Introduction
Galectin-1 is expressed in anaplastic large cell
lymphoma (ALCL) as well as Hodgkin lymphoma (HL) (1). However, the biological significance
of galectin-1 still remains unclear. Galectin-1 is known to induce
cell death in human lymphoma and T cells (1–3).
Galectins act as many biological functional molecules (4). Galectin-1 exists in serum and
deposits to extracellular matrix (5). In addition, galectin-1 regulates cell
adhesion of ovarian cancer cells (6). Cell surface sialic acid appeared to
modulate cell adhesive capacity of lymphoma cells (7). In this study, we analyzed cell
adhesive and invasive capacity to galectin-1 using human ALCL cell
line to clarify the biological roles of galectin-1 in ALCL. We
applied the cell surface lectin array in analysis of cell surface
glycan modifications (8).
Materials and methods
Cell line
Human anaplastic large cell lymphoma cell line,
H-ALCL was established in our laboratory. The H-ALCL cells were
grown in the culture medium of RPMI-1640 containing 15% fetal calf
serum in 5% CO2 at 37°C. The H-ALCL cell line expresses
the galectin-1 receptors, CD45RA [(leukocyte common antigen (LCA)]
and CD45RO (UCHL-1) on the flow cytometric analysis (data not
shown).
Cell surface lectin array analysis
We applied the cell surface lectin array analysis to
detecet the cell surface glycosylations according to Landemarre
et al with several modifications (8). The H-ALCL cells were treated with or
without neuraminidase from Arthrobacter ureafaciens (no.
10269611001, Roche, Germany) at 0.2 U/ml, at 37°C for 30 min, then
the cells were cytospun and cytospin cell preparations were stained
by PNA lectin as described previously (9). Arachis hypogaea (PNA),
Artocarpus integrifolia (Jacalin), Glycine max (SBA),
Helix pomatia (HPA), Vicia villosa (VVA), Ulex
europaeus (UEA-1), Triticum vulgaris (WGA), Canavalia
ensiformis (ConA), Phaseolus vulgaris-L (L-PHA),
Phaseolus vulgaris-E4 (E-PHA), Datura stramonium
(DSA) lectins were from EY Laboratory. The 96-well plate was coated
by each lectin and air-dried. The neuraminidase treated or
non-treated H-ALCL lymphoma cells (1×106/2 ml) were
applied to each well (100 μl/well) and incubated at 37°C for
60 min. After aspiration of the medium, PBS was added to each well
and then aspirated to remove non-adhered cells. Then 100 μl
of 3.7% formaldehyde was added to each well to fix the adhesive
cells at RT for 40 min. After aspiration of formaldehyde, 100
μl of 0.1% crystal violet was added to each well and the
plates were incubated at RT for 40 min. After washing twice, 100
μl of 10% acetic acid was added to each well and the
absorbance at 570–655 or 570 nm was determined using an ELISA plate
reader (7). To analyze the effect
of O-glycosylation cells were treated with the O-glycosylation
inhibitor benzyl 2-acetamido-2-deoxy-3-O-β-D-galactopyranosyl
-α-D-galactopyranoside (benzyl-α-GalNAc B5019, Sigma) (BZ) at a
concentration of 2 mM in culture medium for 72 h at 37°C. To
analyze the effect of N-glycosylation cells were treated with the
N-glycosylation inhibitor SW at a concentration of 1 μg/ml
in culture medium for 96 h at 37°C. BZ treated or non-treated
H-ALCL lymphoma cells (1×106/2 ml) were applied to each
well (100 μl/well), or SW treated or non-treated H-ALCL
lymphoma cells (2×106/2 ml) were applied to each well
(100 μl/well) and incubated as described above.
Cell adhesion assay
Tissue culture plates with 96-wells were coated with
bovine splenic galaptin (a synomym of galectin-1, Sigma, G8777) (10
μg/well) or human recombinant galectin-1 (ATGP0385, ATGen
Co. Ltd.) (10 μg/well) and dried at room temperature
overnight. Each well was washed with 100 μl PBS and was
filled with RPMI-1640 containing 15% bovine serum albumin (BSA) 15%
FCS and the plates were incubated at 37°C for 60 min. After
aspiration of the medium, H-ALCL cells with or without
neuraminidase (from Arthrobacter ureafaciens (AU): final
concentration 0.2 U/ml, at 37°C for 30 min, α2,3-neuraminidase
(BioLabs, P0728S, 50,000 U/ml): final concentration 0.2 U/ml, at
37°C for 30 min, neuraminidase from Newcastle disease virus (NDV):
0.2 U/ml, Prozyme, at 37°C for 30 min) treatment were added to each
well and incubated at 37°C for 1 h. After aspiration of the medium,
PBS was added to each well and then aspirated to remove non-adhered
cells. Then 100 μl of 3.7% formaldehyde was added to each
well to fix the adhesive cells at RT for 40 min. After aspiration
of formaldehyde, 100 μl of 0.1% crystal violet was added to
each well and the plates were incubated at RT for 40 min. After
washing with PBS, 100 μl of 10% acetic acid was added to
each well and the absorbance at 570–655 nm or 570 nm was determined
using an ELISA plate reader (7).
To analyze the effect of O-glycosylation and N-glycosylation cells
are treated with O-glycosylation inhibitor BZ and N-glycosylation
inhibitor SW as described above. To analyze the steric hindrance of
N-glycosylation in cell adhesion to galectin-1, we performed lectin
blocking assay in galectin-1 adhesion assay. To analyze involvement
of anaplastic large cell lymphoma kinase (ALK), CD30, CD45, CD45RO,
epithelial membrane antigen (EMA) in cell adhesion to galectin-1,
we carried out the inhibition assay using anti-ALK (ALK1, Dako,
M7195), CD30 (BerH2, Dako, M0751), CD45 [leukocyte common antigen
(LCA), Nichirei, H0108, Japan], CD45RO (UCHL-1, Dako, M0742), EMA
(E29, Dako, M0613) antibody on galectin-1 adhesion assay. For
isotype control experiment, the mouse IgG isotype control antibody
(BD Pharmingen, no. 554721) was used. These antibodies were used as
5 μl in 1 ml cell suspension. ALK, CD30, CD45, CD45RO and
EMA are expressed on the cell surface of H-ALCL cells determined by
flow cytometric analysis (data not shown). Anti-CD30, CD45, CD45RO
and EMA antibodies enhanced cell adhesion to galectin-1 suggesting
that the protein portion of these molecules may be involved in the
interaction between galectin-1 (in our preliminary data, not
shown).
On the resialylation assay, the H-ALCL cells were
desialylated by neuraminidase treatment and then, the cells were
resialylated by recombinant ST6Gal1 (R&D Systems, 5924-GT) at a
condition ST6Gal1 5 μg/100 μl with CNP-Neu5Ac (Sigma,
C8271) 1 mM, at 37°C for 120 min.
Knockdown of ST6Gal1
In order to analyze the regulatory mechanism of cell
surface sialylation by ST6Gal1, siRNA transfection was performed as
described previously with several modifications (25). For transfection, INTERFERin
(Polyplus transfection, USA) was used according to the
manufacturer’s instructions. For knockdown experiments, siRNA (cat.
no. 12842 called Type 42, sense: AGACAGUUUGUACA AUGAAtt, antisense:
UUCAUUGUACAAACUGUCUtt, or cat. no. s12843 called Type 43, sense:
ACCACUCAGAUAU CCCAAAtt, antisense: UUUGGGAUAUCUGAGUGGUat, Ambion
Japan) was used. For control experiments, Ambion Silencer™ select
negative control no. 1 siRNA (cat. no. 4390843) was applied. After
24-h incubation, the immunohistochemical staining was performed by
anti-ST6Gal1 antibody (dilution ×100, R&D Systems, AF5924) and
knockdown effect was validated by inhibition of ST6Gal1 protein
expression in the cytoplasm of H-ALCL cells (9).
