Introduction
The p53 protein is a sequence-specific transcription
factor controlling the expression of multiple genes, thus
participating in the regulation of the cell cycle, apoptosis,
senescence and genome stability. This protein acts as tumor
suppressor protecting cells from oncogenic transformation. p53
dysfunction is a prerequisite for tumor development. Somatic p53
mutations are present in almost half of all cancer cases in almost
all types of cancer. Germ-line p53 mutations are associated with
the Li-Fraumeni syndrome, a rare cancer syndrome predisposing the
patients to a broad spectrum of tumors.
There are multiple ways of functional inactivation
of p53, including aberrations of its upstream or downstream
mediators and overexpression of its negative regulators. Direct
mutation of the p53 gene is the most frequent mechanism of
p53 inactivation. According to the latest version of the IARC TP53
mutation database (R15 from November 2010 at http://www-p53.iarc.fr) (1), the most common are single-base
substitutions representing >80% of all detected p53 mutations.
They are distributed throughout the coding region with strong
prevalence in its part coding for the central DNA-binding
domain.
The functional impact of the p53 mutations is
complex. Although many mutations cause complete loss of p53
function, some result in its reduction or modification. Modified
p53 often exhibits an altered spectrum of transactivated genes or
changes in dependency on external conditions, such as temperature
or pH (2,3). Partial loss of activity is an
important property of some p53 mutants and is associated with a
milder impact on the clinical course of the Li-Fraumeni syndrome
compared to complete p53 inactivation (4). The studies on p53 performed in
pediatric adrenocortical carcinoma (5,6)
provided formulation of ‘the p53 mutation gradient effect’, a
concept connecting the degree of p53 activity with its impact on
tumors (7).
The partially functional mutations comprise
approximately 10–11% of all p53 mutations (8,9). The
temperature-dependent (td) p53 mutants, the prototypes of the p53
mutants with incomplete and conditionally lost activity have
attracted great interest as they represent potential targets for
the pharmacological rescue of p53 function (10,11).
Several studies deal with the detection and analysis of td p53
mutations; they present a reasonably long list of td p53 mutations
and provide several interesting insights into understanding their
significance. It has been shown that td p53 mutations are not as
rare as had previously been thought and that they comprise
approximately 10% of p53 mutations detected in tumors and cause
single amino-acid substitutions. The td mutations are usually
clustered in the β-sandwich-coding region causing predominantly
hydrophobic large-to-small substitutions. Different td mutants are
inactivated to a different extent and consequently their permissive
temperatures also differ (3,12–16).
In our previous report, we set up a panel of 23
partially inactivated p53 mutants derived from various human
tumors. We described their conformation and transactivation
activity in yeast, their temperature dependency and capacity to
become reactivated by amifostine (17). Subsequently, we analyzed their
transactivation activity and temperature dependency in human H1299
cells (18). Functional analysis
of separated alleles in yeast (FASAY) (19,20)
used for routine detection of p53 mutations allowed us to collect
several new td mutants. In this study, we describe the
transactivating activity of nine of them in yeast cells. We
extensively analyzed their temperature dependency, discriminativity
and capacity to become reactivated by amifostine. Analysis of six
of the p53 td mutants was also performed in transiently transfected
human H1299 cells.
Materials and methods
FASAY
FASAY was performed as previously described
(20,21). Total RNA was purified using RNeasy
Mini Kit (Qiagen). cDNA was synthesized by SuperScript II (Life
Technologies) using oligo(dT)12 as a primer. PCR was
performed using primers P3
(5′-CCT-TGC-CGT-CCC-AAG-CAA-TGG-ATG-AT-3′), P4
(5′-ACC-CTT-TTT-GGA-CTT-CAG-GTG-GCT-GGA-GT-3′), and Pfu DNA
Polymerase (Stratagene). Yeast cells were co-transformed with the
PCR product, linearized pSS16, and the salmon sperm DNA carrier
(Life Technologies) by the lithium acetate procedure (19). Transformed yeast cells were plated
on minimal medium lacking leucine and with a low amount of adenine
(5 μg/ml), followed by incubation at 35°C for 2–3 days, and then at
room temperature for 2–3 days.
Recovery of the p53-expressing plasmid
from yeast and sequencing of the p53 cDNA
Yeast cells of individual yeast colonies were
harvested, resuspended in TSN (2% Triton X-100, 1% SDS, 100 mM
NaCl, 10 mM Tris pH 8.0, 1 mM EDTA), and grinded by vortexing with
glass beads; plasmid DNA was extracted by the phenol/chloroform
procedure. The p53 cDNA was amplified using the P3 and P4
primers and Taq polymerase (Life Technologies) and subjected
to agarose gel electrophoresis. The PCR product was purified by the
MinElute PCR purification kit (Qiagen) and sequenced by the BigDye
Terminator v3.1 cycle sequencing kit (Applied Biosystems) using ABI
PRISM 3100 Genetic Analyzer (Applied Biosystems).
Plasmid construction, cell culture and
transfection
The p53 cDNAs extracted from the yeast cells were
cloned into the p53-pT-REx-DEST30 plasmids using the
Gateway® technology (Invitrogen). H1299 cell lines were
cultured in RPMI-1640 medium (L-glutamine, NaHCO3)
supplemented with 10% fetal calf serum and 1%
penicillin/streptomycin at 5% CO2 and 32 or 37°C. To get
the H1299/p53 cells expressing the p53 variants, the cells were
co-transfected with p53-pT-REx-DEST30 (prepared according to the
manufacturer’s instructions) and pcDNA6/TR repressing p53
expression (Invitrogen) in a ratio of 1:7 using Lipofectamine™ 2000
(Invitrogen).
Luciferase reporter assay
H1299 cells were grown to 75% confluency,
co-transfected with p53-pT-REx-DEST30 (1 μg), cmv-βgal internal
control plasmid (1 μg) and the luciferase reporter plasmids (1 μg)
containing various p53-response elements: p21luc, baxluc or RGCluc
(22) using Lipofectamine 2000 and
incubated in parallel at 32 and 37°C for 42 h. Harvested cells were
resuspended in 0.25 M Tris-Cl (pH 7.5) and lysed by three cycles of
freezing and thawing. Luciferase and β-galactosidase assays were
performed as previously described (23). Relative transactivation activity
was expressed as a ratio of normalized luciferase activity of
individual p53 mutant and p53 wild-type (wt) activity at 37°C on
the same responsive element (RE).
