Introduction
Hepatocellular carcinoma (HCC), which shows a close
relationship with HBV- and HCV-induced inflammation (1), is associated with a high potential
for vascular invasion, metastasis and recurrence (2). Although inflammation is closely
associated with hepatic cell carcinogenesis (3), the involvement of pro-inflammatory
cytokines, such as tumor necrosis factor-α (TNF-α), in liver
carcinoma metastasis and nutritional status has also been
documented (4,5), indicating that TNF-α plays a
particularly important role in the tumor microenvironment and
promotes tumor cell migration and invasion (6). As tumor tissues are composed of
cancer cells, fibroblasts, vasculature and immune cells, the
finding that activated macrophages secrete TNF-α and promote tumor
metastasis (3) does not exclude
the possibility that other components of the liver cancer
microenvironment might secrete TNF-α and promote HCC metastasis in
an autocrine or paracrine manner. Hence, the role of
microenvironment components in TNF-α-facilitated HCC metastasis
remains elusive.
Extracellular matrix (ECM) degradation and basement
membrane destruction are prerequisites for tumor invasion and
metastasis, which are known to involve matrix metalloproteinases
(MMPs) (7,8). Stromelysin-1 (MMP-3) is an MMP with
broad substrate specificity that has been shown to degrade fiber
mucin (FN), laminin (LN), casein, and proteoglycans and to activate
gelatinase B (MMP-9)/collagenase. MMP-3 has been found to be
associated with early oral squamous cell carcinoma and breast
cancer (9,10). Furthermore, involvement of MMP-3 in
hepatocyte growth factor-induced invasion has also been documented
(11,12). Additionally, human collagenase 3
(MMP-13), which represents the third member of the collagenase
subfamily, has been detected in gastric cancer and breast cancer
(10,13,14).
MMP-13 can be activated by MMP-2/MMP-3, and activated MMP-13, in
turn, has the ability to activate MMP-2 and MMP-9 (8), indicating that MMP-13 plays a central
role in the MMP activation cascade (15). However, few studies conducted thus
far have explored the roles of MMP-13 and MMP-3 in
inflammation-associated HCC metastasis.
Tumor necrosis factor-α-induced protein 8-like 2
(TNFAIP8L2, TIPE2), which belongs to the TNF-α-induced protein 8
family, could inhibit the release of pro-inflammatory cytokines,
such as TNF-α, IL-4, IL-12 and IFN-γ (16). TIPE2 inhibition of MMP-9 expression
(17) and reduction of F-actin
polymerization (18) have also
been documented, indicating that TIPE2 might play an important role
in tumor metastasis. However, little is currently known concerning
the effect of TIPE2 on inflammation-associated metastasis, which is
clearly an important issue for treating HCC metastasis.
The aim of the present study was to identify the
functions of TNF-α and TIPE2 in HCC metastasis by studying changes
in MMP expression and to examine their relationships with Erk1/2
and NF-κB pathway activation. For this purpose, we investigated the
effects of both TNF-α and TIPE2 on MMP-13/MMP-3 expression and cell
migration as well as Erk1/2-NF-κB activation under exogenous TNF-α
conditions. We found that TNF-α was able to increase both
MMP-3/MMP-13 expression and enhance the migratory ability of HepG2
cells by activating the Erk1/2-NF-κB pathways. However, when TIPE2
was overexpressed, these TNF-α-induced effects were inhibited.
These results indicated that TIPE2 might play a negative role in
TNF-α-accelerated HCC metastasis, suggesting that TIPE2 could be a
potential new target for the clinical treatment of liver
cancer.
Materials and methods
Reagents
Lipopolysaccharides (LPS) were obtained from
Sigma-Aldrich (St. Louis, MO, USA). Recombinant human TNF-α,
brefeldin A solution, and fluorescein-conjugated antibodies against
mouse or human TNF-α were purchased from eBioscience (San Diego,
CA, USA). Western blot antibodies and the Erk1/2 inhibitor U0126
were acquired from Cell Signaling Technology (Beverly, MA, USA).
The NF-κB inhibitor Bay-7082 was purchased from Cayman Chemical
(Ann Arbor, MI, USA). SYBR® Premix Ex Taq™, TRIzol and
PrimeScript reverse transcriptase were purchased from Takara
(Dalian, China). Goat anti-rabbit IgG-FITC and IgG-PE were obtained
from Abcam (San Francisco, CA, USA). A transwell migration system
was purchased from Corning (Corning, NY, USA). Lipofectamine 2000
was purchased from Invitrogen (Grand Island, NY, USA). DMEM and
fetal bovine sera were obtained from Hyclone (Logan, UT, USA). The
NE-PER nuclear and cytoplasmic extraction reagents were purchased
from Pierce (Rockford, IL, USA).
Cell lines
The human hepatoblastoma cell line HepG2 was
obtained from the Shanghai Cell Bank (Shanghai, China). Cells were
cultured in DMEM supplemented with 10% fetal bovine serum at 37°C
under 5% CO2. Cells were synchronized by serum
starvation for at least 12 h before TNF-α or LPS treatment.
Establishment of a TIPE2-overexpressing
HepG2 cell line
TIPE2-overexpressing HepG2 cells were established as
described previously (18).
Briefly, the pEGFP-C1-TIPE2 plasmid was generated by inserting
full-length TIPE2 into the pEGFP-C1 vector. HepG2 cells plated at a
density of 6×105 cells/well were transfected with the
plasmid using Lipofectamine 2000, and resistant cells were selected
for using 500 μg/ml G418 antibiotic. Separated cell clones were
confirmed via RT-PCR and stored for further experiments.
Flow cytometric measurements
The effects of LPS or TIPE2 on TNF-α expression were
assayed as described previously (19). Briefly, cells plated at a density
of 2×104 cells/well were stimulated with LPS for 4 h.
For PBLs, the final concentration of LPS was 100 ng/ml. For other
cells, the final concentration of LPS was 1 μg/ml. BFA was added to
inhibit TNF-α secretion. After LPS treatment, the cells were
permeabilized and fixed for 30 min using BD Cytofix/Cytoperm™
fixation/permeabilization solution (BD Biosciences, San Jose, CA,
USA). Then, the cells were washed with Perm/Wash buffer and stained
with specific antibodies for 30 min on ice. Flow cytometry was
performed using a FACSCalibur flow cytometer, and the data were
analyzed using CellQuest software.
Confocal immunofluorescence assays
The effects of TNF-α or TIPE2 on p65, Erk1/2
phosphorylation and c-Fos expression were investigated using
immunofluorescence assays (19).
Briefly, cells were treated with TNF-α (2 ng/ml), fixed and
permeabilized in 100% methanol. Then, the cells were blocked with
10% non-fat milk and incubated with primary antibodies
(anti-phospho-p65, anti-phospho-Erk, or anti-c-Fos; 1:100 dilution)
overnight at 4°C, followed by staining with FITC-conjugated IgG
(1:100 dilution) or PE-conjugated IgG (1:1,000 dilution). Finally,
the cells were mounted in mounting medium containing DAPI. Images
were captured using a confocal fluorescence microscope at 488 or
546 nm.
RT-PCR and qPCR
The effects of TIPE2 on MMP-3/MMP-13 and TNF-α
expression were investigated via RT-PCR and real-time PCR analyses
as described previously (20).
