Introduction
Malignant gliomas are the most common primary
neoplasms of the central nervous system in adults. The median
survival time is about a year and half from diagnosis, despite the
modern treatment including surgery, postoperative radiotherapy
combined with temozolomide-based chemotherapy (1,2).
Although improvement of survival time by maximum tumor resection
has been reported (3,4), complete surgical removal is not
possible because of the invasive nature of gliomas to the
surrounding eloquent brain. Residual tumor cells are usually
resistant to standard radiotherapy and efficient chemotherapy
cannot be delivered because of the presence of the blood-brain
barrier and systemic toxicity. Most patients thus die of tumor
proliferation after a certain period. To kill those residual tumor
cells by increasing the concentration of chemotherapeutic agents in
the tumor cells without increasing systemic toxicity, local
treatment with biodegradable 1,3-bis (2-chloroethyl)-1-nitrosourea
wafers (Gliadel wafers) was developed and prolonged survival in
patients with recurrent glioblastoma multiforme, the most malignant
phenotype of gliomas, was demonstrated (5). However, spread of the drug from
implanted Gliadel wafers to the surrounding brain tissues is still
limited within millimeter order (6).
In order to expand the treatment area, our previous
studies have focused on testing neural stem cells (NSCs) and
mesenchymal stem cells (MSCs) that have potent migratory activity
toward brain lesions including tumors (7,8). Rat
brain tumors were successfully treated by an intratumoral injection
of genetically engineered NSCs and MSCs (9,10),
and this efficacy was still potent even when the treatment cells
were injected at distant intracranial sites from the tumor
(11). Therefore, treatment
strategies using such stem cells as vehicles for antitumor agents
may be promising against malignant glioma because it is a
tumor-specific delivery system that spares healthy brain tissue
(12,13). However, NSCs and MSCs proliferate
relatively slowly and it is not easy to prepare a sufficient amount
of cells for treatment.
In 2007, induced pluripotent stem cells (iPSCs) were
established. The usefulness of iPSC-based therapies in various
fields including regenerative medicine has been investigated
(14,15). Because of high proliferative
activity of iPSCs, sufficient amount of treatment cells can be
prepared. Our previous study demonstrated in vitro migratory
activity of mouse iPSCs to glioma-conditioned media and
tumor-associated specific growth factors (16). In the present study, in vivo
tumor tropic activity of iPSCs and iPSC-derived neural stem-like
cells were investigated using a mouse intracranial glioma
model.
Materials and methods
iPSC and neural induction
The mouse iPSCs, iPS-MEF-Ng-20D-17 established by
Okita et al (15) and
transfected with Nanog-GFP as undifferentiation marker, were
obtained from Riken BioResource Center (Tsukuba, Japan) and were
cultured on mitomycin-treated mouse embryonic fibroblasts feeder
cell layers in Dulbecco’s modified Eagle’s medium (DMEM;
Sigma-Aldrich Japan, Tokyo, Japan) high glucose 1× (Invitrogen,
Tokyo, Japan) supplemented with 15% fetal bovine serum (FBS;
Sigma-Aldrich Japan), 0.1 mM MEM non-essential amino acids
(Invitrogen), 0.1 mM 2-mercaptoethanol (Sigma-Aldrich Japan) and
1,000 U/ml murine leukemia inhibitory factor (mLIF, ESGRO;
Millipore, Temecula, CA, USA) according to the protocol previously
reported (17). The iPSCs cultured
on the gelatin-coated dish without mouse embryonic fibroblasts
(passages 10–20) were used in the following experiments. The iPSCs
were dissociated at 37°C for 2 min using 0.25% trypsin with 1 mM
EDTA.
The adherent monoculture method was used as a
differentiation multistep protocol to generate neural stem-like
cells from iPSCs (18,19). The mouse iPSCs were dissociated and
plated onto 0.1% gelatin-coated 100-mm dish at a density of
1.2×106/dish and cultured in N2B27 medium that contained
a 1:1 mixture of DMEM/F12 supplemented with 1% N2 (Invitrogen) and
neurobasal medium supplemented with 2% B27 (Invitrogen). Medium was
renewed every day. Cells were maintained for 12 days and then were
dissociated and plated on 0.1% gelatin-coated 100-mm dish in NSC
expansion medium consisting DMEM/F12 medium supplemented with
GlutaMAX (Invitrogen), 2% B27, 50 U/ml penicillin, 50 μg/ml
streptomycin, 20 ng/ml basic fibroblast growth factor
(Sigma-Aldrich Japan) and 20 ng/ml epidermal growth factor
(Invitrogen). Half of the cell-culture medium was changed every 2
days and cells were split at the ratio of 1:2 when reached 90%
confluence using Accutase (Sigma-Aldrich Japan). In the following
experiment, the iPSC-derived neural stem-like cells during 10–20
passages were used (named as iPS-NSCs).