Immunoblotting
Yeast or human cells were lysed with buffer
containing 150 mM NaCl, 50 mM NaF, 50 mM Tris (pH 8.0), 5 mM EDTA,
1% NP40 and 1 mM phenylmethylsulfonylfluoride protease inhibitor on
ice for 30 min, and the cell extract was centrifuged at 17,000 g
for 30 min to remove cell debris. Equal amounts of the supernatant
proteins were resolved by 10% sodium dodecylsulfate polyacrylamide
gel electrophoresis (SDS-PAGE) and transferred onto a
nitrocellulose membrane. Blots were blocked in 0.1% Tween-20 and 5%
low-fat milk in PBS for 1 h, probed with anti-p53 antibody DO-1
(kindly provided by B. Vojtesek). Blots were developed with Dako
peroxidase-conjugated rabbit anti-mouse immunoglobulin using the
ECL chemiluminiscent detection kit (Amersham Pharmacia
Biotech).
Reactivation by amifostine
Amifostine (WR2721, Ethyol®,
S-2[3-aminopropylamino]-ethyl phosphorothioic acid;
Schering-Plough), dissolved in water, was spread on the agar plates
at a final concentration of 8 mM 30 min before seeding yeast cells.
The parallel plates with and without amifostine were then incubated
at 25, 30 and 35°C for 3–4 days.
Results
Collection of the temperature-dependent
p53 mutants
FASAY can determine the functional status of p53
according to the color of the yeast colonies formed by transformed
cells on the selection medium. Expression of functional p53 results
in the formation of large white colonies, while inactive p53 leads
to the development of smaller red colonies. Expression of the td
p53 causes formation of pink or two-colored (white-red) colonies
(20,24). Based on phenotypes of the yeast
colonies, we identified nine p53 td mutants that were not included
in our collection of 23 mutants described earlier (17,18).
Five of these mutations were recognized as temperature-sensitive
ones by others (11,14,15,25),
the next four (M133T, V157A, R196G, and R283C), are described here
as temperature-dependent for the first time (Table I). The mutants originated from
several different tumors, namely colorectal carcinoma, chronic
lymphocytic leukemia, diffuse large B-cell lymphoma and
glioblastoma. Seven mutations (78%) were localized in the part of
the p53 gene coding for the hydrophobic β-sandwich structure
of the DNA-binding domain. The T211I mutation caused amino acid
