Introduction
Breast cancer is the most common malignancy among
females, accounting for nearly 1 in 3 cancers diagnosed in women
worldwide, and it is the second leading cause of cancer death after
lung cancer (1). The major
treatment procedures for breast cancer patients are surgery,
radiotherapy and chemotherapy; however, the cure rates are not
satisfactory. New agents acting on novel targets in breast cancer
are currently under investigation.
The role of dietary flavonoids in cancer prevention
is widely discussed. Convincing data from laboratory studies,
epidemiological investigations, and human clinical trials indicate
that flavonoids have imperative effects on cancer chemoprevention
and chemotherapy (2). Quercetin
(3,3′,4′,5,7-pentahydroxyavone), a member of the flavonoids family,
is one of the most prominent dietary antioxidants present in human
diet (3,4) and exerts diverse biological
activities in a variety of cancer cell model, including ovarian,
endometrial, lymphoma, prostate, liver and gastric cancer (5–7).
Although precise molecular mechanisms underlying quercetin-mediated
cellular responses on breast cancer remain poorly defined, prior
research has shown that it can modulate diverse proteins involved
in signal transduction pathways associated with cell survival,
apoptosis and proliferation (8–10).
Current studies have shown the importance of several
signaling pathways in breast carcinogenesis and progression
(11,12) including the Wnt/β-catenin pathway
(13–15). The Wnt/β-catenin pathway plays a
vital role in mammary tumorigenesis since overexpression of Wnt1
gene in the mammary epithelium is sufficient for mammary gland
hyperplasia and adenocarcinomas (16). The canonical Wnt ligands,
exemplified by Wnt1 or Wnt3a, bind to frizzled receptor (Fz) and
the low-density lipoprotein receptor-related protein-5/6 (LRP5/6),
and prevent phosphorylation and degradation of β-catenin by the
GSK3β/APC/axin destruction complex. Subsequent accumulation of
cytosolic and nuclear β-catenin bound to T cell factor (TCF)
transcription factors results into activation of downstream signals
which are important for proliferation and matrix remodeling
(17). The elevated levels of
nuclear and/or cytoplasmic β-catenin in breast carcinomas
correlated with the expression of its target gene cyclin D1 and
poor patient prognosis (18). In a
microarray based study, Huang et al (19) showed that inhibition of
Wnt/β-catenin pathway leads to apoptosis in HeLa cells by
upregulating the expression of several pro-apoptotic genes,
involved in apoptotic cell death pathways, such as PTEN-PI3K-AKT
pathway, NFκB pathway and p53 pathway. Furthermore, studies in mice
strongly imply that deregulated β-catenin signaling increases risk
of breast cancer by inducing stem cell and early progenitor cell
accumulation (20,21). Several studies have accounted for
increased cytoplasmic and nuclear β-catenin in primary breast
cancers, especially basal-like breast cancers, and correlated with
poor prognosis and survival (22,23).
Thus, it needs to be investigated whether the activation of
Wnt/β-catenin signaling can be a potential target for the
chemoprevention and treatment of breast cancer.
To identify molecular mechanism for
quercetin-induced apoptosis, we used the murine mammary carcinoma
cell line, 4T1, which is highly tumorigenic and invasive (24). Contrary to most tumor models, 4T1
cells injected into the mammary fat pads of syngeneic BALB/c mice
grow spontaneously into multiple distant organs such as lymph
nodes, liver, lung, brain and bone (25). Therefore, 4T1 is widely used as one
of the most aggressive type of breast cancer cell for the study of
breast cancer. Although quercetin has been reported to stimulate
cell cycle arrest and apoptosis in breast cancer cells (26) the exact molecular mechanism of
action still remains unclear. In this study, we show that quercetin
induces apoptosis of mammary cancer cells through DKK-dependent
inhibition of the Wnt/β-catenin signaling pathway.
Materials and methods
Cell culture and reagents
Murine mammary cancer cell line 4T1 (ATCC, CRL-2539)
was cultured in RPMI-1640 (Invitrogen, Carlsbad, CA) supplemented
with 10% fetal bovine serum (FBS), 100 U/ml penicillin and 100 U/ml
streptomycin (Lonza, Basel, Switzerland) at 37°C and 5%
CO2. Quercetin was purchased from Sigma (St. Louis, MO,
USA). A stock solution (25 mM) of quercetin was prepared in
dimethyl-sulfoxide (DMSO). Recombinant proteins, Wnt3a and DKK1,
were purchased from R&D Systems (Minneapolis, MN).
MTT assay
MTT assay was carried out to evaluate the
viabilities of 4T1 cells, and performed using
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT;
Sigma). 4T1 cells (1.5×104 cells/well) were plated in
96-well plates and allowed to attach overnight. Cells were then
treated with quercetin at various concentrations (5, 10, 20 and 40
μM) or the vehicle (0.16% of DMSO) and incubated at 37°C
with 5% CO2 for 48 h. Fresh MTT (5 mg/ml) was added to
growing cells in 96-well plate and incubated at 37°C for 2 h.
Following the removal of supernatant, the insoluble formazan
crystals were dissolved in 200 μl of DMSO and optical
density was measured at wavelength of 570 nm.
Assessment of cell apoptosis
4T1 cells were treated with quercetin (20 μM)
for 48 h, and then apoptotic cells were quantified by using an
Annexin V-FITC/PI double staining assay kit (BD Pharmingen, San
Diego, CA). Both, early and late apoptotic changes in 4T1 cells
were analyzed. Briefly, cells (1×106) were collected and
washed twice with phosphate-buffered saline (PBS) and suspended in
400 μl of binding buffer (added with 5 μl of Annexin
V-FITC and 5 μl of PI). The combination of Annexin V-FITC
and PI staining permitted the simultaneous quantification of vital,
apoptotic and necrotic cells. Thereafter, the samples were
incubated in the dark for 15 min at room temperature, and then
analyzed on the flow cytometer. The number of Annexin
V-FITC-positive and PI-positive cells in each field was determined
by counting the cells directly. Experiments were performed in
triplicate for accuracy.
RNA isolation and real-time RT-PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen). RNA purity was verified by measuring 260/280
absorbance ratio. The first strand of cDNA was synthesized with 2
mg of total RNA using SuperScript II (Invitrogen), and one-tenth of
the cDNA was used for each PCR mixture containing Express
SYBR-Green qPCR Supermix (BioPrince, Seoul, Korea). Real-time PCR
was performed using a Rotor-Gene Q (Qiagen, Hilden, Germany). The
reaction was subjected to 40-cycle amplification at 95°C for 20
sec, at 60°C for 20 sec and at 72°C for 25 sec. Relative mRNA
expression of selected genes was normalized to GAPDH and quantified
using the DDCT method. The sequences of the PCR primers are listed
in Table I.
 | Table I.Primers for real-time RT-PCR. |
Table I.
