Introduction
Endometrial cancer is the sixth most common incident
cancer in women worldwide, and this prevalence may reflect recent
changes in lifestyles of women (1). Endometrial cancer is classified into
Types I and II based on etiology and clinical behavior (2,3).
Type I tends to appear with age, either before or after menopause,
and comprises ~80% of all cases of endometrial cancer. Type I
tumors are estrogen-dependent and well-differentiated
adenocarcinomas that occur against a background of endometrial
hyperplasia and have a favorable prognosis. PTEN and
K-ras gene mutations are connected to their development.
Type II tumors are poorly-differentiated adenocarcinomas that tend
to occur at a relatively advanced age and include clear cell and
serous adenocarcinomas. These tumors are not estrogen-dependent,
occur de novo against a background of endometrial atrophy,
and have a poor prognosis. P53 gene mutation and high
chromosomal stability are connected to their development. Although
the clinicopathogenic backgrounds of Types I and II differ, the
treatments are similar. Type II adenocarcinomas are resistant to
current therapies and these tumors continue to have a poor
prognosis. Thus, improved treatment for endometrial cancer requires
improved understanding of the carcinogenic mechanism and
development of therapeutic strategies that are specific to each
patient’s condition.
The human Aurora kinase family includes three
subtypes: A, B, and C. Aurora kinase A (AURKA) and B are
overexpressed in many human cancer cell-derived cell lines and
cancer tissues, and are connected to carcinogenesis (4). AURKA is a G2/M phase serine/threonine
kinase that mainly accumulates at centrosomes during late G2 phase
anaphase and plays a role in centrosome separation and bipolar
spindle formation and stabilization (5,6).
AURKA is regulated to ensure proper mitosis, and its overexpression
induces an increase in centrosome number and aneuploid cell
formation, leading to a significant risk of carcinogenesis
(4,7–9).
AURKA overexpression occurs in chromosomal region 20q13, at which
gene amplification is seen in many human cancers; and is involved
in colorectal (10), bladder
(11), pancreatic (12), gastric (13) and breast (14) cancers. In ovarian cancer that is a
poor prognostic gynecological cancer, AURKA overexpression is also
found in cell lines and cancer tissues and is associated with poor
prognosis in cancer patients (15,16).
AURKA overexpression also increases resistance to taxanes, which
are the principal chemotherapeutic drugs for gynecologic
malignancies (17). Recent reports
showed the potential efficacy of combining AURKA inhibitor with
taxanes in epithelial ovarian cancer (18). AURKA has been noted to be a novel
therapeutic target for the gynecological malignancies that are
particularly resistance to taxanes. However, only a few reports
have described a role for AURKA in endometrial cancer. Kurai et
al found significantly increased expression of AURKA and AURKB
in endometrial cancer compared to normal proliferative tissue, with
particularly high expression of AURKB in poorly-differentiated
endometrial cancer and correlation of this expression with
worsening prognosis (19). In a
microarray analysis of endometrial cancer tissue, Moreno-Bueno
et al showed that AURKA is highly expressed in Type II
adenocarcinoma (20). Thus,
abnormalities in cell cycle checkpoint mechanisms may play a role
in carcinogenesis of particular endometrial cancers. However, the
significance of its expression in endometrial cancer is not fully
understood. The aim of this study was to clarify the significance
of AURKA expression in endometrial cancer.
Materials and methods
Patients and tissue samples
Tissues were obtained from 162 patients with
endometrial carcinoma and from 30 women with normal endometrium who
underwent surgery at Keio University (Tokyo, Japan) from 2003 to
2006. All specimens were fixed in 10% phosphate-buffered formalin
and embedded in paraffin. Sections of 3 μm were stained with
hematoxylin and eosin to confirm the presence of a tumor and to
assess the tumor histological characteristics. The items for
immunohistochemistry are summarized in Table I. Written informed consent was
obtained from the patients regarding use of samples for research.
The study was approved by the Ethics Committee of Keio University
(approval no. 20130159).
 | Table IRelationship between overexpression
of AURKA and clinicopathological factors. |
Table I
Relationship between overexpression
of AURKA and clinicopathological factors.
| Overexpression of
AURKA | Negative
(n=110) | Positive
(n=82) | aP |
|---|
| Endometrial tissues
(NEM vs. EC) | | | 0.000 |
| NEM | | | |
| Proliferative
phase | 13 | 2 | |
| Secretory
phase | 15 | 0 | |
| EC | 82 | 80 | |
| FIGO surgical stage
(I, II vs. III, IV) | | | 0.307 |
| I | 63 | 53 | |
| II | 3 | 6 | |
| III | 14 | 18 | |
| IV | 2 | 3 | |
| Histological type
(Non-EA vs. EA) | | | 0.026 |
| Non-EA | | | |
| Serous
adenocarcinoma | 2 | 4 | |
| Clear cell
adenocarcinoma | 0 | 5 | |
| EA | 80 | 71 | |
| Grade (G1 and 2 vs.
G3) | | | 0.005 |
| G1 | 48 | 34 | |
| G2 | 23 | 16 | |
| G3 | 9 | 21 | |
| Recurrence rate
(%) | 19.5 | 31.25 | 0.086 |
Immunohistochemical staining
Samples were deparaffinized in xylene and rehydrated
in a graded series of ethanol. Antigen retrieval was performed with
a 10-min autoclave treatment in 10 mM citrate buffer, pH 7.0. After
blocking of endogenous peroxidase activity by dipping sections in
0.3% H2O2 in PBS for 5 min, sections were
incubated overnight with primary antibodies at 4°C in a humid
chamber. A primary antibody against AURKA (Trans Genic, Inc.,
Kumamoto, Japan) was applied at a dilution of 1:200 and anti-Ki67
monoclonal antibody (Dako Denmark A/S, Glostrup, Denmark) was used
at a dilution of 1:100. Indirect immunohistochemical staining was
performed by the avidin-biotin-peroxidase complex method using a
Vectastain Elite ABC kit (Funakoshi Co., Ltd., Tokyo, Japan), using
3,3′-diaminobenzidine as a chromogen and
H2O2. Sections were counterstained with
hematoxylin, dehydrated in a graded series of ethanol, dried and
coverslipped. TUNEL staining was performed using an In Situ
Cell Death Detection kit (Roche Diagnostics GmbH, Mannheim,
Germany) according to the manufacturer’s instructions.
Evaluation of immunohistochemical
staining
AURKA staining was mainly seen in the nucleus.
Overexpression of AURKA was defined as >30% of tumor cells or
normal endometrial cells showing nuclear immunoreactivity in five
hyper-power fields in each section as previously reported (21). Slides were independently evaluated
by two investigators in a blinded manner. In TUNEL and Ki67
staining, positive cells were counted and the percentage of
positive cells out of the total number of cancer cells was
calculated.
