1. Introduction
Breast cancer (BC) is the most frequently occurring
malignant disease among women worldwide, accounting for nearly one
fourth of the total new cancer cases (1). Despite marked progress in therapeutic
strategies advanced BC remains a deadly disease, prompting efforts
to advance our understanding of BC biology, with the aim of
developing innovative approaches. Substantial research has been,
and are conducted in the field of cross-talk between cancer cells
and their microenvironment (2).
Several tumor cell and stromal cell-derived factors have been
identified and involved in these complex interactions, among them
the macrophage migration inhibitory factor (MIF) (3). MIF is an evolutionarily highly
conserved pleiotropic chemokine identified in 1966 by Bloom and
Bennett as a protein secreted by T cells and inhibiting the
migration of macrophages. Subsequently, it was shown that this
chemokine is not only secreted by lymphocytes but also by other
immune cells such as macrophages and non-immune cells such as
endothelial and epithelial cells (4). In 1994, Paralkar and Wistow described
the MIF gene, which is a small gene of <1 Kb, localized
on chromosome 22. MIF is a protein of 115 amino acids, with a
molecular weight of 12.5 kDa, in which the active form is a trimer
(5). MIF has an enzymatic activity
called tautomerase, situated within an N-terminal proline (Pro1),
which catalyzed the tautomerization of
p-hydroxyphenylpyruvate and D-dopachrome. Although, highly
conserved this enzymatic activity has no natural substrate
identified and its contribution to MIF biological properties is
uncertain (6). MIF is stored in
the intracellular compartment and released upon stimulation via
unconventional pathway, where the Golgi-associated protein p115
could be involved (7).
The discovery that MIF was released from
corticotrophic pituitary cells and override the anti-inflammatory
effects of corticoids suggested a pivotal role in inflammatory
diseases. Its involvement was later described in sepsis, autoimmune
diseases such as rheumatoid arthritis and systemic lupus
erythematous. It plays a master role in regulating the complex
formation of atherogenesis and is elevated in serum of patients
with metabolic syndrome (8,9).
There is growing evidence that MIF is involved in carcinogenesis.
MIF is often overexpressed in tissues and cancer cells. Indeed,
Kindt et al (10) showed
that MIF expression increased during the tumor progression of head
and neck cancer but also in hepatocellular carcinoma (11), ovarian cancer (12) and other malignancies (13,14).
Currently, there is a general consensus that MIF promotes tumor
growth by several mechanisms by acting on the cancer cells
themselves but also on the neighboring or even distant cancerous
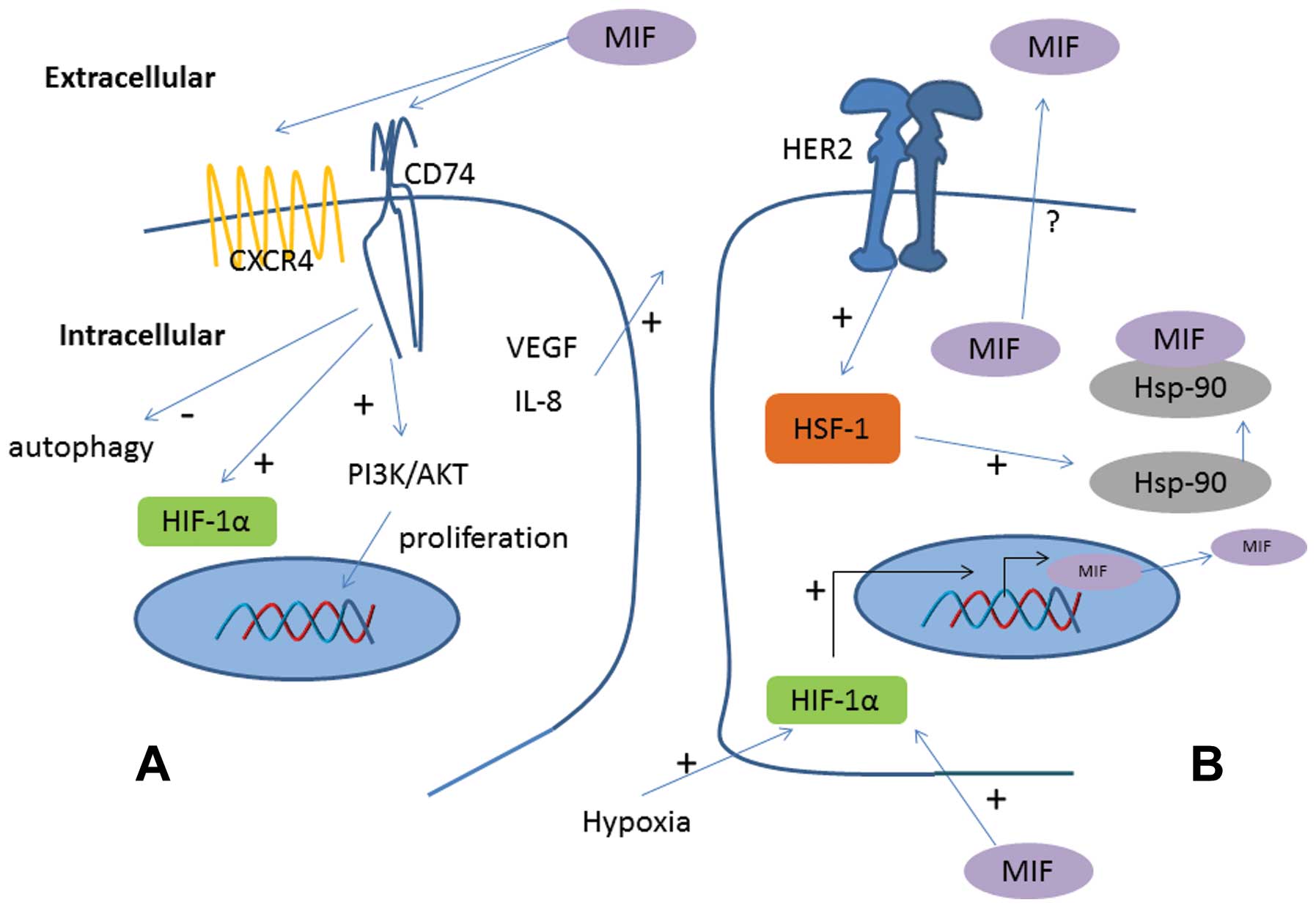
and non-cancerous cells. MIF interacts with its receptor, CD74 to
induce the activation of several pathways such as MAPK and
PI3K/Akt. This interaction plays a role in cell proliferation via
CD44 as describe by Shi et al (15) in 2006. MIF could also interact with
CXCR4 to promote cell migration (16). MIF inhibits induction of
p53-dependent apoptosis, increases production of vascular
endothelial growth factor (VEGF) and inhibits the antitumor immune
response (4,17). In colorectal cancer cell lines MIF
modulates the expression of cyclooxygenase-2 (COX-2), an important
contributor to carcinogenesis (18). In the C4-2b prostate cancer cell
line, NS-398 a COX-2 inhibitor increases MIF expression by NF-κB
transcription factor leading to a more aggressive cell
differentiation (19). In
esophageal squamous cell carcinoma, aspirin and NS-398
significantly inhibit MIF expression (20). Notably, elevated COX-2 expression
in breast cancer is associated with a poor prognosis (21) and a meta-analysis of clinical
studies suggests a slightly protective effect of COX-2 inhibitors
against breast cancer (22).