Primers for real-time RT-PCR.
| Gene | Forward primer
(5′→3′) | Reverse primer
(3′→5′) | Size (bp) | Accession no. |
|---|
| DKK1 |
TCAGGTCCATTCTGGCCAACTCTT |
TGGGCATTCCCTCCCTTCCAATAA | 132 | NM_010051.3 |
| DKK2 |
ATGGCAGAATCTAGGAAGGCCACA |
CGAACCCTTCTTGCGTTGTTTGGT | 184 | NM_020265.4 |
| DKK3 |
AGCTGATGGAAGACACTCAGCACA |
TCCTGGTGCACATGGACTGTGTTA | 175 | NM_015814.2 |
| DKK4 |
ATGGTACTGGTGACCTTGCTTGGA |
TCCGCGGAGCTCTTGATGTTGTTA | 92 | NM_145592.2 |
| GAPDH |
TCAACAGCAACTCCCACTCTTCCA |
ACCCTGTTGCTGTAGCCGTATTCA | 115 | NM_008084.2 |
Luciferase reporter assay
4T1 cells were plated at a density of
2×104 cells/well in 48-well plates, and transfected
using Genefectine transfection reagent (Genetrone Biotech Co.,
Korea) according to the manufacturer’s protocol. The TopFlash
(Addgene, Cambridge, MA) luciferase reporter (100 ng) and Renilla
luciferase thymidine kinase construct (Invitrogen) (50 ng) were
used to determine luciferase activity. Luciferase activity was
measured by a luminometer (Glomax, Promega, Sunnyvale, CA), using a
Dual-Luciferase assay kit (Promega), according to the
manufacturer’s recommendations. Total value of reporter activity in
each sample was normalized to Renilla luciferase activity.
Protein isolation and western blot
analysis
Cells were lysed in RIPA buffer (20 mM Tris-HCl, pH
7.5, 200 mM NaCl, 1% Triton X-100, 1 mM dithiothreitol) containing
protease inhibitor cocktail (Roche). The concentration of protein
was measured with a Protein assay kit (Bio-Rad) following the
manufacturer’s protocol. Total protein was subjected to
SDS-polyacrylamide gel electrophoresis and transferred to a PVDF
membrane. The blot was probed with primary antibody; anti-β-catenin
(Cell Signaling Technology). As a loading control, anti-β-actin
antibody (Santa Cruz biotechnology) was used. Subsequently, the
blots were washed in TBST (10 mM Tris-HCl, 50 mM NaCl, 0.25%
Tween-20) and incubated with a horseradish peroxidase-conjugated
secondary antibody. The presence of target proteins was detected
using the enhanced chemiluminescence reagents (BioNote Inc.,
Hwaseong, Korea).
Lactate dehydrogenase (LDH) activity
assay
Measurement of LDH activity was performed with
cytotoxicity detection kit (Roche) according to the manufacturer’s
protocol as follows. Cell culture media (10 μl) from each
experimental sample was added to a 96-well plate containing 40
μl of PBS. Next, 50 μl of LDH reagent was added to
each well and plates were incubated for 45 min at 25°C in dark, and
then enzymatic reaction was stopped by adding the stop solution (50
μl). Absorbance was read at wavelength of 492 nm. Total cell
lysate served as a positive control of cell death.
Statistical analysis
All the statistical data were analyzed by GraphPad
Prism 5.0 (GraphPad Software, San Diego, CA) and evaluated by
two-tailed Student’s t-test. Value of P<0.05 was considered to
indicate statistical significance.
Results
The treatment of quercetin delays the
growth and induces apoptosis of 4T1 cells
Initially, we attempted to determine whether
quercetin could exert anti-proliferative effect on 4T1 mouse
mammary cancer cell line as reported in case of diverse breast
cancer cell lines (27–29). After treatment of quercetin at
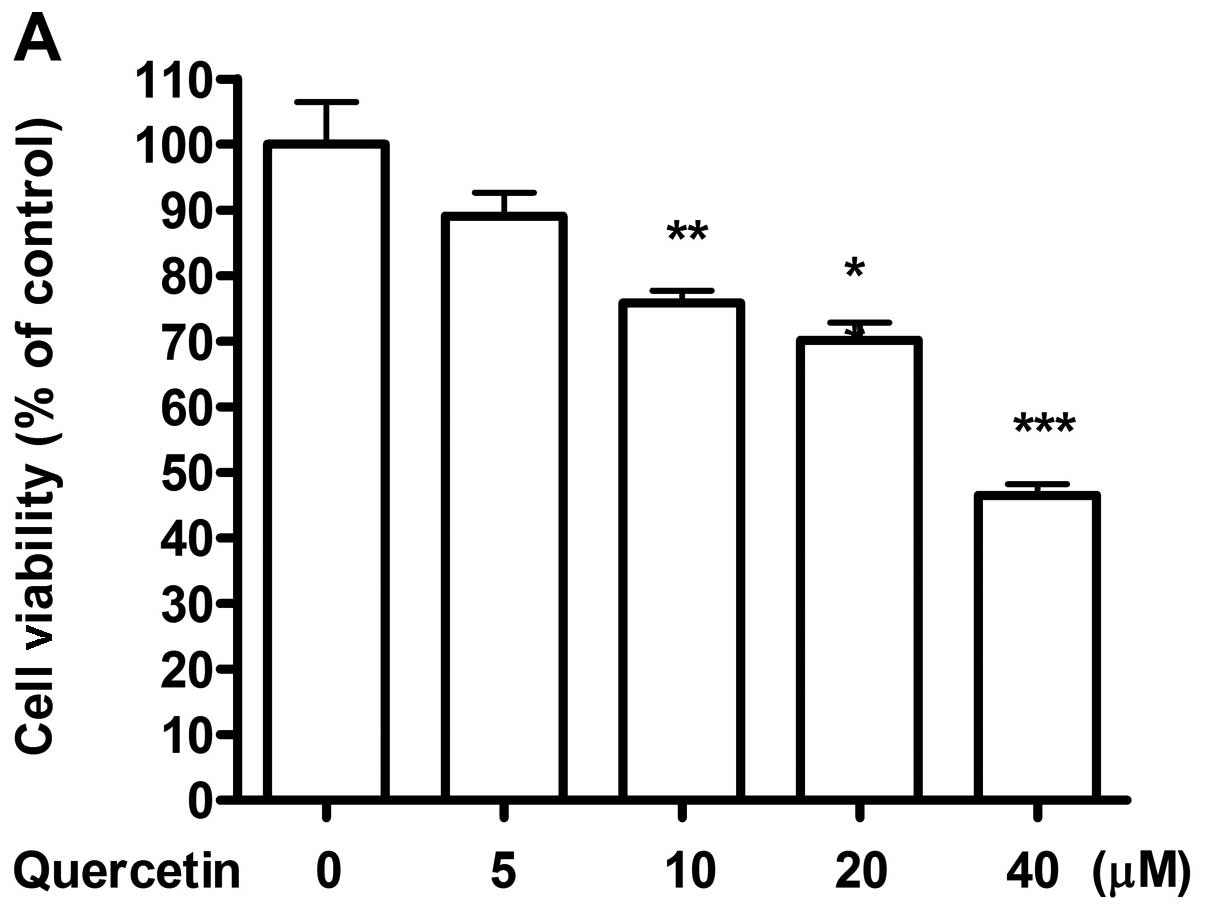
various doses for 48 h, a dose-dependent decrease in cell viability
of 4T1 cells was observed (Fig.
