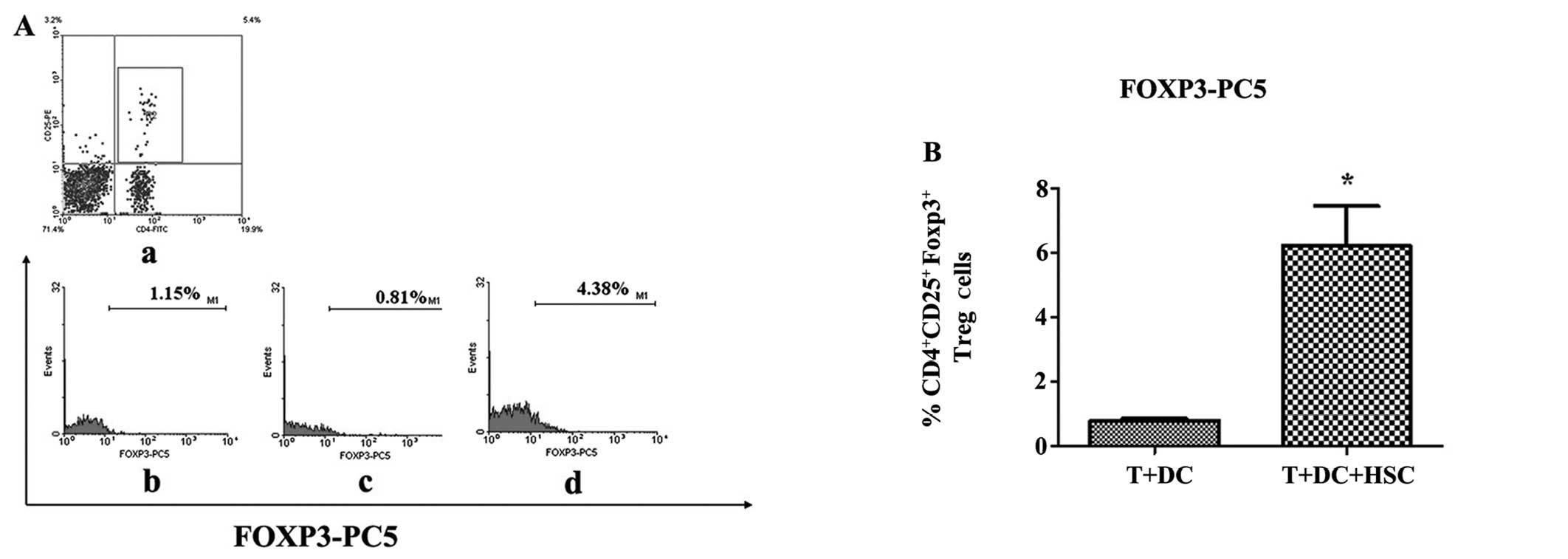
HSCs promote the expansion of Treg
cells
Treg cells suppress T-cell responses in vitro
via cell-cell contact and infectious tolerance. In vivo,
Treg cells can also create a regulatory milieu via the secretion of
IL-10 and/or TGF-β, which results in both antigen-specific and
bystander immunosuppression (19).
To determine whether the HSC-mediated immunosuppression occurred
through Treg cells, we examined Treg cells using MLRs. As shown in
Fig. 4A, the percentage of
CD4+CD25+ FoxP3+ cells in the
experimental group was higher than in the control group (4.38 vs.
0.81%, p<0.05). This result confirms that HSCs can expand the
Treg cell population.