Invasion assay
The invasion assay (haptotaxis) was performed based
on the methods of Albini et al (10) with several modifications. The
24-well culture plate was filled with 600 μl the culture
medium RPMI-1640 containing 15% BSA 15% FCS. The lower surfaces of
the membranes of transwell chamber, chemotaxicell (Krabo, Japan)
with an 8-μm pore membrane were coated with 10 μl
galectin-1 (1.0 mg/ml, ATGen, no. ATGP0385) and dried at RT. Then
coated chemotaxicells were inserted into each well. H-ALCL cells,
100 μl of 2.4×106/ml, were inserted into each
chemotaxicell and incubated at 37°C for 24 h. After incubation, the
invaded cells at lower level of each well were counted by
trypan-blue exclusion methods. The cell count was performed using
duplicate wells with two experiments (n=4), or triplicate wells
with at least two independent experiments. On preliminary assay the
amount of invaded cells in non-coating chemotaxicell was more than
that in galectin-1 coating chemotaxicell (data not shown)
suggesting that H-ALCL cells can invade to lower chamber through
cell ontact to galectin-1 matrix component. To analyze the effect
of cell surface sialylation, H-ALCL cells were treated with
neuraminidase from Arthrobacter ureafaciens (AU) (final
concentration 0.2 U/ml) at 37°C for 30 min. For analysis of
phosphatidylinositol 3 kinase (PI3K) inhibitor, wortmannin (681675,
Calbiochem) and mitogen-activated protein kinase (MAPK) inhibitor,
PD98059 (513000, Calbiochem) or Rho inhibitor (C3 transferase)
cells were pre-incubated with wortmannin at 1.7 μM or
PD98059 at 25 μM for 30 min, or C3 transferase at 2.0
μg/ml, 2 h. Then the cell adhesion assay or invasion assay
were performed. We confirmed the expression of PI3K, MAPK and Rho
in the tumor cells of H-ALCL on immunohistochemical staining (data
not shown). For analysis of cytochalasin B (Sigma), cells were
pre-treated with cytochalasin B at 4 μM for 30 min.
Galectin-1 and ST6 Gal1 expression
H-ALCL cells were cytospun to the slide glass and
the specimen was fixed with 100% alcohol. The immunohistochemical
staining by anti-galectin-1 antibody (100X dilution, Santa Cruz,
clone S-14) or anti-ST6Gal1 antibody (100X dilution, R&D
Systems, AF5924). Then, the preparations were incubated with
biotinylated anti-goat immunoglobulin. After washing in PBS three
times, the preparations were incubated at room temperature for 20
min with avidin-biotin peroxidase complex kit (Dako, Tokyo, Japan).
Then, they were incubated for 5 min at room temperature with
diaminobenzidine (DAB)-H2O2 solution (60
μg DAB in 150 ml PBS). The preparations were counterstained
with hematoxylin and mounted.
Galectin-1 detection by ELISA
The supernatant of the conditioned culture medium in
H-ALCL was prepared by centrifugation at 11,000 rpm, for 20 min, at
4°C. Galectin-1 was measured by ELISA assay kit (USCN Life Science
Inc. E9032 Hu) according to the manufacturer’s instructions.
Cell death induction and CD45 activity
assay by galectin-1 treatment
The H-ALCL cells were treated with or without 12
μM galectin-1 for 72 h. Then the number of viable cells was
counted by trypan-blue exclusion test. We analyzed the CD45 protein
tyrosine phosphatase (PTP) activity by the CD45 PTP drug discovery
kit, AK-812 (Biomol) according to the manufacturer’s instructions
with modification. For analysis, the cell lysate was prepared by 1%
NP-40 PBS buffer. The PTP inhibitor, sodium orthovanadate (V)
(vanadate) (Wako, Japan) treatment was performed at 100 μM
for 16 h at 37°C using H-ALCL cell line for cell death analysis,
and 400 μM for 2 h for CD45 PTP activity analysis. The
inhibition of CD45 activity by vanadate was validated on CD45
activity assay using pure CD45 protein mixed with vanadate (data
not shown).
Results
Sialylation and cell adhesion assay
Treatment of neuraminidase which cleaves cell
surface sialic acid enhanced PNA, HPA and L-PHA lectin reactivity
suggesting that neuraminidase removes cell surface sialic acid from
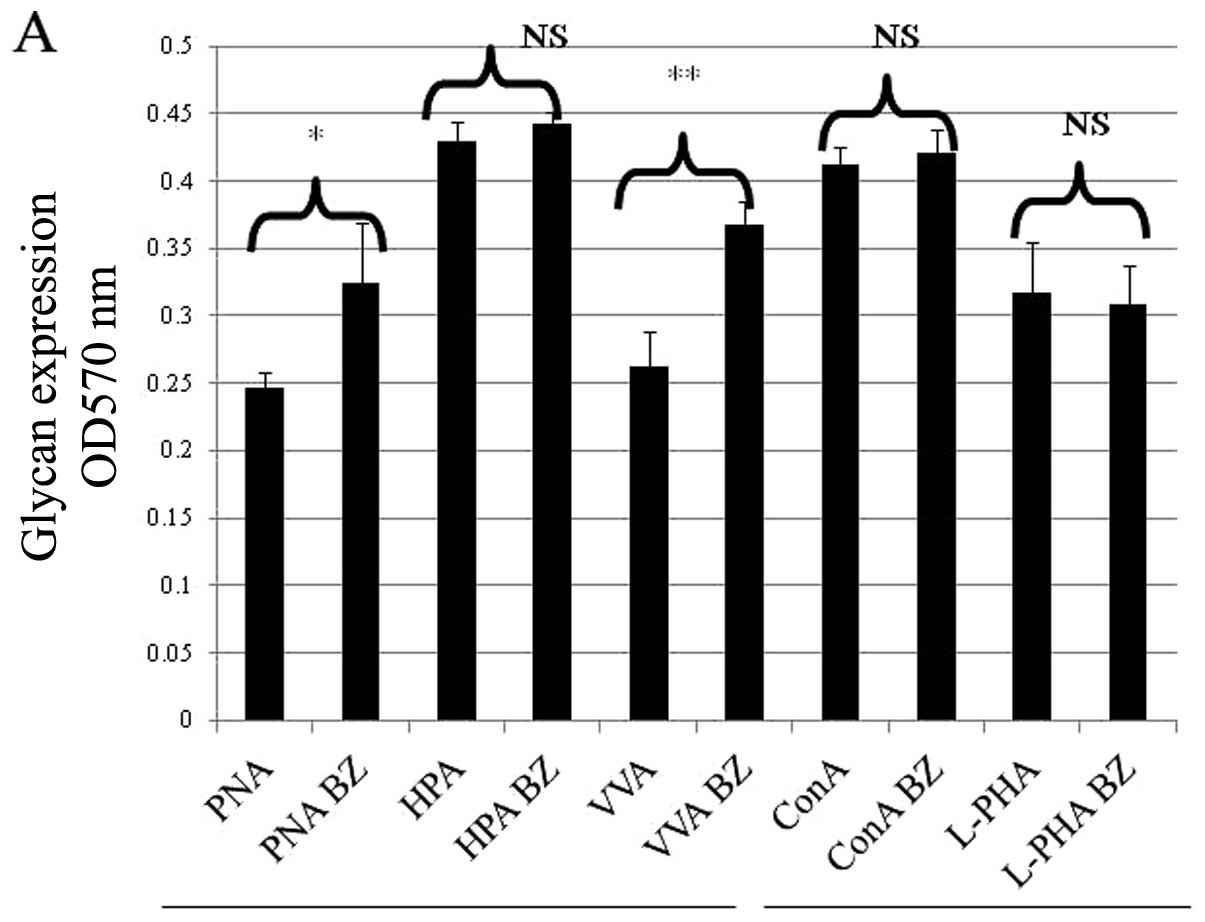
O- and N-glycans (Fig. 1A).
