Introduction
Pancreatic cancer is the fourth most common cause of
cancer-related mortality in the Western world. In the United
States, approximately 36,800 people died from pancreatic cancer in
2010 (1). The overall prognosis
for patients with pancreatic cancer remains poor: the five-year
survival rate is less than 5% (2).
A number of studies have evaluated various chemotherapeutic agents;
however, only a few have produced results showing a significant
improvement in survival (3).
Gemcitabine (GEM), a pyrimidine-based
anti-metabolite, remains the first-line standard drug for the
treatment of advanced human pancreatic cancer either alone or in
combination with other chemotherapeutic agents (4–6). GEM
is metabolized to gemcitabine triphosphate (dFdCTP) in cells and is
incorporated into the DNA resulting in the induction of apoptosis
due to the inhibition of DNA synthesis (7). However, the intrinsic and/or acquired
resistance of pancreatic cancer to GEM presents a major challenge.
Tumor cells acquire resistance to GEM by various mechanisms,
including alterations in transport, drug targets and metabolism or
in the genes regulating cell survival (8). The discovery of cancer stem cells
(CSCs) as cancer-initiating components in leukemia and solid tumors
has presented an attractive approach for treatment (9). Previous studies have shown that
pancreatic CSCs are enriched in GEM-resistant cells (10,11).
Most standard chemotherapy treatments destroy most of the tumor
population; however, CSCs, which have intrinsic drug detoxifying
and resistant mechanisms, can easily escape standard chemotherapy
treatments.
Aldehyde dehydrogenase (ALDH) is a family of
intra-cellular enzymes that participate in cellular detoxification,
differentiation and drug resistance through the oxidation of
cellular aldehydes (12). One of
17 ALDH isoforms, ALDH1A1 is a detoxifying enzyme responsible for
oxidizing aldehydes to carboxylic acids and converting retinol to
retinoic acid. It also holds the distinction of being a potential
marker of CSCs and potentially playing a role in the biology of
tumor-initiating cells (13–16).
Tumor-initiating cell-enriched populations have been identified in
multiple malignancies: breast, colon, pancreatic, lung, liver and
ovarian cancer by using the AldeFluor assay, a functional flow
cytometric assay that identifies cells with active ALDH1A1
(17–26).
In the present study, we performed experiments to
determine whether the direct targeting of ALDH1A1 by small
interfering RNA (siRNA) enhances the chemosensitivity of pancreatic
cancer cells to GEM. Our results suggest that ALDH1A1 is a
potentially important therapeutic target for human pancreatic
ductal carcinoma cells.
Materials and methods
Cell culture and reagents
MIA PaCa-2, Panc-1, CFPAC-1 and BxPC-3 cells were
purchased from the American Type Culture Collection (ATCC;
Manassas, VA), and AsPC-1 and Colo-357 cells were obtained from the
Tissue Culture Shared Resource of Georgetown University Lombardi
Comprehensive Cancer Center (Washington, DC). The human pancreatic
ductal epithelial cell line, HPDE6-C7, was acquired from Dr M.S.
Tsao (27). AsPC-1, BxPC-3 and
Colo-357 cells were cultured in RPMI-1640 medium supplemented with
fetal bovine serum (FBS; 20% for AsPC-1, 10% for Colo-357 and
BxPC-3 cells), 100 U/ml penicillin, 100 μg/ml streptomycin
and 1% sodium pyruvate. MIA PaCa-2 cells were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) containing 10% FBS, 2.5%
horse serum (HS), 100 U/ml penicillin, and 100 μg/ml
streptomycin. Panc-1 and CFPAC-1 cells were cultured in DMEM
containing 10% FBS, 10 U/ml penicillin, and 10 μg/ml
streptomycin. HPDE6-C7 cells were cultured in keratinocyte
serum-free (KSF) medium supplemented by an epidermal growth factor
and bovine pituitary extract and 1X antibiotic-antimycotic. Cell
culture reagents were purchased from BioWhittaker (Walkersville,
MD) and Invitrogen (Carlsbad, CA). GEM was obtained from Sigma (St.
Louis, MO).
Generation of GEM-resistant MIA PaCa-2
cells (MIA PaCa-2/GR cells)
The well-characterized pancreatic adenocarcinoma
cell line, MIA PaCa-2 was used as the parental line (MIA PaCa-2/P)
from which the GEM-resistant cell line was developed. The MIA
PaCa-2/P cells were serially subcultured through incrementally
increasing GEM concentrations, starting with 0.1 μM for six
months. MIA PaCa-2/GR cells retained the capacity for proliferation
when returned to medium containing GEM (0.5 μM).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
A total of either 2,000 MIA PaCa-2/P or MIA
PaCa-2/GR cells were plated in 96-well flat bottom plates and then
exposed to various concentrations of chemotherapeutic agents. At
the indicated times, 10 μl of 1 mg/ml MTT (Sigma) in
phosphate-buffered saline (PBS) were added to each well for 4 h.
After centrifugation and removal of the medium, 150 μl of
dimethylsulphoxide (DMSO) (Sigma) were added to each well to
dissolve the formazan crystals. The absorbance was measured at 560
nm using an ELx808 Absorbance Microplate Reader (BioTek
Instruments, Inc., Winooski, VT). The absorbance of untreated cells
was designated at 100% and cell survival was expressed as a
percentage of this value. Triplicate wells were assayed for each
condition and standard deviation (SD) was determined.
Western blot (WB) analysis
Cells were grown to ∼70% confluence and reagents
were added at the indicated concentrations. After exposure to
control-siRNA or ALDH1A1-siRNA with GEM, the cells were lysed in
cell lysis buffer containing 20 mM Tris-HCl, 0.5 M NaCl, 0.25%
Triton X-100, 1 mM EDTA, 1 mM EGTA, 10 mM β-glycerophosphate, 10 mM
NaF, 300 μM Na3VO4, 1 mM benzamidine,
2 μM PMSF, and 1 mM DTT. The protein concentration was
determined by a BCA protein assay kit (Thermo Scientific, Rockford,
IL). Proteins were separated on SDS-PAGE, transferred onto a PVDF
membrane, blocked in 1X blocking buffer (Sigma) and probed with the
following primary antibodies: ALDH1A1 (Abcam, Cambridge, UK),
poly(ADP-ribose) polymerase (PARP; BD Biosciences, Franklin, NJ),
and α-tubulin (Sigma). The membranes were then incubated with
horseradish peroxidase (HRP)-conjugated secondary antibodies
(Sigma) and visualized with a chemiluminescence kit (Santa Cruz
Biotechnology, Santa Cruz, CA) according to the manufacturer’s
recommended protocol and exposed to X-ray film (American X-ray
& Medical Supply, Jackson, CA).
Flow cytometry
MIA PaCa-2 cells were collected after transfection
and treatment with GEM by trypsinization, washed with PBS and fixed
overnight in 70% ethanol at −20°C. The cells were then incubated
with 20 μg/ml propidium iodide and 40 μg/ml RNase A
in 1X PBS. The cells were analyzed on a FACSCalibur flow cytometer
(Becton-Dickinson, San Jose, CA) at the Flow Cytometry and Cell
Sorting Shared Resource, Lombardi Comprehensive Cancer Center,
Georgetown University. The acquired data were analyzed by CellQuest
Pro Analysis software (Becton-Dickinson).
