Introduction
Ewing's sarcoma (ES) is the second most common
primary bone malignancy in children and young adults (1). Patients with nonmetastatic ES have
improved outcomes, however, 30–40% of recovering ES patients
experience recurrence or relapse (2). Traditional therapeutic approaches
include local control of the primary lesion by surgery and/or
radiation therapy, and treatment of disseminated disease with
multi-agent cytotoxic chemotherapy. These approaches have led to
significant improvements in outcomes in recent decades,
particularly in patients with localized disease (3). However, the high percentage of
relapse, metastasis, and poor prognosis in ES patients has led to
renewed interest in novel treatments.
miRNAs play important roles in cancer development
and progression (4). Altered miRNA
expression profiles have been reported in various types of cancer,
including liver, colon, and prostate (5,6).
Functionally, aberrant miRNA expression can affect cell
proliferation (7), apoptosis
(8), chemo- and
radio-sensitivities (9), and
development of metastases (10).
Recent studies indicated that miR-301a is significantly
overexpressed in many kinds of cancer, including pancreatic,
breast, hepatocellular, and small-cell lung cancers (11–14).
However, the role of miR-301a in ES cell proliferation remains
unclear.
Phosphatase and tensin homologue (PTEN) is
considered as a tumor suppressor gene associated with the p53
signaling pathway (15,16). PTEN plays a vital role in
regulating the cell cycle and inhibiting cell growth and division
(17,18). Recently, several reports
demonstrated PTEN involvement in apoptosis in human hepatocellular
carcinoma cells (19), and in cell
proliferation, migration, and invasion in gastric cancer (20).
In this study, we analyzed genome-wide microarray
expression data for both miRNAs and mRNAs in five human ES cell
lines and human mesenchymal stem cells (hMSCs). The results
indicated increased miR-301a expression and repressed PTEN
expression in all five ES cell lines compared to hMSCs. Based on
the inverse correlation observed between miR-301a and PTEN
expression, we hypothesized that the effect of PTEN in ES cells
might be directly or indirectly mediated in part by miR-301a.
Therefore, the purpose of our study was to assess whether PTEN
expression was repressed by miR-301a and the role of this pathway
in ES cell tumorigenesis.
Materials and methods
Cell lines
The human ES cell lines, SKES1, RDES, SKNMC and SCCH
were obtained from JCRB Cell Bank (Tokyo, Japan), and WE68 was
kindly provided Dr Frans van Valen (Westfalische-Wilhelms
University, Münster, Germany). Human mesenchymal stem cells (hMSCs)
were purchased from Takara Biotechnology (Otsu, Japan). Each line
was authenticated as to genotype and phenotype by the source
company. RDES and SKNMC cells were cultured in Dulbecco's modified
eagle medium (DMEM) high glucose medium (Invitrogen, Grand Island,
NY, USA) with 10% FBS and 1% penicillin and streptomycin. SKES1
cell were cultured in RPMI-1640 (Invitrogen) supplemented with 10%
FBS. SCCH cells were grown in minimal essential medium (MEM)
supplemented with 10% fetal bovine serum (FBS; Invitrogen) and 0.1
mmol/l nonessential amino acids (NEAA). hMSC were cultured in
mesenchymal stem cell basal medium, chemically defined (MSCBM-CD)
with MSCGM-CD SingleQuats (Takara Biotechnology). The cells were
maintained at 37°C in an incubator with 5% CO2 and
passaged every 2 to 3 days.
Ethics statement
The animal experimental protocol was approved by the
Ethics Review Committee for Animal Experimentation of Oita
University, and all mice used in this study were anesthetized with
ketamine/xylazine or isoflurane/oxygen for experiments and
euthanized with cervical dislocation under anesthesia. All efforts
were made to minimize suffering.
Mice
BALB/c nu/nu mice (n=28, 6-week old), were acquired
from the Kyodo Laboratory (Tosu, Japan). After quarantine, all mice
were kept in a pathogen-free environment on a standard 12 h-day/12
h-night cycle and were fed a standard sterilized pellet diet and
water ad libitum. All mice were continuously monitored
during daytime from Monday to Friday, and twice daily during
daytime on Saturdays, Sundays, and holidays for signs of poor
health.
RNA isolation
mRNAs were prepared from the triplicate cell
cultures using RNeasy kit (Qiagen, Valencia, CA, USA) according to
the manufacturer's instruction. The RNA quality was ensured, before
labeling, using RNA 6000 Nano kit and Bioanalyzer 2100 (Agilent,
Santa Clara, CA, USA). miRNAs were prepared from triplicate cell
cultures using the miRNeasy Mini kit (Qiagen) according to the
manufacturer's instructions.
Genome-wide miRNA expression
microarray
GeneChip miRNA 3.0 array (Affymetrix, Santa Clara,
CA, USA) was used for miRNA expression profiling in five ES cell
lines and hMSCs. Small RNA (1 μg) including miRNA from each sample
was labeled with biotin using the FlashTag Biotin HSR kit
(Genisphere, Hatfield, PA, USA). Array hybridization, washing and
scanning of the slides were carried out according to the
manufacturer's recommendations. The data were extracted from the
images, quantile-normalized, summarized (median polish), and
log2-transformed with miRNA QC software (Affymetrix). GeneSpring GX
11.0 (Agilent) was used to analyze the array results. Analysis of
variance was used to determine the probe sets significantly
different between the two groups. The gene list was filtered with a
fold-change cutoff of 2 resulting in the output of a list with
genes that have significant differential expression at ≥2-fold.
Pathway analysis was performed using KEGG pathway Database
(http://www.genome.jp/kegg/pathway.html).
Analysis of mRNA expression by cDNA
arrays
GeneChip Genome HG U133 Plus 2.0 Array (Affymetrix)
was used for mRNA expression profiling in 5 ES cell lines and hMSC.
Biotinylated cRNA was synthesized from total RNA using the 3′ IVT
Express kit (Affymetrix) according to the manufacturer's protocols.
In brief, double stranded cDNA was generated by reverse
transcription from 1 ng of total RNA an oligo (dT) primer bearing a
T7 promoter. The double-strand cDNA was used as a template for
in vitro transcription to generate biotin-labeled cRNA.
After fragmentation, 12.5 μg of cRNA was hybridized to GeneChip
array for 16 h. The arrays were washed and stained using GeneChip
Fluidics Station 450 (Affymetrix) and then scanned with the
GeneChip Scanner 3000 (Affymetrix). The entire experiment was
performed twice. Array hybridization, washing, and scanning of the
slides were carried out according to the manufacturer's protocols.
The microarray numerical values were analyzed using GeneSpring GX
11.0 software: quantile normalization, filter by flags (detected),
filter by expression on the normalized data (20.0–100.0th
percentile). Analysis of variance was used to determine the probe
sets significantly different between the two groups. The gene list
was filtered with a fold-change cutoff of 2, resulting in output of
a list with genes with significant differential expression at
≥2-fold.
Target prediction of miRNAs
Basic Local Alignment Search Tool (BLAST),
TargetScan 6.0, microRNA.org, was used to search for the predicted
target genes of miRNAs.
Prediction of binding site and mature
miRNA transfection
Among the predicted target genes of miR-301a in the
TargetScan (http://www.targetscan.org/), DIANA (http://diana.cslab.ece.ntua.gr/microT/),
and PicTar (http://pictar.mdc-berlin.de/) databases, we found that
PTEN was one of the top candidates. One day prior to the
transfection, cells were seeded onto 6-well plates
(1×105 cells/well) and incubated with the complete
medium without antibiotics (2 ml/well). Actinomycin D (10 μg/ml,
Sigma-Aldrich) was used to inhibit nascent RNA synthesis. The
transfection of miR-301a-3p mimic (5′-CAGUG CA AUAGUAU UGUCA A AG
C-3′) (20 nM), miR-301a-3p mutant (5′-CUCACGUAUAGUAUUG UCAAAGC-3′)
(20 nM) and negative control (NC) mRNAs (20 nM) (Invitrogen) was
performed using Lipofectamine 2000 reagent (Invitrogen) in
antibiotics-free OptiMEM (Invitrogen) according to the
manufacturer's instructions. After 48 h of incubation following the
transfection, the cells were harvested and processed for further
analysis.