Briefly, whole cellular RNA was extracted, and
reverse-transcription was performed using PrimeScript reverse
transcriptase. PCR amplification was conducted using the following
conditions: 95°C for 3 min, followed by 30 cycles of 95°C for 5
sec, 60°C for 30 sec and 72°C for 60 sec and a final extension at
72°C for 10 min. β-actin was used as an internal control. The PCR
products were run on 1.5% agarose gels and analyzed under
ultraviolet (UV) light after ethidium bromide staining. To quantify
gene amplification, real-time PCR analysis was performed using an
ABI 7000 Sequence Detection System in the presence of SYBR Green.
The cycling parameters were 95°C for 3 min, followed by 40 cycles
of 95°C for 5 sec, 60°C for 30 sec, and 72°C for 60 sec, with a
final extension at 72°C for 10 min; a melting curve analysis was
subsequently conducted. The relative expression levels (defined as
fold-changes) of the target genes were normalized to the fold-value
of the corresponding control cells. The following primer sequences
were used in these assays (Table
I).
 | Table IPrimer sequence table. |
Table I
Primer sequence table.
| Genes | F/R | Sequence |
|---|
| β-actin | F |
5′-ACCGTGGAGAAGAGCTACGA-3′ |
| β-actin | R |
5′-GTACTTGCGCTCAGAAGGAG-3′ |
| TIPE2 | F |
5′-CACCGCAATGGCTCCTTT-3′ |
| TIPE2 | R |
5′-CACCAACTCTAGCAGCACATC-3′ |
| TNF-α | F |
5′-GAAAGCATGATCCGGGACGTG-3′ |
| TNF-α | R |
5′-GATGGCAGAGAGGAGGTTGAC-3′ |
| MMP-13 | F |
5′-TTGTTGCTGCGCATGAGTTCG-3′ |
| MMP-13 | R |
5′-GGGTGCTCATATGCAGCATCA-3′ |
| MMP-3 | F |
5′-CCTGCTTTGTCCTTTGATGC-3′ |
| MMP-3 | R |
5′-TGAGTCAATCCCTGGAAAGTC-3′ |
Isolation of cytoplasmic and nuclear
extracts
Cytoplasmic and nuclear extracts were prepared from
whole-cell extracts as described previously (19). Briefly, following TNF-α (2 ng/ml)
treatment, the cells were suspended and vortexed in CER
(cytoplasmic extraction reagent) buffer. The cytosolic fraction was
separated via centrifugation (16,000 × g, 5 min, 4°C), and nuclear
proteins were separated by incubating the insoluble fraction with
NER (nuclear extraction reagent) for 40 min, followed by
centrifugation at 16,000 × g for 10 min. The obtained protein
concentration was estimated, and the nuclear extract was further
analyzed by western blot analysis.
Western blot analysis
The phosphorylation of Erk1/2-c-Fos and NF-κB and
the expression of related proteins were determined via western blot
analysis (21). Briefly, cells
were pretreated with kinase inhibitors 2 h before TNF-α (2 ng/ml)
stimulation, and whole cellular protein was extracted. Then, the
protein was loaded onto 10–12% SDS-PAGE gels and transferred onto a
PVDF membrane. After blocking with 5% non-fat milk, the membrane
was incubated with primary antibodies, followed by HRP-conjugated
secondary antibodies. The bands were developed using an ECL
chemiluminescence substrate. β-actin, tubulin or histone 3 was used
as a loading control.
Transwell migration assay
The effects of TNF-α and TIPE2 on cell migration
were investigated in a transwell migration assay as previously
described (17). Briefly,
4×103 cells were seeded into the top chamber in
serum-free medium (12-well insert, 12 μm; Corning) and pretreated
with related kinase inhibitors 2 h before TNF-α (2 ng/ml)
stimulation. After 48 h of incubation, cell migration was
determined by calculating the number of migrated cells in three
visual fields per well via microscopy at ×100 magnification.
ELISA
To determine the effect of TNF-α on MMP-3 and MMP-13
activities, cells (3×106) were treated with TNF-α (2
ng/ml) for 24 h, and the supernatant was collected. MMP-3/MMP-13
activities were determined via ELISA (22). Briefly, the plates were pre-coated
with a coating antibody at 4°C overnight, washed with PBST, and
blocked with assay buffer. Then, the samples, MMP-3/MMP-13 standard
proteins, assay buffer, and biotin-conjugate detector antibodies
were added to the wells, followed by incubation for 2 h. After
incubation, each plate was washed with PBST, and streptavidin-HRP
was added, followed by incubation for 1 h. The color reaction was
developed with TMB solution, and absorbance was measured at 450
nm.
Statistical analysis
All experiments were repeated at least three times
to confirm the results. The data are presented as the mean ± SEM.
Student’s t-test, one-way ANOVA with the Newman-Keuls post-test or
two-way ANOVA was applied. Differences were considered significant
at p<0.05.
Results
TNF-α enhances the migration of
hepatocellular carcinoma cells
MMPs are involved in the processes of angiogenesis,
tumor progression and metastasis (10). In this study, the effect of TNF-α
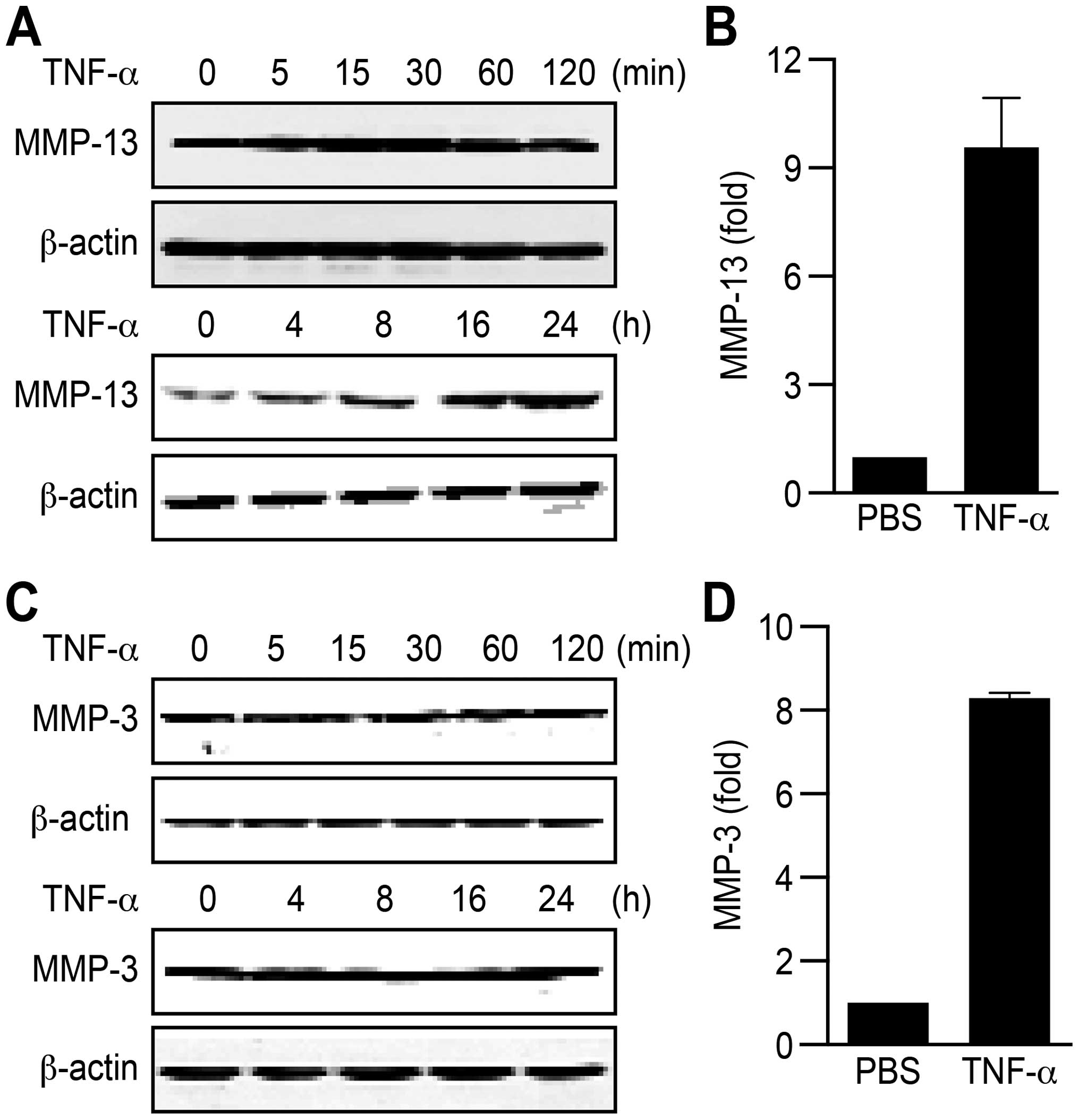
on MMP-13 and MMP-3 expression was determined. Compared with
controls, TNF-α efficiently upregulated MMP-13/MMP-3 expression at
both the protein (Fig. 1A and C)
and RNA levels (Fig. 1B and D).
