Introduction
Hepatocellular carcinoma (HCC) is the dominant
pathologic type of primary liver cancer, which is the third leading
cause of cancer-related death worldwide (1). Despite recent advances in the
diagnosis and treatment of HCC, including hepatectomy and liver
transplantation, the prognosis for HCC patients remains poor due to
the high recurrence rate and early metastasis (2). However, the detailed mechanism
underlying the development and progression of HCC is still not
clear (3). Therefore, it is urgent
to clarify the molecular mechanisms of HCC and identify novel
prognostic biomarkers to provide potential therapeutic targets for
the patients with HCC.
Increasing evidence suggests that microRNAs
(miRNAs), a class of non-coding RNAs composed of ~22 nucleotides,
can act as suitable biomarkers with diagnostic, prognostic, and
predictive potential (4). miRNAs
may function as oncogenes or tumor suppressors by regulating
protein expression by interacting with complementary sites within
the 3′-untranslated region (UTR) of target mRNA transcripts
(5). Emerging studies have
documented that aberrant miRNAs play important roles in various
biological processes (6),
including cell proliferation, differentiation, apoptosis,
drug-resistance, migration and invasion (7–10).
Recently, miRNAs have shown high stability in tissues and body
fluids, which reveal their potential as tumor markers. In addition,
it has been widely recognized that dysregulation of miRNAs
contribute to the development and progression of HCC.
miR-155 is located within a region known as B cell
integration cluster on chromosome 21, which plays a critical role
in the progression of gastric cancer (11), colorectal carcinoma (12), lung cancer (13), breast cancer (14), bladder cancer (15) and B-cell lymphoma (16). Previous studies show that miR-155
functions as a regulator in the pathogenesis of cell proliferation,
apoptosis, drug-resistance, migration, invasion and
epithelial-mesenchymal transition (EMT). miR-155 promotes oral
squamous cell carcinoma metastasis and correlates a poor prognosis
(17). miR-155 promotes the B-cell
lymphoma cell proliferation and inhibits cell apoptosis by
targeting NIAM phenocopies (16).
Moreover, direct quantitative detection for cell-free miR-155 in
urine could be a potential novel biomarker in diagnosis and
prognosis for non-muscle invasive bladder cancer (15). miR-155 mediates anti-Warburg effect
of rosmarinic acid in colorectal carcinoma and gastric cancer
(18). miR-155 presents
tamoxifen-resistance by modulating SOCS6-STAT3 signaling pathway in
breast cancer (19). However, the
clinical significance of miR-155 and the underlying mechanisms
involved in the development of HCC remain to be investigated.
In this study, we demonstrated that the expression
of miR-155 was upregulated in HCC tissues and its high expression
was associated with poor clinicopathological features and the
reduced survival of HCC patients. miR-155 promoted cell
proliferation, cell cycle and apoptosis resistance in vitro.
Moreover, the downregulation of miR-155 inhibited tumor growth of
HCC in vivo. Notably, AT-rich interactive domain 2 (ARID2)
was identified as a direct target of miR-155. The results showed a
new role for miR-155 in prediction of prognosis and promoting tumor
growth of HCC.
Materials and methods
Clinical samples and cell lines
HCC samples (124) and matched normal tumor-adjacent
samples (>2 cm distance from the margin of the resection) were
obtained during surgery and used after obtaining informed consent.
All patients underwent resection of their primary HCC in the
Department of General Surgery at The Second Affiliated Hospital of
Xi'an Jiaotong University, from January 2006 to December 2008. None
of the patients received preoperative chemo- or radiotherapy. The
stage of cancer was determined according to the cancer staging
system published in 2010 by the Union for International Cancer
Control (UICC). The Xi'an Jiaotong University Ethics Committee
approved all protocols according to the 1975 Helsinki
Declaration.
Human HCC cell lines (Hep3B, Bel-7402, MHCC-97L,
HepG2, SMMC-7721) and human immortalized normal hepatic cell line
LO2 were obtained from the Institute of Biochemistry and Cell
Biology, Chinese Academy of Sciences (Shanghai, China). The cells
were maintained in Dulbecco's modified Eagle's medium (DMEM, Gibco,
Grand Island, NY, USA) containing 10% fetal bovine serum (FBS,
Gibco) with 100 U/ml penicillin and 100 μg/ml streptomycin (Sigma,
St. Louis, MO, USA) and cultured in a humidified 5% CO2
incubator at 37°C.
Real-time quantitative reverse
transcription polymerase chain reaction (qRT-PCR)
Total RNA was extracted from clinical specimens or
HCC cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's instructions. cDNA was synthesized
from 1 μg RNA with the PrimeScript RT Master Mix (Takara, Osaka,
Japan). The PCR amplification for the quantification of the miR-155
and U6 was performed using the TaqMan miRNA Reverse Transcription
kit (Applied Biosystems, Foster City, CA, USA) and TaqMan Human
miRNA Assay kit (Applied Biosystems). The relative expression of
miR-155 was shown as the fold difference relative to U6. qPCR
primer against mature miRNA miR-155 (HmiRQP0221) and Homo
sapiens snRNA U6 qPCR Primer (HmiRQP9001) were purchased from
GeneCopoeia (Guangzhou, China).
Western blot analysis
Total protein was extracted from whole cells and 40
μg of isolated protein was separated by 10% SDS-PAGE and
transferred onto a PVDF membrane (Bio-Rad Laboratories, Hercules,
CA, USA). The membranes were probed with antibodies: anti-Akt,
anti-p-Akt, anti-ARID2, anti-Cyclin D, and anti-p27 primary
antibodies (Cell Signaling, Danvers, MA, USA) overnight. Then the
membranes were incubated with the HRP-conjugated goat anti-mouse or
anti-rabbit IgG antibody (ZSGB-BIO, China). Protein bands were
visualized using an enhanced chemiluminescence kit (Amersham,
Little Chalfont, UK).
Immunohistochemical staining
Immunohistochemistry was performed on
paraformaldehyde-fixed paraffin sections. ARID2 (1:100, #13594 Cell
Signaling Technology, Inc.) antibody was used in
immunohistochemistry by a streptavidin peroxidase-conjugated
(SP-IHC) method. The percentage of positive tumor cells was graded
as: 0, <10%; 1, 10–30%; 2, 31–50%; 3, >50%.
