1. Introduction
Data from genome-wide transcriptional analysis in
humans have shown that the amount of protein coding transcripts
accounts for approximately 2% of the entire genome, while the
non-coding RNAs (ncRNAs) correspond to around 98% of all the
genomic output (1,2). Interestingly, it has been reported
that the proportion of non-coding regions in the genome increases
according to the complexity of the organism, suggesting a important
role for these sequences in physiology and development of the
organisms (3,4). For this reason, much attention has
been given to the studies on these non-protein-coding RNAs in many
fields, especially in cancer, leading to new hypothesis about
cancer biology (5). Additionally,
the identification of the circulating microRNAs (miRNAs) in bodily
fluids makes them potential non-invasive biomarkers for cancer
diagnosis and prognosis.
The comprehension of the mechanisms involved in the
interactions between tumor cells and the surrounding environment is
relevant for tumor biology elucidation and for the improvement of
innovative and more efficient therapy approaches (6). The role of extracellular vesicles in
cell-to-cell communication in cancer has been the focus of several
studies. MiRNAs are one of the most studied exosomal cargos due to
their potential role in tumor diagnosis, prognosis and therapy.
In this review, we summarize the role of ncRNAs in
cancer, focusing on miRNAs. Additionally, we focus on the role of
exosomes in intercellular communication and their potential use in
providing diagnostic opportunities, unraveling new therapeutic
targets and predicting therapeutic responses.
2. World of non-coding RNAs
The ncRNAs can be divided in two main groups,
according to their sizes: long non-coding RNAs (lncRNAs), which are
greater than 200 nucleotides and small non-coding RNAs with no more
than 200 nucleotides (7,8). These two main categories also show
some subgroups, based on the structure and biological function of
the transcripts, as long intergenic ncRNAs, pseudogenes, enhancer
RNAs, transcribed ultra-conserved region, repeated-associated
ncRNAs and antisense RNA in the lncRNAs group. In the small ncRNAs,
miRNAs, tiny transcription initiation RNAs, small interfering RNAs,
promoter-associated short RNAs, antisense termini associates short
RNAs and retrotransposon-derived RNAs have been reported in the
literature (5,8).
The miRNAs are the most widely described ncRNA in
the literature, since the first small ncRNA lin-4 was described in
C. elegans more than 20 years ago (9,10).
The synthesis of these evolutionarily conserved endogenous short
single-stranded RNAs (18–20 nucleotides in length) begins in the
nucleus, when the transcription of miRNA-coding genes generate long
primary transcript (pri-miRNA) with stem-loop, which will be
detached by the RNase III Drosha/Pasha/DGCR8 complex, and then
producing a 70-nucleotide precursor (pre-miRNA). After being
transported to the cytoplasm by the protein Exportin-5 (XPO5), the
pre-miRNA will be converted in mature miRNA by the action of the
Dicer and binding to Argonaute 2 (Ago2) to form the RNA-induced
silencing complex (RISC) (11,12).
Overall, miRNAs regulate gene expression post-transcriptionally,
most commonly through the binding to a specific sequence at the
3′-untranslated region (3′-UTR) of a target protein-coding mRNA,
causing a translational repression or cleavage of the target
transcripts (13). Thus, miRNAs
have a relevant role in many pathological and physiological
processes, such as cell proliferation, differentiation, development
and apoptosis, acting as oncogenes or tumor suppressors, depending
on which genes they regulate (14).
The involvement of miRNA genes in cancer was first
described in 2002, when the authors reported that two miRNAs
(miR-15a and miR-16-1) are mapped at 13q14, a chromosomal region
frequently deleted in B-cell chronic lymphocytic leukemia (B-CLL)
and that both genes are down-regulated in a high proportion of the
cases (15). Since then, the
number of studies on miRNAs and cancer has been increasing
considerably, adding novel insights into the role of the miRNAs in
human tumor such as in hematological malignancies (16–19),
colorectal (20–23), breast (24–28),
head and neck (29–32) and gastric cancer (33–36).
The lncRNAs are transcripts longer than 200
nucleotides, a cutoff based on RNA purification protocols that
exclude small RNAs rather than for its functional role (37). The lncRNAs were first described in
a study involving large-scale sequencing and annotation of
full-length cDNA libraries in mouse (38), and the number of reports about
characterization and functions of the lncRNAs has been constantly
increasing in the literature (39). The lncRNAs have many features in
common with mRNAs, as transcription by RNA polymerase II,
polyadenilation and splicing mechanism. This category of ncRNAs
composes a heterogeneous group, which makes the lncRNAs
classification difficult (40).
Most commonly, the lncRNAs can be classified as sense or antisense,
divergent or convergent and intronic or intergenic, depending on
their position relative to the neighboring protein-coding genes
(7,41). Due to lncRNA structure
heterogeneity, it is also difficult to assign a specific function
to this group and still requires further studies. Evidences suggest
that lncRNAs act mainly in regulation of protein-coding genes
transcription, but in more complex ways than the miRNAs (42). lncRNAs can repress the
transcription of target genes involving epigenetic modifications
like chromatin remodeling, since some lncRNAs have been reported to
interact with many chromatin modifiers (43). Additionally, lncRNAs can either
play a role as putative gene enhancers or decoy RNAs in
transcriptional control (41).
Some lncRNAs (antisense ncRNA) also play a role in
post-transcriptional regulation by interfering with the RNA
splicing mechanism (44).
Due to their roles in the functions mentioned above,
the lncRNAs have been related to many human cancers, contributing
to tumor development and progress (40). Many lncRNAs have been mapped at
cancer risk loci in the human genome, such as PTCSC3 (14q13.3) in
thyroid cancer (45,46), PCA3 (9q21–22) in prostate cancer
(47,48), ANRIL (9p21) in prostate and breast
cancers, leukemia and melanoma (49–52),
MALAT1 (11q13) in liver, colorectal, prostate, bladder and lung
cancers (53–56).
The role of ncRNAs in many human tumor types has
been exhaustingly studied in the past few years and its relevance
in mechanisms involved in cancer development and progress is
unquestionable. Additionally, the discovery of stable miRNAs in
bodily fluids introduced new insights in the ncRNAs comprehension
and can represent a new diagnostic approach using less invasive
methods (57). The use of
circulating miRNAs as tumor biomarkers has many advantages since
these transcripts are conserved across species, shows tissue or
disease-specific expression and their levels can be quantified by
various methods, as microarray profiling, northern blot analysis,
in situ hybridization, high-throughput sequencing and
qRT-PCR, which is the most used method due to its sensitivity,
specificity and reliability (58–61).
The first evidence of the presence of miRNAs in serum was reported
by Lawrie et al (2008), who showed the higher serum levels
of miR-21 in large B-cell lymphoma (62). Since then, many studies have
reported the presence of different circulating miRNAs in various
tumor types, such as in colorectum (63,64),
esophagus (65,66), breast (67,68),
stomach (69,70) and ovary (71,72).
Also, the circulating miRNAs have been reported in many other
fluids such as plasma, urine, saliva and cerebrospinal fluid
(58,73). Considering this discovery, it is
possible that other types of ncRNAs similar to lncRNAs can also be
identified in bodily fluids (40).
3. World of extracellular vesicles
The intercellular communication can occur by a
direct cell-to-cell contact including the adhesion junctions, or by
the releasing of soluble signaling molecules or by the exchange of
cellular fragments as extracellular vesicle (EV) (74,75).
The EVs are bilayered membrane vesicles secreted by all cell types,
and released in the interstitial space or into circulating bodily
fluids, where they can travel long distances until they are up
taken by receptor cells (76).
Different terminology is used to describe EVs based on their
morphology and diameter. Exosomes, microvesicles, ectosomes,
microparticles and others, are classified based on their size,
shape and membrane surface composition (77). The most accepted classification in
the literature shows two major groups of EVs, based on their
mechanism of biogenesis and sizes: exosomes and microvesicles (or
ectosomes). Additionally, apoptotic bodies have been considered by
some as a third category of EVs (78–80).
In this review, we will focus on the exosomes and
microvesicles.