Taken together, these findings indicate that pro-inflammatory
cytokines, such as TNF-α, play an important role in HCC
metastasis.
TNF-α activates the Erk1/2 and NF-κB
pathways
Although the MAPK and NF-κB pathways are involved in
TNF-α-augmented cell migration in VSMCs and macrophages (23,24),
the exact mechanism of TNF-α-augmented cell migration remains
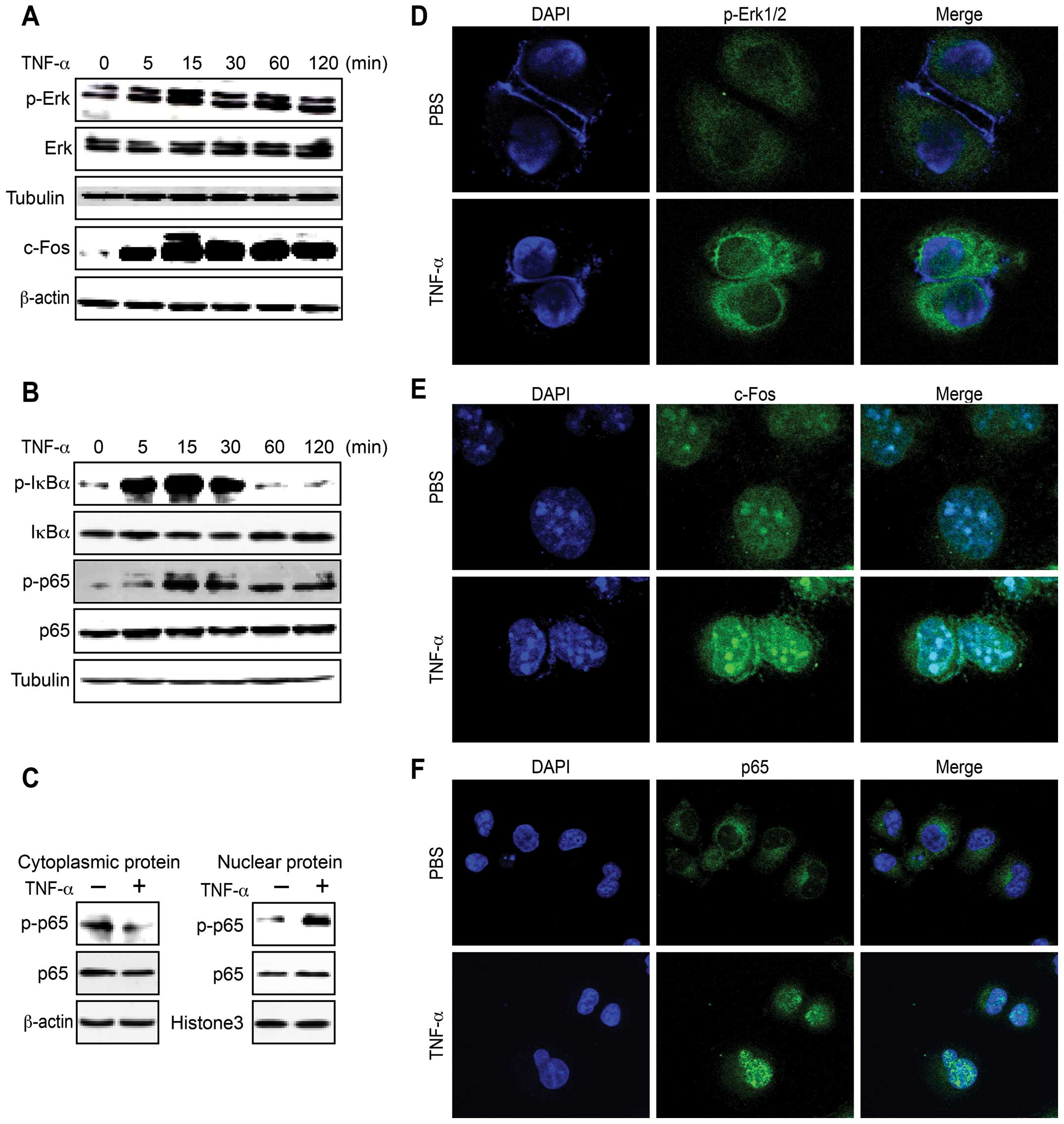
uncertain. Our results revealed that Erk1/2 phosphorylation and
c-Fos expression were clearly increased following TNF-α treatment
(Fig. 2A), as were the levels of
IκBα and p65 phosphorylation (Fig.
2B) and p65 translocation (Fig.
2C). The confocal microscopy results led to a similar
conclusion (Fig. 2D–F), and taken
together, these findings indicate that TNF-α activates the
Erk1/2-c-Fos and NF-κB pathways in HepG2 cells.