Cell line and culture
Four human endometrial cancer cell lines were used:
SNG-M and HHUA were kindly provided by Professor Shiro Nozwa and Dr
Isamu Ishiwata; and HEC-1B and HEC-108 were purchased from the
Health Science Research Resources Bank. All cells were maintained
in Ham’s F12 (Sigma-Aldrich, St. Louis, MO, USA) supplemented with
10% FBS with appropriate antibiotics at 37°C in a 5% CO2
humidified incubator.
RT-PCR analysis
Total RNA from HEC-108 and -1B, as well as HHUA, and
SNG-M cells was extracted for investigation of expression of AURKA
using a RNeasy Mini kit (Qiagen, Tokyo, Japan). cDNA was
synthesized from 1 μg of total RNA using SuperScript II Reverse
Transcriptase (Invitrogen Life Technologies, Carlsbad, CA, USA).
AURKA expression was analyzed in a RT-PCR assay using 1 μl of
first-stand cDNA as template. AmpliTaq Gold and 10X PCR
buffer/MgCl2 with dNTP were used in the PCR assay, with
analysis using a GeneAMP PCR system 9700 (Applied Biosystems,
Foster City, CA, USA). The primer sequences were 5′-ATT GCA GAT TTT
GGG TGG T-3′ (sense), and 5′-AAA CTT CAG TAG CAT GTT CCT GTC-3′
(antisense), 472 bp. PCR was performed for 30 cycles (94, 57 and
72°C for 30 sec, respectively).
Western blot analysis
Western blot analysis was performed to confirm the
effect of AURKA inhibition by transfection of AURKA siRNA.
siRNA-transfected endometrial cancer-derived cells were rinsed with
PBS twice, trypsinized, and centrifuged at 1,000 rpm for 5 min at
room temperature. Cells were lysed using a Mammalian Cell
Extraction kit (BioVision Research Products, Mountain View, CA,
USA). The sample was mixed with sample buffer containing the
equivalent volume of 5% β-mercaptoethanol (both from Bio-Rad,
Hercules, CA, USA) and the mixture was boiled at 100°C for 5 min.
After boiling, the mixture was electrophoresed on a 10%
polyacrylamide gel and the proteins were transferred to
nitrocellulose membranes (Bio-Rad). The membranes were soaked in
PBS containing 1% BSA and 0.1% Tween-20 and incubated at room
temperature for 1 h for blocking. They were then reacted with
anti-β-actin antibody (1:5,000 diluted, AC-74; Sigma-Aldrich) and
anti-AURKA antibody (1:100 diluted; Trans Genic, Inc.) at 4°C
overnight, followed by rinsing three times with PBS containing 0.1%
Tween (PBS-T) for 10 min each. Anti-β-actin samples were reacted
with anti-mouse IgG antibody (PK-6102) and anti-AURKA samples were
reacted with anti-rabbit IgG antibody (PK-6101) (both from Vector
Laboratories, Burlingame, CA, USA) at room temperature for 1 h. The
membranes were rinsed with PBS-T three times and reacted with ABC
complex (pre-reacted at 4°C for 30 min, PK-6100; Vector
Laboratories) at room temperature for 1 h, then rinsed with PBS-T
twice and PBS once, and visualized with DAB (Sigma-Aldrich).
siRNA AURKA inhibition
siRNA duplexes (siAURKA sense, 5′-AUG CCC UGU CUU
ACU GUC ATT-3′; and control sense, 5′-ATC CGC CGC ATA GTA CGT A-3′)
were selected and synthesized (Cosmo Bio Co., Ltd., Tokyo, Japan).
Transfection of double-stranded siRNA was performed using siFECTOR
(B-Bridge International, Inc., Sunnyvale, CA, USA). HEC-1B cells
were seeded in 6-cm plates 48 h before transfection. In each plate,
300 pmol of Aurora A siRNA or negative control siRNA and 18 μl of
siFECTOR were added to MEM and mixed. After incubation, the siRNA
and siFECTOR solutions were mixed gently and added to the plates
according to manufacturer’s instructions. The plate was incubated
for 48 h until it was ready for further assay.
Colony formation assay
For colony formation, transfected cells were plated
at a density of 1×104 cells/10 cm dish. After 10 days,
colonies were stained with 0.1% crystal violet in 50% methanol and
photographed with an inverted phase contrast microscope.
Migration and invasion assay
Migration and invasion assays were performed after
transfection of AURKA siRNA for 48 h. BD BioCoat Control Insert
Chambers, 24-well plates with an 8-μm pore size (no. 353097) and BD
BioCoat Matrigel invasion chambers (no. 354480) (all from BD
Biosciences, Franklin Lakes, NJ, USA) were used for migration and
invasion assays. In the respective assays, 1 and 2×105
cells/well were plated in the upper compartment in 0.5 ml of
serum-free medium. The lower compartment contained 0.75 ml of
medium with 10% FCS. Cells were incubated at 37°C in a 5%
CO2/95% air incubator for 30 h. Cells in the upper
compartment were carefully removed with a cotton swab. Cells that
migrated or invaded the lower surface of the membrane were fixed
and stained using a Diff-Quik kit (Sysmex Corp., Kobe, Japan) and
invading cells were counted in five randomly selected microscope
fields.
Chemosensitivity analysis
HEC-1B cells were seeded in 96-well plates for 48 h
at 37°C in a 5% CO2-humidified incubator. These cells
were treated with various concentrations of paclitaxel, cisplatin
or adriamycin with or without AURKA siRNA transfection. Viable
cells were quantified 48 h after administration of anticancer drugs
using a Cell Counting kit (Dojindo, Kumamoto, Japan). Cytotoxicity
was measured by determining the IC50, the concentration
of drug inducing a 50% reduction in cell growth compared with
control.