RNA extraction, cDNA synthesis, and
quantitative real-time PCR
Total RNA was extracted from treated with the TRIzol
reagent (Invitrogen) and cDNA was synthesized according to the
manufacturer's protocol (Roche). Quantitative real-time PCR
(qRT-PCR) was performed using a Light Cycler 480 Probe Master
System (Roche), and PCR-specific amplification was conducted in the
LightCycler® Nano (Roche). The relative expression of
genes (PTEN and GAPDH) was calculated with the 2−ΔΔCt
method. The primers used were as listed: (qRT-PCR; PTEN-forward
5′-AAGACAAAGCCAACCGATAC-3′, PTEN-reverse
5′-GAAGTTGAACTGCTAGCCTC-3′; GAPDH-forward
5′-CCTCTATGCCAACACAGTGC-3′, GAPDH-reverse
5′-GTACTCCTGCTTGCTGATCC-3′.
miRNA inhibitor transfection
One day prior to the transfection, SKES1 and RDES
were seeded onto 6-well plates (1×105 cells/well) and
incubated with the complete medium without antibiotics (2 ml/well).
The transfection of hsa-miR-301a inhibitor (5, 10, 20, 40, 80 nM)
or negative control miRNAs (Control-miR) (5, 10, 20, 40, 80 nM)
(Invitrogen) was performed using Lipofectamine 2000 reagent
(Invitrogen) in antibiotics-free OptiMEM (Invitrogen) according to
the manufacturer's instructions. After 48 h of incubation following
the transfection, the cells were harvested and processed for
further analysis.
Cell proliferation assay
The PTEN expression plasmid (SC119965) was obtained
from OriGene Technologies Inc. (IA, USA). The SKES1 and RDES were
plated in 6-well plates (1×105 cells per well), and were
transfected with or without anti-miR-301a inhibitor, negative
control miRNA, PTEN plasmid and Mock plasmid vector using
Lipofectamine 2000. Then the cells were incubated in
antibiotic-free OptiMEM. After 48 h of the cultivation, the cells
were counted using TC10 Automated Cell Counter (Bio-Rad).
Western blotting
Total cellular protein (15 μg) was resolved on a
precast 10% Tris-HCl Criterion 10-well gel (Bio-Rad) at 200 V (300
mAmp) for 30 min. The gel was wet-transferred to a PVDF membrane
for 1 h, and blocked with PBST containing 5% instant dry non-fat
milk for 30 min at room temperature. Primary antibodies (x1000)
Rabbit source against PTEN (#9552), β-actin (#4970), Akt (#4691),
p-Akt (#4060), p21 (#2947), Bad (#9292) and p-Bad (#9291) proteins
were obtained from Cell Signal Technology (Tokyo, Japan).
Immunocomplexes were visualized with horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G antibodies
(x1000) (GE Healthcare, Tokyo, Japan), developed the blots using
ECL Prime system (GE Healthcare) with a ChemiDoc camera (ImageQuant
LAS 4000mini; GE Healthcare). The quantification of western blot
signals was performed by the densitometry with ImageQuant TL
software (GE Healthcare). All western blot experiments were
repeated at least three times.
Cell cycle analysis
For cell cycle analysis, SKES1 cells were stained
with propidium iodide using Cycletest Plus DNA Reagent kit (BD
Biosciences) following the manufacturer's protocol, and the cell
cycle distribution was analyzed by FACSVerse flow cytometer (BD
Biosciences). The percentages of cells in G0/G1, S and G2/M phases
were counted and compared. The experiments were carried out in
triplicate.
Apoptosis assay
The changes in the expression of apoptotic proteins
were analyzed by western blot analysis using antibodies against
PAR/poly (ADP-ribose) polymerase (PARP) (#9542) and cleaved PARP
(#9541) (Cell Signaling Technology, Tokyo, Japan) as an index of
apoptosis.
The quantification of cell death was determined by
fluorescence activated cell sorting (FACS) using Annexin V-FITC
apoptosis detection kit (BD Bioscience) according to the
manufacturer's instructions. Briefly, 1×106 SKES1 cells
were seeded and incubated for 24 h, then anti-miR-301a or PTEN
expression plasmid was added to the cells followed by incubation
for 48 h. The cells were washed with PBS, suspended in Annexin V
binding buffer, then added to an Annexin V-FITC solution and
propidium iodide (PI) for 20 min at room temperature. The samples
were analyzed by FACSVerse using FACSuite analysis software (BD
Bioscience). To verify the ability for induction of apoptosis,
SKES1 cells treated with low dose ADM at 5 μg/ml for 24 h were
used.
In vivo tumor-bearing nude mouse
models
The experimental metastasis model was established by
injection of 2×106 cells transfected with anti-miR-301a
suspended in 100 μl of normal saline into the gluteal region of
nude mice (BALB/c nu/nu, Kudo, Tosu, Japan). Three groups were
generated: i) untreated control (n=7); ii) transfected with
negative control-miRNA (n=7); iii) transfected with anti-miR-301a
(n=7); and transfected with PTEN expression vector (n=7). All mice
were fed in standard conditions with weight monitoring and
sacrificed 6 weeks after the cell inoculation. Tumor size was
measured in two perpendicular dimensions parallel with the surface
and the depth of the tumor in mice using a caliper. The tumor
volume was estimated using the formula V = (Length ×
Width2)/2.
Immunofluorescence microscopy
To determine the effect of anti-miR-301a and PTEN
expression vector on the protein level of PTEN, we also performed
immunofluorescence staining with PTEN (CST) or p-Akt antibodies
(CST). After 48 h, the transfected SKES1 cell lines were fixed with
4% formaldehyde for 20 min, then incubated with 0.5% Triton X-100.
A rabbit anti-Human PTEN/p-Akt antibody was used for
immunofluorescence staining. Following three washes with PBS, the
cells were incubated with a goat anti-rabbit Alexa Fluor 594
antibody (Life Technology).
Statistical analysis
Statistical analysis was carried out using SPSS 18.0
software (SPSS Japan Inc., Tokyo, Japan). Two-tailed Student's
t-test was used for analysis of continuous variables. We determined
the differences among more than 3 groups using a nonrepeated
measures analysis of variance (ANOVA) and Scheffe test. Results are
expressed as the mean ± standard deviation, and p<0.01 was
considered to indicate a statistically significant difference.