Finally, MIF modulates metastatic behavior of tumor cells and
affects tumor stromal cells in a complex manner (3,23,24).
Here, we review the clinical and experimental
published data on MIF in breast cancer and discuss the potential
clinical implications.
2. MIF and breast cancer: experimental
studies
MIF and its receptor complex CD74/CD44
and CXCR4
In 2009, Verjans et al (25) studied the expression of MIF in
invasive and non-invasive breast cancer cell lines. Non-invasive
ductal breast cancer cell lines (MDA-MB-468 and ZR-75-1) showed an
upregulation of MIF compared to benign epithelial breast cells
(MCF-12A). Surprisingly, MIF was less expressed in invasive cells
(MDA-MB-231). They observed no constitutive MIF release in
non-cancerous MCF-12A cells contrary to the cancerous cell lines
MDA-MB-468 and MDA-MB-231. They next studied the effect of
exogenous rMIF on the secretion rate of endogenous MIF by cancerous
and non-cancerous cells and showed an obvious upregulation, the
most dramatic effect was seen in MDA-MB-231 cells, suggesting that
MIF secretion could be influenced by auto- and paracrine MIF in the
tumor microenvironment (25). The
mechanisms of MIF overexpression are not well known, but Schulz
et al (26) suggested that
one of them could be the result of MIF protein stabilization by the
tumor-activated HSP90 chaperone complex. Moreover, MIF gene has
been shown to be upregulated by hypoxia in human breast cancer
cells MCF-7 (27). However, Larsen
et al (28) suggested that
hypoxia MIF upregulation in MCF-7 is dependent on the cell genotype
and that its secretion from hypoxic MCF-7 cells is regulated at the
level of transcription independently of HIF. CD74, the main MIF
receptor, is overexpressed, but not prominently, in invasive
MDA-MB-231 cells, compared with non-cancerous MCF-12A cells
(25). CXCR-4, the receptor of
chemokine CXCL-12, can also bind to MIF. Indeed, JNK activation
occurred when MIF interacts with CXCR-4 in Jurkat T cells (29). In breast cancer cell line, CXCR-4
expression is high in MDA-MB-231 aggressive cells and low in less
aggressive MCF-7 cells (30).
These in vitro data confirmed the involvement of the MIF
complex in the carcinogenesis of breast cancer.
MIF promotes cell proliferation
Lue et al (31) showed on various breast cancer cell
lines that the interaction of MIF with its membrane receptor CD74
induced activation of PI3K/Akt signaling pathway, promoting cell
survival. They also found stronger MIF expression in breast cancer
cell lines MCF-7 and ZR 75-1 compared to normal breast cell line
MCF-12A but also in MDA-MB-468 cancer cells. They correlated MIF
expression to Akt phosphorylation ratio and studied the response of
these cell lines to exogenous rMIF. Breast cancer cells are
unresponsive to exogenous rMIF when the Akt pathway is maximally
activated or functionally blocked and when endogenous MIF is
already high (31). Verjans et
al (25) have also showed that
recombinant MIF stimulates the proliferation of non-invasive and
invasive cancer cell lines. Moreover, Simpson et al
(32) demonstrated that the
depletion of MIF has no impact on the growth of cancer cells in
vitro. Lim et al (33)
studied the regulation of MIF by epidermal growth factor receptor
(EGFR) in the MCF10AT model of breast cancer progression. After EGF
stimulation the level of MIF expression decreased in normal breast
mammary epithelial cells but increased in MCF10AT1k preneoplastic
and MCF10CA1h low-grade breast cancer cells and was unchanged in
high grade cancer cells. Inversely, the silencing of MIF decreased
EGF-stimulated CA1h cell proliferation. Further to these results,
they proposed a proliferative pathway linking EGFR stimulating MEK
and finally MIF (33).
