Introduction
PN is a sesquiterpene lactone, derived from the
medicinal plant feverfew Tanacetum parthenium, which is
conventionally used to treat migraine and rheumatoid arthritis
(1). In addition it has been shown
that PN exhibits anticancer activity in several cancer cell types,
such as colorectal cancer (2),
pancreatic cancer (3), prostate
cancer (4), multiple myeloma
(5), osteosarcoma and melanoma
(6).
Recently we have demonstrated that PN exerted a
marked cytotoxic effect on MDA-MB-231 cells, a triple-negative
breast cancer cell line (7). This
effect was not prevented by the caspase inhibitor z-VAD-fmk, while
was counteracted by the ROS scavenger NAC. Cytotoxic action was
primarily correlated with generation of ROS, which in turn were
responsible for induction of autophagy, dissipation of Δψm and
necrotic events.
Epidermal growth factor receptor (EGFR), also known
as human epidermal growth factor receptor (HER1 or erbB1), is a
member of a family of tyrosine kinase receptors which includes
three additional receptors: HER2 (erbB2), HER3 (erbB3) and HER4
(erbB4) (8). In response to EGF
binding, EGFR forms homo- or heterodimers with other HER family
members, followed by autophosphorylation of key tyrosine residues
located within the tyrosine kinase domain (8). Once activated, EGFR can promote
proliferation and survival such as migration, invasion,
differentiation, inhibition of apoptosis and angiogenesis (9–11).
EGFR and related family members have been found to
be mutated or amplified in a number of human lung and breast
cancers and this event has been associated with poor prognosis,
short survival and poor response to therapy (12,13).
Data from several research groups have shown that
activation of EGFR can be involved in the production of ROS
(14–16). Moreover, it has been reported that
endogenous hydrogen peroxide (H2O2), produced
as a consequence of EGF binding to EGFR, can be utilized by the
cells as a secondary messenger to regulate physiological signal
transduction (17,18).
The aim of this study was to ascertain whether the
activation of NADPH oxidase (NOX) induced by PN in MDA-MB-231 cells
can be mediated by EGFR activation. The results demonstrate that
treatment of MDA-MB-231 cells with PN increases the level of the
phosphorylated form of EGFR (phospho-EGFR) and that this event is
correlated with the activation of NOX and superoxide
generation.
Materials and methods
Chemicals and reagents
All reagents were purchased from Sigma-Aldrich
(Milan, Italy), except for recombinant human EGF (rhEGF), which was
supplied from Clontech (Palo Alto, CA, USA) and AG1478, supplied
from Calbiochem-Merk Millipore (Milan, Italy). PN, apocynin and
AG1478 were solubilized in dimethyl sulfoxide (DMSO) and diluted to
final concentration in the culture medium. Final concentration of
DMSO employed as vehicle never exceeded 0.04% and had no
discernible effects on MDA-MB-231 cells in comparison with the
control.
Cell cultures
Human breast carcinoma cell line MDA-MB-231 was
provided by ‘Istituto Scientifico Tumori’ (Genoa, Italy). Cells
were grown as monolayer in DMEM medium supplemented with 10% (v/v)
heat inactivated fetal calf serum (FCS), 2 mM glutamine and 1%
non-essential amino acids, at 37°C in a humidified atmosphere
containing 5% CO2. After plating on 96- or 6-well
plates, cells were allowed to adhere overnight and then treated
with chemicals or vehicle only.
Western blot analysis
Whole cell lysates were prepared as previously
reported (19) and protein
concentration was determined by Lowry assay (20). Equal amounts of protein samples (50
μg/lane) were run in an SDS polyacrylamide gel
electrophoresis, then transferred to a nitrocellulose membrane.
Analyses of EGFR and phopho-EGFR were performed using specific
antibodies which were provided by Santa Cruz Biotechnology (Santa
Cruz, CA, USA). In particular, for phopho-EGFR an antibody
recognizing phosphorylated Tyr1173 was used. Then the detection was
developed by using a secondary antibody conjugated with alkaline
phosphatase. Protein bands were visualized using nitroblue
tetrazolium and 5-bromo-4-chloro-3-indoyl-phosphate and their
intensity was quantified by densitometric analysis using SMX Image
software. The correct protein loading was ascertained by red
Ponceau staining and immunoblotting for β-actin. All the blots
shown are representative of at least three separate
experiments.