1A). Cells taken as control showed increase in confluence from
initial 60 to 100% after 48 h of culture. However, the cells
treated with 10 and 20 μM of quercetin reached only 70–80%
of confluence. Moreover, a dose of 40 μM of quercetin showed
50% decrease in initial cell number. Hence, for further
experiments, 20 μM of quercetin was selected as an ideal
dose to detect the effect of quercetin. Since cell apoptosis may be
one of the consequences of growth arrest, we stained the cells with
Annexin V-FITC and PI, and analyzed the apoptotic effect of
quercetin on 4T1 cells using flow cytometry. Fig. 1B showed an increased apoptotic
effect of quercetin at a dose of 20 μM (37%) as compared to
control (18%). The population of early apoptotic cells increased
∼3-fold compared to control whereas, significant increase in late
apoptotic cells were also observed.
Wnt/β-catenin signaling activity is
decreased by quercetin
As Wnt/β-catenin signaling pathway is implicated in
regulating tumor growth (30), the
effect of quercetin on Wnt/β-catenin signaling pathway of 4T1 cells
needed defining. To detect the activity of Wnt/β-catenin signaling
pathway, we used the luciferase reporter assay. 4T1 cells were
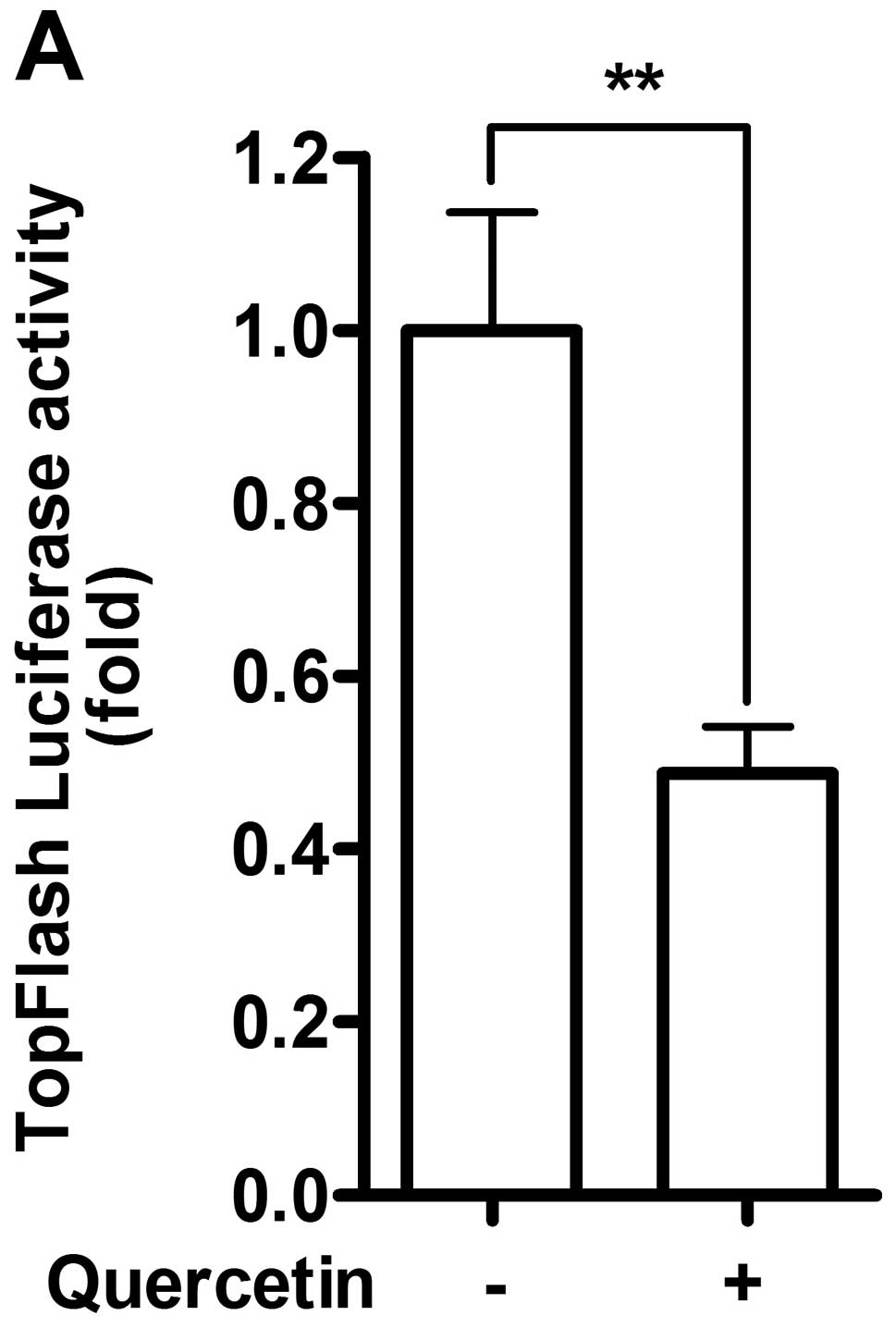
transfected with TopFlash-luc for Wnt/β-catenin signaling pathway.