Treatment of neuraminidase from Arthrobacter ureafaciens
markedly enhanced cell adhesion to galectin-1 (using galaptin)
(Fig. 1B). Treatment of
neuraminidase from Arthrobacter ureafaciens markedly
enhanced cell adhesion to galectin-1 (using recombinant
galectin-1). Treatment of neuraminidase from Newcastle disease
virus inhibited cell adhesion to galectin-1 and α2,3-neuraminidase
did not enhance cell adhesion to galectin-1 (Fig. 1C). On resialylation assay, ST6Gal1
enhanced cell adhesion to WGA, and inhibited the desialyated H-ALCL
cell binding capacity to L-PHA lectin and galectin-1 (Fig. 1D). On knockdown experiments,
ST6Gal1 dramatically disappeared in the cytoplasm of H-ALCL cells
and knockdown of ST6Gal1 enhanced cell adhesion to galectin-1
(Fig. 1E and F).
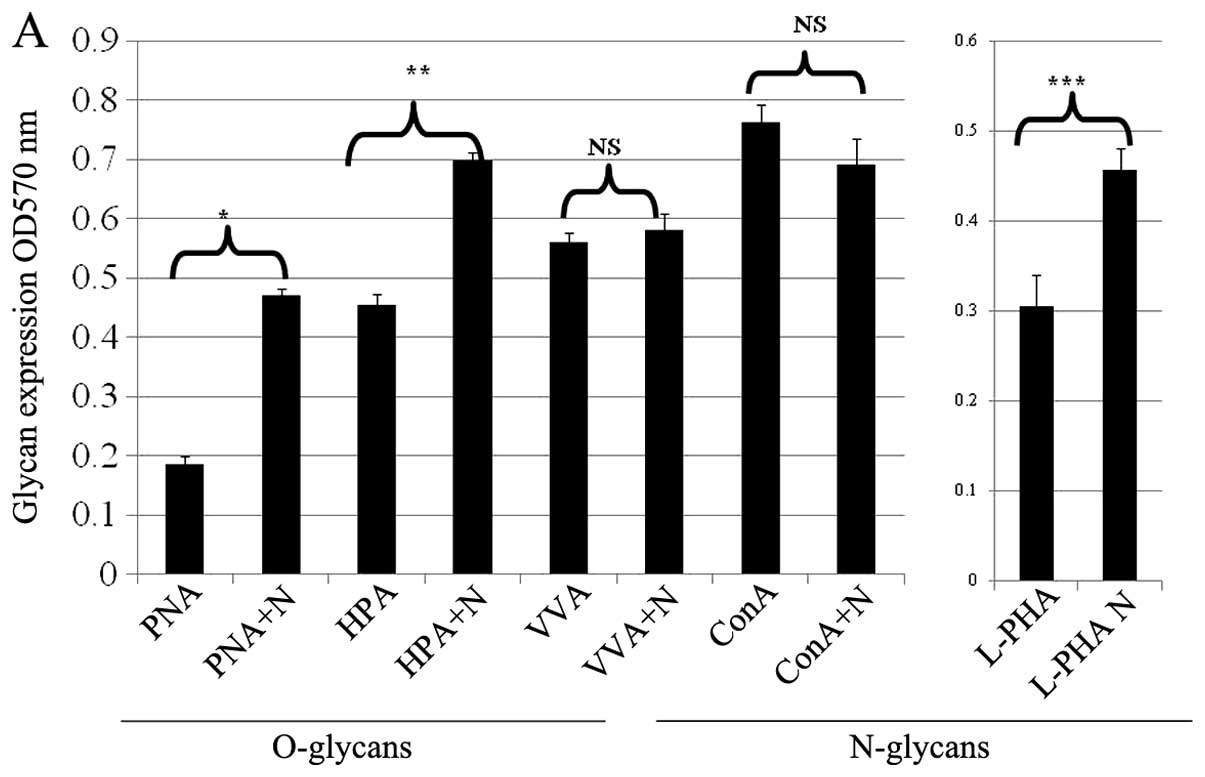 | Figure 1.The treatment of neuraminidase which
cleaves cell surface sialic acid enhanced PNA, HPA and L-PHA lectin
reactivity suggesting that neuraminidase removes cell surface
sialic acid from O- or N-glycans (PNA, *P<0.00001;
HPA, **P<0.0001; L-PHA, ***P=0.002). N,
neuraminidase pre-treatment (A). Treatment of neuraminidase
markedly enhanced cell adhesion to galectin-1 (using galaptin)
(*P<0.0009). Neu, neuraminidase pre-treatment (B).
Treatment of neuraminidase from Arthrobacter ureafaciens
markedly enhanced cell adhesion to galectin-1 (using recombinant
galectin-1). Treatment of Newcastle disease virus neuraminidase
dramatically inhibited cell adhesion to galectin-1 and α2,3
neuraminidase did not enhance cell adhesion to galectin-1 (C)
(*P=0.0000069; **P= 0.0001; NS, not
significant). AU, neuraminidase from Arthrobacter
ureafaciens; NDV, neuraminidase from Newcastle disease virus;
23 NEU, α2,3 specific neuraminidase. On resialylation assay, ST6Gal1 enhanced cell
adhesion to WGA, and inhibited the desialyated H-ALCL cells binding
capacity to L-PHA lectin and galectin-1 (*P=0.003;
**P=0.002; ***P=0.01) (D). On knockdown
experiments, expression of ST6Gal1 on the cytoplasm of H-ALCL cells
immunohistochemically (E) and knockdown of ST6Gal1 dramatically
enhanced cell adhesion to galectin-1 (*P=0.018;
**P=0.0017) (F). Representative results from two or
three independent experiments in triplicate are shown. |
O-glycosylation inhibitor and cell
adhesion assay
O-glycosylation inhibitor, benzyl-GalNAc (BZ)
treatment resulted in the enhancement of PNA and VVA lectin
reactivity suggesting the inhibition of elongation of
O-glycosylation (Fig. 2A). ConA
and L-PHA lectin binding activity which is related to N-glycans was
not dramatically changed. Treatment of BZ did not show alteration
of cell adhesive capacity to human recombinant galectin-1 (Fig. 2B).
N-glycosylation inhibitor and adhesion
assay
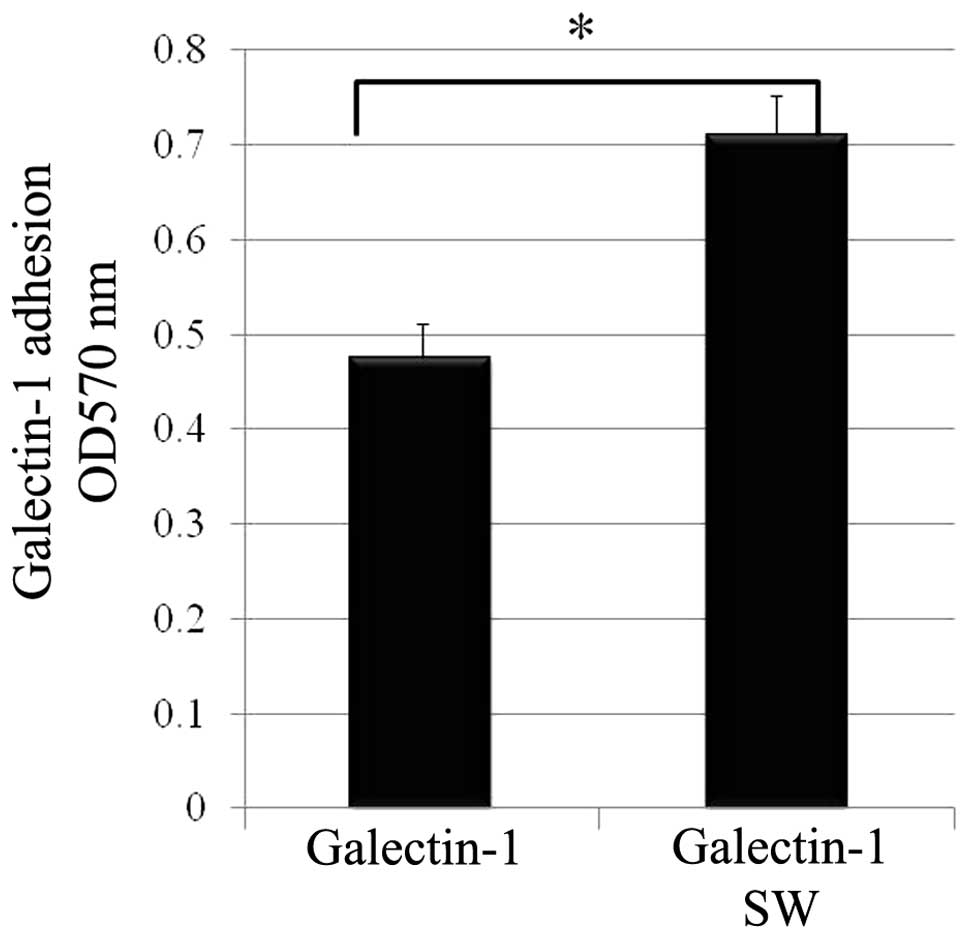
Treatment of SW markedly enhanced the cell adhesive
capacity to human recombinant galectin-1 (Fig. 3).