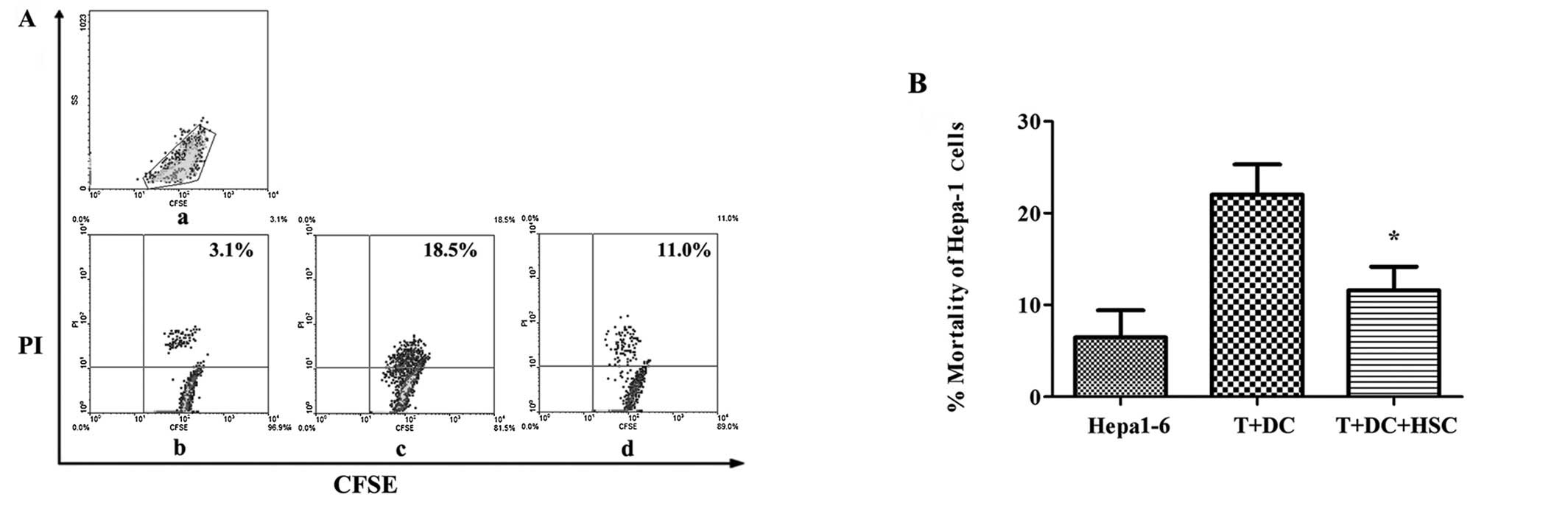
HSCs inhibit the cytotoxic T lymphocyte
response
Our data demonstrated that activated HSCs could
suppress T cell responses. However, whether they can inhibit
activated T cells remains unknown. To test this, Hepa1-6 cells were
labelled using CFSE and used as target cells. As shown in Fig. 5, after gating the CFSE+
cells, the proportion of CFSE/PI double-positive cells was lower in
the cultures containing HSCs (11.6% ± 2.6%) compared with the
no-HSCs group (22% ± 3.2%; 47% decrease; p<0.05). These results
demonstrate that activated HSCs markedly reduce the cytotoxic
activity of activated T cells.
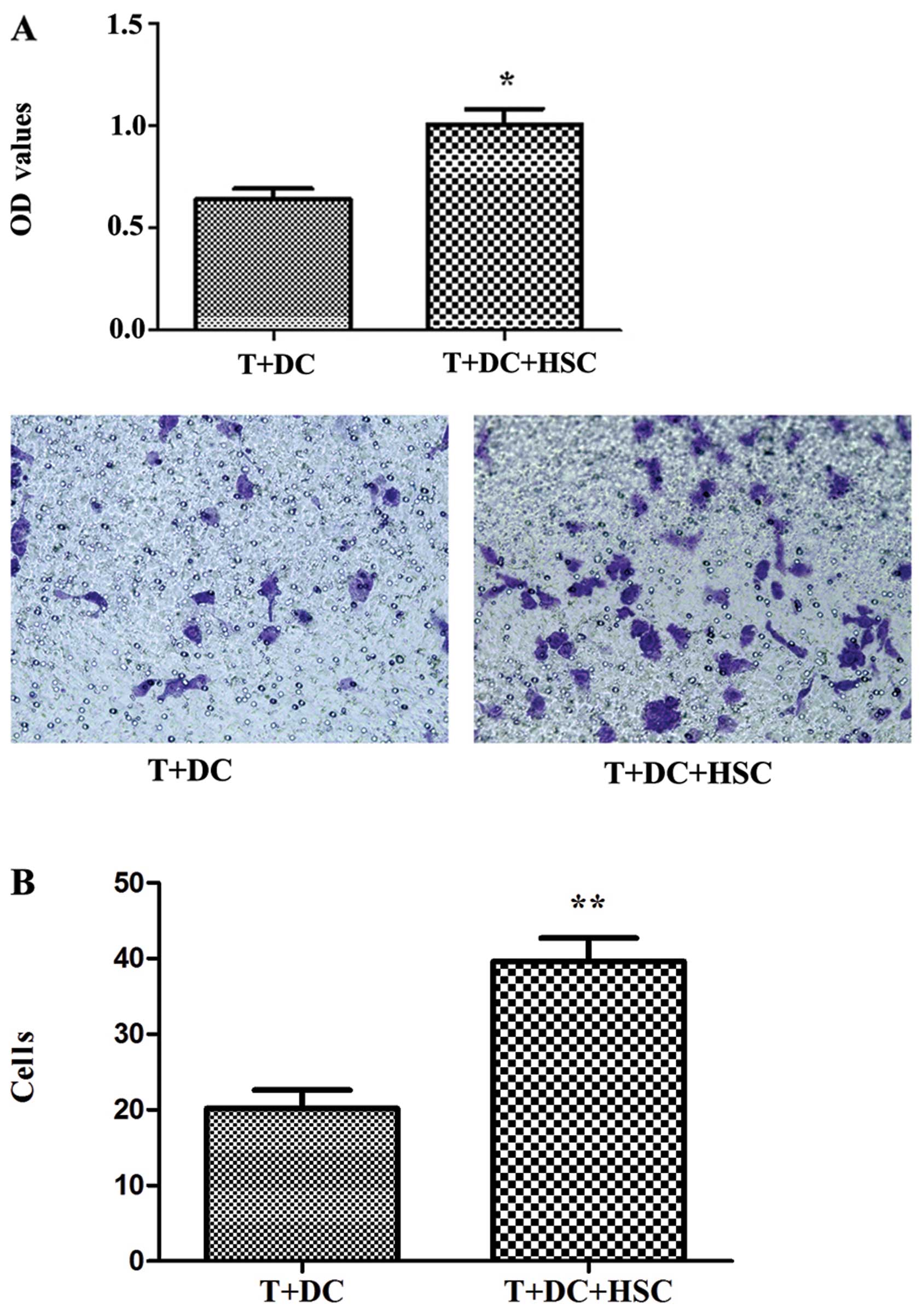
The supernatant from MLRs promotes the
proliferation and migration of tumour cells
Since the activated HSCs attenuated the function of
T cells, we evaluated the effect of supernatants from MLRs on the