MIF and microenvironment modulation
MIF as an immunomodulator
In the 4T1 murine model of breast cancer, Simpson
et al (32) showed that MIF
promotes tumor growth and metastasis by increasing the prevalence
of a highly immune suppressive subpopulation of myeloid-derived
suppressor cells (MDSCs) within the tumor. These results are in
line with the study of Drews-Elger et al (34) demonstrating that, in pre-clinical
models, the systemic recruitment of S100A8+ myeloid
cells (including MDSCs) is promoted by MIF among others
tumor-derived factors, S100A8 being a pro-inflammatory mediator
secreted by MDSC. In vitro, MIF knockdown does not alter the
migration ability and growth rate of MDA-MB-231 cells but, in
vivo, depletion of MIF decreased primary tumor growth and
metastatic burden in the lungs, correlated with a reduction of the
number of S100A8+ infiltrating cells. They suggested
that MIF produced by the tumor promotes tumor progression by
recruiting S100A8+ cells acting as potent
immunosuppressors and driving invasion, migration and angiogenesis
(34).
MIF and neo-angiogenesis
In 2008, Xu et al (35) demonstrated not only that the MIF
expression increased in cancerous cell lines compared to normal
epithelial cells but also that addition of exogenous MIF in cell
culture induced a dose-dependent increase in VEGF and IL-8
secretion, two factors implicated in angiogenesis. Other studies
have showed that MIF was linked to hypoxia. Indeed, MCF-7 cells,
under hypoxia condition, present an upregulation of MIF mRNA after
16 and 24 h compare to normoxic condition (28). The effect of MIF on HIF-1 activity
was investigated in human breast cancer MCF-7 and MDA-MB-231 cells
by Oda et al (36). They
demonstrated that intracellular overexpression or extracellular
administration of MIF enhances activation of HIF-1 under hypoxic
conditions in MCF-7 cells, involving the CD74 receptor.
MIF and migration
Martinez et al (24) demonstrated in their study that
peripheral blood plasma from breast cancer patients induced a
higher transendothelial migration of MDA-MB-231 and MCF-7 cells
than plasma from healthy volunteers. They also found significantly
higher MDA-MB-231 cell proliferation. These results could be linked
to the higher levels of MIF and others cytokines in peripheral
blood plasma of breast cancer patients compared to the healthy
volunteers.
MIF and autophagy
Autophagy is a process that maintains cellular
homeostasis under stress conditions. The microtubule-associated
protein 1 light chain 3 (LC3-II) is proportional to the number of
autophagic vacuole formation and its expression level can be used
to quantify autophagy. In their study, Wu et al (37), demonstrated that the diminution of
MIF expression by siRNA induce increase of LC3-II expression. In
the same study, they showed that SRC-3 (steroid receptor
coactivator 3), an oncogene that favors drug resistance in cancer,
targets MIF. Indeed, knockdown of SRC-3 in MCF-7 cells induces a
decrease of MIF expression. This was reversed by exogenous
recombinant MIF. They suggested that MIF could inhibit autophagy in
breast cancer cells.
MIF and HER2
In ErbB2 transgenic model of human HER2-positive
breast cancer mice, Schultz et al (38) demonstrated that genetic MIF
deletion protected from the development of breast cancer and led to
extended survival, suggesting a link between HER2 status and MIF.
ErbB2 cancers in MIF+/+ mice showed overexpression of
MIF in malignant breast epithelium. Delayed tumor progression in
MIF−/− ErbB2 mice seemed to be linked to slower tumor
growth with decreased proliferation as showed by lower Ki-67
staining. They later showed that specific inhibition of HER2
signaling in HER2 overexpressing breast cancer cells reduces MIF
levels by abolishing its stabilization. Indeed, overexpressed HER2
activates HSF1 (heat-shock factor1), transcriptional regulator of
HSP90 which stabilizes MIF (26).
MIF and drug resistance
Wu et al (37) showed that autophagic cell death
induced by MIF knockdown increased the chemo-sensitivity of breast
cancer cells to doxorubicin, apoptosis being not affected. The aza
derivatives of resveratrol, a natural phytoalexin, showed
anti-proliferative effect in the MCF-7 cell line and inhibits the
MIF tautomerase activity (39). It
was also shown to reverse partially the paclitaxel resistance in
MDA-MB-231/PacR cell lines (40).
Based on these data, we suggest a schematic model of
MIF involvement in breast cancer cells (Fig. 1).