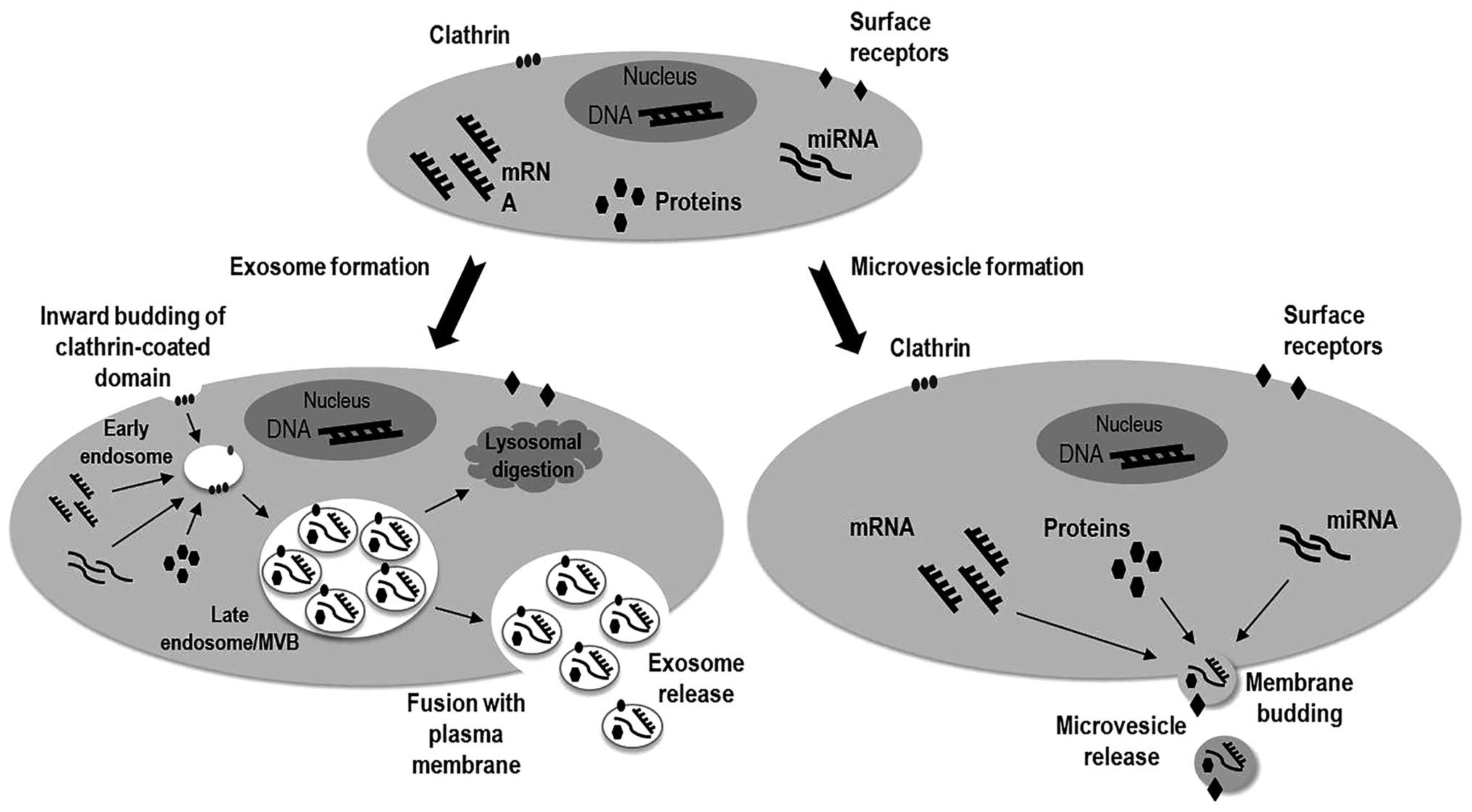
Exosomes are 40–140 nm diameter bilayered-membrane
vesicles of endocytic origin, with a cup-shaped morphology, showing
densities ranging between 1.13–1.19 g/ml (81). The exosomes are originated by the
inward budding of clathrin-coated domains in the plasma membrane,
generating the multivesicular bodies (MVBs) containing intraluminal
vesicles (ILVs) in the late endosome. The formation of ILVs occurs
during the endosome maturation, when specific cytosolic proteins
are incorporated into these vesicles inside de MVBs. These initial
steps occur under control of the ESCRT (endosomal sorting complex
required for transport) machinery. Later, the MVBs fuse with
lysosomes for degradation or with the cell membrane releasing the
exosomes to the extracellular space, process regulated by the RAB
family (76,82–84).
Microvesicles (or ectosomes) are larger than exosomes, with size
ranging between 100 and 1,000 nm in diameter and heterogeneous in
morphology. Differently from the exosomes, microvesicles (MVs) are
originated from the plasma membrane through direct outward budding
into the extracellular space. During this process, the newly
originated vesicle captures the donor cellular cytosolic content
and the receptors on the plasma membrane (Fig. 1). The regulation of MVs biogenesis
is intracellular calcium-dependent and it is the result of the
activation of cell surface receptors, phospholipid redistribution
and cytoskeletal protein contraction (84,85).
The apoptotic bodies (ABs) are membrane vesicles, heterogeneous in
shape, showing sizes ranging between 50–500 nm in diameter. The ABs
are released from the outward protrusion of the plasma membrane
during the late phase of cell death by apoptosis and are featured
by the presence of organelles inside the vesicles (85,86).
The EV cargo specificity
The interaction between the EVs and the target cells
can occur by different mechanisms, as direct interaction of the
surface proteins of the EVs with the receptors on the target cells,
triggering the activation of the intracellular pathways. EVs can
also be engulfed by the target cells through membrane fusion or by
endocytosis/phagocytosis, with transfer and release of their cargo.
Transcripts as mRNAs and miRNAs contained inside de EVs can be
transferred to the target cells and be functional (6).
The EVs carry specific contents (cargo) as membrane
receptors, ligands, proteins, nucleic acids and infectious agents,
depending on the cell of origin and how they were originated from
the donor cell (75). There is no
consensus regarding the specific content of different EVs, but it
seems that MVs are characterized by the presence of cell-surface
proteins from the donor cells such as receptors and adhesion
proteins. In turn, exosomes have been found to be characterized by
proteins associated to their endosomal origin and MVBs formation
(84). Some specific markers have
been associated to exosomes as tetraspanins (CD9, CD63, CD81 and
CD82), major histocompatibility complex class I and II, LAMP1 and
LAMP2, flotilins, annexins, TSG101 and heat shock proteins
(83,87,88).
The protein content of MVs seems to be more heterogeneous,
containing cell membrane markers, phosphatidylserine (PS) residues,
integrins, selectins and CD40 ligands (84), and high levels of cholesterol and
signaling complexes known as lipid rafts (76). Most importantly, it has been shown
that the population of exosomes secreted by cancer cells contains a
representation of the entire genome of the cell of origin,
providing exciting opportunities of using exosomes as a liquid
biopsy (89).
Tumor-derived exosomes
The exosomes are by far the most extensively studied
due to their characteristics (as presence in bodily fluids and
expression of specific markers) that can contribute not only to
intercellular communication but also to their potential role in
diagnosis (82). The release of
exosomes can be a response to different cellular stress conditions
common in cancer, such as hypoxia, acidic pH, heat shock and
oxidative stress, resulting in alterations of the tumor
microenvironment and distal organs activating angiogenesis and
promoting migration and leading to metastasis (90–92).
An important step before considering using the
exosomal content for study or diagnosis purposes is a reliable
exosome isolation method, to insure the quality of the results.
Exosomes can be collected from fluids or cell supernatant by a
series of sequential centrifugations to remove larger cellular
debris and filtration through 100–220 nm filters to exclude larger
EVs, including MVs and apoptotic bodies. Then, the exosomes are
pelleted by ultracentrifugation and/or suspension in a sucrose
gradient for the completely remove of protein contamination
(77,93,94).