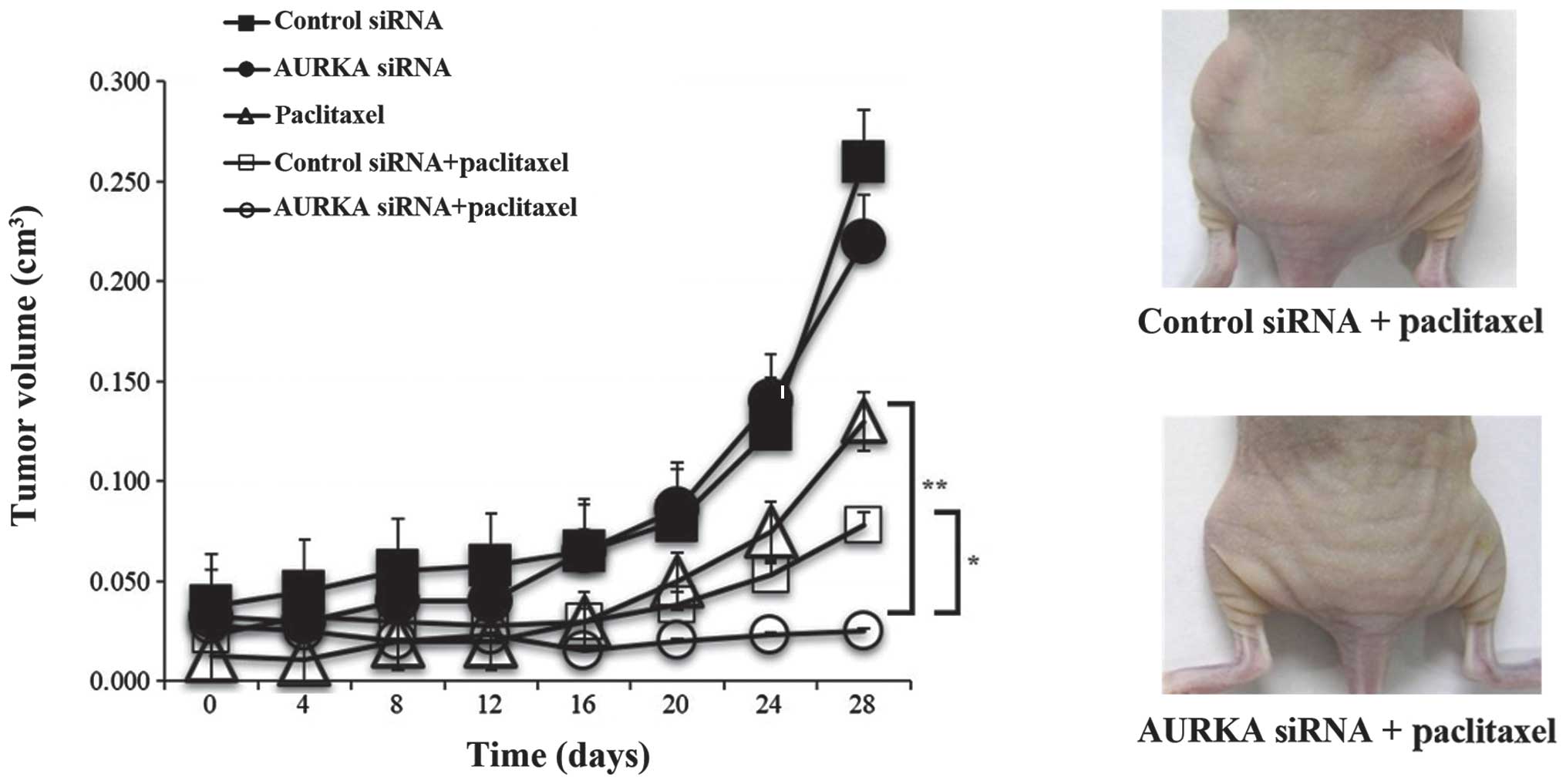
In vivo experiments
The care and use of animals in the study were
approved by the Animal Research Center at Keio University. HEC-1B
cells (1×107) were subcutaneously injected at two sites
in the flank of female nude mice obtained from CLEA Japan, Inc.
(Tokyo, Japan). Twenty days later, mice with tumor formation were
randomly divided into five groups treated with control siRNA, AURKA
siRNA, paclitaxel, control siRNA and paclitaxel, and AURKA siRNA
and paclitaxel. Paclitaxel was administered intraperitoneally at 20
mg/kg and a mixture of AURKA siRNA and atelocollagen (Koken Co.,
Ltd., Tokyo, Japan) was injected at the tumor site on alternate
days. Tumor diameters were measured every 4 days and the tumor
volume (mm3) was calculated using the following formula:
V = length × (wide)2 × 1/2. Five tumors were used in
each group. Apoptosis was quantified by TUNEL analysis and
proliferative index was measured by Ki67 staining.
Statistical analysis
Correlations between overexpression of AURKA and
clinicopathological factors were evaluated by χ2 test.
The Kaplan-Meier method was used to estimate the probability of
disease-free survival (DFS) and a log-rank test was used to compare
DFS between groups. P<0.05 was considered to indicate
statistical significance. All analyses were performed using IBM
SPSS Statistics 21.0 (IBM Japan, Ltd., Tokyo, Japan).
Results
Relationship between overexpression of
AURKA and clinicopathological factors
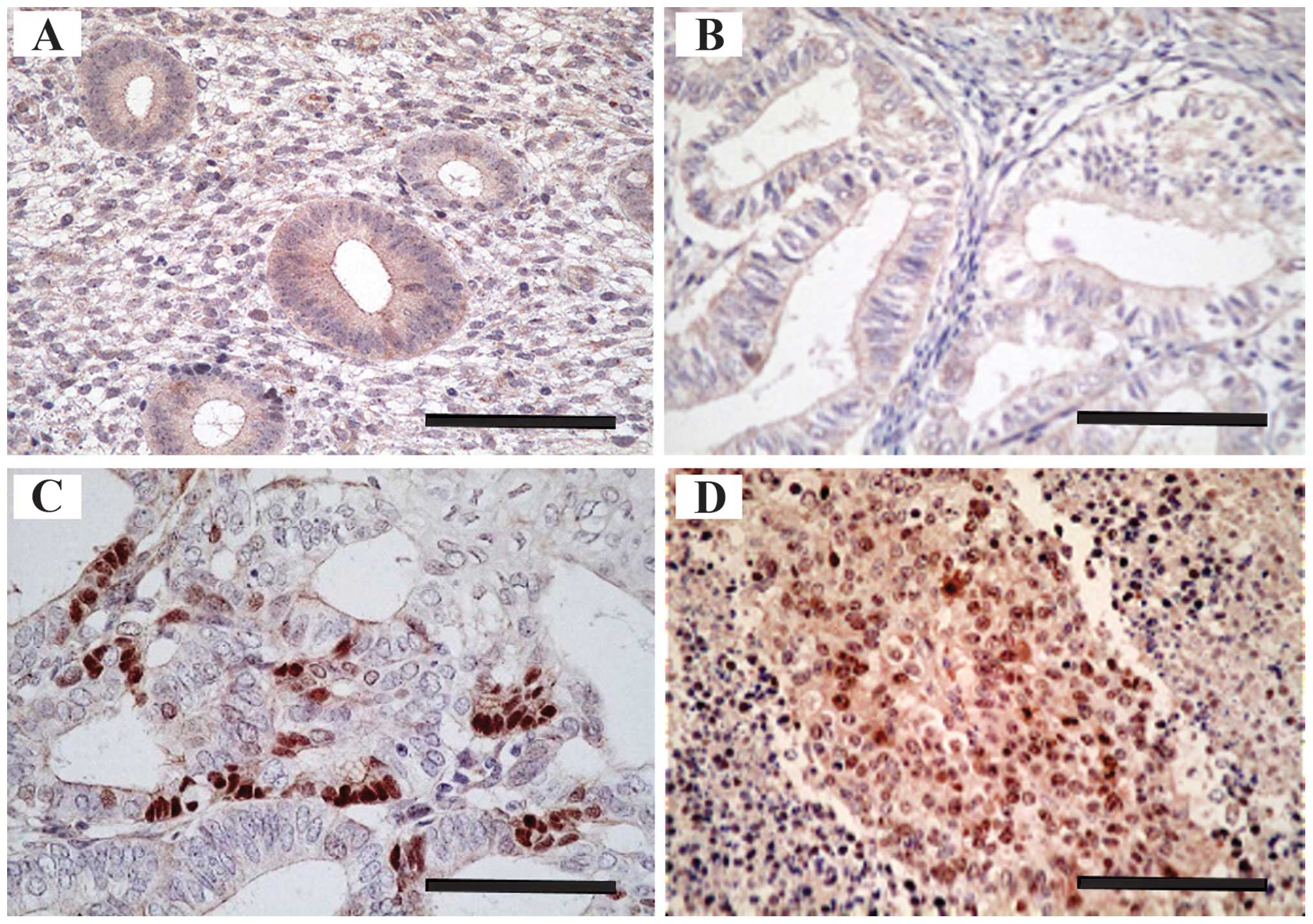
AURKA was expressed mainly in the nucleus in normal
endometrium and endometrial cancer tissues (Fig. 1A–D). As described in Table I, overexpression of AURKA occurred
at a significantly higher rate in endometrial carcinoma than in
normal endometrial tissues (49 vs. 3%). Normal endometrial samples
that showed overexpression of AURKA were all proliferative phase
endometria. In endometrial cancer tissues, overexpression of AURKA
was significantly higher in non-endometrioid than in endometrioid
adenocarcinoma (81 vs. 47%, P=0.026) and in poorly-differentiated
(grade 3) tumors compared with well- (grade 1) or moderately-
(grade 2) differentiated tumors in endometrioid adenocarcinoma (70
vs. 41%, P=0.005). However, surgical stage (FIGO 2008) was not
associated with overexpression of AURKA (stage I, II vs. III, IV:
47 vs. 57%, P=0.307).