Caspase-3 activity assay
Caspase-3 activity assay was carried out by using a
caspase-3/CPP32 colorimetric assay kit (BioVision, Mountain View,
CA) according to the manufacturer’s instructions. MIA PaCa-2 cells
transfected with siRNAs were treated with GEM for 48 h.
Approximately 100 μg of protein were incubated with 200
μM Asp-Glu-Val-Asp-p-nitroanilide (DEVD-pNA) and 10 mM DTT
at 37°C for 2 h. Absorbance was measured at 405 nm using an ELx808
Absorbance Microplate Reader (BioTekInstruments, Inc.). Increased
CPP32 activity was determined by calculating these results
according to the percentage of un-induced control samples.
AldeFluor assay
Active ALDH1A1 was identified with the AldeFluor
assay according to manufacturer’s instructions (StemCell
Technologies, Durham, NC). The ALDH1A1-positive population was
defined by cells with an increased FITC signal, with gates
determined by diethylaminobenzaldehyde (DEAB)-treated cells (DEAB
being an inhibitor of ALDH1A1 activity). The AldeFluor-positive
cell population was measured by a FACSCalibur flow cytometer
(Becton-Dickinson) and analyzed as described above.
siRNAs
For the purpose of the RNA interference experiments,
ALDH1A1-siRNA-1, 5′-GAACAGUGUGGGUGAAUUG-3′; ALDH1A1-siRNA-2,
5′-AGAGUACGGUUUCCAUGAA-3′; and control-siRNA,
5′-GACGAGCGGCACGUGCACA-3′, were purchased from Dharmacon Inc.
(Lafayette, CO). The ALDH1A1-siRNA-1, ALDH1A1-siRNA-2 or
control-siRNA were subsequently transfected into the MIA PaCa-2/P
and MIA PaCa-2/GR cells using Lipofectamine™ 2000 (Invitrogen)
according to the instructions of the manufacturer. The transfected
cells were then processed for cell cycle analysis, WB analysis,
caspase-3 activity, AldeFluor activity and cell proliferation.
Trypan blue exclusion assay
Cells were collected after trypsinization of the
cell monolayer, resuspended in serum-containing medium, stained
with trypan blue and counted. Cell viability was evaluated via the
trypan blue exclusion test using the Luna Cell Counter (Logos
Biosystems, Gyunggi-do, Republic of Korea).
Statistical methods
Statistical comparisons were made using the
two-tailed Student’s t-test where appropriate. In all the
experiments, the values, P<0.05, P<0.01 and P<0.001, were
considered to indicate statistically significant differences. Data
are expressed as the means ± SD.
Results
ALDH1A1 is differentially expressed in
human pancreatic cancer cell lines and an immortal human pancreatic
duct epithelial cell line
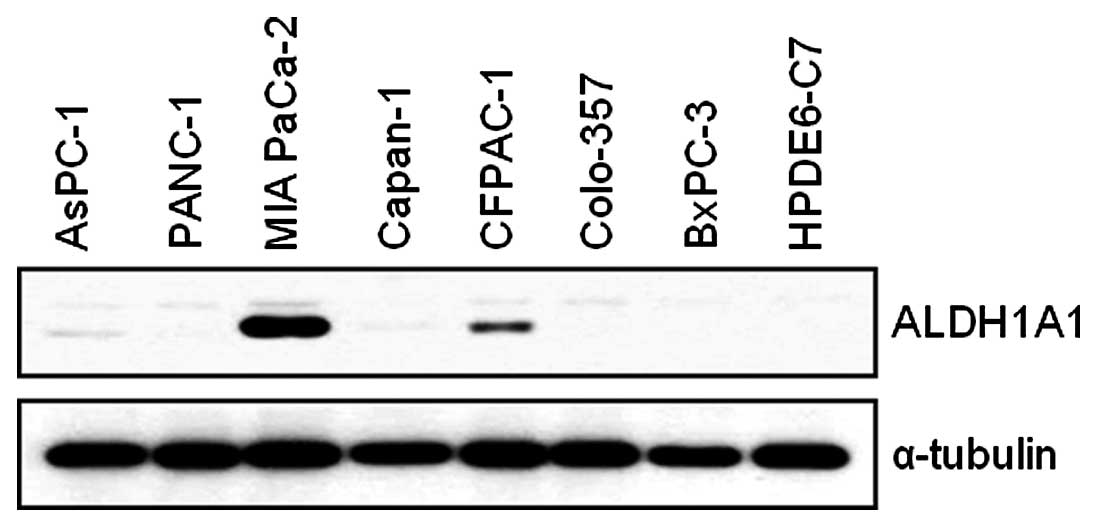
We first assessed the basal expression level of
ALDH1A1 in AsPC-1, Panc-1, MIA PaCa-2, Capan-1, CFPAC-1, Colo-357,
BxPC-3 and HPDE6-C7 cells. MIA PaCa-2 and CFPAC-1 cells expressed
higher levels of ALDH1A1 than the other cell lines (Fig. 1). As we were interested in the
potential contribution of ALDH1A1 to GEM-induced cytotoxicity, we
performed further studies with ALDH1A1-positive MIA PaCa-2
cells.
