Introduction
The essential characteristics of cancer are the
ability to invade surrounding tissues and metastasize to distal
tissues, known as cancer metastasis, which is the major cause of
mortality in cancer patients. In non-small cell lung cancer (NSCLC)
patients, the median overall survival of patients with metastatic
lung cancer (stage IIIb or IV in the TNM classification) is limited
(only 8 or 5 months, respectively) (1); therefore, suppression of cancer
metastasis results in the improved survival of lung cancer
patients.
Cancer metastasis consists of several steps:
intravasation, attachment to a vessel, extravasation, angiogenesis
and growth in distal tissues (2–5).
Among them, epithelial-to-mesenchymal transition (EMT) is involved
in an early step of metastasis (6,7). EMT
is a phenomenon in which cobblestone-like epithelial cells change
into spindle-like mesenchymal cells with down-regulation of
epithelial markers such as E-cadherin and also upregulation of
mesenchymal markers such as N-cadherin (8). In addition to metastasis, this
physiological phenomenon is important for resistance to apoptosis,
maintenance of cancer stem cells and production of extracellular
matrix. Thus, EMT would be an attractive therapeutic target in
metastatic cancers.
Juzentaihoto, which is a Japanese Kampo
medicine, has been widely used for the decline of physical
strength, general debility, cold hands and feet, fatigue, night
sweats, circulatory problems and anemia (9). Interestingly, our previous studies
showed that Juzentaihoto indirectly inhibited cancer
metastasis through the activation of macrophages and T cells in
mouse models (10–12). Although Juzentaihoto could
suppress cancer metastasis, it is not well known how
Juzentaihoto can directly affect tumor cells during the
metastasis process and which herbal ingredients of
Juzentaihoto are involved in the regulation of EMT.
Here we firstly report that Cinnamomi Cortex (CC),
one of the herbal medicines from Juzentaihoto, inhibits
TGF-β-induced EMT phenotypes. After fractionation of CC extract,
the content of catechin trimer was well-associated with the
inhibitory activities of CC extract. Finally, procyanidin C1 from
CC extract is newly identified as the responsible molecule for the
EMT inhibition.
Materials and methods
Herbal medicine extraction
Astagali Radix (AsR), Glycyrrhizae Radix (GlR),
Cinnamomi Cortex (CC), Rehmanniae Radix (RR), Paeoniae Radix (PR),
Cnidii Rhizoma (CR), Angelicae Radix (AnR), Ginseng Radix (GiR),
Hoelen (Ho), and Atractylodis Lanceae Radix (ALR) were purchased
from Uchida-Wakanyaku Co., Ltd. (Tokyo, Japan). In this study,
Cinnamomi Cortex was added to an appropriate volume of distilled
water (w/v, 1:10) and extracted at 100°C for 1 h. The extracted
solution was filtered and then freeze dried to obtain dried powder.
A voucher sample of this extract (INM 10000007, University of
Toyama) was preserved in the Research Promotion Office, Institute
of Natural Medicine, University of Toyama, Toyama, Japan. Six other
Cinnamomi Cortex samples were collected from different regions of
Vietnam and China, coded as CC-1 (Vietnam), CC-2 (Guang Xi, China),
CC-3 (Guang Xi, China), CC-4 (Guang Dong, China), CC-5 (Guang Xi,
China), CC-6 (Vietnam). Water extracts of the above-mentioned six
CC samples were prepared and provided by the National Institute of
Biomedical Innovation, Osaka, Japan, and a part of each extract was
deposited at our institute (voucher specimen no. INM
10000001–10000006).
Cell cultures
Human lung adenocarcinoma A549 cells were cultured
in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) with 10% fetal
bovine serum (FBS; ICN Biomedicals, Aurora, OH, USA), 2 mM
L-glutamine (Invitrogen), 100 U/ml penicillin and 100 μg/ml
streptomycin in 5% CO2 at 37°C. The cells were treated
with recombinant human TGF-β (5 ng/ml) (Peprotech, London, UK) for
various times as indicated, after pretreatment with TGF-β receptor
I kinase inhibitor (10 μM) (Merck, Whitehouse, NJ, USA),
each herbal component from Juzentaihoto, or Cinnamomi Cortex
extracts (CC, or CC-1 to CC-6) for 30 min.