Inhibition of Erk1/2 and NF-κB activation
abrogates the effects of TNF-α on cell migration
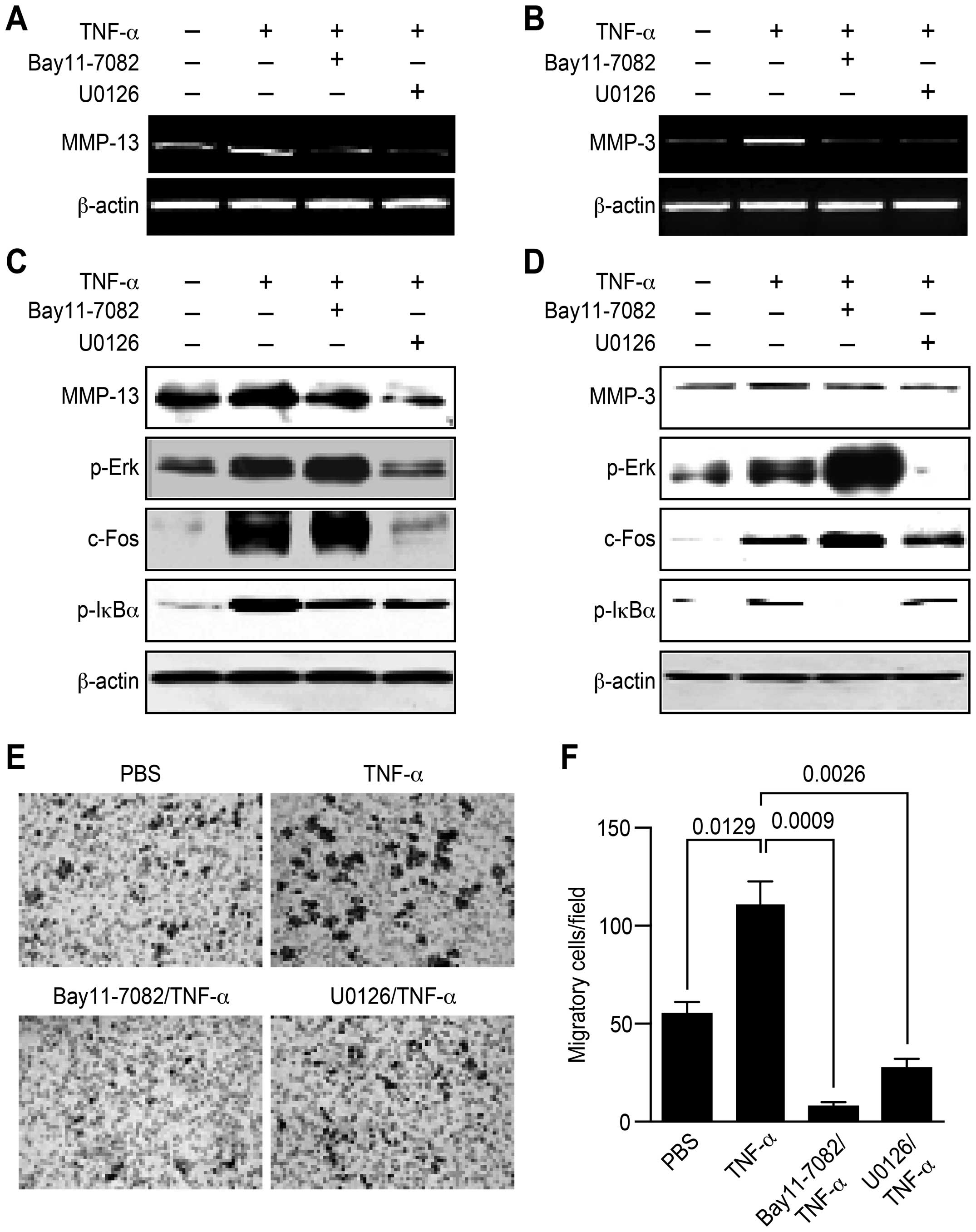
To understand the role of Erk1/2 and NF-κB
activation in cell migration, inhibitors were used to inhibit
related kinase activities. These inhibitors were invariably able to
abolish Erk1/2 or NF-κB kinase activity. The inhibition of Erk1/2
or NF-κB activity clearly abrogated the effect of TNF-α on
MMP-13/MMP-3 expression at both the transcriptional (Fig. 3A and B) and translational (Fig. 3C and D) levels, indicating that
TNF-α augments MMP-13/MMP-3 expression by activating the Erk1/2 and
NF-κB pathways. Additionally, the inhibition of Erk1/2 or NF-κB
activity led to a 77.8 or 93.1% rate of inhibition of cell
migration, respectively (Fig. 3E,
p<0.05, p<0.01, p<0.001, one-way ANOVA with the
Newman-Keuls post-test). The observation that inhibition of Erk1/2
or NF-κB activity abolished the effect of TNF-α on MMP-13/MMP-3
expression and cell migration indicates that the Erk1/2 and NF-κB
pathways are involved in MMP-13/MMP-3-mediated cell migration in
HepG2 cells.
TIPE2 abolishes the effect of TNF-α on
cell migration by downregulating MMP-13/MMP-3 expression
HCC exhibits lower TIPE2 expression than ambient
tissues (18), and the finding
that TIPE2 inhibits F-actin depolymerization (18) and downregulates MMP-9 expression
(17) therefore raises the
question of whether TIPE2 plays an important role in HCC
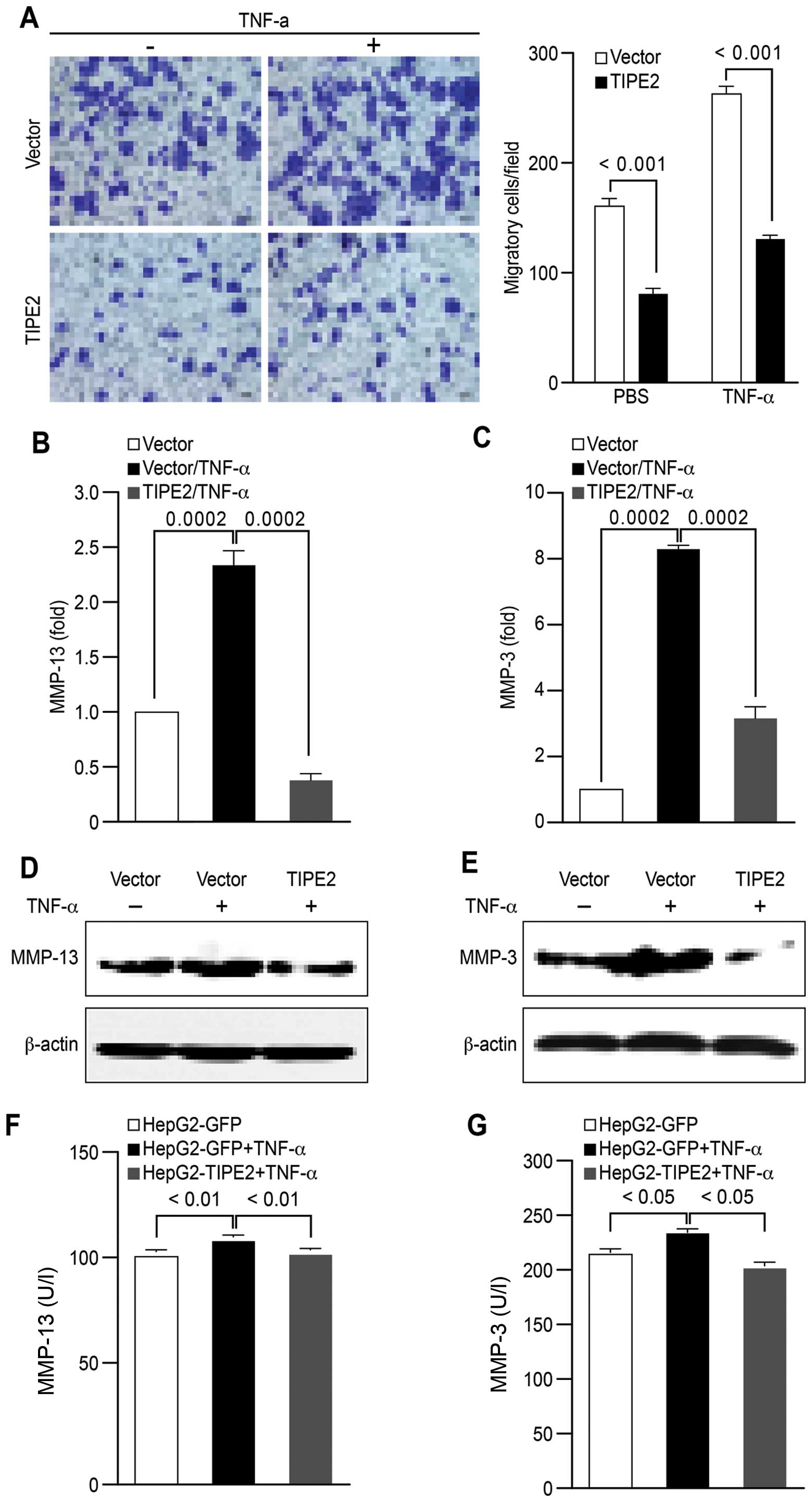
metastasis. Although TNF-α clearly augmented cell migration, TIPE2
overexpression led to approximately 50% inhibition of cell
migration (Fig. 4A). Moreover,
TIPE2 efficiently abrogated the effect of TNF-α on the expression
of MMP-13 and MMP-3 at both the RNA (Fig. 4B and C) and protein (Fig. 4D and E) levels. Additionally, TIPE2
significantly inhibited MMP-13 and MMP-3 activities (Fig. 4F and G) (p<0.01, p<0.001,
one-way ANOVA with the Newman-Keulspost test). The data presented
in this study indicate that TIPE2 inhibits the effect of TNF-α on
HCC metastasis by inhibiting MMP expression.
