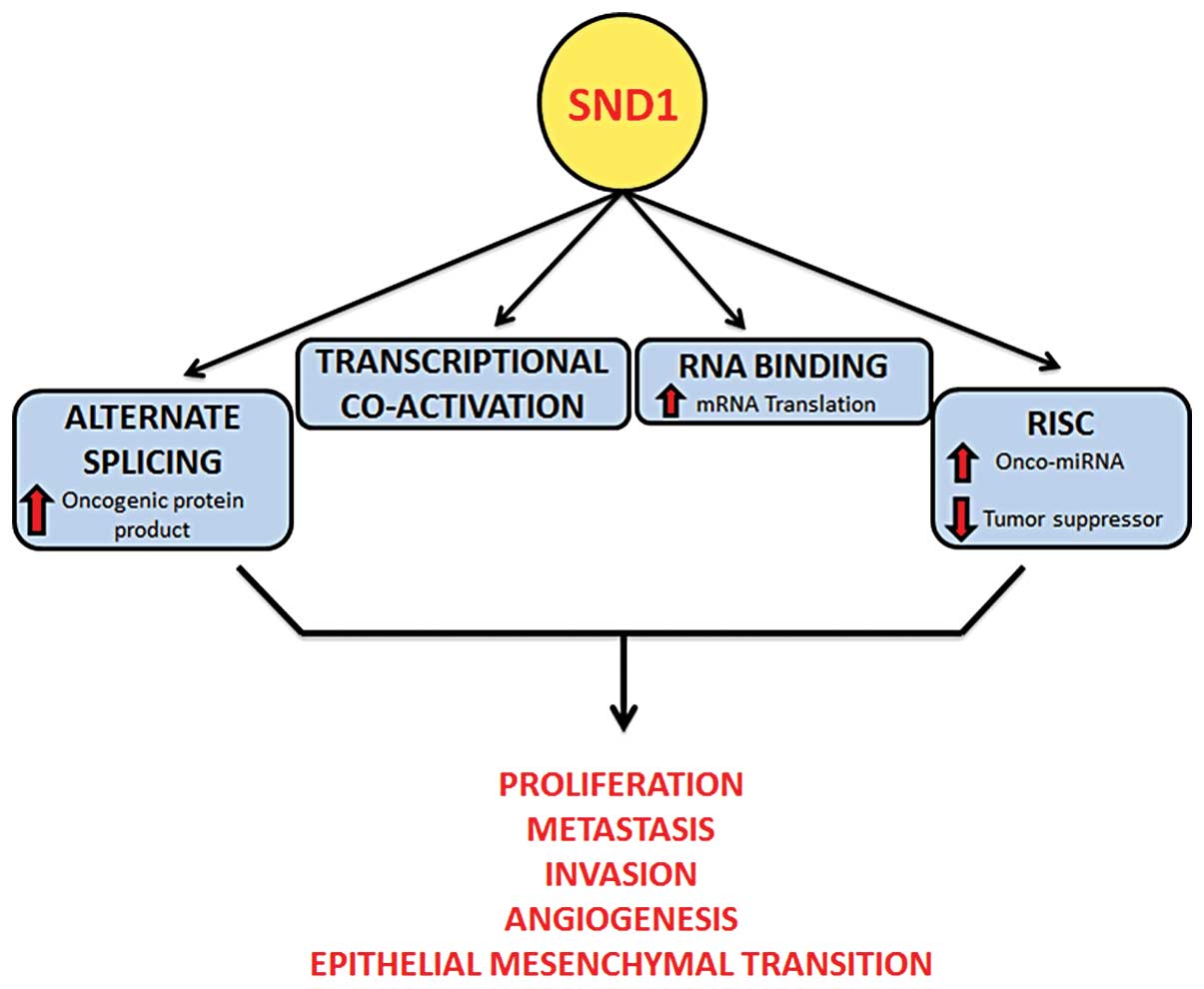
1. Introduction
Cancer is a highly aggressive disease characterized
by interplay of mutations that cause cellular transformation and
abnormal cell growth. Over the years, state-of-the-art studies have
provided significant insights into the pathogenesis of this highly
complex disease. Cancer is now well-defined by six hallmarks:
sustained proliferative signaling, evasion of growth suppression,
invasion and metastasis, replicative immortality, angiogenesis and
resistance to apoptosis (1).
Changes in expression patterns of any gene controlling these
pathways initiate tumorigenesis that has the potential of
developing malignancy. With increasing number of mutations being
identified each day, that play a role in cancer development,
molecular understanding of this disease gets more complicated.
Further adding to the complexity is the effect of environmental
factors in disease pathogenesis at each stage. Mutations arising in
cells are mainly governed by genetic predisposition to changes in
DNA structure, stability and expression, epigenetic changes and
environmental factors (2).
Epigenetic variations broadly include methylation pattern, microRNA
expression disorders and chromatin organization (2). Diversity in cancer is not just
restricted to the factors leading to cancer development, but is
also affected by the tumor microenvironment, broadly classified as
intertumor heterogeneity and intra-tumor heterogeneity (2). With the advent of new technologies,
the molecular changes, which are responsible for such alterations,
are being identified and a subset of these can serve as potential
therapeutic targets.
The staphylococcal nuclease and tudor domain
containing 1 (SND1) is involved in transcriptional activation, RNA
splicing, editing and stability, and RNAi function (3–10).
These processes are relevant for regulation of gene expression.
SND1 is predicted to manifest a dynamic role modulating multiple
molecular networks that control gene expression. Structural
analysis has confirmed that the SND1 protein functions include
nucleic acid interaction along with protein-protein interactions
(11). Several studies have
illustrated the significant association of SND1 with different
types of cancers, including liver, breast, prostate and colorectal
cancers and malignant glioma (12–20).
It is overexpressed in these cancers and known to promote
manifestation of aforementioned hallmarks of carcinogenesis. The
present review provides a comprehensive description of the
functional aspects of SND1 that are relevant to cancer development
and progression.
2. SND1 protein structure
The human SND1 gene was assigned to chromosome
7q31.3 band location employing restriction fragment and
fluorescence in situ hybridization (FISH) analyses (21). Genomic gain in 7q, especially 7q31,
has been demonstrated in prostate, renal and colorectal carcinoma
(22–24). SND1 is overexpressed in prostate
and colorectal carcinoma (18,19)
and genomic amplification might be an underlying mechanism for this
overexpression.