Protein preparation and western
blotting
Whole cell lysates were collected in lysis buffer
supplemented with some protease and phosphatase inhibitors as
described previously (13). Equal
amounts of protein were resolved by electrophoresis on acrylamide
gels and transferred to PVDF membranes. Antibodies against
phospho-specific Smad-2 (Ser 465/467), E-cadherin, and N-cadherin
were purchased from Cell Signaling Technology (Beverly, MA, USA)
and an antibody against PCNA was purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA).
Cell morphology and cell migration
Cell morphology was determined by BZ-8000 (Keyence,
Osaka, Japan) after staining with hematoxylin and eosin. For the
migration assay, Transwell cell culture chambers were used as
described previously (14).
Briefly, the filters were precoated with 1.25 μg fibronectin
on the lower surfaces. The cell suspension (3×104
cells/100 μl) in serum-free medium was added to the upper
compartment and incubated for 6 h. The migrated cells were stained
with hematoxylin and eosin, and counted under the microscope in
three predetermined fields at a magnification of ×400.
Fractionation of CC and identification of
procyanidin C1 from CC
CC extract, 2.0 g, was subjected to reversed-phase
silica gel (Cosmosil 75C18-OPN; Nacalai Tesque Inc.,
Kyoto, Japan) using medium pressure liquid chromatography (MPLC;
Buchi, Flawil, Switzerland) with a H2O-CH3CN
gradient system (98:2→96:4→92:8→90:10→80:20→60:40→40:60 →10:90) to
obtain eight fractions (fr. 1, 435 mg; fr. 2, 282 mg; fr. 3, 123
mg; fr. 4, 212 mg; fr. 5, 105 mg; fr. 6, 305 mg; fr. 7, 205 mg; fr.
8, 105 mg). The bioactive fraction 4 (fr. 4, 200 mg) was further
subjected to preparative HPLC (Discovery C18 column; 10×250 mm
i.d., 5 μm particle size; Supelco, PA, USA) with
H2O-CH3CN (92:8) containing 0.01%
trifluoroacetic acid (TFA) at a flow rate of 2 ml/min to yield a
procyanidin trimer (1, 5.5 mg, tR 23.3
min). The molecular formula of compound 1 was determined by
HR-TOF-MS to be C45H38O18
[m/z 865.2003 (M - H)+]. Its chemical structure
was further identified to be
epicatechin-(4β→8)-epicatechin-(4β→8)-epicatechin
(procyanidin C1, Fig. 3C) by
comparing the [1H] nuclear magnetic resonance (NMR),
[13C] NMR, and circular dichroism (CD) spectral data
with those in the literature (15).
Liquid chromatography-mass spectrometry
(LC-MS) analysis
For chemical profiling of 7 CC extracts, liquid
chromatography-mass spectrometry (LC-MS) analysis was performed
with a Shimadzu LC-IT-TOF mass spectrometer (Kyoto, Japan) equipped
with an ESI interface (Shimadzu). The ESI parameters were as
follows: source voltage +4.5 kV, capillary temperature 200°C and
nebulizer gas 1.5 l/min. The mass spectrometer was operated in
positive ion mode scanning from m/z 200 to 2,000. A Waters
Atlantis T3 column (2.1 mm i.d. × 150 mm, 3 m; Milford, MA, USA)
was used and the column temperature was maintained at 40°C. The
mobile phase was a binary eluent of (A) 5 mM ammonium acetate
solution and (B) CH3CN under the following gradient
conditions: 0–30 min linear gradient from 10 to 100% B, 30–40 min
isocratic at 100% B. The flow rate was 0.15 ml/min. Mass
spectrometry data obtained from the extract were deposited in the
MassBank Database and stored with pharmacological information on
the extract in the Wakan-Yaku Database System, Institute of Natural
Medicine, University of Toyama.
Results
Cinnamomi Cortex (CC) extract suppresses
TGF-β-induced EMT
Our previous studies showed that
Juzentaihoto, a Japanese Kampo medicine, prevents metastasis
in mouse models (10–12). Because Juzentaihoto contains
ten (‘Ju’ means ‘ten’ in Japanese) kinds of herbal ingredients, we
thus investigated whether each component can suppress TGF-β-induced
EMT. We firstly screened the expression of an epithelial marker,
E-cadherin, in a human non-small-cell lung cancer cell line, A549
cells, treated with TGF-β, TGF-β receptor kinase inhibitor (TGFRi),
or herbal medicines (Fig. 1A).