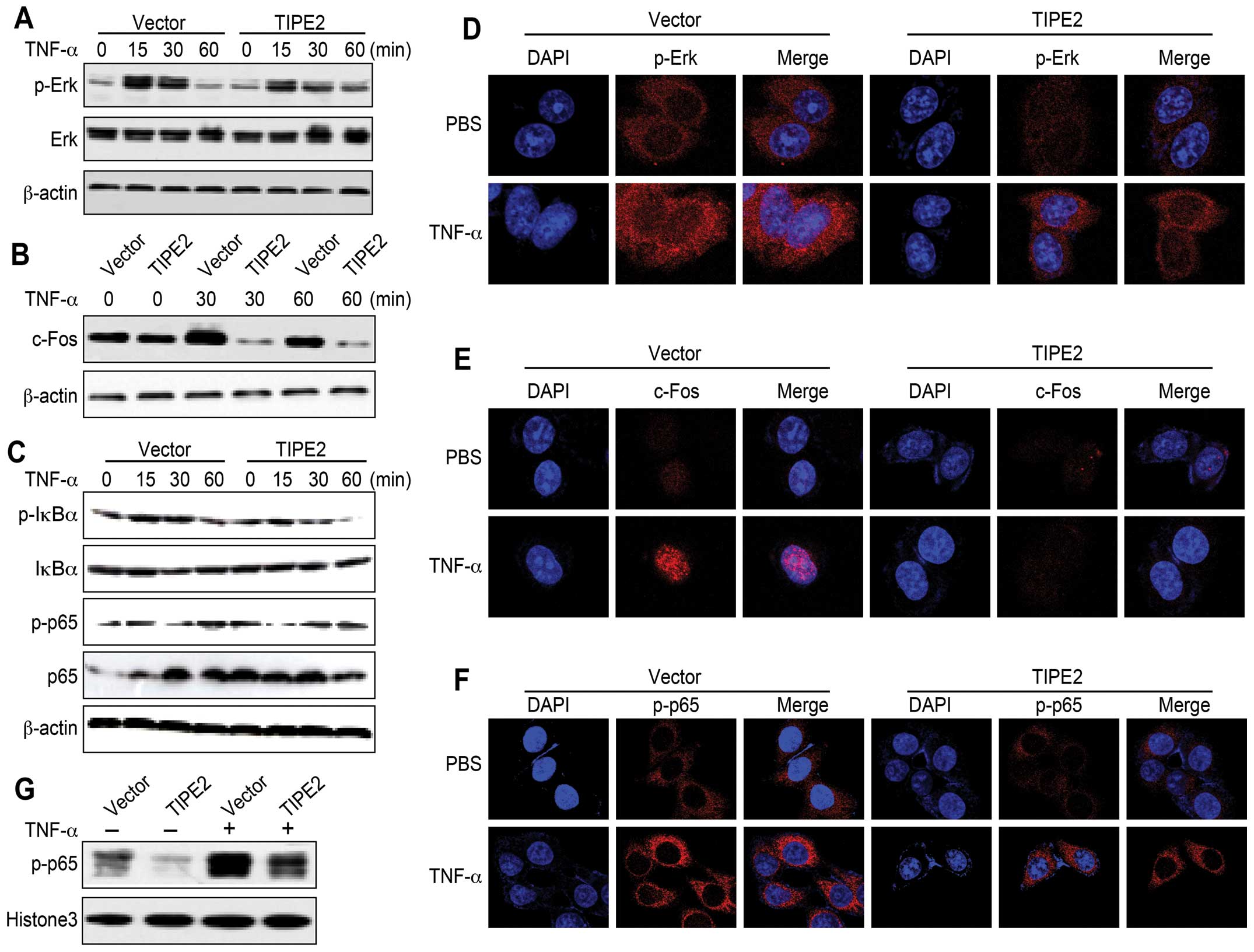
TIPE2 inhibits the effect of TNF-α on
Erk1/2 and NF-κB activation
The Erk/AP-1 and NF-κB pathways play important roles
in TNF-α-induced MMP-13/MMP-3 expression and cell migration
(23,24). Hence, we examined the effect of
TIPE2 on TNF-α-augmented Erk1/2-NF-κB activation. As shown in
Fig. 5, TIPE2 efficiently
inhibited the effect of TNF-α on Erk1/2 (Fig. 5A), c-Fos expression (Fig. 5B) and IκBα/p65 phosphorylation
(Fig. 5C). Confocal microcopy
demonstrated similar results (Fig.
5D–F). As phosphorylated p65 translocation was impeded by TIPE2
(Fig. 5G), the data presented in
this study indicate that TIPE2 abolishes the effect of TNF-α on
Erk1/2-NF-κB activation in HepG2 cells.
The primary components of the HCC
microenvironment express TNF-α following LPS treatment
Kupffer cells and pit cells (hepatic natural killer
cells), which are primary immune cells of the HCC microenvironment,
contribute to elevated serum TNF-α levels (25). To elucidate the source of TNF-α,
the primary components of the tumor microenvironment were treated
with LPS, and the expression of TNF-α was analyzed. Consistent with
another report (25), LPS
treatment increased TNF-α expression in macrophages (Fig. 6A and B), fibroblasts (Fig. 6C and D) and human peripheral blood
lymphocyte (PBL) (Fig. 6E and F)
at both the transcriptional (Fig. 6A,
C and E) and translational (Fig.
6B, D and F) levels. To our surprise, the expression of TNF-α
in HepG2 (Fig. 6G and H) was
notably upregulated following LPS treatment, indicating that liver
cancer itself secretes TNF-α in an autocrine manner and promotes
metastasis under LPS treatment conditions.