Results
Upregulation of miR-301a expression in ES
cell lines
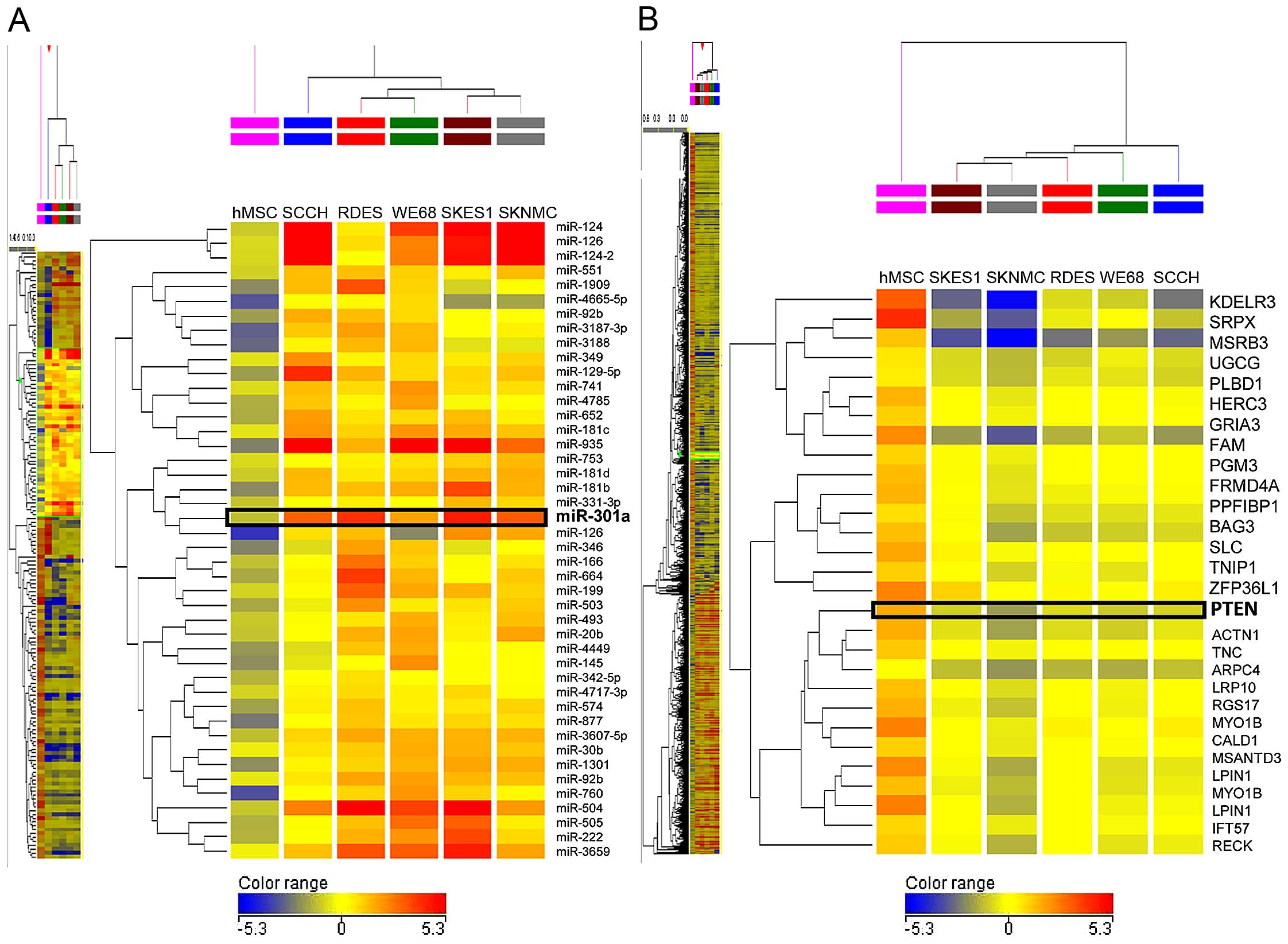
The genome-wide miRNA expression profiling using
five ES cell lines was carried out to identify miRNAs specifically
expressed in ES cells. The array analysis showed that the
expression levels of 1054 miRNAs in ES cells were changed
(fold-change >2.0) in comparison with hMSCs (Fig. 1A). Among 1054 miRNAs, 228 were
significantly upregulated, whereas 705 were significantly
downregulated in all tested ES cells compared to hMSCs. The
remaining 121 miRNAs were up- or down-regulated among five ES cell
lines. In ES cell lines, the increased expression of miR-301a by
2.63 to 40.21-fold compared with hMSCs was observed.
Downregulation of PTEN expression in ES
cell lines
The cDNA array analysis demonstrated that the
expression levels of 3043 mRNAs were significantly changed between
five ES cell lines and hMSCs (Fig.
1B). We found that 1062 genes were significantly upregulated,
whereas 1884 genes were significantly downregulated and the
remaining 97 genes were up- or down-regulated in five ES cell lines
compared to hMSCs. The expression of PTEN was decreased by −2.69 to
−6.55-fold in five ES cell lines.
PTEN as a direct miR-301a target in ES
cell lines
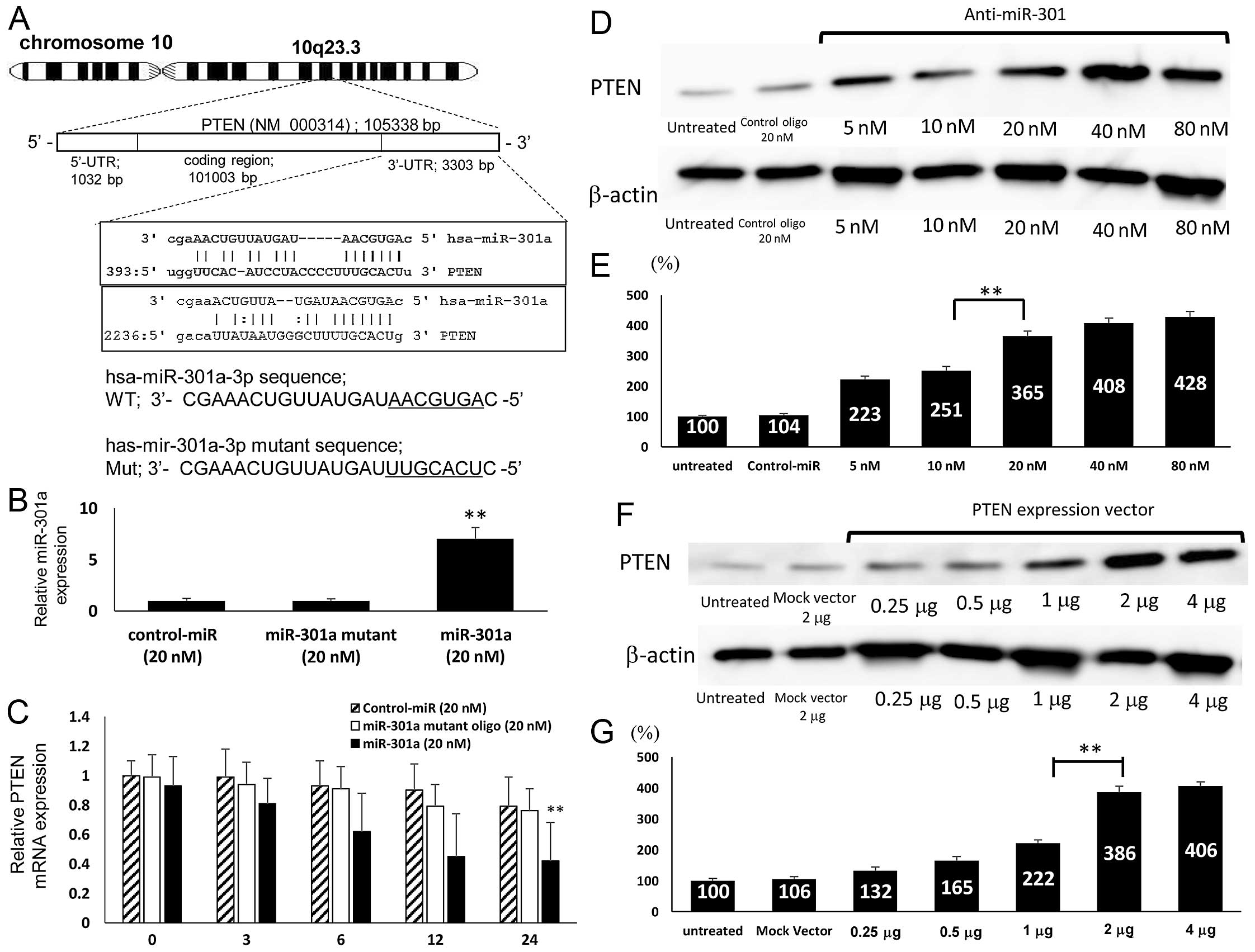
The region complementary to the miR-301a seed region
was found in the 3′-UTR of human PTEN (Fig. 2A). A considerable complementarity
between sequences within the seed regions of miR-301a and sequences
in the 3′ untranslated region (UTR) of PTEN was predicted, using
the algorithms in BLAST and TargetScan. The results suggested that
miR-301a might affect the expression of PTEN genes by binding to
3′UTR of PTEN. We blocked de novo mRNA transcription using
actinomycin D (10 μg/ml), an inhibitor of mRNA transcription to
determine changes in miRNA or mRNA stability. To test whether
miR-301a expression affected endogenous PTEN expression, we
transfected the miR-301a and miR-301a mutant oligonucleotides, as
well as the negative control-miR, into SKES1 cells. We observed an
increased miR-301a expression by 7.03±1.09-fold compared with
miR-93 mutant (0.99±0.19) or control-miR (1.00±0.21) (p<0.01)
(Fig. 2B) and decreased PTEN
expression at the mRNA level following transfection with the
miR-301a-3p oligonucleotide 0.42±0.26 compared with miR-301a mutant
(0.76±0.15) or control-miR (0.79±0.2) (p<0.01) (Fig. 2C).