The exosomes isolation can also be performed using specific
filters, immune isolation by magnetic beads or microfluidic
separation (95). Recently, some
commercial isolation kits are available based on polymer-based
precipitation and on the magnetic bead isolation. Then, some
additional procedures are necessary to confirm the purity of the
isolated exosomes. One of them is to verify the size and shape of
the exosomes by electron microscopy analysis. Vesicles diameter and
morphology can also be assessed by specific instruments that can
visualize, characterize and measure small vesicles. Another
important factor that must be evaluated is the protein content,
that can be assessed by flow cytometry and western blot analysis
for markers as CD63, CD81, Tsg101 and flotilin (77,96).
4. The fusion of the two worlds
Exosomal miRNAs
In the bodily fluids, the miRNAs have been reported
to play a role at intercellular communication, and can act at short
and long distant sites in a hormone-like behavior (14,87).
The transport of circulating miRNAs can be carried by protein
transporters or by exosomes. It is known that serum contains
ribonucleases, suggesting that the circulating miRNAs are protected
from the RNase action within extravesicles. The miRNA recruitment
to the exosomes depends on the attachment of RNA-induced silencing
complexes (RISCs) to the ESCRT components. However, the release of
exosomal miRNAs is under control of a ceramide-dependent machinery,
as reported by Kosaka et al (73). These authors showed that the
inhibition of neutral sphingomyelinase 2 (a regulator of the
ceramide biosynthesis) resulted in lower levels of miRNA secretion
(73).
The first evidence of the existence of miRNAs in
exosomes was reported by Valadi et al, showing that these
vesicles contain both mRNA and miRNAs, which can be transferable to
another cell, where the transcripts can be functional (97). Since then, the number of studies
regarding the identification of exosomal miRNAs in cancer has been
increasing in the literature. A summary of some studies in the
literature in this field is in Table
I. Most of these reports are based on in vitro studies
involving a variety of cancer cell lines, identifying the exosomal
miRNA content as in breast cancer (98,99),
leukemia (100), melanoma
(101,102), prostate cancer (103), ovarian (104) and gastric cancer (105). The transcripts content of the
exosomes usually can differ from that in the donor cells, and the
exosomal miRNA profile of tumor cells can also differ from that
released by normal controls (106).
 | Table ISummary of the studies reporting the
identification of exosomal miRNAs in cancer. |
Table I
Summary of the studies reporting the
identification of exosomal miRNAs in cancer.
| Tumor | Sample | Exosome
extraction | miRNA | Refs. |
|---|
| Breast cancer | Cell line | SC, 0.22 μm
filtering and UC | miR-233 | (108) |
| Breast cancer | Cell line | SC and UC or
ExoQuick (System Biosciences) | miR-210 | (116) |
| Breast cancer | Cell line | SC, 0.22 μm
filtering and UC | miR-100, miR-17,
miR-222, miR-342-3p, miR-451, miR-30a | (114) |
| Breast cancer | Cell line | SC, 0.22 μm
filtering, UC and sucrose gradient | miR-198, miR-26a,
miR-34a, miR-49a, let-7a, miR-328, miR-130a, miR-149, miR-602 and
miR-92b | (99) |
| Breast cancer | Serum samples,
tumor samples, cell line, animal models | SC, UC | miR-105 | (113) |
| Cervical
cancer | Cervicovaginal
lavage fluid | SC and UC | miR-21 and
miR-146a | (126) |
| Cholangiocarcinoma
(biliary tree) | Bile sample | SC and 0.22 μm
filtering | miR-222, miR-126,
miR-486-3p, miR-484, miR-19a, miR-19b, miR-16, miR-191, miR-31,
miR-1274b, miR-618, miR-486-3p, miR-16, miR-1274b, miR-484,
miR-191 | (125) |
| Colorectal
cancer | Cell line | SC and 0.22 μm
filtering | miR-21, miR-192,
miR-221 | (107) |
| Colorectal
cancer | Serum samples | SC, 0.22 μm
filtering and UC | let-7a, miR-1229,
miR-1246, miR-150, miR-21, miR-223 and miR-23 | (121) |
| Esophageal
cancer | Serum samples | SC, 0.45 μm
filtering and ExoQuick (System Biosciences) | miR-21 | (120) |
| Gastric cancer | Cell line | SC, 0.1 μm
filtering and UC | let-7 family (a, b,
c, d, e, f, g, i) | (105) |
| Glioblastoma | Tumor samples,
serum samples | SC, 0.22 μm
filtering and UC | let-7a, miR-15b,
mR-16, miR-19b, miR-21, miR-26a, miR-27a, miR-92, miR-93, miR-320,
miR-20 | (118) |
| Glioblastoma | Tumor samples, cell
lines, animal models | SC, 0.22 μm
filtering, and UC | miR-1 | (140) |
| Leukemia | Cell line | ExoQuick (System
Biosciences) | miR-19a,
miR-146-5p, miR-454, miR-18b, miR-574-3p, miR-21, miR-431, miR-345,
miR-210, miR-197, miR-20a, miR-24, miR-19b, miR-130b, miR-106b,
miR-224, miR-210, miR-652, miR-379, miR-185 | (117) |
| Leukemia | Cell line | SC, 0.22 μm
filtering and ExoQuick (System Biosciences) | miR-17–92 cluster,
miR-24, miR-222 | (109) |
| Leukemia | Cell line | SC, 0.22 μm
filtering and UC | miR-1908 and
miR-298 | (100) |
| Lung
adenocarcinoma | Plasma samples | Size exclusion by
chromatography, magnetic beads (EpCAM) | miR-17-3p, miR-21,
miR-106a, miR-146, miR-155, miR-191, miR-192, miR-203, miR-205,
miR-210, miR-212, miR-214 | (122) |
| Lung
adenocarcinoma | Plasma samples | ExoQuick (System
Biosciences) | miR-378a, miR-379,
miR-139-5p, miR-200-5p, miR-151-5p, miR-30a-3p, miR-200b-5p,
miR-629, miR-100 and miR-154-3p | (123) |
| Lung
adenocarcinoma | Cell line | SC, 0.22 μm
filtering, and UC | miR-192 | (112) |
| Lung cancer | Cell line | UC and ExoQuick
(System Biosciences) | miR-21, miR-98,
miR-133b, miR-138, miR-181a, miR-200c | (115) |
| Lung cancer | Plasma sample,
bronchoalveolar lavage fluid | SC, 0.22 μm
filtering, and UC | miR-222, miR-126,
miR-144, miR-302a, miR-302c | (124) |
| Melanoma | Cell line | SC, 0.22 μm
filtering and UC; SC and ExoQuick (System Biosciences) | miR-181b, miR-181a,
miR-4802-3p, miR-23b, miR22, miR-107, miR-103a, miR-9,
miR-338-3p | (101) |
| Melanona and colon
carcinoma | Cell line | SC, 0.22 μm
filtering, prominin-1 based immuno-magnetic selection | miR-216b, miR-889,
miR-4307, miR-4272, miR-203, miR-4289, miR-3149, miR-203, miR-3145,
miR-1911, miR-513a-3p, miR-3916, miR-886-3p, miR-1182, miR-3613-5p,
let-7i, miR-3132, miR-3914, miR-3618, miR-1307, miR-3614-3p,
miR-519c-3p, miR-3160, miR-3153, miR-4278, miR-3646, miR-3926,
miR-515-5p, miR-3169, miR-590-3p, miR-525-5p, miR-548g, miR-365,
miR-525-3p, miR-320d | (102) |
| Multiple
myeloma | Cell line | 0.22 μm filtering,
UC and ExoQuick (System Biosciences) | miR-125q-3p,
miR-128, miR-15a, miR-185, miR-192, miR-212, miR-324-3p,
miR-331-5p, miR-345, miR-422a, miR-429, miR-511, miR-576-3p,
miR-618, miR-9, miR-1271, miR-139, miR-148, miR-151-3p, miR-15b,
miR-19b-1, miR-21, miR-34b, miR-378, miR-589, miR-592, miR-625,
miR-93 | (111) |
| Ovarian cancer | Serum samples, cell
line | Magnetic activated
cell sorting (MACS) - EpCAM | miR-21, miR-141,
miR-200a, miR-200c, miR-200b, miR-203, miR-205, miR-214 | (119) |
| Ovarian cancer | Cell line | SC, UC and sucrose
gradient | let-7 | (104) |
| Prostate
cancer | Cell line | SC, 0.22 μm
filtering, and UC | miR-143 | (110) |
| Protaste
cancer | Cell line | SC and UC | miR-1280, miR-720
and miR-1260b | (103) |
Studies have reported evidence of the intercellular
transfer of the exosomal content between different cells. Chiba
et al showed that exosomes derived from colorectal cancer
cells can be transferred to hepatoma and lung cancer cells
(107). In addition, some of
these reports have demonstrated that the transferred exosomal
content can be functional in the receptor cells. Yang et al
reported the presence of a specific miRNA for IL-4-activated
macrophage - miR-233, in the co-cultured breast cancer cells and it
can enhance the invasiveness potential of the receptor cells
(108). The transfer of the
leukemia cell line-derived exosomal miR-92a to the endothelial
cells affected the endothelial cell migration and tube formation in
the receptor cells (109). Kosaka
et al showed that exosomal miR-143 derived from a normal
prostate cell line act as a tumor suppressor by inhibiting the
growth in the target prostate cancer cells (110). Similar results were reported by
Roccaro et al in a study demonstrating that the exosomal
miR-15 from the normal bone marrow mesenchymal stromal cells causes
a tumor suppressor effect when transferred to tumor cells, where
this miRNA is downregulated (111). In a recent study, Valencia et
al demonstrated that the exosomal miR-192 derived from lung
adenocarcinoma cell lines repressed the angiogenic activity in the
co-cultured endothelial cells by the inhibition of the
proangiogenic factors (112).