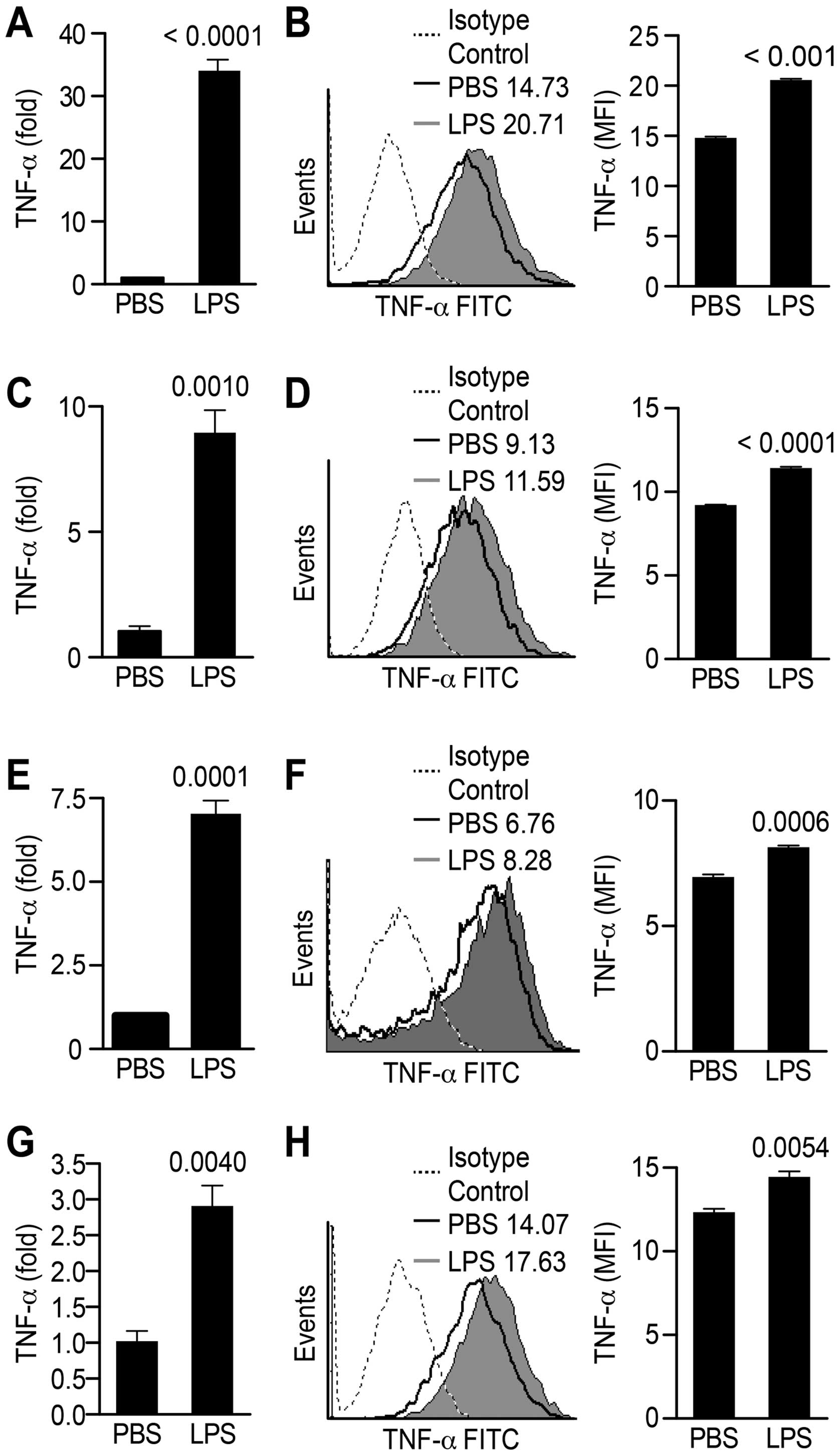 | Figure 6The components of the tumor
microenvironment secrete TNF-α following LPS treatment. The primary
components of the HCC microenvironment, such as Raw 264.7 (A and
B), NIH-3T3 (C and D) and HepG2 (G and H) cells and peripheral
blood lymphocytes (PBLs) (E and F) were exposed to LPS, and TNF-α
expression was determined via real-time PCR (A, C, E and G), and
flow cytometry analyses (B, D, F and H). For the flow cytometry
analysis, the numbers in the histogram indicate the geometric mean
fluorescence (MFI) of the tested samples. Positive percentages of
TNF-α expression and the MFI are shown. The data are presented as
the mean ± SEM, p<0.05, p<0.01, Student’s t-test.
Representative results from 1 of 3 independent experiments are
shown. β-actin was used as a loading control. LPS,
lipopolysaccharide. |
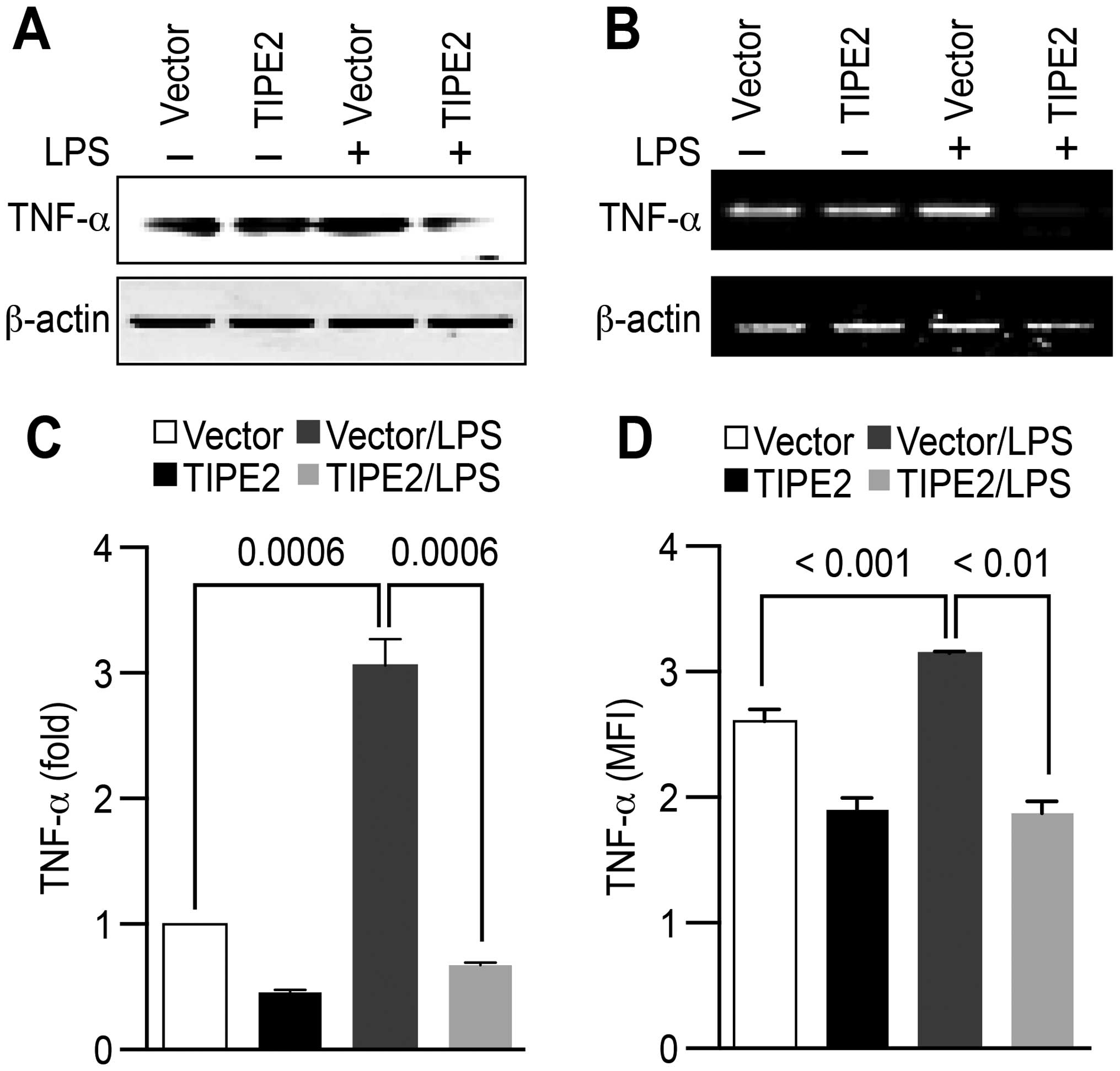
TIPE2 suppresses the effect of LPS on
TNF-α expression in HCC cells
TIPE2, which is a negative regulator of immune
homeostasis, can inhibit the release of the pro-inflammatory
cytokine TNF-α (16). The finding
that TIPE2 is downregulated during HCC carcinogenesis (18) raises the question of whether TIPE2
could be a potential candidate for managing TNF-α-promoted HCC
metastasis. Although LPS treatment significantly augmented TNF-α
expression, TIPE2 successfully abrogated the effect of LPS on TNF-α
expression in HepG2 (Fig. 7).
