Introduction
Colon cancer remains a significant global health
concern; it is the third most common malignancy and the fourth most
common cause of cancer mortality (1–4).
Annually, it accounts for approximately 600,000 deaths worldwide.
However, the vast majority (~80%) of these cases can be ascribed to
environmental causes and are therefore potentially preventable
(5). For instance, physical
inactivity has been reported to account for 10% of all colon cancer
cases (6), whereas physical
activity has been associated with reduced risk for incidence of
colon cancer. This inverse relationship between physical activity
and colon cancer risk is supported by epidemiological studies as
well as controlled experimental studies using rodent models. For
example, 9 weeks of treadmill running has been reported to decrease
the total number of intestinal polyps as well as the number of
large polyps in the ApcMin/+
mouse model of intestinal tumorigenesis (7). Similarly a recent study in a
multiethnic colon cancer screening population reported that
exercising for 1 h per week was associated with a lower prevalence
of polyps and adenomas when compared to those who exercised less or
not at all (8).
The mechanisms responsible for a preventative effect
of exercise on colon cancer risk are complex and multifaceted. An
exercise-induced alteration in immune system function is one
possible mechanism that has not been widely explored. Macrophages,
cells of the innate immune system, have recently emerged as major
components in the development of colon cancer given their ability
to produce a wide array of inflammatory mediators with pro-tumoral
functions (9,10). In general, these cells have been
associated with poor prognosis in colon cancer (9–14).
For example, one study reported a reduction in macrophage
infiltration that was consistent with a decrease in the size of
colon polyps in an MCP-1 receptor deficient mouse model of
colitis-associated colon cancer (15). Further, in a clinical study it was
documented that macrophage accumulation within the tumor was
positively correlated with stage of progression (14). Consistent with these reports, we
have documented that MCP-1 deficient
ApcMin/+ mice have decreased
macrophage number in both the polyps and surrounding intestinal
tissue and this was associated with a reduction in total polyp and
large polyp number. Exercise has been reported to influence
macrophage behavior in various disease models. For instance, it has
been shown that exercise can reduce macrophage-mediated
inflammation in an obese mouse model (16). Similarly, aerobic exercise reverses
the arterial inflammation and macrophage infiltration that is
associated with aging (17).
However, while it is well established that macrophages play a
significant role in the pathogenesis of colon cancer and that
exercise can influence the macrophage response, there is very
little evidence on the benefits of exercise on macrophage behavior
in the presence of colon cancer.
T cells also play a significant role in
tumorigenesis and have been shown to be altered by exercise
(18,19). It is well recognized that cytotoxic
T lymphocytes (CTLs) constitute one of the most important effector
mechanisms of antitumor immunity (20,21).
For example, adoptive transfer of CD8+ cells controls
the growth of B16 melanoma in mice (22). On the other hand, regulatory T
cells (Tregs) have been associated with accelerated tumor growth
and immune evasion through their inhibitory actions on CTLs and
helper T cells (23,24). The T cell response to exercise has
been reported to be highly variable and is likely dependent on the
mode, intensity and duration of exercise. However, to date there is
no evidence on the effects of exercise on T cells in a mouse model
of colon cancer.
The purpose of this investigation was to examine the
effects of exercise on expression of markers associated with
macrophages and certain T cell subsets in a mouse model of
intestinal tumorigenesis and to relate this to polyp
characteristics. We used the
ApcMin/+ mouse, the most widely
used genetic mouse model for cancer studies that involve the
gastro-intestinal tract (25).
Since the Apc gene is mutated in a large percentage of human
colon cancer cases, this is a common model for studying factors
that may influence progression of colon cancer. The exercise
protocol was designed to mimic a lifestyle that encompasses daily
physical activity; mice were exercised for 1 h/day, 6 days/week,
for a 12-week period. We hypothesized that daily exercise would
reduce the expression of markers associated with macrophages and
alter the T cell expression profile in the tumor environment, and
that this would be associated with a reduction in polyp burden.
Materials and methods
Animals
ApcMin/+ mice on
a C57BL/6 background were originally purchased from Jackson
Laboratories (Bar Harbor, ME). All experimental mice
(ApcMin/+) were bred in the
University of South Carolina’s Center for Colon Cancer Research
(CCCR) Mouse Core Facility. Specifically,
ApcMin/+ male mice were bred
with female C57BL/6 mice to generate
ApcMin/+ mice. Offspring were
genotyped as heterozygotes by RT-PCR for the Apc gene by
taking tail snips at weaning. The primer sequences were sense,
5′-TGAGAAAGACAGAAGTTA-3′; and antisense, 5′-TTCCACTTTGGCATAAGGC-3′.
Male ApcMin/+ mice were used in
this experiment and were maintained on a 12:12 h light-dark cycle
in a low-stress environment (22°C, 50% humidity and low noise) and
provided food (AIN-76A) and water ad libitum. All animal
experimentation was approved by the University of South Carolina’s
Institutional Animal Care and Use Committee.
Exercise protocol
Male ApcMin/+
mice were randomly assigned to either the sedentary (Sed; n=9) or
exercise (Ex; n=6) treatment. Mice in the Ex group ran on the
treadmill at 15 m/min and at a 5% incline for 1 h per day (starting
at 7 pm), 6 days per week from 4 to 16 weeks of age. Mice in the
Sed group remained in their cages in the treadmill room throughout
the exercise bouts but were exposed to similar handling and noise
in an attempt to control for extraneous stresses, and were deprived
of food and water during the exercise sessions.