Glycomic analysis on galectin-1 cell
adhesion assay
Briefly the H-ALCL cells were treated with
neuraminidase from AU. Then the H-ALCL cells were treated with PNA,
Jacalin, SBA, HPA, VVA, UEA-1, WGA, L-PHA, E-PHA and DSA lectins
and these lectins modulate the adhesive properties to galectin-1 of
H-ALCL cells (Fig. 4).
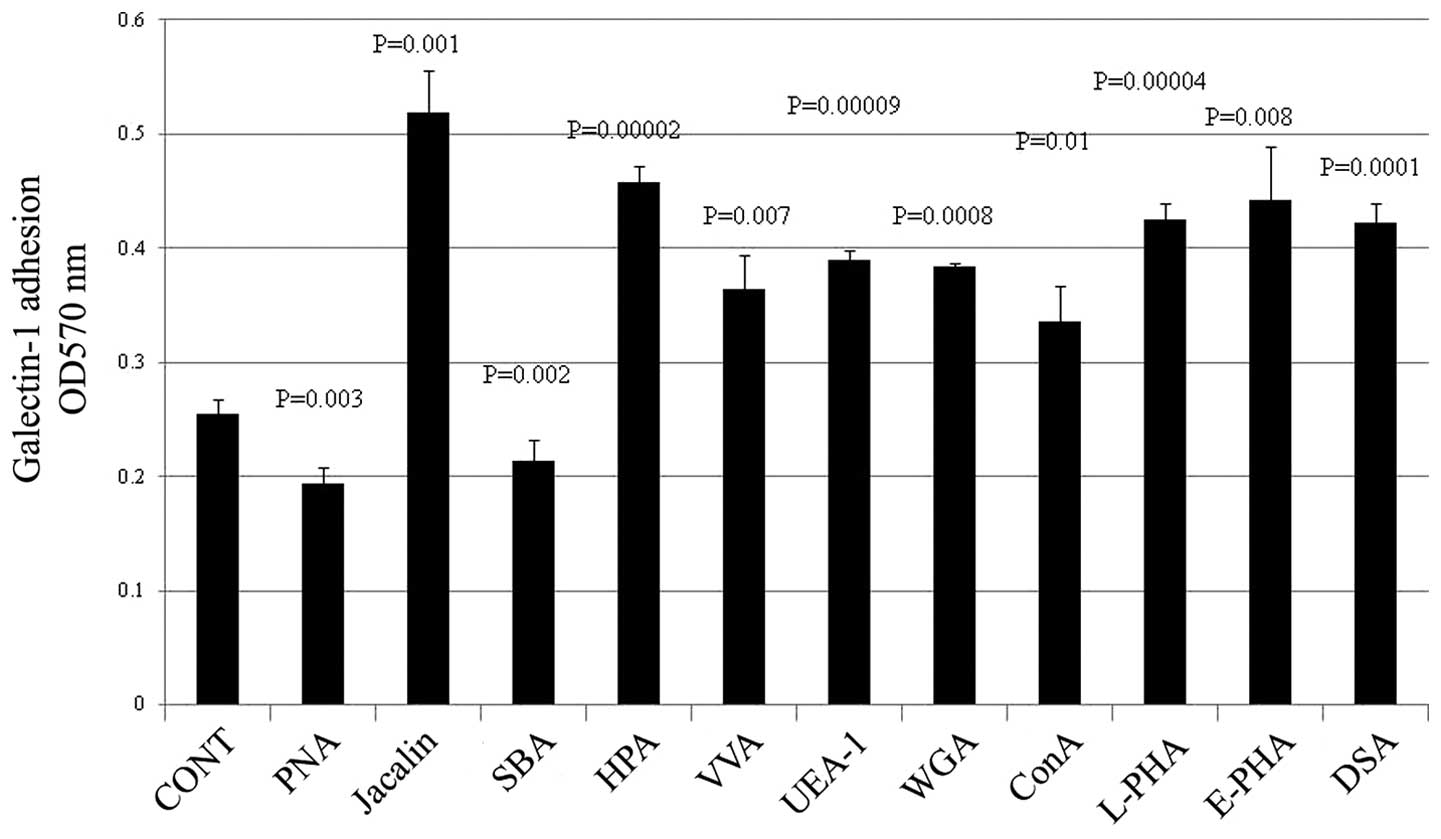 | Figure 4.The H-ALCL cells were treated with
neuraminidase from AU. The H-ALCL cells were treated with PNA,
Jacalin, SBA, HPA, VVA, UEA-1, WGA, L-PHA, E-PHA and DSA lectins
and these lectins modulate the adhesive properties to galectin-1 of
H-ALCL cells (PNA, P=0.003; Jacalin, P=0.001; SBA, P=0.002; HPA,
P=0.00002; VVA, P=0.0075; UEA-1, P=0.00009; WGA, P=0.0008; L-PHA,
P=0.00004; E-PHA, P=0.008; DSA, P=0.0001; NS, not significant).
Representative results from two independent experiments in
triplicate are shown. |
Sialylation regulates cell invasion
through galectin-1
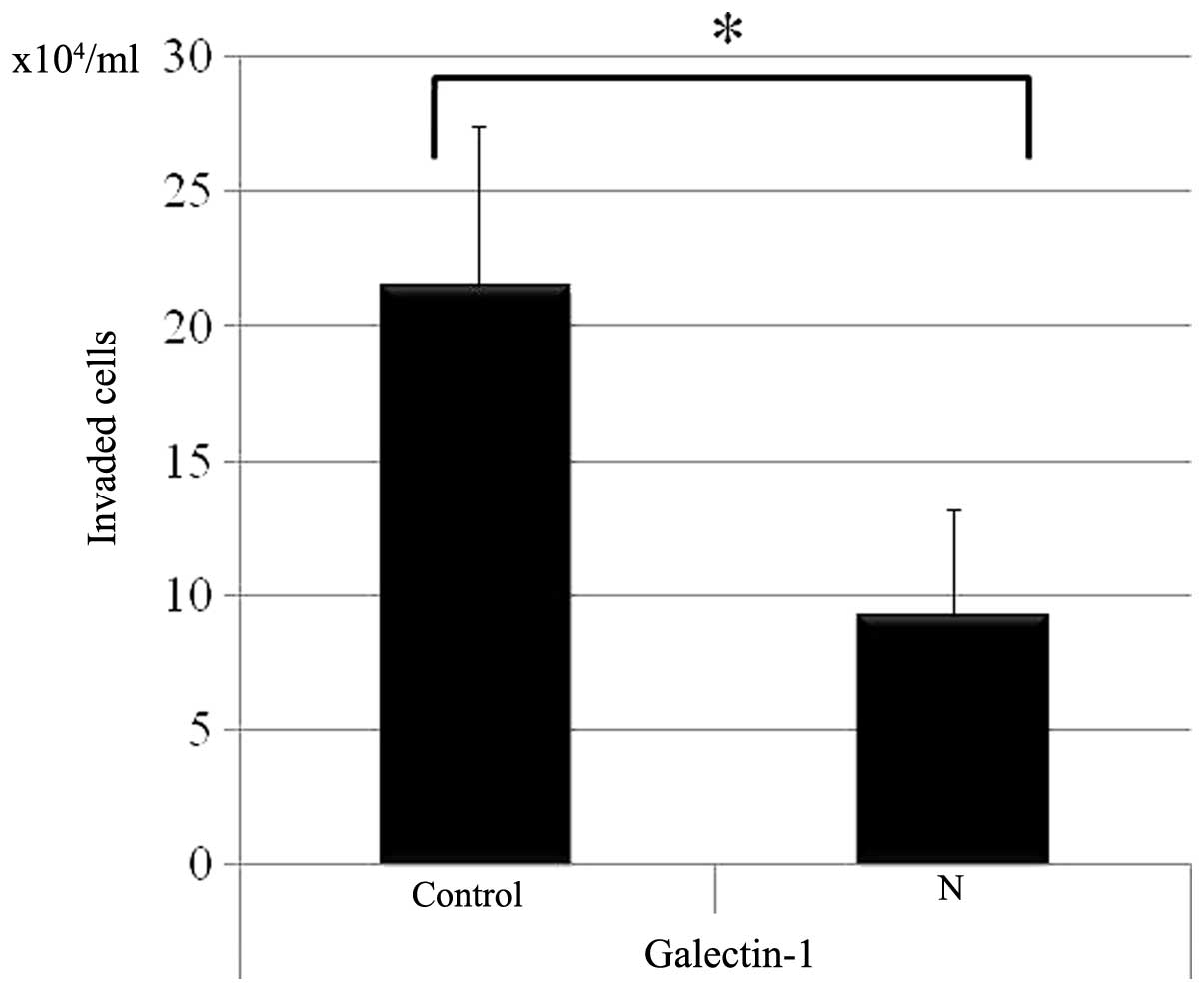
Treatment of neuraminidase markedly inhibited cell
invasive capacity to galectin-1 (Fig.