Mouse glioma sphere cells
The mouse glioma cell line GL261 and the rat glioma
cell line C6 were purchased from the Health Science Research
Resources Bank (Osaka, Japan), and the human glioma cell lines U87
from the American Type Culture Collection (ATCC, Manassas, VA,
USA). The cells were grown in DMEM supplemented with 10% FBS,
penicillin (100 IU/ml) and streptomycin (100 μg/ml) at 37°C in a
humidified atmosphere of 5% CO2. The glioma cell lines
were dissociated using 0.25% trypsin with 1 mM EDTA for 3 min.
For establishment of GL261 sphere cells (20,21),
GL261 cells were seeded in 24-well plates at 2×104
cells/well (BD Falcon) containing 500 μl DMEM, supplemented with
10% FBS, 100 U/ml penicillin G and 100 μg/ml streptomycin for
around 18 h. Culture medium (250 μl) was replaced with an equal
volume of NSC expansion medium with 1,000 U/ml mLIF and vincristine
(5 ng/ml; Wako). This procedure was repeated every 24 h until day
7. Then all culture medium was discarded and the wells were filled
with 1 ml fresh NSC expansion medium with 1,000 U/ml mLIF (without
vincristine) on day 7 and changed every 3 days. On day 15 after
filled with NSC expansion medium with 1,000 U/ml mLIF, the spheres
were dissociated and single cells were seeded in 96-well plates in
0.1 ml/well of serum-free NSC medium. Every 3 days, 0.02 ml NSC
medium was added to the culture. The cells from the well containing
spheres with >50 cells were dissociated and cultured for
expansion in NSC expansion medium with 1,000 U/ml mLIF.
Migration of iPSCs and iPS-NSCs towards
the glioma-conditioned media
The in vitro migratory capacity of iPSCs was
assessed using the 24-well Matrigel invasion chamber (BD
Biosciences Discovery Labware, Bedford, MA, USA), which contained
an 8-μm pore size PET membrane treated with Matrigel basement
membrane matrix in the insert (16). First, 0.5 ml DMEM was added to the
interior of the inserts and the bottom of the wells and allowed to
rehydrate for 2 h at 37°C in a 5% CO2 humidified
atmosphere after the gelatin coated both membranes. The DMEM was
then carefully removed without disturbing the layer of Matrigel
matrix on the membrane. The mouse iPSCs were washed twice in
phosphate-buffered saline (PBS) and resuspended to 2×106
cells/ml. The cell suspension (0.5 ml) (1×106 cells) was
then added to the upper insert. The lower chamber was filled with
0.75 ml of conditioned medium (CM) of the glioma cell lines as well
as unconditioned DMEM as a control. CM was obtained by collecting,
centrifuging and filtering medium from GL261, C6 and U87 clones
(1×106), which were cultured in 10 ml of DMEM without
FBS for 48 h. Following incubation of Matrigel invasion chambers
for 24 h at 37°C in a 5% CO2 humidified atmosphere, the
non-invading cells and/or Matrigel matrix were removed from the
upper surface of the membrane in the inserts with a cotton swab.
The cells migrating to the lower surface of the membrane were
stained with Diff-Quick kit (International Reagents, Hyogo, Japan),
which was achieved by sequentially transferring the inserts to air
dry. The nuclei of migrated cells were counted in 4 high-power
fields (HPF) per membrane with ×200 magnification. The images of
Nanog-GFP expression in the migrated cell were acquired with EVOS
fluorescence microscope (LMS Co., Ltd., Tokyo, Japan). All
experiments were conducted in triplicate and results were expressed
as the mean number of cells migrating per field ± SD.
Immunofluorescence for characterization
of iPS-NSC
Cultured cells were fixed with 4% paraformaldehyde
for 15 min at room temperature. For labeling intracellular
antigens, cells were permeabilized in PBS containing 0.5% Triton
X-100. After washing in PBS, cells were blocked with 5% FBS in PBS
for 1 h and incubated overnight in 3% FBS in PBS with following
primary antibodies: Oct 4 (1:100; Santa Cruz Biotechnology), Sox2
(1:100; Chemicon), Pax6 (1:100; MBL Eppendorf) and Nestin (1:200;
Abcam). After two washes in PBS, appropriate secondary antibodies
conjugated to Alexa Fluorophores 488 or 568 (Invitrogen) were
diluted at 1:500 in blocking solution and mixed with DAPI (1 μg/ml;
Invitrogen) to counterstain the nuclei. The cells were then washed
twice in PBS. Images were acquired with an EVOS microscope.
In vivo migration assay using
5-bromo-2-deoxyuridine (BrdU)-labeled iPSCs and iPS-NSCs
The following experiments were performed according
to the Rules of Animal Experimentation and the Guide for the Care
and Use of Laboratory Animals of Hamamatsu University School of
Medicine and approved by the Institutional Animal Care and Use
Committee of Hamamatsu University School of Medicine
(H23-012-2011031).
For the in vivo transplantation experiments
(22), iPSCs or iPS-NSCs cultured
for 5 days were pulsed for 48 h with 5 μM BrdU (Sigma) in each
appropriate medium before harvest. iPSCs were harvested by
incubation with 0.25% trypsin for 3 min and iPS-NSCs, with Accutase
for 5 min at room temperature followed by gentle scraping and
resuspended in PBS for implantation.