Tissue collection
Body weight was measured weekly from 4 to 16 weeks
of age. Mice were sacrificed for tissue collection as previously
described (26,27). Briefly, sections 1 and 4 of the
small intestine and the large intestine (section 5) were fixed in
10% buffered formalin (Fisher Scientific, Pittsburg, PA) for 24 h.
For intestinal section 3, mucosal scrapings were performed in
TRIzol reagent (Invitrogen, Carlsbad, CA). Samples were stored at
−80°C until analysis for the expression of markers associated with
macrophage phenotype and certain T cell subsets. Previously
reported findings from our laboratory have shown that intestinal
section 3 has a greater polyp incidence than section 2 (26), and therefore we included only
section 3 for these analyses. Visceral fat pads (retroperitoneal,
epididymal and mesentery) were dissected and weighed for the
measurement of total visceral fat as it has been reported that
ApcMin/+ mice become cachectic
with disease progression (28).
Spleen was also weighed as splenomegaly has been shown to be
associated with increased polyp number in this model (29). Blood was collected from the
inferior vena cava using a heparinized syringe and blood parameters
were examined immediately on fresh whole blood using a Vetscan
blood analyzer (Abaxis, Union City, CA).
Polyp counts
Formalin-fixed intestinal sections from all animals
were rinsed in deionized water, briefly stained in 0.1% methylene
blue, and counted by the same investigator who was blinded to the
treatments. Polyps were counted under a dissecting microscope and
were categorized according to size (>2, 1–2 and <1 mm).
mRNA gene expression
Quantification of gene expression for total
macrophages (F4/80), markers associated with macrophage phenotype