These results indicate that TIPE2 might be a potential therapeutic
target for pro-inflammatory cytokine-associated HCC metastasis.
Discussion
HCC patients and metastatic liver carcinomas display
elevated serum TNF-α levels (5,26),
indicating that metastasis is influenced by the tumor
microenvironment (27). TNF-α,
which is a key regulator of the tumor microenvironment, has been
reported to promote malignant invasion and metastasis in bladder
cancer and breast cancer (14,28).
The present study demonstrated that TNF-α upregulates MMP-13/MMP-3
expression and augments cell migration in HepG2 cells (Figs. 1 and 2). Moreover, inhibition of Erk1/2-NF-κB
activities abrogates the effect of TNF-α on cell migration and MMP
expression, indicating that TNF-α might be a candidate therapeutic
target in inflammation-associated metastasis. As tumor tissues are
composed of tumor cells, fibroblasts, and immune cells, the finding
that Kupffer cells synthesize and secrete TNF-α cannot exclude the
possibility that other components of liver cancer tissues might
contribute to the elevation of serum TNF-α. In this study, all of
the components of cancer tissues, such as cancer cells,
fibroblasts, PBLs and macrophages, were shown to secrete TNF-α
under LPS treatment conditions, indicating that TNF-α promotes
metastasis in both a paracrine and autocrine manner in HCC.
Although TNF-α was found to contribute to the
resistance of pathogen infection and inhibit carcinogenesis
(29), a role of TNF-α in
metastasis in bladder cancer and breast cancer has also been
documented (14,28). These contrasting findings might be
due to the concentration of TNF-α used in these studies. While a
higher concentration of TNF-α (25 ng/ml) induces mouse blastocyst
death (30), a lower concentration
of TNF-α (1–10 ng/ml) clearly promotes prostate cancer metastasis
and affects the survival of chronic lymphocytic leukemia patients
(31,32).
MMPs, which are ECM degradation enzymes, play
important roles in tumor migration and metastasis (10). MMP-3 has been found to directly
degrade the extracellular matrix, to activate other MMPs, such as
MMP-1, MMP-7 and MMP-9 (12,33),
and to be related to the prognosis of HCV-associated liver cancer
(12), indicating that MMP-3 is a
primary MMP involved in HCC metastasis. Moreover, the ability of
MMP-13, which can be activated by MMP-3, to directly degrade the
extracellular matrix and to indirectly activate other MMP members,
such as MMP-2/MMP-9, has also been documented, indicating that
MMP-13 is another MMP involved in extracellular matrix remodeling
(34). Hence, in this study, the
effect of TNF-α on MMP-3 and MMP-13 expression was explored.
TIPE2 has been found to reduce F-actin
polymerization and to inhibit cell migration (18). As actin polymerization is essential
for the nuclear translocation of RhoA and protein trafficking
(35,36), the effects of TIPE2 on MMP
expression and cell migration might be due to the modulation of
F-actin polymerization and epithelial-mesenchymal transition by
TIPE2. In this study, we demonstrate that TIPE2 inhibits the effect
of TNF-α on Erk1/2-NF-κB activation. TNF-α-induced protein 3
(TNFAIP3, A20), which also belongs to the TNF-α-induced protein
family, shows an effect similar to that of TIPE2 on NF-κB
activation (37). The N-terminus
of A20 encodes a deubiquitinating (DUB) domain that mediates the
deubiquitination of K63-polyubiquitinated NF-κB signaling proteins,
such as TRAF6 and RIP1, whereas the C-terminus of A20 encodes seven
zinc-finger (ZF) motifs and confers E3 ubiquitin ligase activity
(38). More recently, the binding
of A20 to unanchored K63-linked polyubiquitin chains and to NEMO
was shown to block the upstream kinase of IKK, TAK1 (39), indicating that the effect of A20 on
NF-κB activation is a non-catalytic mechanism-dependent effect.
Because TIPE2 and A20, which belong to the TNFAIP family,
negatively regulate immune function and NF-κB activation,
determining whether TIPE2 encodes deubiquitinase enzyme domains or
antagonizes the ubiquitination of the NEMO-IκB kinase complex (IKK)
regulatory subunit by interacting with NEMO requires further
exploration.
Taken together, our experiments reveal that TNF-α
increases MMP-13/MMP-3 expression and augments cell migration by
activating the Erk1/2 and NF-κB pathways. The observed inhibition
of the effects of TNF-α on MMP expression, cell migration and
Erk1/2-NF-κB activation by TIPE2 indicates that TIPE2 and TNF-α
might be potential targets for managing HCC metastasis.
Acknowledgements
This study was supported by grants from the State
Key Laboratory of Oncogenes and Related Genes (no. 90-14-05) and by
grants from the National Natural Science Foundation of China (no.
81273203).
References
|
1
|
Arzumanyan A, Reis HM and Feitelson MA:
Pathogenic mechanisms in HBV- and HCV-associated hepatocellular
carcinoma. Nat Rev Cancer. 13:123–135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim DY and Han KH: Epidemiology and
surveillance of hepatocellular carcinoma. Liver Cancer. 1:2–14.
2012. View Article : Google Scholar
|
|
3
|
Shirabe K, Mano Y, Muto J, et al: Role of
tumor-associated macrophages in the progression of hepatocellular
carcinoma. Surg Today. 42:1–7. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang YY, Lo GH, Lai KH, et al: Increased
serum concentrations of tumor necrosis factor-α are associated with
disease progression and malnutrition in hepatocellular carcinoma. J
Chin Med Assoc. 66:592–597. 2003.
|
|
5
|
Ding J, Huang S, Wang Y, et al:
Genome-wide screening reveals that miR-195 targets the TNF-α/NF-κB
pathway by down-regulating IκB kinase alpha and TAB3 in
hepatocellular carcinoma. Hepatology. 58:654–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Teixeira AC, Mendes CT Jr, Marano LA, et
al: Alleles and genotypes of polymorphisms of IL-18, TNF-α and
IFN-γ are associated with a higher risk and severity of
hepatocellular carcinoma (HCC) in Brazil. Hum Immunol.