5).
Galectin-1 and ST6Gal1 expression
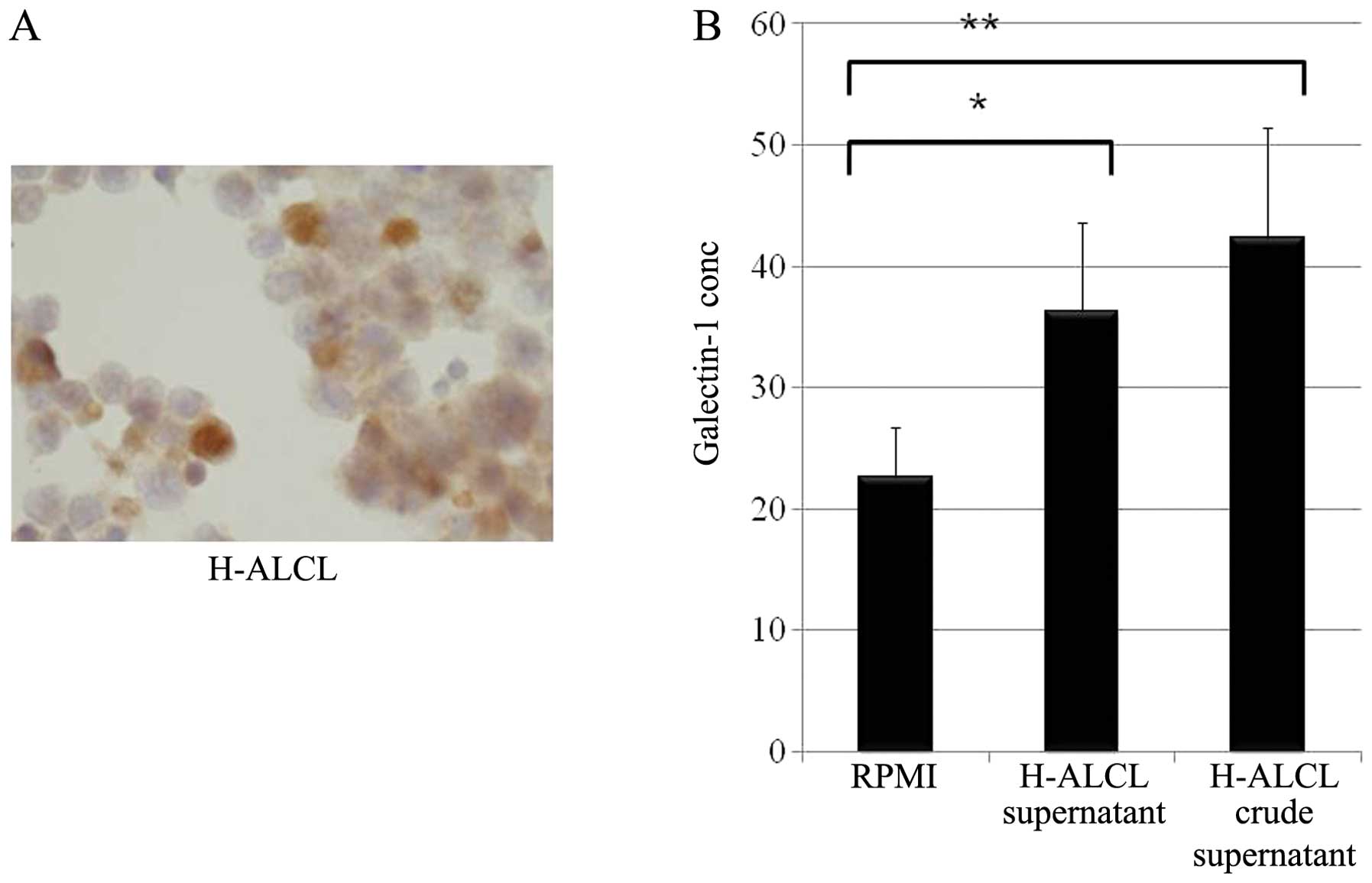
H-ALCL cells showed galectin-1 expression in the
cytoplasm (Fig. 6A). Galectin-1
was produced in autocrine fashion in H-ALCL cell line (Fig. 6B). ST6Gal1 was expressed in the
cytoplasm of H-ALCL cells (Fig.
6C).
Effect of wortmannin, PD98059,
cytochalasin B and Rho inhibitor
The amount of cell adhesion of H-ALCL cells was
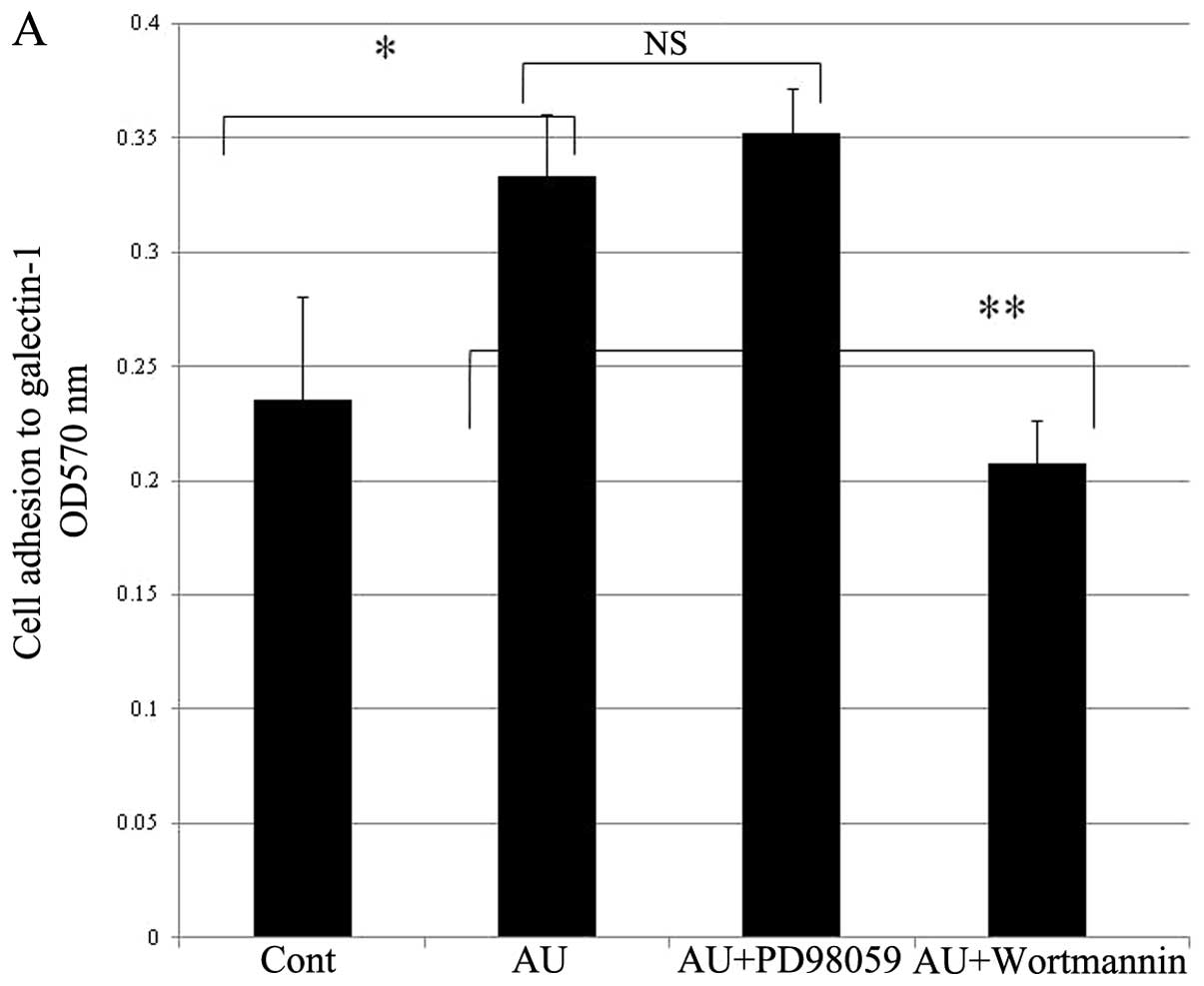
inhibited by treatment of wortmannin (Fig. 7A). The amount of cell invasive
capacity was inhibited by treatment of wortmannin and PD98059
(Fig. 7B). The amount of cell
adhesion of H-ALCL cells was enhanced by treatment of cytochalasin
B (Fig. 7C). The amount of cell
invasion to galectin-1 was dramatically inhibited by treatment of
cytochalasin B (Fig. 7D). The
amount of cell invasion to galectin-1 was dramatically inhibited by
treatment of Rho inhibitor, C3 transferase (Fig. 7E).