Known as EMT, the reduction of E-cadherin expression in A549 cells
was detected after TGF-β treatment in A549 cells. Among the ten
herbal components, only Cinnamomi Cortex (CC) extract strikingly
suppressed TGF-β-induced EMT, as indicated by the restoration of
E-cadherin. We could not detect any inhibition of the cell
viabilities by CC extract (data not shown).
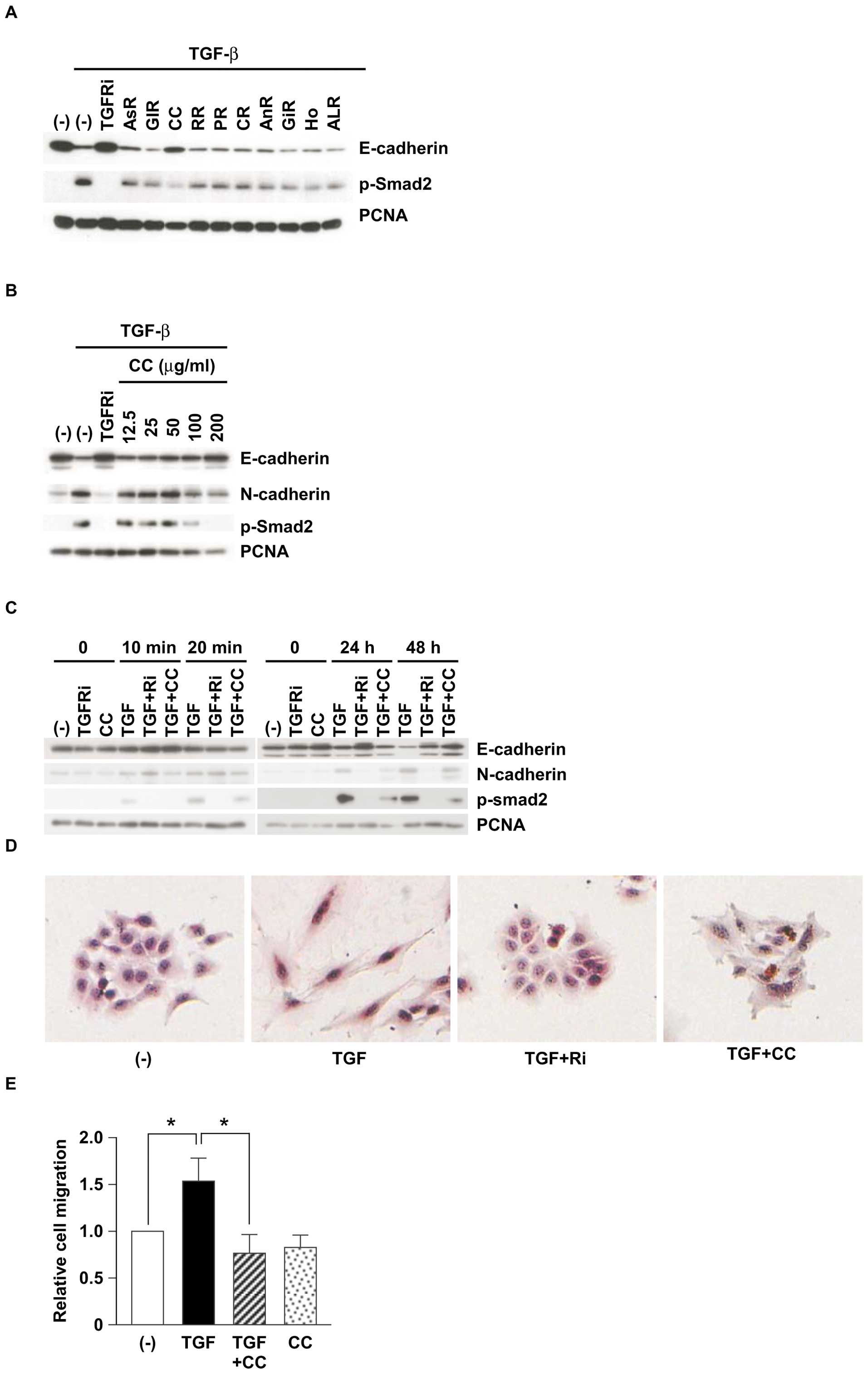 | Figure 1.Cinnamomi Cortex (CC) suppresses
TGF-β-induced EMT. (A) Effects of ten herbal components from
Juzentaihoto. AsR, Astagali Radix; GlR, Glycyrrhizae Radix;
CC, Cinnamomi Cortex; RR, Rehmanniae Radix; PR, Paeoniae Radix; CR,
Cnidii Rhizoma; AnR, Angelicae Radix, GiR, Ginseng Radix; Ho,
Hoelen; ALR, Atractylodis Lanceae Radix. A549 cells were pretreated
with various herbal ingredients (100 μg/ml) or TGF-β
receptor I kinase inhibitor (TGFRi; 10 μM) for 30 min and
stimulated with TGF-β (5 ng/ml) for 48 h. Expression of each
protein was detected by western blotting. (B) Dose-dependent
inhibition of TGF-β-induced EMT with CC treatment. A549 cells were
pretreated with various concentrations of CC (12.5, 25, 50, 100 and
200 μg/ml) or TGF-β receptor I kinase inhibitor (TGFRi; 10
μM) for 30 min and stimulated with TGF-β (5 ng/ml) for 48 h.
Expression of each protein was detected by western blotting. (C)
Time-dependent inhibition of TGF-β-induced EMT with CC treatment.
A549 cells were pretreated with CC (100 μg/ml) for 30 min