substitution in the loop and the R283C in the α-helix structure of
the DNA-binding domain.
 | Table IList of temperature-dependent p53
mutations used in this study. |
Table I
List of temperature-dependent p53
mutations used in this study.
| Mutation | DNA sequence | Source | Exon | Structural
motifa | Reported as
somatic/germline mutationb | Reference |
|---|
| M133T | ATG-ACG | CRC | 5 | S2 | 7/3 | - |
| V157F | GTC-TTC | CLL | 5 | S4 | 188/0 | Dearth et al
(15) |
| V157A | GTC-GCC | DLBCL | 5 | S4 | 2/0 | - |
| R196G | CGA-GGA | CLL | 6 | S5 | 2/0 | - |
| T211I | ACT-ATT | CLL | 6 | L | 19/0 | Shiraishi et
al (14) |
| Y234H | TAC-CAC | glio | 7 | S8 | 27/2 | Campomenosi et
al (25) |
| I255F | ATC-TTC | CLL | 7 | S9 | 39/0 | Shiraishi et
al (14) |
| V272M | GTG-ATG | CLL | 8 | S10 | 103/1 | Maurici et
al (11); Dearth et al
(15) |
| R283C | CGC-TGC | CLL | 8 | H2 | 27/4 | - |
Transactivation by the
temperature-dependent p53 mutants in yeast
Td p53 mutants often discriminate among their target
genes (3,12,15).
For routine detection of p53 mutations, we used the yeast yIG397
cells containing the ribosomal gene cluster (RGC)-derived p53-RE
placed upstream of the ADE2 reporter gene (20). To analyze discriminativity and
temperature-dependency, we also used the YPH-p21 and YPH-bax
strains containing the p53-specific REs derived from the regulatory
regions of the p21 and bax genes, respectively,
placed upstream of the ADE2 (25). The yIG397, YPH-p21 and YPH-bax
cells transformed with td p53 mutants, p53wt and the hot-spot R175H
mutant as controls, were grown on selection media and incubated at
25, 30 and 35°C for 4 days. Relative transactivation by p53 was
assessed semi-quantitatively according to the color of the
resulting yeast colonies using an empirically established
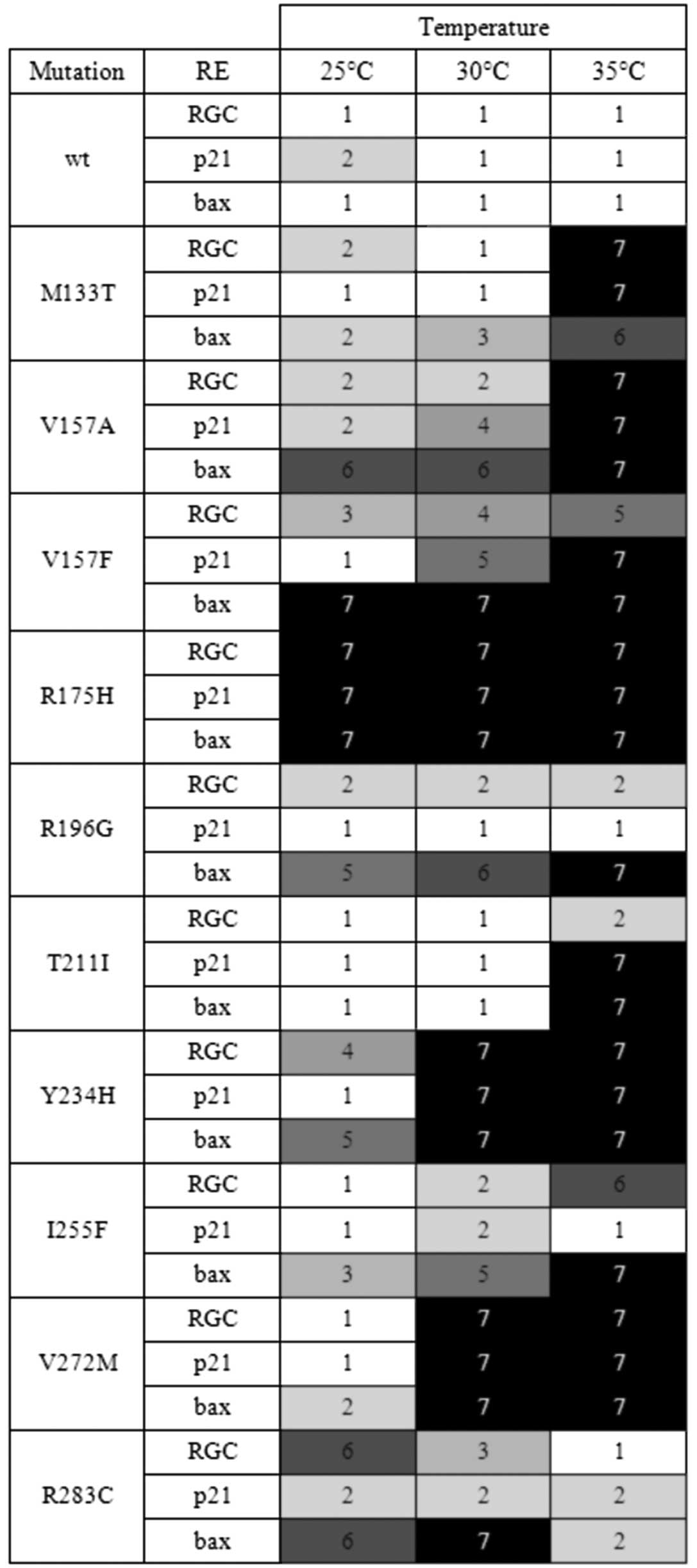
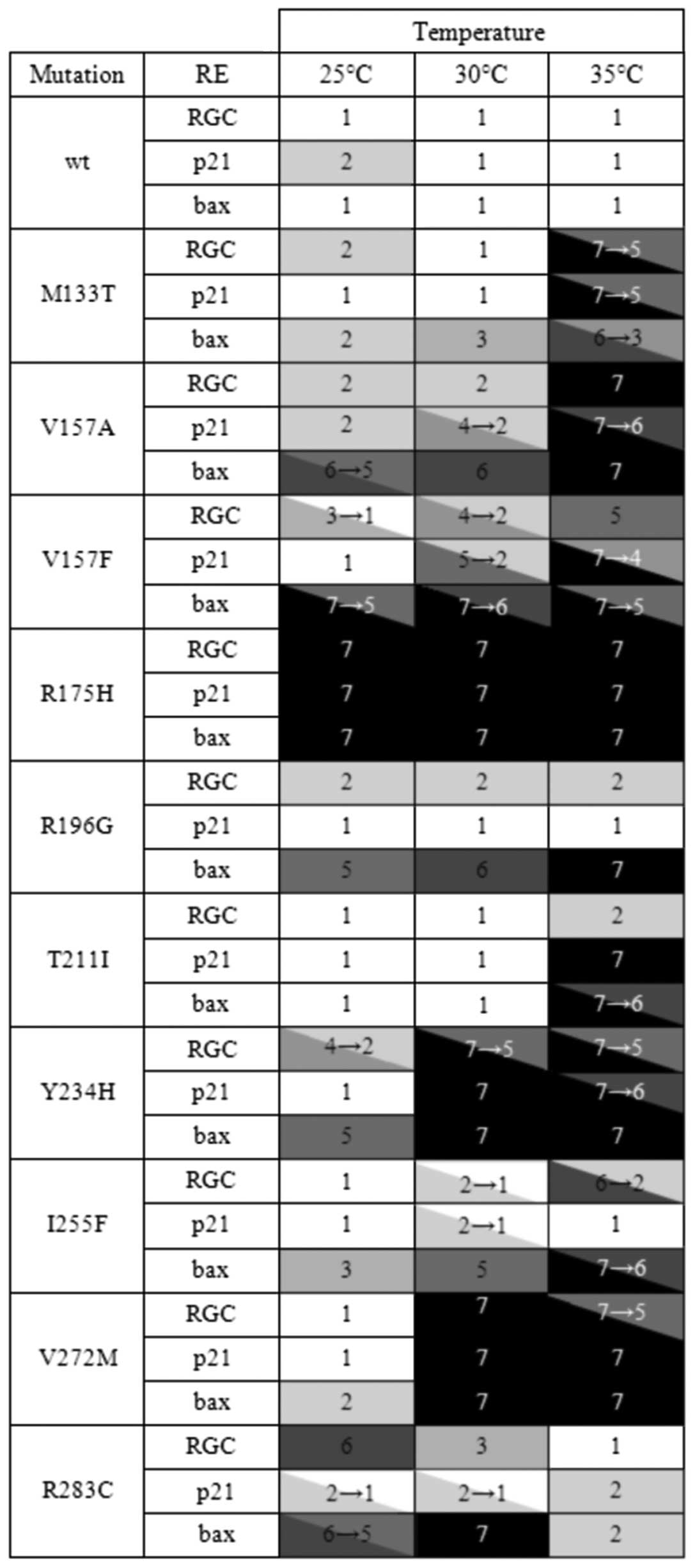
seven-step color scale (Fig. 1).
The scale comprises the tones of red color exhibited by the yeast
colonies depending on the transactivation rate of p53. The
endpoints of the scale are represented by the white color of the
colonies expressing fully active p53wt (step 1) and the dark red
color of the colonies expressing inactive hot spot mutant R175H
(step 7) (17).