Plasmids and cell transfection
miRNA vectors, including miR-155 expression vector
(HmiR0358-MR02), the control vector for miR-155 (CmiR0001-MR04 and
miR-control), miR-155 inhibitor (HmiR-AN0220-AM03 and anti-miR-155)
and the negative control for the miR-155 inhibitor
(CmiR-AN0001-AM04 and anti-miR-NC), and ARID2 expression plasmid
were purchased from GeneCopoeia. The targeted sequences for ARID2
siRNA sense, 5′-AGCTCCAATTCCTTGTGAAGTTTT-3′ and antisense,
5′-ACTTCACAAGGAATTGCAGCTTTT-3′ or a non-specific duplex
oligonucleotide as a negative control were produced by Sangon
Biotech Co., Ltd. (Shanghai, China). The cells were transfected
with the vectors mentioned above using Lipofectamine 2000 according
to the manufacturer's instructions (Invitrogen).
Cell cycle, proliferation and detection
of apoptosis
Flow cytometry was performed using the
fluorescence-activated cell sorting (FACS) Calibur and CellQuest
software (both from Becton-Dickinson, San Jose, CA, USA). For cell
cycle analysis, the cells were seeded in 6-well plates at
2×105/well. Forty-eight hours after transfection, the
cells were fixed in 70% ethanol at 4°C for 24 h and stained with 50
μg/ml propidium iodide (Keygen, Nanjing, China). An Annexin V-Fluor
Staining kit (Roche) was used to analyze apoptosis levels. For the
proliferation assay, bromodeoxyuridine labeling and
immunofluorescence was used. Cells grown on coverslips (Fisher,
Pittsburgh, PA, USA) were incubated with bromodeoxyuridine (BrdU)
for 1 h and stained with anti-BrdU antibody (Sigma) according to
the manufacturer's instructions. Gray level images were acquired
under a laser scanning microscope (Axioskop 2 plus, Carl Zeiss Co.
Ltd., Jena, Germany).
Luciferase reporter assay
The 3′-UTR sequence of ARID2 predicted to interact
with miR-155 or the mutated sequence within the predicted target
sites was synthesized and inserted into the pGL3 control vector
(Promega, Madison, WI, USA). These constructs were named as wt
ARID2-3′UTR or mt ARID2-3′UTR, respectively. Then, SMMC-7721 cells
(1×105) were seeded into 24-well plates, and were
cultured in OptimMEM reduced serum media (Life Technologies) as per
the recommended conditions, and were cotransfected with 200 ng of
each luciferase reporter construct (the wt or mt 3′-UTR of ARID2
mRNA) and miR-155 expression vector, miR-155 inhibitor, control
vector or negative control (50 nM) using FuGENE (Promega). After 48
h, the cells were harvested and luciferase activity was measured
using the dual-luciferase reporter assay system (Promega). Firefly
luciferase activity was normalized to the Renilla luciferase
activity. Results were obtained from three independent experiments
performed in triplicate.
In vivo experiments
Four-to-six-week-old female BALB/c nude mice (Centre
of Laboratory Animals, The Medical College of Xi'an Jiaotong
University, Xi'an, China) were used to establish the nude mouse
xenograft model. SMMC-7721 (5×106) cells that were
transfected with anti-miR-155 or anti-miR-NC vectors were mixed in
150 μl of Matrigel and were inoculated subcutaneously into the
flank of nude mice. The tumor volume for each mouse was determined
by measuring two of its dimensions and then calculated as tumor
volume = length × width × width/2. After 3 weeks, the mice were
sacrificed by cervical dislocation under anesthesia with ether and
the xenograft tumor tissue was explanted for examination. Animal
protocols were approved by the Institutional Animal Care and Use
Committee of Xi'an Jiaotong University.
Statistical analysis
Data are presented as the mean ± SD from at least
three independent replicates. SPSS software, 16.0 (SPSS, Inc,
Chicago, IL, USA) was used to conduct the analysis, and a
two-tailed Student's t-test was employed to analyze the differences
between two groups. Pearson's correlation analysis was used to
analyze the correlation between two indices. Survival curves were
plotted by the Kaplan-Meier method and compared by the log-rank
test. Differences were considered statistically significant at
P<0.05.
Results
miR-155 is upregulated in HCC tissues and
cells
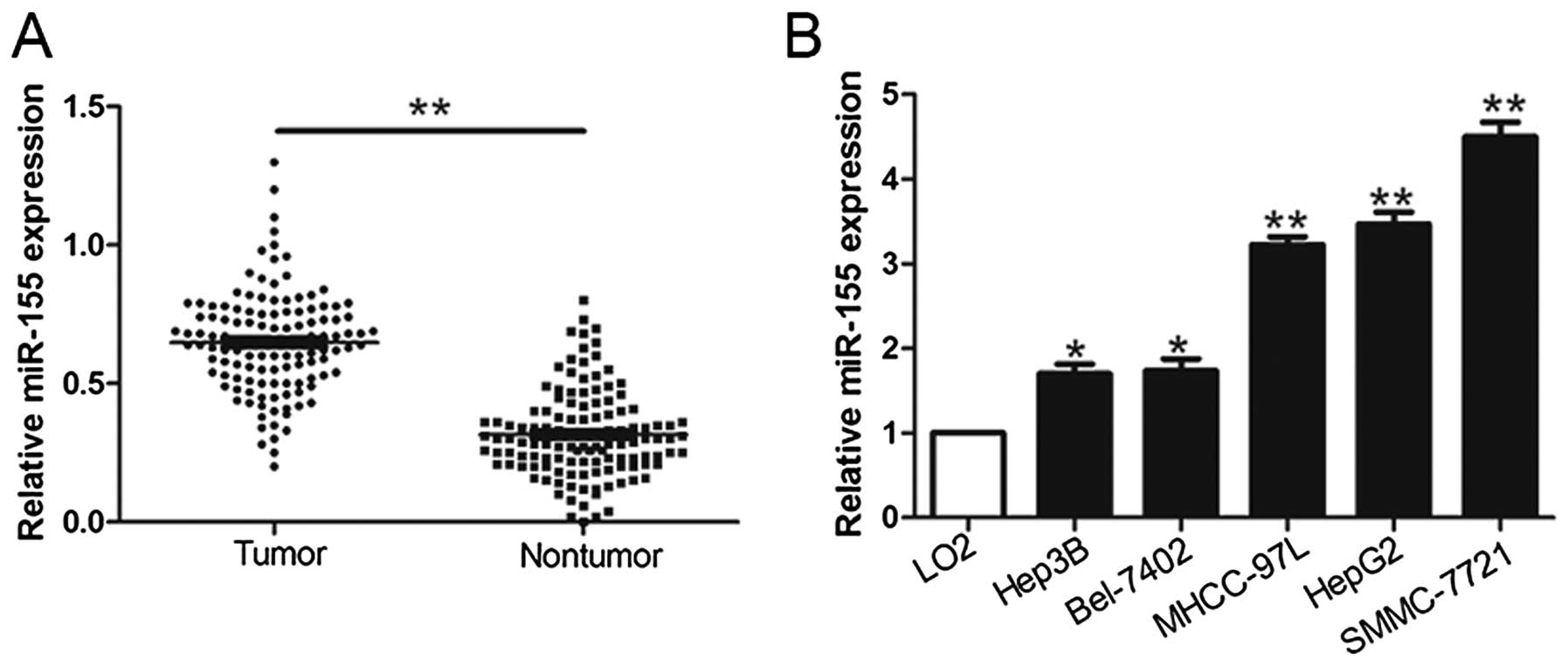
To evaluate the potential role of miR-155 in HCC, we
first quantified miR-155 in 124 pairs of HCC tissues and matched
adjacent non-tumor tissues using qRT-PCR methods. As shown in
Fig. 1A, miR-155 was significantly
increased in the HCC tissues compared with adjacent non-tumor
tissues. Furthermore, we determined the expression level of miR-155
in HCC cell lines and the normal hepatocyte cell line LO2.