Cell death induction by galectin-1 and
CD45PTP
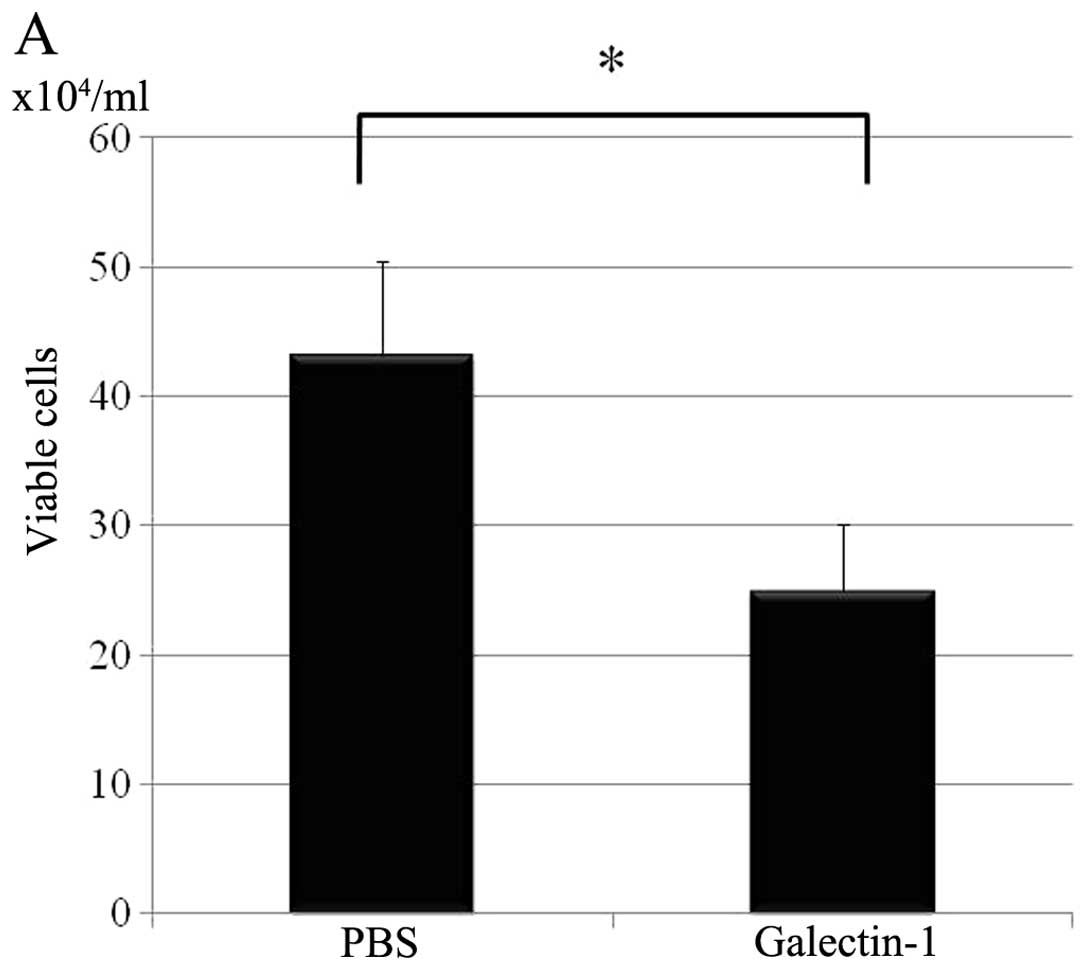
Measurement of viable cells by trypan-blue exclusion
method revealed the number of H-ALCL cells decreased with
galectin-1 treatment after 3 days (Fig. 8A). On treatment with 20 μM
galectin-1 for 1 h, the CD45 PTP activity was inhibited (Fig. 8B). Vanadate can completely inhibit
the recombinant CD45 PTP activity (data not shown). Treatment with
vanadate inhibited CD45 PTP activity after 2 h at a concentration
of 400 μM (Fig. 8C) and
induced cell death in H-ALCL cell line after 16 h at a
concentration of 100 μM (Fig.
8D).
Discussion
Galectin-1 acts as extracellular matrix (6) as well as a modulator of cell adhesion
and invasion to extracellular matrix in tumor cell lines (11–14).
The present data suggested that cell surface sialylation may
inhibit cell adhesion to galectin-1 in H-ALCL cells and suggested
that cell surface desialylation resulted in enhancement of cell
adhesion to galectin-1 and reduction in cell invasion through
galectin-1. Previously we reported that α2,6-linked sialic acid is
involved in masking effect on interaction between galectin-1 and
cell surface glycans (7,15). In the present study α2,6-linked
sialic acid is similarly involved. The data confirmed α2,6-linked
sialic acid is able to modulate interactions between galectin-1 and
cell surface glycans in human lymphoma. On the other hand, the
neuraminidase from NDV which can cleave α2,3, 2,8, 2,9-linked
sialic acids, not 2,6-linked, inhibited degree of cell adhesion to
galectin-1. The α2,3 neuraminidase did not enhance cell adhesion to
galectin-1. Therefore, taken together, α2,6-linked sialic acids may
be essential to inhibit cell adhesion to galectin-1. Several recent
reports suggested that α2,3-linked sialic acid also may be
associated with tumor metastasis (16,17).
In future investigations we will clarify biological functions of
α2,3-linked sialic acid in malignant lymphoma. Furthermore, the
H-ALCL cells expressed ST6Gal1 in the cytoplasm, and neuraminidase
treatment enhanced cell adhesion to L-PHA lectin and the
α2,6-linked sialylation by ST6Gal1 resulted in inhibition of cell
adhesion to L-PHA lectin suggesting that ST6Gal1 resialylates the
terminal galactose residue of N-glycans. Treatment with ST6Gal1
inhibits cell adhesion to galectin-1 suggesting that α2,6-linked
sialylation of terminal residue of N-glycans by ST6Gal1 inhibits
cell adhesion to galectin-1. The data are consistent with our
previous reports using DLBCL cell line, HBL-2, as described above
(7). Therefore, α2,6-sialylation
of cell surface glycans may be associated with inhibitory effect of
cell adhesion to galectin-1 regardless of histological types of
human lymphoma cells. α2,6-sialylation by ST6Gal1 is expressed in
colon cancer tissues compared to that of normal tissue and
expression of ST6Gal1 is correlated to the high risk group in
pediatric acute leukemia (18,19).