The pattern of transactivation abilities of
individual mutants was considerable variable. The
temperature-dependency was confirmed for all tested mutants. Most
of them were temperature-sensitive exhibiting elevated or fully
restored activity at a lower temperature. The mutants M133T, T211I,
I255F, and V272M exhibited the strongest temperature-sensitivity
causing the lowest activity at an elevated temperature. The R283C
was the only mutant showing clear cold-sensitive (cs) phenotype. It
was almost fully active at 35°C, while losing transactivation
activity towards the RGC- and especially the bax-derived RE
at a lower temperature. Activity towards the p21-derived RE
was temperature-independent and comparable to the activity of
p53wt. Thus, this mutant was also clearly discriminating. In
addition to the R283C mutant, the R196G and I255F mutants also
belonged to the strongest discriminators. By contrast, the T211I,
V272, and M133T mutants discriminated only weakly or almost not at
all. The discriminators transactivated more efficiently from the
p21- than the bax-derived RE. Considering the overall
transactivating activity, there are also reasonable differences
among individual mutants. Some mutants, such as Y234H or V272M, can
weakly transactivate only at 25°C and are fully inactive at 30 or
35°C, while some other mutants, such as M133T, T211I and I255F are
inactive at 35°C and either fully or almost fully active at
30°C.
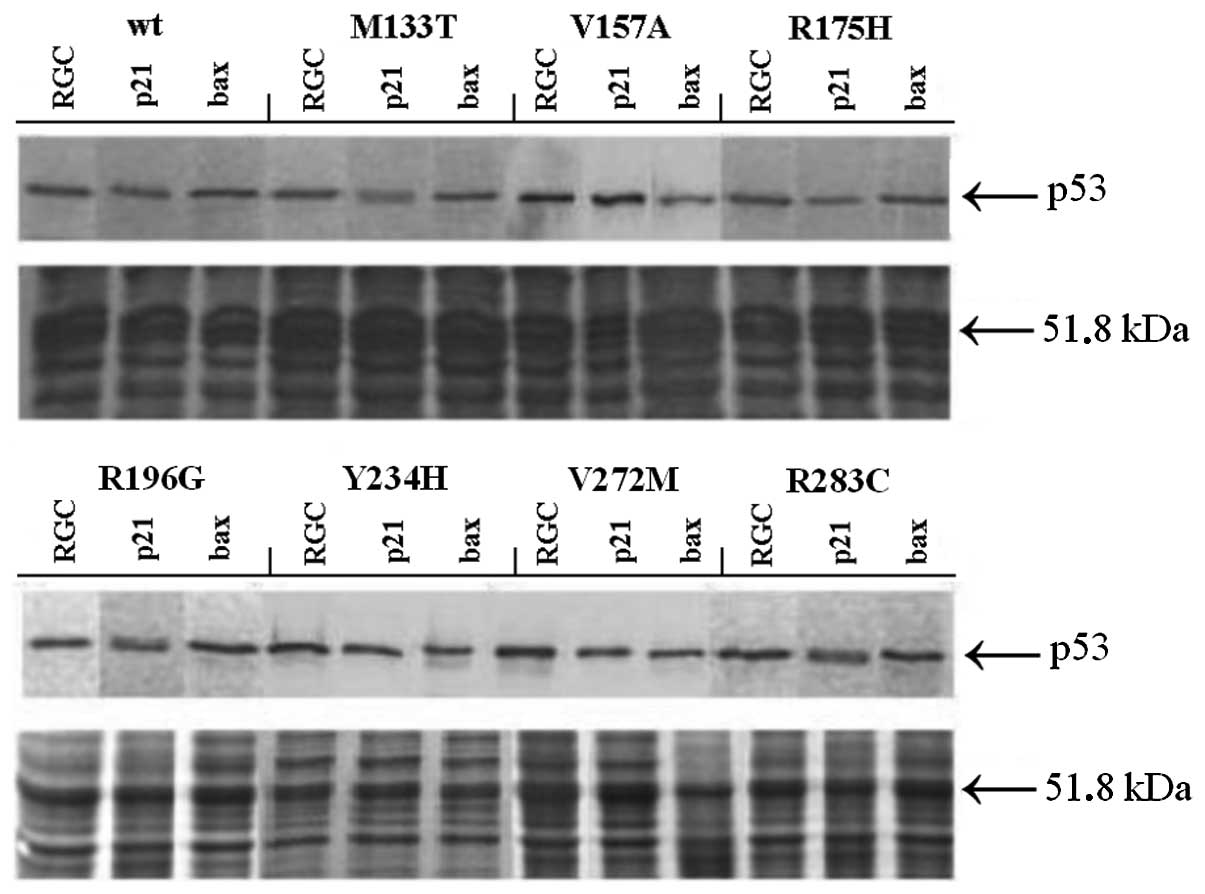
The p53 transactivation activity often depends on
the intracellular concentration of the p53 protein (3). Therefore, we assessed the expression
of all studied p53 variants in all three yeast strains by
immunoblotting using the p53-specific antibody DO-1 (Fig. 2). We found that the level of the
p53 protein was comparable in all yeast strains and temperatures
tested. This documents that the phenotype of the yeast colonies was
not affected by the level of p53 in cells.