Measurement of superoxide production
The superoxide radical production was assessed by
dihydroethidium (DHE) staining. DHE is a fluorochrome that is
oxidized by superoxide to fluorescent ethidium that intercalates
with nuclear DNA, staining the nucleus with a bright red
fluorescence. MDA-MB-231 cells (8×103/well) were
incubated with PN or other effectors, then cells were centrifuged
at 120 × g for 5 min and culture medium was replaced with a
solution of DHE (20 μM) in PBS. The incubation was
protracted for 10 min and DHE-positive cells were analysed by
fluorescence microscopy using an excitation wavelength of 596 nm
and an emission wavelength of 620 nm. All the images were acquired
by Leica Q Fluoro Software. Percentage of DHE-positive cells was
evaluated by using ImageJ software.
Statistical analysis
Results are presented as means ± standard error.
Data were analysed using Student’s t-test.
Results
PN stimulates EGFR phosphorylation
In the present study we evaluated whether the
exposure of MDA-MB-231 cells to PN resulted in the phosphorylation
and activation of EGFR and consequently in the induction of NOX
activity.
It is well known that activation of EGFR depends on
phosphorylation of the amino acid residue Tyr1173 (21). Therefore, MDA-MB-231 cells were
treated with 25 μM PN for different times and subjected to
western blot analysis to evaluate the level of phospho-EGFR by
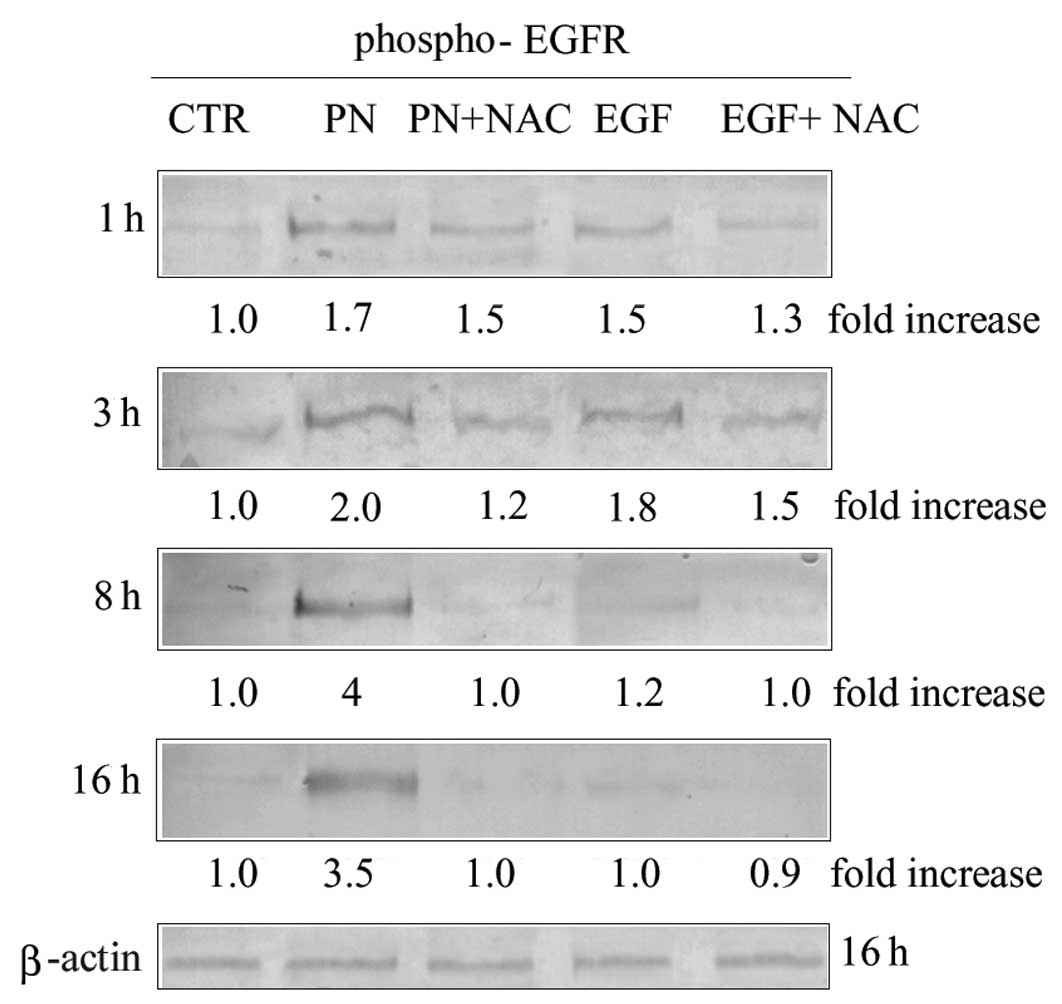
using an antibody recognizing phospho-Tyr1173. As shown in Fig. 1, the level of phospho-EGFR was
increased already at 1 h of treatment with PN. The effect was
enhanced with time reaching the maximum at about 8 h of treatment,
and remaining elevated up to 16 h. Increment of EGFR
phosphorylation was prevented by 2 mM NAC, whatever the time of
treatment was (Fig. 1), thus
suggesting that the effect was a consequence of oxidative stress
induction.