Upregulation of PTEN expression by
anti-miR-301a and PTEN expression vector
To examine the correlation between miR-301a and PTEN
in ES cells, miR-301a inhibitor anti-miR-301a was transfected into
SKES1 cells. Western blot analysis showed that the expression
levels of PTEN markedly increased in anti-miR-301a transfected
cells compared with untreated or negative control oligo-transfected
cells (Fig. 2D). The protein
expression level of PTEN in the anti-miR-301a (20 nM) transfected
cells was increased to 3.65±0.17-fold of that in the control cells
(p<0.01) (Fig. 2E). To further
confirm the effects of PTEN on the growth of ES cells, the
transfection with PTEN expression vector was carried out. Although
the expression level of PTEN protein in the cells transfected with
Mock vector was not significantly affected, that in the cells
transfected with PTEN expression vector was significantly reduced,
as determined by western blotting (Fig. 2F). Compared to the control cells
(100%), PTEN vector (2 μg) transfected cells exhibited
significantly higher PTEN expression level by 3.86±0.19-fold
(p<0.01) (Fig. 2G).
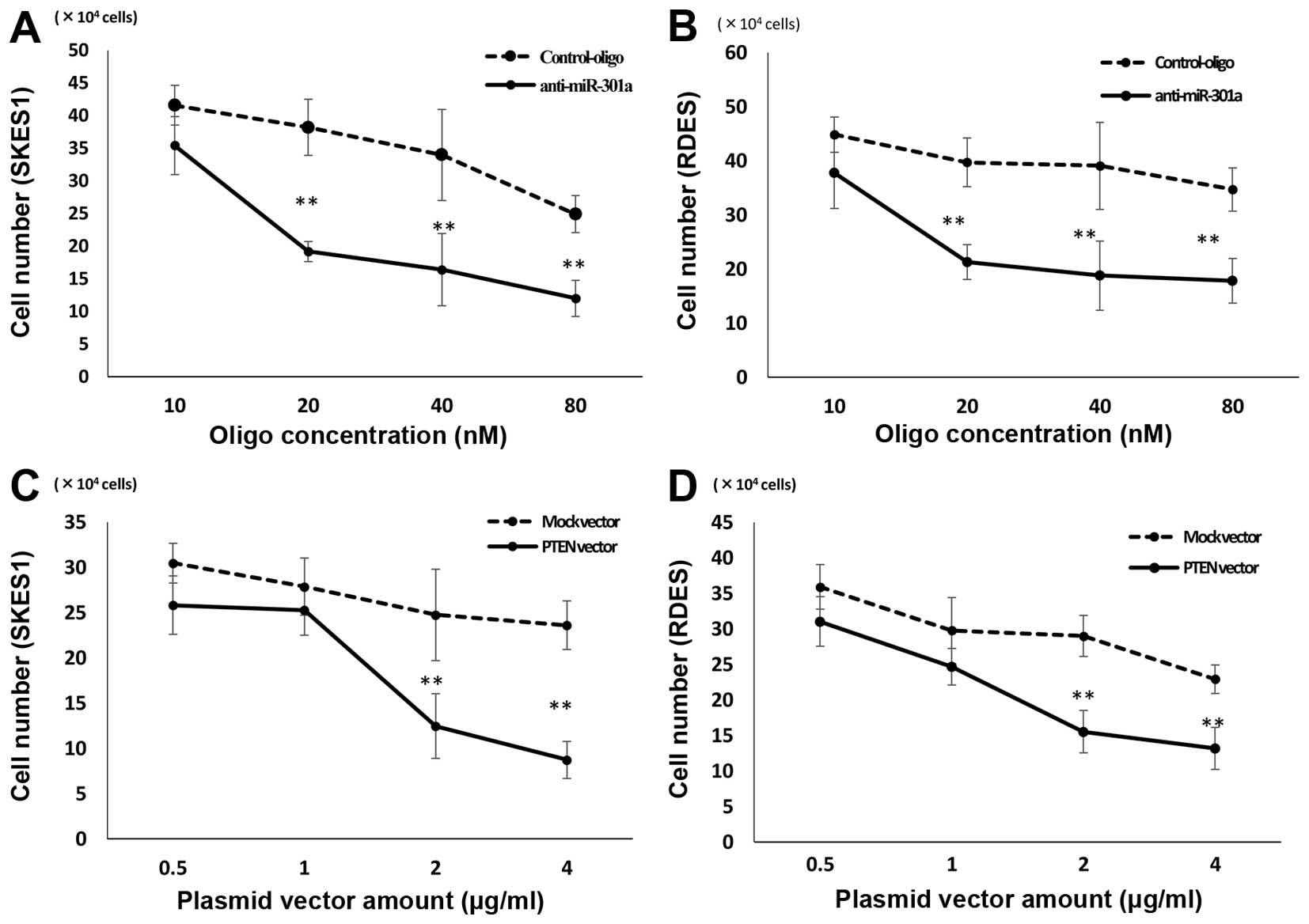
Suppression of ES cell growth by
transfection of anti-301a-miR and PTEN expression vector
PTEN is known to play important roles in the
regulation of cell proliferation. Since the transfection of
anti-miR-301a resulted in the reduction of PTEN expression, we next
examine the effects of anti-miR-301a on the proliferation of ES
cells. The cell growth of SKES1 was inhibited by the transfection
of anti-miR-301a as determined by cell counting in comparison with
negative control-miRNA transfected cells (Fig. 3A). The cell growth of RDES was
inhibited by the transfection of anti-miR-301a as determined by
cell counting in comparison with negative control-miRNA transfected
cells (Fig. 3B). The cell growth
of SKES1 was inhibited by the transfection of PTEN expression
vector as determined by cell counting in comparison with negative
control-miRNA transfected cells (Fig.
3C). The cell growth of RDES was inhibited by the transfection
of PTEN expression vector as determined by cell counting in
comparison with negative control-miRNA transfected cells (Fig. 3D).