More recently, Zhou et al showed that the transfer of
exosomal miR-105 to non-metastatic breast cancer cells induces
metastasis and vascular permeability by targeting the cellular
tight junction protein ZO-1 (113).
The intercellular transfer of the exosome cargo can
also affect the resistance or sensitivity of cancer cells to
therapy. The transfer of the exosomes derived from chemoresistant
breast cancer cell lines can also spread the resistance potential
to receptor chemosensitive cell lines, possibly due to the action
of the exosomal content as miR-100, miR-222 and miR-30a (114). Similarly, in lung cancer, Xiao
et al reported that after miR-21- and miR-133-enriched
exosome transfer from the chemoresistant tumor cells, the
chemosensitive target cells acquire resistance to the drug exposure
(115).
It is well known that hypoxia is an important factor
that triggers angiogenesis and metastasis formation and evidence
has been presented for the involvement of the exosome in this
mechanism. King et al found an increased concentration of
released exosomes and higher expression of exosomal miR-210
secreted by breast cancer cells under hypoxic conditions when
compared to normoxic cells (116). In leukemia, the miR-210 can also
be found in a subset of miRNAs upregulated in exosomes released by
tumor cells under hypoxic conditions (117).
The exosomal miRNA expression profiling from serum
and plasma samples has also been assessed in glioblastoma, where
the expression of 11 miRNAs known to be upregulated was slightly
lower in exosomes than in the donor cells but still reflecting the
tumor profile (118). When the
serum samples from ovarian cancer patients were evaluated, a
distinct exosomal miRNA profile was identified from that of benign
disease (119). Another study
reporting a potential use of exosomal miRNAs as diagnostic markers
showed a higher expression of miR-21 released from esophageal
cancer serum samples when compared to non-tumoral samples, and it
correlated with advanced tumor stages, lymph node involvement and
metastasis (120). Exosomal
let-7a, miR-1229, miR-1246, miR-150, miR-21, miR-223 and miR-23a
from colorectal tumor samples and cancer cell lines are more highly
expressed than those from healthy controls samples and normal colon
cell lines, and the expression levels of these miRNAs are
significantly decreased in exosomes samples collected after tumor
resection, indicating the cancer status (121).
The exosomal miRNA profiling from plasma samples was
assessed to develop a diagnostic screening method for lung
adenocarcinoma. The expression pattern of 12 specific upregulated
miRNAs (miR-17-3p, miR-21, miR-106a, miR-146, miR-155, miR-191,
miR-192, miR-203, miR-205, miR-210, miR-212, miR-214) in tumor
samples was similar in the tumor plasma-derived exosomes and
distinct from the control samples, indicating exosomal miRNAs could
be relevant as a screening method for this tumor (122). In another report, the exosomal
miRNAs miR-378a, miR-379, miR-139-5p and miR-200-5p were identified
as possible markers to distinguish tumor from normal samples in
lung adenocarcinoma (123). In
addition, the miRNAs miR-151-5p, miR-30a-3p, miR-200b-5p, miR-629,
miR-100 and miR-154-3p are possible markers to discriminate lung
adenocarcinoma from granulomas (123). More recently, Rodríguez et
al evaluated the exosomes derived from bronchoalveolar lavage
(BAL) and plasma samples from lung cancer, in which the exosomal
miRNA content derived from tumor plasma samples is more elevated
than in the BAL, suggesting that the a higher concentration of
exosomal miRNAs are released in the plasma than in the
bronchoalveolar fluid (124).
The use of exosome as a diagnostic tool has also
been evaluated in other fluids than plasma and serum samples.
Recently, a report identified the exosomal miRNAs in bile from
cholangiocarcinoma patients and a potential diagnostic panel that
includes miR-486-3p, miR-16, miR-1274b, miR-484 and miR-191 as
predictive markers (125). In a
study involving cervicovaginal lavage fluids, the miR-21 and
miR-146a were highly expressed in fluids from cervical cancer
samples when compared to those from HPV(+) and HPV(−) normal
samples (126).
Exosomal lncRNAs
The study of lncRNAs is a relatively new field on
cancer research, and many questions about their expression and
functions remain unclear, like the presence of these ncRNAs in the
bodily fluids. Since the use of circulating miRNAs in diagnostic
screening methods and therapeutics have been intensively evaluated
in many tumor types, the presence of released lncRNAs in bodily
fluids, specially within extravesicles as exosomes, could be a
source of novel potential biomarkers for diagnosis, prognosis and
therapeutics purposes. Of our knowledge, there are only few data
about this particular aspect of the lncRNAs (Table II). The ucRNA (ultranconserved
lncRNA) TUC399 was identified in exosomes derived from
hepatocellular cancer cell lines, and the intercellular transfer of
exosomal TUC399 can contribute to tumor growth and progression
(127). More recently, the same
group demonstrated that the expression of lncRNAs linc-RoR (long
intergenic non-coding RNA, regulator of reprogramming) in
hepatocellular cancer is responsive to hypoxic conditions and the
transfer of exosomal linc-RoR can modulate the intercellular
response to hypoxia (128).
| Table IISummary of the reports of the
circulating lncRNAs in cancer. |
Table II
Summary of the reports of the
circulating lncRNAs in cancer.
| Tumor | Sample | Extravesicles
isolation | Long ncRNA | Refs. |
|---|
| Gastric cancer | Plasma samples,
cell lines | NA | H19, HOTAIR,
MALAT1 | (129) |
| Hepatocellular
cancer | Cell line | UC and density
gradient separation | TUC399 | (127) |
| Hepatocellular
cancer | Tissue samples,
plasma samples | NA | HULC | (132) |
| Hepatocellular
cancer | Cell line, animal
model | SC and UC | linc-RoR | (128) |
| Leukemia and
multiple myeloma | Plasma samples | NA | TUG1, MALAT1,
HOTAIR, lincRNA-p21, GAS5 | (133) |
| Prostate
cancer | Tissue samples,
plasma samples | NA | MALAT-1 and
PCA3 | (130) |
At present, reports regarding the circulating
lncRNAs in bodily fluids are also scant in the literature. In a
study evaluating the expression of the lncRNAs H19, HOTAIR and
MALAT1 in gastric cancer plasma samples, Arita et al (2013)
showed that only H19 is higher expressed in tumor samples than in
the controls, and reported significanly decreased expression levels
in post-operative tumor samples, indicating that the release of
lncRNAs into the plasma can reflect the disease status (129). In another report, Ren et
al identified the MALAT1-derived mini-RNA (MD-miniRNA) as
potential novel plasma biomarker in prostate cancer (130). Some reports have demonstrated the
use of lncRNA PCA3 as a specific and reliable marker detectable in
urine samples from patients of prostate cancer, instead of the
standardized use of the prostate-specific antigen (PSA). The
evidence that highly upregulated in liver cancer (HULC) lncRNA
expression is significantly higher in plasma tumor samples than in
the healthy controls indicates the use of this lncRNA as potential
circulating biomarker for diagnosis in hepatocellular cancer
(131,132). The evaluation of lncRNAs
expression in plasma samples from leukemia and multiple myeloma
showed that TUG1, MALAT1, HOTAIR and GAS5 are more highly expressed
in leukemia than in the control samples, and only lincRNA-p21 is
upregulated in multiple myeloma (133).