The effects of PN on MDA-MB-231 cells were compared
with that exerted by recombinant human EGF (rhEGF), a direct EGFR
ligand. Treatment of cells with 200 ng/ml rhEGF caused
phosphorylation of EGFR with a maximum effect found between 1 and 3
h. Such an effect was much lower than that observed with PN and was
only partially reduced by the addition of 2 mM NAC (Fig. 1). Finally treatment with rhEGF, in
contrast with PN, did not exert any cytotoxic effect (not
shown).
It is noteworthy that treatment of MDA-MB-231 cells
with PN or rhEGF did not modify the level of the EGFR (not shown).
Therefore, after PN or rhEGF treatment the amount of EGFR remained
unchanged, whereas its tyrosine phosphorylation increased.
PN induces production of superoxide
anion
ROS include superoxide anion
(O2.−), which is a highly reactive oxygen
species and hydrogen peroxide (H2O2). High
doses of ROS, in particular superoxide anion, result in oxidative
stress, which is implicated in the pathogenesis of a number of
diseases and in the cytotoxic action of many drugs (22). NADPH oxidases (NOXs), which are
present in phagocytes, but also in other somatic cells, are
multicomponent enzymes that catalyze the generation of superoxide
anion from oxygen and NADPH (23).
NOX4 has been identified as the principal source of superoxide
production in MDA-MB-231 cells (24).
In order to ascertain whether PN stimulated NOX
activation in MDA-MB-231 cells, we evaluated the effect of the drug
on superoxide production by using DHE, a specific fluorescent
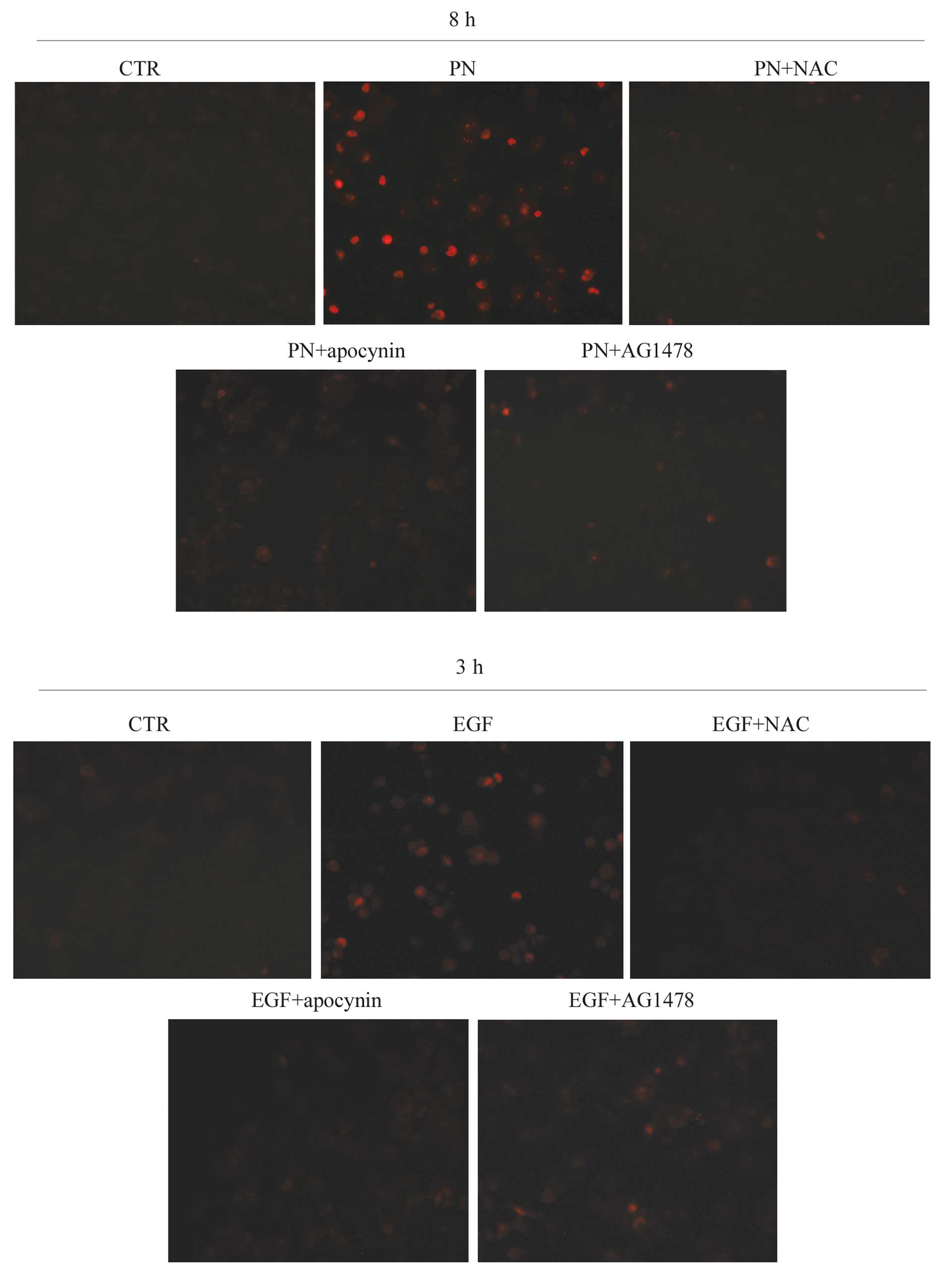
probe. It was observed that PN induced the appearance of intense
red fluorescence, suggesting that the drug induced production of
superoxide anion by stimulating NOX activity (Fig. 2). Such an effect was abrogated by
either NAC or apocynin, a specific inhibitor of NOX activity
(25).