and stimulated with TGF-β (5 ng/ml) for the indicated times.
Expression of each protein was detected by western blotting. (D)
Cell morphologies with CC treatment. A549 cells were stained by
H&E after treatment as in (C) for 48 h. (E) Migration with CC
treatment. A549 cells treated as in (D) were seeded in Transwell
chambers for 6 h. No. of migrated cells per field (×400) was
counted. Relative cell migration with TGF-β (TGF, filled bar),
TGF-β with CC extract (TGF+CC, shadow bar), or CC extract (CC,
dotted bar) was normalized by untreated cell migration (open bar).
Data are the mean ± SD of four independent experiments.
*p<0.01, compared among each pair, by one-way ANOVA
and Bonferroni's multiple comparision test. |
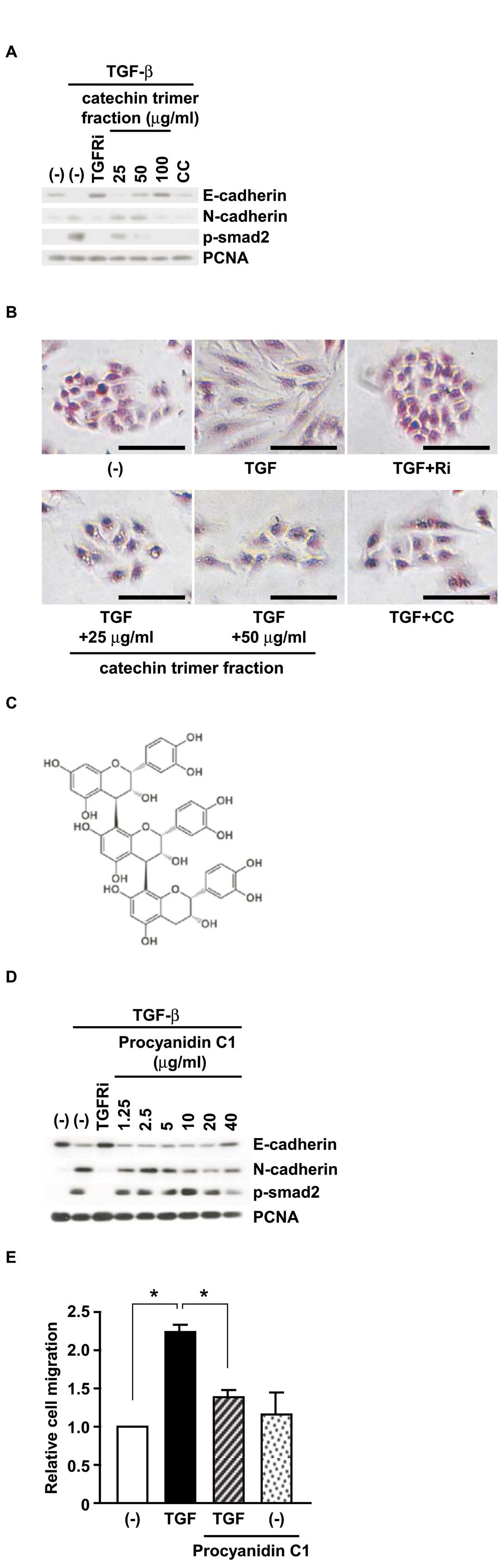
In order to further determine the inhibition of
TGF-β-induced EMT by CC extract, E-cadherin, N-cadherin, and
phosphorylated Smad-2 were examined. Similarly to Fig. 1A, restoration of E-cadherin
expression by CC extract was detected in a
concentration/time-dependent manner (Fig. 1B and C). Interestingly,
phosphorylation of Smad-2 by TGF-β, a downstream molecule of TGF-β
signaling, was suppressed by CC extract after the early phase (10
and 20 min) as well as the late phase (24 and 48 h). We also
detected the inhibition of TGF-β-induced EMT by CC extract at mRNA
levels of snail, E-cadherin and fibronectin (data not
shown). In addition to expression levels of EMT markers, we