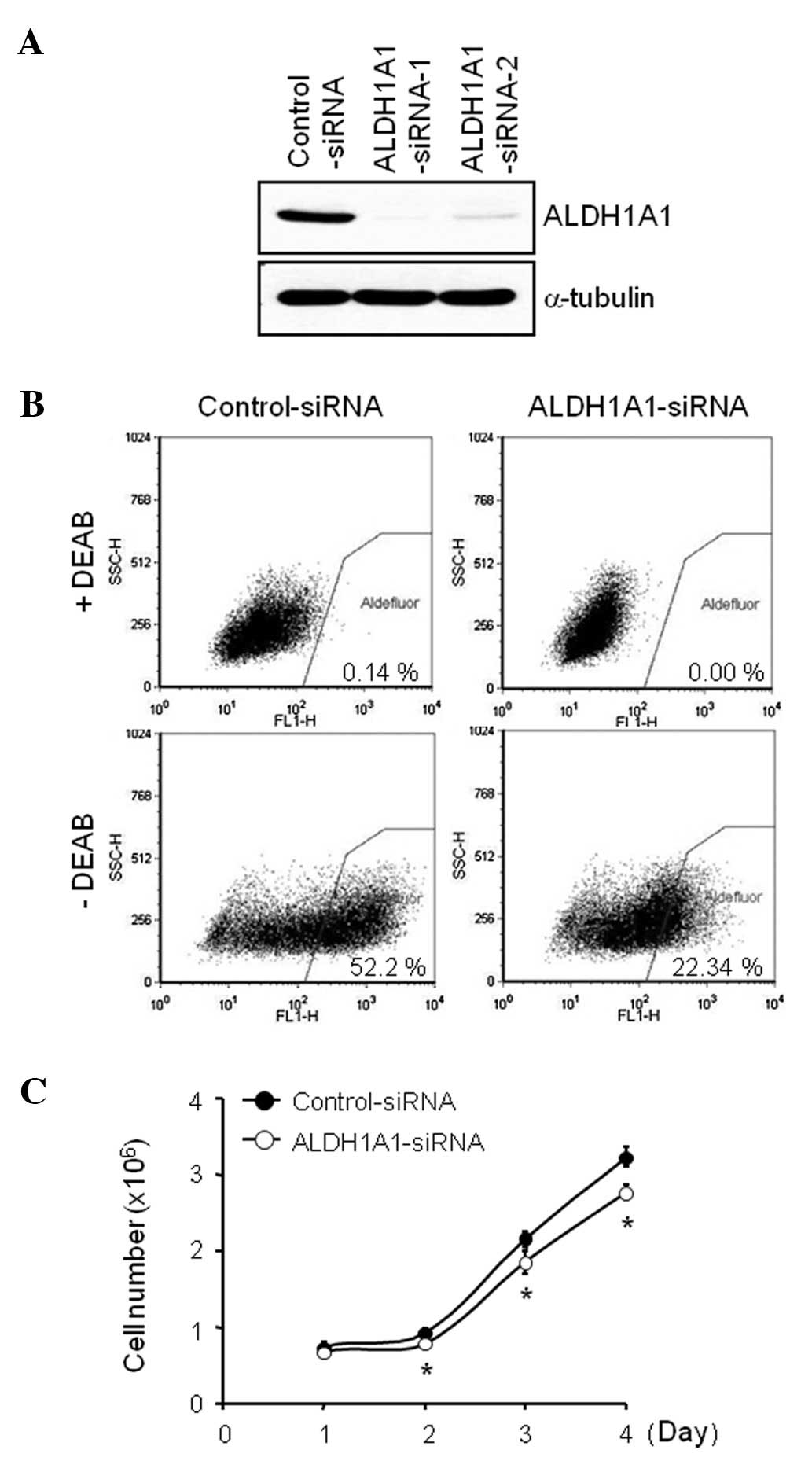
ALDH1A1 knockdown affects expression and
activity of ALDH1A1 and cell proliferation
In order to investigate the significance of
endogenous ALDH1A1 on GEM-induced cytotoxicity, we used the
siRNA-based knockdown system. ALDH1A1-specific-siRNAs, targeting
two different regions of ALDH1A1 sequences were designed and
tested. Our transfection experiments with 100 nM of siRNA for 72 h
demonstrated that ALDH1A1-siRNA-1 was more effective in reducing
ALDH1A1 expression level than ALDH1A1-siRNA-2 (Fig. 2A). We then performed an AldeFluor
assay, a functional flow cytometric assay that identifies cells
with active ALDH1A1, to determine the effect of ALDH1A1-siRNA on
ALDH1A1 activity. ALDH1A1 knockdown markedly reduced the
AldeFluor-positive cell population from 52.2% by control-siRNA to
22.3% by ALDH1A1-siRNA at 72 h after administration (Fig. 2B). Moreover, the ALDH1A1 knockdown
inhibited cell proliferation in a time-dependent manner (0, 1, 2, 3