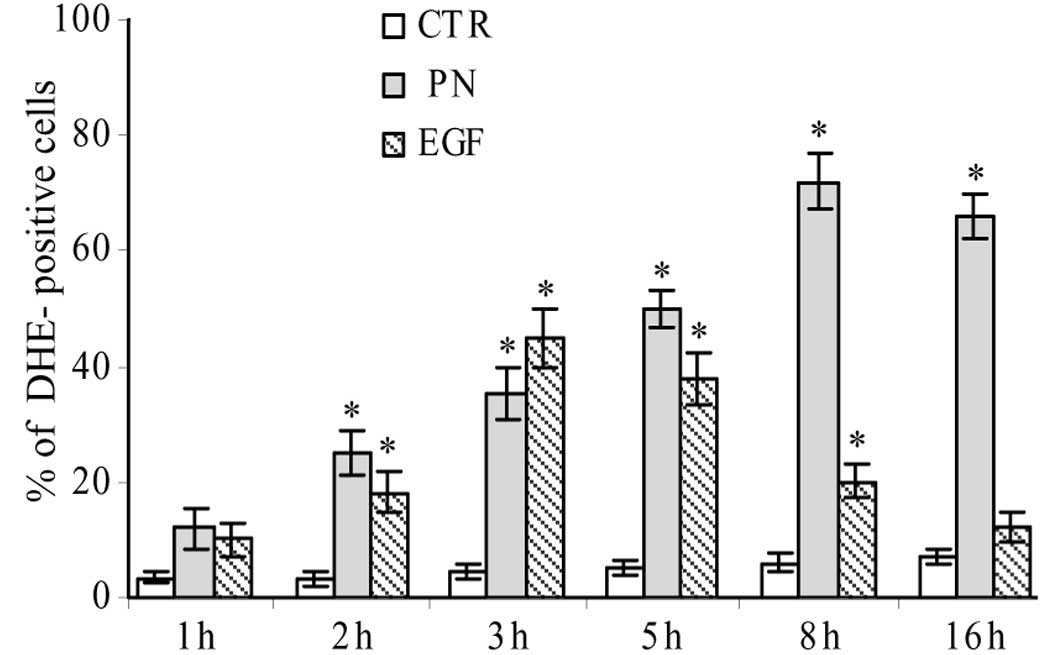
A time course study showed that the production of
superoxide anion increased already after 1 h of treatment with 25
μM PN, reached the maximum at 8 h, when 72% of cells showed
red fluorescence, and maintained a considerable level until 16 h
(Fig. 3).
We also evaluated whether rhEGF treatment induced
the production of superoxide anion. As shown in Fig. 2, the effect was observed
particularly at 3 h of treatment and was lower than that found in
PN-treated MDA-MB-231 cells.
Production of superoxide anion induced by
PN is correlated with EGFR activation
In order to ascertain a possible correlation between
superoxide anion production and EGFR activation induced by PN
treatment, we performed experiments using AG1478, which is a
specific inhibitor of the kinase activity of EGFR (26).