3. MIF and breast cancer: clinical
studies
MIF serum levels in human breast cancer
patients
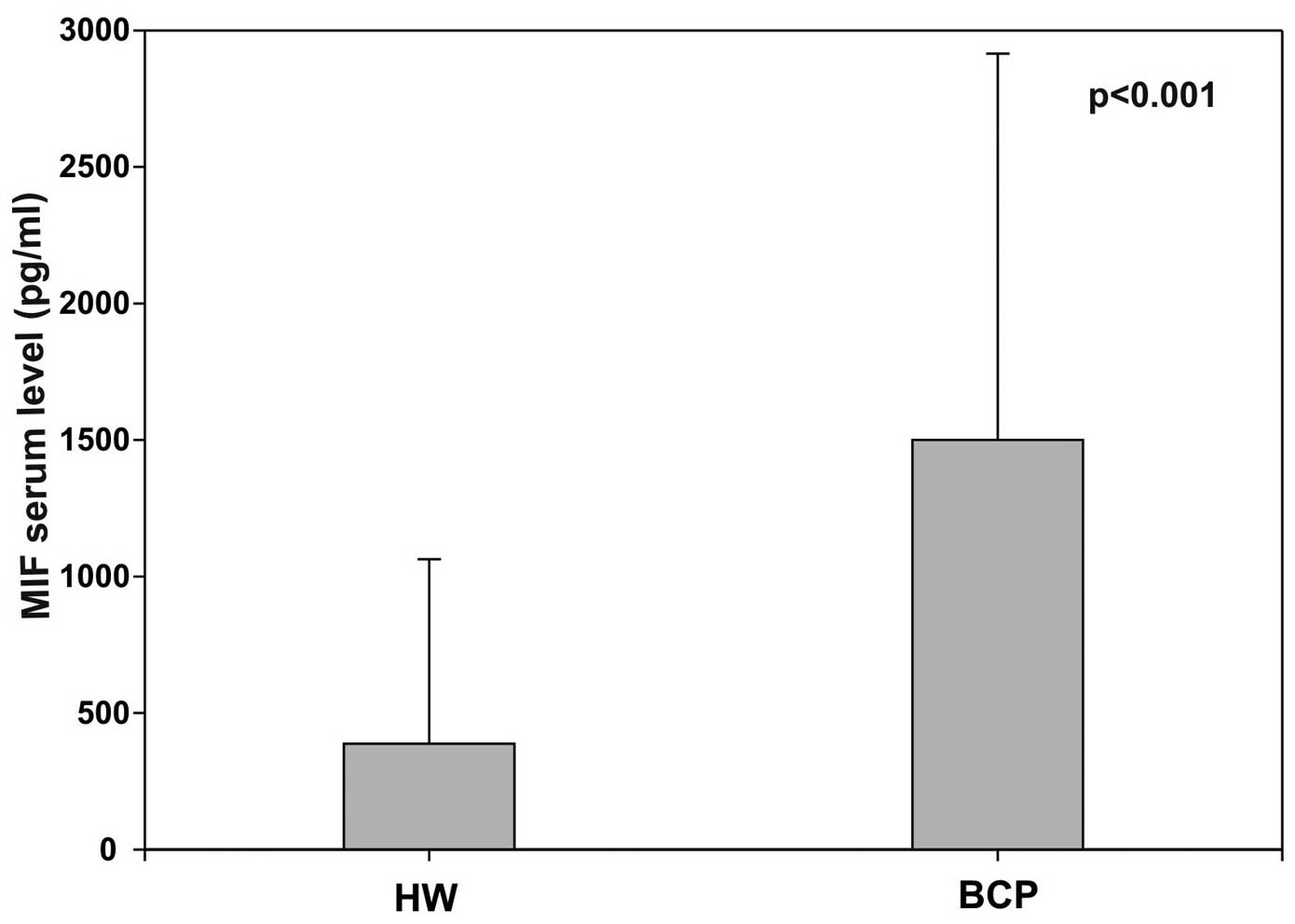
Five studies evaluated MIF serum levels in breast
cancer patients (BCP) compared to healthy volunteers (HV): all
showed significant increased levels in BCP (24,35,41–43).
Our data are showed in Fig. 2
(42). Using a set of serum
proteins, two others studies found a non-diagnostic MIF serum
elevation in BCP (44,45). Correlations with invasive tumor
characteristics and patients prognostic are poorly documented:
Bando et al (27) found no
significant correlation with MIF intratumoral levels and
clinico-pathological factors but a significant inverse correlation
with nodal status. In the study of Fersching et al (43) pre-therapeutic levels of MIF tended
to be higher in non-responsive patients undergoing preoperative
chemotherapy. No data are available concerning MIF serum levels and
tumor recurrence. These sets of data suggest that the increased
serum level of MIF in BCP could be a non-specific signature of a
systemic response to breast cancer.
MIF and CD74 expression in breast cancer
tissue
MIF expression in breast cancer tissue has been
evaluated in five studies, using immunohistochemical techniques in
four, with different scoring scales, and western blot analysis and
RT-PCR in one (25,35,41,42,46).
All found a high level of MIF expression in the tumor tissue. Xu
et al (35) showed higher
microvessels density in tumors with higher MIF expression.
Correlations with classical tumor characteristics are elusive
(Table I). MIF expression level
was evaluated in glandular compartment only except in our study
where we have evaluated stromal and glandular compartments
separately. Bando et al (41) found an inverse correlation with
nodal involvement, Verjans et al (25) a positive correlation with
progesterone and estrogen receptor expression. A negative
correlation between MIF expression and tumor size was shown by
Verjans. MIF expression significantly correlated with HER-2 for Xu
et al (35). Choi et
al (46) confirmed this result
in a study evaluating the expression of a set of proteins in
invasive breast cancer. These observations could be put in
perspective with the experimental data describe earlier (MIF and
HER2).
 | Table IMIF tumor tissue expression level and
histoprognostic factors. |
Table I
MIF tumor tissue expression level and
histoprognostic factors.