Association between AURKA expression and
patient prognosis
The follow-up period of the 162 patients ranged from
4 to 127 months, with a median of 86 months. Twelve patients had
relapse of the disease and 16 died due to the disease. The
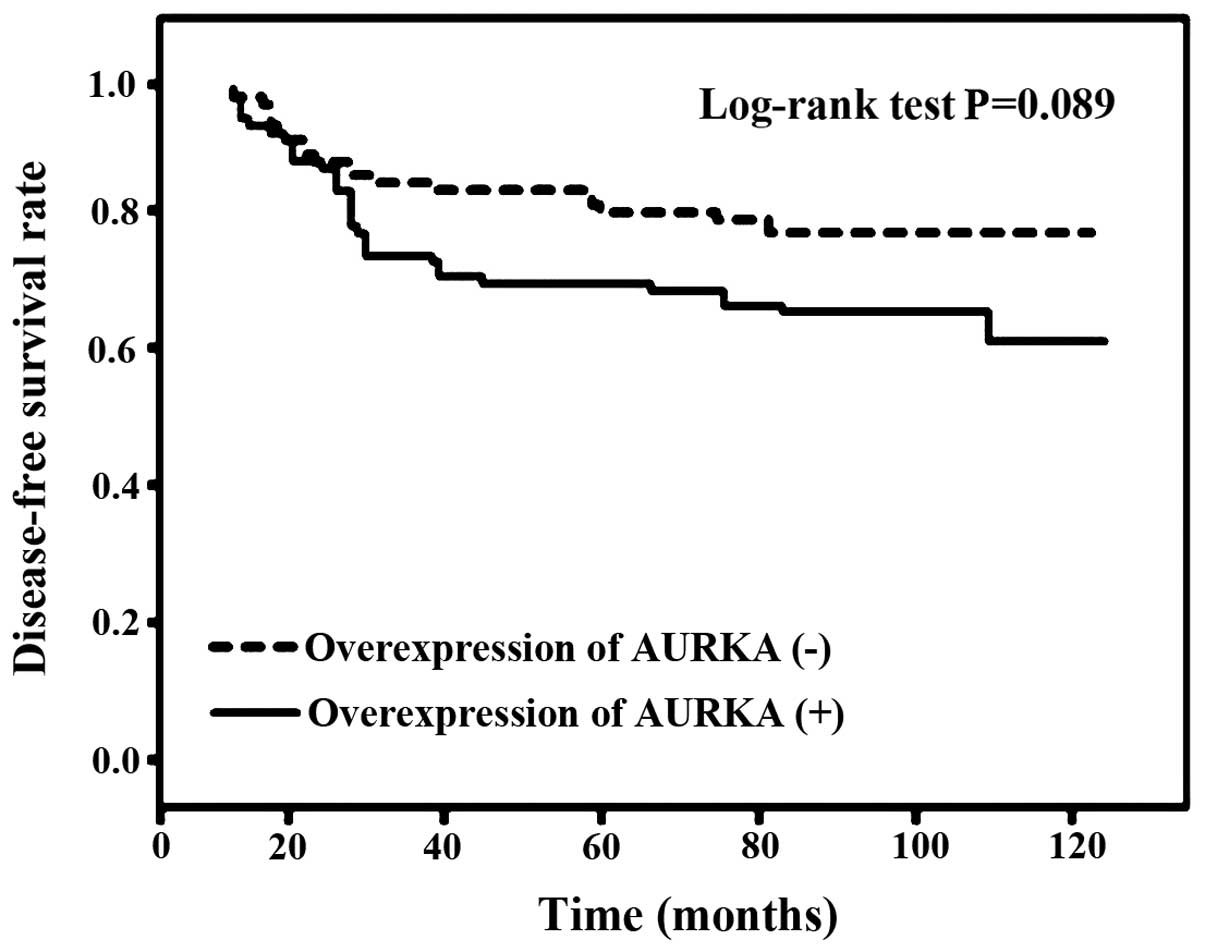
recurrence rate tended to be higher (P=0.086; Table I) and DFS tended to be shorter
(P=0.087; Fig. 2) in patients with
overexpression of AURKA compared to those without
overexpression.