However, some limitations in using exosomal ncRNAs
in diagnostics have been pointed out by many authors. The
specificity and sensitivity of exosomal tumor marker detection in
bodily fluids is still challenging. For example, serum and
plasma-derived extravesicles as exosomes can be released by other
than tumor cells, such as different blood cell types, affecting the
purity of the tumor-derived exosome samples. In addition, the
release of these exosomes depends on the age of the patient,
infection or inflammation status of the disease, possibly
introducing a bias in comparison analysis if not appropriately
normalized to these conditions. Another issue that must be
considered is the need of a standardized protocol for collecting
and handling of the samples, as well as the exosomes isolation
method (95,106).
Relevance in therapy
Since the miRNA is able to target multiple genes and
signaling networks simultaneously, acting like oncogenes or tumor
suppressor factors, it makes them a suitable tool for therapeutics
interventions. A well and highly specific design is necessary for a
successful result and to prevent undesirable targets. However, one
of the principal limitations of this approach is the nuclease
activity, causing the degradation before the miRNAs can achieve the
targets. The use of vesicles for the delivery of exogenous
therapeutic molecules to the targets has been intensively
considered as a new promising therapeutic intervention. As
mentioned before, the exosomes have the ability to transfer
functional proteins and transcripts as perfect non-immunogenic
carriers of therapeutic agents to target cells, making them
suitable as therapeutic tool (134).
Considering that the exosome content can act as
modulator of the microenvironment, facilitating tumor growth and
metastasis, the blockade of the production, release and uptake by
receptor cells could reverse the influence of the increased levels
of exosomes in tumor progression (95). Based on this, focusing on the
inhibition of key components of the extravesicle production and
release, such as the members of the ESCRT machinery, could be a
useful strategy for therapy (6).
A third possible direction is represented by the
drug or gene delivery by extravesicles to the target sites.
Considering the elucidation of the intercellular transfer by
exosomes, many reports have demonstrated the use of the
extravesicles as small RNA carriers (135,136). Intercellular transfer by exosomes
can be used as miRNA carriers to restore miRNA expression in the
target cells, where they play a therapeutic role as tumor
suppressor factors. The targeted delivery of miRNAs by exosomes was
demonstrated in a study in breast cancer cells expressing high
levels of EGFR. This was achieved by the engineering of the donor
cells to modify the surface of the exosomes to express the
transmembrane domain of the PDGFR fused to the GE11 peptide. Then,
the modified exosomes can deliver the let-7a miRNA after
intravenous injection to EGFR-expressing xenograft breast cancer
tissue in immunodeficient mice (137). The ability of the miRNAs to
target multiple genes can be a limitation for the specificity of
this method as a selective approach for targeted therapy. The use
of synthetic siRNA has been exploited as a more selective
therapeutic tool. In an interesting study reported by
Alvarez-Erviti et al, dendritic cells expressing a specific
protein of the exosomal membrane Lamp2b fused to a
neuron-penetrating RVG peptide were isolated from mice, and the
exosomes derived from these cells were loaded with exogenous siRNA
to GAPDH by electroporation. Subsequently, these RVG-targeted
exosomes were intravenously injected, which delivered GAPDH siRNA
to specific cells in the brain, leading to a selective gene
knockdown (138). Similarly,
another report showed the delivery of siRNA into monocytes and
lymphocytes by exosomes as gene delivery vector, reflecting a
selective gene silencing of MAPK1 (139).
5. Concluding remarks
The discovery of the intercellular communication by
the extravesicles has opened a new field for tumor biology.
Exosomes can be found in the bodily fluids in a variety of tumor
types and many reports have proved that the exosomal content as
proteins, mRNA, miRNA and DNA can reflect the disease status,
making them suitable for biomarkers for non-invasive diagnostic and
prognosis purposes. With the advance of the engineering that
permits the manipulation of the exosomal content and surface
markers, many studies have been focusing on the development of
therapeutic approaches in various tumor types, involving more
specific delivery to the target tumor cells with more selective and
efficient results. However, despite the efforts focusing on the
study of the extracellular vesicles, specially exosomes, there are
many aspects of the their biology that still need to be elucidated
so that it would improve the advantages of the use of this
promising approach in tumors.
Acknowledgements
G.A.C. is The Alan M. Gewirtz Leukemia &
Lymphoma Society Scholar. Study in G.A.C.’s laboratory is supported
in part by the NIH/NCI grants 1UH2TR00943-01 and 1 R01 CA182905-01,
Developmental Research Awards in Prostate Cancer, Multiple Myeloma,
Leukemia (P50 CA100632) and Head and Neck (P50 CA097007) SPOREs, a
SINF MDACC_ DKFZ grant in CLL, a SINF grant in colon cancer, a
Kidney Cancer Pilot Project, the Duncan Family Institutional Seed
Funds, The Blanton-Davis Ovarian Cancer - 2013 Sprint for Life
Research Award, the Laura and John Arnold Foundation, the RGK
Foundation and the Estate of C. G. Johnson, Jr and by the CLL
Global Research Foundation.
References
|
1
|
Mattick JS: Non-coding RNAs: the
architects of eukaryotic complexity. EMBO Rep. 2:986–991. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Morceau F, Chateauvieux S, Gaigneaux A,
Dicato M and Diederich M: Long and short non-coding RNAs as
regulators of hematopoietic differentiation. Int J Mol Sci.
14:14744–14770. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mattick JS and Makunin IV: Non-coding RNA.
Hum Mol Genet. 15(Spec 1): R17–R29. 2006. View Article : Google Scholar
|
|
4
|
Amaral PP and Mattick JS: Noncoding RNA in
development. Mamm Genome. 19:454–492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Setoyama T, Ling H, Natsugoe S and Calin
GA: Non-coding RNAs for medical practice in oncology. Keio J Med.
60:106–113. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vader P, Breakefield XO and Wood MJA:
Extracellular vesicles: emerging targets for cancer therapy. Trends
Mol Med. 20:385–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Prensner JR and Chinnaiyan AM: The
emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee RC, Feinbaum RL and Ambros V: The
C. elegans heterochronic gene lin-4 encodes small RNAs with
antisense complementarity to lin-14. Cell. 75:843–854. 1993.
View Article : Google Scholar
|
|
10
|
Wightman B, Ha I and Ruvkun G:
Posttranscriptional regulation of the heterochronic gene lin-14 by
lin-4 mediates temporal pattern formation in C. elegans.
Cell. 75:855–862. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang W, Dahlberg JE and Tam W: MicroRNAs
in tumorigenesis: a primer. Am J Pathol. 171:728–738. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Siomi H and Siomi MC: Posttranscriptional
regulation of microRNA biogenesis in animals. Mol Cell. 38:323–332.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Raisch J, Darfeuille-Michaud A and Nguyen
HTT: Role of microRNAs in the immune system, inflammation and
cancer. World J Gastroenterol. 19:2985–2996. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ling H, Fabbri M and Calin GA: MicroRNAs
and other non-coding RNAs as targets for anticancer drug
development. Nat Rev Drug Discov. 12:847–865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Calin GA, Dumitru CD, Shimizu M, et al:
Frequent deletions and down-regulation of micro-RNA genes miR15 and
miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci
USA. 99:15524–15529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yendamuri S and Calin GA: The role of
microRNA in human leukemia: a review. Leukemia. 23:1257–1263. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Balatti V, Pekarky Y, Rizzotto L and Croce
CM: miR deregulation in CLL. Adv Exp Med Biol. 792:309–325. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gordon JEA, Wong JJ-L and Rasko JEJ:
MicroRNAs in myeloid malignancies. Br J Haematol. 162:162–176.