Human SND1 is a 910-a.a. containing protein with
highly conserved domains found even in Caenorhabditis
elegans. SND1 comprises tandem repeats of four nuclease (SN)
domains and a fifth domain containing fusion of Tudor and partial
nuclease domains (TSN) (11)
(Fig. 1). The nuclease domains
share 20–30% sequence homology (3). These domains have been well
characterized and found to be structurally related to
staphylococcal nuclease (SN) domains (3). These are thermonucleases that
hydrolyze DNA and RNA in a calcium-dependent manner (3). However, hydrophobic cluster analysis
shows that the SN domains of SND1 lack the specific amino acid
residues involved in calcium-dependent catalytic activity (3). SN domains are included in the
oligonucleotide/oligosaccharide binding (OB) fold superfamily,
which comprises a large number of proteins, involved in nucleic
acid binding (3). OB fold proteins
are critical for DNA replication, DNA recombination, DNA repair,
telomeric maintenance and cold shock response (25). OB fold domains range from 70 to 150
a.a. in length and consist of variable loops between conserved
secondary domains (25). Other
than staphylococcal nucleases, OB fold is also observed in
bacterial enterotoxins, nucleic acid binding proteins and inorganic
pyrophosphates (25). Many of the
proteins in OB fold family, lack catalytic activity but carry out
several other functions, such as transcriptional activation or
repression, chromatin modification and DNA repair (3,25).
Tudor domains are highly conserved domains across eukaryotic
species and studied in great detail employing Drosophila as
a model system (26). Tudor domain
containing proteins are involved in DNA interactions, specifically
in epigenetic regulation, gene expression as well as snRNP, miRNA
and siRNA biogenesis (26). The
presence of these versatile SN and Tudor domains confer SND1 with
its diverse multifunctional properties (Fig. 2).
3. Regulation of SND1 expression
The isolated 5′-flanking region of human SND1 gene
spans >3.8 kb, including transcription start site and
translation initiation codon (27). Bioinformatics analysis revealed
presence of putative binding motifs for NF-κB, Sp1 and NF-Y
transcription factors (27). The
core promoter region does not contain TATA box, but is rich in CpG
islands to form pre-initiation transcription complexes. The lack of
a TATA box, an initiator sequence for transcription, is
complimented by the presence of a CCAAT box in the reverse
orientation, i.e., ATTGG at position −61 and −28 (27). The CCAAT box and a GC box, at −48,
act as positive regulatory elements for SND1 expression (27). Inverted CCAAT sequences are often
binding targets for the NF-Y transcription factor. Luciferase
assays with promoter deletion mutants show that −274 to −112,
containing potential binding sites for NF-κB and Sp1, are crucial
for SND1 expression (27).
Interaction of NF-κB, Sp1 and NF-Y with the SND1 promoter was also
detected by chromatin immnunoprecipitation (27). Mutation in GC box or CCAAT box
reduced SND1 expression by 55–75%. As expected, TNFα, an activator
of NF-κB, induced SND1 transcription, thereby suggesting the
functional role of NF-κB in SND1 transcription regulation (27).
Promoter regions of the human, mouse and rat SND1
gene show approximately 80–85% sequence homology (27). Similar findings have been observed
in promoter analysis of rat homologue of SND1, p102 (28,29).
Putative binding sites for CCAAT/enhancer binding protein (C/EBP),
STATs and upstream stimulatory factor (USF) have been identified
(28). NF-Y binds at the CCAAT box
(−370, −366), whereas the GC rich regions were identified as
putative Sp1 binding sites (29).
DNA elements that matched the consensus binding sequences for liver
specific transcription factors, such as HNF-4, were also identified
(28), suggesting that SND1 might
be differentially expressed in a tissue-specific manner.
Fashe et al investigated cellular
localization and tissue specific expression of SND1 in mice. With
an exception of muscle tissues, SND1 is ubiquitously expressed in
mice and the expression patterns are mostly consistent to that
reported in humans (30). SND1 is
noticeably upregulated in active secretory organs including
pancreas, liver and mammary glands (30). Studies have reported upregulated
levels of SND1 in lipid droplets of milk secreting mammary
epithelial cells (31). SND1
levels were higher in exocrine pancreatic cells as compared to
endocrine cells (30). SND1
expression is lower in hepatocytes, however, it is overexpressed in
sinusoid endothelial cells (30).
Actively proliferating cells such as crypts of Lieberkuhn and basal
keratinocytes of skin and hair follicles show high SND1 protein
levels compared to more differentiated or terminally differentiated
cells in the same tissue. SND1 protein levels are much higher in
spermatogonial cells than in spermatocytes and Sertoli cells. No
protein is detected in terminally differentiated spermatids and
mature spermatozoa. In ovary, SND1 levels are high in follicular
cells compared to that in stromal cells. There is no SND1 protein
in oocytes. SND1 protein levels are moderately expressed in human
brain tissue, in neuronal as well as glial cells. In mouse kidney,
SND1 is comparatively overexpressed in endothelial cells of
Bowman’s capsule than podocytes and mesangial cells. This protein
is also expressed in bronchiolar epithelium, alveolar cells and
pneumocytes of lung. Within gastrointestinal system, SND1 is
expressed in ileum, deuodenum and colon. Lower SND1 levels are
observed in more differentiated cells like villi and Paneth cells
in the ileum and absorptive cells of the colon. Overall, SND1 is
overexpressed in rapidly proliferating or precursor cells and
downregulated in terminally differentiated cells (30). SND1 is upregulated in T-cells and
co-localizes with CD3, as observed in lymphoid organs (30). SND1 expression is not evident in
macrophages, in lymphoid organs or in tissue residing macrophages
like Kuppfer’s cells in liver and alveolar macrophages in lungs.
Further, SND1 levels in the red pulp of spleen are found to be
higher than that in white pulp. Differential expression of SND1 in
T lymphocytes and macropahges suggests a potential role of SND1 in
regulating immunity.
4. Functions of SND1
Regulation of transcription
SND1, also called TudorSN or p100, was first
identified as a transcriptional co-activator in an attempt to
identify proteins interacting with Epstein-Barr nuclear antigen
(32). EBNA2 specifically
activates transcription of genes that mediate B lymphocyte
transformation (32). Along with
known interacting factors such as TFIIB, TFIIH and TAF40, SND1
specifically interacts with EBNA2 acidic domain (32). This interaction is mediated by
SND1-TFIIE interaction such that SND1 acts as an adapter protein
between EBNA2 and the transcriptional machinery (32). Pim kinase, found to be upregulated
during Epstein-Barr virus infection, also interacts with SND1 and
mediate cellular transformation by cooperating to enhance c-Myb
activity (9). It was illustrated
that Pim-1 phosphorylates SND1 and forms a stable complex, leading
to an induction in c-Myb activity (9). Ectopic expression of Ras and Pim-1
also induce c-Myb responsive genes, since c-Myb is a downstream
target in this signaling cascade (9). This research study thus revealed that
SND1 is a vital link in Ras- and Pim-1-mediated induction of c-Myb
(9). Lack of SND1 or dominant
negative alleles of SND1 failed to cause an induction in c-Myb
activity (9). c-Myb is linked to
proliferation and differentiation, and also known to mediate
cellular transformation. Hence, a role of SND1 in regulating c-Myb
expression via Ras or Pim-1 is significant in understanding its
functional impact in oncogenesis.