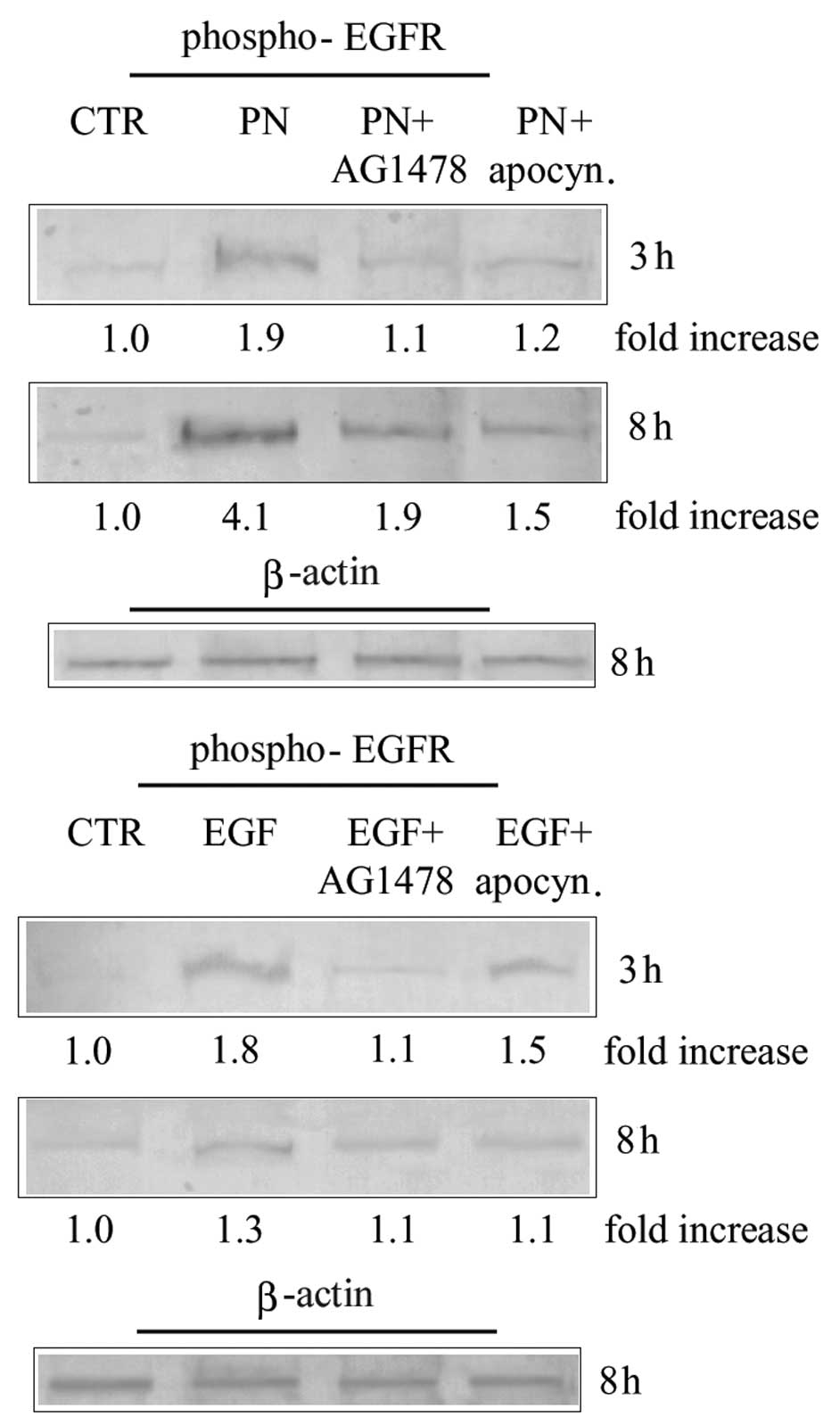
MDA-MB-231 cells were treated for 3 and 8 h with 25
μM PN in the presence or absence of 20 μM AG1478. In
order to confirm the specificity of the inhibitory action exerted
by AG1478, PN was substituted in some samples with 200 ng/ml rhEGF.
Results reported in Fig. 4 show
that the increase in the level of phospho-EGFR induced by both PN
and rhEGF was prevented by AG1478. Then we evaluated the effect of
AG1478 on the production of superoxide anion in MDA-MB-231 cells
stimulated with PN or rhEGF. Results shown in Fig. 2 demonstrate that the addition of
AG1478 to PN or rhEGF prevented, after incubation with DHE, the
production of cells with red fluorescence. These results suggest
that phosphorylation and activation of EGFR induced by PN were
closely correlated with stimulation of NOX and production of
superoxide anion.
Discussion
EGFR is a member of the ErbB family of tyrosine
kinases, which is involved in the regulation of proliferation and
differentiation primarily of epithelial cell types (27). Following ligand binding, the EGFR
dimerizes and the intracellular tyrosine kinase region is
activated, causing receptor tyrosine autophosphorylation and
transphosphorylation of another receptor monomer (27). These events lead to the recruitment
and phosphorylation of several intracellular substrates and the
subsequent transmission of extracellular signals to the nucleus via
an intracellular signaling network (27).