proliferation and migration of tumour cells. As shown in Fig. 6A, the supernatant from MLRs
containing HSCs clearly promoted the proliferation of Hepa1–6 cells
(p<0.05). In addition, the presence of HSCs markedly increased
the migration of Hepa1–6 cells (Fig.
6B).
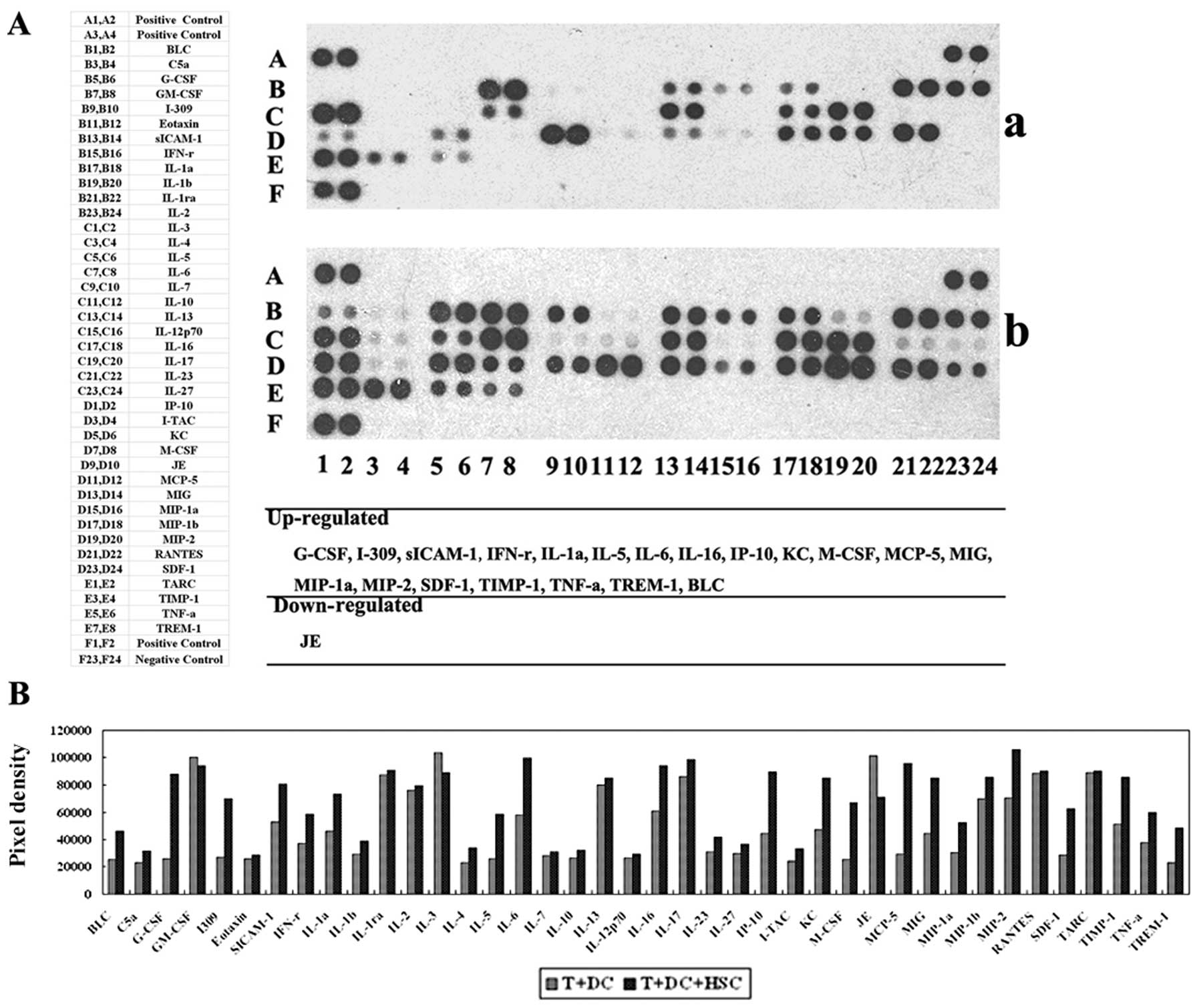
The expression of suppressive cytokines
is altered by HSCs
HSCs play a role in immunosuppression and
immunoregulation not only through their ability to inhibit T-cell
responses but also through the secretion of cytokines (19). We evaluated the cytokines in the
supernatants from MLRs using a mouse cytokine panel and detected
cytokines that induce tolerance in addition to TGF-β. As shown in
Fig. 7, the following cytokines
were expressed at higher levels in the supernatant from MLRs
containing HSCs: B-lymphocyte chemoattractant (BLC), granulocyte
colony-stimulating factor (G-CSF), soluble inter-cellular adhesion
molecule-1 (sICAM/CD54), interleukin-1α (IL-1α), interleukin-6
(IL-6), chemokine ligand-1 (CCL1/I309), interleukin-16 (IL-16),
interferon-γ (IFN-γ), monocyte chemoattractant protein-5
(MCP-5/CCL12), interferon-inducible protein (IP-10/CXCL-10),
keratinocyte chemoattractant (KC), monokine induced by IFN-γ
(MIG/CXCL-9), macrophage inflammatory protein (MIP-2/CXCL2),
MIP-1α, stromal cell-derived factor-1 (SDF-1), tissue inhibitor of
metalloproteinase-1 (TIMP-1), tumour necrosis factor α (TNF-α) and
triggering receptor expressed on myeloid cells 1 (TREM-1). This
suggests that HSCs can also influence the response and cytotoxic
capacity of T cells by increasing the expression of some cytokines
in the MLRs. These factors may be responsible for the
immunosuppressive and immunoregulatory ability of HSCs.