After 48 h of treatment, quercetin suppressed ∼50% of basal level
of TopFlash luciferase activity, demonstrating inhibitory effect of
quercetin on Wnt/β-catenin signaling pathway (Fig. 2A). Canonical Wnt signaling acts
through stabilization of β-catenin, which translocate into the
nucleus to regulate the expression of genes related to tumor
growth, such as cyclin D1 and survivin (31). Thus, in order to confirm the
inhibitory role of quercetin on Wnt/β-catenin signaling pathway as
observed by luciferase assay, the stability of β-catenin was
detected by western blot analysis. We observed reduction of
β-catenin stabilization in quercetin treated cells after 1 h of
treatment, confirming inhibitory effect of quercetin on
Wnt/β-catenin signaling pathway (Fig.
2B).
Quercetin induces the gene expression of
DKK1, 2, 3 and 4
Wnt/β-catenin signaling pathway is regulated by
several processes. Secreted proteins such as secreted
frizzled-related protein (sFRP) and Dickkopf (DKK) act as
extracellular antagonists and are among various mechanisms to
regulate Wnt/β-catenin signaling pathway (32,33).
To investigate whether various negative regulators of Wnt/β-catenin
signaling pathway is being upregulated in 4T1 cells by quercetin,
we next examined the expression of known negative regulators by
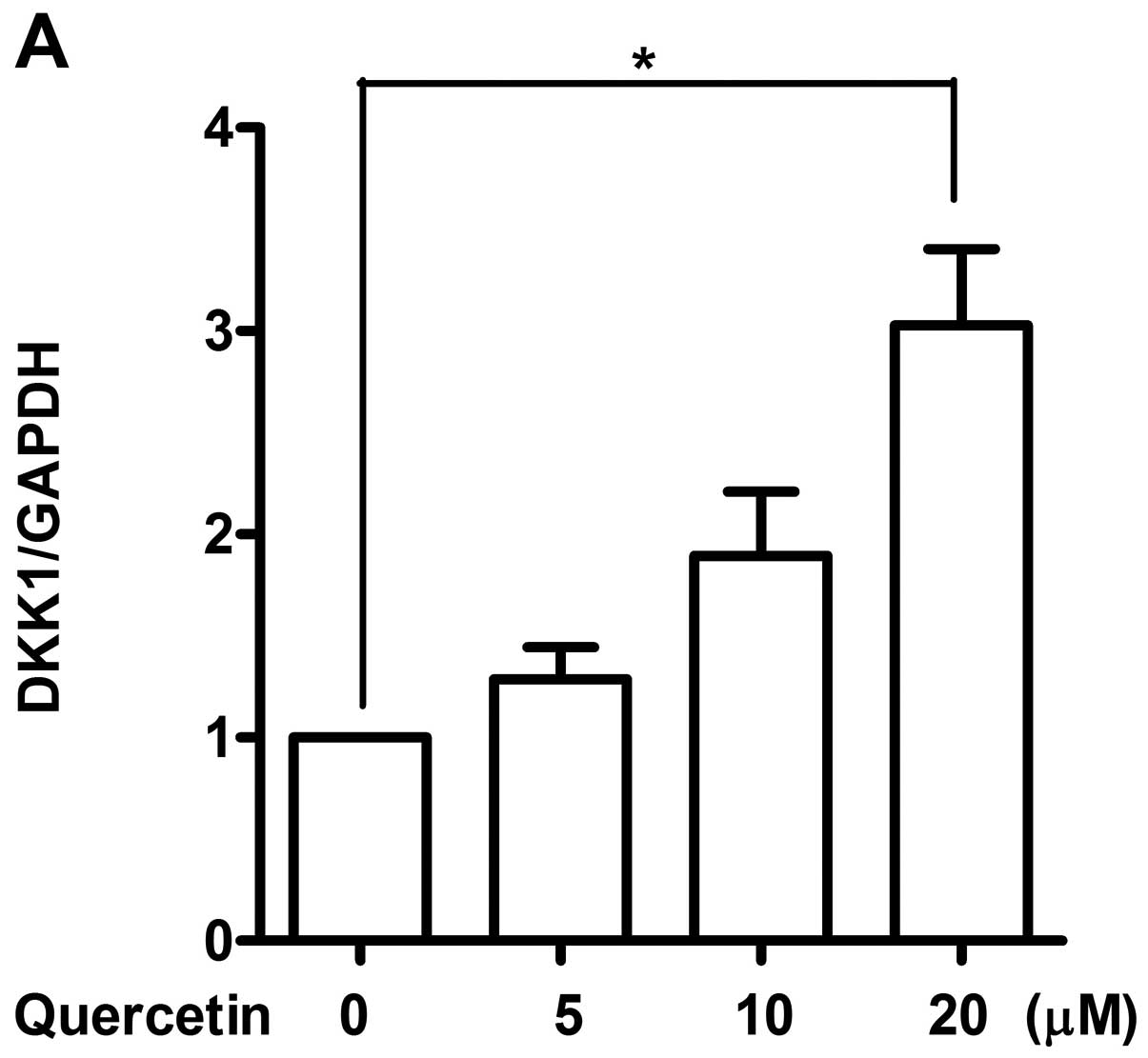
real-time RT-PCR. Among various antagonists screened for
Wnt/β-catenin signaling pathway, mRNA expression level of DKK1, 2,
3 and 4 was found to be elevated dose-dependently (Fig. 3), while any altered expression of
sFRP family was not observed (data not shown). Expression of DKK1
increased more than 3-fold while increase in DKK2 and 3 was about
2-fold. A non-significant induction of DKK4 was observed when
treated with 20 μM of quercetin. Induced expressions of DKK
family by quercetin suggest their involvement in regulating the
Wnt/β-catenin signaling pathway in our system as expected.
Exogenous treatment of DKK1 shows
inhibitory effect on cell growth
Among DKK1, 2, 3 and 4, expression of DKK1 have been
strongly implicated to proliferation ability of breast cancer cells
via regulation of Wnt/β-catenin signaling pathway (34,35).