EGFR activation plays important roles in the
stimulation of cell growth, survival, migration and differentiation
(9,11). EGFR and related family members have
been found to be mutated or amplified in a number of human lung and
breast cancers (12,13,28,29).
Our study was performed on MDA-MB-231 cells, a ‘triple-negative’
breast cancer cell line which lacks estrogen receptor α (ERα),
progesterone receptor and HER2 (29). Instead MDA-MB-231 cells are known
to contain high levels of EGFR (HER1) (29,30).
This observation has been correlated with the absence in these
cells of ERα, because an inverse relationship has been demonstrated
between the presence of ERα and the level of EGFR (31).
In the present study we provide evidence that in
MDA-MB-231 cells PN increased the level of the phosphorylated form
of EGFR. This effect was remarkable between 8 and 16 h of
treatment. Furthermore, the production of phospho-EGFR was
prevented by NAC, a general ROS scavenger. Such a result suggests
that PN effect was mediated by ROS generation. This conclusion
agrees with our previous finding (7) that in MDA-MB-231 cells PN strongly
induced production of ROS already in the first hours of treatment
(1–3 h).
It has been shown also previously that a correlation
exists between enhancement of ROS and phosphorylation of EGFR
(32), and suggested that ROS
could be responsible for the reversible oxidization of thiol groups
of the catalytic site of protein phosphatases, involved in
dephosphorylation of EGFR (33,34).
As a consequence phosphatases are inactivated and EGFR is
maintained in the phosphorylated active form (34). However, other authors demonstrated
that H2O2 directly can modify a specific
cysteine residue (Cys797) of EGFR to sulfenic acid in the active
site of the receptor, enhancing tyrosine kinase activity of EGFR
(35).
Our results also show that PN increased the
production of anion superoxide, reaching very high levels between 8
and 16 h. The observation that the production of superoxide anion
induced by PN was prevented by the addition of apocynin, an
inhibitor of NOX, suggests the involvement of NOX in the production
of superoxide.
Our study provides evidence that addition of AG1478,
a specific inhibitor of EGFR tyrosine kinase, was capable of
inhibiting the effects of PN both on EGFR phosphorylation and on
the production of superoxide anion. Therefore it seems that
phosphorylation of EGFR and activation of NOX are closely
correlated. Such a correlation has been suggested by many other
authors. In particular Tamàs et al (36) have shown that EGFR activates Rac1,
a member of Rac GTPase involved in activation of the non-phagocytic
NADPH oxidases (23), by
phosphorylating and associating with Vav2 factor. Moreover, Sheng
et al (37) reported that
ofloxacin caused intracellular ROS production by activating
EGFR-Rac1-NOX2 pathway.
Collectively, our results led us to the following
conclusions: PN generates already in the first phase of treatment
(1–3 h) production of ROS; ROS activate phosphorylation of EGFR and
this causes stimulation of NOX and new production of ROS. These
considerations suggest that the mechanism induced by PN is based on
the creation of a positive feedback loop, which through the
involvement of the EGFR sustains for a long period of time (16 h)
activation of NOX and production of ROS.
Abbreviations:
|
DHE
|
dihydroethidium
|
|
EGF
|
epidermal growth factor
|
|
rhEGF
|
recombinant human EGF
|
|
EGFR
|
epidermal growth factor receptor
|
|
ERα
|
estrogen receptor α
|
|
HER
|
human epidermal growth factor
receptor
|
|
NAC
|
N-acetylcysteine
|
|
NOX
|
NADPH oxidase
|
|
PN
|
parthenolide
|
|
PR
|
progesterone receptor
|
|
ROS
|
reactive oxygen species
|
|
TNBC
|
triple-negative breast cancer
|
Acknowledgements
This study was supported by grants
from: Italian Ministry of Education, University and Research (MIUR)
ex-60%, 2007, and partially funded by European Regional Development
Fund, European Territorial Cooperation 2007–2013, CCI 2007 CB 163
PO 037, OP Italia-Malta 2007–2013. Dr D. Carlisi is a recipient of
a grant by ‘Italian Ministry of Education, University and Research’
(MIUR). Dr G. Buttitta is a PhD student supported by ‘Italian
Ministry of Education, University and Research’ (MIUR).
References
|
1.
|
Ghantous A, Sinjab A, Herceg Z and
Darwiche N: Parthenolide: from plant shoots to cancer roots. Drug
Discov Today. 18:894–905. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Kim SL, Trang KT, Kim SH, Kim IH, Lee SO,
Lee ST, Kim DG and Kim SW: Parthenolide suppresses tumor growth in
a xenograft model of colorectal cancer cells by inducing
mitochondrial dysfunction and apoptosis. Int J Oncol. 41:1547–1553.