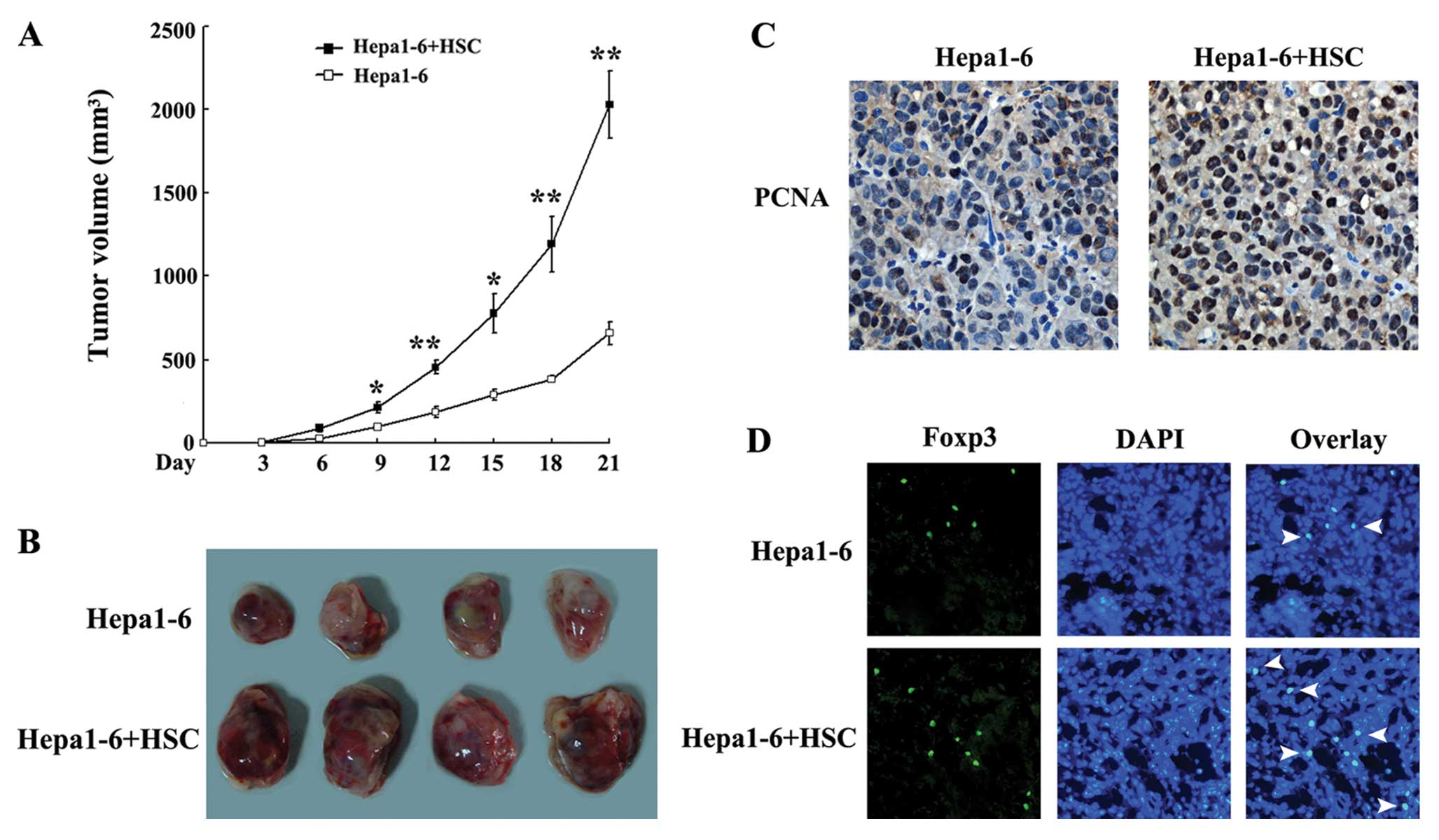
Activated HSCs promote HCC growth and
Treg cells expansion in vivo
To determine the role of HSCs in the development of
HCC, we established an in vivo model of HCC in mice via the
subcutaneous injection of Hepa1–6 cells (control group) or a
mixture of Hepa1–6 cells with activated HSCs (experimental group).