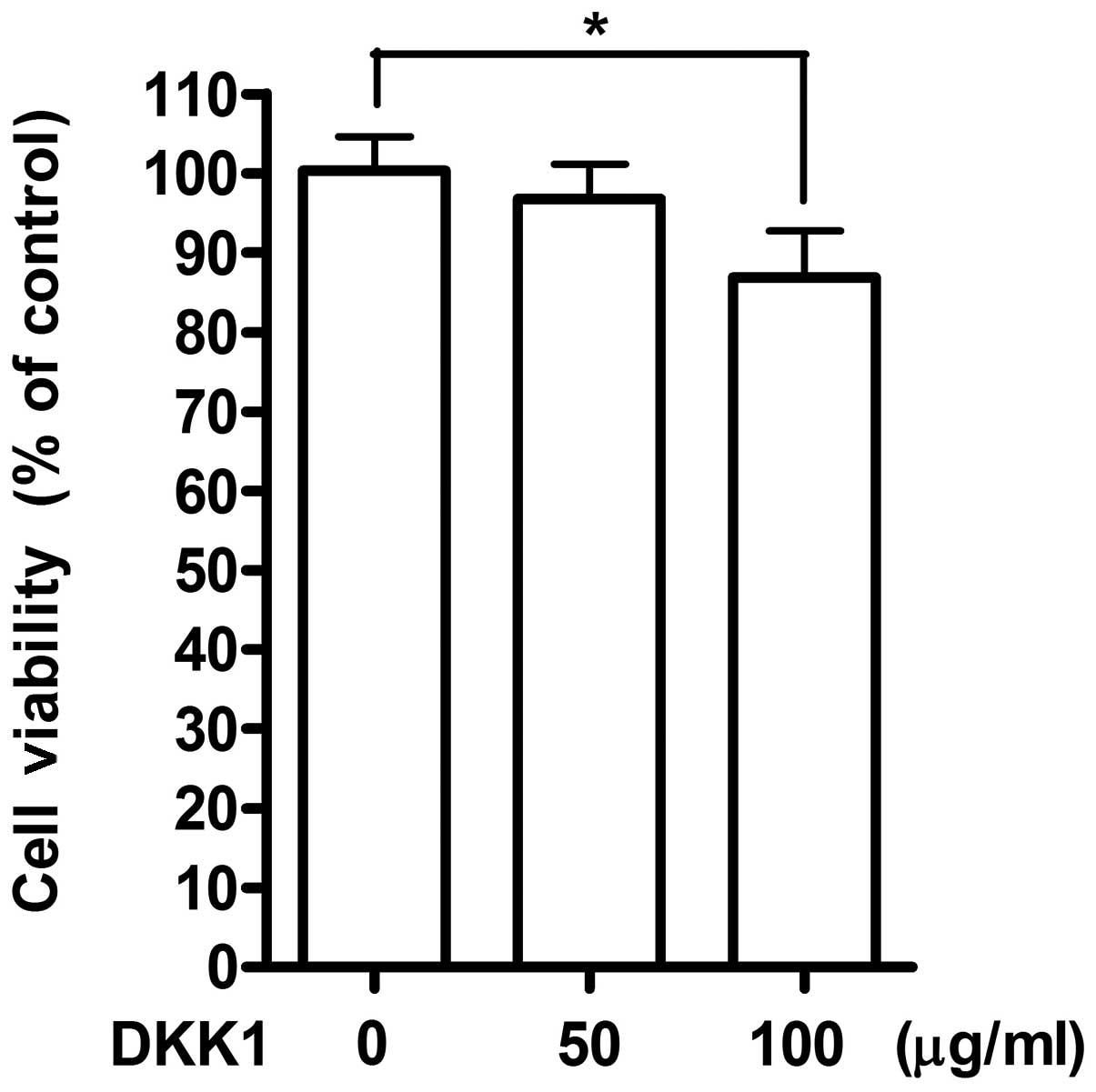
Therefore, to confirm involvement of DKK1 as an antagonist for
Wnt/β-catenin signaling pathway after treatment with quercetin, 4T1
cells were stimulated with recombinant DKK1 protein instead of
quercetin. Stimulation with 100 μg/ml of recombinant DKK1
showed suppressive effect on cell growth as detected by MTT assay
(Fig. 4). The inhibition on cell
growth by DKK1 was partial (∼10%) as compared to the effect of
quercetin, suggesting the possibility of involvement of several
other regulatory mechanisms by quercetin for the inhibition of cell
growth along with DKK1.
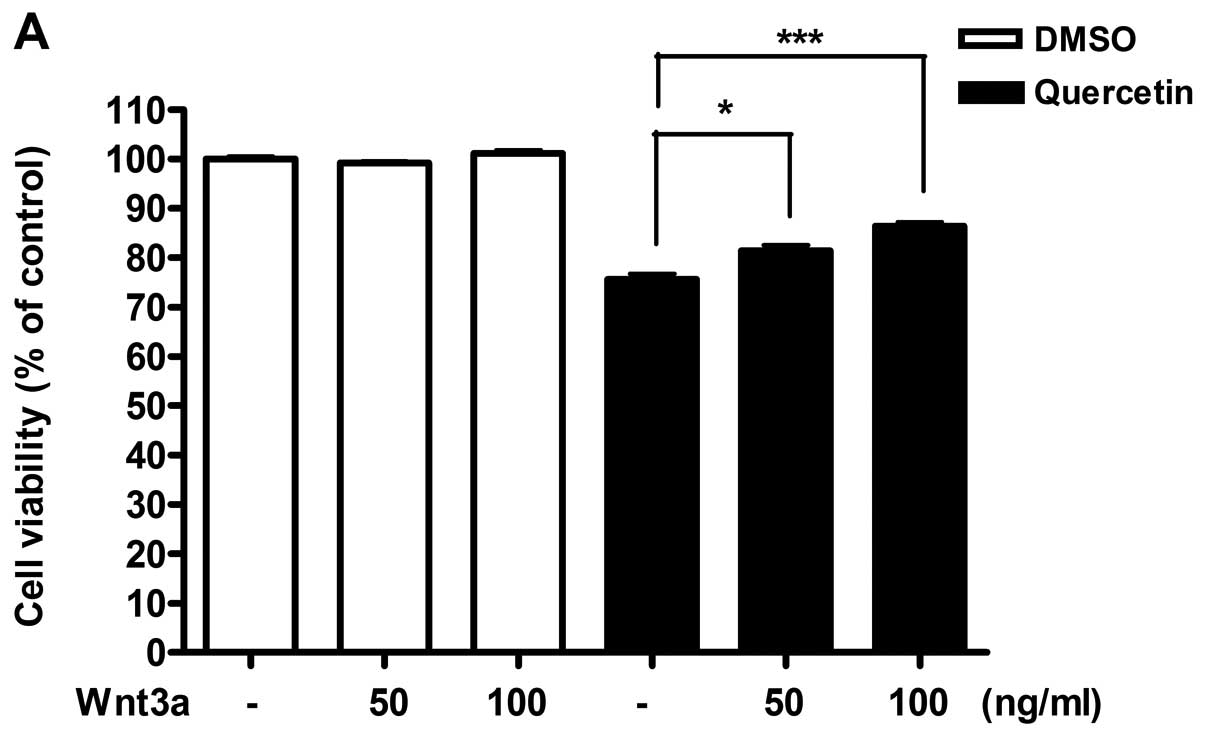
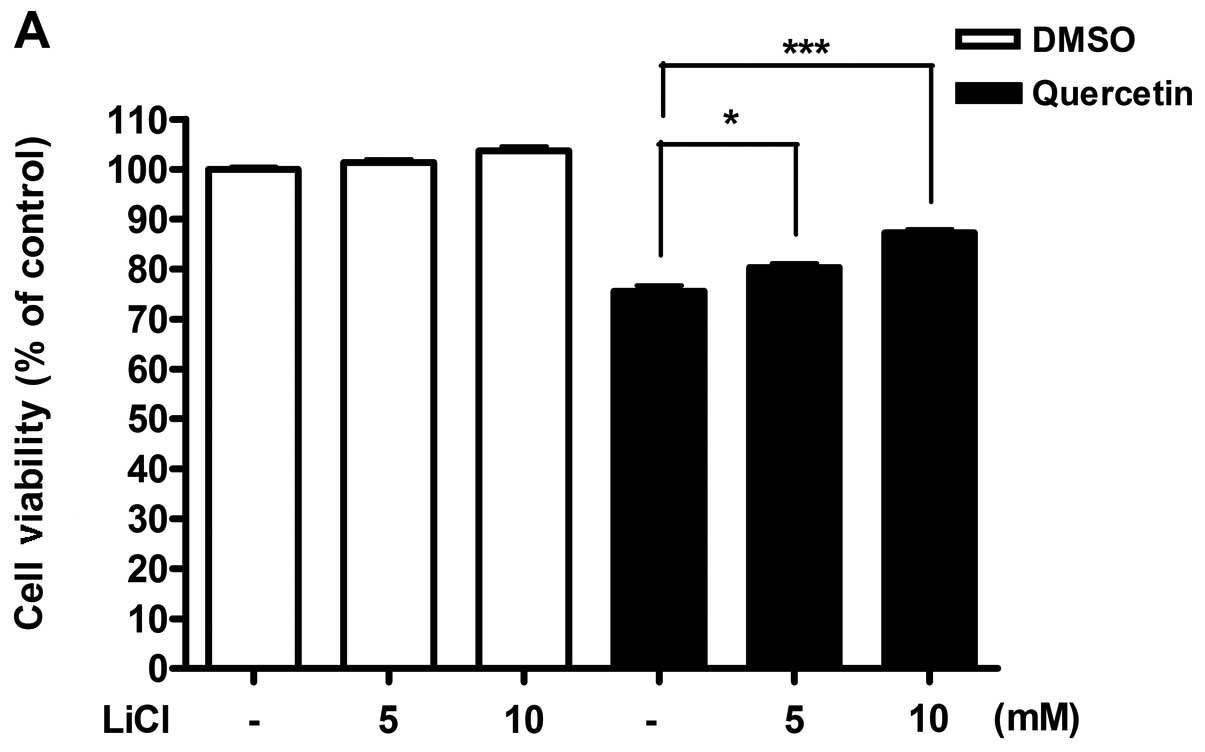
Stimulation of Wnt/β-catenin signaling
pathway with Wnt3a or LiCl restored the effect of quercetin
From results obtained, we hypothesized that
Wnt/β-catenin signaling activity might be reduced by the induction
of the secreted antagonists in response to quercetin. To
investigate this possibility, we potentiated the activity of
Wnt/β-catenin signaling pathway by exogenous stimulation and
observed whether activation of Wnt/β-catenin pathway can
effectively restore the inhibition of cell viability in 4T1 cells
treated with quercetin. 4T1 cells were incubated with quercetin
along with recombinant Wnt3a (Fig.
5), or LiCl (Fig. 6). After 48
h, cell viability was measured by MTT assay and cytotoxicity was
detected by LDH assay. Both stimulators for Wnt/β-catenin signaling
pathway were able to restore the suppressed cell viability by
quercetin in a dose-dependent manner, as well as inhibition of the
increased cytotoxicity (Figs. 5
and 6). However, Wnt3a or LiCl did
not show any individual effect on cell viability and cytotoxicity
in absence of quercetin.
Discussion
The aim of the present study was to investigate the
inhibitory role of quercetin on tumorigenic potential of murine
mammary cancer cell line, 4T1, and to evaluate its ability for
modulating Wnt/β-catenin signaling pathway.
Quercetin is one of the major dietary flavonoids