Six male C57BL/6 mice (19–21 g, 6-weeks old; Nippon
SLC, Hamamatsu, Japan) were anesthetized with 0.4 ml/100 g
equithesin and placed in a stereotaxic apparatus (Narishige
Scientific Instrument Laboratory, Tokyo, Japan). The bregma and
lambda were placed in the same horizontal plane. The scalp was
incised at the midline and retracted laterally to make a burr hole
at an appropriate location (coordinates with respect to bregma: 1
mm posterior, 2 mm right). A 25-gauge needle was inserted to the
point of 4 mm ventral from the dura, left in position for 1 min,
and then withdrawn to the point of 3 mm ventral from the dura,
where 1×104 GL261 sphere cells per 5 μl PBS were infused
with a 50 μl microsyringe (Hamilton Company, Reno, NV, USA) and a
microinjector (Harvard Apparatus Inc., South Natick, MA, USA) for 5
min. The needle was kept in the point of 2 mm ventral from the dura
for an additional minute before removal. After that, a
contralateral burr hole (bregma: 1 mm posterior, 2 mm left) was
made and 1×105 BrdU-labeled iPSCs (n=3) or iPS-NSCs
(n=3) per 5 μl were injected as stated above. The animals were kept
under the same laboratory conditions. All the animals were
euthanized by injection of high dose of pentobarbital on day 7
after the implantation and the removed brains were fixed with 4%
paraformaldehyde for 2 h. After the fixation, the brains were kept
in 20% sucrose 4°C overnight then blocked in OCT embedding compound
and snap-frozen at −78°C using Histo-Tek PINO (Sakura Finetek Japan
Co., Ltd., Tokyo, Japan). Serial coronal cryosections (10 μm) were
obtained and stained with hematoxylin and eosin. Sister sections
were prepared for dual-filter immunofluorescence.
For the immunofluorescence (23), these sections were treated with
citric acid at >97°C for 15 min and allowed to cool at room
temperature for 1 h to denature DNA. After washing in PBS, the
sections were blocked with 1% bovine serum albumin (BSA) in PBS for
15 min and incubated overnight in 1% BSA in PBS with anti-BrdU
antibody (1:100; Sigma) at 4°C. For the conformation of GFP (Nanog)
expression, the sections of mice injected with iPSCs were
additionally stained with anti-GFP antibody conjugated to Alexa
Fluorophores 488 (1:500; Molecular Probes-Invitrogen). After two
washes in PBS, appropriate secondary antibody conjugated to Alexa
Fluorophores 568 (1:500; Molecular Probes-Invitrogen) and DAPI (1
μg/ml; Molecular Probes-Invitrogen) in blocking solution was
applied for 1 h at room temperature. The sections were examined and
photographed under an FV1000 fluorescent microscope.
Statistical analysis
The data (means ± SD) were analyzed using the
two-tailed unpaired Student’s t-test with 95% confidence interval
for a two-group comparison. Differences were considered significant
at P<0.05.
Results
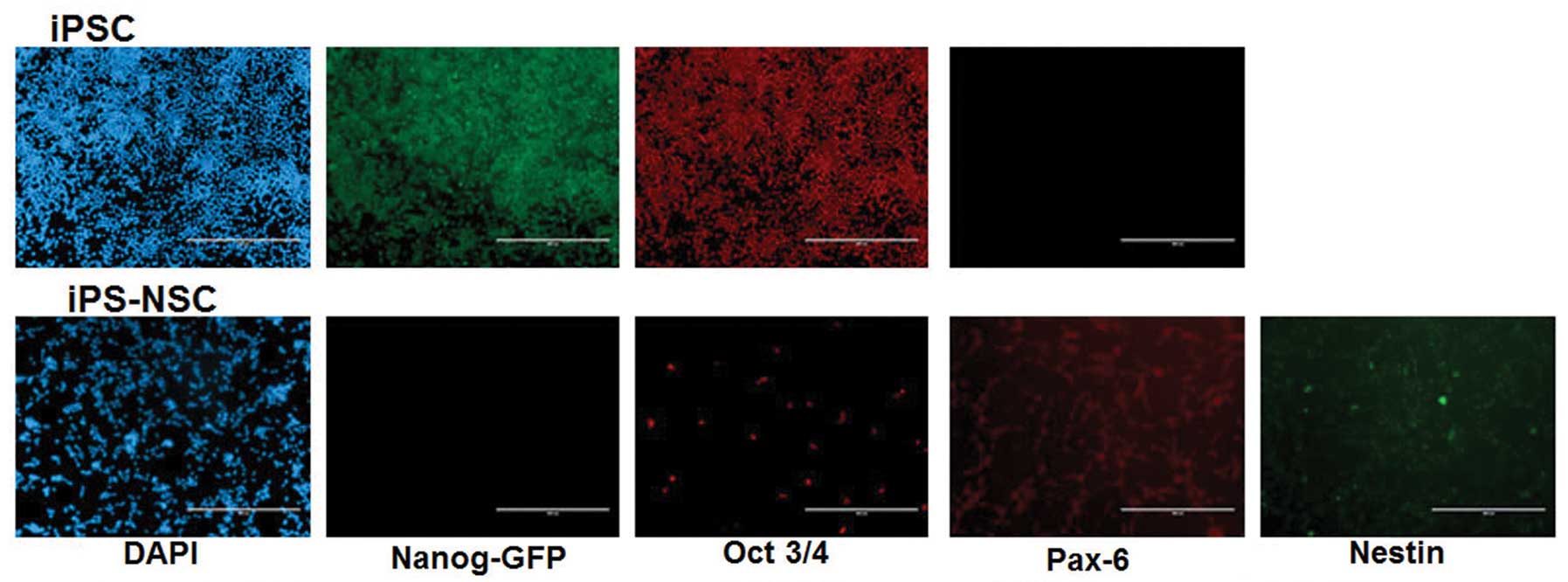
Characterization of iPSC and iPS-NSC
Immunocytochemical characterization of iPS-NSCs was
Pax6 (+), nestin (+) and Oct3/4 (−), Nanog-GFP (−), while that of
iPSCs was positive for Oct3/4 (+), Nanog-GFP (+) and Pax 6 (−)
(Fig. 1).