2013. View Article : Google Scholar
|
|
19
|
Srivastava S, Tsongalis GJ and Kaur P:
Recent advances in microRNA-mediated gene regulation in chronic
lymphocytic leukemia. Clin Biochem. 46:901–908. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rossi S, Kopetz S, Davuluri R, Hamilton SR
and Calin GA: MicroRNAs, ultraconserved genes and colorectal
cancers. Int J Biochem Cell Biol. 42:1291–1297. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hong L, Han Y, Zhou Y and Nita A:
Angiogenesis-related microRNAs in colon cancer. Expert Opin Biol
Ther. 13:77–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hutchison J, Cohen Z, Onyeagucha BC, Funk
J and Nelson MA: How microRNAs influence both hereditary and
inflammatory-mediated colon cancers. Cancer Genet. 206:309–316.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Menéndez P, Villarejo P, Padilla D,
Menéndez JM and Rodríguez-Montes JA: Implications of the
histological determination of microRNAs in the screening, diagnosis
and prognosis of colorectal cancer. J Surg Oncol. 108:70–73.
2013.PubMed/NCBI
|
|
24
|
Ferracin M, Querzoli P, Calin GA and
Negrini M: MicroRNAs: toward the clinic for breast cancer patients.
Semin Oncol. 38:764–775. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harquail J, Benzina S and Robichaud GA:
MicroRNAs and breast cancer malignancy: an overview of
miRNA-regulated cancer processes leading to metastasis. Cancer
Biomark. 11:269–280. 2012.PubMed/NCBI
|
|
26
|
Zhang ZJ and Ma SL: miRNAs in breast
cancer tumorigenesis (Review). Oncol Rep. 27:903–910.
2012.PubMed/NCBI
|
|
27
|
Mulrane L, McGee SF, Gallagher WM and
O’Connor DP: miRNA dysregulation in breast cancer. Cancer Res.
73:6554–6562. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singh R and Mo Y-Y: Role of microRNAs in
breast cancer. Cancer Biol Ther. 14:201–212. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
John K, Wu J, Lee B-W and Farah CS:
MicroRNAs in head and neck cancer. Int J Dent. 2013:6502182013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagadia R, Pandit P, Coman WB,
Cooper-White J and Punyadeera C: miRNAs in head and neck cancer
revisited. Cell Oncol (Dordr). 36:1–7. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nohata N, Hanazawa T, Kinoshita T, Okamoto
Y and Seki N: MicroRNAs function as tumor suppressors or oncogenes:
aberrant expression of microRNAs in head and neck squamous cell
carcinoma. Auris Nasus Larynx. 40:143–149. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tu HF, Lin SC and Chang KW: MicroRNA
aberrances in head and neck cancer: pathogenetic and clinical
significance. Curr Opin Otolaryngol Head Neck Surg. 21:104–111.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Song JH and Meltzer SJ: MicroRNAs in
pathogenesis, diagnosis, and treatment of gastroesophageal cancers.
Gastroenterology. 143:35–47.e2. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang F, Sun GP, Zou YF, Hao JQ, Zhong F
and Ren WJ: MicroRNAs as promising biomarkers for gastric cancer.
Cancer Biomark. 11:259–267. 2012.PubMed/NCBI
|
|
35
|
Gao M, Yin H and Fei ZW: Clinical
application of microRNA in gastric cancer in Eastern Asian area.
World J Gastroenterol. 19:2019–2027. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Song S and Ajani JA: The role of microRNAs
in cancers of the upper gastrointestinal tract. Nat Rev
Gastroenterol Hepatol. 10:109–118. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kapranov P, Cheng J, Dike S, et al: RNA
maps reveal new RNA classes and a possible function for pervasive
transcription. Science. 316:1484–1488. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Okazaki Y, Furuno M, Kasukawa T, et al:
Analysis of the mouse transcriptome based on functional annotation
of 60,770 full-length cDNAs. Nature. 420:563–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar
|
|
40
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: a new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tang JY, Lee JC, Chang YT, Hou MF, Huang
HW, Liaw CC and Chang HW: Long noncoding RNAs-related diseases,
cancers, and drugs. Sci World J. 2013:9435392013.PubMed/NCBI
|
|
42
|
Van Roosbroeck K, Pollet J and Calin GA:
miRNAs and long noncoding RNAs as biomarkers in human diseases.
Expert Rev Mol Diagn. 13:183–204. 2013.PubMed/NCBI
|
|
43
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: a new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fitzgerald KA and Caffrey DR: Long
noncoding RNAs in innate and adaptive immunity. Curr Opin Immunol.
26:140–146. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jendrzejewski J, He H, Radomska HS, et al:
The polymorphism rs944289 predisposes to papillary thyroid
carcinoma through a large intergenic noncoding RNA gene of tumor
suppressor type. Proc Natl Acad Sci USA. 109:8646–8651. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fan M, Li X, Jiang W, Huang Y, Li J and
Wang Z: A long non-coding RNA, PTCSC3, as a tumor suppressor and a
target of miRNAs in thyroid cancer cells. Exp Ther Med.
5:1143–1146. 2013.PubMed/NCBI
|
|
47
|
Hessels D and Schalken JA: The use of PCA3
in the diagnosis of prostate cancer. Nat Rev Urol. 6:255–261. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Day JR, Jost M, Reynolds MA, Groskopf J
and Rittenhouse H: PCA3: from basic molecular science to the
clinical lab. Cancer Lett. 301:1–6. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Pasmant E, Laurendeau I, Héron D, Vidaud
M, Vidaud D and Bièche I: Characterization of a germ-line deletion,
including the entire INK4/ARF locus, in a melanoma-neural system
tumor family: identification of ANRIL, an antisense noncoding RNA
whose expression coclusters with ARF. Cancer Res. 67:3963–3969.
2007. View Article : Google Scholar
|
|
50
|
Iacobucci I, Sazzini M, Garagnani P, et
al: A polymorphism in the chromosome 9p21 ANRIL locus is associated
to Philadelphia positive acute lymphoblastic leukemia. Leuk Res.
35:1052–1059. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cheetham SW, Gruhl F, Mattick JS and
Dinger ME: Long noncoding RNAs and the genetics of cancer. Br J
Cancer. 108:2419–2425. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cheng W, Zhang Z and Wang J: Long
noncoding RNAs: new players in prostate cancer. Cancer Lett.
339:8–14. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ji P, Diederichs S, Wang W, et al:
MALAT-1, a novel noncoding RNA, and thymosin beta4 predict
metastasis and survival in early-stage non-small cell lung cancer.