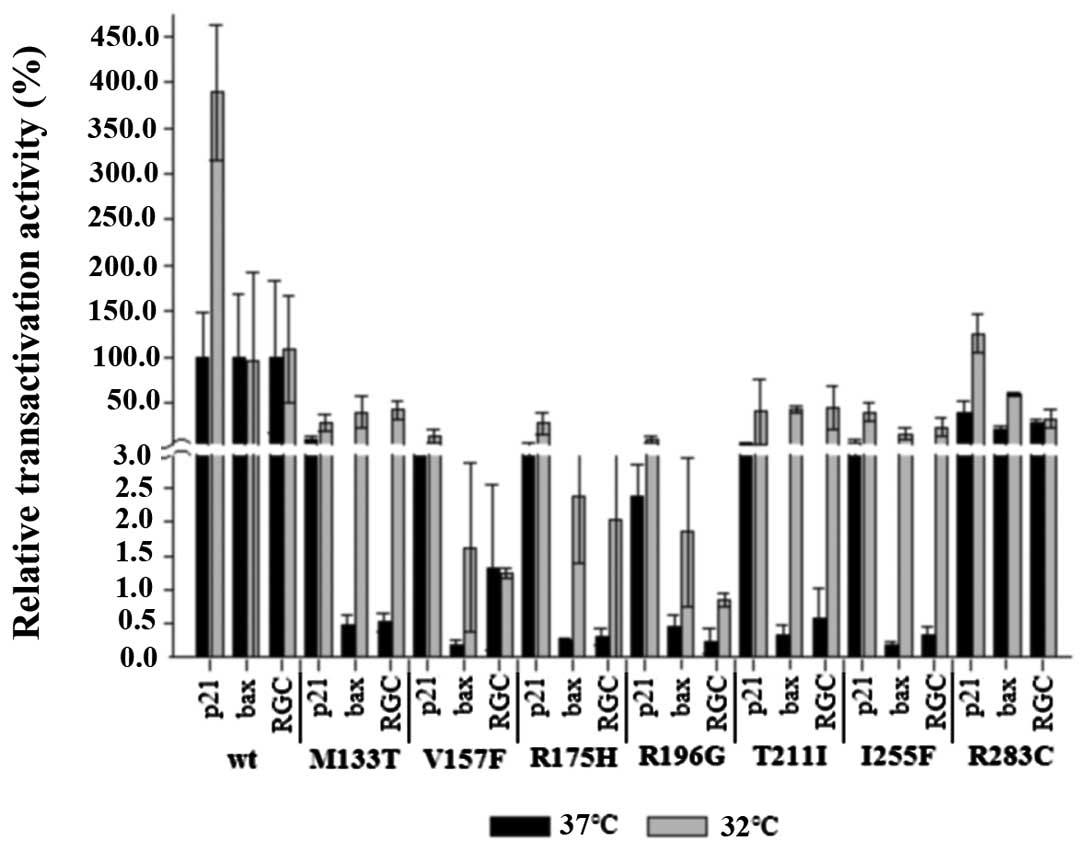
Transactivation by p53 variants in human
and yeast cells
To evaluate the significance of the data obtained in
yeast cells, we measured the transactivation ability of some p53 td
mutants (M133T, V157F, R196G, T211I, I255F, and R283C) and control
variants in human cells. cDNAs coding for the p53 variants were
cloned into the pT-REX-DEST30 expression vector. Subsequently, the
p53-null H1299 cells were transiently co-transfected with the
p53-expression plasmid, internal control plasmid (cmv-βgal) and one
of the RE-luc reporter plasmids containing p21-, bax-
or RGC REs upstream of the luciferase gene. Transfected cells were
cultured at 32 and 37°C for 42 h, then harvested and used for
transactivation assay. Luciferase activity was normalized according
to the transfection efficiency using β-galactosidase as internal
control. Relative transactivation activity of individual p53
mutants is shown in Fig. 3. As in
the yeast cells, we found considerable differences in the pattern
of transactivation activities of individual p53 variants. The
mutants differed in their overall activity, temperature
sensitivity, as well as discriminativity. The R283C mutant was the
most active of all td mutants tested, reaching 34.4% overall
activity of wt p53; it did not exhibit the cold-sensitive pattern
observed in transformed yeast cells and its temperature-sensitivity
was only weak, reaching reasonable activity even at 37°C. The R283C
mutant transactivated strongly and almost temperature-independently
from the p21-derived RE in yeast cells. In human cells, it
retained a rather strong transactivation ability towards this RE at
37°C and this activity was substantially elevated at 32°C,
exceeding even the activity of standard p53, thus fulfilling the
criteria of the super-trans mutant (27). The T211I, I255F and M133T mutants
exhibited the strongest temperature-sensitivity and they were all
transactivation competent. Generally, the low-active V157F and
R196G mutants also possessed weak temperature-sensitivity. At the
same time, these mutants were the strongest discriminators. The
temperature-sensitivity of the M133T, T211I and I255F mutants was
host-independent occurring both in yeast and human H1299 cells.
Similarly, the strongest discriminators in human cells, the mutants
V157F and R196G, were strong discriminators in yeast cells as well.
These phenomena were not dependent on the overall activity in yeast
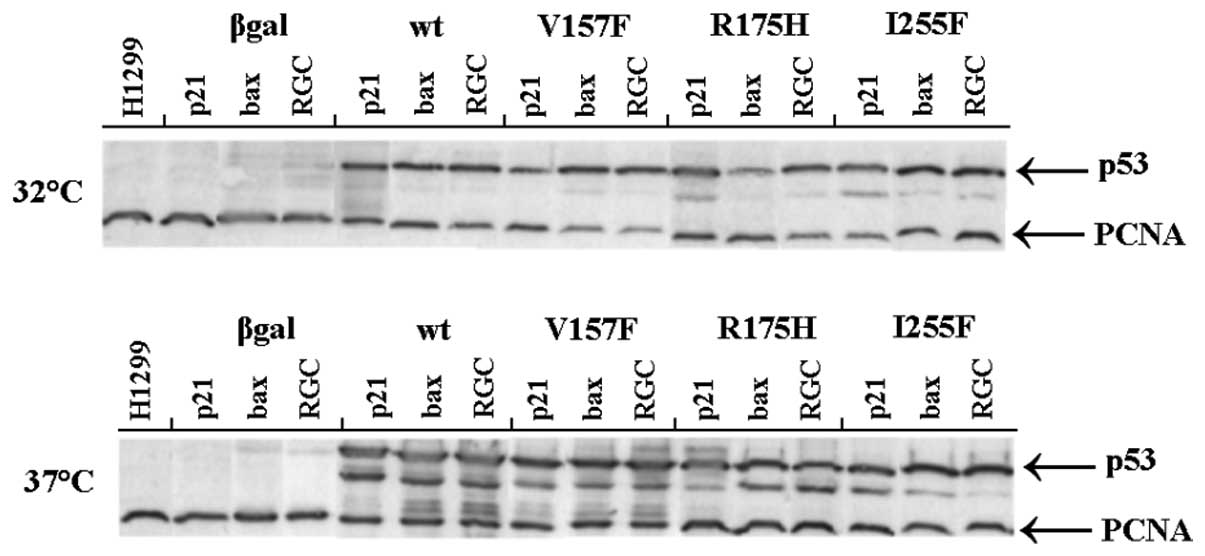
cells. To test whether or not the transactivation ability of the
p53 variants was affected by the p53 protein concentration in
H1299cells, we assessed the expression of all studied p53 variants
in transiently transfected cells by immunoblotting using the
p53-specific antibody DO-1 (Fig.
4). We verified that the level of the p53 proteins was
comparable in all tested cells.