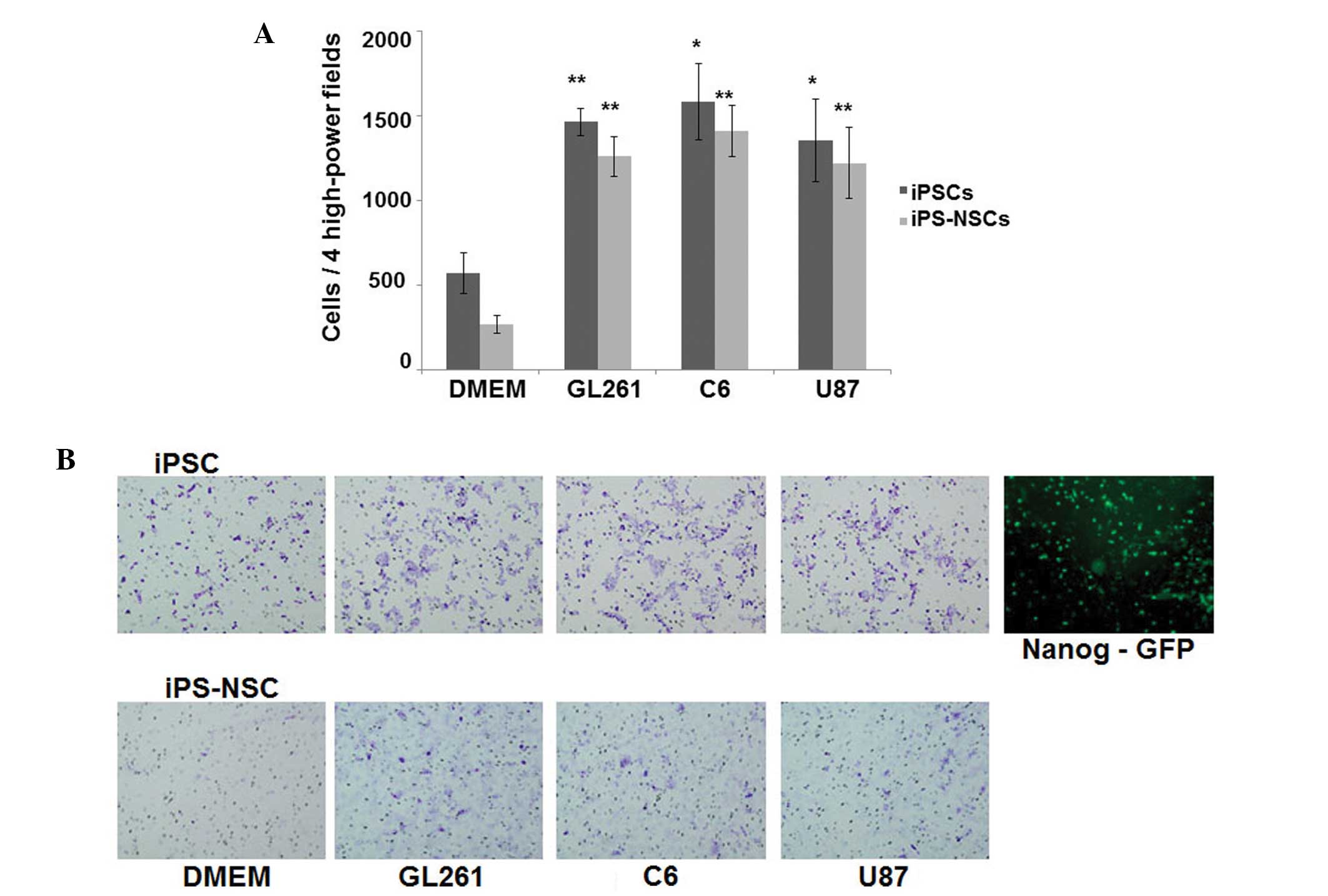
Migration of iPSCs and iPS-NSCs towards
the glioma-CM in vitro
In vitro migratory capacity of iPSCs and
iPS-NSCs toward the CM prepared from 3 glioma cell lines (GL261, C6
and U87) was analyzed using the 24-well Matrigel invasion chamber.