Another important class of transcription factors
that are known to interact and cooperate with SND1 is the signal
transducers and activator of transcription, i.e., STATs (4,8,33).
STAT proteins are sequestered to Janus kinases (JAK), which upon
stimulation by cytokines like IFN-γ, undergo phosphorylation
(34). Phosphorylation of STAT
proteins and subsequent dimerization leads to nuclear localization
where it activates transcription of target genes (34). Constitutive activation of the
JAK-STAT pathway has been implicated in many cancers (34). There are seven STAT proteins, which
share structural and functional homology. Of these, SND1 is known
to act as a transcriptional co-activator for STAT5 and STAT6. SND1
co-immunoprecipitates with STAT5, confirming the protein-protein
interaction, but does not affect the phosphorylation status of
STAT5 (8). Interaction with STAT5
is mediated via the SN domains as well as Tudor domains of SND1,
whereas STAT6-SND1 interactions involve only SN domains. SND1 acts
as an adapter molecule, allowing functional bridging between CREB
binding proteins (CBP) and STAT6 (33). It interacts with the D3-D4 domains
of CBP and STAT6 transactivation domain (TAD). Histone acetylase
activity of CBP is stimulated by SND1, thereby allowing
transcriptional activation (33).
A recent study in a breast cancer model demonstrated
that SND1 significantly interacts with the promoter regions of
several genes in the TGFβ signaling pathway, including Smad 1–4 and
TGFβ (35). We have shown that
SND1 upregulation also correlates with TGFβ signaling in HCC, as
described later in this review (16).
Post-transcriptional regulation of gene
expression
Regulation of RNA-induced silencing
complex (RISC) activity
Post-transcriptional regulation of gene expression
can be mediated by several mechanisms including nucleocytoplasmic
localization, mRNA stability, mRNA processing and translation. SND1
functions as a nuclease in RNA-induced silencing complex (RISC)
that plays a significant role in modulation of gene expression at a
post-transcriptional level (6).
Our studies have shown that astrocyte elevated gene-1 (AEG1), an
important oncogene (36,37), in association with SND1 and other
proteins, forms a stable RISC complex (12). RISC incorporates one strand of a
small interfering RNA (siRNA) or microRNA (miRNA) and uses the
siRNA or miRNA as a template for recognizing complementary mRNA.
Argonaute proteins are activated in RISC when a complementary mRNA
is identified which then cleaves the mRNA. SND1 functions as a
nuclease in RISC along with the Argonaute proteins, while AEG-1
functions as a scaffold protein for proper assembly of this
complex. Both SND1 and AEG-1 are overexpressed in multiple cancers
and together they facilitate functions of oncogenic miRNAs
(onco-miRNA). Indeed RISC activity in cancer cells was found to be
higher than that in normal cells (12). The increased activity of
onco-miRNAs leads to increased suppression of their target tumor
suppressor genes. In HCC cells it was documented that
overexpression of AEG-1 or SND1 resulted in decreased expression of
several tumor suppressor genes that are targets of oncomiRNAs,
e.g., PTEN, target of miR-221 and miR-21; CDKN1C (p57), target of
miR-221; CDKN1A (p21), target of miR-106b; SPRY2, target of miR-21;
and TGFBR2, target of miR-93 (12). The reverse finding was observed
upon knockdown of SND1 or AEG-1 (12). These findings suggest that
increased RISC activity conferred by SND1 and AEG-1 might
contribute to the carcinogenic process (12).
In pancreatic cancer, synaptogamin-11 interacts with
RISC via SND1 binding (38). It is
hypothesized that this protein is the missing link between membrane
trafficking and miRNA-mediated gene regulation (38). Synthesis of mature mir17–92 cluster
is also inhibited by SND1 protein, thereby affecting several
downstream target genes (39).
Regulation of mRNA stability
SND1 interaction with mRNA transcript can be
independent of the RISC. Studies show that SND1 interacts with
3′UTR of angiotensin II type 1 receptor (AT1R), a G protein coupled
receptor mediating the action of angiotensin (7). Here, SND1 increases mRNA stability
and translational efficiency by increasing AT1R mRNA half-life,
resulting in elevated protein levels (7). A recent finding shows that SND1 and
AT1R mRNA colocalize in stress granules followed by oxidative
stress and SND1 is required for efficient protein-RNA aggregation
(40). These findings imply that
SND1 increases stability of specific mRNAs, crucial for cellular
stress response (40). Using HCC
cell lines, we have demonstrated that SND1-mediated increased
activity of AT1R activates the TGFβ signaling cascade thereby
promoting epithelial-mesenchymal transition (EMT) and an increase
in migration and invasion (16).
Studies have also established interaction of SND1 and Dengue virus
3′UTR, leading to increased viral replication (41). Further studies are required to
determine whether SND1 plays a similar role in promoting
replication of hepatitis B or C viruses (HBV or HCV), the most
common cause of HCC.
Regulation of mRNA splicing
Splicing is an important post-transcriptional event,
involved in excluding the non-coding intronic regions of mRNA
transcript and thereby allowing translation of exonic regions into
a functional polypeptide (42). A
large macromolecular complex, driven by several proteins is
required for this processing. Splicing is tightly regulated and
functionally coupled with transcription (42). Because differential regulation of
genes influencing cell growth and proliferation are critical for
carcinogenesis, splicing is speculated to play a vital role in
establishing pathogenesis (43).
The spliceosome complex is comprised of five major small
ribonucleoproteins - U1, U2, U4/U6 and U5 along with several small
non-snRNP (42,44). SND1 interacts with the U5 component
of the spliceosome and other non-snRNPs (44). Immunoprecipitation studies with
GST-TSN fusion protein (lacking SN domains) and GST-SN fusion
protein (lacking TSN domain) demonstrated that this interaction
occurs specifically via TSN domain (44). In vitro, exogenously added
SND1 accelerated the kinetics of spliceosome assembly, detected by
a ligated mRNA product, in a dose-dependent manner (44). However, no difference was observed
in the amount of splicing products with or without SND1 (44). SND1 improved the efficiency of
pre-spliceosomal complex assembly and accelerated the formation of
complex B and complex C (44).
Alternative splicing, observed in eukaryotes, allows
translation of multiple polypeptides from the same gene transcript
by selection of specific exons to be included in the processed mRNA
(43). Recent reports indicate
that deregulation of alternative splicing is associated with cancer
development and progression (43).