Similarly, miR-155 was significantly upregulated in all HCC cell
lines compared with LO2 cells (P<0.05, Fig. 1B). These results suggest that
miR-155 expression is upregulated in HCC and may contribute to the
development of HCC.
Clinical significance of elevated miR-155
expression in HCC specimens
To investigate the clinical significance of miR-155
in HCC, the patients were classified into high and low miR-155
expression subgroups with the median level of miR-155 as the
cut-off. As shown in Table I,
higher levels of miR-155 were significantly correlated with a large
tumor size (≥5 cm; P=0.001), high histological grade
(Edmondson-Steiner grade III + IV; P=0.021) and advanced tumor
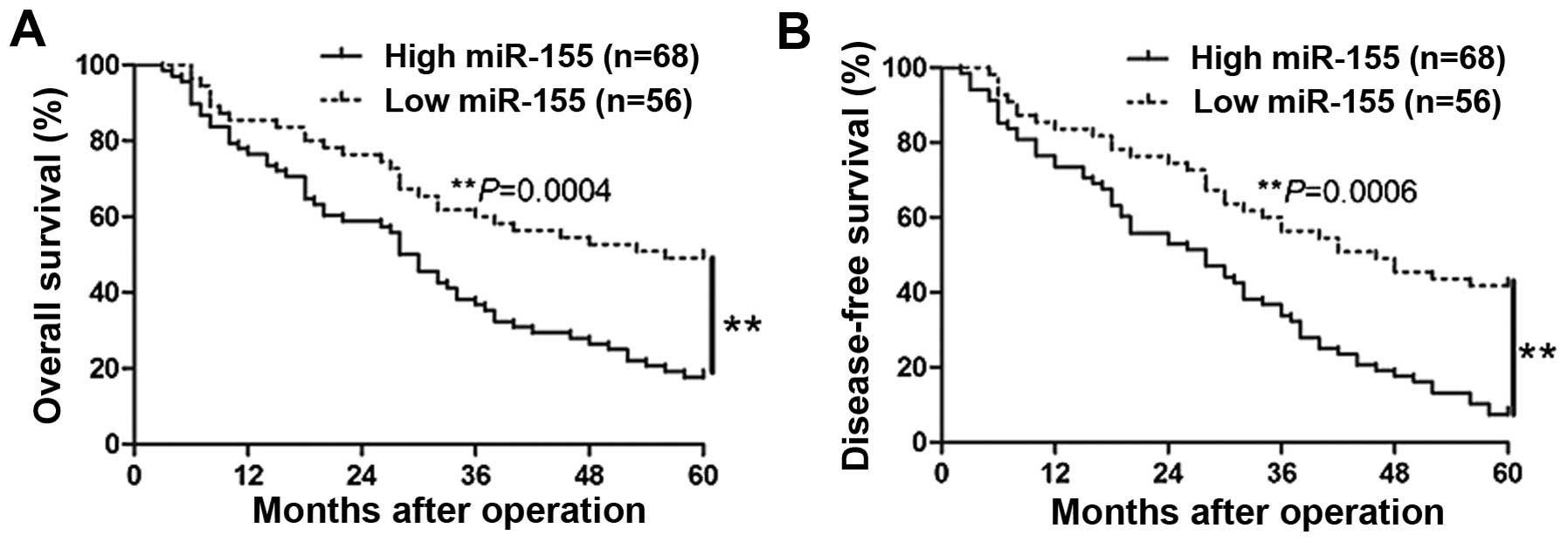
stage (TNM stage III + IV; P=0.020) (Table I). To further evaluate the
significant contribution of miR-155 expression in the prognosis of
patients with HCC, we constructed Kaplan-Meier survival curves
using the overall 5-year patient survival data to analyze cases
with high and low miR-155 expression. As shown in Fig. 2, patients with high miR-155
expression exhibited worse overall survival (P=0.0004) and
disease-free survival (P=0.0006). These data indicate that miR-155
could serve as a valuable indicator for predicting the prognosis of
HCC.
 | Table IClinical correlation of miR-155
expression in HCC (n=124). |
Table I
Clinical correlation of miR-155
expression in HCC (n=124).