α2,6-sialylation of tumor cells may be associated with
carcinogenesis in colon cancer cells. ST6Gal1 is reported to be
secreted into cell culture media (20). In our speculation a soluble form of
ST6Gal1 may be a regulator to resialylate cell surface of lymphoma
cells as indicated in the present results using a recombinant
ST6Gal1.
In the present data neuraminidase treatment enhanced
cell adhesion to PNA, HPA and L-PHA suggesting that desialylation
resulted in exposure of β-galactose (ligand for PNA) or GalNAc
(ligand for HPA) or lactosamine (ligand for L-PHA) residues of O-
or N-glycans on the cell surface. Therefore, there is a possibility
that GalNAc may influence cell adhesion to galectin-1 as well as
one of the galectin-1 receptor, β-galactose residue. Cell surface
sialic acid is known to be closely related to invasive capacity of
lymphoma cells (21). Furthermore,
the cases with the highly sialylated type glycan showed a high
clinical stage in human lymphoma (9). These data suggested that the high
sialylation may be associated with an advanced clinical stage of
lymphoma cells which is related to invasiveness of tumor cells.
From the present study galectin-1 mediated cell invasive capacity
appeared to be regulated by cell surface sialylation and
oversialylation may facilitate cell invasion through galectin-1.
The biological role of galectin-1 such as extracellular matrix in
ALCL has not yet been published. This is the first report that
galectin-1 affects cell adhesion or invasion as extracellular
matrix in ALCL.
The CD45 is a candidate glycoprotein of galectin-1
receptor (22). In our preliminary
data from the antibody inhibition assay, we demonstrated that
anti-CD30, CD45, CD45RO and EMA (one of galectin-3 receptors)
antibodies enhanced cell adhesion to galectin-1 suggesting that
these antibodies possess agonistic effects and the protein portions
of ALK, CD30, CD45, CD45RO and EMA molecules may contribute to
galectin-1 mediated cell adhesion. There is a possibility that the
protein portions of glycoproteins can affect interaction between
galectin-1 and cell surface glycoproteins.
On the kinase inhibitor analysis, wortmannin showed
complete inhibition of cell adhesion to galectin-1, and PD98059
showed marked inhibition of invasion to galectin-1. The PI3K is
known to regulate cell adhesion (23) and the MAPK is known to regulate
cell migration (24). Therefore,
PI3K may regulate the galectin-1-mediated cell adhesion and MAPK
may regulate the galectin-1-mediated cell migration in H-ALCL
cells. As cytochalasin B disrupts polymerization of actin which is
a cytoskeleton, effect of cytochalasin B on cell adhesion to
galectin-1 implies that cell adhesion to galectin-1 may be mediated
by actin polymerization in H-ALCL cells. Rho (Ras homologue gene
family member) is a protein which is associated with lymphocyte
migration (25). In our present
data Rho inhibitor, C3-transferase dramatically inhibits cell
invasion through galectin-1. Rho may be a candidate regulator for
cell migration of H-ALCL cells contacting galectin-1.
Sialylation was reported to regulate interaction
between galectin-3 and ligand in tumor cells and the mechanisms are
crucial in regulating adhesive and de-adhesive events in the
invasive capacity of metastatic cells (26). We hypothesized that the masking
effect of ligand by sialylation may be important in cell adhesion
and invasion to galectins which are known to deposit in the tumor
environment. Sialylation may act as switch on/off mechanism in the
interaction between galectin and its oligosaccharide ligands. This
hypothesis may provide a new scientific foundation in
understandings the mechanisms of lymphoma cell biology, especially
lymphoma cell adhesion and invasion.
Treatment of SW, a potent N-glycosylation inhibitor
showed marked enhancement of cell adhesive capacity to galectin-1
suggesting that mature N-glycans which have sialylated glycans
inhibit cell adhesion to galectin-1 in ALCL. Previously, we
reported that SW treatment showed enhancement of lymphoma cells to
galectin-1 in the human diffuse large B cell lymphoma cell line
HBL-2 (7). Collectively, these
data suggest that cell surface N-glycans can modulate cell adhesive
capacity to galectin-1 in lymphoma cells, regardless of
histological subtypes.
In eukaryotic cells the glycans mathematically show
more than one million structures and due to this complexity the
glycomic analysis is required to glycobiological investigation
(27). In the present glycomics
analysis, cell adhesion to galectin-1 was modulated by several
types of lectin blocking suggesting that galectin-1-mediated cell
adhesion may be regulated by several types of glycans as well as
β-galactose residue. PNA, Jacalin, SBA, HPA, VVA, UEA-1, WGA,
L-PHA, E-PHA and DSA lectin blocking showed modulation of cell
adhesive properties to galectin-1. These findings may be supported
by the hypothesis that several cell surface glycans may interfere
with β-galactose by physical disturbance, such as steric hindrance.
PNA and SBA lectin reactive β-galactose may be a receptor for
galectin-1. On the other hand, the glycans reactive for Jacalin,
UEA-1, WGA, ConA, L-PHA, E-PHA and DSA lectins may interfere the
binding activity of H-ALCL cell to galectin-1 by steric hindrance.
The steric hindrance by several different glycans may modulate cell
adhesive properties to galectin-1. This may be a new concept of the
extremely complex glycobiology.
In Hodgkin lymphoma galectin-1 is a predictive
marker for evaluation of prognosis of the patients (28). The expression of galectin-1 is
reported and galectin-1 may facilitate the immunosuppressive
microenvironment (29,30). Previously some reports show that
galectin-1 can induce apoptosis in T cells (18) and tumor galectin-1 suppresses T
cell immunity through induction of apoptosis of T cells (31). Galectin-1 induced cell death is
caspase-independent (32), and in
our preliminary data, the cell death induced by galectin-1 in
H-ALCL was caspase-independent (data not shown). In the present
study galectin-1 induced cell death after 72 h. In other research,
the expression of galectin-1 in ALCL and caspase-dependent
apoptosis induction by galectin-1 was reported recently (1). The data implies that galectin-1 is a
useful tool for therapy of ALCL. The galectin-1 receptor, CD45PTP
activity was inhibited by treatment with galectin-1 in H-ALCL
cells. The data are consistent with the previous reports showing
the interaction between galectin-1 and CD45 in Burkitt’s lymphoma
cell line (33). Our present
report is the first showing interaction between galectin-1 and
CD45PTP activity in ALCL cells.
Acknowledgements
We are grateful to Ms. M. Satoh and
Mrs. H. Kaneko for their technical assistant and advice.
References
|
1.
|
Suzuki O, Hirsch B, Abe M, Dürkop H and
Stein H: Galectin-1-mediated cell death is increased by
CD30-induced signaling in anaplastic large cell lymphoma cells but
not in Hodgkin lymphoma cells. Lab Invest. 92:191–199. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Suzuki O, Nozawa Y and Abe M: Regulatory
roles of altered N-and O-glycosylation of CD45 in
galectin-1-induced cell death in human diffuse large B cell
lymphoma. Int J Oncol. 26:1063–1068. 2005.
|
|
3.
|
Earl LA, Bi S and Baum LG: N- and
O-glycans modulate galectin-1 binding, CD45 signaling, and T cell
death. J Biol Chem. 285:2232–2244. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Rabinovich GA, Rubinstein N and Toscano
MA: Role of galectins in inflammatory and immunomodulatory
processes. Biochim Biophys Acta. 1572:274–284. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Allen HJ, Sucato D, Gottstine S, Kisailus
E, Nava H, Petrelli N, Castillo N and Wilson D: Localization of
endogenous beta-galactoside-binding lectin in human cells and
tissues. Tumor Biol. 12:52–60. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Skrincosky DM, Allen HJ and Bernacki RJ:
Galaptin-mediated adhesion of human ovarian carcinoma A121 cells
and detection of cellular galaptin-binding glycoproteins. Cancer
Res. 53:2667–2675. 1993.PubMed/NCBI
|
|
7.
|
Suzuki O, Nozawa Y and Abe M: The
regulatory roles of cell surface sialylation and N-glycans in human
B cell lymphoma cell adhesion to galectin-1. Int J Oncol.