| Réfs. | No. of
patients | Tumor size | Lymph node
status | Histological
grade | PR/ER
expression | HER2 status | TRN status | Ki-67 level |
|---|
| Bando et al
(41) | 93 | NS | 0.039
(inverse) | - | NS | - | - | - |
| Xu et al
(35) | 121 | NS | 0.063 | NS | - | 0.03 | - | - |
| Verjans et
al (25) | 175 | 0.007
(inverse) | NS | NS | 0.006
0.028 | - | - | - |
| Choi et al
(46) | 276 | NS | NS | NS | NS | <0.001 | - | - |
| Richard et
al (42) | 96 | 0.02a (inverse) | NS | NS | NS | NS | NS | NS |
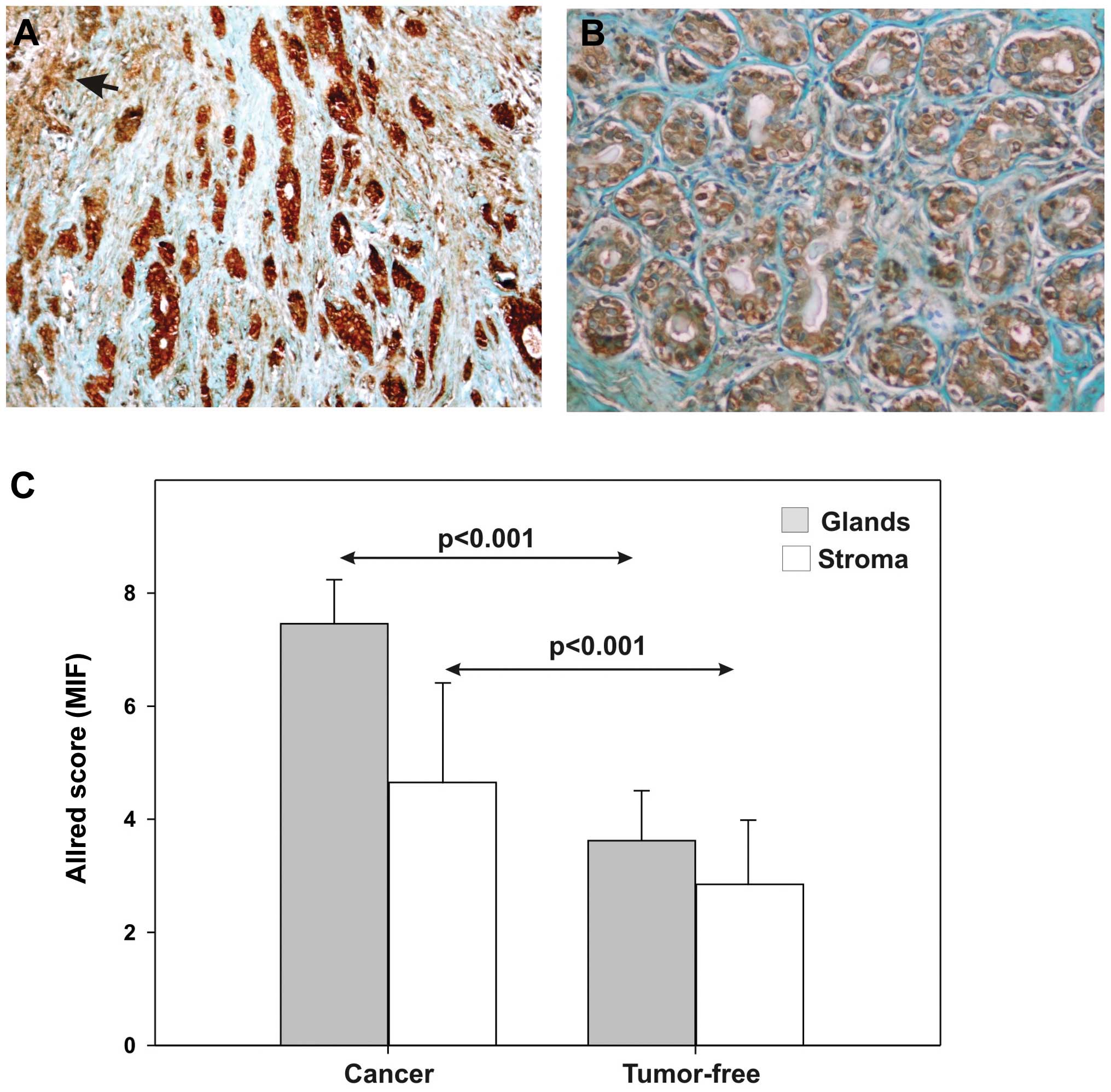
Our study is the only one evaluating the expression
of MIF and its receptor CD74 on the same breast cancer biopsies and
normal breast tissue (42). MIF
tissue expression, quantified by a modified Allred score, was
strongly increased in carcinoma compared to tumor-free specimens,
in the cancer cells and in the peritumoral stroma, with fibroblasts
the most intensely stained (Fig.
3). We did not find any significant correlation with
histoprognostic factors, except for a significant inverse
correlation between tumor size and MIF stromal positivity. These
findings have to be put in perspective with the hypothesis that MIF
could modulate the tumor size by inhibiting recruitment of
cancer-associated fibroblasts (CAFs)/myofibroblasts (23) and several recent publications
underscoring the pivotal role of CAFs in tumor progression
(3,47,48,49).
CD74 staining was heterogeneous and significantly decreased in
cancer cells but increased in the surrounding stroma, namely in
lymphocytes, macrophages and vessel endothelium. There was no
significant variation according to classical histoprognostic
factors, except that CD74 stromal expression was significantly
correlated with triple-negative receptor (TRN) status and the
absence of estrogen receptors (42). This finding could be compared with
the growing importance of evaluating stromal lymphocytes
infiltration of the TRN tumor (50).
Only Xu et al (25) and Verjans et al (35) have
reported data concerning MIF expression levels and patients
prognosis. The results are discordant with positive MIF expression
correlating with worse disease-free survival for Xu et al,
but a better overall and recurrence-free survival for Verjans et
al. However, MIF scoring scales and cut-off for positivity were
not the same. These data show an increased expression of MIF in
breast cancer tissue and suggest that this cytokine contribute to
shape and modulate the tumor aggressivity.
We did not find any published data concerning MIF
and CD74 expression in non-invasive breast cancer tissue.
4. Conclusions and perspectives
MIF is a pleiotropic chemokine involved in
carcinogenesis. Experimental and clinical studies suggest that MIF
could have a multi-functional role in human breast cancer. Firstly,
MIF has auto- and paracrine effects on cancer cells promoting
proliferation, migration and inhibiting autophagy and apoptosis.