IL-12, IL-23 and Nos2 (M1 macrophage phenotype), CD206, IL-10,
IL-4, CCL17, CCL22 and Arg-1 (M2 macrophage phenotype), macrophage
chemoattractants (MCP-1, fetuin A and CXCL14), and T cell subsets
(CD8 and Foxp3) were performed as previously described (26,30).
Briefly, mucosal tissue was homogenized under liquid nitrogen with
a polytron, and total RNA was extracted using TRIzol reagent
(Invitrogen). The extracted RNA (2.5 μl of sample) was dissolved in
diethyl pyrocarbonate (DEPC)-treated water and quantified
spectrophotometrically at 260 nm wavelength. RNA was reverse
transcribed into cDNA and quantitative RT-PCR analysis was done per
manufacturer’s instructions (Applied Biosystems, Foster City, CA)
using TaqMan® Gene Expression Assays. Quantification of
cytokine gene expression was calculated using the ΔΔCT method
(31).
Statistical analysis
Analysis was done using commercially available
software (SigmaStat, SPSS, Chicago, IL). A two-way repeated
measures ANOVA was performed on body weight measurements. T-tests
were performed on all other data. Statistical significance was set
with an α-value of P<0.05. Data are presented as mean ± SEM.
Results
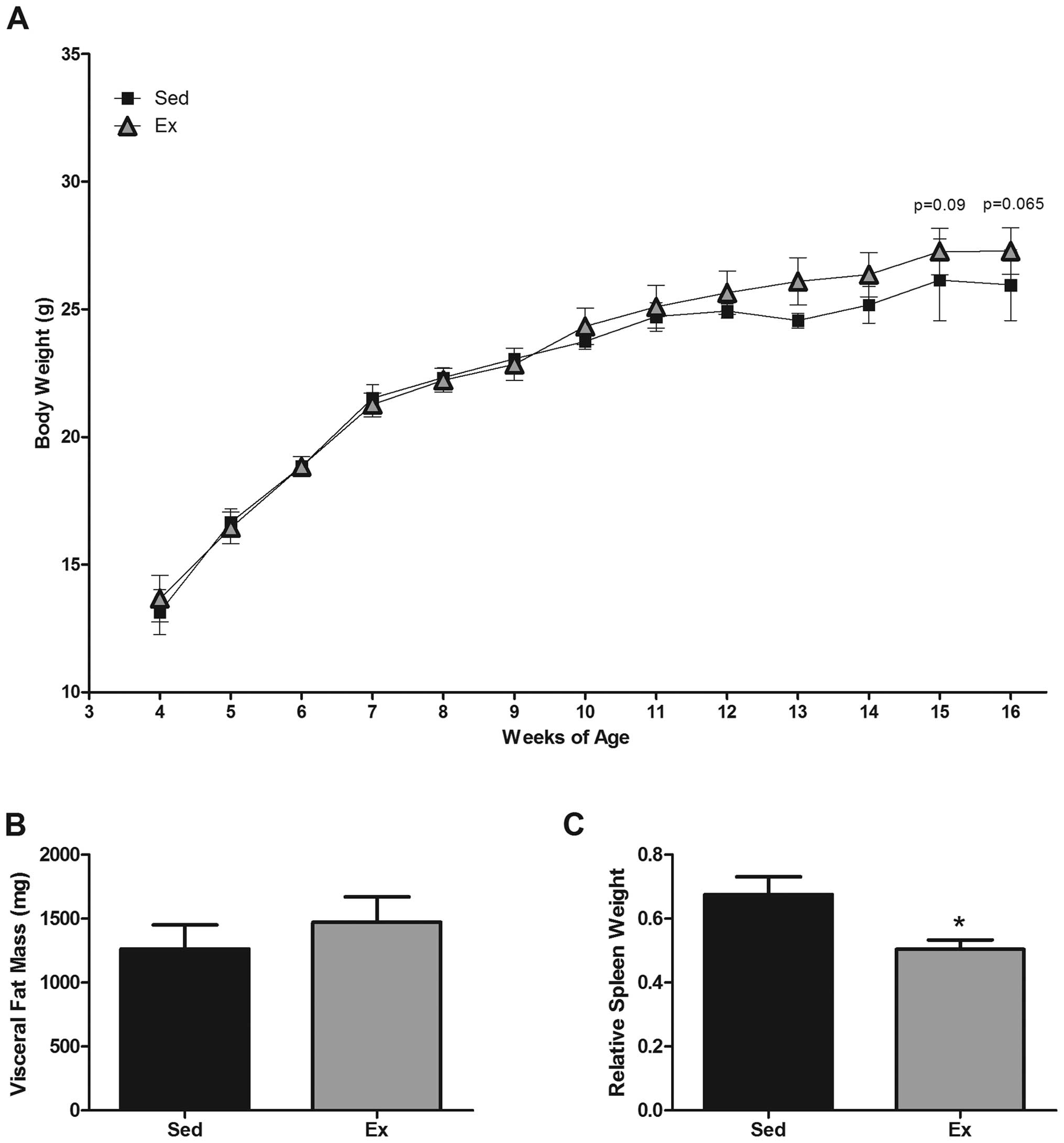
Body weight and visceral fat mass
It has been reported that
ApcMin/+ mice develop cachexia
that is positively correlated with polyp burden (32). Therefore, we examined the potential
benefits of exercise on body weight and visceral fat mass in the
ApcMin/+ mouse. At 15 and 16
weeks of age, there was an apparent difference (~1–2 g) between the
groups; the Sed group weighed 26.1±1.6 and 25.9±1.4 g at 15 and 16
weeks, respectively, whereas the Ex group weighed 27.3±0.9 and
27.3±0.9 g, respectively (P<0.1) (Fig. 1A). However, this did not quite
reach statistical significance. Visceral fat tissue