SND1 plays a role in this biological process. SND1 is identified as
an interacting partner for SAM68, a prooncogenic RNA binding
protein that is upregulated in prostate cancer and supports
cellular proliferation (45).
SAM68 is involved in alternative splicing of CD44, specifically
favoring inclusion of exon v5 of this gene (45). Prior studies indicate that
inclusion of variable exons (v5) in CD44 mRNA correlates with
cancer development (45). SND1
functions as a positive regulator of the alternative splicing of
CD44 via SAM68. Knocking down SND1 inhibits inclusion of upstream
variable exons (v4, v5 and v7), whereas downstream variable exons
(v8-v10) and constitutive exons are not affected (45). These findings suggest that SND1
co-ordinates transcriptional and post-transcriptional events to
regulate gene expression. Future studies focused on splicing
activity of SND1 might uncover insights into the molecular events
governing malignancy.
Regulation of RNA editing
Adenosine deaminase (ADAR) proteins function in RNA
A-to-I editing by deaminating Adenosine to Inosine, which is read
as guanosine on an mRNA transcript (46). Such processes regulate protein
translation by functionally changing the mRNA sequence (46). This post-transcriptional regulation
also plays a role in gene expression and miRNA processing (46). Interestingly, ADAR protein
expression is tightly regulated during embryogenesis, where it is
highest in the oocytes and zygote and diminished in the embryo
stages (10). A strong correlation
was observed between ADAR and SND1 levels during mouse
fertilization (10). While ADAR
marks hyper-edited transcripts, SND1 is responsible for degradation
of hyper-edited mRNA transcripts as well as miRNA precursors
(10). The data suggests that SND1
and ADAR1 are functionally synchronized in A-to-I editing and
eliminate most miRNA precursors, progressively from oocyte to
zygote (10). However, this
function is prohibited during embryogenesis. Thus, SND1 plays a
crucial role in early stage embryogenesis and cell differentiation
(10). Analysis of a SND1 knockout
mouse will provide insights into the role of SND1 in regulating
gene expression during the developmental stage.
SND1 and stress response
SND1 plays a key role in cellular stress response
via stress granule formation (47). Several environmental stimuli can be
stressful for cellular growth. In response to such stimuli, cells
undergo reprogramming in gene expression that allows cell survival
(47,48). Initial studies illustrated that
SND1 is a component of the cytoplasmic stress granules and the SN
domain is crucial for this function (47,48).
SND1 interacts with Ras GTPase activating protein SH3 domain
binding protein (G3BP) and adenosine deaminase (ADAR1). While G3BP
has been shown to be essential for assembly of stress granules
(47), ADAR1 has been linked to
apoptosis and stress response (48). ADAR1 levels are also induced by
interferon and its deficiency causes defective hematopoiesis
(48). Such studies implicate
ADAR1 in cell survival and immune response towards stress. SND1 was
found to directly interact with ADAR1. These proteins co-localized
within cytoplasmic stress granules following oxidative stress, heat
shock or interferon induction (48). Knockdown of SND1 prohibited
assembly of smaller stress foci into larger granules, emphasizing
its role in this cellular function (47). These studies need to be extended
in vivo using appropriate experimental models to confirm the
essential role of SND1 to cope with stress responses.
Regulation of cell viability
SND1 is an essential protein in normal programmed
cell death, a process mediated by caspases (49). SND1 is cleaved by caspase 3 during
drug-induced apoptosis. A non-cleavable SND1 mutant increased cell
viability and knocking down SND1 promoted drug-induced apoptosis in
HeLa cells (49). Incubation with
caspases completely blocked RNase activity of SND1 indicating that
SND1 enzymatic activity is required for maintaining cell viability
or protection from apoptosis (49).
5. Oncogenic functions of SND1
Role of SND1 in hepatocellular carcinoma
(HCC)
HCC is the primary liver malignancy, characterized
as a highly aggressive form of cancer (50). In most cases, HCC develops over a
preexisting condition, such as liver cirrhosis, non-alcoholic
steatohepatitis and viral hepatitis infection (50). It is the fifth most common cancer
and ranks third in cancer-related deaths worldwide (50). Current management options include
surgical resection, and transarterial chemoembolization (TACE),
though there is virtually no cure for this cancer (51). Molecular approaches such as the
multikinase inhibitor Sorafenib help to moderately increase the
survival of HCC patients (52). To
curb the increasing mortality rate, there is an urgent need for
identification of potential therapeutic targets that can be
employed for translational applications.
Studies demonstrate multiple ways by which SND1
contributes to hepatocarcinogenesis. Immunohistochemical analysis
has demonstrated that SND1 is overexpressed in a high percentage of
HCC patients and SND1 levels correlate with HCC stage (12). Stable clones of human HCC cells,
with either overexpression or knockdown of SND1, in nude mouse
xenograft studies confirmed the positive role of SND1 in regulating
cell viability and growth (12).
Since SND1 is also an integral component of RISC complex, we
investigated if RISC activity correlated with HCC development.
Increased RISC activity was observed in HCC cell lines, which
consequently led to an increased downregulation of tumor
suppressors via RNA interference (12). As pointed out earlier, SND1
interacts and cooperates with AEG-1 to form RISC along with other
known RISC proteins and inhibition of SND1 activity diminished the
AEG-1 activity (12). Further
investigation established that increased SND1 protein levels
trigger a cascade of molecular events that promote invasion,
proliferation, migration and angiogenesis (12,16,17).
We unraveled a linear pathway in which NF-κB activation by SND1
augments miR-221 levels (17).
Target genes of miR-221, Angiogenin and CXCL16, were observed to
promote angiogenesis (17).
Analysis of global gene expression profiles of SND1 knockdown HCC
clones identified 123 genes drastically downregulated, many of
which are in the TGFβ signaling pathway (16). We documented that increased
stabilization of AT1R mRNA by SND1 activates AT1R downstream
signaling, such as activation of ERK1/2, culminating in activation
of the TGFβ signaling pathway. The TGFβ signaling pathway closely
regulates EMT (16). Indeed, in
vitro analysis demonstrated increased invasion and migration in
SND1-overexpressing clones and vice versa in SND1 knockdown clones
(16) and expression of EMT marker
proteins, such as N-cadherin, Slug and Snail and Vimentin, was
found to be congruent with our hypothesis (16). A tissue microarray on 50 HCC cases
showed statistically significant correlation between AT1R and SND1
levels further establishing a causative relationship between SND1,
AT1R and TGFβ in hepatocarcinogenesis (16).
IGF pathways are reported to be deregulated in HCC
and found to be a cause of the aggressive tumorigenesis.