and/or 4 days), compared to the control-siRNA (Fig. 2C). The overall number of attached
cells was decreased by the ALDH1A1-siRNA knockdown (data not
shown), and the loss of cell proliferation was observed from day
one post-transfection (Fig.
2C).
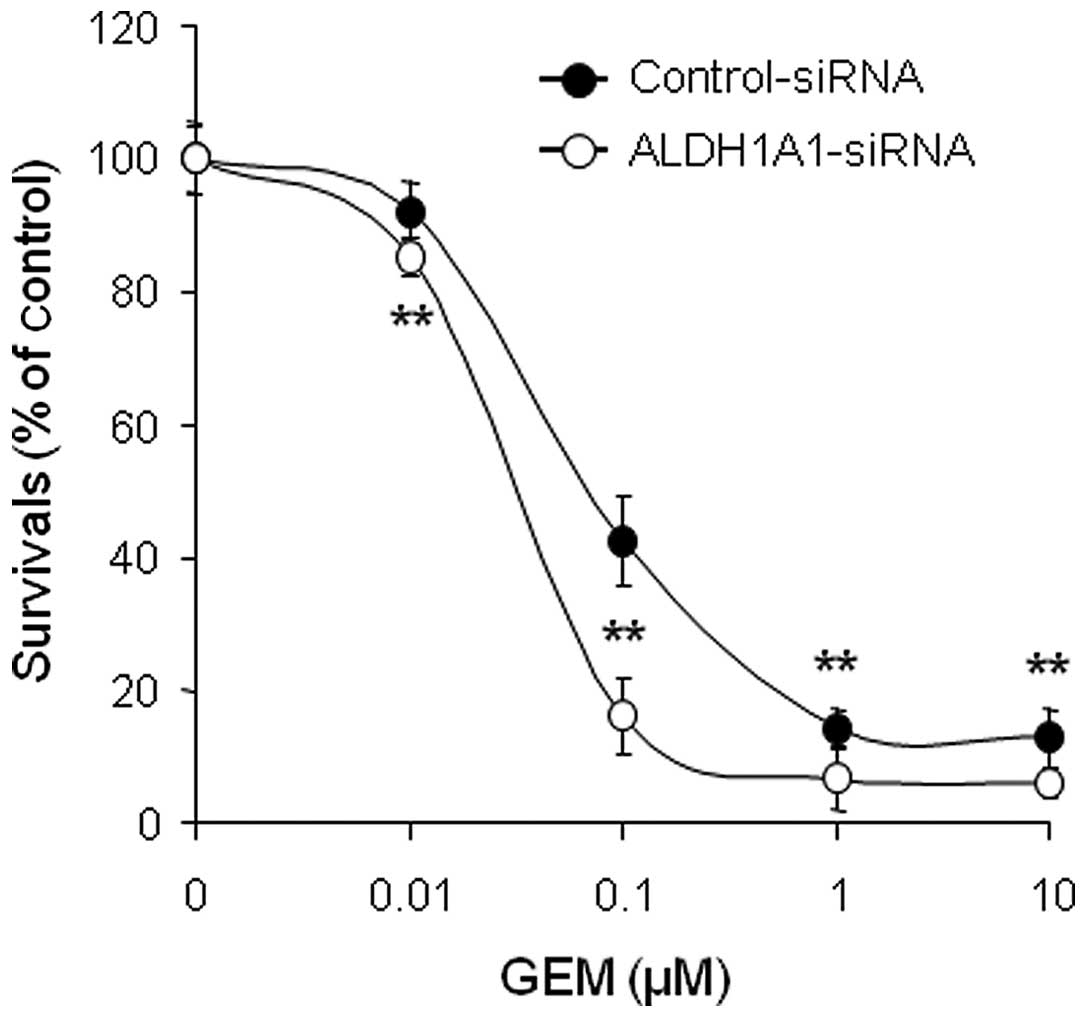
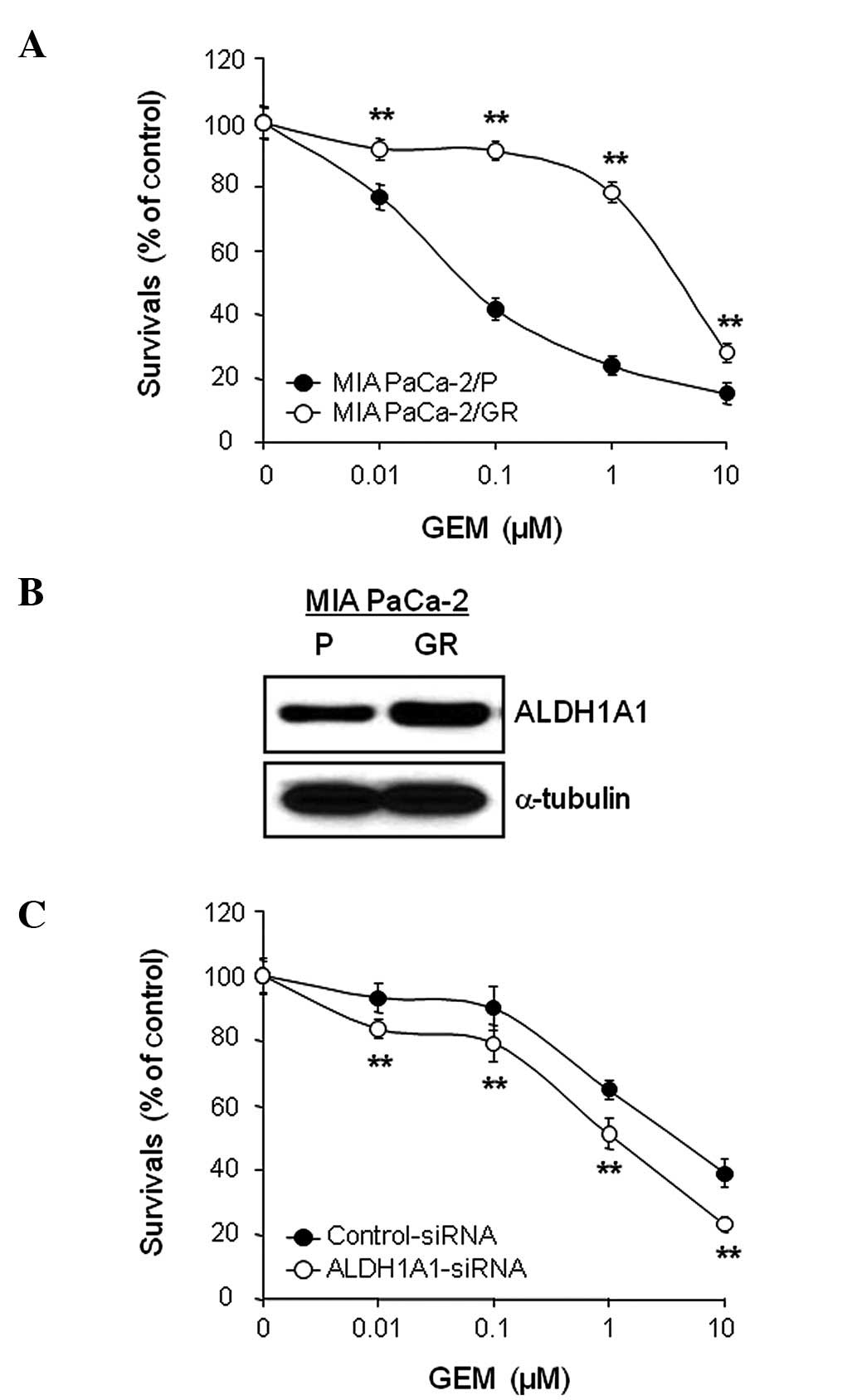
ALDH1A1 knockdown enhances cytotoxicity
and apoptotic cell death induced by GEM
In order to investigate whether controlling ALDH1A1
expression levels can alter sensitivity to GEM, MIA PaCa-2 cells
pre-treated with ALDH1A1- or control-siRNA for 48 h were incubated
with various concentrations of GEM (0, 0.01, 0.1, 1 and/or 10
μM) for 72 h and cell viability was determined by MTT assay.
The results showed that the combination of ALDH1A1-siRNA plus GEM
was significantly more effective at reducing survival than
control-siRNA plus GEM (Fig. 3).
To further evaluate the interaction between ALDH1A1-siRNA and GEM,
we determined the half maximal inhibitory concentration