(retroperitoneal, epididymal and mesentery) was collected at
sacrifice and weighed to determine any influence of exercise on
cachexia-related fat mass. There was no protective effect of
exercise on total visceral fat pad weight (Sed, 1262.2±189.2 mg;
Ex, 1471.2±197.7 mg) (Fig.
1B).
Spleen weight
Increased spleen weight has been positively
associated with polyp burden in this model (29). Therefore, spleens were harvested
during sacrifice at 16 weeks of age, weighed and expressed relative
to body weight. Exercise significantly decreased spleen weight
versus the Sed group (0.5±0.0 versus 0.68±0.01 mg/kg, respectively)
(P=0.05) (Fig. 1C).
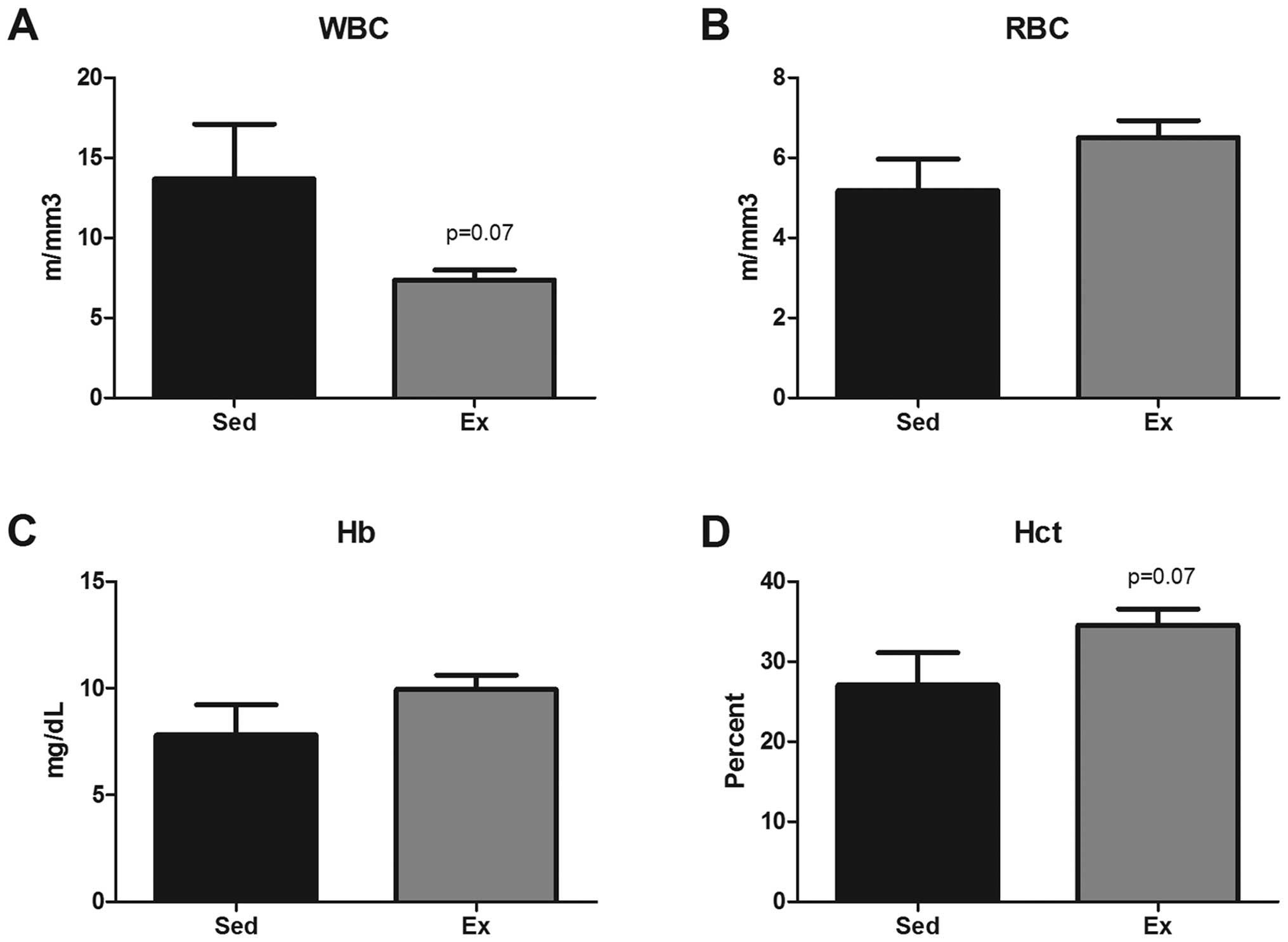
Complete blood count
A complete blood count was performed at sacrifice as
both white blood cell (WBC) and red blood cell (RBC) counts have
been shown to be altered during progression of intestinal
tumorigenesis in this mouse model (Fig. 2) (27). WBC count at 16 weeks of age tended
to be decreased by exercise (13.7±3.2 versus 7.4±0.6
m/mm3) (P=0.07). Similarly, there was a trend for an
exercise-induced increase in hematocrit (Hct) compared to Sed
(34.6±2.0 versus 27.1±3.8%) (P=0.07). But there were no apparent
differences among the groups for RBCs or hemoglobin (Hb).
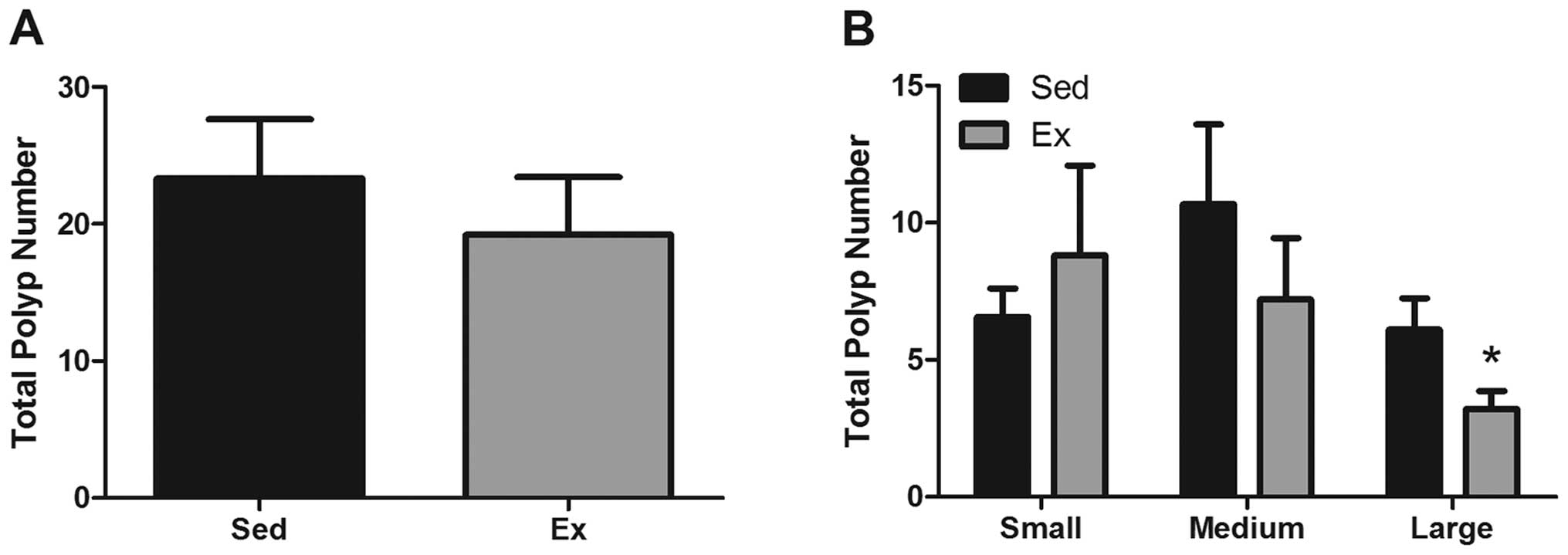
Polyp incidence
At 16 weeks of age, mice (Sed and Ex) were
sacrificed, intestinal tissue was harvested and polyps were counted
on formalin-fixed, methylene blue-stained sections. Overall polyp
number (sections 1, 4 and 5) was not significantly changed by
exercise (23.3±4.3 versus 19.2±4.2 for Sed and Ex, respectively)
(Fig. 3A). To examine polyp size
(Fig. 3B), we counted and
classified polyps as being large (>1 mm in diameter), medium
(<2>1 mm in diameter) or small (<1 mm in diameter).