Insulin-like growth factor binding protein 3, a negative regulator
of IGF pathway, was reported to be significantly overexpressed in
SND1 knockdown HCC clones (53).
Thus SND1 might contribute to HCC by inhibiting IGFBP3 and
promoting IGF-1 activity (53).
SND1 promotes invasion and metastasis in
breast cancer
Cancer cells possess the ability to invade
circulatory system and establish tumors in tissues distant from the
primary site of tumorigenesis (54). Angiogenesis enables them to acquire
nourishment and growth factors for constitutive proliferation.
Metastatic potential of a tumor is often a measure of the fatality
of the cancer. Metastasis has been long known to be a major cause
of disease relapse and cancer related deaths. More patients succumb
to metastatic malignancy than by the primary cancer. An efficient
prognosis and diagnostic tool has not been discovered, in spite of
several attempts at analyzing differential expression patterns in
tumorous tissue.
iTRAQ based proteomic analysis of a breast cancer
metastasis model revealed that SND1 levels are upregulated with
carcinoma progression (13). This
investigation aimed at studying gene signatures in the breast
cancer metastasis model. Out of the 197 proteins differentially
regulated in cancerous tissue as compared to normal tissue, only
those that have not been reported in association with metastasis
were prioritized for further scrutiny (13). This approach helped identify 10
novel proteins significantly associated with metastasis, employing
Mass spectrometry and immunohistochemistry. SND1 was one of these
proteins, deregulated in breast cancer metastasis and showed
significant differential expression in normal versus breast cancer
tissue (13). Immunohistochemical
analysis of a tissue microarray showed upregulation of SND1 in the
majority of the cases (13).
AEG-1, also called Metadherin, is a known oncogene
associated with most oncogenic phenotype such as metastasis,
invasion, angiogenesis, chemoresistance and apoptosis (55–59).
A study aimed at exploring role of AEG1 in breast cancer
metastasis, identified SND1 as an AEG-1 interacting protein
(14). Role of SND1 in metastasis
was further emphasized based on the global gene expression
profiling of SND1 knockdown clones. The list of gene sets globally
enriched in SND1-expressing control cells versus SND1-KD cells was
strikingly dominated by those involving genes upregulated in some
component of metastatic or oncogenic signaling (14). Genes such as ANGPTL, ID1 and EREG,
known to promote specifically lung metastasis in breast cancer
model, were significantly enriched in this gene set (14). An experimental lung metastasis
study in nude mice using a highly metastatic breast cancer cell
line with SND1 knock down showed dramatic reduction in pulmonary
metastatic burden in vivo (14). Although SND1 did not promote
invasion, it was found to augment chemoresistance to cells
(14). SND1-KD cells demonstrated
sensitivity towards chemotherapeutic drug-induced apoptosis.
Microarray data from SND1-KD cells indicated upregulation of the
KiSS1 gene (14). Reporter assays
later confirmed that SND1 directly downregulates the expression of
the KiSS1 gene that is known to suppress metastasis (60). Analysis of clinical data set of
breast cancer patients with metastasis revealed that SND1 levels
strongly correlate with lung metastasis incidence and
metastasis-free survival (14).
Thus, SND1 is established to be a pro-metastasis protein. A recent
finding shows significant relationship between AEG-1-SND1
interactions and mammary tumorigenesis (15). AEG-1 knockout cells show reduced
tumor initiation and sphere formation in vivo (15). This effect could be completely
rescued by ectopic expression of AEG-1 in these cells (15). However, knocking down SND1 in these
cells completely abolished the rescue effect of ectopic AEG-1
(15). Knocking down SND1 in
AEG1+/+ cells reduced the sphere formation in
vitro and tumor formation in vivo, resembling the
phenotype of AEG1−/− cells (15). Thus, SND1 is essential for
pro-oncogenic properties of AEG-1 in breast cancer (15).
Early studies have established that SND1 cooperates
with c-Myb, a differentiation and growth factor for immature
hematopoietic stem cells promoting lymphocyte transformation
(9). Studies in breast cancer
patients show overexpression of c-Myb as well as SND1 (61). A study focused on identifying
target genes of c-Myb revealed that the SND1 promoter is one of the
target sites of c-Myb (61). There
might be a positive regulatory mechanism to maintain levels of
c-Myb and SND1 that potentially maintain tumorigenesis in breast
cancer.
SND1: an efficient diagnostic marker for
prostate cancer and colorectal cancer
Studies focused on prostate cancer have identified
SND1 as an efficient diagnostic marker (18). In 174 prostate cancer patients,
SND1 levels correlated with histological grade of the tumor
(18). SND1 protein levels were
comparable to α-methylacyl-coA racemase (AMACR) protein levels, a
currently employed marker protein for prostate cancer diagnosis
(18). Though there were some
cases where SND1 and AMACR protein levels differed, there was
robust overexpression of SND1 in tumorigenic tissue (18,45).
It was suggested that multiple protein markers, including SND1 and
AMACR, should be used for better diagnosis of the disease (18). Knocking down SND1 reduced
proliferation of prostate cancer cells demonstrating the importance
of SND1 in maintaining prostate cancer viability (18). SND1 also promotes prostate cancer
development by positively regulating CD44 alternative splicing,
allowing inclusion of variable exon v5 that is known to be
pro-oncogenic (45).
SND1 is significantly associated with colorectal
cancer. A recent genome wide analysis of methylation patterns in
CRC patients revealed a pool of genes that are differentially
methylated in cancerous tissue in comparison to adjacent normal
tissue (62). The study also
included tissue from normal, healthy patients with no familial
history of CRC, as a control. A CpG site located in SND1 gene was
identified with highest discriminative accuracy, highlighting role
of this protein in oncogenesis (62). Most of the hypermethylated CpG
sites were within the SND1 gene regions, whereas methylation in the
promoter region of SND1 was observed to be comparatively lower.
Accordingly, SND1, along with other genes was identified as a
potential diagnostic candidate gene (62). Significant correlation was observed
in nodal stage, pathological stage and co-expression of SND1 and
AEG-1 in colon cancer (63).
Immunohistochemical analyses of 196 colon cancer cases establish
that cytoplasmic expression of AEG1 and SND1 protein positively
correlates with tumor grade and cancer progression, but negatively
correlates with post-operative survival of patients (63). The study suggests potential
application of these proteins as prognostic markers for colon
cancer (63).