Secondly, MIF contributes to shape the immune and non-immune tumor
microenvironment leading to immunomodulation and angiogenesis
promoting cancer cells growth, invasion and metastases. Thirdly,
MIF acts systemically leading to metabolic disturbances such as
metabolic syndrome and negative immune implications promoting tumor
growth and metastases development. Even if many questions remain
unresolved regarding the mechanisms of cell modulation by MIF
inside the primary or metastatic tumor, therapeutic targeting of
this cytokine could provide clinical benefits. Several experimental
data have been published exploring MIF pathway inhibition. HSP90
stabilizes MIF and its pharmacologic inhibition by
17-N-allylamino-17-demethoxygeldanamycin (17-AAG, Tanespimycin) or
suberoylanilide-hydroxamic-acid (SAHA, vorinostat) destabilizes MIF
protein leading to death of human breast cancer cells lines. This
is reversed by excess ectopic MIF. In an ErbB2 transgenic model of
human HER2-positive breast cancer, systemic administration of 17AAG
reduces MIF expression and inhibits growth of MIF-expressing, but
not MIF-deficient tumors (40).
Ganetespib, an HSP90 inhibitor has been recently tested with
success in metastatic HER2 breast cancer in combination with
paclitaxel (51). Part of the
activity of these molecules could be linked to MIF inactivation.
Human anti-MIF antibodies, low molecular weight inhibitors of the
tautomerase activity of MIF protein, blocking agents of MIF
receptors have been tested in diverse human cancer cell lines with
some positive results but we did not find any specific breast
cancer data. A phase I trial with anti-MIF antibody is being tested
in advanced solid tumor (52).
Anti-CD74 strategy with milatuzumab has been successfully
investigated in hematologic malignancies (53). MIF being a potent endogenous
mediator of COX-2 expression and this enzyme being linked to breast
cancer progression, anti COX-2 molecules could be tested as
indirect anti-MIF strategies (21,54).
Using a highly metastatic syngenic murine C3L5 breast cancer model,
Majunder et al (55) showed
that indomethacin, a COX-1/COX-2 inhibitor inhibited cell
proliferation and migration. In vitro, celecoxib, a COX-2
inhibitor reduced tumor growth and spontaneous lung and lymph node
metastases. However published data on the impact of COX-2
inhibition on MIF expression and its biological consequences are
lacking in breast cancer cells.
In conclusions, MIF pathway is obviously involved in
the breast cancer biology and interfering with it remains an
attractive but complex challenge.
Acknowledgements
N.K. is the recipient of a grant (Televie) from the
National Fund for Scientific Research.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pietras K and Östman A: Hallmarks of
cancer: Interactions with the tumor stroma. Exp Cell Res.
316:1324–1331. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mitchell RA and Yaddanapudi K:
Stromal-dependent tumor promotion by MIF family members. Cell
Signal. 26:2969–2978. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Conroy H, Mawhinney L and Donnelly SC:
Inflammation and cancer: Macrophage migration inhibitory factor
(MIF) - the potential missing link. QJM. 103:831–836. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Donn RP and Ray DW: Macrophage migration
inhibitory factor: Molecular, cellular and genetic aspects of a key
neuroendocrine molecule. J Endocrinol. 182:1–9. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Swope M, Sun HW, Blake PR and Lolis E:
Direct link between cytokine activity and a catalytic site for
macrophage migration inhibitory factor. EMBO J. 17:3534–3541. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li XJ, Luo Y and Yi YF: P115 promotes
growth of gastric cancer through interaction with macrophage
migration inhibitory factor. World J Gastroenterol. 19:8619–8629.
2013. View Article : Google Scholar :
|
|
8
|
Zernecke A, Bernhagen J and Weber C:
Macrophage migration inhibitory factor in cardiovascular disease.
Circulation. 117:1594–1602. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim H, Lee S, Kim HJ, Kong MH, Kim YR,
Kang SH, Lee K, Leng L, Lee B, Park CG, et al: Elevated levels of
macrophage migration inhibitory factor in women with metabolic
syndrome. Horm Metab Res. 43:642–645. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kindt N, Preillon J, Kaltner H, Gabius HJ,
Chevalier D, Rodriguez A, Johnson BD, Megalizzi V, Decaestecker C,
Laurent G, et al: Macrophage migration inhibitory factor in head
and neck squamous cell carcinoma: Clinical and experimental
studies. J Cancer Res Clin Oncol. 139:727–737. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang D, Luo L, Chen W, Chen LZ, Zeng WT,
Li W and Huang XH: Significance of the vascular endothelial growth
factor and the macrophage migration inhibitory factor in the
progression of hepatocellular carcinoma. Oncol Rep. 31:1199–1204.
2014.
|
|
12
|
Krockenberger M, Kranke P, Häusler S,
Engel JB, Horn E, Nürnberger K, Wischhusen J, Dietl J and Hönig A:
Macrophage migration-inhibitory factor levels in serum of patients
with ovarian cancer correlates with poor prognosis. Anticancer Res.
32:5233–5238. 2012.PubMed/NCBI
|
|
13
|
Tan L, Ye X, Zhou Y, Yu M, Fu Z, Chen R,
Zhuang B, Zeng B, Ye H, Gao W, et al: Macrophage migration
inhibitory factor is overexpressed in pancreatic cancer tissues and
impairs insulin secretion function of β-cell. J Transl Med.
12:922014. View Article : Google Scholar
|
|
14
|
He XX, Yang J, Ding YW, Liu W, Shen QY and
Xia HH: Increased epithelial and serum expression of macrophage
migration inhibitory factor (MIF) in gastric cancer: Potential role
of MIF in gastric carcinogenesis. Gut. 55:797–802. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi X, Leng L, Wang T, Wang W, Du X, Li J,
McDonald C, Chen Z, Murphy JW, Lolis E, et al: CD44 is the
signaling component of the macrophage migration inhibitory
factor-CD74 receptor complex. Immunity. 25:595–606. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee CY, Su MJ, Huang CY, Chen MY, Hsu HC,
Lin CY and Tang CH: Macrophage migration inhibitory factor
increases cell motility and up-regulates αvβ3 integrin in human
chondrosar-coma cells. J Cell Biochem. 113:1590–1598. 2012.
|
|
17
|
Mitchell RA: Mechanisms and effectors of
MIF-dependent promotion of tumourigenesis. Cell Signal. 16:13–19.