Interestingly, we found a significant reduction in the number of
large polyps with exercise; the Ex group had 48% fewer large polyps
than the Sed group (3.2±0.7 versus 6.1±1.1) (P<0.05) but there
were no significant differences in the number of small or medium
polyps.
Macrophage number and phenotypic
markers
Gene expression of macrophage phenotypic markers
IL-12, IL-23 and Nos2 (M1 macrophage phenotype), CD206, IL-10,
IL-4, CCL17, CCL22 and Arg-1 (M2 macrophage phenotype), were
examined in the mucosal scrapings (Fig. 4A) of intestinal tissue. Data were
normalized to fold-change from Sed mice. There was a significant
decrease in mRNA expression of F4/80, a general macrophage marker
(P<0.05). Similarly, there was a decrease in mRNA expression of
the M2 associated macrophage markers, CD206, CCL22 and Arg in the
Ex mice (P<0.05), and a trend for a decrease in CCL17
(P<0.06). Even though not all markers associated with M2
macrophages were significantly reduced with exercise, all were
consistently decreased. IL-12, a marker associated with the M1
macrophage phenotype was also decreased in the Ex mice (P<0.05)
but there was no change in IL-23 or Nos2. Gene expression of
macrophage-associated chemokines MCP-1, fetuin A and CXCL14 were
also examined in the mucosal scrapings. While mRNA expression of
each of these markers was decreased with exercise (Fig. 4B), statistical significance was not
reached.
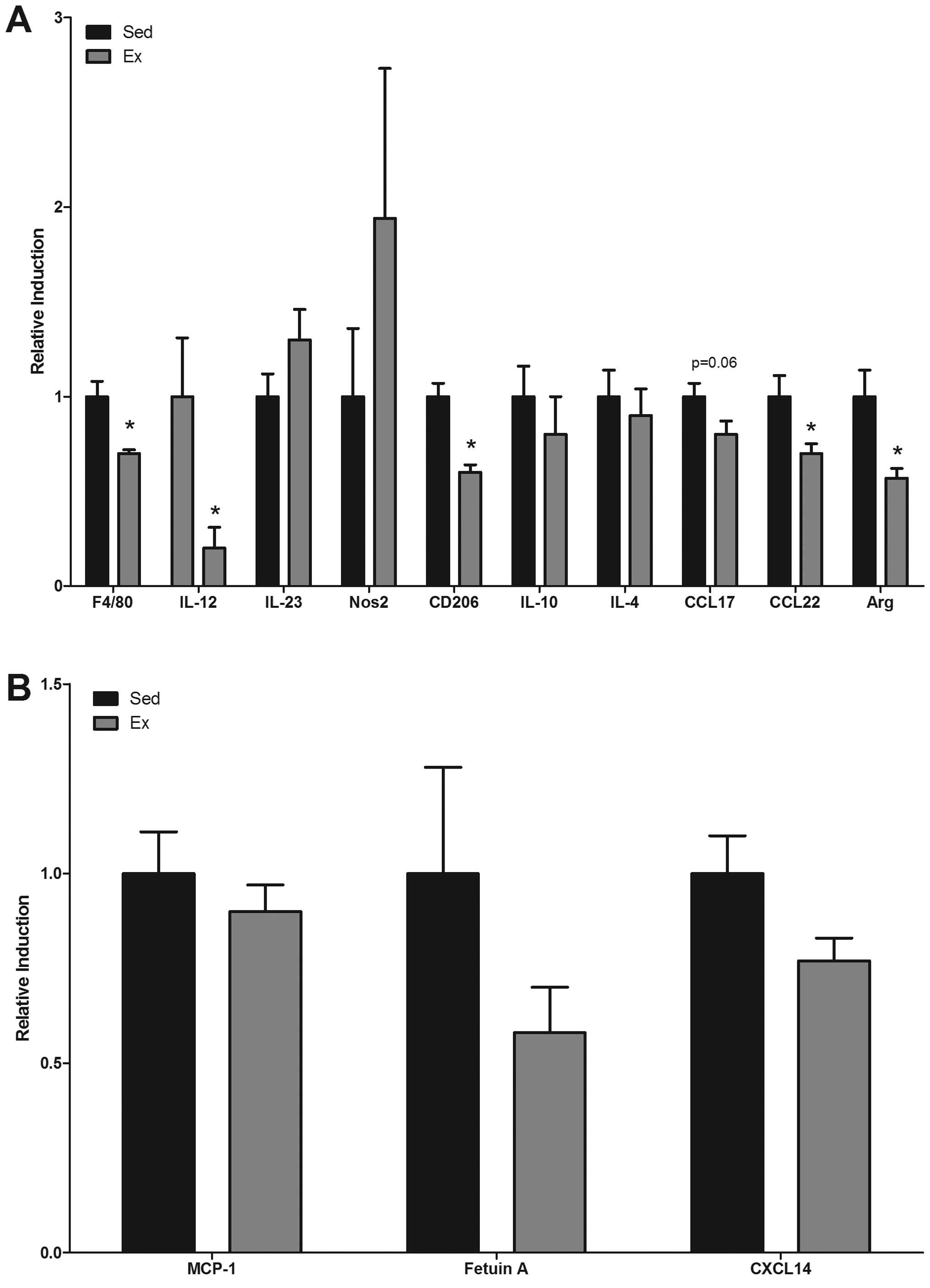 | Figure 4Effects of exercise on gene
expression of M1 and M2 associated phenotypic macrophage markers in
ApcMin/+ mice. Differences in
gene expression of (A) F4/80, M1 macrophage markers (IL-12, IL-23,
and NOS2), M2 macrophage markers (CD206, IL-10, IL-4, CCL17, CCL22
and Arg) and (B) macrophage chemattractants (MCP-1, Fetuin A, and
CXCL14) were examined in sedentary (Sed) and exercised (Ex)
ApcMin/+ mice (n=6–9/group) at
16 weeks of age in mucosal scrapings. Values are means ± SEM.