The tumours grew more rapidly and larger in the experimental group
than in the control group (Fig. 8A and
B). As HSCs promoted Hepa1–6 proliferation in vitro
(Fig. 6A), the in vivo
pro-proliferative response of HSCs was then assessed by analysing
tumour samples using PCNA immunostaining. The number of
PCNA-positive cells was significantly increased in the experimental
group compared with the control group (Fig. 8C). Thus, Treg cells play an
important role in tumour immune tolerance. As HSCs increased the
expansion of Treg cells in vitro, we assessed the effect of
HSCs on Treg cells in tumours. As shown in Fig. 8D, the number of Foxp3-positive Treg
cells in the experimental group was much higher than the number in
the control group.
Discussion
Following liver injury, HSCs undergo a
transformation from quiescent cells containing large retinoid
droplets to activated proliferating myofibroblast-like cells.
Activated HSCs can promote the development of HCC in vitro
and in vivo(14). However,
the mechanisms underlying the impact of HSCs on T cells and the
development of HCC remain unclear. We hypothesised that activated
HSCs may inhibit the ability of immune cells, especially T cells,
to kill cancer cells. Furthermore, HSCs can change the cytokine
milieu, which is involved in controlling the immune response to
HCC. To gain a better understanding of the underlying mechanism, we
utilised one-way MLRs to investigate the effect of HSCs on T
cells.
Our findings demonstrated that T-cell proliferation
was markedly inhibited in the presence of HSCs compared with
control MLRs containing no HSCs and that this effect was dependent
on the number of HSCs added. Moreover, we examined the apoptosis of
T cells in MLRs via staining with Annexin V and PI and found that
most of the T cells were Annexin V/PI double-positive in the MLRs
with HSCs. These findings are consistent with other reports
(9,11) and demonstrate that activated HSCs
suppress the proliferation of T cells and induce the apoptosis of
activated T cells. These findings could represent one mechanism
through which HSCs modulate the activity of T cells responding to
HCC.
A previous study reported that CD4+ T
cells could be converted to Treg cells through exposure to vitamin
A or TGF-β (20). Notably, HSCs
store vitamin A and secrete TGF-β in response to
inflammation-induced injury. Therefore, it has been suggested that
HSCs may exhibit tolerogenic functions in addition to their
immunosuppressive ability (21).
Our studies confirmed that the number of Treg cells
(CD4+CD25+FoxP3+) in the MLRs with
HSCs was significantly higher than in the MLRs without HSCs. In
addition, the tumours that developed in mice implanted with both
HCC cells and HSCs contained more Foxp3-positive cells. These
results indicate that HSCs induce an increase in the number of Treg
cells, which partially elicit the suppression of the immune
response of T cells against cancer cells.
In our study, activated T cells exhibited
cytotoxicity against the allogeneic target cells (Hepa1–6 cells)
after they were stimulated with DCs exposed to Hepa1–6 lysates.
However, this cytotoxicity was blocked by the HSCs. This important
finding confirms that activated HSCs attenuate the cytotoxicity of
T cells against cancer cells.
The role of activated HSCs in the development of HCC
is not only to directly affect T cell function but also to change
the expression of cytokines. We analysed the supernatant from the
MLRs and found increased expression of a number of cytokines in
MLRs containing HSCs, such as IL-6, G-CSF, sICAM-1, SDF-1, IFN-γ
and TNF-α. IL-6 is a proinflammatory factor that plays a critical
role in the natural history of some malignancies, such as human
plasma cell neoplasms, colon cancer, and HCC. IL-6 can mediate
autoimmune disease and tumour growth through the IL-6/STAT-3
signalling pathway (22–24).
G-CSF and GM-CSF and their receptors are
constitutively expressed in numerous solid tumours, such as skin
and head and neck squamous cell carcinomas, gliomas, and
meningiomas. Moreover, G-CSF and GM-CSF have previously been shown
to stimulate tumour cell growth and migration in
vitro(25,26).
As a member of the immunoglobulin superfamily, it
has been reported that sICAM-1 is immunosuppressive and local
release of ICAM-1 appears to promote local immune tolerance and
cancer cell immune escape (21,27).
ICAM-1 was constitutively expressed on HSCs and can be induced by
TNF-α and IFN-γ. ICAM-1 deficient HSCs can partially reverse HSCs
immune inhibitory activity both in vitro and in
vivo(9,28). As shown in Fig. 7, higher IFN-γ and TNF-α expression
were found in supernatant from MLRs containing the HSCs group.