Knockdown of AURKA in HEC-1B cells
decreases invasion, migration and colony formation
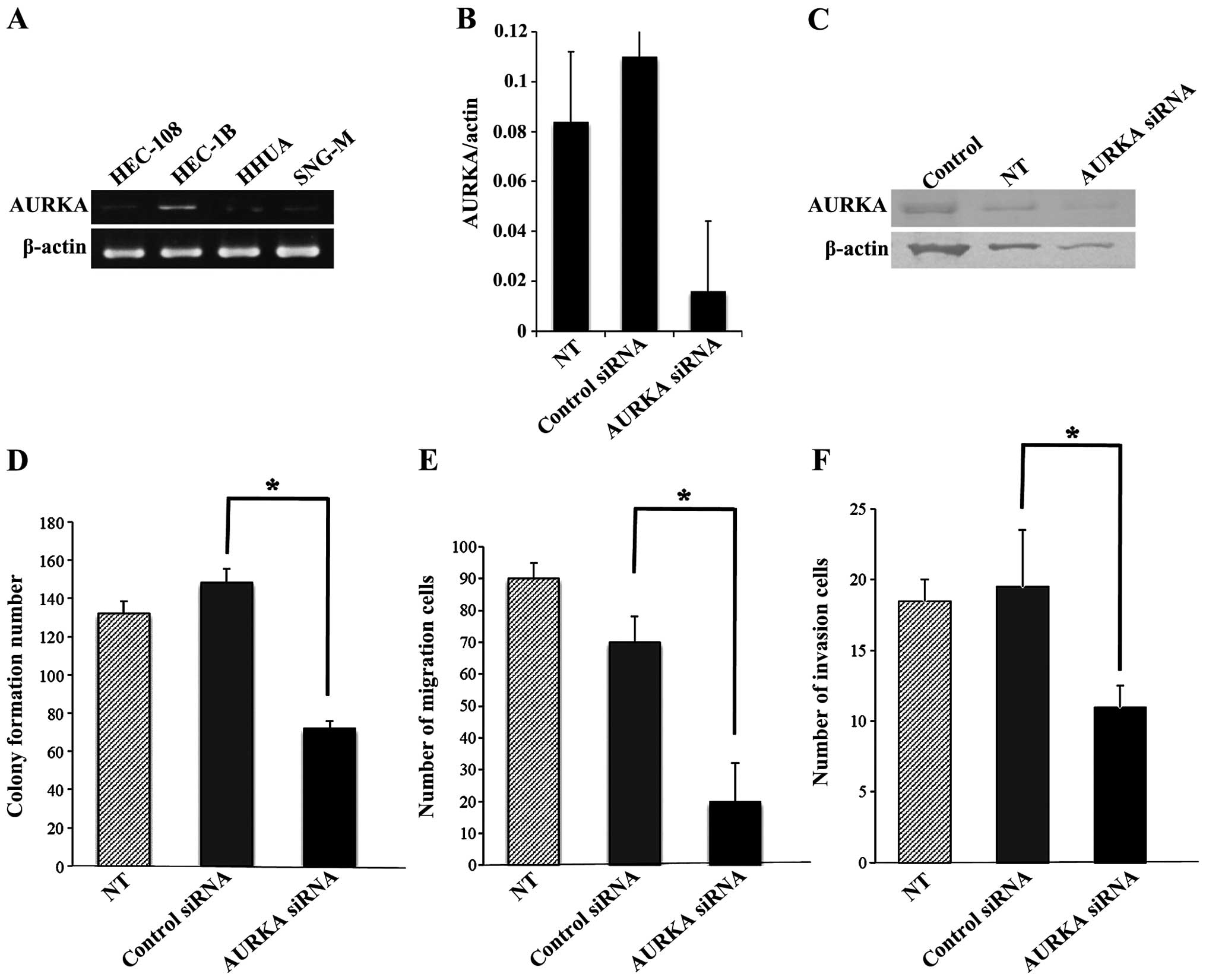
We screened endogenous AURKA expression in a panel
of four endometrial cancer cell lines: HEC-108
(poorly-differentiated), HEC-1B and HHUA
(moderately-differentiated), and SNG-M (well-differentiated).
RT-PCR showed that HEC-1B cells had a high level of AURKA mRNA
expression (Fig. 3A) and these
cells were selected for evaluation of the effects of AURKA
inhibition or overexpression. To determine whether AURKA has a role
in the behavior of endometrial cancer cells, in vitro
experiments were performed to analyze the effects of AURKA loss of
function on cell proliferation, invasion and migration.
siRNA-targeting AURKA produced efficient knockdown of AURKA in
HEC-1B cells, as shown by RT-PCR and western blot analysis: the
AURKA mRNA level decreased to 13% of that with scrambled control
siRNA (Fig. 3B) and the AURKA
protein level also decreased (Fig.
3C). Transfection with AURKA siRNAs significantly decreased
cell proliferation, invasion and migration of HEC-1B cells,
compared with transfection of control siRNA (Fig. 3D–F).
AURKA knockdown by siRNA enhances the
chemosensitivity of HEC-1B cells to paclitaxel
Recent reports showed that the upregulation of AURKA
contributes to chemoresistance in a human cell line, pancreatic
esophageal, breast and colon carcinoma cells (22–26).
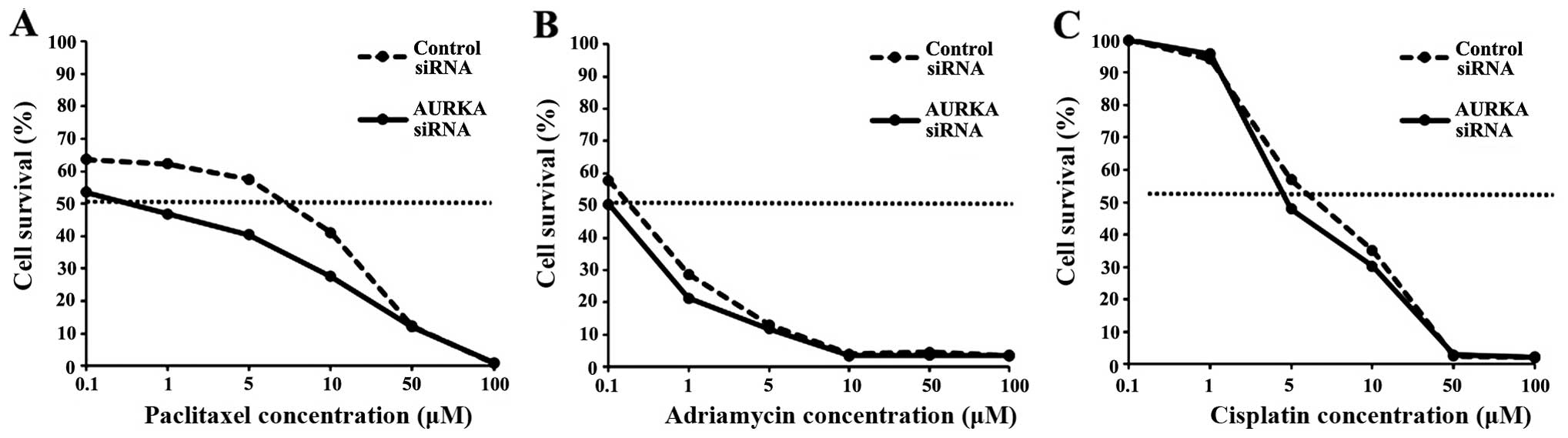
To determine whether AURKA knockdown had an effect on
chemosensitivity, a cytotoxicity assay was performed to measure
IC50 values of paclitaxel, adriamycin, and cisplatin,
which are widely used in gynecological cancer chemotherapy, before
and after AURKA knockdown in HEC-1B cells. Only the IC50
for paclitaxel changed after AURKA knockdown (Fig. 4, Table II), indicating that AURKA
expression is correlated with sensitivity to paclitaxel. These
results suggest that AURKA siRNA and paclitaxel in combination may
be effective for treatment of endometrial cancer.
| Table IIIC50 in HEC-1B cells with
or without AURKA siRNA. |
Table II
IC50 in HEC-1B cells with
or without AURKA siRNA.
| IC50
(μM) |
|---|
|
|
|---|
| Control siRNA | AURKA siRNA |
|---|
| Paclitaxel | 2.0 | 40.0 |
| Adriamycin | 0.20 | 0.10 |
| Cisplatin | 7.0 | 6.0 |
AURKA siRNA and paclitaxel in combination
enhances chemosensitivity in vivo
We analyzed antitumor activity in nude mice bearing
established HEC-1B tumors using treatment with control siRNA, AURKA
siRNA, paclitaxel, control siRNA and paclitaxel, and AURKA siRNA
and paclitaxel. In comparison with other treatments, the
combination of AURKA siRNA and paclitaxel showed a tendency to
inhibit tumor growth (P<0.1; Fig.