*P<0.05 significant difference. |
Changes in T cell expression
Given the role of T cell subsets in tumorigenesis,
we also performed gene expression analysis of markers associated
with CTLs (CD8) and Tregs (Foxp3) (Fi. 5). CD8, a marker for CTLs
that represents one of the most important effector mechanisms of
antitumor immunity, was increased with exercise (P<0.05).
Conversely, Foxp3, a marker for Tregs that are known to suppress
immune function and that have been associated with increased
tumorigenesis, was decreased with exercise (P<0.05).
Discussion
There is an inverse relationship between physical
activity and colon cancer risk (33). A multitude of mechanisms, including
immune function dysregulation, have been implicated in this
response. Macrophages and T cells play a significant role in the
pathogenesis of colon cancer and exercise can influence the actions
of these cells; however, there is very little evidence on the
benefits of exercise on macrophage and T cell responses in the
settings of colon cancer. We examined the effects of exercise on
markers associated with macrophages and select T cell subsets in a
mouse model of intestinal tumorigenesis in relation to polyp
characteristics. Overall, certain markers associated with both the
M1 and the M2 macrophage phenotype were reduced in
ApcMin/+ mice following
exercise. Additionally, exercise resulted in an increased
expression of CD8 and decreased expression of Foxp3, markers for
CTLs and Tregs, respectively. These alterations in immune cell
parameters following exercise training were accompanied by a
decrease in the percentage of large polyps.
Animal models provide a tool to examine the effects
of exercise on colon cancer in an experimental environment in which
the type and intensity of exercise can be controlled. They allow
for detailed study of stage-specific responses to exercise, and
help to identify the optimal mode, intensity and duration of
exercise. The benefits of exercise on colon cancer risk have been
well documented in the ApcMin/+
mouse model of intestinal tumorigenesis (7,28,34,35).
For example, 9 weeks of treadmill running has been reported to
decrease the total number of intestinal polyps by 29% as well as
the number of large polyps (38%) in male mice in this model
(36). Similarly, exercise was
reported to reduce total intestinal polyp number by 50% and the
number of large polyps by 67% in this same model (34). Our findings are somewhat consistent
with these investigations in that we report a 48% reduction in the
number of large polyps. In contrast to the findings by Baltgalvis
et al (7), we did not find
a significant reduction in the number of total polyps; however,
this is likely due to the smaller sample size in our study and/or
to the slightly lower intensity of the exercise protocol (15 versus
18 m/min). Nonetheless, the benefits of regular exercise training
in the ApcMin/+ mouse model of
intestinal tumorigenesis are evident across studies and it is clear
from our findings and those of others that exercise plays a larger
role in reducing the progression of growth as opposed to the
initiation of development, at least in this model.
In addition to polyp characteristics, we measured
body weight, fat mass, spleen weight and markers of anemia. These
outcomes have been associated with increased tumorigenesis and
ultimately poorer prognosis in the
ApcMin/+ mouse (27,32).
Previous published data have shown that
ApcMin/+ mice develop cachexia
that is positively correlated with polyp burden (32). In our study, the exercise mice
tended to be heavier than the sedentary mice at 15 and 16 weeks of
age, although this did not reach statistical significance. Further,
there were no differences in fat mass between the groups. However,
the lack of positive findings is likely due to the timing of
sacrifice as mice were sacrificed prior to the onset of severe
cachexia. This was done to eliminate any possible influence of
cachexia or illness on the ability to perform the exercise
protocol. Therefore, it is not surprising that we did not see a
statistically significant effect of exercise at these time points.
Spleen weight has been associated with increased polyp number and
systemic inflammation (36). Our
data indicate a reduction in spleen weight with exercise. This is
consistent with previously reported literature; Baltgalvis et
al (36) also reported a
reduction in spleen weight in male
ApcMin/+ mice following
exercise. We have recently reported an increase in markers of
anemia in this mouse model (27).
While exercise did prevent the characteristic decrease in Hct in
these mice, the effect was not found to be statistically
significant. Again, this is likely due to the timing of sacrifice
as in our previous study mice were sacrificed at 18 weeks, a time
in which the disease is much more severe. Thus, any benefits of
exercise would likely be more evident had the mice been sacrificed
at a later time point.
We next examined the effects of exercise on markers
associated with macrophages in the mucosal tissue. Macrophages can
represent up to 50% of the tumor mass producing a wide array of
inflammatory mediators with pro-tumoral functions (9,10).
Further, abundance of tumor associated macrophages has been
associated with poor prognosis in colon cancer (9–14).