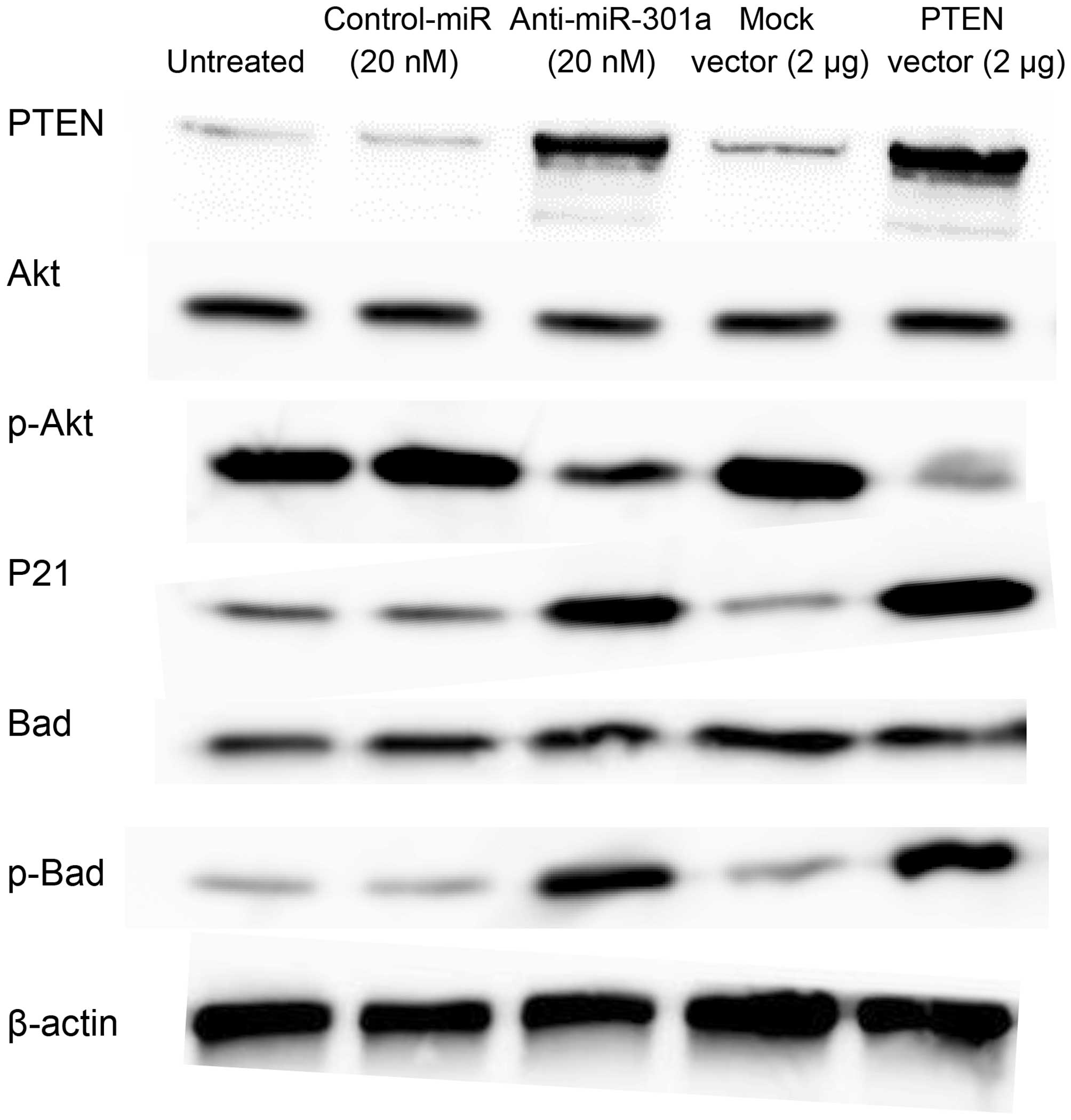
PTEN restoration induced the expression
change of Akt
To examine the correlation between PTEN and Akt, the
expression of PTEN, Akt, and their downstream factors were
investigated in SKES1 cells. SKES1 transfected miR-301a inhibitor
and PTEN vector showed increase in expression of PTEN, p21 and
phospho (p)-Bad. Western blot analysis showed that the expression
levels of p-Akt markedly decreased in anti-miR-301a and PTEN vector
transfected cells compared with untreated, negative control oligo
and Mock vector transfected cells (Fig. 4).
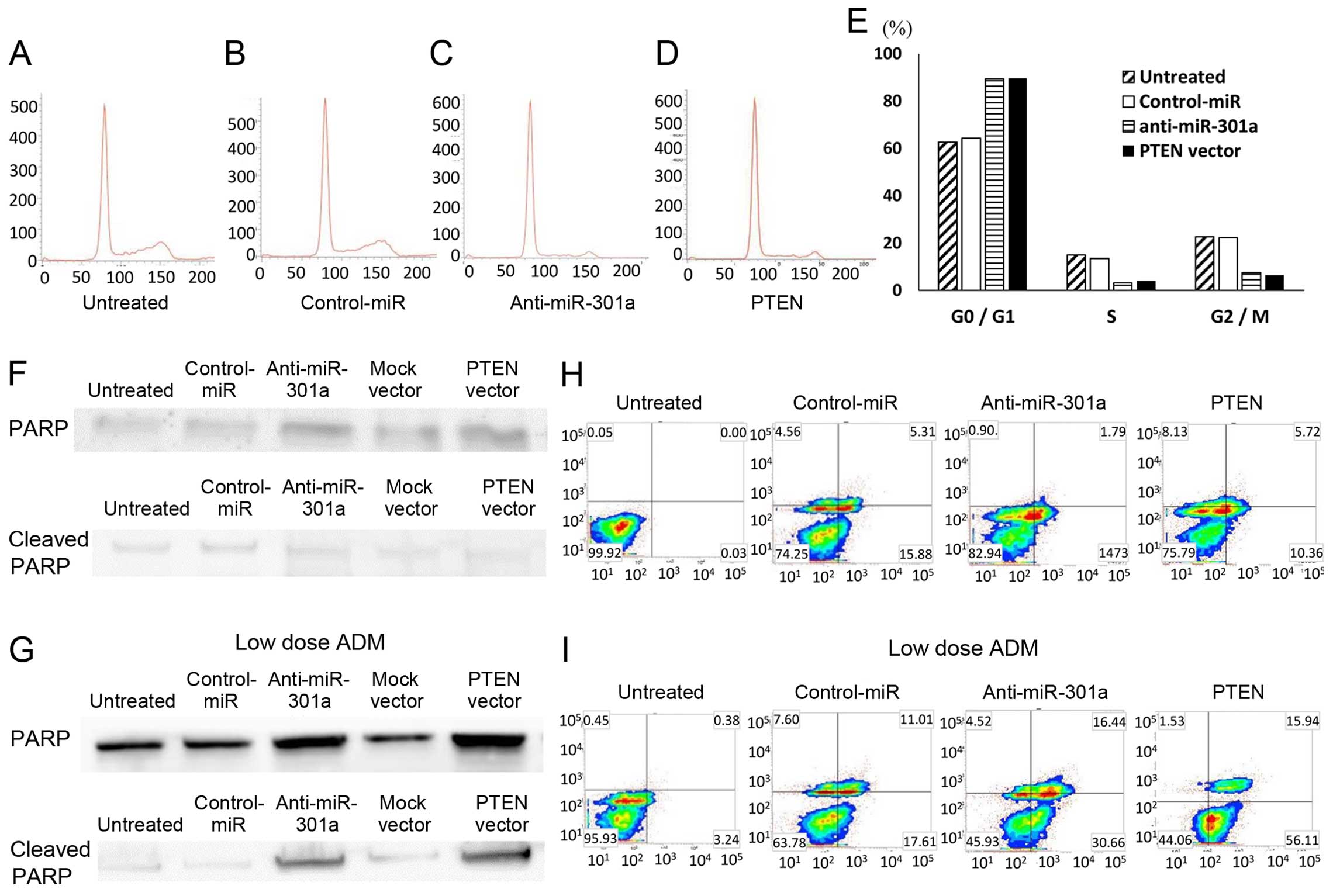
Induction of cell cycle arrest at G0/G1
phase by anti-miR-301a
Since the introduction of anti-miR-301a
significantly inhibited cell proliferation of SKES1 cells, we
hypothesized that anti-miR-301a might reduce the cell cycle
progression and/or induce-apoptosis of the cells. To monitor the
cell cycle distribution, FACS analyses were carried out using
anti-miR-301a and PTEN plasmid transfected cells (Fig. 5A–D). Both in anti-miR-301a and PTEN
plasmid transfected cell lines, the number of the cells in G2/M and
G0/G1 phase was significantly lower, and higher than that in the
control cells, respectively (Fig.