2012.PubMed/NCBI
|
|
3.
|
Yip-Schneider MT, Nakshatri H, Sweeney CJ,
Marshall MS, Wiebke EA and Schmidt CM: Parthenolide and sulindac
cooperate to mediate growth suppression and inhibit the nuclear
factor-kappa B pathway in pancreatic carcinoma cells. Mol Cancer
Ther. 4:587–594. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Sun Y, St Clair DK, Xu Y, Crooks PA and St
Clair WH: A NADPH oxidase-dependent redox signaling pathway
mediates the selective radiosensitization effect of parthenolide in
prostate cancer cells. Cancer Res. 70:2880–2890. 2010. View Article : Google Scholar
|
|
5.
|
Wang W, Adachi M, Kawamura R, Sakamoto H,
Hayashi T, Ishida T, Imai K and Shinomura Y: Parthenolide-induced
apoptosis in multiple myeloma cells involves reactive oxygen
species generation and cell sensitivity depends on catalase
activity. Apoptosis. 11:2225–2235. 2006. View Article : Google Scholar
|
|
6.
|
D’Anneo A, Carlisi D, Lauricella M,
Emanuele S, Di Fiore R, Vento R and Tesoriere G: Parthenolide
induces caspase-independent and AIF-mediated cell death in human
osteosarcoma and melanoma cells. J Cell Physiol. 228:952–967.
2013.PubMed/NCBI
|
|
7.
|
D’Anneo A, Carlisi D, Lauricella M, Puleio
R, Martinez R, Di Bella S, Di Marco P, Emanuele S, Di Fiore R,
Guercio A, Vento R and Tesoriere G: Parthenolide generates reactive
oxygen species and autophagy in MDA-MB-231 cells A soluble
parthenolide analogue inhibits tumour growth and metastasis in a
xenograft model of breast cancer. Cell Death Dis. (In press).
|
|
8.
|
Bazley LA and Gullick WJ: The epidermal
growth factor receptor family. Endocr Relat Cancer. 12:17–27. 2005.
View Article : Google Scholar
|
|
9.
|
Hölsken A, Gebhardt M, Buchfelder M,
Fahlbusch R, Blümcke I and Buslei R: EGFR signaling regulates tumor
cell migration in craniopharyngiomas. Clin Cancer Res.
17:4367–4377. 2011.PubMed/NCBI
|
|
10.
|
Zhang J, Li H, Wang J, Dong Z, Mian S and
Yu FS: Role of EGFR transactivation in preventing apoptosis in
Pseudomonas aeruginosa-infected human corneal epithelial
cells. Invest Ophthalmol Vis Sci. 45:2569–2576. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
van Cruijsen H, Giaccone G and Hoekman K:
Epidermal growth factor receptor and angiogenesis: opportunities
for combined anticancer strategies. Int J Cancer. 118:883–888.
2006.PubMed/NCBI
|
|
12.
|
Macias A, Azavedo E, Hagerstrom T,
Klintenberg C, Perez R and Skoog L: Prognostic significance of the
receptor for epidermal growth factor in human mammary carcinomas.
Anticancer Res. 7:459–464. 1987.PubMed/NCBI
|
|
13.
|
Sequist LV, Joshi VA, Jänne PA, Muzikansky
A, Fidias P, Meyerson M, Haber DA, Kucherlapati R, Johnson BE and
Lynch TJ: Response to treatment and survival of patients with
non-small cell lung cancer undergoing somatic EGFR mutation
testing. Oncologist. 12:90–98. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Bae YS, Kang SW, Seo MS, Baines IC, Tekle
E, Chock PB and Rhee SG: Epidermal growth factor (EGF)-induced
generation of hydrogen peroxide. Role in EGF receptor-mediated
tyrosine phosphorylation. J Biol Chem. 272:217–221. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Cuddihy SL, Winterbourn CC and Hampton MB:
Assessment of redox changes to hydrogen peroxide-sensitive proteins
during EGF signaling. Antioxid Redox Signal. 15:167–174. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Truong TH and Carroll KS: Redox regulation
of epidermal growth factor receptor signaling through cysteine
oxidation. Biochemistry. 51:9954–9965. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Chiarugi P, Pani G, Giannoni E, Taddei L,
Colavitti R, Raugei G, Symons M, Borrello S, Galeotti T and Ramponi
G: Reactive oxygen species as essential mediators of cell adhesion:
the oxidative inhibition of a FAK tyrosine phosphatase is required
for cell adhesion. J Cell Biol. 161:933–944. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Jagadeesha DK, Takapoo M, Banfi B, Bhalla
RC and Miller FJ Jr: Nox1 transactivation of epidermal growth
factor receptor promotes N-cadherin shedding and smooth muscle cell
migration. Cardiovasc Res. 93:406–413. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Giuliano M, Lauricella M, Calvaruso G,
Carabillò M, Emanuele S, Vento R and Tesoriere G: The apoptotic
effects and synergistic interaction of sodium butyrate and MG132 in
human retinoblastoma Y79 cells. Cancer Res. 59:5586–5595.