observed morphological changes with CC extract (Fig. 1D). A549 cells treated with TGF-β
showed spindle-like shapes as compared with untreated cells. On the
other hand, TGF-β-stimulated cells treated together with CC extract
showed cobblestone-like shapes similar to the untreated cells or
the cells with TGFRi and TGF-β. Moreover, consistent with cell
morphology, TGF-β-induced cell migration was also suppressed with
CC extract (Fig. 1E). These
results support that CC extract suppresses TGF-β-induced EMT in
A549 cells and consequently inhibits cell migration.
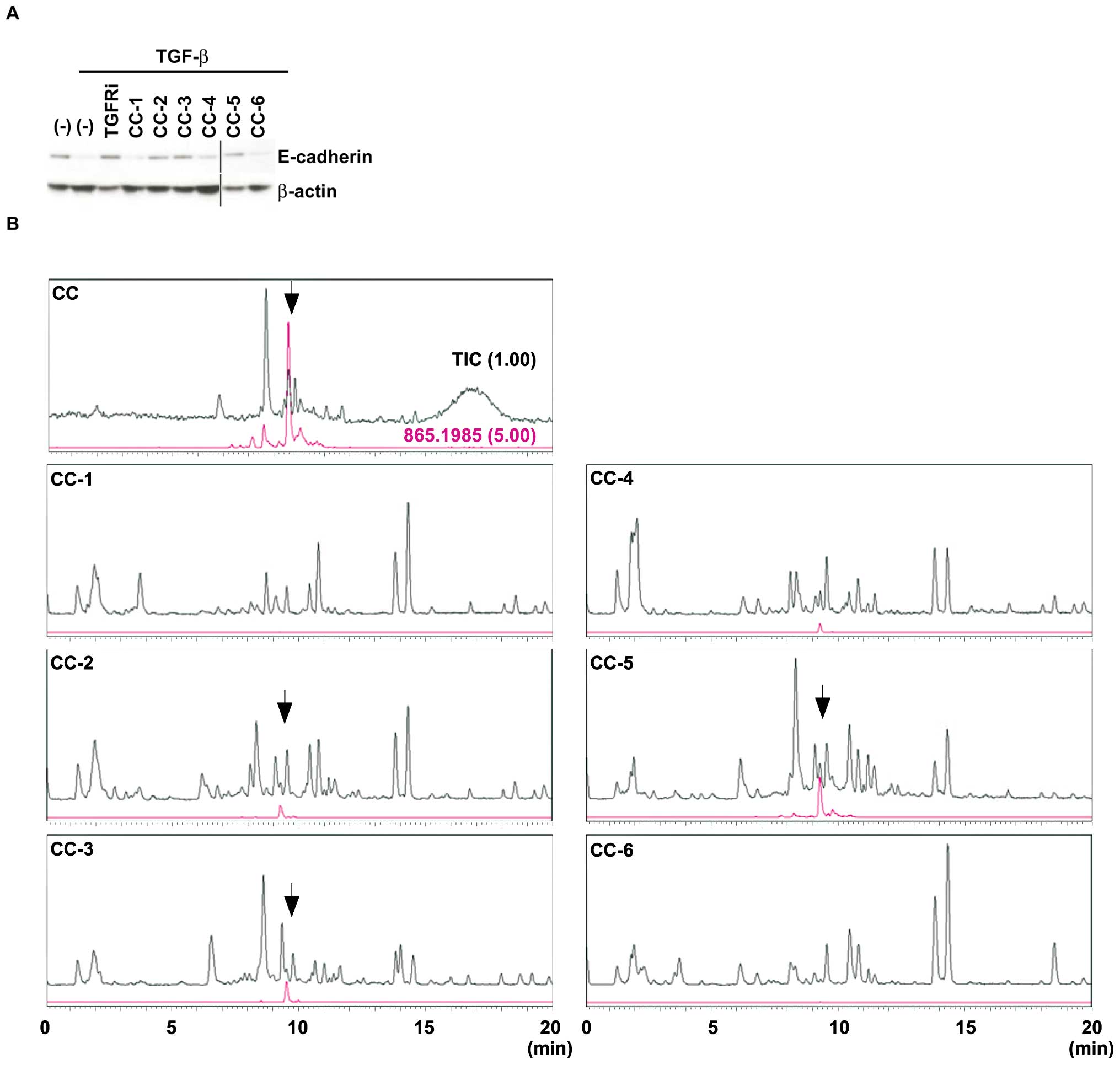
Identification of an active fraction
inhibiting TGF-β-induced EMT
To gain insight into the variations of CC extracts
in EMT-inhibitory activities, we examined six additional CC (CC-1
to -6) extracts, which differ according to the harvest location
(Fig. 2A). CC-2, CC-3 and CC-5
extracts showed restoration of E-cadherin expression stronger than
CC-1, CC-4, and CC-6 extracts; therefore, an active component(s)
for the inhibition of EMT might be included in CC-2, CC-3 and CC-5
extracts more than in CC-1, CC-4 and CC-6.
To identify the active chemical compound(s) that is
able to inhibit TGF-β-induced EMT, total ion chromatogram analysis
was performed in CC and CC-1 to CC-6 extracts. Notably, the peaks
of catechin-catechin-catechin trimer (catechin trimer)
significantly overlapped with EMT inhibitory activities in CC-2,
CC-3 and CC-5 extracts, but were not detected in CC-1, CC-4, and
CC-6 extracts (Fig. 2B),