| | Expression level | |
|---|
| |
| |
|---|
| Clinical
parameters | Cases (n) |
miR-155high (n=68) | miR-155low
(n=56) | P-value
(p<0.05)a |
|---|
| Age | | | | |
| <50 years | 39 | 20 | 19 | 0.698 |
| ≥50 years | 85 | 48 | 37 | |
| Gender | | | | |
| Male | 92 | 50 | 42 | 0.852 |
| Female | 32 | 18 | 14 | |
| Tumor size
(cm) | | | |
0.001a |
| <5 cm | 88 | 40 | 48 | |
| ≥5 cm | 36 | 28 | 8 | |
| Tumor number | | | | 0.947 |
| Solitary | 106 | 58 | 48 | |
| Multiple | 18 | 10 | 8 | |
| Edmondson | | | | |
| I+II | 77 | 36 | 41 |
0.021a |
| III+IV | 47 | 32 | 15 | |
| TNM stage | | | |
0.020a |
| I+II | 102 | 51 | 51 | |
| III+IV | 22 | 17 | 5 | |
| Capsular
infiltration | | | | 0.826 |
| Present | 81 | 45 | 36 | |
| Absent | 43 | 23 | 20 | |
| Venous
infiltration | | | | 0.668 |
| Present | 15 | 9 | 6 | |
| Absent | 109 | 59 | 50 | |
| AFP | | | | 0.569 |
| <400 ng/ml | 41 | 21 | 20 | |
| ≥400 ng/ml | 83 | 47 | 36 | |
| HBsAg | | | | 0.964 |
| Positive | 115 | 63 | 52 | |
| Negative | 9 | 5 | 4 | |
miR-155 promotes cell cycle progression,
cell proliferation and inhibits apoptosis in HCC cells
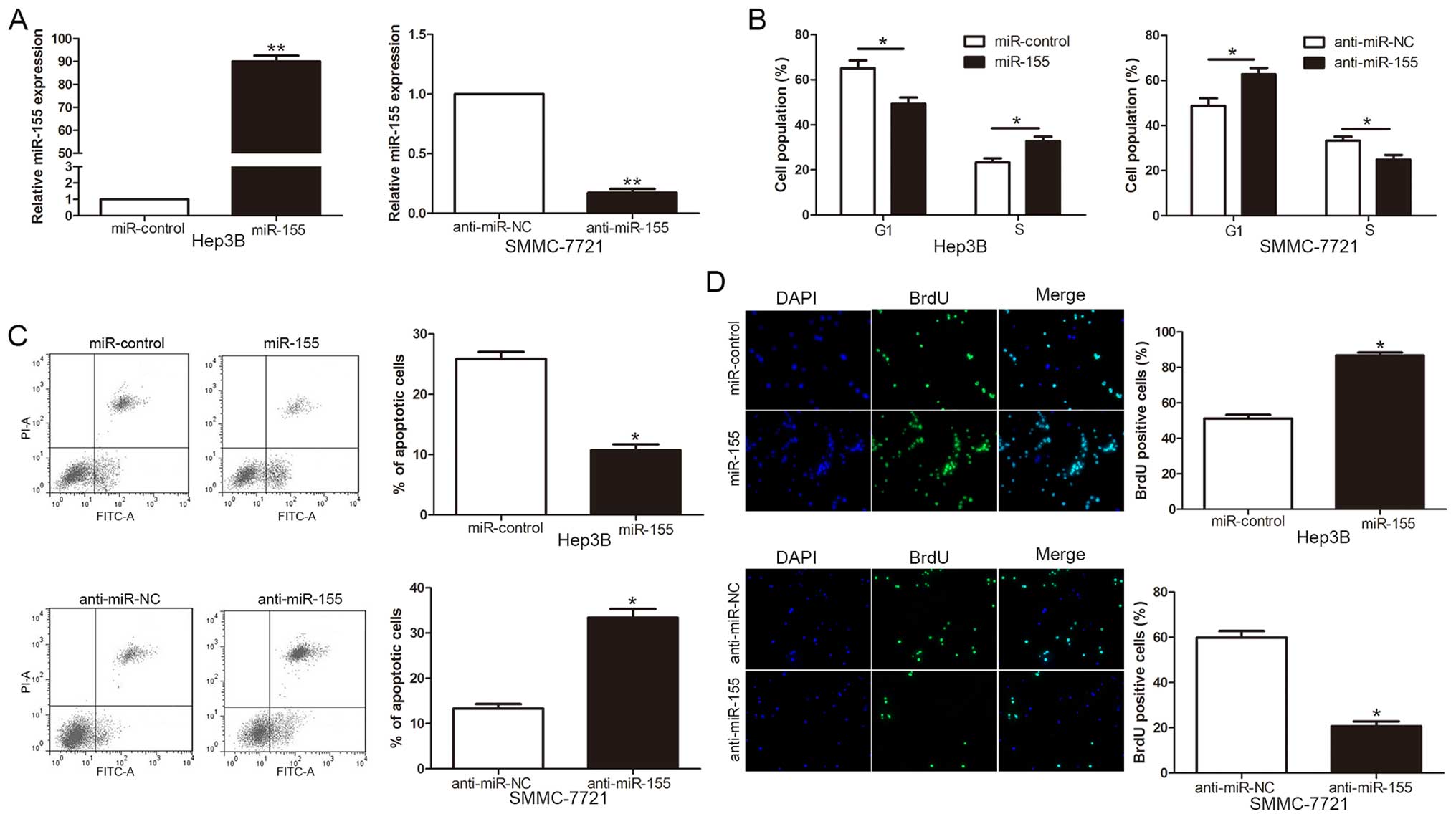
To investigate the biological function of miR-155 in
the development and progression of HCC, we transduced a miR-155
expression vector or a miR-155 inhibitor (anti-miR-155) into Hep3B
and SMMC-7721 cells, respectively. As measured by qRT-PCR, miR-155
expression vector significantly increased the level of miR-155 in
Hep3B cells, while the anti-miR-155 vector significantly reduced
the expression of miR-155 in SMMC-7721 cells (P<0.01, Fig. 3A). As determined by flow cytometric
analysis, the upregulation of miR-155 promoted cell cycle
transition from G1 to S phase (P<0.05, Fig. 3B) and apoptosis resistance
(P<0.05, Fig. 3C) in Hep3B
cells. Furthermore, the overexpression of miR-155 significantly
increased cell proliferation examined with incorporation assay in
Hep3B cells (P<0.05, Fig. 3D).
By contrast, the downregulation of miR-155 resulted in G1 arrest,
apoptosis promotion and proliferation reduction in SMMC-7721 cells
(P<0.05, respectively, Fig.
3B–D). These results demonstrated that miR-155 regulates the
cell cycle progression, apoptosis and proliferation of HCC
cells.