Significantly higher numbers of iPSCs and iPS-NSCs were migrated
toward not only mouse GL261-CM but also rat C6-CM and human U87-CM,
whereas only a few cells toward the unconditioned medium (Fig. 2A). Representative photomicrographs
of the micropore membranes are shown (Fig. 2B, left panels). Most of the
migrated iPSCs were positive for Nanog-GFP, suggesting that the
migrated iPSCs were not differentiated (Fig. 2B, right panels).
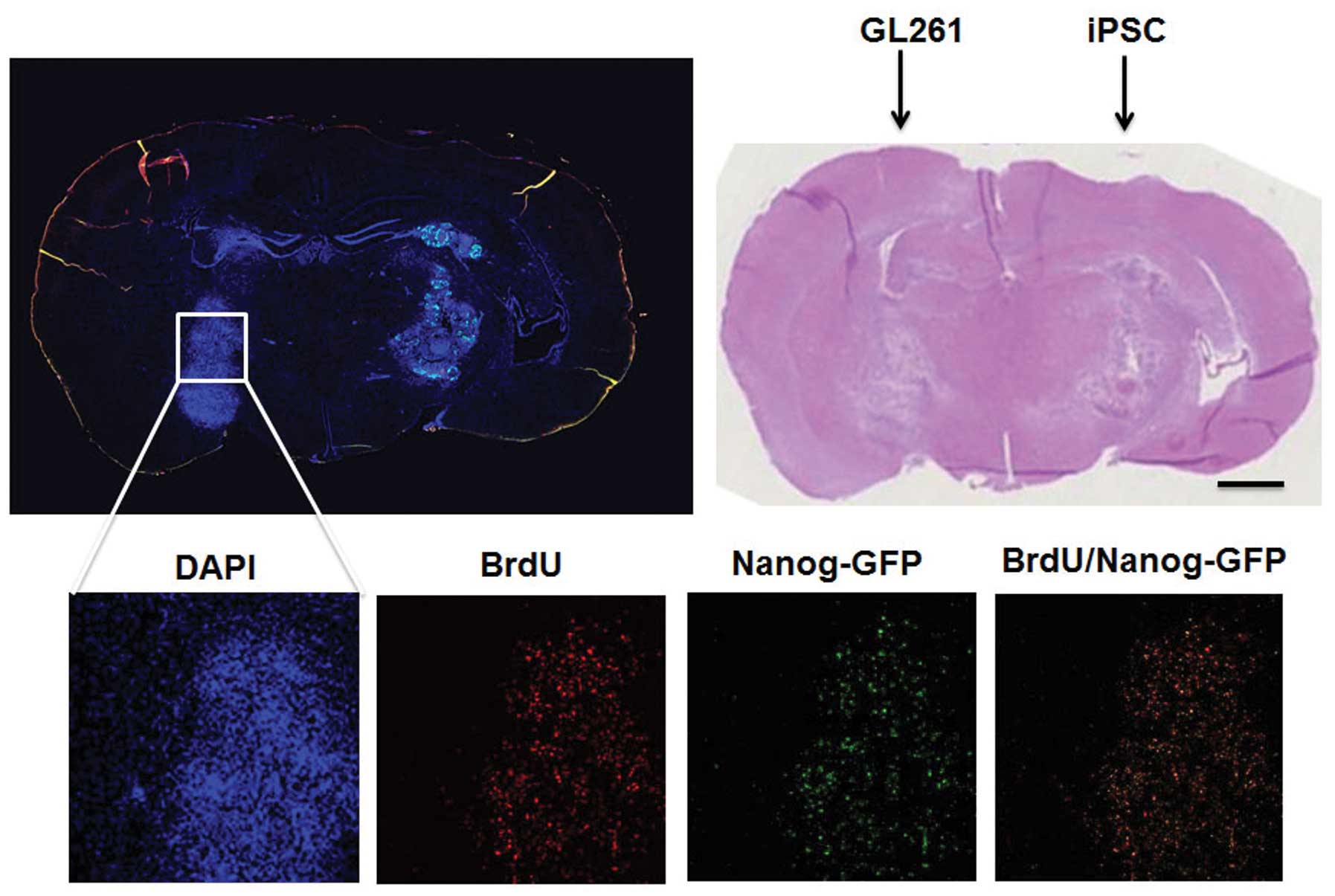
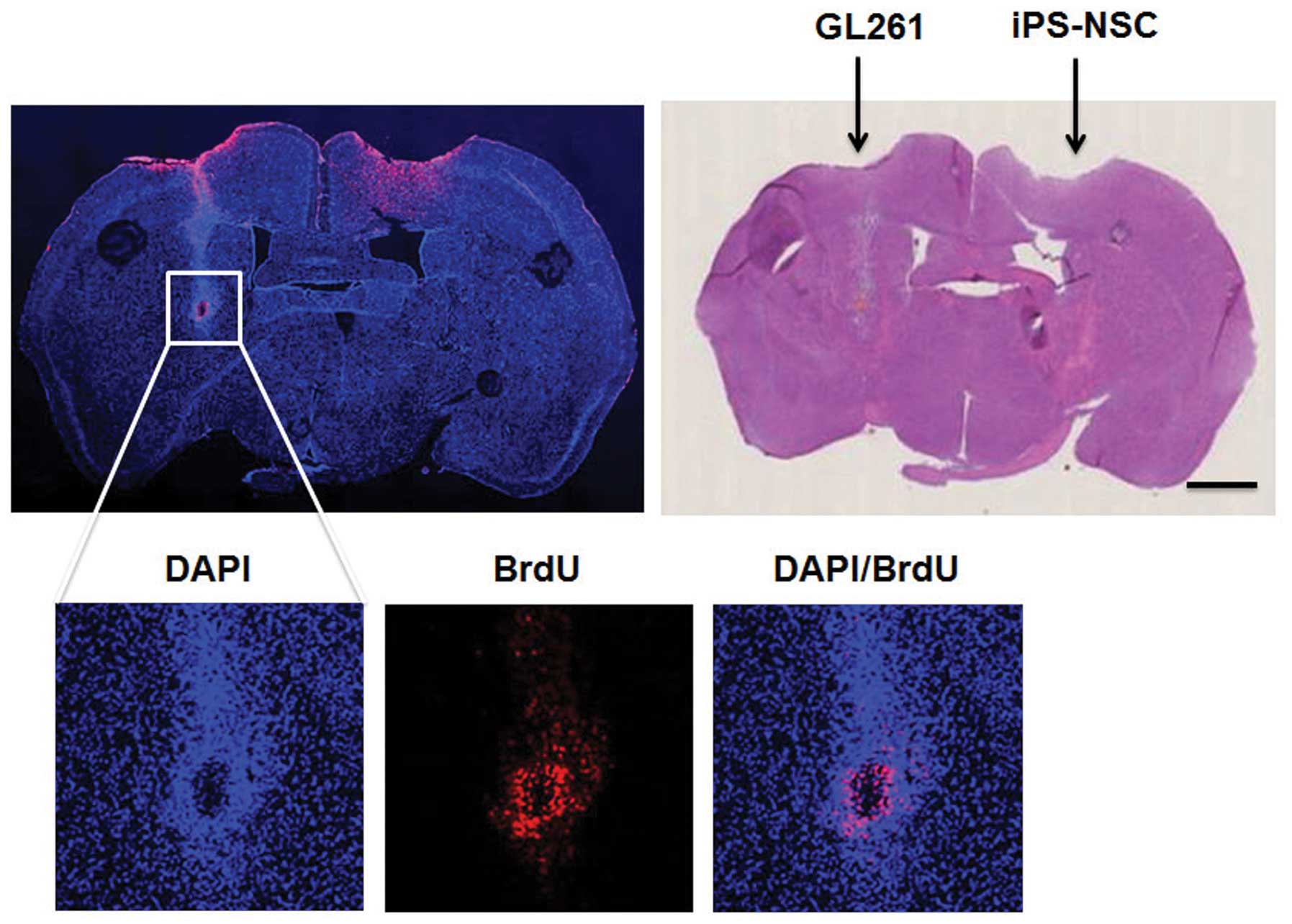
Migration of iPSCs and iPS-NSCs in the
mouse glioma model
BrdU-labeled iPSCs or iPS-NSCs were implanted into
the C57BL/6 mouse brain contralateral to the GL261 sphere cell
implantation (n=3 for each group) to assess in vivo
migratory capability of iPSCs and iPS-NSCs. Substantial amount of
BrdU-labeled iPSCs was observed in the GL261 tumor area, most of
which were positive for Nanog-GFP, suggesting that the migrated
iPSCs were undifferentiated (Fig.
3). When the iPS-NSCs were used, a similar migratory activity
was observed (Fig. 4).
Discussion
NSCs and MSCs are noteworthy alternative therapeutic
tools for malignant glioma because these cells have a strong glioma
tropism and immunosuppressive properties to ensure long time
reservoirs of therapeutic agents at the tumor sites. Since Aboody
and colleagues (24) found an
extensive tropism of NSCs for intracranial gliomas, several
laboratories have investigated these phenomena using various kinds
of stem-like cells. Those cells displayed a unique intrinsic
capacity to ‘home’ the tumor site and to migrate along
metastatic/invasive tumor borders even far from the initial site of
transplantation. In our previous studies, we demonstrated that NSCs
and MSCs have a strong glioma tropism as well as a sufficient
bystander effect for a suicide gene therapy using the herpes
simplex virus-thymidine kinase gene and a prodrug, ganciclovir
(9–11,25).