suggesting that the catechin trimer may have crucial roles in
suppressing TGF-β-induced EMT.
Procyanidin C1 can suppress TGF-β-induced
EMT
Next to assess whether catechin trimer fraction of
CC extract has activity to inhibit EMT, we examined the protein
expression levels with the catechin trimer fraction from CC
(Fig. 3A). As predicted, the
catechin trimer fraction inhibited E-cadherin reduction and
N-cadherin induction. Similarly to protein levels, the change of
cell morphology (Fig. 3B) also
supported the inhibition of TGF-β-induced EMT.
We finally purified the catechin trimer from CC and
identified its structure, which was an
epicatechin-epicatechin-epicatechin trimer, procyanidin C1
(Fig. 3C). To check the inhibition
of EMT by procyanidin C1, the protein expression and cell migration
activities were examined (Fig. 3D and
E). Strikingly, EMT markers represented the inhibition of EMT
in western blotting, and TGF-β-induced cell migration was
suppressed by procyanidin C1.
Discussion
In this study, we firstly identified Cinnamomi
Cortex (CC) extract in 10 herbal components from
Juzentaihoto as the only inhibitory herbal ingredient of
TGF-β-induced epithelial-to-mesenchymal transition (EMT) in a human
non-small cell lung cancer cell line, A549 cells. The inhibitory
effects of CC extracts could be derived from the procyanidin C1
because of the correlation of the content of procyanidin C1 in CC
extract with their inhibitory activities.