(IC50) and found strong synergistic anti-tumor effects.
The IC50 of GEM decreased from 0.16 μM in the
control-siRNA-treated cells to 0.035 μM in the
ALDH1A1-siRNA-treated cells.
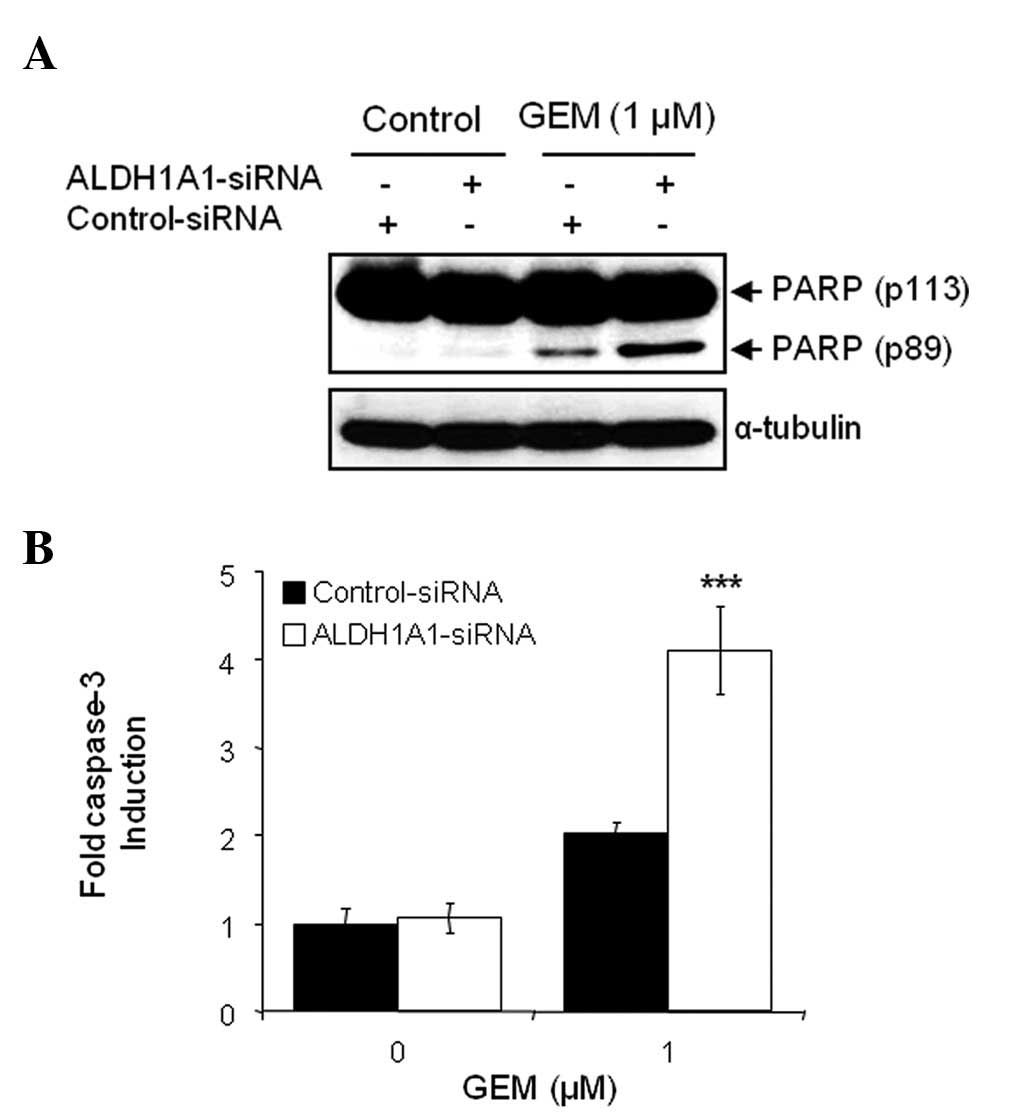
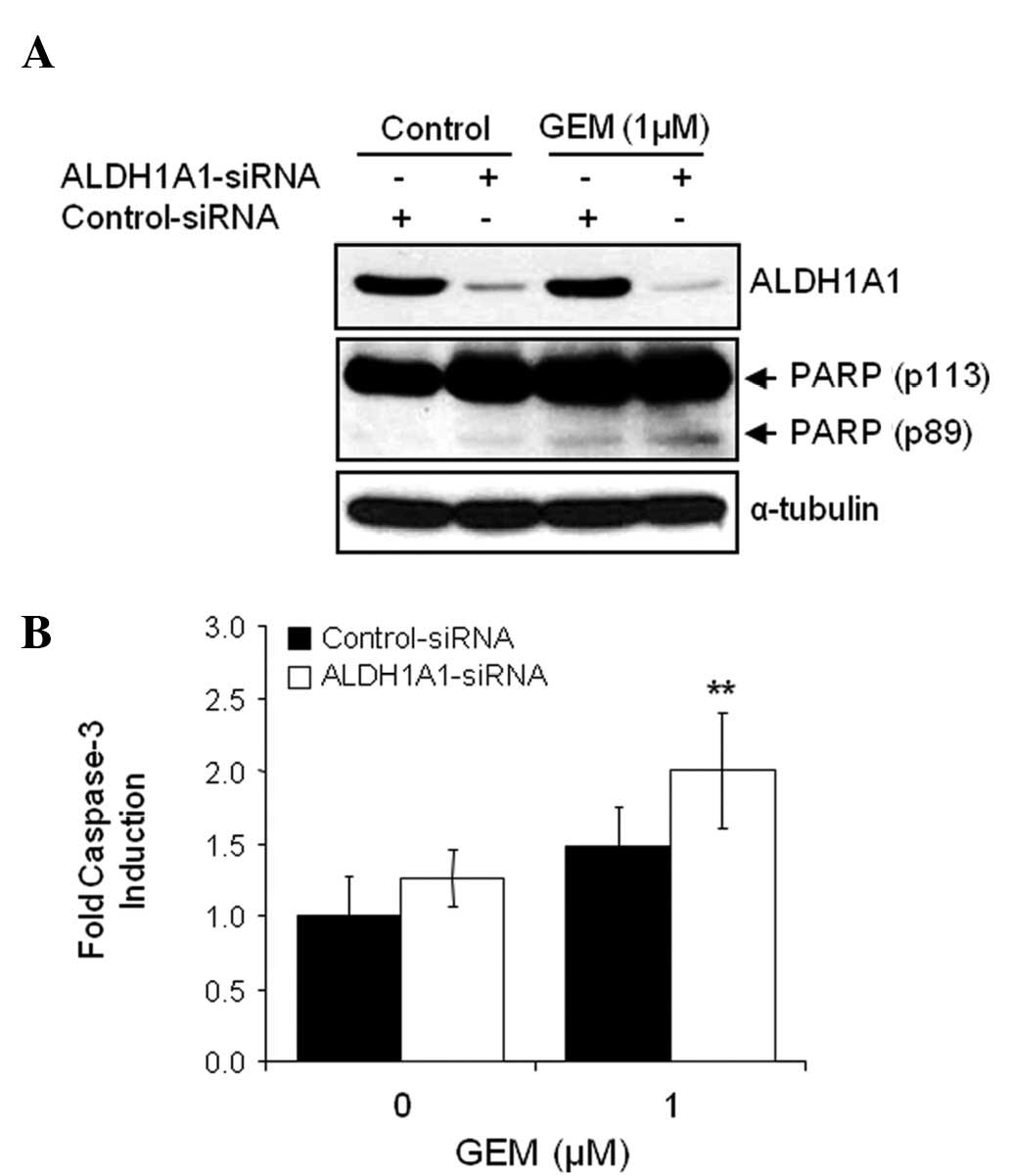
The combination of ALDH1A1-siRNA and GEM on the
induction of apoptosis was also investigated. Cells pre-treated
with siRNA for 48 h were exposed to 1 μM GEM for 48 h.
Apoptotic cell death was detected by WB analysis with the molecular
biomarker of apoptosis, PARP cleavage. When comparing treatment
with control-siRNA alone, control-siRNA plus GEM, ALDH1A1-siRNA
alone or ALDH1A1-siRNA plus GEM, the latter treatment led to a more
dramatic increase in cleaved PARP (Fig. 4A). We also performed a caspase-3
activity assay to confirm the effects of the ALDH1A1 knockdown on
GEM-mediated apoptotic cell death. As expected, the combination of
ALDH1A1-siRNA plus GEM most significantly increased caspase-3
activity (Fig. 4B).