SND1 downregulates adenomateous polyposis coli
protein levels by post-transcriptional modification, without
altering the mRNA levels of this gene, as reported in colon cancer
(19). APC is a tumor suppressor
in colon carcinogenesis and also associated with maintaining cell
polarity and cell-cell adhesion by regulating E-cadherin
localization (19). Loss of APC
protein leads to stabilization and subsequent accumulation of
β-catenin by preventing its proteasomal degradation with subsequent
loss of contact inhibition and increased proliferation (19). These findings suggested that SND1
might be involved in early stage colon carcinogenesis. However,
whether miRNAs or RISC is involved in SND1-mediated downregulation
of APC protein remains to be studied.
SND1 in malignant glioma
Malignant gliomas are the most frequent malignant
brain tumor in adults (64).
Current multi-modality therapies include surgery, radiation and
chemotherapy, nonetheless the prognosis of malignant glioma remains
extremely poor (65). The rapid
growth and highly invasive nature of malignant glioma, favors its
infiltration into surrounding normal brain parenchyma and
facilitates recurrence after therapy (66). This aggressive disease progression
necessitates further understanding of molecular mechanism involved
in glioma growth and invasion that ultimately will lead to
identification of novel therapeutic targets of translational
relevance.
Our recent studies suggest that SND1 may provide a
novel target for malignant glioma treatment (20). In this study we found higher SND1
mRNA and protein in human malignant glioma tissue as compared to
normal brain. TCGA data showed a similar trend of high SND1
expression in human astrocytoma and glioblastoma samples as
compared to normal brain. Additionally Rembrandt data (https://caintegrator.nci.nih.gov/Rembrandt) supports
the prognostic significance of SND1 expression in which patients
with intermediate levels of SND1 survived longer than patients
showing elevated SND1 expression. Overexpression (OE) and knock
down (KD) studies were performed to unravel the functional
significance of SND1 in glioma progression. Overexpression of SND1
in immortalized primary human fetal astrocytes (IM-PHFA) (low SND1)
significantly enhanced invasion and colony formation as compared to
parental IM-PHFA cells. Conversely when SND1 was knocked down in
multiple glioma cell lines (high SND1) it significantly decreased
invasion and colony formation, both in monolayer and in soft agar.
Interestingly SND1-KD primary glioma cells demonstrated enhanced
sensitivity towards Temozolomide, an FDA approved drug used with
radiation therapy as a standard of treatment for GBM patients
(67). Knock down of SND1 in
malignant glioma cells resulted in a flat-shaped cells, which
stained positive with β-galactosidase indicating induction of
cellular senescence. Further studies documented a potential
involvement of STAT-3 in SND-1-mediating glioma invasion and
senescence-induced cell death. An intracranial xenograft study in
nude mice using a highly invasive patient-derived malignant glioma
cell line, showed a significant improved survival in SND1-KD group
(20). This was associated with a
significant decrease in proliferation marker, activated STAT3, and
angiogenesis marker and an enhanced expression of apoptotic marker.
The observation that SND1 regulates several important determinants
of glioma progression supports the rationale of using
SND1-inhibition as a means of treating glioma patients.
6. Specific inhibitors of SND1
BLAST search reveals that SND1 is the only
eukaryotic protein with a Tudor and SN fusion domain. Hence, this
quaternary fold can be employed for targeted therapeutic
approaches, developing SND1 specific inhibitors. A similar protein
domain is observed in Plasmodium falciparum, a parasite
causing malarial infections in humans. Studies with P.
falciparum SND1 have confirmed that the SN domain is involved
in nuclease activity, whereas the Tudor domain carries out the
function of RNA binding (68). 3′,
5′-Deoxythymidine bisphosphate (pdTp) is a competitive chemical
inhibitor against SNases (6). In
P. falciparum, it not only blocks the nuclease activity, but
also inhibits RNA/protein interaction of SND1 (68). pdTp inhibited growth of both
chloroquine-sensitive and chloroquine-resistant strains of P.
falciparum at a concentration of 100–200 μM. Nevertheless, it
did not inhibit the growth of human HCC cell line Huh7-D12 at a
concentration as high as 800 μM suggesting that pdTp might be
developed as an anti-malarial drug (68). However, studies performed in our
laboratory using multiple human HCC cell lines demonstrated that
pdTp treatment resulted in significant reduction in cell viability
and colony forming potential (12). This effect was observed at high
concentrations, ranging from 100 to 200 μM thus rendering it
ineffective for clinical applications. Further high throughput
screening assays need to be performed to develop clinically
relevant chemical inhibitors against SND1 enzymatic activity.
Employing the C terminus fusion domain of SND1 might prove to be a
productive effort in designing targeted molecular therapy.
7. Conclusion and future direction
SND1 is known to orchestrate a series of changes
that affect global gene expression and confer cellular
transformation. It regulates gene expression employing multiple
unique mechanisms that function at transcriptional as well as
post-transcriptional levels. With increasing relevance of studies
on SND1 in the context of carcinogenesis it is important to
elucidate the molecular mechanisms underlying SND1 activity.
Studies so far suggest that it is a promising molecule for clinical
investigation and targeted therapeutic management of cancer.
Considering the pleiotrophic functions of SND1, one important
question that has not been addressed is the role of SND1 in
maintaining normal physiological function(s) including growth and
development. An SND1 knockout mouse, conditional and global, will
not only provide comprehensive insights into the physiological
functions of SND1, but also will be an ideal model to interrogate
the role of SND1 in immortalization, transformation, metastasis and
overall cancer development and progression. Conversely,
organ-specific SND1 transgenic mouse will also provide useful
insight into the oncogenic function of overexpressed SND1. Is SND1
overexpression alone sufficient for transforming normal cells into
cancer cells? Can SND1 function as a driver for tumorigenesis or is
it a passenger and expediter of transformation following initial
mutagenic events? Which particular aspects of SND1 function are
most relevant to confer its oncogenic properties? Future studies
need to focus on and address these questions, which will provide
important insights into the precise role of SND1 in normal and
abnormal physiology.
Acknowledgements
The present study was supported in part by The James
S. McDonnell Foundation and National Cancer Institute Grants R01
CA138540 and R21 CA183954 (D.S.), National Institutes of Health
grant R01 CA134721 (P.B.F.). D.S. is a Harrison Scholar in the VCU
Massey Cancer Center. P.B.F. holds the Thelma Newmeyer Corman Chair
in Cancer Research.
References
|
1
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Almendro V, Marusyk A and Polyak K:
Cellular heterogeneity and molecular evolution in cancer. Annu Rev
Pathol. 8:277–302. 2013. View Article : Google Scholar
|
|
3
|
Callebaut I and Mornon JP: The human
EBNA-2 coactivator p100: multidomain organization and relationship
to the staphylococcal nuclease fold and to the tudor protein
involved in Drosophila melanogaster development. Biochem J.