Usually, IFN-γ is a positive regulator in immune reactions.
However, after being stimulated by IFN-γ, HSCs become activated,
upregulate inhibitory surface molecules B7H1, expand the population
of Treg cells, and exhibit profound immunosuppressive activity
against the adaptive immune response (11,29).
TNF-α is a multifunctional cytokine involved in
apoptosis and cell survival as well as in inflammation and immunity
(30,31). Although recognized for its
antitumor activity, studies in the pathogenesis of many
neurological conditions have demonstrated that TNF-α has
immunosuppressive functions during the chronic phase of the
disease, suggesting a dual role for TNF-α (32). As a pleiotropic chemokine, SDF-1
has recently been reported to participate in inducing immunological
tolerance (33). Although some of
the cytokines detected in our study have proinflammatory roles,
based on our results, the inhibitory action of these molecules
overrides their proinflammatory functions, resulting in T-cell
hyporesponsiveness and the metastasis of cancer cells.
In summary, our study demonstrates that activated
HSCs can induce the death of activated T cells, modify the type of
T cells present (expanding Treg cells), and can reduce the
cytotoxicity of cancer-specific T cells. HSCs can also affect the
cytokines present in MLRs, resulting in the increased proliferation
and migration of cancer cells. Additionally, consistent with the
in vitro results, activated HSCs can induce HCC
proliferation and Treg cell expansion in vivo. These results
demonstrate that HSCs inhibit the activity of T cells and promote
the immune escape of HCC.
Acknowledgements
The authors thank Ms. Lili Liu and Mr.
Zhigang Liu for their help with flow cytometry, Ms. Yuehong Ma for
her help with the immunofluorescence and Mr. Yongzhi Wang for the
technical assistance with the MLRs. This study was supported by
grants from the National Key Sci-Tech Special Project of China (no.
2012ZX10002011-005) and the National Natural Science Foundation of
China (no. 81171967).
Reference
|
1
|
Kim WR and Kremers WK: Benefits of ‘the
benefit model’ in liver transplantation. Hepatology. 48:697–698.
2008.
|
|
2
|
Gressner AM, Weiskirchen R, Breitkopf K
and Dooley S: Roles of TGF-beta in hepatic fibrosis. Front Biosci.
7:d793–d807. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Friedman SL: Liver fibrosis - from bench
to bedside. J Hepatol. 38:S38–S53. 2003. View Article : Google Scholar
|
|
4
|
Friedman SL: Mechanisms of hepatic
fibrogenesis. Gastroenterology. 134:1655–1669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bomble M, Tacke F, Rink L, Kovalenko E and
Weiskirchen R: Analysis of antigen-presenting functionality of
cultured rat hepatic stellate cells and transdifferentiated
myofibroblasts. Biochem Biophys Res Commun. 396:342–347. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Friedman SL: Hepatic stellate cells:
protean, multifunctional, and enigmatic cells of the liver. Physiol
Rev. 88:125–172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Benseler V, McCaughan GW, Schlitt HJ,
Bishop GA, Bowen DG and Bertolino P: The liver: a special case in
transplantation tolerance. Semin Liver Dis. 27:194–213. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen CH, Kuo LM, Chang Y, et al: In vivo
immune modulatory activity of hepatic stellate cells in mice.
Hepatology. 44:1171–1181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yin Z, Jiang G, Fung JJ, Lu L and Qian S:
ICAM-1 expressed on hepatic stellate cells plays an important role
in immune regulation. Microsurgery. 27:328–332. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen CH, Shu KH, Su YH, et al:
Cotransplantation of hepatic stellate cells attenuates the severity
of graft-versus-host disease. Transplant Proc. 42:971–975. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu MC, Chen CH, Liang X, et al: Inhibition
of T-cell responses by hepatic stellate cells via B7-H1-mediated
T-cell apoptosis in mice. Hepatology. 40:1312–1321. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amann T, Bataille F, Spruss T, et al:
Activated hepatic stellate cells promote tumorigenicity of
hepatocellular carcinoma. Cancer Sci. 100:646–653. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mikula M, Proell V, Fischer AN and
Mikulits W: Activated hepatic stellate cells induce tumor
progression of neoplastic hepatocytes in a TGF-beta dependent
fashion. J Cell Physiol. 209:560–567. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao W, Zhang L, Yin Z, et al: Activated
hepatic stellate cells promote hepatocellular carcinoma development
in immunocompetent mice. Int J Cancer. 129:2651–2661. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Friedman SL: Seminars in medicine of the
Beth Israel Hospital, Boston. The cellular basis of hepatic
fibrosis. Mechanisms and treatment strategies. N Engl J Med.