2004. View Article : Google Scholar
|
|
18
|
Xin D, Rendon BE, Zhao M, Winner M, McGhee
Coleman A and Mitchell RA: The MIF homologue D-dopachrome
tautomerase promotes COX-2 expression through β-catenin-dependent
and -independent mechanisms. Mol Cancer Res. 8:1601–1609. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meyer-Siegler K: COX-2 specific inhibitor,
NS-398, increases macrophage migration inhibitory factor expression
and induces neuroendocrine differentiation in C4-2b prostate cancer
cells. Mol Med. 7:850–860. 2001.
|
|
20
|
Xia HH, Zhang ST, Lam SK, Lin MC, Kung HF
and Wong BC: Expression of macrophage migration inhibitory factor
in esophageal squamous cell carcinoma and effects of bile acids and
NSAIDs. Carcinogenesis. 26:11–15. 2005. View Article : Google Scholar
|
|
21
|
Ristimäki A, Sivula A, Lundin J, Lundin M,
Salminen T, Haglund C, Joensuu H and Isola J: Prognostic
significance of elevated cyclooxygenase-2 expression in breast
cancer. Cancer Res. 62:632–635. 2002.PubMed/NCBI
|
|
22
|
de Pedro M, Baeza S, Escudero MT,
Dierssen-Sotos T, Gómez-Acebo I, Pollán M and Llorca J: Effect of
COX-2 inhibitors and other non-steroidal inflammatory drugs on
breast cancer risk: A meta-analysis. Breast Cancer Res Treat.
149:525–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tarnowski M, Grymula K, Liu R, Tarnowska
J, Drukala J, Ratajczak J, Mitchell RA, Ratajczak MS and Kucia M:
Human Rhabdomyosarcomas secrete MIF that modulates metastatic
behavior of tumor cells and inhibits recruitment of cancer
associated fibroblasts. Mol Cancer Res. 8:1328–1343. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Martinez LM, Vallone VBF, Labovsky V, Choi
H, Hofer EL, Feldman L, Bordenave RH, Batagelj E, Dimase F,
Villafañe AR, et al: Changes in the peripheral blood and bone
marrow from untreated advanced breast cancer patients that are
associated with the establishment of bone metastases. Clin Exp
Metastasis. 31:213–232. 2014. View Article : Google Scholar
|
|
25
|
Verjans E, Noetzel E, Bektas N, Schûtz AK,
Lue H, Lennartz B, Hartmann A, Dahl E and Bernhagen J: Dual role of
macrophage migration inhibitory factor (MIF) in human breast
cancer. BMC Cancer. 9:2302009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schulz R, Streller F, Scheel AH, Rüschoff
J, Reinert MC, Dobbelstein M, Marchenko ND and Moll UM: HER2/ErbB2
activates HSF1 and thereby controls HSP90 clients including MIF in
HER2-overexpressing breast cancer. Cell Death Dis. 5:e9802014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bando H, Toi M, Kitada K and Koike M:
Genes commonly upregulated by hypoxia in human breast cancer cells
MCF-7 and MDA-MB-231. Biomed Pharmacother. 57:333–340. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Larsen M, Tazzyman S, Lund EL, Junker N,
Lewis CE, Kristjansen PE and Murdoch C: Hypoxia-induced secretion
of macrophage migration-inhibitory factor from MCF-7 breast cancer
cells is regulated in a hypoxia-inducible factor-independent
manner. Cancer Lett. 265:239–249. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lue H, Dewor M, Leng L, Bucala R and
Bernhagen J: Activation of the JNK signalling pathway by macrophage
migration inhibitory factor (MIF) and dependence on CXCR4 and CD74.
Cell Signal. 23:135–144. 2011. View Article : Google Scholar
|
|
30
|
Dewan MZ, Ahmed S, Iwasaki Y, Ohba K, Toi
M and Yamamoto N: Stromal cell-derived factor-1 and CXCR4 receptor
interaction in tumor growth and metastasis of breast cancer. Biomed
Pharmacother. 60:273–276. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lue H, Thiele M, Franz J, Dahl E,
Speckgens S, Leng L, Fingerle-Rowson G, Bucala R, Lüscher B and
Bernhagen J: Macrophage migration inhibitory factor (MIF) promotes
cell survival by activation of the Akt pathway and role for
CSN5/JAB1 in the control of autocrine MIF activity. Oncogene.
26:5046–5059. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Simpson KD, Templeton DJ and Cross JV:
Macrophage migration inhibitory factor promotes tumor growth and
metastasis by inducing myeloid-derived suppressor cells in the
tumor microenvironment. J Immunol. 189:5533–5540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lim S, Choong LY, Kuan CP, Yunhao C and
Lim YP: Regulation of macrophage inhibitory factor (MIF) by
epidermal growth factor receptor (EGFR) in the MCF10AT model of
breast cancer progression. J Proteome Res. 8:4062–4076. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Drews-Elger K, Iorns E, Dias A, Miller P,
Ward TM, Dean S, Clarke J, Campion-Flora A, Rodrigues DN,
Reis-Filho JS, et al: Infiltrating S100A8+ myeloid cells
promote metastatic spread of human breast cancer and predict poor
clinical outcome. Breast Cancer Res Treat. 148:41–59. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu X, Wang B, Ye C, Yao C, Lin Y, Huang X,
Zhang Y and Wang S: Overexpression of macrophage migration
inhibitory factor induces angiogenesis in human breast cancer.