ARID2 is a direct target of miR-155 in
HCC
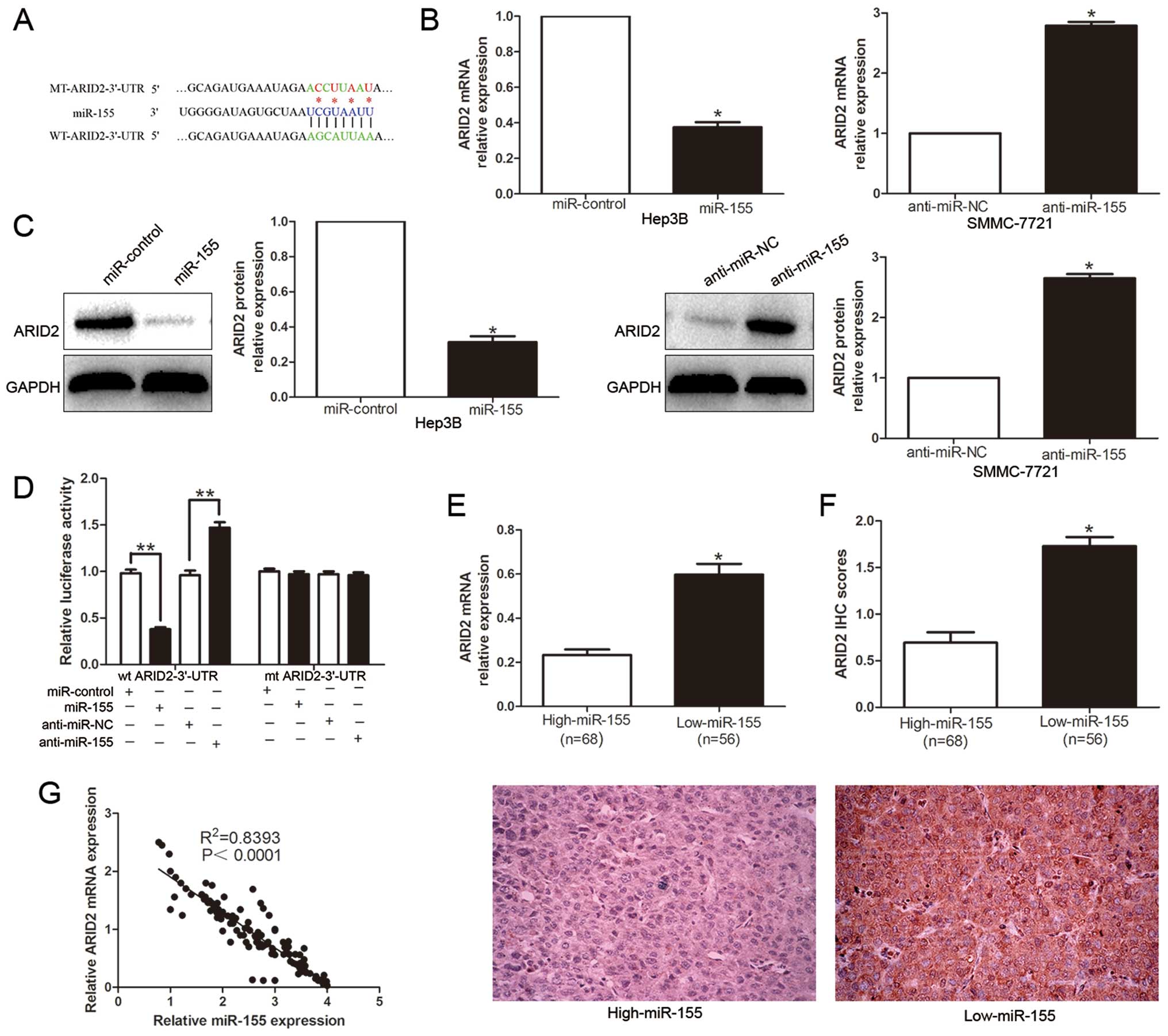
To explore the mechanism of miR-155 regulation in
HCC, we used publicly available databases TargetScan 6.2 and
miRanda to search predicted genes. Among them, the binding sites
for miR-155 on the 3′-UTR of ARID2 were conserved among species
(Fig. 4A). To verify the
regulation role of miR-155 on ARID2, qRT-PCR and western blotting
were performed to detect the effect of miR-155 on ARID2 mRNA and
protein levels. Ectopic expression of miR-155 markedly decreased,
while inhibition of miR-155 increased the ARID2 mRNA (P<0.05,
Fig. 4B) and protein (P<0.05,
Fig. 4C). In addition, the
over-expression of miR-155 prominently inhibited the luciferase
activity of ARID2 containing a wild-type (wt) 3′-UTR but did not
suppress the activity of ARID2 with a mutant (mt) 3′-UTR
(P<0.01, Fig. 4D). Suppression
of miR-155 by anti-miR-155 increased the luciferase activity of wt
ARID2 3′-UTR (P<0.01, Fig. 4D).
However, with the mt ARID2 3′-UTR constructs, there was no relative
increase in activity. Moreover, the expression levels of ARID2 mRNA
and protein in the high miR-155 expression tumors were
significantly lower than those in the low miR-155 expression tumors
(P<0.05, respectively, Fig. 4E and
F). Notably, the expression level of miR-155 was inversely
correlated with the level of ARID2 mRNA in HCC tissues
(R2=0.8393, P<0.0001, Fig. 4G). On the basis of these data, we
conclude that ARID2 is a direct target gene for miR-155 and that
miR-155 downregulates ARID2 expression.