5E). The data suggested that the repression of miR-301a and
restoration of PTEN might have resulted in G0/G1 retardation in ES
cells. Then the cellular expression of PARP and its cleaved product
was assayed by immunoblotting in SKES1 cells and their
transfectants (Fig. 5F). The
cleavage of PARP protein, a marker of caspase-mediated apoptosis,
was not observed in both anti-miR-301a and PTEN transfectants or
the untreated cells and negative control or in mock plasmid vector
transfectants. Adriamycin (ADM) is commonly used for the induction
of apoptosis as positive control. Additional low dose ADM treatment
was able to induce expression of cleaved PARP in both anti-301a-miR
and PTEN transfectant, this was not observed in untreated cells,
the negative control or the mock plasmid vector transfectants
(Fig. 5G). Furthermore, in flow
cytometry analysis using Annexin V-FITC/PI double staining, there
was no significant difference in the distribution patterns between
untreated, negative control miRNA, anti-miR-301a or PTEN plasmid
transfected cells (Fig. 5H).
Programmed cell death was induced by low dose ADM treatment in
anti-miR-301a or PTEN expression vector in SKES1 cells (Fig. 5I).
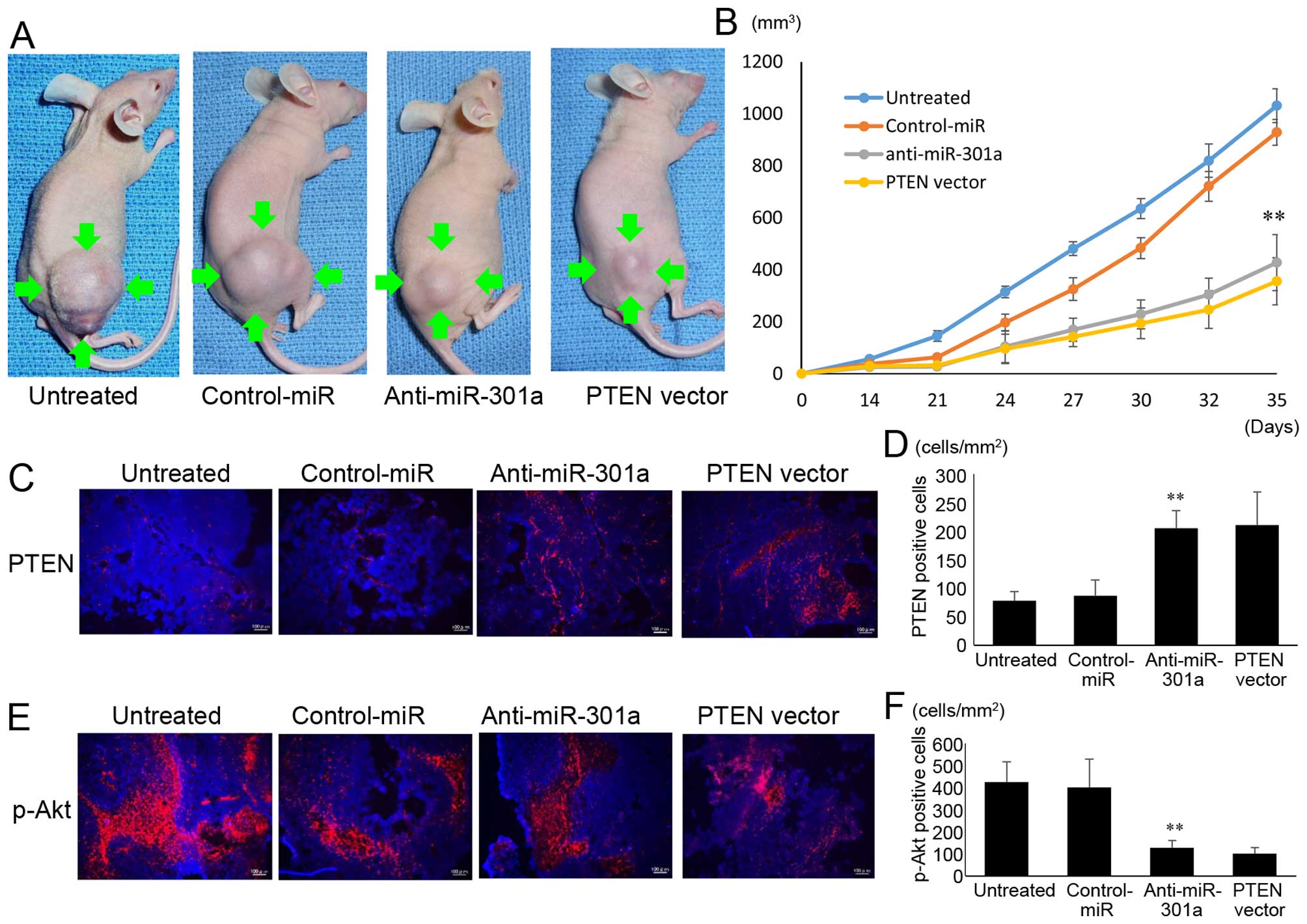
Inhibition of tumor growth in a nude
mouse xenograft model by anti-miR-301a
We investigated the efficacy of anti-miR-301a
against tumor growth in vivo. The introduction of
anti-miR-301a into SKES1 cells resulted in decreased growth of
subcutaneous xenografted tumors in nude mice (Fig. 6A). SKES1 cells transfected with
anti-miR-301a (426.7±107.98 mm3) and PTEN expression
vector (355.4±90.2 mm3) showed statistically smaller
tumors in mice compared to untreated (1030.4±65.4 mm3)
and negative control miRNA (927.9±49.9 mm3) transfected
groups (Fig. 6B), indicating that
anti-miR-301a also inhibited the growth of ES cells in
vivo.
The expression of PTEN (Fig. 6C) and p-AKT (Fig. 6E) within the primary tumor lesion
was reduced in the anti-miR-301a and PTEN expression vector
transfected tumor tissues. The number of PTEN positive cells per
unit area was higher (p<0.01) in the mice with anti-miR-301a
(205±30.8 cells/mm2) and PTEN expression vector
(210.5±57.9 cells/mm2) than in those that received
untreated (77.7±16.6 cells/mm2) and control-miR
86.5±28.1 cells/mm2) (Fig.
6D). The number of p-Akt positive cells per unit area was lower
(p<0.01) in the mice that anti-miR-301a (127.6±32.5
cells/mm2) and PTEN expression vector (100.8±28.1
cells/mm2) than in those that received untreated
(424±92.2 cells/mm2) and control-miR (401.2±127
cells/mm2) (Fig.
6F).
Discussion
MicroRNAs are small (19–25 nucleotides) noncoding
RNAs capable of modulating the expression of their cognate target
genes by binding to the 3′-UTR of target mRNAs and causing either
translational inhibition or mRNA cleavage (21). Aberrant miRNA expression
contributes to the proliferation, invasion, or metastatic behavior
of human cancers (22). One miRNA
is capable of targeting multiple genes in order to globally
regulate a number of biological processes. Moreover, aberrant miRNA
expression in human cancer cells causes destruction of
miRNA-mediated mRNA networks. Abnormalities in these networks could
influence the expression of tumorigenic proteins. To identify
important miRNA-mRNA relationships in ES, we performed genome-wide
miRNA microarray analysis, as well as cDNA microarray analysis, in
ES cells.
Herein, miRNA microarray results demonstrated that
the expression of miR-301a was upregulated in all five ES cell
lines. Several studies showed that miR-301a was upregulated and
involved in various biological and pathological processes,
including development, differentiation, inflammation, apoptosis,
and cancer metastasis (11–13).
The function of miR-301a is important in the ecology of other types
of cancer through the targeting of PTEN mRNA (23–25).
However, the biological role of miR-301a, including its
relationship to PTEN in ES cells, has not been clarified. Our
results indicated that the expression of miR-301a was coordinately
upregulated in ES cell lines, leading us to investigate genome-wide
mRNA profiling by cDNA microarray in order to detect the possible
targets of miR-301a in ES cells.