Reactivation of td p53 mutants by
amifostine in yeast
We previously showed that the majority of td p53
mutants can be reactivated by amifostine in yeast cells (17). In this study, we tested the ability
of all nine td mutants to be reactivated by 8 mM amifostine in 25,
30 and 35°C towards all three tested REs. The yeast yIG397, YPH-p21
and YPH-bax cells expressing the tested p53 variants were plated in
selection medium either containing or lacking 8 mM amifostine and
incubated at 25, 30 and 35°C for 3–4 days. Then, the relative
transactivation by p53 was assessed according to the color of the
resulting colonies. The variable effect of amifostine was detected
in eight of them (Fig. 5). The
effect of amifostine was reproducibly temperature- and
RE-dependent. Only the R196G mutant was completely resistant to
amifostine under these conditions. On the other hand, the most
sensitive V157F mutant exhibited reactivation to some extent at all
tested temperatures and using all tested REs. The rest of the p53
td mutants scored in between these two possibilities. The level of
reactivation by amifostine did not follow any rule and was specific
for each p53 mutant.
Extent of interference with the p53
transactivation activity depends on the type of amino acid
exchange
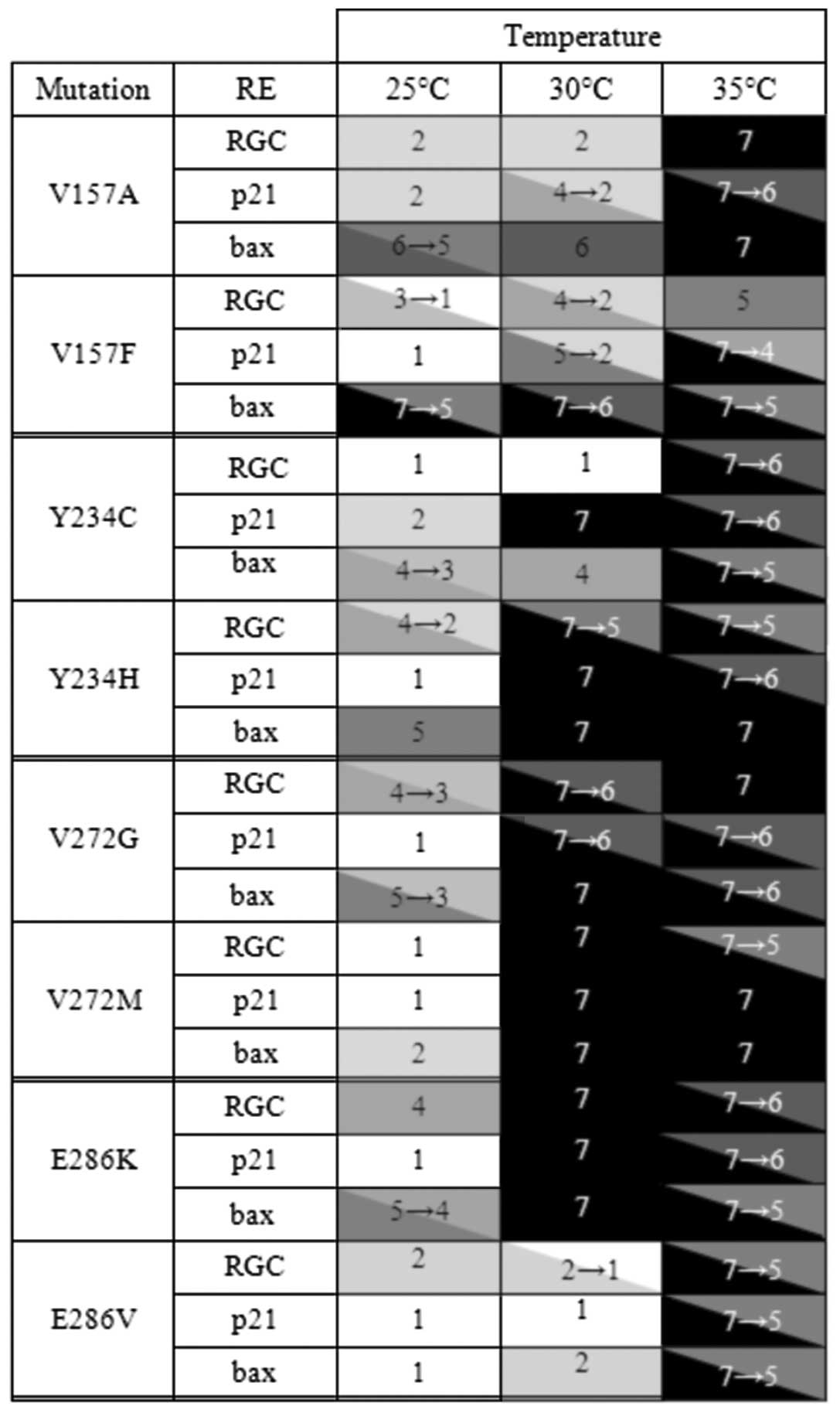
Our database of 32 td p53 mutants derived from
various types of tumors comprises already four pairs of mutants
localized in the same position, namely in codons 157 (causing
exchange of valine for alanine or phenylalanine), 234 (tyrosine for
cysteine or histidine), 272 (valine for glycine or methionine), and
286 (glutamic acid for lysine or valine). It allowed us to assess
the impact of the specific amino acid exchange on the activity of
p53 and its capacity to become reactivated by amifostine in yeast
cells (Fig. 6). The results
clearly show that the transactivation by p53, its temperature
dependency and discriminativity, as well as the capacity to become
reactivated by amifostine depend not only on the position of amino
acid substitution in the p53 protein but also on its specificity of
exchanged amino acids.
Discussion
In the present study we described nine
temperature-dependent (td) p53 mutants that were detected in
several different tumor types during routine analysis of the p53
status by FASAY. Temperature-dependency of five of these mutants
was reported earlier (11,14,15,25).
Four of them, M133T, V157A, R196G, and R283C, are described as
temperature-dependent in this study for the first time.
Nevertheless, there are other p53 mutants in the same positions
that have been recognized as temperature-dependent. For example,
the mutant bearing leucine instead of arginine in position 196
(R196L) and two mutants in position 283 (R283H, R283P) were
reported as temperature-sensitive (3,12,14,15,26,28).
We also present the V157F mutation previously described (15).