In 2010, a clinical trial for evaluating the efficacy/safety of the
transplantation of human NSCs genetically modified to express
another suicide gene, cytosine deaminase, which convert
5-fuorocytosine (prodrug) to 5-fluorouracil (active drug) at the
tumor site, was started for malignant glioma patients in
California, USA (13). Though NSCs
can potentially be obtained from the adult brain, this process is
very complicated and time-consuming. As an alternative, most
studies use a stable cell line of immortalized NSCs originally
obtained from embryonic stem cells. Use of embryonic stem cells
obviously implies a serious ethical concern, which can be
circumvented by using iPSCs. In addition, iPSCs can be obtained
from the patients themselves. In fact, suicide gene therapies using
iPSC-derived NSCs as cellular vehicles have been already studied
(18,26).
In our previous study, mouse iPSCs exerted in
vitro tropism to glioma CM as well as various growth factors
(16). The present in vivo
study was designed to examine whether iPSCs had a glioma tropic
activity in the brain as in the case of in vitro study. The
implanted iPSCs may migrate or stay at the site of implantation.
When the implanted iPSCs migrate, the migrated cells may not be
iPSCs, because implanted iPSCs may easily change to more
differentiated forms. Our findings demonstrated that the
intracranially implanted iPSCs migrated to the tumor site in the
contralateral hemisphere. Interestingly, most of the migrated cells
were still iPSCs, which was confirmed by positive staining for GFP
(Nanog), a marker for iPSC.
Because iPSCs are known to form teratomas when
implanted in the brain (27).
Safety issues are thus more important than efficacy of the
treatment in clinical application of iPSCs. We then tested
migratory activity of the safer iPS-NSCs which were transduced from
iPSCs after neural induction. Our in vitro study
demonstrated that iPS-NSCs displayed a potent migratory activity
similar to that of iPSCs and the migrated iPS-NSCs were negative
for GFP (Nanog) (data not shown), suggesting no substantial
contamination of iPSCs in the prepared iPS-NSCs fraction. Our in
vivo study also demonstrated a potent tropism of GFP
(Nanog)-negative iPS-NSCs to the glioma site in the contralateral
hemisphere.
Since the potent migratory activity of iPS-NSCs was
similar to iPSCs, use of genetically engineered iPS-NSCs might be a
safer strategy for treating the tumor recurrence as a glioma gene
therapy. Namely, iPSCs are obtained from the patients themselves,
transduced with therapeutic genes and expanded. Those genetically
engineered iPSCs are intratumorally implanted after neural
differentiation (iPS-NSCs) at the time of tumor recurrence.
Alternatively, established allogeneic iPS-NSC lines transduced with
therapeutic genes may also be useful, because rejection of the
treatment cells after a certain period may not disturb treatment
efficacy in the case of cancer gene therapies. In this setting, the
treatment strategy can be also applied at the time of the initial
surgery. One of the limitations of the present in vivo study
is lack of quantitative evaluation of migrated cells. Now, we are
developing an in vivo method for semi-quantitative
evaluation of ‘tumor homing therapeutic cells’ using radioactive
tracers (28).
In conclusion, the present study demonstrated that
both iPSCs and iPS-NSCs present a similarly potent tumor tropism
under both in vivo and in vitro conditions. In
addition, the iPSCs did not differentiate during migration, since
they were positive for GFP (Nanog) in the tumor site. To the best
of our knowledge, the present study is the first report to explore
the biological behavior of iPSCs after being transplanted. These
findings suggest that iPSCs and their derivatives can be feasible
candidates as vehicles in stem cell-based gene therapy for the
patients with malignant glioma.
Acknowledgements
The present study was supported by the grants from
the Japan Society for the Promotion of Science (grant-in-aid for
Scientific Research [B; General], #25293306 and [C; General],
#23592119).
References
|
1
|
Grossman SA, Ye X, Piantadosi S, et al:
Survival of patients with newly diagnosed glioblastoma treated with
radiation and temozolomide in research studies in the United
States. Clin Cancer Res. 16:2443–2449. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Johnson DR, Ma DJ, Buckner JC and Hammack
JE: Conditional probability of long-term survival in glioblastoma:
a population-based analysis. Cancer. 118:5608–5613. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lacroix M, Abi-Said D, Fourney DR, et al:
A multivariate analysis of 416 patients with glioblastoma
multiforme: prognosis, extent of resection, and survival. J
Neurosurg. 95:190–198. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sanai N, Polley M, McDermott M, Parsa A
and Berger M: An extent of resection threshold for newly diagnosed
glioblastomas. J Neurosurg. 115:3–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Westphal M, Hilt D, Bortey E, et al: A
phase 3 trial of local chemotherapy with biodegradable carmustine
(BCNU) wafers (Gliadel wafers) in patients with primary malignant
glioma. Neuro Oncol. 5:79–88. 2003.PubMed/NCBI
|
|
6
|
Fleming A and Saltzman W: Pharmacokinetics
of the carmustine implant. Clin Pharmacokinet. 41:403–419. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aboody K, Brown A, Rainov NG, et al:
Neural stem cells display extensive tropism for pathology in adult
brain: evidence from intracranial gliomas. Proc Natl Acad Sci USA.