28:155–160. 2006.PubMed/NCBI
|
|
8.
|
Landemarre L, Cancellieri P and Duverger
E: Cell surface lectin array: parameters affecting cell glycan
signature. Glycoconj J. 30:195–203. 2013. View Article : Google Scholar
|
|
9.
|
Suzuki O, Nozawa Y, Kawaguchi T and Abe M:
Phaseolus vulgaris leukoagglutinationg lectin-binding reactivity in
human diffuse large B cell lymphoma and its relevance to the
patient’s clinical outcome: lectin histochemistry and lectin blot
analysis. Pathol Int. 49:874–880. 1999.PubMed/NCBI
|
|
10.
|
Albini A, Iwamoto Y, Kleinman HK, Martin
GR, Aaronson SA, Kozlowski JM and McEwan RN: A rapid in vitro assay
for quantitating the invasive potential of tumor cells. Cancer Res.
47:3239–3245. 1987.PubMed/NCBI
|
|
11.
|
Horiguchi N, Arimoto K, Mizutani A,
Endo-Ichikawa Y, Nakada H and Taketani S: Galectin-1 induces cell
adhesion to the extracellular matrix and apoptosis of non-adherent
human colon cancer Colo201 cells. J Biochem. 134:869–874. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Espelt MV, Croci DO, Bacigalupo ML,
Carabias P, Manzi M, Elola MT, Muñoz MC, Dominici FP,
Wolfenstein-Todel C, Rabinovich GA and Troncoso MF: Novel roles of
galectin-1 in hepatocellular carcinoma cell adhesion, polarization,
and in vivo tumor growth. Hepatology. 53:2097–2106. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Sanchez-Ruderisch H, Detjen KM, Welzel M,
André S, Fischer C, Gabius HJ and Rosewicz S: Galectin-1 sensitizes
carcinoma cells to anoikis via the fibronectin receptor
α5β1-integrin. Cell Death Differ. 18:806–816. 2011.PubMed/NCBI
|
|
14.
|
Zhao XY, Chen TT, Xia L, Guo M, Xu Y, Yue
F, Jiang Y, Chen GQ and Zhao KW: Hypoxia inducible factor-1
mediates expression of galectin-1: the potential role in
migration/invasion of colorectal cancer cells. Carcinogenesis.
31:1367–1375. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Zhuo Y and Bellis SL: Emerging role of
alpha2,6-sialic acid as a negative regulator of galectin binding
and function. J Biol Chem. 286:5935–5941. 2011. View Article : Google Scholar
|
|
16.
|
Cui H, Lin Y, Yue L, Zhao X and Liu J:
Differential expression of the α2,3-sialic acid residues in breast
cancer is associated with metastatic potential. Oncol Rep.
25:1365–1371. 2011.
|
|
17.
|
Wang FL, Cui SX, Sun LP, Qu XJ, Xie YY,
Zhou L, Mu YL, Tang W and Wang YS: High expression of alpha 2,
3-linked sialic acid residues is associated with the metastatic
potential of human gastric cancer. Cancer Detect Prev. 32:437–443.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Dall’Olio F, Chiricolo M, Ceccarelli C,
Minni F, Marrano D and Santini D: Beta-galactoside alpha2,6
sialyltransferase in human colon cancer: contribution of multiple
transcripts to regulation of enzyme activity and reactivity with
Sambucus nigra agglutinin. Int J Cancer. 88:58–65. 2000.PubMed/NCBI
|
|
19.
|
Mondal S, Chandra S and Mandal C: Elevated
mRNA level of hST6Gal I and hST3Gal V positively correlates with
the high risk of pediatric acute leukemia. Leuk Res. 34:463–470.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Lee M, Park JJ, Ko YG and Lee YS: Cleavage
of ST6Gal I by radiation-induced BACE1 inhibits golgi-anchored
ST6Gal I-mediated sialylation of integrin β1 and migration in colon
cancer cells. Radiat Oncol. 7:472012.PubMed/NCBI
|
|
21.
|
Suzuki O, Nozawa Y, Kawaguchi T and Abe M:
UDP-GlcNAc2-epimerase regulates cell surface sialylation and cell
adhesion to extracellular matrix in Burkitt’s lymphoma. Int J
Oncol. 20:1005–1011. 2002.PubMed/NCBI
|
|
22.
|
Perillo NL, Pace KE, Seilhamer JJ and Baum
LG: Apoptosis of T cells mediated by galectin-1. Nature.
378:736–739. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Yamamoto H, Nishi N, Shoji H, Itoh A, Lu
LH, Hirashima M and Nakamura T: Induction of cell adhesion by
galectin-8 and its target molecules in Jurkat T-cells. J Biochem.
143:311–324. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Huang C, Jacobson K and Schaller MD: MAP
kinases and cell migration. J Cell Sci. 117:4619–4628. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Tybulewicz VL and Henderson RB: Rho family
GTPases and their regulators in lymphocytes. Nat Rev Immunol.
9:630–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26.
|
de Oliveira JT, de Matos AJ, Santos AL,
Pinto R, Gomes J, Hespanhol V, Chammas R, Manninen A, Bernardes ES,
Albuquerque Reis C, Rutteman G and Gärtner F: Sialylation regulates
galectin-3/ligand interplay during mammary tumour progression - a
case of targeted uncloaking. Int J Dev Biol. 55:823–834.
2011.PubMed/NCBI
|
|
27.
|
Varki A, Cummings RD, Esko JD, et al:
Glycomics. Essentials of Glycobiology. 2nd edition. Cold Spring
Harbor Laboratory Press; Cold Spring Harbor, NY: 2009
|
|
28.
|
Kamper P, Ludvigsen M, Bendix K,
Hamilton-Dutoit S, Rabinovich GA, Møller MB, Nyengaard JR, Honoré B
and d’Amore F: Proteomic analysis identifies galectin-1 as a
predictive biomarker for relapsed/refractory disease in classical
Hodgkin lymphoma. Blood. 117:6638–6649. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Rodig SJ, Ouyang J, Juszczynski P, Currie
T, Law K, Neuberg DS, Rabinovich GA, Shipp MA and Kutok JL:
AP1-dependent galectin-1 expression delineates classical Hodgkin
and anaplastic large cell lymphomas from other lymphoid
malignancies with shared molecular features. Clin Cancer Res.
14:3338–3344. 2008. View Article : Google Scholar
|
|
30.
|
Juszczynski P, Ouyang J, Monti S, Rodig
SJ, Takeyama K, Abramson J, Chen W, Kutok JL, Rabinovich GA and
Shipp MA: The AP1-dependent secretion of galectin-1 by Reed
Sternberg cells fosters immune privilege in classical Hodgkin
lymphoma. Proc Natl Acad Sci USA. 104:13134–13139. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Banh A, Zhang J, Cao H, Bouley DM, Kwok S,
Kong C, Giaccia AJ, Koong AC and Le QT: Tumor galectin-1 mediates
tumor growth and metastasis through regulation of T-cell apoptosis.
Cancer Res. 71:4423–4431. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Hahn HP, Pang M, He J, Hernandez JD, Yang
R-Y, Li L, Wang X, Liu F-T and Baum LG: Galectin-1 induces nuclear
translocation of endonuclease G in caspase- and cytochrome
c-independent T cell death. Cell Death Differ. 11:1277–1286. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Fouillit M, Joubert-Caron R, Poirier F,
Bourin P, Monostori E, Levi-Strauss M, Raphael M, Bladier D and
Caron M: Regulation of CD45-induced signaling by galectin-1 in
Burkitt lymphoma B cells. Glycobiology. 10:413–419. 2000.
View Article : Google Scholar : PubMed/NCBI
|