The data from cDNA microarray analysis showed that
PTEN mRNA expression was consistently decreased in five ES cell
lines as compared with hMSCs. Furthermore, sequence analysis
suggested possible association of miR-301a with the 3′-UTR of PTEN
mRNA. PTEN is one of the most frequently mutated tumor
suppressor genes in human cancers, leading to the activation of the
phosphoinositide 3-kinase (PI3K)/ protein kinase B (Akt) signaling
pathway and increased cell survival and oncogenesis (26,27).
Numerous studies suggested that the transcriptional role of PTEN
involves a key position in the complex network of tumor suppressor
and oncogenes that regulate cellular transformation. PTEN
downregulation was found in lung cancer (28), hepatocellular carcinoma (29), and prostate cancer (30). PTEN level might be related to the
biology of Ewing's sarcoma cells and loss of PTEN could result in
clinically more aggressive tumors (31). Our data in ES cells are consistent
with previous studies reporting that PTEN downregulation may
contribute to malignant potentials (ref. ?).
Although miR-301a might influence the expression of
many genes, we focused on PTEN mRNA as a miR-301a target in ES
cells. Several genes have been reported as targets of miR-301a,
including Bim, RUNX3, Gax and Nkrf (11,12,32,33).
Our cDNA microarray analysis demonstrated that PTEN was the only
miR-301a-target gene whose expression was uniformly upregulated in
all five ES cell lines, whereas the expression of other candidate
genes was different among the ES cells. Analysis using multiple
algorithms, such as BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and
TargetScan (www.targetscan.org/), also suggested
that PTEN was the putative target of miR-301a. Thus, we
analyzed whether miR-301a might contribute to anticancer activities
by targeting PTEN mRNA in ES cells. It is possible that miR-301a
may have downregulated PTEN expression via indirect pathways. In
order to examine whether miR-301a inhibits PTEN mRNA
translation, we inhibited PTEN transcription using
actinomycin D, and then introduced miR-301a and a miR-301a variant
with a mutated seed region. The miR-301a mutant did not result in
any changes in PTEN levels, while non-mutated miR-301a resulted in
reductions in PTEN mRNA. Therefore, these results suggested that
miR-301a directly inhibited PTEN mRNA function, indicating that
miR-301a affects PTEN translation.
We examined the functions of miR-301a in the
regulation of its possible target gene, PTEN, and the
subsequent changes in the biological characteristics of an ES cell
line. The forced expression of miR-301a resulted in the repression
of PTEN protein translation, indicating that miR-301a might
function as an oncogene in ES cells. Ma et al reported that
PTEN, as a cell cycle and proliferation regulator, was the direct
target of miR-301a in breast cancer (23). Our results suggested that a similar
PTEN regulatory mechanism might exist via miR-301a in ES cells.
Our data regarding the cell cycle showed that
miR-301a promoted ES cell proliferation by inhibiting cell cycle
progression at the G1/G0 phase. These observations were consistent
with previous studies demonstrating that PTEN is necessary for the
control of the normal G1/S cell cycle checkpoint (34). It was shown that PTEN also played
important roles in regulating the G1/S-phase transition and
cellular transformation, and that restoring PTEN expression
resulted in cell cycle arrest in G1 phase (35). Therefore, we can assume that
miR-301a upregulation might affect ES cell cycle progression via
miR-301a-mediated control of PTEN expression. It is noteworthy that
PTEN upregulation following transfection of anti-301a-miR or a
PTEN-expression vector induced apoptosis in ES cells,
indicating that the repression of ES cell growth following
restoration of PTEN expression implied that PTEN was
necessary for cell cycle inhibition, as well as apoptosis
induction.
Provided that PTEN acts as a negative regulator of
the PI3K/Akt signaling pathway, PTEN repression by miR-301a might
play an important role in proliferation of other types of cells
(36). Akt activates the mTOR
signaling pathway (37) and
represses p21, as well as Bax, to enhance cell proliferation
preventing apoptosis (38,39). Therefore, Akt downregulation by
PTEN in ES cells induced the upregulation of p21 and Bax, which
might have resulted in cell cycle inhibition and apoptosis in ES
cells.
Furthermore, miR-301a repression resulted in the
inhibition of ES tumor growth in vivo. Consistent with the
data from in vitro experiments, xenograft ES models also
indicated that miR-301a repression was capable of inhibiting ES
tumor development in vivo following restoration of
PTEN expression and translation.
In summary, our results indicated a novel, inversely
correlated relationship between miR-301a and PTEN in ES cells. Our
in vitro and in vivo results provided evidence that
the expression levels of miR-301a in ES cells were significantly
increased and inversely correlated with PTEN expression levels, and
that miR-301a played important roles in ES cell proliferation and
tumorigenesis by targeting PTEN mRNA. Silenced PTEN has been
observed in several human cancers, wherein PTEN inhibition
promotes cell growth and proliferation (40,41).
Our data suggested that PTEN is one of the crucial factors involved
in enhancing tumor proliferation in ES, as well as other malignant
tumors. Although confirmation of the results demonstrated in the
present study are required using clinical ES samples, these novel
findings regarding the link between miR-301a and PTEN in ES cells
will be beneficial for understanding ES oncogenesis, and may
provide strategies for clinical application.
Acknowledgements
We would like to thank Katsuhiro Hanada Ph.D, and
Takashi Kobayashi, Ph.D, for helpful discussion for this study.
This work was supported in part by National Cancer Center Research
and Development Fund (26-A-4), the Grants-in-Aid for Scientific
Research (no. 24592250) and 15K10451 from Japan Society for the
Promotion of Science, and the Grants-in-Aid for Scientific Research
from Japan Agency for Medical Research and Development
(15Ack0106087h0002).
References
|
1
|
Linet MS, Ries LA, Smith MA, Tarone RE and
Devesa SS: Cancer surveillance series: Recent trends in childhood
cancer incidence and mortality in the United States. J Natl Cancer
Inst. 91:1051–1058. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Leavey PJ and Collier AB: Ewing sarcoma:
Prognostic criteria, outcomes and future treatment. Expert Rev
Anticancer Ther. 8:617–624. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Womer RB, West DC, Krailo MD, Dickman PS,
Pawel BR, Grier HE, Marcus K, Sailer S, Healey JH, Dormans JP, et
al: Randomized controlled trial of interval-compressed chemotherapy
for the treatment of localized Ewing sarcoma: A report from the
Children's Oncology Group. J Clin Oncol. 30:4148–4154. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Motoyama K, Inoue H, Takatsuno Y, Tanaka
F, Mimori K, Uetake H, Sugihara K and Mori M: Over- and
under-expressed microRNAs in human colorectal cancer. Int J Oncol.
34:1069–1075. 2009.PubMed/NCBI
|
|
6
|
Tong AW, Fulgham P, Jay C, Chen P, Khalil
I, Liu S, Senzer N, Eklund AC, Han J and Nemunaitis J: MicroRNA
profile analysis of human prostate cancers. Cancer Gene Ther.
16:206–216. 2009.
|
|
7
|
Hiyoshi Y, Kamohara H, Karashima R, Sato
N, Imamura Y, Nagai Y, Yoshida N, Toyama E, Hayashi N, Watanabe M,
et al: MicroRNA-21 regulates the proliferation and invasion in
esophageal squamous cell carcinoma. Clin Cancer Res. 15:1915–1922.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma J, Dong C and Ji C: MicroRNA and drug
resistance. Cancer Gene Ther. 17:523–531. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou P, Jiang W, Wu L, Chang R, Wu K and
Wang Z: miR-301a is a candidate oncogene that targets the homeobox
gene Gax in human hepatocellular carcinoma. Dig Dis Sci.