The temperature-dependent character of the p53
mutants is not obvious, and even if assessed, its impact is
unclear. Although there is no doubt that these p53 mutants
represent a specific group of mutants, the overwhelming majority of
clinical trials does not take this property into consideration.
First, with the exception of some functional methods, FASAY in
particular, temperature-dependency is not detectable by
conventional analyses of the p53 status. Second, even exact
determination of the temperature-dependency of the p53 variants
found in tumors is not unambiguous. The extent of damage of the p53
activity resulting from a temperature-dependent mutation ranges
from slight, leaving activity of the resulting p53 protein almost
unrecognizable from its standard variant, up to extensive,
resulting in protein with very low activity. This lack of clarity
is well documented by authors attempting to clearly and measurably
delimit and define criteria to select td p53 mutants (14). In addition, even p53wt has been
recognized as a temperature-sensitive protein and this fact
illustrates how difficult it is to define the
temperature-dependency of its mutants (17,29,30).
Altogether, the td p53 mutants do not comprise a uniform group.
Therefore, behavior of each mutant has to be tested and examined
individually. The data presented in this study clearly support this
view. Therefore, our findings contribute to the general conclusions
outlined below.
It has previously been postulated that the majority
of td mutations is localized in the β-sandwich-coding region of the
p53 gene. Out of the nine td mutants analyzed in this work,
seven were indeed mapped in this region.
The majority of the discriminating p53 mutants have
been repeatedly shown to prefer the p21- to the
bax-derived RE. This has even led to the conclusion that
abrogation of apoptosis represents the main target of p53
inactivation during oncogenic transformation (12,26).
Our discriminators clearly accomplished this rule. Nevertheless, we
detected a few exceptions in our previous collection of td p53
mutants, namely Y234C and P98A. Markedly, if a p53 mutant prefers
the bax-derived RE or uses both REs equally in yeast cells,
it usually keeps this pattern also in human cells. This is the case
of the P98A and Y126C mutants and, also to some extent, of the
T211I, V272M, Y234C and R337C mutants (17,18).
The cold-sensitive p53 mutants are rather rare. In
this study, we described R283C exhibiting a clear cold-sensitive
pattern in yeast cells. In human cells, R283C was only weakly
temperature-dependent; it was the most active of the td mutants
tested and it reached reasonable activity even at 37°C. Notably,
the P98A, P222L, R337C and L344R mutants classified as
cold-sensitive in yeast were the only mutants from the previous
collection of 23 td mutants which retained some transactivating
ability in human cells towards all three promoters also at 37°C.
The P98A mutant transactivated the p21-derived RE with the
same efficiency as p53wt at 37°C and with higher efficiency at this
temperature than at 32°C (17,18).
It seems that although it is not cold-sensitivity that
characterizes this group of mutants in human cells, once they are
cold-sensitive in yeast cells, it categorizes them into a specific
group of mutants exhibiting a specific pattern of transactivation
activity.
Specificity of amino acid exchange in p53 mutants is
important. Our previous panel of 23 td p53 mutants included two
mutants with replaced glutamic acid in position 286 for either
lysine or valine. These two mutants differed considerably in their
transactivation abilities in yeast (17) as well as in human cells (18). The mutant containing valine was
significantly more active than the mutant that had lysine in the
same position. In the present study, we confirmed this result and
further extended it for the next three pairs of mutants. We found
similarly strong differences in the activities of Y234C and Y234H,
while pairs of mutants V157A/F and V272G/M behaved more similarly,
though not equally. The impact of specific amino acid exchange on
the p53 protein activity is a function not only of the size of the
amino acid but also of its reactivity and flexibility. Similar
results have previously been reached and in several cases even
connecting these differences with a different clinical impact of
the mutants (7).
Despite some differences and minor discrepancies,
there is substantial concordance between the activity patterns of
different p53 mutants in yeast and human H1299 cells. This
conclusion confirms our own previous results as well as results
reported by others (15,18). This strongly justifies employment
of the yeast cell-derived systems, such as FASAY, for reliable
detection of p53 mutations of human origin.
In conclusion, we can confirm that despite some
similar activity patterns, there is no steady rule to categorize
the td p53 mutants in relation to their transactivation activity or
their reactivation capacity. Each missense mutation affects
specific amino acid interactions within the p53 molecule or its
contact with DNA in a unique way or alters the overall conformation
of the DNA-binding domain (8).
Thus, it is necessary to analyze each p53 mutant individually.
Acknowledgements
This study was supported by grant
NS/10448-3 of the Internal Grant Agency of the Ministry of Health
of the Czech Republic.
References
|
1.
|
Petitjean A, Mathe E, Kato S, Ishioka C,
Tavtigian SV, Hainaut P and Olivier M: Impact of mutant p53
functional properties on TP53 mutation patterns and tumor
phenotype: lessons from recent developments in the IARC TP53
database. Hum Mutat. 28:622–629. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Blagosklonny MV: P53 from complexity to
simplicity: mutant p53 stabilization, gain-of-function, and
dominant-negative effect. FASEB J. 14:1901–1907. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Resnick MA and Inga A: Functional mutants
of the sequence-specific transcription factor p53 and implications
for master genes of diversity. Proc Natl Acad Sci USA.
100:9934–9939. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Monti P, Ciribilli Y, Jordan J, Menichini
P, Umbach DM, Resnick MA, Luzatto L, Inga A and Fronza G:
Transcriptional Functionality of germ line p53 mutants infuences
cancer phenotype. Clin Cancer Res. 13:3789–3795. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Varley JM, McGown G, Thorncroft M, James
LA, Margison GP, Forster G, Evans DGR, Harris M, Kelsey AM and
Birch JM: Are there low-penetrance TP53 alleles? Evidence from
childhood adrenocortical tumors. Am J Hum Genet. 65:995–1006. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Ribeiro RC, Sandrini F, Figueiredo B,
Zambetti GP, Michalkiewicz E, Lafferty AR, DeLacerda L, Rabin M,
Cadwell C, Sampaio G, et al: An inherited p53 mutation that
contributes in a tissue-specific manner to pediatric adrenal
cortical carcinoma. Proc Natl Acad Sci USA. 98:9330–9335. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Zambetti GP: The p53 mutation ‘gradient
effect’ and its clinical implications. J Cell Physiol. 213:370–373.