The inhibitory effect of CC extract on EMT
phenotypes we showed here might be supported by the evidence gained
from Ninjinyoeito and Keishibukuryogan, both of which
are other Kampo medicines containing CC. Ninjinyoeito has
also been reported to suppress cancer metastasis (16). On the other hand,
Keishibukuryogan has been shown to suppress renal fibrosis,
which is caused by TGF-β-induced EMT (17). Despite the inhibitory activity of
CC extract, the possibility can not be excluded that other herbal
medicines might play a role in concert with CC extract, because of
the harmonization effects among various herbal medicines (11,12).
We identified procyanidin C1 as an active compound
for the inhibition of EMT, but CC extract contains various chemical
compounds, such as phenyl propanoids (cinnamic acid,
cinnamaldehyde), terpenoids (cinnamonol, cassioside) and tannin
(epicatechin) (18). There are
many reports on the biological activities of cinnamic acid or
cinnamaldehyde, for example, induction of ROS-mediated apoptosis
(19). In addition, the
procyanidin C1 we purified is known for its anti-inflammatory
effects (20,21); however, it is the first time to
show that procyanidin C1 has anti-metastatic or anti-EMT effects.
Among the catechin sub-family, epigallocatechin-3 gallate in green
tea has been reported to inhibit EMT in human melanoma cells
(22,23). These reports raise the possibility
that another catechin sub-family is involved in the inhibition of
EMT besides procyanidin C1. The inhibition of cell migration by
procyanidin C1 (Fig. 3E) and the
clear linkage of the amount of procyanidin C1 to the inhibition of
TGF-β-induced EMT (Fig. 2B)
suggest that procyanidin C1 is still the main compound in CC
extracts to inhibit cell migration and EMT.
Although we need more efforts to identify the target
molecule of procyanidin C1, the phosphorylation of Smad-2 (Fig. 3D) and its transcriptional activity
(data not shown) were suppressed. Indeed, CC extract and
procyanidin C1 did not affect basal cell migration in A549 cells
(CC alone or procyanidin C1 alone in Figs. 1E and 3E), suggesting that the inhibition of EMT
by CC extract and procyanidin C1 is dependent on inhibition of the
TGF-β signal pathway and that mesenchymal-to-epithelial transition
is not induced by CC extract or procyanidin C1, which might affect
basal levels of cell migration.
Collectively, our data identified CC in
Juzentaihoto as the only herbal component inhibiting
TGF-β-induced EMT, which could be worthy of clinical study in a
variety of settings associated with EMT, including cancer
metastasis and/or tissue fibrosis. As procyanidin C1 showed
inhibition of EMT, it is suggested that the utilization of
procyanidin C1 as a lead compound would be attractive for the
development of cancer metastasis inhibitors.
Abbreviations:
|
EMT
|
epithelial-to-mesenchymal
transition
|
|
CC
|
Cinnamomi Cortex
|
Acknowledgements
This study was supported in part by
Grants-in-Aid for Challenging Exploratory Research 24659348 (I.S.),
by Grant-in-Aid 24700971 for Young Scientists (B) (S.Y.) from the
Ministry of Education, Culture, Sports, Science, and Technology
(Japan) and by a Grant-in-Aid for the Cooperative Research Project
from Joint Usage/Research Center (Joint Usage/Research Center for
Science-Based Natural Medicine), Institute of Natural Medicine,
University of Toyama in 2013.
References
|
1.
|
Ou SH and Zell JA: Validation study of the
proposed IASLC staging revisions of the T4 and M non-small cell
lung cancer descriptors using data from 23,583 patients in the
California Cancer Registry. J Thorac Oncol. 3:216–227. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Fidler IJ: The evolution of biological
heterogeneity in metastatic neoplasms. Cancer Invasion and
Metastases: Biologic and Therapeutic Aspects. Nicolson GL and Milas
L: Raven Press; New York, NY: pp. 5–26. 1984
|
|
3.
|
Nicolson GL: Tumor cell instability,
diversification, and progression to the metastatic phenotype from
oncogene to oncofetal expression. Cancer Res. 47:1473–1487.
1987.PubMed/NCBI
|
|
4.
|
Hart IR: Seed and soil revisited:
mechanisms of site specific metastasis. Cancer Metastasis Rev.