Oncogene. 22:8031–8041. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gutschner T and Diederichs S: The
hallmarks of cancer: a long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li CH and Chen Y: Targeting long
non-coding RNAs in cancers: progress and prospects. Int J Biochem
Cell Biol. 45:1895–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Martens-Uzunova ES, Olvedy M and Jenster
G: Beyond microRNA-novel RNAs derived from small non-coding RNA and
their implication in cancer. Cancer Lett. 340:201–211. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen X, Ba Y, Ma L, et al:
Characterization of microRNAs in serum: a novel class of biomarkers
for diagnosis of cancer and other diseases. Cell Res. 18:997–1006.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gilad S, Meiri E, Yogev Y, et al: Serum
microRNAs are promising novel biomarkers. PLoS One. 3:e31482008.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
De Planell-Saguer M and Rodicio MC:
Analytical aspects of microRNA in diagnostics: a review. Anal Chim
Acta. 699:134–152. 2011.PubMed/NCBI
|
|
60
|
Ajit SK: Circulating microRNAs as
biomarkers, therapeutic targets, and signaling molecules. Sensors
(Basel). 12:3359–3369. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Schwarzenbach H, Nishida N, Calin GA and
Pantel K: Clinical relevance of circulating cell-free microRNAs in
cancer. Nat Rev Clin Oncol. 11:145–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lawrie CH, Gal S, Dunlop HM, et al:
Detection of elevated levels of tumour-associated microRNAs in
serum of patients with diffuse large B-cell lymphoma. Br J
Haematol. 141:672–675. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Luo X, Stock C, Burwinkel B and Brenner H:
Identification and evaluation of plasma microRNAs for early
detection of colorectal cancer. PLoS One. 8:e628802013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang J, Huang S-K, Zhao M, et al:
Identification of a circulating microRNA signature for colorectal
cancer detection. PLoS One. 9:e874512014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Komatsu S, Ichikawa D, Takeshita H, et al:
Circulating microRNAs in plasma of patients with oesophageal
squamous cell carcinoma. Br J Cancer. 105:104–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wu C, Wang C, Guan X, et al: Diagnostic
and prognostic implications of a serum miRNA panel in oesophageal
squamous cell carcinoma. PLoS One. 9:e922922014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chan M, Liaw CS, Ji SM, et al:
Identification of circulating microRNA signatures for breast cancer
detection. Clin Cancer Res. 19:4477–4487. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zearo S, Kim E, Zhu Y, Zhao JT, Sidhu SB,
Robinson BG and Soon PS: MicroRNA-484 is more highly expressed in
serum of early breast cancer patients compared to healthy
volunteers. BMC Cancer. 14:2002014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tsujiura M, Ichikawa D, Komatsu S, et al:
Circulating microRNAs in plasma of patients with gastric cancers.
Br J Cancer. 102:1174–1179. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhu C, Ren C, Han J, et al: A
five-microRNA panel in plasma was identified as potential biomarker
for early detection of gastric cancer. Br J Cancer. 110:2291–2299.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zheng H, Liu JY, Song FJ and Chen KX:
Advances in circulating microRNAs as diagnostic and prognostic
markers for ovarian cancer. Cancer Biol Med. 10:123–130.
2013.PubMed/NCBI
|
|
72
|
Shapira I, Oswald M, Lovecchio J, et al:
Circulating biomarkers for detection of ovarian cancer and
predicting cancer outcomes. Br J Cancer. 110:976–983. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kosaka N, Iguchi H and Ochiya T:
Circulating microRNA in body fluid: a new potential biomarker for
cancer diagnosis and prognosis. Cancer Sci. 101:2087–2092. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ahmed KA and Xiang J: Mechanisms of
cellular communication through intercellular protein transfer. J
Cell Mol Med. 15:1458–1473. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Ogorevc E, Kralj-Iglic V and Veranic P:
The role of extracellular vesicles in phenotypic cancer
transformation. Radiol Oncol. 47:197–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lee TH, D’Asti E, Magnus N, Al-Nedawi K,
Meehan B and Rak J: Microvesicles as mediators of intercellular
communication in cancer - the emerging science of cellular
‘debris’. Semin Immunopathol. 33:455–467. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang HG and Grizzle WE: Exosomes: a novel
pathway of local and distant intercellular communication that
facilitates the growth and metastasis of neoplastic lesions. Am J
Pathol. 184:28–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Choi DS, Kim DK, Kim YK and Gho YS:
Proteomics, transcriptomics and lipidomics of exosomes and
ectosomes. Proteomics. 13:1554–1571. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
El Andaloussi S, Mäger I, Breakefield XO
and Wood MJA: Extracellular vesicles: biology and emerging
therapeutic opportunities. Nat Rev Drug Discov. 12:347–357.
2013.PubMed/NCBI
|
|
80
|
Raposo G and Stoorvogel W: Extracellular
vesicles: exosomes, microvesicles, and friends. J Cell Biol.
200:373–383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Van der Pol E, Böing AN, Harrison P, Sturk
A and Nieuwland R: Classification, functions, and clinical
relevance of extracellular vesicles. Pharmacol Rev. 64:676–705.
2012.PubMed/NCBI
|
|
82
|
Simpson RJ, Jensen SS and Lim JWE:
Proteomic profiling of exosomes: current perspectives. Proteomics.
8:4083–4099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Nazarenko I, Rupp A-K and Altevogt P:
Exosomes as a potential tool for a specific delivery of functional
molecules. Methods Mol Biol. 1049:495–511. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Principe S, Hui ABY, Bruce J, Sinha A, Liu
FF and Kislinger T: Tumor-derived exosomes and microvesicles in
head and neck cancer: implications for tumor biology and biomarker
discovery. Proteomics. 13:1608–1623. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Akers JC, Gonda D, Kim R, Carter BS and
Chen CC: Biogenesis of extracellular vesicles (EV): exosomes,
microvesicles, retro-virus-like vesicles, and apoptotic bodies. J
Neurooncol. 113:1–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Mathivanan S, Ji H and Simpson RJ:
Exosomes: extracellular organelles important in intercellular
communication. J Proteomics. 73:1907–1920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Kosaka N, Yoshioka Y, Hagiwara K, Tominaga
N, Katsuda T and Ochiya T: Trash or treasure: extracellular
microRNAs and cell-to-cell communication. Front Genet. 4:1732013.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Martins VR, Dias MS and Hainaut P:
Tumor-cell-derived microvesicles as carriers of molecular
information in cancer. Curr Opin Oncol. 25:66–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Kahlert C, Melo SA, Protopopov A, et al:
Identification of double-stranded genomic DNA spanning all
chromosomes with mutated KRAS and p53 DNA in the serum exosomes of
patients with pancreatic cancer. J Biol Chem. 289:3869–3875. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Fang DY, King HW, Li JY and Gleadle JM:
Exosomes and the kidney: blaming the messenger. Nephrology
(Carlton). 18:1–10. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Gajos-Michniewicz A, Duechler M and Czyz
M: MiRNA in melanoma-derived exosomes. Cancer Lett. 347:29–37.
2014. View Article : Google Scholar
|
|
92
|
Peinado H, Lavotshkin S and Lyden D: The
secreted factors responsible for pre-metastatic niche formation:
old sayings and new thoughts. Semin Cancer Biol. 21:139–146. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Théry C, Amigorena S, Raposo G and Clayton
A: Isolation and characterization of exosomes from cell culture
supernatants and biological fluids. Curr Protoc Cell Biol. Chapter
3(Unit 3): 222006.PubMed/NCBI
|
|
94
|
Lässer C: Identification and analysis of
circulating exosomal microRNA in human body fluids. Methods Mol
Biol. 1024:109–128. 2013.PubMed/NCBI
|
|
95
|
Gonda DD, Akers JC, Kim R, Kalkanis SN,
Hochberg FH, Chen CC and Carter BS: Neuro-oncologic applications of
exosomes, microvesicles, and other nano-sized extracellular
particles. Neurosurgery. 72:501–510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Lässer C, Eldh M and Lötvall J: Isolation
and characterization of RNA-containing exosomes. J Vis Exp.
e30372012.
|
|
97
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Jenjaroenpun P, Kremenska Y, Nair VM,
Kremenskoy M, Joseph B and Kurochkin IV: Characterization of RNA in
exosomes secreted by human breast cancer cell lines using
next-generation sequencing. Peer J. 1:e2012013. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Kruger S, Abd Elmageed ZY, Hawke DH, et
al: Molecular characterization of exosome-like vesicles from breast
cancer cells. BMC Cancer. 14:442014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Feng DQ, Huang B, Li J, et al: Selective
miRNA expression profile in chronic myeloid leukemia K562
cell-derived exosomes. Asian Pac J Cancer Prev. 14:7501–7508. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xiao D, Ohlendorf J, Chen Y, et al:
Identifying mRNA, microRNA and protein profiles of melanoma
exosomes. PLoS ONE. 7:e468742012. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Rappa G, Mercapide J, Anzanello F, Pope RM
and Lorico A: Biochemical and biological characterization of
exosomes containing prominin-1/CD133. Mol Cancer. 12:622013.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Hessvik NP, Phuyal S, Brech A, Sandvig K
and Llorente A: Profiling of microRNAs in exosomes released from
PC-3 prostate cancer cells. Biochim Biophys Acta. 1819.1154–1163.