97:12846–12851. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee J, Elkahloun AG, Messina SA, et al:
Cellular and genetic characterization of human adult bone
marrow-derived neural stem-like cells: a potential antiglioma
cellular vector. Cancer Res. 63:8877–8889. 2003.PubMed/NCBI
|
|
9
|
Amano S, Li S, Gu C, et al: Use of
genetically engineered bone marrow-derived mesenchymal stem cells
for glioma gene therapy. Int J Oncol. 35:1265–1270. 2009.PubMed/NCBI
|
|
10
|
Li S, Tokuyama T, Yamamoto J, Koide M,
Yokota N and Namba H: Bystander effect-mediated gene therapy of
gliomas using genetically engineered neural stem cells. Cancer Gene
Ther. 12:600–607. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li S, Gao Y, Tokuyama T, et al:
Genetically engineered neural stem cells migrate and suppress
glioma cell growth at distant intracranial sites. Cancer Lett.
251:220–227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amano S, Gu C, Koizumi S, Tokuyama T and
Namba H: Tomoricidal bystander effect in the suicide gene therapy
using mesenchymal stem cells does not injure normal brain tissues.
Cancer Lett. 306:99–105. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bovenberg MS, Degeling MH and Tannous BA:
Advances in stem cell therapy against gliomas. Trends Mol Med.
19:281–291. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nishikawa S, Goldstein RA and Nierras CR:
The promise of human induced pluripotent stem cells for research
and therapy. Nat Rev Mol Cell Biol. 9:725–729. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Okita K, Ichisaka T and Yamanaka S:
Generation of germline-competent induced pluripotent stem cells.
Nature. 448:313–317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koizumi S, Gu C, Amano S, et al: Migration
of mouse-induced pluripotent stem cells to glioma-conditioned
medium is mediated by tumor-associated specific growth factors.
Oncol Lett. 2:283–288. 2011.PubMed/NCBI
|
|
17
|
Takahashi K, Okita K, Nakagawa M and
Yamanaka S: Induction of pluripotent stem cells from fibroblast
cultures. Nat Protoc. 2:3081–3089. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee E, Lam D, Wu C, et al: Glioma gene
therapy using induced pluripotent stem cell derived neural stem
cells. Mol Pharm. 8:1515–1524. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Onorati M, Camnasio S, Binetti M, Jung CB,
Moretti A and Cattaneo E: Neuropotent self-renewing neural stem
(NS) cells derived from mouse induced pluripotent stem (iPS) cells.
Mol Cell Neurosci. 43:287–295. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pellegatta S, Poliani PL, Corno D, et al:
Neurospheres enriched in cancer stem-like cells are highly
effective in eliciting a dendritic cell-mediated immune response
against malignant gliomas. Cancer Res. 66:10247–10252. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu SC, Ping YF, Yi L, et al: Isolation and
characterization of cancer stem cells from a human glioblastoma
cell line U87. Cancer Lett. 265:124–134. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao Y, Gu C, Li S, et al: p27 modulates
tropism of mesenchymal stem cells toward brain tumors. Exp Ther
Med. 1:695–699. 2010.PubMed/NCBI
|
|
23
|
Tang X, Falls DL, Li X, Lane T and Luskin
MB: Antigen-retrieval procedure for bromodeoxyuridine
immunolabeling with concurrent labeling of nuclear DNA and antigens
damaged by HCl pretreatment. J Neurosci. 27:5837–5844. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Aboody K, Najbauer J and Danks M: Stem and
progenitor cell-mediated tumor selective gene therapy. Gene Ther.
15:739–752. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li S, Tokuyama T, Yamamoto J, Koide M,
Yokota N and Namba H: Potent bystander effect in suicide gene
therapy using neural stem cells transduced with herpes simplex
virus thymidine kinase gene. Oncology. 69:503–508. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang J, Lam DH, Goh SS, et al: Tumor
tropism of intravenously injected human-induced pluripotent stem
cell-derived neural stem cells and their gene therapy application
in a metastatic breast cancer model. Stem Cells. 30:1021–1029.
2012. View Article : Google Scholar
|
|
27
|
Kawai H, Yamashita T, Ohta Y, et al:
Tridermal tumorigenesis of induced pluripotent stem cells
transplanted in ischemic brain. J Cereb Blood Flow Metab.
30:1487–1493. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pei Z, Lan X, Cheng Z, et al:
Multimodality molecular imaging to monitor transplanted stem cells
for the treatment of ischemic heart disease. PLoS One.
9:e905432014. View Article : Google Scholar : PubMed/NCBI
|