ALDH1A1 knockdown enhances induction of
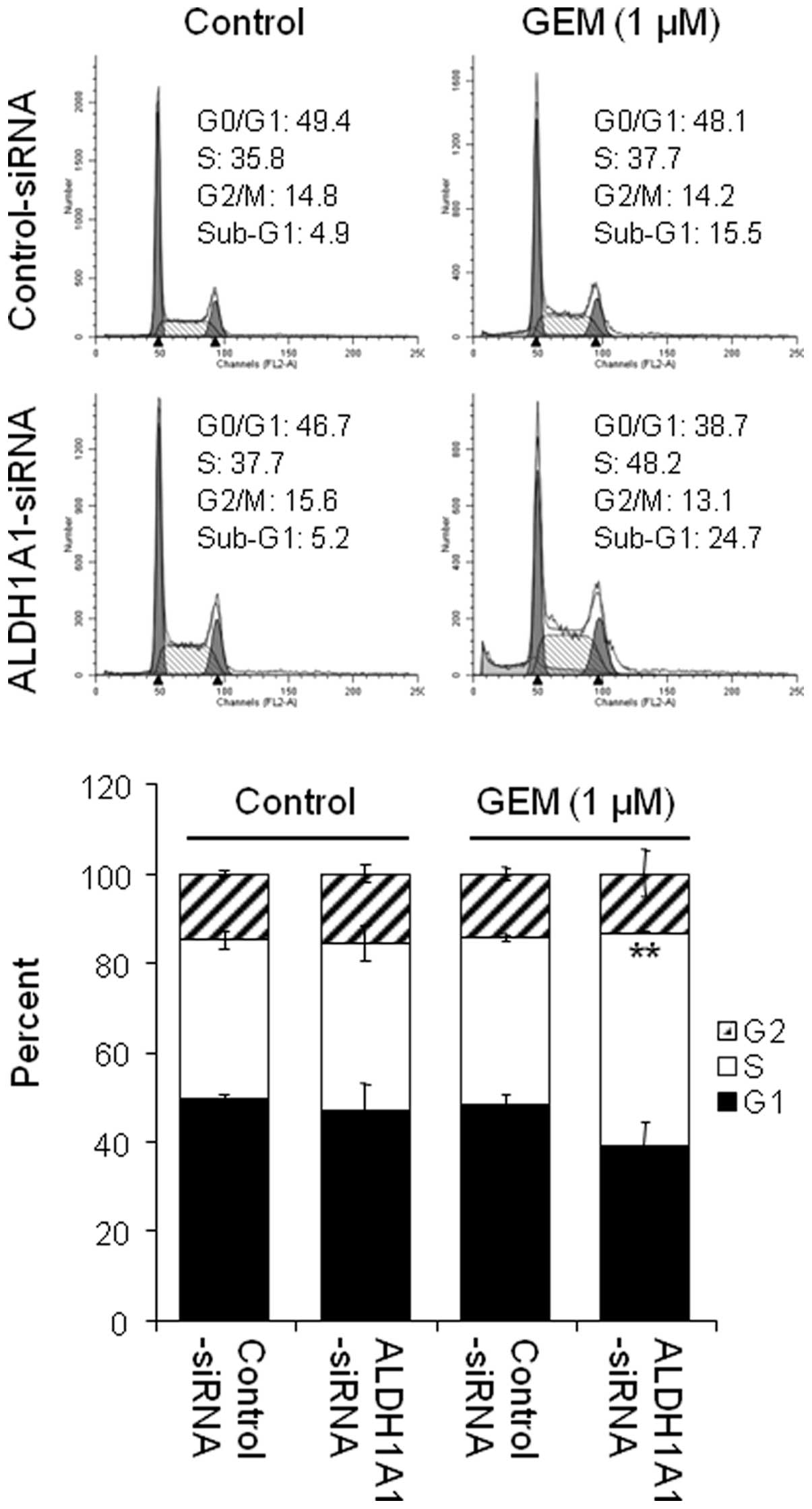
S-phase arrest by GEM
As GEM is known to accumulate in the S-phase of the
cell cycle, we were interested in the effects of the ALDH1A1
knockdown on GEM-induced S-phase arrest. Cells pre-treated with
ALDH1A1- or control-siRNA for 48 h were exposed to 1 μM GEM
for 48 h and their cell cycle profiles were assessed by FACS
analysis. The data showed that the knockdown of ALDH1A1 or
treatment with 1 μM GEM alone induced a small increase in
the accumulation of cells in the S-phase (from 35.8% by
control-siRNA to 37.7% by ALDH1A1-siRNA or to 37.7% by 1 μM
GEM) (Fig. 5). However, the
combined effects of ALDH1A1-siRNA plus GEM significantly increased
the cell population in the S-phase (from 37.7% by control-siRNA
plus GEM to 48.2% by ALDH1A1-siRNA plus GEM) (Fig. 5).
ALDH1A1 knockdown enhances apoptotic cell
death by GEM in MIA PaCa-2/GR cells
MIA PaCa-2/GR cells were generated from MIA Paca-2/P
cells that had been continuously exposed to increasing
concentrations of GEM as described in Materials and methods. MIA
PaCa-2/P and MIA PaCa-2/GR cells were treated with a various
concentrations of GEM (0, 0.01, 0.1, 1 and/or 10 μM) for 72
h and cell viability was evaluated by the MTT assay. The results
showed that MIA PaCa-2/GR cells were relatively resistant to GEM
with an IC50 value of 4.43 μM, whereas MIA
PaCa-2/P cells were relatively sensitive to GEM with an
IC50 value of 0.11 μM (Fig. 6A). The MIA PaCa-2/GR cells
demonstrated a 40.3-fold higher resistant index towards GEM than
MIA PaCa-2/P cells.
In order to compare the expression levels of ALDH1A1
between MIA PaCa-2/P and MIA PaCa-2/GR cells, cell lysates were
obtained from exponentially growing cells and used for WB analysis.
The expression level of ALDH1A1 was higher in MIA PaCa-2/GR cells
than in the control MIA PaCa-2/P cells (Fig. 6B). Similarly, the
AldeFluor-positive cell population was also higher in MIA PaCa-2/GR
cells (data not shown). Finally, we investigated whether the
increase in the ALDH1A1 expression level or activity in MIA
PaCa-2/GR cells correlated with GEM resistance. MIA PaCa-2/GR cells
pre-treated with ALDH1A1- or control-siRNA for 48 h were exposed to
various concentrations of GEM (0, 0.01, 0.1, 1 and/or 10 μM)
for 72 h and cell viability was determined by MTT assay. The
ALDH1A1-siRNA plus GEM combination demonstrated the most
synergistic effect. The IC50 of GEM decreased from 4.54
μM in the control-siRNA-treated cells to 0.94 μM in
the ALDH1A1-siRNA-treated cells (Fig.
6C). Moreover, we evaluated the effects of the ALDH1A1
knockdown on GEM-induced apoptosis in GEM-resistant cells. MIA
PaCa-2/GR cells pre-treated with ALDH1A1- or control-siRNA for 48 h
were exposed to 1 μM GEM for 48 h. Treatment with
ALDH1A1-siRNA plus GEM most significantly increased the level of
cleaved PARP and caspase-3 activity in MIA PaCa-2/GR cells
(Fig. 7A and B).
Discussion
In this study, we investigated the significant role
of ALDH1A1 in the chemoresistance of human pancreatic
adenocarcinoma to GEM. We found that i) MIA PaCa-2 and CFPAC-1
cells express higher levels of ALDH1A1 than the other cell lines;
ii) MIA PaCa-2/GR cells express higher levels of ALDH1A1 expression
and activity than MIA PaCa-2/P cells; iii) the ALDH1A1 knockdown
markedly reduced ALDH1A1 expression and activity and inhibited cell
proliferation in MIA PaCa-2 cells; iv) the ALDH1A1 knockdown
enhanced GEM-inhibited cell proliferation and GEM-induced apoptotic
cell death not only in MIA PaCa-2/P cells, but also in MIA
PaCa-2/GR cells and v) the ALDH1A1 knockdown enhanced GEM-induced
S-phase arrest. These results strongly support the hypothesis that
ALDH1A1 is an important determinant of chemoresistance to GEM. As
far as we know, this is the first data showing that a combination
of ALDH1A1-siRNA and GEM has synergistic anti-tumor effects in
human pancreatic cancer cells.