1999.PubMed/NCBI
|
|
20.
|
Lowry OH, Rosebrough NJ, Farr AL and
Randall RJ: Protein measurement with the Folin phenol reagent. J
Biol Chem. 193:265–275. 1951.PubMed/NCBI
|
|
21.
|
Voldborg BR, Damstrup L, Spang-Thomsen M
and Poulsen HS: Epidermal growth factor receptor (EGFR) and EGFR
mutations, function and possible role in clinical trials. Ann
Oncol. 8:1197–1206. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Paletta-Silva R, Rocco-Machado N and
Meyer-Fernandes JR: NADPH oxidase biology and the regulation of
tyrosine kinase receptor signaling and cancer drug cytotoxicity.
Int J Mol Sci. 14:3683–3704. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Hordijk PL: Regulation of NADPH oxidases:
the role of Rac proteins. Circ Res. 98:453–462. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Boudreau HE, Casterline BW, Rada B,
Korzeniowska A and Leto TL: Nox4 involvement in TGF-beta and
SMAD3-driven induction of the epithelial-to-mesenchymal transition
and migration of breast epithelial cells. Free Radic Biol Med.
53:1489–1499. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Wu F, Tyml K and Wilson JX: iNOS
expression requires NADPH oxidase-dependent redox signaling in
microvascular endothelial cells. J Cell Physiol. 117:207–214. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Zhua XF, Liua ZC, Xiea BF, Lia ZM, Fenga
GK, Yangb D and Zeng YX: EGFR tyrosine kinase inhibitor AG1478
inhibits cell proliferation and arrests cell cycle in
nasopharyngeal carcinoma cells. Cancer Lett. 169:27–32. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Schlessinger J: Ligand-induced,
receptor-mediated dimerization and activation of EGF receptor.
Cell. 110:669–672. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Kosaka T, Yatabe Y, Endoh H, Kuwano H,
Takahashi T and Mitsudomi T: Mutations of epidermal growth factor
receptor gene in lung cancer: biological and clinical implications.
Cancer Res. 64:8919–8923. 2004. View Article : Google Scholar
|
|
29.
|
Rampaul RS, Pinder SE, Nicholson RI,
Gullick WJ, Robertson JF and Ellis IO: Clinical value of epidermal
growth factor receptor expression in primary breast cancer. Adv
Anat Pathol. 12:271–273. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Lauricella M, Ciraolo A, Carlisi D, Vento
R and Tesoriere G: SAHA/TRAIL combination induces detachment and
anoikis of MDA-MB231 and MCF-7 breast cancer cells. Biochimie.
94:287–299. 2012. View Article : Google Scholar
|
|
31.
|
Lee CS, Hall RE, Alexander IE, Koga M,
Shine J and Sutherland RL: Inverse relationship between estrogen
receptor and epidermal growth factor. Growth Factors. 3:97–103.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Gamou S and Shimizu N: Hydrogen peroxide
preferentially enhances the tyrosine phosphorylation of epidermal
growth factor receptor. FEBS Lett. 357:161–164. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Haj FG, Markova B, Klaman LD, Bohmer FD
and Neel BG: Regulation of receptor tyrosine kinase signaling by
protein tyrosine phosphatase-1B. J Biol Chem. 278:739–744. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Chiarugi P: PTPs versus PTKs: the redox
side of the coin. Free Radic Res. 39:353–364. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Paulsen CE, Truong TH, Garcia FJ, Homann
A, Gupta V, Leonard SE and Carroll KS: Peroxide-dependent
sulfenylation of the EGFR catalytic site enhances kinase activity.
Nat Chem Biol. 8:57–64. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Tamas P, Solti Z, Bauer P, Illes A, Sipeki
S, Bauer A, Farago A, Downward J and Buday L: Mechanism of
epidermal growth factor regulation of Vav2, a guanine nucleotide
exchange factor for Rac. J Biol Chem. 278:5163–5171. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Sheng ZG, Huang W, Liu YX, Yuan Y and Zhu
BZ: Ofloxacin induces apoptosis via β1 integrin-EGFR-Rac1-Nox2
pathway in microencapsulated chondrocytes. Toxicol Appl Pharmacol.
267:74–87. 2013.PubMed/NCBI
|