Cancer Lett. 261:147–157. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Oda S, Oda T, Nishi K, Takabuchi S,
Wakamatsu T, Tanaka T, Adachi T, Fukuda K, Semenza GL and Hirota K:
Macrophage migration inhibitory factor activates hypoxia-inducible
factor in a p53-dependent manner. PLoS One. 3:e22152008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu MY, Fu J, Xu J, O'Malley BWO and Wu RC:
Steroid receptor coactivator 3 regulates autophagy in breast cancer
cells through macrophage migration inhibitory factor. Cell Res.
22:1003–1021. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schulz R, Marchenko ND, Holembowski L,
Fingerle-Rowson G, Pesic M, Zender L, Dobbelstein M and Moll UM:
Inhibiting the HSP90 chaperone destabilizes macrophage migration
inhibitory factor and thereby inhibits breast tumor progression. J
Exp Med. 209:275–289. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fujita Y, Islam R, Sakai K, Kaneda H, Kudo
K, Tamura D, Aomatsu K, Nagai T, Kimura H, Matsumoto K, et al:
Aza-derivatives of resveratrol are potent macrophage migration
inhibitory factor inhibitors. Invest New Drugs. 30:1878–1886. 2012.
View Article : Google Scholar
|
|
40
|
Sprouse AA and Herbert BS: Resveratrol
augments paclitaxel treatment in MDA-MB-231 and
paclitaxel-resistant MDA-MB-231 breast cancer cells. Anticancer
Res. 34:5363–5374. 2014.PubMed/NCBI
|
|
41
|
Bando H, Matsumoto G, Bando M, Muta M,
Ogawa T, Funata N, Nishihira J, Koike M and Toi M: Expression of
macrophage migration inhibitory factor in human breast cancer:
Association with nodal spread. Jpn J Cancer Res. 93:389–396. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Richard V, Kindt N, Decaestecker C, Gabius
HJ, Laurent G, Noël JC and Saussez S: Involvement of macrophage
migration inhibitory factor and its receptor (CD74) in human breast
cancer. Oncol Rep. 32:523–529. 2014.PubMed/NCBI
|
|
43
|
Fersching DM, Nagel D, Siegele B, Salat C,
Heinemann V, Holdenrieder S and Stoetzer OJ: Apoptosis-related
biomarkers sFAS, MIF, ICAM-1 and PAI-1 in serum of breast cancer
patients undergoing neoadjuvant chemotherapy. Anticancer Res.
32:2047–2058. 2012.PubMed/NCBI
|
|
44
|
Jesneck JL, Mukherjee S, Yurkovetsky Z,
Clyde M, Marks JR, Lokshin AE and Lo JY: Do serum biomarkers really
measure breast cancer? BMC Cancer. 9:1642009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Opstal-van Winden AW, Rodenburg W,
Pennings JLA, van Oostrom CTM, Beijnen JH, Peeters PHM, van Gils CH
and de Vries A: A bead-based multiplexed immunoassay to evaluate
breast cancer biomarkers for early detection in pre-diagnostic
serum. Int J Mol Sci. 13:13587–13604. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Choi J, Jung WH and Koo JS:
Metabolism-related proteins are differentially expressed according
to the molecular subtype of invasive breast cancer defined by
surrogate immunohistochemistry. Pathobiology. 80:41–52. 2013.
View Article : Google Scholar
|
|
47
|
Catteau X, Simon P, Vanhaeverbeek M and
Noël JC: Variable stromal periductular expression of CD34 and
smooth muscle actin (SMA) in intraductal carcinoma of the breast.
PLoS One. 8:e577732013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Catteau X, Simon P and Noël JC:
Myofibroblastic stromal reaction and lymph node status in invasive
breast carcinoma: possible role of the TGF-β1/TGF-βR1 pathway. BMC
Cancer. 4:4992014. View Article : Google Scholar
|
|
49
|
Catteau X, Simon P and Noël JC:
Myofibroblastic reaction is a common event in metastatic disease of
breast carcinoma: A descriptive study. Diagn Pathol. 9:1962014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Salgado R, Denkert C, Demaria S, Sirtaine
N, Klauschen F, Pruneri G, Wienert S, Van den Eynden G, Baehner FL,
Penault-Llorca F, et al: The evaluation of tumor-infiltrating
lymphocytes (TILs) in breast cancer: Recommendations by an
International TILs Working Group 2014. Ann Oncol. 26:259–271. 2015.
View Article : Google Scholar
|
|
51
|
Jhaveri K, Chandarlapaty S, Lake D,
Gilewski T, Robson M, Goldfarb S, Drullinsky P, Sugarman S,
Wasserheit-Leiblich C, Fasano J, et al: A phase II open-label study
of ganetespib, a novel heat shock protein 90 inhibitor for patients
with metastatic breast cancer. Clin Breast Cancer. 14:154–160.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Baxalta US Inc. (sponsor). Phase I Study
of anti-Macrophage Migration Inhibitory Factor antibody in solid
Tumors. ClinicalTrials.gov identifier: NCT01765790. https://clinicaltrials.gov/ct2/show/NCT01765790.
|
|
53
|
Berkova Z, Tao RH and Samaniego F:
Milatuzumab - a promising new immunotherapeutic agent. Expert Opin
Investig Drugs. 19:141–149. 2010. View Article : Google Scholar
|
|
54
|
Howe LR: Inflammation and breast cancer.
Cyclooxygenase/ prostaglandin signaling and breast cancer. Breast
Cancer Res. 9:2102007. View Article : Google Scholar
|
|
55
|
Majumder M, Xin X, Liu L, Girish GV and
Lala PK: Prostaglandin E2 receptor EP4 as the common target on
cancer cells and macrophages to abolish angiogenesis,
lymphangiogenesis, metastasis, and stem-like cell functions. Cancer
Sci. 105:1142–1151. 2014. View Article : Google Scholar : PubMed/NCBI
|