57:1171–1180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z, Chen LY, Dai HY, Wang P, Gao S and
Wang K: miR-301a promotes pancreatic cancer cell proliferation by
directly inhibiting Bim expression. J Cell Biochem. 113:3229–3235.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shi W, Gerster K, Alajez NM, Tsang J,
Waldron L, Pintilie M, Hui AB, Sykes J, P'ng C, Miller N, et al:
MicroRNA-301 mediates proliferation and invasion in human breast
cancer. Cancer Res. 71:2926–2937. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Miko E, Czimmerer Z, Csánky E, Boros G,
Buslig J, Dezso B and Scholtz B: Differentially expressed microRNAs
in small cell lung cancer. Exp Lung Res. 35:646–664. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jones N, Bonnet F, Sfar S, Lafitte M,
Lafon D, Sierankowski G, Brouste V, Banneau G, Tunon de Lara C,
Debled M, et al: Comprehensive analysis of PTEN status in breast
carcinomas. Int J Cancer. 133:323–334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mayo LD and Donner DB: The PTEN, Mdm2, p53
tumor suppressor-oncoprotein network. Trends Biochem Sci.
27:462–467. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chu EC and Tarnawski AS: PTEN regulatory
functions in tumor suppression and cell biology. Med Sci Monit.
10:RA235–RA241. 2004.PubMed/NCBI
|
|
18
|
Zeng N, Yang KT, Bayan JA, He L, Aggarwal
R, Stiles JW, Hou X, Medina V, Abad D, Palian BM, et al: PTEN
controls β-cell regeneration in aged mice by regulating cell cycle
inhibitor p16ink4a. Aging Cell. 12:1000–1011. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Li Z, Li N, Qi J, Fan K, Yin P, Zhao
C, Liu Y, Yao W, Cai X, et al: MAGI2 enhances the sensitivity of
BEL-7404 human hepatocellular carcinoma cells to
staurosporine-induced apoptosis by increasing PTEN stability. Int J
Mol Med. 32:439–447. 2013.PubMed/NCBI
|
|
20
|
Yang TS, Yang XH, Wang XD, Wang YL, Zhou B
and Song ZS: MiR-214 regulate gastric cancer cell proliferation,
migration and invasion by targeting PTEN. Cancer Cell Int.
13:682013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi Z, Zhang J, Qian X, Han L, Zhang K,
Chen L, Liu J, Ren Y, Yang M, Zhang A, et al: AC1MMYR2, an
inhibitor of dicer-mediated biogenesis of Oncomir miR-21, reverses
epithelial-mesenchymal transition and suppresses tumor growth and
progression. Cancer Res. 73:5519–5531. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma F, Zhang J, Zhong L, Wang L, Liu Y,
Wang Y, Peng L and Guo B: Upregulated microRNA-301a in breast
cancer promotes tumor metastasis by targeting PTEN and activating
Wnt/β-catenin signaling. Gene. 535:191–197. 2014. View Article : Google Scholar
|
|
24
|
Dou L, Wang S, Sui X, Meng X, Shen T,
Huang X, Guo J, Fang W, Man Y, Xi J, et al: MiR-301a mediates the
effect of IL-6 on the AKT/GSK pathway and hepatic glycogenesis by
regulating PTEN expression. Cell Physiol Biochem. 35:1413–1424.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Masliah-Planchon J, Pasmant E, Luscan A,
Laurendeau I, Ortonne N, Hivelin M, Varin J, Valeyrie-Allanore L,
Dumaine V, Lantieri L, et al: MicroRNAome profiling in benign and
malignant neurofibromatosis type 1-associated nerve sheath tumors:
Evidences of PTEN pathway alterations in early NF1 tumorigenesis.
BMC Genomics. 14:4732013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Di Cristofano A, De Acetis M, Koff A,
Cordon-Cardo C and Pandolfi PP: Pten and p27KIP1 cooperate in
prostate cancer tumor suppression in the mouse. Nat Genet.
27:222–224. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Podsypanina K, Ellenson LH, Nemes A, Gu J,
Tamura M, Yamada KM, Cordon-Cardo C, Catoretti G, Fisher PE and
Parsons R: Mutation of Pten/Mmac1 in mice causes neoplasia in
multiple organ systems. Proc Natl Acad Sci USA. 96:1563–1568. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu ZL, Wang H, Liu J and Wang ZX:
MicroRNA-21 (miR-21) expression promotes growth, metastasis, and
chemo- or radio-resistance in non-small cell lung cancer cells by
targeting PTEN. Mol Cell Biochem. 372:35–45. 2013. View Article : Google Scholar
|
|
29
|
Jiang J, Zhang Y, Yu C, Li Z, Pan Y and
Sun C: MicroRNA-492 expression promotes the progression of hepatic
cancer by targeting PTEN. Cancer Cell Int. 14:952014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim RJ, Bae E, Hong YK, Hong JY, Kim NK,
Ahn HJ, Oh JJ and Park DS: PTEN loss-mediated Akt activation
increases the properties of cancer stem-like cell populations in
prostate cancer. Oncology. 87:270–279. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Niemeyer BF, Parrish JK, Spoelstra NS,
Joyal T, Richer JK and Jedlicka P: Variable expression of PIK3R3
and PTEN in Ewing Sarcoma impacts oncogenic phenotypes. PLoS One.
10:e01168952015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang M, Li C, Yu B, Su L, Li J, Ju J, Yu
Y, Gu Q, Zhu Z and Liu B: Overexpressed miR-301a promotes cell
proliferation and invasion by targeting RUNX3 in gastric cancer. J
Gastroenterol. 48:1023–1033. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu Z and Li Y, Takwi A, Li B, Zhang J,
Conklin DJ, Young KH, Martin R and Li Y: miR-301a as an NF-κB
activator in pancreatic cancer cells. EMBO J. 30:57–67. 2011.
View Article : Google Scholar :
|
|
34
|
Guo SL, Ye H, Teng Y, Wang YL, Yang G, Li
XB, Zhang C and Yang X, Yang ZZ and Yang X: Akt-p53-miR-365-cyclin
D1/ cdc25A axis contributes to gastric tumorigenesis induced by
PTEN deficiency. Nat Commun. 4:25442013. View Article : Google Scholar
|
|
35
|
Ou Y, Ma L, Ma L, Huang Z, Zhou W, Zhao C,
Zhang B, Song Y, Yu C and Zhan Q: Overexpression of cyclin B1
antagonizes chemotherapeutic-induced apoptosis through PTEN/Akt
pathway in human esophageal squamous cell carcinoma cells. Cancer
Biol Ther. 14:45–55. 2013. View Article : Google Scholar :
|
|
36
|
Campos M, Kool MM, Daminet S, Ducatelle R,
Rutteman G, Kooistra HS, Galac S and Mol JA: Upregulation of the
PI3K/Akt pathway in the tumorigenesis of canine thyroid carcinoma.
J Vet Intern Med. 28:1814–1823. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Z, Hou X, Shao C, Li J, Cheng JX,
Kuang S, Ahmad N, Ratliff T and Liu X: Plk1 inhibition enhances the
efficacy of androgen signaling blockade in castration-resistant
prostate cancer. Cancer Res. 74:6635–6647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li J, Chen Y, Wan J, Liu X, Yu C and Li W:
ABT-263 enhances sorafenib-induced apoptosis associated with Akt
activity and the expression of Bax and p21((CIP1/WAF1)) in human
cancer cells. Br J Pharmacol. 171:3182–3195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vadlakonda L, Pasupuleti M and Pallu R:
Role of PI3K-AKT-mTOR and Wnt signaling pathways in transition of
G1-S phase of cell cycle in cancer cells. Front Oncol. 3:852013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen ZX, Sang QT, Du YG and Xin YY:
Silence of PTEN in colorectal cancer cells via siRNA inhibits cell
growth. J Environ Pathol Toxicol Oncol. 33:233–237. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nishioka C, Ikezoe T, Yang J and Yokoyama
A: Long-term exposure of leukemia cells to multi-targeted tyrosine
kinase inhibitor induces activations of AKT, ERK and STAT5
signaling via epigenetic silencing of the PTEN gene. Leukemia.
24:1631–1640. 2010. View Article : Google Scholar : PubMed/NCBI
|