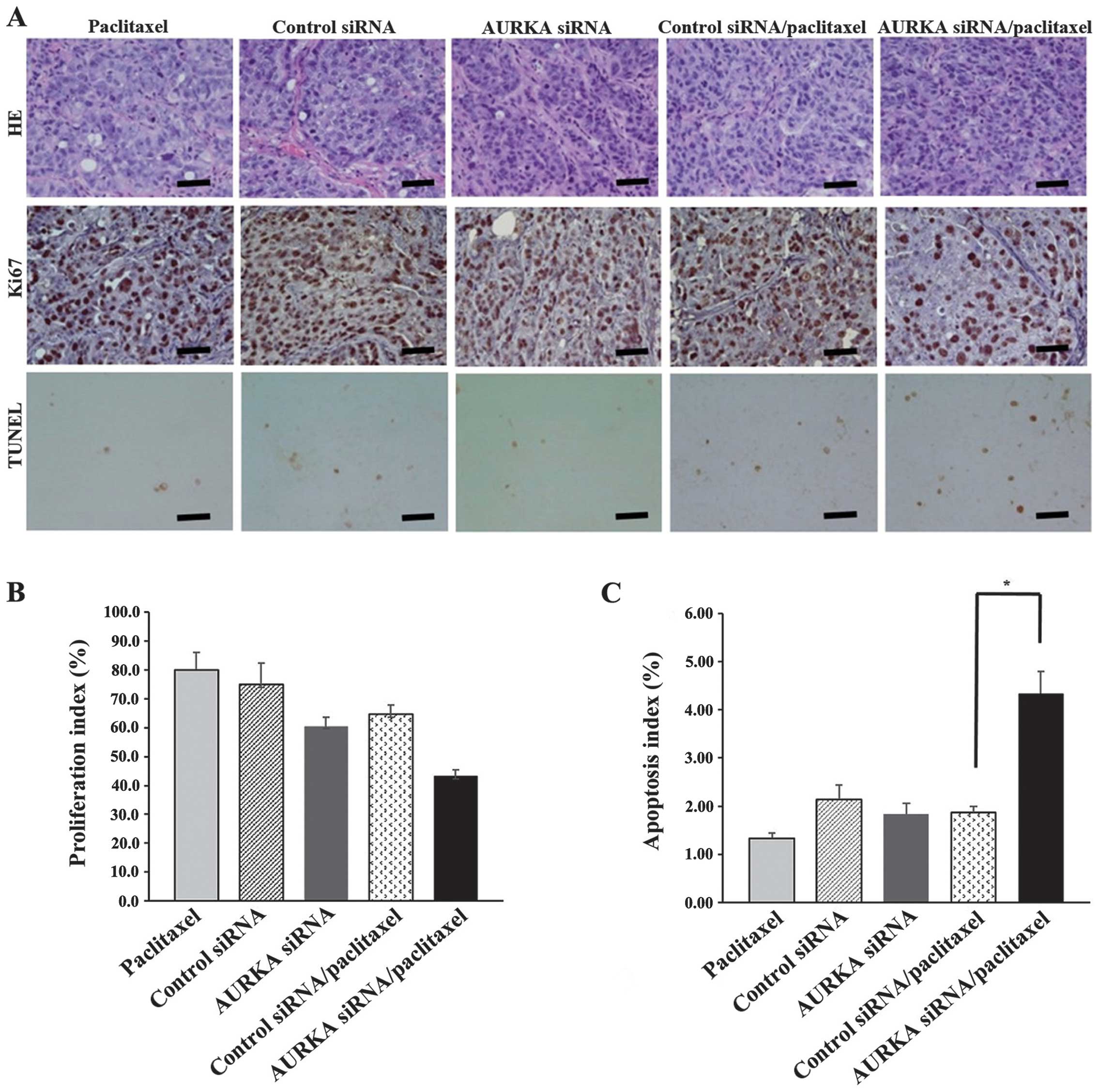
5). Immunohistochemical analysis of apoptosis (TUNEL analysis)
and proliferation (Ki67) after treatment (on day 28) showed that
the combination of AURKA siRNA and paclitaxel significantly
increased the number of TUNEL-positive cells (P<0.05) and showed
a trend for decreasing the number of Ki67-positive cells (Fig. 6A–C). Therefore, these in
vivo data indicate enhanced antitumor activity of AURKA siRNA
and paclitaxel combination in endometrial cancer.
Discussion
AURKA, also called STK15, is a serine/threonine
kinase that maintains cell division by regulating centrosome
separation, bipolar spindle assemble, and chromosome segregation
(5,6,27).
AURKA is also linked to the processes of G2-M arrest, apoptosis and
ectopic expression leading to bypass of G2-M in the DNA
damage-activated checkpoint system (9,28).
Aberrant amplification of AURKA occurs in human malignancies such
as breast (27), pancreatic
(12,29), colorectal (10), gastric (13), and ovarian carcinomas (27,30)
and in some cases is associated with a poor prognosis (31–33).
Although AURKA is a potential new oncogenic target, the role of
this protein in endometrial cancer is unclear.
In this study, we found an association of
overexpression of AURKA with clinicopathological factors in
endometrial cancer. Immunohistochemistry showed overexpression of
AURKA in endometrial cancer tissues compared with normal
endometrium, indicating that upregulation of AURKA is a frequent
abnormality in endometrial cancer. Overexpression of AURKA was
associated with tumor grade and histological type in endometrial
cancer tissues in χ2 tests and a tendency for this
association remained in logistic regression analysis, but this was
not significant (data not shown). Patients with overexpression of
AURKA also tended to have shorter DFS and a higher recurrence rate.
These results suggest that elevated AURKA tumor expression may be
an indicator of rapid cell division, rather than the cause of a
malignant phenotype.
Correlation of overexpression of AURKA with
malignant phenotypes has been shown in several cancers (11,12,34,35).