Our data show a reduction in the expression of F4/80, an overall
macrophage marker, with exercise. This is consistent with a
previous study by Baltgalvis et al that reported a reduction
in macrophage number following exercise training in this model
(7). Because it is now well
accepted that macrophages constitute an extremely heterogeneous
population that is divided into two main classes (M1 and M2)
(10), we next examined the
effects of exercise on expression of markers associated with both
the M1 and M2 phenotype. In general, it is thought that M1
macrophages are cytotoxic against neoplastic cells, whereas M2
macrophages exert pro-tumoral functions (10). We report the novel finding that
exercise reduces the expression of certain markers that are
associated with the M1 (IL-12) and M2 (CD206, CCL17, CCL22 and
Arg-1) phenotype in the mucosal tissue. It is important to note
that given the limited available tissue, macrophage markers were
not measured in the polyps themselves. However, previous data from
our group show a similar response for these outcomes when comparing
the mucosal tissue and polyp tissue in this model (27). While our data suggest that a
reduction in both M1 and M2 macrophages with exercise is associated
with a reduction in polyp growth in this model, a greater
understanding of the roles of each macrophage subset within the
tumor microenvironment is necessary.
Given the reduction in macrophage markers with
exercise, we next examined the effects of exercise on macrophage
chemoattractants. MCP-1 is a major player in macrophage chemotaxis
in the ApcMin/+ mouse (27). In fact, we recently reported a link
between macrophages and MCP-1 in this mouse model (27). Although our data indicate an
exercise-induced decrease in the expression of macrophage
associated markers in the mucosal tissue, we did not find a
significant decrease in MCP-1. Therefore, we also examined fetuin A
and CXCL14, both of which have been implicated in macrophage
recruitment. Fetuin A is a recently characterized macrophage
chemoattractant that is also known to play a role in macrophage
polarization (37). While our
results show a decrease in the expression of fetuin A that is
consistent with our macrophage findings, this did not reach
statistical significance. Similarly, there was no effect of
exercise on CXCL14, a chemoattractant for activated tissue
macrophages. It is important to point out that only a subset of
macrophage chemoattractants were measured in this study, thus it is
possible that exercise may have impacted other chemokines.
One of the most important effector mechanisms of
antitumor immunity is the activities of CTLs (20,21).
For example, growth of B16 melanoma cells can be controlled in mice
following the transfer of CD8+ cells (22). On the other hand, Tregs have been
linked to accelerated tumor growth and immune evasion due to their
inhibitory actions on CTLs and helper T cells (23,24).
The findings of the current study show an increased expression of
CD8, and conversely, a decreased expression of Foxp3 in the mucosal
tissue following exercise training. While this is the first report
of a favorable effect of exercise on CTLs and Tregs in a model of
colon cancer, the evidence supporting exercise-induced alterations
in immune function is far reaching. In general, regular moderate
exercise is thought to enhance immune function; however, these
effects are likely dependent on a multitude of factors including
individual characteristics, the type, intensity and duration of
exercise, and the stage of cancer. Although these findings support
a positive effect of exercise on the T cell profile in a cancer
model, it is important to point out that whether this is due to a
direct effect of the exercise on these cell populations or an
indirect effect that results from the reduction in large polyp
number resulting from an entirely different mechanism could not be
determined from this study.
Consistent with previous reports, we show a benefit
of exercise training on reducing large polyp number in the
ApcMin/+ mouse model of
intestinal tumorigenesis. This was associated with alterations in
the expression of immune markers in the mucosal tissue including a
reduction in markers associated with M1 and M2 macrophages, an
increase CD8 and a decrease in Foxp3. Overall this data provide
important new information on immune regulation as a possible
mechanism for the benefits of exercise training on reducing colon
cancer progression.
Acknowledgements
This study was supported by a grant from the
American Institute for Cancer Research (10A070) to E.A.M.
References
|
1
|
Tenesa A and Dunlop MG: New insights into
the aetiology of colorectal cancer from genome-wide association
studies. Nat Rev Genet. 10:353–358. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Center MM, Ward E, et al: Cancer
occurrence. Methods Mol Biol. 471:3–29. 2009. View Article : Google Scholar
|
|
3
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics, 2008. CA Cancer J Clin. 58:71–96. 2008. View Article : Google Scholar
|
|
4
|
Howlader N, Noone AM, Krapcho M, et al:
SEER Cancer Statistics Review, 1975–2008. National Cancer
Institute; Bethesda, MD: 2010
|
|
5
|
Rustgi AK: The genetics of hereditary
colon cancer. Genes Dev. 21:2525–2538. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee IM, Shiroma EJ, Lobelo F, et al:
Effect of physical inactivity on major non-communicable diseases
worldwide: an analysis of burden of disease and life expectancy.
Lancet. 380:219–229. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Baltgalvis KA, Berger FG, Pena MM, et al:
Effect of exercise on biological pathways in ApcMin/+ mouse
intestinal polyps. J Appl Physiol (1985). 104:1137–1143. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sanchez NF, Stierman B, Saab S, et al:
Physical activity reduces risk for colon polyps in a multiethnic
colorectal cancer screening population. BMC Res Notes. 5:3122012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mantovani A and Sica A: Macrophages,
innate immunity and cancer: balance, tolerance, and diversity. Curr
Opin Immunol. 22:231–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Solinas G, Germano G, Mantovani A, et al:
Tumor-associated macrophages (TAM) as major players of the
cancer-related inflammation. J Leukoc Biol. 86:1065–1073. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jedinak A, Dudhgaonkar S and Sliva D:
Activated macrophages induce metastatic behavior of colon cancer
cells. Immunobiology. 215:242–249. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kaler P, Augenlicht L and Klampfer L:
Macrophage-derived IL-1beta stimulates Wnt signaling and growth of
colon cancer cells: a crosstalk interrupted by vitamin D3.