enriched in various fruits and vegetables. Recent studies report
that quercetin exert a broad range of pharmacological effects and
is a promising chemopreventive agent in a variety of cancer models
(36,37). Previous experimental studies
indicate that quercetin possesses antitumor activity and induces
apoptotic cell death of breast cancer cells by the activation of
caspaseor p53-dependent pathway (38–40).
Consistent with previous reports, our study showed that quercetin
reduces viability and induces apoptosis even in murine mammary
cancer cell line, 4T1 (Fig. 1).
Though quercetin has attracted much attention in relation to its
anticancer activities in many cancer cell models, the molecular
mechanisms underlying quercetin-mediated cellular responses remain
poorly defined.
The Wnt/β-catenin signaling pathway has been
demonstrated to play an important role in tumorigenesis (30). Initially, Wnt/β-catenin signaling
activation, as defined by β-catenin nuclear expression and
overexpression of the cyclin D1, was reported to be associated with
a poorer prognosis in breast cancer patients (27). Recent studies have confirmed the
Wnt/β-catenin signaling in both the pathogenesis of breast cancer
and the regulation of normal mammary epithelial stem cell processes
(41–43). A possible role for Wnt/β-catenin
signaling pathway in cancer was first described three decades ago
in mouse models of mammary cancer and in human and mouse colon
cancer. Moreover, different stages of mammary gland development
have been shown to get controlled and regulated by Wnt/β-catenin
signaling (44). Despite the
cancer preventive potential of quercetin and involvement of
Wnt/β-catenin signaling pathway in cancer progression, little is
known about the modulation of Wnt/β-catenin signaling by quercetin
in breast cancer. According to our results of decreased TopFlash
luciferase activity and β-catenin stability (Fig. 2), we suggest that regulation of
Wnt/β-catenin signaling may be one of the mechanisms responsible
for suppression of cell viability in breast cancer by
quercetin.