Amplification of AURKA causes chromosomal instability (6) that may help tumor cells acquire
invasive and metastatic phenotypes. Our results showed that
specific inhibition of AURKA by siRNA suppressed endometrial cancer
cell growth, migration and invasion. The mechanisms through which
AURKA influences cell migration and invasion are not completely
defined, but previous studies suggest roles for p53 (8,36,37),
RAS (38), AKT (39,40),
and MAPK (41). The level of p53
protein, which is a key player in this checkpoint, is increased in
AURKA-overexpressing cells and apoptosis is inhibited by deletion
of p53. Given that malignant tumor formation does not occur in a
mouse model with AURKA overexpression after a long latency period,
additional factors such as p53 inactivation and expression of
pro-survival proteins are likely to be required for tumorigenesis
(4). This hypothesis is supported
by the clinical observation that AURKA overexpression is correlated
with p53 mutation in hepatocellular carcinomas, and that tumors
with both AURKA overexpression and p53 mutation have a worse
prognosis than those with p53 mutation alone (42). In this study we showed that
overexpression of AURKA was significantly higher in
non-endometrioid adenocarcinoma and in grade 3 tumors that are
classified Type II tumors. From these findings, we speculated that
overexpression of AURKA may be associated with p53 mutation, and
caused poor prognosis in Type II tumors.
Several reports have shown that upregulation of
AURKA results in resistance to anticancer agents including
paclitaxel (18,22,43,44),
and docetaxel (23,44) in various cancers. Our in
vitro data showed that AURKA expression was correlated with
sensitivity to paclitaxel and our in vivo results suggested
that paclitaxel and AURKA siRNA in combination had significantly
enhanced antitumor efficacy. Taxanes such as paclitaxel bind to
microtubules and inhibit dissociation of tubulin subunits. This
inhibition of microtubule depolymerization by paclitaxel in tumor
cells prevents reconstruction of microtubules and formation of the
spindle, generating aberrant cell division. The spindle formation
checkpoint recognizes this abnormality and triggers apoptosis of
the tumor cell, causing the tumor to shrink (7,45,46).
Overexpression of Aurora kinases causes dysfunction of checkpoints
in cell division and permits the cell to enter anaphase in an
improper state (22). Thus, in the
presence of overexpressed Aurora kinases, taxane-based anticancer
agents cannot induce apoptosis of aberrant cells and have reduced
sensitivity. Conversely, drugs that inhibit Aurora kinases may
suppress resistance to apoptosis induced by taxanes and enhance
antitumor action. For this reason, several small-molecule Aurora
kinase inhibitors have been developed that exhibit preclinical
activity against various solid tumors. These include MLN8237,
Hesperadin, VX-680, VE465 and Barasertib, and clinical trials are
ongoing to verify the effects of these inhibitors (47). Combination therapy of paclitaxel
and AURKA inhibition using siRNA or an AURKA inhibitor may also
allow reduction of the dose of paclitaxel, with the result of fewer
side-effects.
In summary, our data on endometrial carcinoma show
that overexpression of AURKA is strongly associated with tumor
grade and histological type, and that there is a correlation
between expression of AURKA and sensitivity to paclitaxel. These
results suggest that AURKA may be a biomarker for identification of
a subgroup of patients with resistance to treatment and a poor
prognosis, and a promising target for novel therapeutics for
endometrial cancer. Combination treatment using AURKA inhibitors
and paclitaxel may be particularly effective for cases of
endometrial cancer that are resistant to conventional
treatment.
Acknowledgements
This study was partially supported by a Keio
University Grant-in-Aid for Encouragement of Young Medical
Scientists; the Japan Society for the Promotion of Science (JSPS)
through a Grant-in-Aid for Scientific Research (KAKENHI), a
Grant-in-Aid for Scientific Research (B) (22390313) and (C)
(22591866), as well as a Grant-in-Aid for Young Scientists (B)
(24791718); the Ichiro Kanehara Foundation (Tokyo, Japan);
Kobayashi Foundation for Cancer Research (Tokyo, Japan); and the
Keio University Medical Science Fund through a Research Grant for
Life Sciences and Medicine.
References
|
1
|
Ferlay J, Shin HR, Bray F, et al: GLOBOCAN
2008 v20, Cancer Incidence and Mortality Worldwide. International
Agency for Research on Cancer; Lyon: 2010, http://globocan.iarc.fr.
|
|
2
|
Murali R, Soslow RA and Weigelt B:
Classification of endometrial carcinoma: more than two types.
Lancet Oncol. 15:e268–e278. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bokhman JV: Two pathogenetic types of
endometrial carcinoma. Gynecol Oncol. 15:10–17. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marumoto T, Zhang D and Saya H: Aurora-A -
a guardian of poles. Nat Rev Cancer. 5:42–50. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Glover DM, Leibowitz MH, McLean DA and
Parry H: Mutations in aurora prevent centrosome separation leading
to the formation of monopolar spindles. Cell. 81:95–105. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou H, Kuang J, Zhong L, et al: Tumour
amplified kinase STK15/BTAK induces centrosome amplification,
aneuploidy and transformation. Nat Genet. 20:189–193. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Katayama H, Brinkley WR and Sen S: The
Aurora kinases: role in cell transformation and tumorigenesis.
Cancer Metastasis Rev. 22:451–464. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Katayama H, Sasai K, Kawai H, et al:
Phosphorylation by aurora kinase A induces Mdm2-mediated
destabilization and inhibition of p53. Nat Genet. 36:55–62. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cazales M, Schmitt E, Montembault E,
Dozier C, Prigent C and Ducommun B: CDC25B phosphorylation by
Aurora-A occurs at the G2/M transition and is inhibited by DNA
damage. Cell Cycle. 4:1233–1238. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nishida N, Nagasaka T, Kashiwagi K, Boland
CR and Goel A: High copy amplification of the Aurora-A gene is
associated with chromosomal instability phenotype in human
colorectal cancers. Cancer Biol Ther. 6:525–533. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sen S, Zhou H, Zhang RD, et al:
Amplification/overexpression of a mitotic kinase gene in human
bladder cancer. J Natl Cancer Inst. 94:1320–1329. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li D, Zhu J, Firozi PF, et al:
Overexpression of oncogenic STK15/BTAK/Aurora A kinase in human
pancreatic cancer. Clin Cancer Res. 9:991–997. 2003.PubMed/NCBI
|
|
13
|
Sakakura C, Hagiwara A, Yasuoka R, et al:
Tumour-amplified kinase BTAK is amplified and overexpressed in
gastric cancers with possible involvement in aneuploid formation.
Br J Cancer. 84:824–831. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sen S, Zhou H and White RA: A putative
serine/threonine kinase encoding gene BTAK on chromosome 20q13 is
amplified and overexpressed in human breast cancer cell lines.