328:1828–1835. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiang J, Gu X, Qian S and Chen Z: Graded
function of CD80 and CD86 in initiation of T-cell immune response
and cardiac allograft survival. Transpl Int. 21:163–168.
2008.PubMed/NCBI
|
|
17
|
Lee WC, Wang HC, Jeng LB, et al: Effective
treatment of small murine hepatocellular carcinoma by dendritic
cells. Hepatology. 34:896–905. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yokoi Y, Namihisa T, Kuroda H, et al:
Immunocytochemical detection of desmin in fat-storing cells (Ito
cells). Hepatology. 4:709–714. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carambia A and Herkel J: CD4 T cells in
hepatic immune tolerance. J Autoimmun. 34:23–28. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Strober W: Vitamin A rewrites the ABCs of
oral tolerance. Mucosal Immunol. 1:92–95. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tiegs G and Lohse AW: Immune tolerance:
what is unique about the liver. J Autoimmun. 34:1–6. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rutsch S, Neppalli VT, Shin DM, et al:
IL-6 and MYC collaborate in plasma cell tumor formation in mice.
Blood. 115:1746–1754. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Knupfer H and Preiss R: Serum
interleukin-6 levels in colorectal cancer patients – a summary of
published results. Int J Colorectal Dis. 25:135–140. 2010.
|
|
24
|
Chau GY, Wu CW, Lui WY, et al: Serum
interleukin-10 but not interleukin-6 is related to clinical outcome
in patients with resectable hepatocellular carcinoma. Ann Surg.
231:552–558. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Obermueller E, Vosseler S, Fusenig NE and
Mueller MM: Cooperative autocrine and paracrine functions of
granulocyte colony-stimulating factor and granulocyte-macrophage
colony-stimulating factor in the progression of skin carcinoma
cells. Cancer Res. 64:7801–7812. 2004. View Article : Google Scholar
|
|
26
|
Gutschalk CM, Herold-Mende CC, Fusenig NE
and Mueller MM: Granulocyte colony-stimulating factor and
granulocyte-macrophage colony-stimulating factor promote malignant
growth of cells from head and neck squamous cell carcinomas in
vivo. Cancer Res. 66:8026–8036. 2006. View Article : Google Scholar
|
|
27
|
Wang HW, Babic AM, Mitchell HA, Liu K and
Wagner DD: Elevated soluble ICAM-1 levels induce immune deficiency
and increase adiposity in mice. FASEB J. 19:1018–1020.
2005.PubMed/NCBI
|
|
28
|
Knittel T, Dinter C, Kobold D, et al:
Expression and regulation of cell adhesion molecules by hepatic
stellate cells (HSC) of rat liver: involvement of HSC in
recruitment of inflammatory cells during hepatic tissue repair. Am
J Pathol. 154:153–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang HR, Chou HS, Gu X, et al: Mechanistic
insights into immunomodulation by hepatic stellate cells in mice: a
critical role of interferon-gamma signaling. Hepatology.
50:1981–1991. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bazzoni F and Beutler B: The tumor
necrosis factor ligand and receptor families. N Engl J Med.
334:1717–1725. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Locksley RM, Killeen N and Lenardo MJ: The
TNF and TNF receptor superfamilies: integrating mammalian biology.
Cell. 104:487–501. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Correale J and Villa A: The
neuroprotective role of inflammation in nervous system injuries. J
Neurol. 251:1304–1316. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Karin N: The multiple faces of CXCL12
(SDF-1alpha) in the regulation of immunity during health and
disease. J Leukoc Biol. 88:463–473. 2010. View Article : Google Scholar : PubMed/NCBI
|