Altering expression of ARID2 influences
the effect of miR-155 on HCC cells
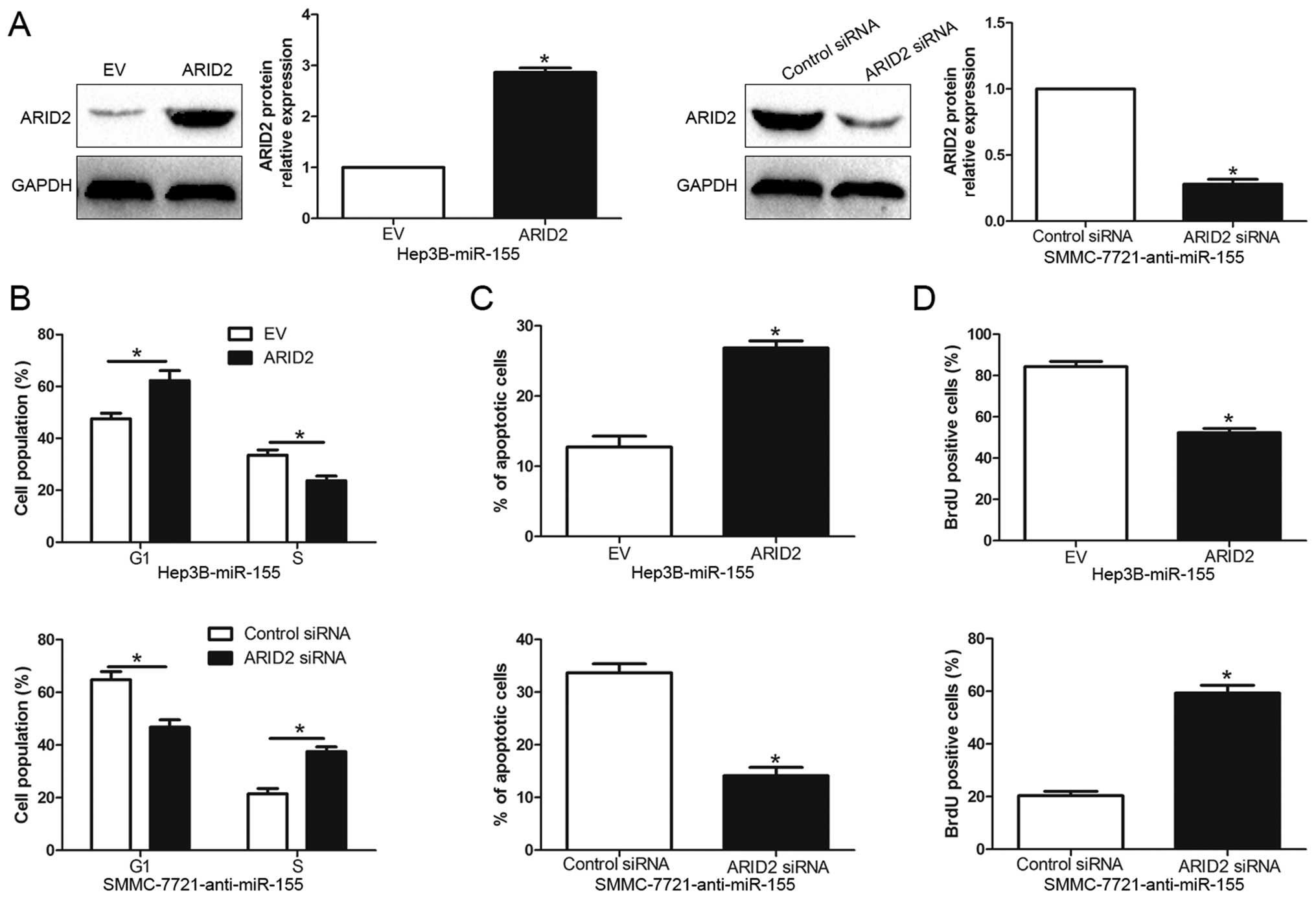
To confirm that ARID2 is a functional target of
miR-155, we restored ARID2 expression in Hep3B-miR-155 cells by
transfecting ARID2 expression plasmid (P<0.05, Fig. 5A). Functionally, restoration of
ARID2 expression in Hep3B-miR-155 cells partially abrogated the
effect of exogenous miR-155, resulting in significant increase of
apoptosis (P<0.01, Fig. 5C) and
obvious decrease of cell cycle progression and cell proliferation
(P<0.05, respectively, Fig. 5B and
D). Similarly, silencing of ARID2 in SMMC-7721-anti-miR-155
cells partially abolished the effect of anti-miR-155 on cell cycle,
apoptosis and proliferation (P<0.05, respectively, Fig. 5). These results demonstrate that
ARID2 is a downstream mediator for the function of miR-155 in
HCC.
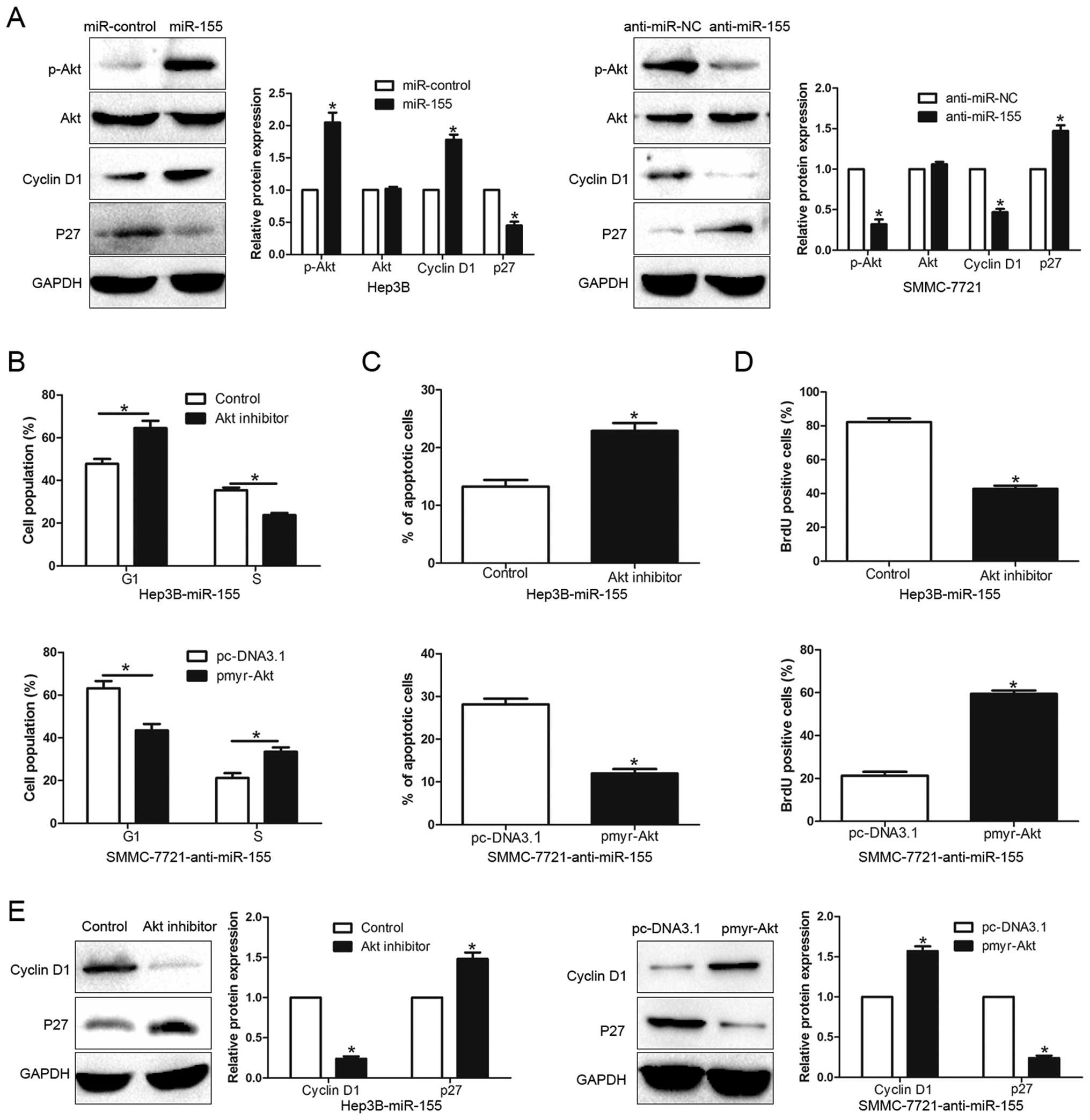
Akt phosphorylation is essential for the
biological function of miR-155 in HCC
Previous studies demonstrated that activation of Akt
signaling played an important role in HCC cell cycle progression,
apoptosis and proliferation (20–22),
so we further investigated the underlying molecular mechanisms of
the miR-155-mediated promotion of HCC biological effects. As shown
in Fig. 6A, ectopic expression of
miR-155 significantly increased, while miR-155 inhibition
decreased, the Akt phosphorylation in HCC cells (P<0.05).