2012.PubMed/NCBI
|
|
104
|
Kobayashi M, Salomon C, Tapia J, Illanes
SE, Mitchell MD and Rice GE: Ovarian cancer cell invasiveness is
associated with discordant exosomal sequestration of Let-7 miRNA
and miR-200. J Transl Med. 12:42014. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Ohshima K, Inoue K, Fujiwara A, et al:
Let-7 microRNA family is selectively secreted into the
extracellular environment via exosomes in a metastatic gastric
cancer cell line. PLoS One. 5:e132472010. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zöller M: Pancreatic cancer diagnosis by
free and exosomal miRNA. World J Gastrointest Pathophysiol.
4:74–90. 2013.PubMed/NCBI
|
|
107
|
Chiba M, Kimura M and Asari S: Exosomes
secreted from human colorectal cancer cell lines contain mRNAs,
microRNAs and natural antisense RNAs, that can transfer into the
human hepatoma HepG2 and lung cancer A549 cell lines. Oncol Rep.
28:1551–1558. 2012.
|
|
108
|
Yang M, Chen J, Su F, et al: Microvesicles
secreted by macrophages shuttle invasion-potentiating microRNAs
into breast cancer cells. Mol Cancer. 10:1172011. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Umezu T, Ohyashiki K, Kuroda M and
Ohyashiki JH: Leukemia cell to endothelial cell communication via
exosomal miRNAs. Oncogene. 32:2747–2755. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Kosaka N, Iguchi H, Yoshioka Y, Hagiwara
K, Takeshita F and Ochiya T: Competitive interactions of cancer
cells and normal cells via secretory microRNAs. J Biol Chem.
287:1397–1405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Roccaro AM, Sacco A, Maiso P, et al: BM
mesenchymal stromal cell-derived exosomes facilitate multiple
myeloma progression. J Clin Invest. 123:1542–1555. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Valencia K, Luis-Ravelo D, Bovy N, et al:
miRNA cargo within exosome-like vesicle transfer influences
metastatic bone colonization. Mol Oncol. 8:689–703. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Zhou W, Fong MY, Min Y, et al:
Cancer-secreted miR-105 destroys vascular endothelial barriers to
promote metastasis. Cancer Cell. 25:501–515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Chen W, Zhong S, Ji M, et al: MicroRNAs
delivered by extracellular vesicles: an emerging resistance
mechanism for breast cancer. Tumour Biol. 35:2883–2892. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Xiao X, Yu S, Li S, et al: Exosomes:
decreased sensitivity of lung cancer A549 cells to cisplatin. PLoS
One. 9:e895342014. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
King HW, Michael MZ and Gleadle JM:
Hypoxic enhancement of exosome release by breast cancer cells. BMC
Cancer. 12:4212012. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Tadokoro H, Umezu T, Ohyashiki K, Hirano T
and Ohyashiki JH: Exosomes derived from hypoxic leukemia cells
enhance tube formation in endothelial cells. J Biol Chem.
288:34343–34351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Skog J, Würdinger T, van Rijn S, et al:
Glioblastoma microvesicles transport RNA and proteins that promote
tumour growth and provide diagnostic biomarkers. Nat Cell Biol.
10:1470–1476. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Tanaka Y, Kamohara H, Kinoshita K, et al:
Clinical impact of serum exosomal microRNA-21 as a clinical
biomarker in human esophageal squamous cell carcinoma. Cancer.
119:1159–1167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Ogata-Kawata H, Izumiya M, Kurioka D, et
al: Circulating exosomal microRNAs as biomarkers of colon cancer.
PLoS One. 9:e929212014. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Rabinowits G, Gerçel-Taylor C, Day JM,
Taylor DD and Kloecker GH: Exosomal microRNA: a diagnostic marker
for lung cancer. Clin Lung Cancer. 10:42–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Cazzoli R, Buttitta F, Di Nicola M,
Malatesta S, Marchetti A, Rom WN and Pass HI: microRNAs derived
from circulating exosomes as noninvasive biomarkers for screening
and diagnosing lung cancer. J Thorac Oncol. 8:1156–1162. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Rodríguez M, Silva J, López-Alfonso A, et
al: Different exosome cargo from plasma/bronchoalveolar lavage in
non-small-cell lung cancer. Genes Chromosomes Cancer. Apr
25–2014.(Epub ahead of print).
|
|
125
|
Li L, Masica D, Ishida M, et al: Human
bile contains microRNA-laden extracellular vesicles that can be
used for cholangiocarcinoma diagnosis. Hepatology. Feb 4–2014.(Epub
ahead of print).
|
|
126
|
Liu J, Sun H, Wang X, Yu Q, Li S, Yu X and
Gong W: Increased exosomal microRNA-21 and microRNA-146a levels in
the cervicovaginal lavage specimens of patients with cervical
cancer. Int J Mol Sci. 15:758–773. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Kogure T, Yan IK, Lin W-L and Patel T:
Extracellular vesicle-mediated transfer of a novel long noncoding
RNA TUC339: A mechanism of intercellular signaling in human
hepatocellular cancer. Genes Cancer. 4:261–272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Takahashi K, Yan IK, Haga H and Patel T:
Modulation of hypoxia-signaling pathways by extracellular linc-RoR.
J Cell Sci. 127:1585–1594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Arita T, Ichikawa D, Konishi H, et al:
Circulating long non-coding RNAs in plasma of patients with gastric
cancer. Anticancer Res. 33:3185–3193. 2013.PubMed/NCBI
|
|
130
|
Ren S, Wang F, Shen J, et al: Long
non-coding RNA metastasis associated in lung adenocarcinoma
transcript 1 derived miniRNA as a novel plasma-based biomarker for
diagnosing prostate cancer. Eur J Cancer. 49:2949–2959. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Panzitt K, Tschernatsch MMO, Guelly C, et
al: Characterization of HULC, a novel gene with striking
up-regulation in hepatocellular carcinoma, as noncoding RNA.
Gastroenterology. 132:330–342. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Xie H, Ma H and Zhou D: Plasma HULC as a
promising novel biomarker for the detection of hepatocellular
carcinoma. Biomed Res Int. 2013:1361062013.PubMed/NCBI
|
|
133
|
Isin M, Ozgur E, Cetin G, Erten N, Aktan
M, Gezer U and Dalay N: Investigation of circulating lncRNAs in
B-cell neoplasms. Clin Chim Acta. 431:255–259. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Lässer C: Exosomal RNA as biomarkers and
the therapeutic potential of exosome vectors. Expert Opin Biol
Ther. 12(Suppl 1): S189–S197. 2012.PubMed/NCBI
|
|
135
|
Lee Y, El Andaloussi S and Wood MJA:
Exosomes and microvesicles: extracellular vesicles for genetic
information transfer and gene therapy. Hum Mol Genet. 21:R125–R134.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Kosaka N, Takeshita F, Yoshioka Y,
Hagiwara K, Katsuda T, Ono M and Ochiya T: Exosomal
tumor-suppressive microRNAs as novel cancer therapy: ‘exocure’ is
another choice for cancer treatment. Adv Drug Deliv Rev.
65:376–382. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Ohno S, Takanashi M, Sudo K, et al:
Systemically injected exosomes targeted to EGFR deliver antitumor
microRNA to breast cancer cells. Mol Ther. 21:185–191. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Alvarez-Erviti L, Seow Y, Yin H, Betts C,
Lakhal S and Wood MJA: Delivery of siRNA to the mouse brain by
systemic injection of targeted exosomes. Nat Biotechnol.
29:341–345. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Wahlgren J, De Karlson LT, Brisslert M,
Vaziri Sani F, Telemo E, Sunnerhagen P and Valadi H: Plasma
exosomes can deliver exogenous short interfering RNA to monocytes
and lymphocytes. Nucleic Acids Res. 40:e1302012. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Bronisz A, Wang Y, Nowicki MO, et al:
Extracellular vesicles modulate the glioblastoma microenvironment
via a tumor suppression signaling network directed by miR-1. Cancer
Res. 74:738–750. 2014. View Article : Google Scholar : PubMed/NCBI
|