2007.
|
|
8.
|
Kato S, Han SY, Liu W, Otsuka K, Shibata
H, Kanamura R and Ishioka C: Understanding the function-structure
and function-mutation relatiohship of p53 tumor suppressor protein
by high-resolution missense mutation analysis. Proc Natl Acad Sci
USA. 100:8424–8429. 2003. View Article : Google Scholar
|
|
9.
|
Olivier M, Hollstein M and Hainaut P: TP53
mutations in human cancers: Origins, consequences, and clinical
use. Cold Spring Harb Perspect Biol. 2:a0010082010. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
North S, Pluquet O, Maurici D,
El-Ghissassi F and Hainaut P: Restoration of wild-type conformation
and activity of a temperature-sensitive mutant of p53 (p53(V272M))
by the cytoprotective aminothiol WR1065 in the esophageal cancer
cell line TE-1. Mol Carcinog. 33:181–188. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Maurici D, Monti P, Campomenosi P, North
S, Frebourg T, Fronza G and Hainaut P: Amifostine (WR2721) restores
transcriptional activity of specific p53 mutant proteins in a yeast
functional assay. Oncogene. 20:3533–3540. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Di Como C and Prives C: Human
tumor-derived p53 proteins exhibit binding site selectivity and
temperature sensitivity for transactivation in yeast-based assay.
Oncogene. 19:2527–2539. 1998.PubMed/NCBI
|
|
13.
|
Bullock AN, Henckel J and Fersht AR:
Quantitative analysis of residual folding and DNA binding in mutant
p53 core domain: definition of mutant states for rescue in cancer
therapy. Oncogene. 19:1245–1256. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Shiraishi K, Kato S, Han SY, Liu W, Otsuka
K, Sakayori M, Ishida T, Takeda M, Kanamaru R, Ochuchi N and
Ishioka C: Isolation of temperature-sensitive p53 mutations from a
comprehensive missense mutation library. J Biol Chem. 279:348–355.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Dearth LR, Qian H, Wang T, Baroni TE, Zeng
J, Chen SW, Yi SY and Brachmann RK: Inactive full-length p53
mutants lacking dominant wild-type p53 inhibition highlight loss of
heterozygosity as an important aspect of p53 status in human
cancers. Carcinogenesis. 28:289–298. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Joerger AC and Fersht AR:
Structure-function-rescue: the diverse nature of common p53 cancer
mutants. Oncogene. 26:2226–2242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Grochova D, Vankova J, Damborsky J,
Ravcukova B, Smarda J, Vojtesek B and Smardova J: Analysis of
transactivation capability and conformation of p53
temperature-dependent mutants and their reactivation by amifostine
in yeast. Oncogene. 27:1243–1252. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Slovackova J, Grochova D, Navratilova J,
Smarda J and Smardova J: Transactivation by temperature-dependent
p53 mutants in yeast and human cells. Cell Cycle. 9:2141–2148.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Ishioka C, Frebourg T, Yan Yx, Vidal M,
Friend SH, Schmidt S and Iggo R: Screening patients for
heterozygous p53 mutations using a functional assay in yeast. Nat
Genet. 5:124–129. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Flaman JM, Frebourg T, Moreau V,
Charbonnier F, Martin C, Chappuis P, Sappino AP, Limacher JM, Bron
L, Benhattar J, et al: A simple p53 functional assay for screening
cell lines, blood, and tumors. Proc Natl Acad Sci USA.
92:3963–3967. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Smardova J, Nemajerova A, Trbusek M,
Vagunda V and Kovarik J: Rare somatic p53 mutation identified in
breast cancer: a case report. Tumor Biol. 22:59–66. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Friedlander P, Haupt Y, Prives C and Oren
M: A mutant p53 that discriminates between p53-responsive genes
cannot induce apoptosis. Mol Cell Biol. 16:4961–4971.
1996.PubMed/NCBI
|
|
23.
|
Navratilova J, Horvath V, Kozubik A, Lojek
A, Lipsick J and Smarda J: p53 arrest growth and induces
differentiation of v-Myb-transformed monoblasts. Differentiation.
75:592–604. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Pavlova S, Mayer J, Koukalova H and
Smardova J: High frequency of temperature-sensitive mutations of
p53 tumor suppressor in acute myeloid leukemia revealed by
functional assay in yeast. Int J Oncol. 23:121–131. 2003.PubMed/NCBI
|
|
25.
|
Campomenosi P, Monti P, Aprile A,
Abbondandolo A, Frebourg T, Gold B, Crook T, Inga A, Resnick MA,
Iggo R and Fronza G: p53 mutants can ften transactivate promoters
containing a p21 but not Bax or PIG3 responsive elements. Oncogene.
20:3573–3579. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Flaman JM, Robert V, Lenglet S, Moreau V,
Iggo R and Frebourg T: Identification of human p53 mutations with
differential effects on the bax and p21 promoters using functional
assay in yeast. Oncogene. 16:1369–1272. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Inga A, Monti P, Fronza G, Darden T and
Resnick M: p53 mutants exhibiting enhanced transcriptional
activation and altered promoter selectivity are revealed using a
sensitive, yeast-based functional assay. Oncogene. 20:501–513.
2001. View Article : Google Scholar
|
|
28.
|
Fulci G, Ishii N, Maurici D, Gernert KM,
Hainaut P, Kaur B and Van Metr EG: Initiation of human astrocytoma
by clonal evolution of cells with progressive los sof p53 functions
in a patient with a 283H TP53 germ-line mutation: Evidence for
precursor lesion. Cancer Res. 62:2897–2905. 2002.PubMed/NCBI
|
|
29.
|
Hansen S, Hupp TR and Lane DP: Allosteric
regulation of the thermostability and DNA binding activity of human
p53 by specific interacting proteins. J Biol Chem. 271:3917–3924.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Xirodimas DP and Lane DP: Molecular
evolution of the thermosensitive PAb1620 epitope of human p53 by
DNA shuffling. J Biol Chem. 274:28042–28049. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Cho Y, Gorina S, Jeffrey PD and Pavletich
NP: Crystal structure of a p53 tumor suppressor-DNA complex:
understanding tumorigenic mutations. Science. 265:346–355. 1994.
View Article : Google Scholar : PubMed/NCBI
|