Consistently, the expression of Cyclin D1 and p27, which are the
downstream effectors of Akt signaling and the key regulators of
cell cycle progression and proliferation in HCC, were also altered
in the up- or down-expression of miR-155 HCC cells (P<0.05).
Furthermore, to confirm that Akt phosphorylation contributed to the
biological function of miR-155-mediated in HCC cells, we used Akt
inhibitor or pmyr-Akt (dominant-active Akt) plasmid to affect Akt
activation. Inactivation of Akt phosphorylation by Akt inhibitor
significantly decreased cell cycle progression (P<0.05, Fig. 6B) and proliferation (P<0.05,
Fig. 6D) and induced apoptosis
(P<0.05, Fig. 6C) in
Hep3B-miR-155 cells. In addition, Akt overexpression of
phosphorylation increased cell cycle progression, cell
proliferation and inhibited apoptosis (P<0.05, respectively,
Fig. 6B–D) in
SMMC-7721-anti-miR-155 cells. Accordingly, the expression of Cyclin
D1 and p27 was also significantly altered (P<0.05, Fig. 6E). Taken together, our results
demonstrate that Akt phosphorylation exerts an important role in
miR-155-mediated HCC cell cycle progression, apoptosis and
proliferation.
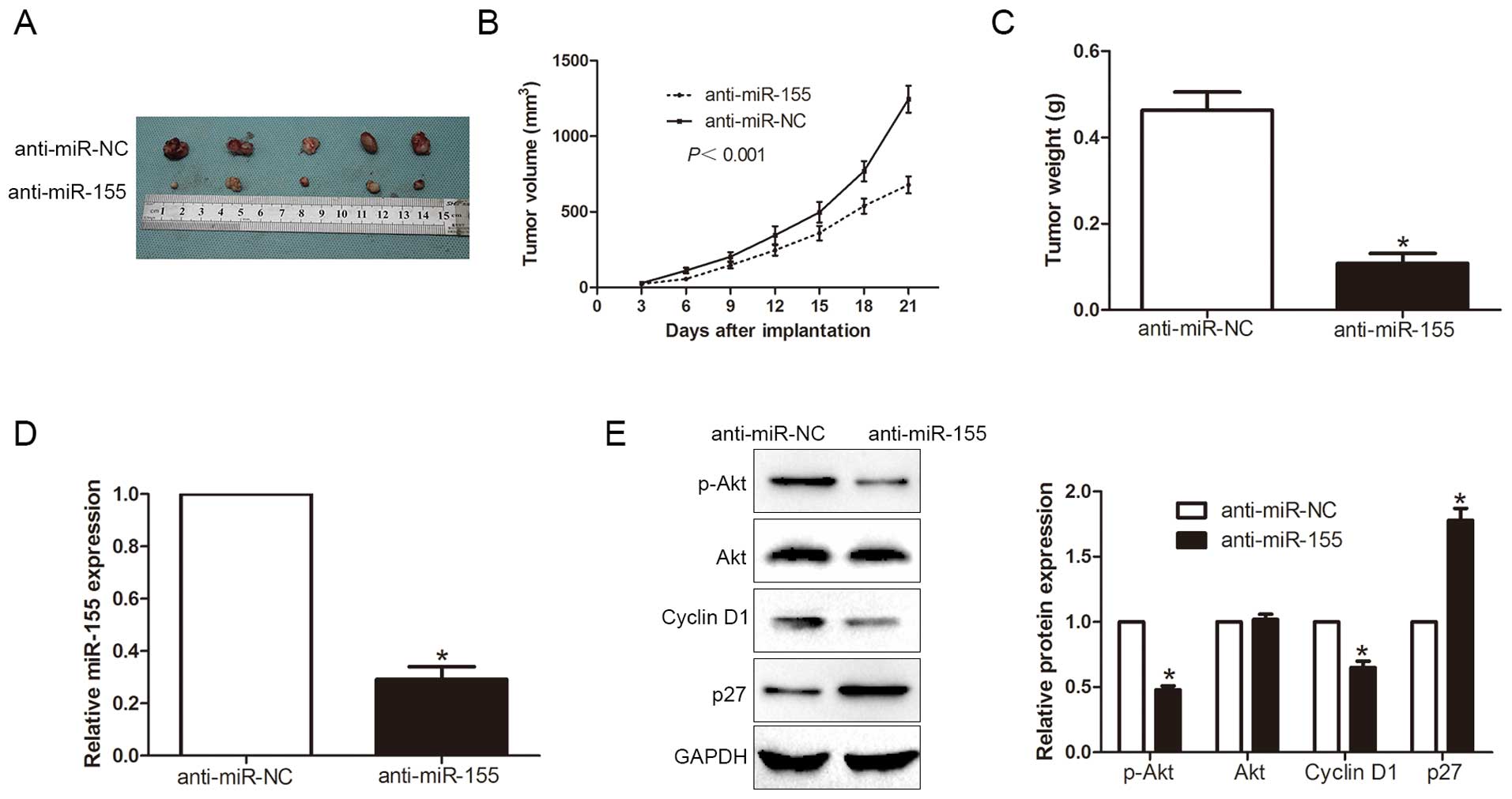
miR-155 promotes HCC cell cycle
progression, proliferation and inhibits apoptosis in vivo
To further confirm our results in vivo,
SMMC-7721 cells were transfected with anti-miR-155 or anti-miR-NC
vector and implanted subcutaneously into nude mice (Fig. 7A). Notably, the tumor growth curve
revealed that knockdown of miR-155 significantly retarded the tumor
growth of human HCC in the subcutaneous nude mouse model
(P<0.001, Fig. 7B). As shown in
Fig. 7C, in the miR-155 knockdown
group, the tumor weight was smaller than that in the control group.