ALDH1A1 expression characterizes a subpopulation of
cells with tumor-initiating or CSC properties in several
malignancies. ALDH1A1 expression has also been found in a
subpopulation of cells with chemoresistance in numerous human
cancer types. Based on genomic and proteomic profiles, a number of
studies have shown that the levels of ALDH1A1 expression are
significantly higher in platinum- or taxane-resistant ovarian
cancer cells (26,28), classical and atypical
multidrug-resistant gastric carcinoma cells (29), cyclophosphamide-resistant human
carcinoma cells (30,31) and oxazaphosphorine-resistant human
malignant blood cell lines (32).
Moreover, ALDH1A1 has been proposed to play a significant role in
the mechanism of resistance to cyclophosphamide in human carcinoma
cells (30,31), oxazaphosphorine in human malignant
blood cells (32) and platinum or
taxane in human ovarian cancer cells (26). Shah et al and Hong et
al showed that GEM-resistant cell lines had an increased
expression of CD44, CD24 and ESA, which were reported as putative
markers of pancreatic CSCs (10,11).
These studies suggest that GEM preferentially targets more
differentiated and rapidly proliferating pancreatic tumor cells,
indicating the enrichment of the pancreatic CSC population in
GEM-resistant pancreatic cancer cells. In the present study, we
observed that the ALDH1A1-positive population in the GEM-resistant
MIA PaCa-2 cells (MIA PaCa-2/GR) was enriched in the long-term
treatment with GEM to establish resistant cell lines. Consistent
with our results, Kallifatidis et al showed that long-term
in vitro treatment with GEM for 21 days induced an
enrichment of ALDH1A1-positive pancreatic CSCs (33). Taken together, these results
suggest a promising strategy for targeting the pancreatic CSC
population by targeting ALDH1A1 to contribute overcoming resistance
to GEM.
Our study demonstrates that ALDH1A1 confers
resistance to GEM in ALDH1A1-positive MIA PaCa-2 cells. ALDH1A1 is
known to oxidize many intracellular aldehydes into carboxylic acids
(34) and detoxify free oxygen
radicals generated by chemotherapeutic agents. The induction of
reactive oxygen species (ROS) has been described to increase
mitochondrial membrane permeability and promote apoptosis. In a
previous study, GEM markedly increased ROS production and the
depletion of ROS significantly decreased GEM-induced growth
suppression, indicating that ROS plays a role in GEM-mediated
cytotoxicity in T3M4 pancreatic cancer cells (35). Thus, the high level of ALDH1A1 may
reduce GEM cytotoxicity by efficiently detoxifying ROS generated by
GEM. Moreover, either the ALDH1A1 knockdown or GEM treatment
induced cell cycle arrest at the S-phase. In addition, the combined
effects of ALDH1A1-siRNA plus GEM induced a greater accumulation of
cells in the S-phase, which is critical for growth inhibition.
Landen et al showed that the ALDH1A1 knockdown induced an
accumulation of cells in the S- and G2-phase in taxane-resistant
but not platinum-resistant ovarian cancer cells (26). However, the molecular mechanism of
the ALDH1A1-siRNA-induced S-phase arrest is not clear at this
point. Further studies are required to understand the function of
ALDH1A1 in the regulation of the cell cycle.
In conclusion, in the present study, we demonstrate
a potential significance of ALDH1A1 in two pancreatic cancer cell
lines (MIA PaCa-2/P and MIA PaCa-2/GR). Reproducing these findings
in other pancreatic cancer cell lines may help to determine whether
the effects are cancer cell line-specific or not. Although
ALDH1A1-positive cells were not isolated in this study, it may be
useful to investigate the correlation between pancreatic CSCs and
GEM resistance. Further studies on animal models will help to
determine the significant role of ALDH1A1 in drug resistance.
Acknowledgements
I.B. was supported by the National
Institutes of Health (1R03CA152530), the National Research
Foundation of Korea [R31-10069; World Class University (WCU)
program] and the Georgetown University Lombardi Comprehensive
Cancer Center (P30-CA051008). We also appreciate BioMedText,
Inc./Dr Rashmi Nemade for helpful discussions and editing.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar
|
|
3
|
Van Laethem JL, Verslype C, Iovanna JL,
Michl P, Conroy T, Louvet C, Hammel P, Mitry E, Ducreux M,
Maraculla T, Uhl W, Van Tienhoven G, Bachet JB, Marechal R,
Hendlisz A, Bali M, Demetter P, Ulich F, Aust D, Luttges J, Peeters
M, Mauer M, Roth A, Neoptolemos JP and Lutz M: New strategies and
designs in pancreatic cancer research: consensus guidelines report
from a European expert panel. Ann Oncol. 23:570–576.
2012.PubMed/NCBI
|
|
4
|
Burris HA III, Moore MJ, Andersen J, Green
MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo
AM, Tarassoff P, Nelson R, Dorr FA, Stephens CD and Von Hoff DD:
Improvements in survival and clinical benefit with gemcitabine as
first-line therapy for patients with advanced pancreas cancer: a
randomized trial. J Clin Oncol. 15:2403–2413. 1997.PubMed/NCBI
|
|
5
|
Conroy T, Desseiqne F, Ychou M, Bouche O,
Guimbaud R, Becouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de
la Fouchardiere C, Bennouna J, Bachet JB, Khemissa-Akouz F,
Pere-Verqe D, Delbaldo C, Assenat E, Chauffert B, Michel P,
Montoto-Grillot C and Ducreux M: FOLFIRINOX versus gemcitabine for
metastatic pancreatic cancer. N Engl J Med. 12:1817–1825. 2011.
View Article : Google Scholar
|
|
6
|
Cunninqham D, Chau I, Stocken DD, Valle
JW, Smith D, Steward W, Harper PG, Dunn J, Tudur-Smith C, West J,
Falk S, Crellin A, Adab F, Thompson J, Leonard P, Ostrowski J,
Eatock M, Scheithauer W, Herrmann R and Neoptolemos JP: Phase III
randomized comparison of gemcitabine versus gemcitabine plus
capecitabine in patients with advanced pancreatic cancer. J Clin
Oncol. 27:5513–5518. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ueno H, Kiyosawa K and Kaniwa N:
Pharmacogenomics of gemcitabine: can genetic studies lead to
tailor-made therapy. Br J Cancer. 97:145–151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Galmarini CM, Mackey JR and Dumontet C:
Nucleoside analogous: mechanisms of drug resistance and reversal
strategies. Leukemia. 15:875–890. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lapidot T, Sirard C, Vormoor J, Murdoch B,
Hoang T, Caceres-Cortes J, Minden M, Paterson B, Caliqiuri MA and
Dick JE: A cell initiating human acute myeloid leukaemia after
transplantation into SCID mice. Nature. 367:645–648. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shah AN, Summy JM, Zhang J, Park SI,
Parikh NU and Gallick GE: Development and characterization of
gemcitabine-resistant pancreatic tumor cells. Ann Surg Oncol.