Next, we focused on to identify the regulatory
mechanism that could answer the decreased Wnt/β-catenin signaling
activity in breast cancer cells after treatment of quercetin. Among
various possible mechanisms, secretion of antagonists upstream of
Wnt/β-catenin signaling pathway could play a vital role in
suppression of cell viability. Wnt/β-catenin pathway is regulated
by a variety of secreted proteins, such as WIF, DKK and sFRP that
can competitively dislodge certain Wnt ligands from their receptors
or contend for receptor binding. In a few cancer models, an
increase in sFRP levels attenuates cancer growth, particularly in
cells that require autocrine Wnt stimulation such as myeloma cells
(45) and subsets of breast cancer
cells (46). As expected, the
expression level of DKK family was increased after stimulation with
quercetin, and treatment with recombinant DKK1 showed suppressive
effect on cell growth (Figs. 3 and
4). Previous studies have reported
that the downregulation of DKK1 is due to the high proliferation
ability of breast cancer cells via loss of control of β-catenin/TCF
transcription cascade, and that DKK1 secreted by mesenchymal stem
cells inhibits growth of breast cancer cells via depression of
Wnt/β-catenin signaling (34,35).
However, as the inhibitory effect of DKK1 on cell growth was much
less than that of quercetin, the possibility of another mechanism
remains to be studied. We cannot rule out the possible
participation of other antagonists in regulation of Wnt/β-catenin
signaling activity. Also, the relative roles of DKK 2, 3 and 4 in
the suppression of cell viability by quercetin remain to be
elucidated. Besides, our data showed that exogenous stimulation of
recombinant Wnt3a or LiCl for the activation of Wnt/β-catenin
signaling pathway can effectively restore the inhibition of cell
viability by quercetin (Figs. 5
and 6), indicating that the effect
of quercetin in regulation of cell viability of cancer can be
mediated by the modulation of Wnt/β-catenin signaling pathway.
Collectively, our findings provide new insight into
the mechanism of quercetin regulation of Wnt/β-catenin signaling.
Consequently, the inhibition of cell viability and induction of
apoptosis in 4T1 mammary cancer cells with quercetin administration
provide evidence for its potential for cancer treatment. Our
results also highlight the importance of Wnt/β-catenin signaling
regulated by DKK family that may serve as a future target for
therapeutic strategies to inhibit the development of breast cancer
cells.
Acknowledgements
This research was supported by a grant
from Basic Science Research Program through the National Research
Foundation of Korea (NRF) funded by the Ministry of Education,
Science and Technology (2012-0000312 and 2011-0015044), Korean
Health Technology R&D Project, Ministry of Health and Welfare,
Republic of Korea (A121370), and Hallym University Medical Center
Research Fund (01-2011-04).
References
|
1.
|
DeSantis C, Siegel R, Bandi P and Jemal A:
Breast cancer statistics, 2011. CA Cancer J Clin. 61:409–418. 2011.
View Article : Google Scholar
|
|
2.
|
Ren W, Qiao Z, Wang H, Zhu L and Zhang L:
Flavonoids: promising anticancer agents. Med Res Rev. 23:519–534.
2003. View Article : Google Scholar
|
|
3.
|
Baowen Q, Yulin Z, Xin W, et al: A further
investigation concerning correlation between anti-fibrotic effect
of liposomal quercetin and inflammatory cytokines in pulmonary
fibrosis. Eur J Pharmacol. 642:134–139. 2010. View Article : Google Scholar
|
|
4.
|
Boots AW, Haenen GR and Bast A: Health
effects of quercetin: from antioxidant to nutraceutical. Eur J
Pharmacol. 585:325–337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Murakami A, Ashida H and Terao J:
Multitargeted cancer prevention by quercetin. Cancer Lett.
269:315–325. 2008. View Article : Google Scholar
|
|
6.
|
Wei YQ, Zhao X, Kariya Y, Fukata H,
Teshigawara K and Uchida A: Induction of apoptosis by quercetin:
involvement of heat shock protein. Cancer Res. 54:4952–4957.
1994.PubMed/NCBI
|
|
7.
|
Yoshida M, Sakai T, Hosokawa N, et al: The
effect of quercetin on cell cycle progression and growth of human
gastric cancer cells. FEBS Lett. 260:10–13. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Choi EJ, Bae SM and Ahn WS:
Antiproliferative effects of quercetin through cell cycle arrest
and apoptosis in human breast cancer MDA-MB-453 cells. Arch Pharm
Res. 31:1281–1285. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Gulati N, Laudet B, Zohrabian VM, Murali R
and Jhanwar-Uniyal M: The antiproliferative effect of quercetin in
cancer cells is mediated via inhibition of the PI3K-Akt/PKB
pathway. Anticancer Res. 26:1177–1181. 2006.PubMed/NCBI
|
|
10.
|
Lee YK and Park OJ: Regulation of mutual
inhibitory activities between AMPK and Akt with quercetin in MCF-7
breast cancer cells. Oncol Rep. 24:1493–1497. 2010.PubMed/NCBI
|
|
11.
|
Eroles P, Bosch A, Perez-Fidalgo JA and
Lluch A: Molecular biology in breast cancer: intrinsic subtypes and
signaling pathways. Cancer Treat Rev. 38:698–707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Grant S: Cotargeting survival signaling
pathways in cancer. J Clin Invest. 118:3003–3006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Cai J, Guan H, Fang L, et al:
MicroRNA-374a activates Wnt/β-catenin signaling to promote breast
cancer metastasis. J Clin Invest. 123:566–579. 2013.PubMed/NCBI
|
|
14.
|
Lu W, Lin C, King TD, Chen H, Reynolds RC
and Li Y: Silibinin inhibits Wnt/beta-catenin signaling by
suppressing Wnt co-receptor LRP6 expression in human prostate and
breast cancer cells. Cell Signal. 24:2291–2296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Wu Y, Ginther C, Kim J, et al: Expression
of Wnt3 activates Wnt/beta-catenin pathway and promotes EMT-like
phenotype in trastuzumab-resistant HER2-overexpressing breast
cancer cells. Mol Cancer Res. 10:1597–1606. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Tsukamoto AS, Grosschedl R, Guzman RC,
Parslow T and Varmus HE: Expression of the int-1 gene in transgenic
mice is associated with mammary gland hyperplasia and
adenocarcinomas in male and female mice. Cell. 55:619–625. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Ryo A, Nakamura M, Wulf G, Liou YC and Lu
KP: Pin1 regulates turnover and subcellular localization of
beta-catenin by inhibiting its interaction with APC. Nat Cell Biol.