Oncogene. 14:2195–2200. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Landen CN Jr, Lin YG, Immaneni A, et al:
Overexpression of the centrosomal protein Aurora-A kinase is
associated with poor prognosis in epithelial ovarian cancer
patients. Clin Cancer Res. 13:4098–4104. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lassmann S, Shen Y, Jütting U, et al:
Predictive value of Aurora-A/STK15 expression for late stage
epithelial ovarian cancer patients treated by adjuvant
chemotherapy. Clin Cancer Res. 13:4083–4091. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Umene K, Banno K, Kisu I, et al: Aurora
kinase inhibitors: potential molecular-targeted drugs for
gynecologic malignant tumors. Biomed Rep. 1:335–340. 2013.
|
|
18
|
Do TV, Xiao F, Bickel LE, et al: Aurora
kinase A mediates epithelial ovarian cancer cell migration and
adhesion. Oncogene. 33:539–549. 2014. View Article : Google Scholar
|
|
19
|
Kurai M, Shiozawa T, Shih HC, et al:
Expression of Aurora kinases A and B in normal, hyperplastic, and
malignant human endometrium: Aurora B as a predictor for poor
prognosis in endometrial carcinoma. Hum Pathol. 36:1281–1288. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moreno-Bueno G, Sánchez-Estévez C, Cassia
R, et al: Differential gene expression profile in endometrioid and
nonendometrioid endometrial carcinoma: STK15 is frequently
overexpressed and amplified in nonendometrioid carcinomas. Cancer
Res. 63:5697–5702. 2003.PubMed/NCBI
|
|
21
|
Tanaka E, Hashimoto Y, Ito T, et al: The
clinical significance of Aurora-A/STK15/BTAK expression in human
esophageal squamous cell carcinoma. Clin Cancer Res. 11:1827–1834.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Anand S, Penrhyn-Lowe S and Venkitaraman
AR: AURORA-A amplification overrides the mitotic spindle assembly
checkpoint, inducing resistance to Taxol. Cancer Cell. 3:51–62.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tanaka E, Hashimoto Y, Ito T, et al: The
suppression of aurora-A/STK15/BTAK expression enhances
chemosensitivity to docetaxel in human esophageal squamous cell
carcinoma. Clin Cancer Res. 13:1331–1340. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Y, Tang K, Zhang H, Zhang Y, Zhou W and
Chen X: Function of Aurora kinase A in Taxol-resistant breast
cancer and its correlation with P-gp. Mol Med Rep. 4:739–746.
2011.PubMed/NCBI
|
|
25
|
Cammareri P, Scopelliti A, Todaro M, et
al: Aurora-a is essential for the tumorigenic capacity and
chemoresistance of colorectal cancer stem cells. Cancer Res.
70:4655–4665. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hata T, Furukawa T, Sunamura M, et al: RNA
interference targeting aurora kinase a suppresses tumor growth and
enhances the taxane chemosensitivity in human pancreatic cancer
cells. Cancer Res. 65:2899–2905. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang F, Guo X, Yang G, Rosen DG and Liu J:
AURKA and BRCA2 expression highly correlate with prognosis of
endometrioid ovarian carcinoma. Mod Pathol. 24:836–845. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Du J and Hannon GJ: Suppression of
p160ROCK bypasses cell cycle arrest after Aurora-A/STK15 depletion.
Proc Natl Acad Sci USA. 101:8975–8980. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Furukawa T, Kanai N, Shiwaku HO, Soga N,
Uehara A and Horii A: AURKA is one of the downstream targets of
MAPK1/ERK2 in pancreatic cancer. Oncogene. 25:4831–4839. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang G, Chang B, Yang F, et al: Aurora
kinase A promotes ovarian tumorigenesis through dysregulation of
the cell cycle and suppression of BRCA2. Clin Cancer Res.
16:3171–3181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Goos JA, Coupe VM, Diosdado B, et al:
Aurora kinase A (AURKA) expression in colorectal cancer liver
metastasis is associated with poor prognosis. Br J Cancer.
109:2445–2452. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yamamoto S, Yamamoto-Ibusuki M, Yamamoto
Y, Fujiwara S and Iwase H: A comprehensive analysis of Aurora A;
transcript levels are the most reliable in association with
proliferation and prognosis in breast cancer. BMC Cancer.
13:2172013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yen CC, Yeh CN, Cheng CT, et al:
Integrating bioinformatics and clinicopathological research of
gastrointestinal stromal tumors: identification of Aurora kinase A
as a poor risk marker. Ann Surg Oncol. 19:3491–3499. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Royce ME, Xia W, Sahin AA, et al:
STK15/Aurora-A expression in primary breast tumors is correlated
with nuclear grade but not with prognosis. Cancer. 100:12–19. 2004.
View Article : Google Scholar
|
|
35
|
Hamada M, Yakushijin Y, Ohtsuka M,
Kakimoto M, Yasukawa M and Fujita S: Aurora2/BTAK/STK15 is involved
in cell cycle checkpoint and cell survival of aggressive
non-Hodgkin’s lymphoma. Br J Haematol. 121:439–447. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen SS, Chang PC, Cheng YW, Tang FM and
Lin YS: Suppression of the STK15 oncogenic activity requires a
transactivation-independent p53 function. EMBO J. 21:4491–4499.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Q, Kaneko S, Yang L, et al: Aurora-A
abrogation of p53 DNA binding and transactivation activity by
phosphorylation of serine 215. J Biol Chem. 279:52175–52182. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tatsuka M, Sato S, Kitajima S, et al:
Overexpression of Aurora-A potentiates HRAS-mediated oncogenic
transformation and is implicated in oral carcinogenesis. Oncogene.
24:1122–1127. 2005. View Article : Google Scholar
|
|
39
|
Guan Z, Wang XR, Zhu XF, et al: Aurora-A,
a negative prognostic marker, increases migration and decreases
radiosensitivity in cancer cells. Cancer Res. 67:10436–10444. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang H, He L, Kruk P, Nicosia SV and Cheng
JQ: Aurora-A induces cell survival and chemoresistance by
activation of Akt through a p53-dependent manner in ovarian cancer
cells. Int J Cancer. 119:2304–2312. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wan XB, Long ZJ, Yan M, et al: Inhibition
of Aurora-A suppresses epithelial-mesenchymal transition and
invasion by downregulating MAPK in nasopharyngeal carcinoma cells.
Carcinogenesis. 29:1930–1937. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jeng YM, Peng SY, Lin CY and Hsu HC:
Overexpression and amplification of Aurora-A in hepatocellular
carcinoma. Clin Cancer Res. 10:2065–2071. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin Y, Richards FM, Krippendorff BF, et
al: Paclitaxel and CYC3, an Aurora kinase A inhibitor, synergise in
pancreatic cancer cells but not bone marrow precursor cells. Br J
Cancer. 107:1692–1701. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Long M, Yin G, Liu L, et al:
Adenovirus-mediated Aurora A shRNA driven by stathmin promoter
suppressed tumor growth and enhanced paclitaxel chemotherapy
sensitivity in human breast carcinoma cells. Cancer Gene Ther.
19:271–281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee HH, Zhu Y, Govindasamy KM and Gopalan
G: Down-regulation of Aurora-A overrides estrogen-mediated growth
and chemoresistance in breast cancer cells. Endocr Relat Cancer.
15:765–775. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Agnese V, Bazan V, Fiorentino FP, et al:
The role of Aurora-A inhibitors in cancer therapy. Ann Oncol.
18(Suppl 6): vi47–vi52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gautschi O, Heighway J, Mack PC, Purnell
PR, Lara PN Jr and Gandara DR: Aurora kinases as anticancer drug
targets. Clin Cancer Res. 14:1639–1648. 2008. View Article : Google Scholar : PubMed/NCBI
|