321:125–132. 1997.PubMed/NCBI
|
|
4
|
Yang J, Aittomaki S, Pesu M, et al:
Identification of p100 as a coactivator for STAT6 that bridges
STAT6 with RNA polymerase II. EMBO J. 21:4950–4958. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gao X, Zhao X, Zhu Y, et al: Tudor
staphylococcal nuclease (Tudor-SN) participates in small
ribonucleoprotein (snRNP) assembly via interacting with
symmetrically dimethylated Sm proteins. J Biol Chem.
287:18130–18141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Caudy AA, Ketting RF, Hammond SM, et al: A
micrococcal nuclease homologue in RNAi effector complexes. Nature.
425:411–414. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Paukku K, Kalkkinen N, Silvennoinen O,
Kontula KK and Lehtonen JY: p100 increases AT1R expression through
interaction with AT1R 3′-UTR. Nucleic Acids Res. 36:4474–4487.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Paukku K, Yang J and Silvennoinen O: Tudor
and nuclease-like domains containing protein p100 function as
coactivators for signal transducer and activator of transcription
5. Mol Endocrinol. 17:1805–1814. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leverson JD, Koskinen PJ, Orrico FC, et
al: Pim-1 kinase and p100 cooperate to enhance c-Myb activity. Mol
Cell. 2:417–425. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Garcia-Lopez J, de Hourcade JD and Del
Mazo J: Reprogramming of microRNAs by adenosine-to-inosine editing
and the selective elimination of edited microRNA precursors in
mouse oocytes and preimplantation embryos. Nucleic Acids Res.
41:5483–5493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li CL, Yang WZ, Chen YP and Yuan HS:
Structural and functional insights into human Tudor-SN, a key
component linking RNA interference and editing. Nucleic Acids Res.
36:3579–3589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoo BK, Santhekadur PK, Gredler R, et al:
Increased RNA-induced silencing complex (RISC) activity contributes
to hepatocellular carcinoma. Hepatology. 53:1538–1548. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ho J, Kong JW, Choong LY, et al: Novel
breast cancer metastasis-associated proteins. J Proteome Res.
8:583–594. 2009. View Article : Google Scholar
|
|
14
|
Blanco MA, Aleckovic M, Hua Y, et al:
Identification of staphylococcal nuclease domain-containing 1
(SND1) as a Metadherin-interacting protein with
metastasis-promoting functions. J Biol Chem. 286:19982–19992. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wan L, Lu X, Yuan S, et al: MTDH- SND1
interaction is crucial for expansion and activity of
tumor-initiating cells in diverse oncogene- and carcinogen-induced
mammary tumors. Cancer Cell. 26:92–105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Santhekadur PK, Akiel M, Emdad L, et al:
Staphylococcal nuclease domain containing-1 (SND1) promotes
migration and invasion via angiotensin II type 1 receptor (AT1R)
and TGFbeta signaling. FEBS Open Bio. 4:353–361. 2014. View Article : Google Scholar
|
|
17
|
Santhekadur PK, Das SK, Gredler R, et al:
Multifunction protein staphylococcal nuclease domain containing 1
(SND1) promotes tumor angiogenesis in human hepatocellular
carcinoma through novel pathway that involves nuclear factor kappaB
and miR-221. J Biol Chem. 287:13952–13958. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuruma H, Kamata Y, Takahashi H, et al:
Staphylococcal nuclease domain-containing protein 1 as a potential
tissue marker for prostate cancer. Am J Pathol. 174:2044–2050.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsuchiya N, Ochiai M, Nakashima K, Ubagai
T, Sugimura T and Nakagama H: SND1, a component of RNA-induced
silencing complex, is up-regulated in human colon cancers and
implicated in early stage colon carcinogenesis. Cancer Res.
67:9568–9576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Emdad L, Janjic A, Al-Zubi M, et al:
Suppression of miR-184 in malignant gliomas upregulates SND1 and
promotes tumor aggressiveness. Neuro Oncol. Sep 12–2014.(Epub ahead
of print). pii: nou220. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lienard P, Riviere M, Van Vooren P,
Szpirer C and Szpirer J: Assignment of SND1, the gene encoding
coactivator p100, to human chromosome 7q31.3 and rat chromosome
4q23 by in situ hybridization. Cytogenet Cell Genet. 90:253–254.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Voeghtly LM, Mamula K, Campbell JL,
Shriver CD and Ellsworth RE: Molecular alterations associated with
breast cancer mortality. PLoS One. 7:e468142012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jenkins RB, Qian J, Lee HK, et al: A
molecular cytogenetic analysis of 7q31 in prostate cancer. Cancer
Res. 58:759–766. 1998.PubMed/NCBI
|
|
24
|
Glukhova L, Lavialle C, Fauvet D, et al:
Mapping of the 7q31 subregion common to the small chromosome 7
derivatives from two sporadic papillary renal cell carcinomas:
increased copy number and overexpression of the MET proto-oncogene.
Oncogene. 19:754–761. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Theobald DL, Mitton-Fry RM and Wuttke DS:
Nucleic acid recognition by OB-fold proteins. Annu Rev Biophys
Biomol Struct. 32:115–133. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ying M and Chen D: Tudor domain-containing
proteins of Drosophila melanogaster. Dev Growth Differ. 54:32–43.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Armengol S, Arretxe E, Rodriguez L, Ochoa
B, Chico Y and Martinez MJ: NF-kappaB, Sp1 and NF-Y as
transcriptional regulators of human SND1 gene. Biochimie.
95:735–742. 2013. View Article : Google Scholar
|
|
28
|
Rodriguez L, Bartolome N, Ochoa B and
Martinez MJ: Isolation and characterization of the rat SND p102
gene promoter: putative role for nuclear factor-Y in regulation of
transcription. Ann NY Acad Sci. 1091:282–295. 2006. View Article : Google Scholar
|
|
29
|
Rodriguez L, Ochoa B and Martinez MJ: NF-Y
and Sp1 are involved in transcriptional regulation of rat SND p102
gene. Biochem Biophys Res Commun. 356:226–232. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fashe T, Saarikettu J, Isomaki P, Yang J
and Silvennoinen O: Expression analysis of Tudor-SN protein in
mouse tissues. Tissue Cell. 45:21–31. 2013. View Article : Google Scholar
|
|
31
|
Keenan TW, Winter S, Rackwitz HR and Heid
HW: Nuclear coactivator protein p100 is present in endoplasmic
reticulum and lipid droplets of milk secreting cells. Biochim
Biophys Acta. 1523:84–90. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tong X, Drapkin R, Yalamanchili R,
Mosialos G and Kieff E: The Epstein-Barr virus nuclear protein 2
acidic domain forms a complex with a novel cellular coactivator
that can interact with TFIIE. Mol Cell Biol. 15:4735–4744.