3:793–801. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Huang M, Wang Y, Sun D, et al:
Identification of genes regulated by Wnt/beta-catenin pathway and
involved in apoptosis via microarray analysis. BMC Cancer.
6:2212006. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Li Y, Welm B, Podsypanina K, et al:
Evidence that transgenes encoding components of the Wnt signaling
pathway preferentially induce mammary cancers from progenitor
cells. Proc Natl Acad Sci USA. 100:15853–15858. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Teissedre B, Pinderhughes A, Incassati A,
Hatsell SJ, Hiremath M and Cowin P: MMTV-Wnt1 and
-DeltaN89beta-catenin induce canonical signaling in distinct
progenitors and differentially activate Hedgehog signaling within
mammary tumors. PLoS One. 4:e45372009. View Article : Google Scholar
|
|
22.
|
Khramtsov AI, Khramtsova GF, Tretiakova M,
Huo D, Olopade OI and Goss KH: Wnt/beta-catenin pathway activation
is enriched in basal-like breast cancers and predicts poor outcome.
Am J Pathol. 176:2911–2920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Lin SY, Xia W, Wang JC, et al:
Beta-catenin, a novel prognostic marker for breast cancer: its
roles in cyclin D1 expression and cancer progression. Proc Natl
Acad Sci USA. 97:4262–4266. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Aslakson CJ and Miller FR: Selective
events in the metastatic process defined by analysis of the
sequential dissemination of subpopulations of a mouse mammary
tumor. Cancer Res. 52:1399–1405. 1992.PubMed/NCBI
|
|
25.
|
Yoneda T, Michigami T, Yi B, Williams PJ,
Niewolna M and Hiraga T: Actions of bisphosphonate on bone
metastasis in animal models of breast carcinoma. Cancer.
88:2979–2988. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Choi JA, Kim JY, Lee JY, et al: Induction
of cell cycle arrest and apoptosis in human breast cancer cells by
quercetin. Int J Oncol. 19:837–844. 2001.PubMed/NCBI
|
|
27.
|
Avila MA, Velasco JA, Cansado J and
Notario V: Quercetin mediates the down-regulation of mutant p53 in
the human breast cancer cell line MDA-MB468. Cancer Res.
54:2424–2428. 1994.PubMed/NCBI
|
|
28.
|
Rodgers EH and Grant MH: The effect of the
flavonoids, quercetin, myricetin and epicatechin on the growth and
enzyme activities of MCF7 human breast cancer cells. Chem Biol
Interact. 116:213–228. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Brusselmans K, Vrolix R, Verhoeven G and
Swinnen JV: Induction of cancer cell apoptosis by flavonoids is
associated with their ability to inhibit fatty acid synthase
activity. J Biol Chem. 280:5636–5645. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Brennan KR and Brown AM: Wnt proteins in
mammary development and cancer. J Mammary Gland Biol Neoplasia.
9:119–131. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Giles RH, van Es JH and Clevers H: Caught
up in a Wnt storm: Wnt signaling in cancer. Biochim Biophys Acta.
1653:1–24. 2003.PubMed/NCBI
|
|
32.
|
Suzuki H, Toyota M, Carraway H, et al:
Frequent epigenetic inactivation of Wnt antagonist genes in breast
cancer. Br J Cancer. 98:1147–1156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Bafico A, Liu G, Goldin L, Harris V and
Aaronson SA: An autocrine mechanism for constitutive Wnt pathway
activation in human cancer cells. Cancer Cell. 6:497–506. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Zhou XL, Qin XR, Zhang XD and Ye LH:
Downregulation of Dickkopf-1 is responsible for high proliferation
of breast cancer cells via losing control of Wnt/beta-catenin
signaling. Acta Pharmacol Sin. 31:202–210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Qiao L, Xu ZL, Zhao TJ, Ye LH and Zhang
XD: Dkk-1 secreted by mesenchymal stem cells inhibits growth of
breast cancer cells via depression of Wnt signalling. Cancer Lett.
269:67–77. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Jeong JH, An JY, Kwon YT, Rhee JG and Lee
YJ: Effects of low dose quercetin: cancer cell-specific inhibition
of cell cycle progression. J Cell Biochem. 106:73–82. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Gibellini L, Pinti M, Nasi M, et al:
Quercetin and cancer chemoprevention. Evid Based Complement
Alternat Med. 2011:5913562011. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Seo HS, Ju JH, Jang K and Shin I:
Induction of apoptotic cell death by phytoestrogens by
up-regulating the levels of phosphop53 and p21 in normal and
malignant estrogen receptor alpha-negative breast cells. Nutr Res.
31:139–146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Chou CC, Yang JS, Lu HF, et al:
Quercetin-mediated cell cycle arrest and apoptosis involving
activation of a caspase cascade through the mitochondrial pathway
in human breast cancer MCF-7 cells. Arch Pharm Res. 33:1181–1191.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Chien SY, Wu YC, Chung JG, et al:
Quercetin-induced apoptosis acts through mitochondrial- and
caspase-3-dependent pathways in human breast cancer MDA-MB-231
cells. Hum Exp Toxicol. 28:493–503. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Nusse R: Wnt signaling in disease and in
development. Cell Res. 15:28–32. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Lindvall C, Bu W, Williams BO and Li Y:
Wnt signaling, stem cells, and the cellular origin of breast
cancer. Stem Cell Rev. 3:157–168. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Anastas JN and Moon RT: WNT signalling
pathways as therapeutic targets in cancer. Nat Rev Cancer.
13:11–26. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Prosperi JR and Goss KH: A Wnt-ow of
opportunity: targeting the Wnt/beta-catenin pathway in breast
cancer. Curr Drug Targets. 11:1074–1088. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Chim CS, Pang R, Fung TK, Choi CL and
Liang R: Epigenetic dysregulation of Wnt signaling pathway in
multiple myeloma. Leukemia. 21:2527–2536. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46.
|
Matsuda Y, Schlange T, Oakeley EJ, Boulay
A and Hynes NE: WNT signaling enhances breast cancer cell motility
and blockade of the WNT pathway by sFRP1 suppresses MDA-MB-231
xenograft growth. Breast Cancer Res. 11:R322009. View Article : Google Scholar : PubMed/NCBI
|