14:3629–3637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hong SP, Wen J, Bang S, Park S and Song
SY: CD44-positive cells are responsible for gemcitabine resistance
in pancreatic cancer cells. Int J Cancer. 125:2323–2331. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moreb J, Schweder M, Suresh A and Zucali
JR: Overexpression of the human aldehyde dehydrogenase class 1
results in increased resistance to 4-hydroperocyclophosphamide.
Cancer Gene Ther. 3:24–30. 1996.PubMed/NCBI
|
|
13
|
Moreb JS: Aldehyde dehydrogenase as a
marker for stem cells. Curr Stem Cell Res Ther. 3:237–246. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang F, Qui Q, Khanna A, Todd NW, Deepak
J, Xing L, Wang H, Liu Z, Su Y, Stass SA and Katz RL: Aldehyde
dehydrogenase 1 is a tumor stem cell-associated marker in lung
cancer. Mol Cancer Res. 7:330–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li T, Su Y, Mei Y, Leng Q, Leng B, Liu Z,
Stass SA and Jiang F: ALDH1A1 is a marker for malignant prostate
stem cells and predictor of prostate cancer patients’ outcome. Lab
Invest. 90:234–244. 2010.PubMed/NCBI
|
|
16
|
Su Y, Qui Q, Zhang Z, Jiang Z, Leng Q, Liu
Z, Stass SA and Jiang F: Aldehyde dehydrogenase 1 A1-positive cell
population is enriched in tumor-initiating cells and associated
with progression of bladder cancer. Cancer Epidemiol Biomarkers
Prev. 19:327–337. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Balicki D: Moving forward in human mammary
stem cell biology and breast cancer prognostication using ALDH1.
Cell Stem Cell. 1:485–487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, Schott A, Hayes D, Birnbaum D, Wicha MS and Dontu G: ALDH1
is a marker of normal and malignant human mammary stem cells and a
predictor of poor clinical outcome. Cell Stem Cell. 1:555–567.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Charafe-Jauffret E, Ginestier C, Iovino F,
Tarpin C, Diebel M, Esterni B, Houvenaeghel G, Extra JM, Bertucci
F, Jacquemier J, Xerri L, Dontu G, Stassi G, Xiao Y, Barsky SH,
Birnbaum D, Viens P and Wicha MS: Aldehyde dehydrogenase 1-positive
cancer stem cells mediate metastasis and poor clinical outcome in
inflammatory breast cancer. Clin Cancer Res. 16:45–55. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Croker AK, Goodale D, Chu J, Postenka C,
Hedley BD, Hess DA and Allan AL: High aldehydrogenase and
expression of cancer stem cell markers selects for breast cancer
cells with enhanced malignant and metastatic ability. J Cell Mol
Med. 13:2236–2252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang EH, Hynes MJ, Zhang T, Ginestier C,
Dontu G, Appelman H, Fields JZ, Wicha MS and Boman BM: Aldehyde
dehydrogenase 1 is a marker for normal and malignant human colonic
stem cells (SC) and tracks SC overpopulation during colon
tumorigenesis. Cancer Res. 69:3382–3389. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carpentino JE, Hynes MJ, Appelman HD,
Zheng T, Sterndler DA, Scott EW and Huang EH: Aldehyde
dehydrogenase-expressing colon stem cells contribute to
tumorigenesis in the transition from colitis to cancer. Cancer Res.
69:8208–8215. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dembinski JL and Krauss S:
Characterization and functional analysis of a slow cycling stem
cell-like subpopulation in pancreas adenocarcinoma. Clin Exp
Metastasis. 26:611–623. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ucar D, Cogle CR, Zucali JR, Ostmark B,
Scott EW, Zori R, Gray BA and Moreb JS: Aldehyde dehydrogenase
activity as a functional marker for lung cancer. Chem Biol
Interact. 178:44–55. 2009. View Article : Google Scholar
|
|
25
|
Ma S, Chan KW, Hu L, Lee TK, Wo JY, Ng IO,
Zheng BJ and Guan XY: Identification and characterization of
tumorigenic live cancer stem/progenitor cells. Gastroenterology.
132:2542–2556. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Landen CN Jr, Goodman B, Katre AA, Steg
AD, Nick AM, Stone RL, Miller LD, Mejia PV, Jennings NB, Gershenson
DM, Bast RC Jr, Coleman RL, Lopez-Berestein G and Sood AK:
Targeting aldehyde dehydrogenase cancer stem cells in ovarian
cancer. Mol Cancer Ther. 9:3186–3199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Furukawa T, Duquid WP, Rosenberg L,
Viallet J, Galloway DA and Tsao MS: Long-term culture and
immortalization of epithelial cells from normal adult human
pancreatic ducts transfected by the E6E7 gene of human papilloma
virus 16. Am J Pathol. 148:1763–1770. 1996.PubMed/NCBI
|
|
28
|
Le Moguen K, Lincert H, Marcelo P,
Lemoisson E, Heutte N, Duval M, Poulain L, Ving J, Gauduchon P and
Baudin B: A proteomic kinetic analysis of JGROV1 ovarian carcinoma
cell line response to cisplatin treatment. Proteomics. 7:4090–4101.
2007.PubMed/NCBI
|
|
29
|
Ludwig A, Dietel M and Lage H:
Identification of differentially expressed genes in classical and
atypical multidrug-resistant gastric carcinoma cells. Anticancer
Res. 22:3213–3221. 2002.PubMed/NCBI
|
|
30
|
Russo JE and Hilton J: Characterization of
cytosolic aldehyde dehydrogenase from cyclophosphamide resistant
L1210 cells. Cancer Res. 48:2963–2968. 1988.PubMed/NCBI
|
|
31
|
Yoshida A, Dave V, Han H and Scanlon KJ:
Enhanced transcription of the cytosolic ALDH gene in
cyclophosphamide resistant human carcinoma cells. Adv Exp Med Biol.
328:63–72. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kohn FR, Landkamer GJ, Manthey CL, Ramsay
NK and Sladek NE: Effect of aldehyde dehydrogenase inhibitors on
the ex vivo sensitivity of human multipotent and committed
hematopoietic progenitor cells and malignant blood cells to
oxazaphosphorines. Cancer Res. 47:3180–3185. 1987.
|
|
33
|
Kallifatidis G, Labsch S, Rausch V,
Mattern J, Gladkich J, Moldenhauer G, Buchler MW, Salnikov AV and
Herr I: Sulforaphane increases drug-mediated cytotoxicity toward
cancer stem-like cells of pancreas and prostate. Mol Ther.
19:188–195. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Riveros-Rosas H, Julian-Sanchez A and Pina
E: Enzymology of ethanol and acetaldehyde metabolism in mammals.
Arch Med Res. 28:453–471. 1997.PubMed/NCBI
|
|
35
|
Donadelli M, Costanzo C, Beqhelli S,
Scupoli MT, Dandrea M, Bonora A, Piacentini P, Budillon A, Caraqlia
M, Scarpa A and Palmieri M: Synergistic inhibition of pancreatic
adenocarcinoma cell growth by trichostatin A and gemcitabine.
Biochim Biophys Acta. 1773:1095–1106. 2007. View Article : Google Scholar : PubMed/NCBI
|