1995.PubMed/NCBI
|
|
33
|
Valineva T, Yang J, Palovuori R and
Silvennoinen O: The transcriptional co-activator protein p100
recruits histone acetyl-transferase activity to STAT6 and mediates
interaction between the CREB-binding protein and STAT6. J Biol
Chem. 280:14989–14996. 2005. View Article : Google Scholar
|
|
34
|
Bromberg J: Stat proteins and oncogenesis.
J Clin Invest. 109:1139–1142. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu X, Dong L, Zhang X, et al:
Identification of p100 target promoters by chromatin
immunoprecipitation-guided ligation and selection (ChIP-GLAS). Cell
Mol Immunol. 8:88–91. 2011. View Article : Google Scholar
|
|
36
|
Sarkar D and Fisher PB: AEG-1/MTDH/LYRIC:
clinical significance. Adv Cancer Res. 120:39–74. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yoo BK, Emdad L, Lee SG, et al: Astrocyte
elevated gene-1 (AEG-1): A multifunctional regulator of normal and
abnormal physiology. Pharmacol Ther. 130:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Milochau A, Lagree V, Benassy MN, et al:
Synaptotagmin 11 interacts with components of the RNA-induced
silencing complex RISC in clonal pancreatic beta-cells. FEBS Lett.
588:2217–2222. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Heinrich EM, Wagner J, Kruger M, et al:
Regulation of miR-17-92a cluster processing by the microRNA binding
protein SND1. FEBS Lett. 587:2405–2411. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gao X, Shi X, Fu X, et al: Human Tudor
staphylococcal nuclease (Tudor-SN) protein modulates the kinetics
of AGTR1–3′UTR granule formation. FEBS Lett. 588:2154–2161. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lei Y, Huang Y, Zhang H, Yu L, Zhang M and
Dayton A: Functional interaction between cellular p100 and the
dengue virus 3′ UTR. J Gen Virol. 92:796–806. 2011. View Article : Google Scholar
|
|
42
|
Wahl MC, Will CL and Luhrmann R: The
spliceosome: design principles of a dynamic RNP machine. Cell.
136:701–718. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
David CJ and Manley JL: Alternative
pre-mRNA splicing regulation in cancer: pathways and programs
unhinged. Genes Dev. 24:2343–2364. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang J, Valineva T, Hong J, et al:
Transcriptional co-activator protein p100 interacts with snRNP
proteins and facilitates the assembly of the spliceosome. Nucleic
Acids Res. 35:4485–4494. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cappellari M, Bielli P, Paronetto MP, et
al: The transcriptional co-activator SND1 is a novel regulator of
alternative splicing in prostate cancer cells. Oncogene.
33:3794–3802. 2014. View Article : Google Scholar
|
|
46
|
Nishikura K: Functions and regulation of
RNA editing by ADAR deaminases. Annu Rev Biochem. 79:321–349. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gao X, Ge L, Shao J, et al: Tudor-SN
interacts with and co-localizes with G3BP in stress granules under
stress conditions. FEBS Lett. 584:3525–3532. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Weissbach R and Scadden AD: Tudor-SN and
ADAR1 are components of cytoplasmic stress granules. RNA.
18:462–471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sundstrom JF, Vaculova A, Smertenko AP, et
al: Tudor staphylococcal nuclease is an evolutionarily conserved
component of the programmed cell death degradome. Nat Cell Biol.
11:1347–1354. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Villanueva A, Hernandez-Gea V and Llovet
JM: Medical therapies for hepatocellular carcinoma: a critical view
of the evidence. Nat Rev Gastroenterol Hepatol. 10:34–42. 2013.
View Article : Google Scholar
|
|
52
|
Llovet JM, Ricci S, Mazzaferro V, et al:
Sorafenib in advanced hepatocellular carcinoma. N Engl J Med.
359:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yin J, Ding J, Huang L, et al: SND1
affects proliferation of hepatocellular carcinoma cell line
SMMC-7721 by regulating IGFBP3 expression. Anat Rec. 296:1568–1575.
2013. View Article : Google Scholar
|
|
54
|
Nguyen DX, Bos PD and Massague J:
Metastasis: from dissemination to organ-specific colonization. Nat
Rev Cancer. 9:274–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Brown DM and Ruoslahti E: Metadherin, a
cell surface protein in breast tumors that mediates lung
metastasis. Cancer Cell. 5:365–374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hu G, Chong RA, Yang Q, et al: MTDH
activation by 8q22 genomic gain promotes chemoresistance and
metastasis of poor-prognosis breast cancer. Cancer Cell. 15:9–20.
2009. View Article : Google Scholar :
|
|
57
|
Yoo BK, Emdad L, Su ZZ, et al: Astrocyte
elevated gene-1 regulates hepatocellular carcinoma development and
progression. J Clin Invest. 119:465–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yoo BK, Chen D, Su ZZ, et al: Molecular
mechanism of chemoresistance by astrocyte elevated gene-1. Cancer
Res. 70:3249–3258. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Emdad L, Lee SG, Su ZZ, et al: Astrocyte
elevated gene-1 (AEG-1) functions as an oncogene and regulates
angiogenesis. Proc Natl Acad Sci USA. 106:21300–21305. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu W, Beck BH, Vaidya KS, et al:
Metastasis suppressor KISS1 seems to reverse the Warburg effect by
enhancing mitochondrial biogenesis. Cancer Res. 74:954–963. 2014.
View Article : Google Scholar
|
|
61
|
Quintana AM, Liu F, O’Rourke JP and Ness
SA: Identification and regulation of c-Myb target genes in MCF-7
cells. BMC Cancer. 11:302011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Naumov VA, Generozov EV, Zaharjevskaya NB,
et al: Genome-scale analysis of DNA methylation in colorectal
cancer using Infinium HumanMethylation450 BeadChips. Epigenetics.
8:921–934. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang N, Du X, Zang L, et al: Prognostic
impact of Metadherin-SND1 interaction in colon cancer. Mol Biol
Rep. 39:10497–10504. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ostrom QT, Gittleman H, Farah P, et al:
CBTRUS statistical report: primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol.
15:1–56. 2013. View Article : Google Scholar
|
|
65
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Lefranc F, Brotchi J and Kiss R: Possible
future issues in the treatment of glioblastomas: special emphasis
on cell migration and the resistance of migrating glioblastoma
cells to apoptosis. J Clin Oncol. 23:2411–2422. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Stupp R, Mason WP, van den Bent MJ, et al:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Hossain MJ, Korde R, Singh S, et al: Tudor
domain proteins in protozoan parasites and characterization of
Plasmodium falciparum tudor staphylococcal nuclease. Int J
Parasitol. 38:513–526. 2008. View Article : Google Scholar
|