Oncogene. 28:3892–3902. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kang JC, Chen JS, Lee CH, et al:
Intratumoral macrophage counts correlate with tumor progression in
colorectal cancer. J Surg Oncol. 102:242–248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bailey C, Negus R, Morris A, et al:
Chemokine expression is associated with the accumulation of tumour
associated macrophages (TAMs) and progression in human colorectal
cancer. Clin Exp Metastasis. 24:121–130. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Popivanova BK, Kostadinova FI, Furuichi K,
et al: Blockade of a chemokine, CCL2, reduces chronic
colitis-associated carcinogenesis in mice. Cancer Res.
69:7884–7892. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawanishi N, Yano H, Mizokami T, et al:
Exercise training attenuates hepatic inflammation, fibrosis and
macrophage infiltration during diet induced-obesity in mice. Brain
Behav Immun. 26:931–941. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lesniewski LA, Durrant JR, Connell ML, et
al: Aerobic exercise reverses arterial inflammation with aging in
mice. Am J Physiol Heart Circ Physiol. 301:H1025–H1032. 2011.
View Article : Google Scholar
|
|
18
|
Rogers CJ, Zaharoff DA, Hance KW, et al:
Exercise enhances vaccine-induced antigen-specific T cell
responses. Vaccine. 26:5407–5415. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Woodland DL, Hogan RJ and Zhong W:
Cellular immunity and memory to respiratory virus infections.
Immunol Res. 24:53–67. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Weigelin B, Krause M and Friedl P:
Cytotoxic T lymphocyte migration and effector function in the tumor
microenvironment. Immunol Lett. 138:19–21. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Swann JB and Smyth MJ: Immune surveillance
of tumors. J Clin Invest. 117:1137–1146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garcia-Hernandez Mde L, Hamada H, Reome
JB, et al: Adoptive transfer of tumor-specific Tc17 effector T
cells controls the growth of B16 melanoma in mice. J Immunol.
184:4215–4227. 2010.PubMed/NCBI
|
|
23
|
Salama P, Phillips M, Grieu F, et al:
Tumor-infiltrating FOXP3+ T regulatory cells show strong
prognostic significance in colorectal cancer. J Clin Oncol.
27:186–192. 2009.
|
|
24
|
Curiel TJ: Tregs and rethinking cancer
immunotherapy. J Clin Invest. 117:1167–1174. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tammariello AE and Milner JA: Mouse models
for unraveling the importance of diet in colon cancer prevention. J
Nutr Biochem. 21:77–88. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McClellan JL, Davis JM, Steiner JL, et al:
Intestinal inflammatory cytokine response in relation to
tumorigenesis in the Apc(Min/+) mouse. Cytokine. 57:113–119. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McClellan JL, Davis JM, Steiner JL, et al:
Linking tumor-associated macrophages, inflammation, and intestinal
tumorigenesis: role of MCP-1. Am J Physiol Gastrointest Liver
Physiol. 303:G1087–G1095. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baltgalvis KA, Berger FG, Pena MM, et al:
Activity level, apoptosis, and development of cachexia in
Apc(Min/+) mice. J Appl Physiol (1985). 109:1155–1161. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Murphy EA, Davis JM, McClellan JL, et al:
Quercetin’s effects on intestinal polyp multiplicity and macrophage
number in the Apc(Min/+) mouse. Nutr Cancer. 63:421–426. 2011.
|
|
30
|
Murphy EA, Davis JM, McClellan JL, et al:
Curcumin’s effect on intestinal inflammation and tumorigenesis in
the ApcMin/+ mouse. J Interferon Cytokine Res. 31:219–226.
2011.
|
|
31
|
Nieman DC, Henson DA, Davis JM, et al:
Quercetin’s influence on exercise-induced changes in plasma
cytokines and muscle and leukocyte cytokine mRNA. J Appl Physiol
(1985). 103:1728–1735. 2007.
|
|
32
|
Baltgalvis KA, Berger FG, Pena MM, et al:
Interleukin-6 and cachexia in ApcMin/+ mice. Am J Physiol Regul
Integr Comp Physiol. 294:R393–R401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Song JH, Kim YS, Yang SY, et al: Physical
activity and other lifestyle factors in relation to the prevalence
of colorectal adenoma: a colonoscopy-based study in asymptomatic
Koreans. Cancer Causes Control. 24:1717–1726. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Baltgalvis KA, Berger FG, Pena MM, et al:
The interaction of a high-fat diet and regular moderate intensity
exercise on intestinal polyp development in Apc Min/+ mice. Cancer
Prev Res (Phila). 2:641–649. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Puppa MJ, White JP, Velazquez KT, et al:
The effect of exercise on IL-6-induced cachexia in the Apc (Min/+)
mouse. J Cachexia Sarcopenia Muscle. 3:117–137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mehl KA, Davis JM, Clements JM, et al:
Decreased intestinal polyp multiplicity is related to exercise mode
and gender in ApcMin/+ mice. J Appl Physiol (1985). 98:2219–2225.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chatterjee P, Seal S, Mukherjee S, et al:
Adipocyte fetuin-A contributes to macrophage migration into adipose
tissue and polarization of macrophages. J Biol Chem.
288:28324–28330. 2013. View Article : Google Scholar : PubMed/NCBI
|