74:1024–1029. 2013. View Article : Google Scholar
|
|
7
|
Rucci N, Sanità P and Angelucci A: Roles
of metalloproteases in metastatic niche. Curr Mol Med. 11:609–622.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wilken R, Veena MS, Wang MB, et al: A
review of anti-cancer properties and therapeutic activity in head
and neck squamous cell carcinoma. Mol Cancer. 10:122011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stokes A, Joutsa J, Ala-Aho R, et al:
Expression profiles and clinical correlations of degradome
components in the tumor microenvironment of head and neck squamous
cell carcinoma. Clin Cancer Res. 16:2022–2035. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu D, Guo H, Li Y, et al: Association
between polymorphisms in the promoter regions of matrix
metalloproteinases (MMPs) and risk of cancer metastasis: a
meta-analysis. PLoS One. 7:e312512012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang M, Dai C, Zhu H, et al: Cyclophilin
A promotes human hepatocellular carcinoma cell metastasis via
regulation of MMP3 and MMP9. Mol Cell Biochem. 357:387–395. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Okamoto K, Ishida C, Ikebuchi Y, et al:
The genotypes of IL-1 beta and MMP-3 are associated with the
prognosis of HCV-related hepatocellular carcinoma. Intern Med.
49:887–895. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nannuru KC, Futakuchi M, Varney ML, et al:
Matrix metalloproteinase (MMP)-13 regulates mammary tumor-induced
osteolysis by activating MMP9 and transforming growth factor-beta
signaling at the tumor-bone interface. Cancer Res. 70:3494–3504.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pivetta E, Scapolan M, Pecolo M, et al:
MMP-13 stimulates osteoclast differentiation and activation in
tumour breast bone metastases. Breast Cancer Res. 13:R1052011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kudo Y, Iizuka S, Yoshida M, et al: Matrix
metalloproteinase-13 (MMP-13) directly and indirectly promotes
tumor angiogenesis. J Biol Chem. 287:38716–38728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun H, Gong S, Carmody RJ, et al: TIPE2, a
negative regulator of innate and adaptive immunity that maintains
immune homeostasis. Cell. 133:415–426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang G, Zhang W, Lou Y, et al: TIPE2
deficiency accelerates neointima formation by downregulating smooth
muscle cell differentiation. Cell Cycle. 12:501–510. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gus-Brautbar Y, Johnson D, Zhang L, et al:
The anti-inflammatory TIPE2 is an inhibitor of the oncogenic Ras.
Mol Cell. 45:610–618. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ke SZ, Ni XY, Zhang YH, et al:
Camptothecin and cisplatin upregulate ABCG2 and MRP2 expression by
activating the ATM/NF-κB pathway in lung cancer cells. Int J Oncol.
42:1289–1296. 2013.PubMed/NCBI
|
|
20
|
Jin HJ, Sui HX, Wang YN, et al: Nicotine
up-regulated 4-1BBL expression by activating Mek-PI3K pathway
augments the efficacy of bone marrow-derived dendritic cell
vaccination. J Clin Immunol. 33:246–254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jin HJ, Li HT, Sui HX, et al: Nicotine
stimulated bone marrow-derived dendritic cells could augment HBV
specific CTL priming by activating PI3K-Akt pathway. Immunol Lett.
146:40–49. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao FG, Li HT, Li ZJ, et al: Nicotine
stimulated dendritic cells could achieve anti-tumor effects in
mouse lung and liver cancer. J Clin Immunol. 31:80–88. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Z, Xu X, Bai L, et al: Epidermal growth
factor receptor-mediated tissue transglutaminase overexpression
couples acquired tumor necrosis factor-related apoptosis-inducing
ligand resistance and migration through c-FLIP and MMP-9 proteins
in lung cancer cells. J BiolChem. 286:21164–21172. 2011.
|
|
24
|
Lin X, Zheng W, Liu J, et al: Oxidative
stress in malignant melanoma enhances tumor necrosis factor-α
secretion of tumor-associated macrophages that promote cancer cell
invasion. Antioxid Redox Signal. 19:1337–1355. 2013. View Article : Google Scholar
|
|
25
|
Sakai T, Liu L, Teng X, et al:
Inflammatory disease and cancer with a decrease in Kupffer cell
numbers in Nucling-knockout mice. Int J Cancer. 126:1079–1094.
2010.PubMed/NCBI
|
|
26
|
Kim MJ, Jang JW, Oh BS, et al: Change in
inflammatory cytokine profiles after transarterial chemotherapy in
patients with hepatocellular carcinoma. Cytokine. 64:516–522. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Budhu A, Forgues M, Ye QH, et al:
Prediction of venous metastases, recurrence, and prognosis in
hepatocellular carcinoma based on a unique immune response
signature of the liver microenvironment. Cancer Cell. 10:99–111.
2006. View Article : Google Scholar
|
|
28
|
Lee EJ, Kim WJ and Moon SK: Cordycepin
suppresses TNF-alpha-induced invasion, migration and matrix
metalloproteinase-9 expression in human bladder cancer cells.
Phytother Res. 24:1755–1761. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kroesen S, Widmer AF, Tyndall A, et al:
Serious bacterial infections in patients with rheumatoid arthritis
under anti-TNF-α therapy. Rheumatology. 42:617–621. 2003.
View Article : Google Scholar
|
|
30
|
Fabian D, Juhás S, Il’ková G, et al: Dose-
and time-dependent effects of TNFα and actinomycin D on cell death
incidence and embryo growth in mouse blastocysts. Zygote.
15:241–249. 2007. View Article : Google Scholar
|
|
31
|
Radhakrishnan P, Chachadi V, Lin MF, et
al: TNFα enhances the motility and invasiveness of prostatic cancer
cells by stimulating the expression of selective glycosyl- and
sulfotransferase genes involved in the synthesis of selectin
ligands. Biochem Biophys Res Commun. 409:436–441. 2011. View Article : Google Scholar
|
|
32
|
Ferrajoli A, Keating MJ, Manshouri T, et
al: The clinical significance of tumor necrosis factor-α plasma
level in patients having chronic lymphocytic leukemia. Blood.
100:1215–1219. 2002.
|
|
33
|
Ye S, Eriksson P, Hamsten A, et al:
Progression of coronary atherosclerosis is associated with a common
genetic variant of the human stromelysin-1 promoter which results
in reduced gene expression. J Biol Chem. 271:13055–13060. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Altadill A, Rodríguez M, González LO, et
al: Liver expression of matrix metalloproteases and their
inhibitors in hepatocellular carcinoma. Dig Liver Dis. 41:740–748.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Smith EC, Teixeira AM, Chen RC, et al:
Induction of mega-karyocyte differentiation drives nuclear
accumulation and transcriptional function of MKL1 via actin
polymerization and RhoA activation. Blood. 121:1094–1101. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hao YH, Doyle JM, Ramanathan S, et al:
Regulation of WASH-dependent actin polymerization and protein
trafficking by ubiquitination. Cell. 152:1051–1064. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang S, Wen F, Wiley GB, et al: An
enhancer element harboring variants associated with systemic lupus
erythematosus engages the TNFAIP3 promoter to influence A20
expression. PLoS Genet. 9:e10037502013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wertz IE, O’Rourke KM, Zhou H, et al:
De-ubiquitination and ubiquitin ligase domains of A20 downregulate
NF-kappaB signalling. Nature. 430:694–699. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Skaug B, Chen J, Du F, et al: Direct,
noncatalytic mechanism of IKK inhibition by A20. Mol Cell.
44:559–571. 2011. View Article : Google Scholar : PubMed/NCBI
|