1:5–16. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Liotta LA, Rao CV and Barsky SH: Tumour
invasion and the extracellular matrix. Lab Invest. 49:636–649.
1983.PubMed/NCBI
|
|
6.
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Guarino M: Epithelial-mesenchymal
transition and tumor invasion. Int J Biochem Cell Biol.
39:2153–2160. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Xu J, Lamouille S and Derynck R:
TGF-β-induced epithelial to mesenchymal transition. Cell Res.
19:156–172. 2009.
|
|
9.
|
Maruyama H, Kawamura H, Takemoto N,
Komatsu Y, Aburaya M and Hosoya E: Effect of Juzentaihoto on
phagocytes. Jpn J Inflammation. 8:461–465. 1988.
|
|
10.
|
Ohnishi Y, Fujii H, Hayakawa Y, et al:
Oral administration of a Kampo (Japanese herbal) medicine
Juzentaihoto inhibits liver metastasis of colon 26-L5
carcinoma cells. Jpn J Cancer Res. 89:206–213. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Ohnishi Y, Yamaura T, Tauchi K, et al:
Expression of the anti-metastatic effect induced by Juzentaihoto is
based on the content of Shimotsu-to constituents. Biol Pharm Bull.
7:761–765. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Saiki I: A Kampo medicine
‘Juzentaihoto’ - prevention of malignant progression and
metastasis of tumor cells and the mechanism of action. Biol Pharm
Bull. 23:677–688. 2000.
|
|
13.
|
Sakurai H, Suzuki S, Kawasaki N, et al:
Tumor necrosis factor-alpha-induced IKK phosphorylation of
NF-kappaB p65 on serine 536 is mediated through the TRAF2, TRAF5,
and TAK1 signaling pathway. J Biol Chem. 278:36916–36923. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Ogasawara M, Murata J, Ayukawa K and Saiki
I: Differential effect of intestinal neuropeptides on invasion and
migration of colon carcinoma cells in vitro. Cancer Lett.
119:125–130. 1997. View Article : Google Scholar
|
|
15.
|
Esatbeyoglu T, Jaschok-Kentner B, Wray V
and Winterhalter P: Structure elucidation of procyanidin oligomers
by low-temperature 1H NMR spectroscopy. J Agric Food
Chem. 59:62–69. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Kamei T, Kumano H, Iwata K, Nariai Y and
Matsumoto T: The effect of a traditional Chinese prescription for a
case of lung carcinoma. J Altern Complement Med. 6:557–579. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Nakagawa T, Tashiro I, Fujimoto M, et al:
Keishibukuryogan reduces renal injury in the early stage of renal
failure in the remnant kidney model. Evid Based Complement Alternat
Med. 2011:9142492011. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Leela NK: 7 Cinnamon and Cassia. Chemistry
of Spices. Parthasarathy VA, Chempakam B and Zachariah TJ: CAB
International; Wallingford, Oxon: pp. 124–145. 2008, View Article : Google Scholar
|
|
19.
|
Ka H, Park HJ, Jung HJ, et al:
Cinnamaldehyde induces apoptosis by ROS-mediated mitochondrial
permeability transition in human promyelocytic leukemia HL-60
cells. Cancer Lett. 196:143–152. 2003. View Article : Google Scholar
|
|
20.
|
Nakano N, Nishiyama C, Tokura T, et al:
Procyanidin C1 from apple extract inhibits FcεRI-mediated mast cell
activation. Int Arch Allergy Immunol. 147:213–221. 2008.PubMed/NCBI
|
|
21.
|
Terra X, Palozza P, Fernandez-Larrea J, et
al: Procyanidin dimer B1 and trimer C1 impair inflammatory response
signalling in human monocytes. Free Radic Res. 45:611–619. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Singh T and Katiyar SK: Green tea
catechins reduce invasive potential of human melanoma cells by
targeting COX-2, PGE2 receptors and epithelial-to-mesenchymal
transition. PLoS One. 6:e252242011. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Chen PN, Chu SC, Kuo WH, Chou MY, Lin JK
and Hsieh YS: Epigallocatechin-3 gallate inhibits invasion,
epithelial-mesenchymal transition, and tumor growth in oral cancer
cells. J Agric Food Chem. 59:3836–3844. 2011. View Article : Google Scholar : PubMed/NCBI
|