Furthermore, the downregulation of miR-155 in the tumor formation
in the subcutaneous model (P<0.05, Fig. 7D), inhibited the activation of Akt
signaling (P<0.05, Fig. 7E).
Taken together, these results demonstrated that miR-155 regulates
the cell cycle progression, apoptosis and cell proliferation of HCC
cells and contributed to the development and progression of
HCC.
Discussion
miRNAs are known to be regulators of many genes at
the post-transcript level, and their aberrant expression is related
to cancer initiation, development and prognosis (23). However, the molecular mechanisms by
which miRNAs modulate the biological function of cancer cells are
still largely unknown. In previous studies, miR-155 controls
affinity-based selection by protecting c-MYC+ B cells from
apoptosis (24). Lerner et
al indicated that miR-155 was upregulated in blood, tissue and
cell lines of head and neck squamous cell carcinoma and could serve
as an independent prognosis predictor for patients and promoted the
cell proliferation and migration (25). Moreover, Hou et al found
that miR-155 promoted non-small cell lung carcinoma proliferation
through inhibition of FoxO1 and the subsequent increase of ROS
generation (26).
In this study, we initially investigated the
expression of miR-155 in 124 paired samples of HCC and non-tumor
tissues. Our data showed that the expression of miR-155 was
elevated in HCC tissues. Furthermore, miR-155 was upregulated in
HCC cell lines as compared with a normal hepatic cell line. These
results indicate that miR-155 may be an oncogene and may play a
critical role in hepatocarcinogenesis. Importantly, its
upregulation significantly correlated with the poorer prognosis of
patients with HCC. In addition, elevated miR-155 expression was
significantly correlated with malignant clinicopathological featurs
of HCC, including large tumor size, high histological grade and TNM
stage. miR-155 has been found to promote proliferation of lung
cancer by repressing FoxO1. We found that the downregulation of
miR-155 inhibited HCC cell proliferation and induced G1 phase
arrest and apoptosis in vitro. By contrast, the
overexpression of miR-155 promoted proliferation and cell cycle
transition to S phase and inhibited apoptosis. These results
suggested that miR-155 is a novel tumor-promoting miRNA that plays
a critical role in the regulation of tumor growth in HCC.
To further understand the underlying mechanisms by
which miR-155 exerts its biological effects on HCC cells, it is
necessary to identify its downstream functional targets. ARID2, a
new tumor suppressor gene in HCC, was initially identified in the
Polybromo-associated BRG1-associated factor (PBAF) complex
(27), an SWI/SNF chromatin
remodeling complex involved in ligand-dependent transcriptional
activation by nuclear receptors (28). Downregulated expression of ARID2
has been observed in liver cancer cells (29). We identified ARID2 as a novel
direct target of miR-155. Firstly, miR-155 negatively regulated
ARID2 mRNA and protein in HCC cells. Secondly, overexpression of
miR-155 decreased, while downregulation of miR-155 increased, the
luciferase reporter activity of ARID2 wt 3′-UTR, but not mt 3′-UTR.
Thirdly, miR-155 was inversely correlated with the levels of both
ARID2 mRNA and protein in HCC tissues. Finally, the effects of
miR-155 alteration on cell cycle progression, apoptosis and
proliferation were also abolished by ARID2 modulation.
Collectively, our results support that miR-155 exerts its
biological function on HCC through targeting ARID2.
It has been well-established that Akt
phosphorylation could promote cell cycle progression and
proliferation through upregulation of Cyclin D1 and downregulation
of p27 (22). As miR-155 plays a
critical role in cell fate, a better understanding of its
regulatory axis might provide new opportunities for developing more
effective therapeutic approaches to treat cancers. In addition, we
discovered that miR-155 activated Akt phosphorylation through
targeting ARID2. Consistently, we performed loss-experiment by Akt
inhibitor, gain-experiment by dominant-active Akt plasmid to
confirm that Akt phosphorylation plays an important role in the
miR-155-mediated biological function. In addition, we discovered
miR-155 activated Akt signaling through targeting ARID2 in
vivo, suggesting miR-155 may represent as a potential
therapeutic target for HCC treatment.
In conclusion, we find that miR-155 is upregulated
in HCC tissues and cells, and its elevated expression is associated
with poor prognostic features. In vitro and in vivo
studies indicate that miR-155 promotes tumor growth by promoting
HCC cell cycle progression, cell proliferation and inhibiting
apoptosis. Mechanistically, we suggest that miR-155 exerts its
biological function through Akt phosphorylation by targeting ARID2.
Therefore, miR-155 has the potential to be a valuable diagnostic
and prognostic biomarker for HCC.
Acknowledgements
This study was supported by the Science and
Technology Research and Development Program of Shaanxi Province
[2013k-12].
References
|
1
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang ZY: Hepatocellular carcinoma surgery
- review of the past and prospects for the 21st century. J Surg
Oncol. 91:95–96. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lujambio A and Lowe SW: The microcosmos of
cancer. Nature. 482:347–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yates LA, Norbury CJ and Gilbert RJ: The
long and short of microRNA. Cell. 153:516–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Corsini LR, Bronte G, Terrasi M, Amodeo V,
Fanale D, Fiorentino E, Cicero G, Bazan V and Russo A: The role of
microRNAs in cancer: Diagnostic and prognostic biomarkers and
targets of therapies. Expert Opin Ther Targets. 16(Suppl 2):
S103–S109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chang RM, Yang H, Fang F, Xu JF and Yang
LY: MicroRNA-331-3p promotes proliferation and metastasis of
hepatocellular carcinoma by targeting PH domain and leucine-rich
repeat protein phosphatase. Hepatology. 60:1251–1263. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tu K, Liu Z, Yao B, Han S and Yang W:
MicroRNA-519a promotes tumor growth by targeting PTEN/PI3K/AKT
signaling in hepatocellular carcinoma. Int J Oncol. 48:965–974.
2016.
|
|
9
|
Dou C, Wang Y, Li C, Liu Z, Jia Y, Li Q,
Yang W, Yao Y, Liu Q and Tu K: MicroRNA-212 suppresses tumor growth
of human hepatocellular carcinoma by targeting FOXA1. Oncotarget.
6:13216–13228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chai ZT, Kong J, Zhu XD, Zhang YY, Lu L,
Zhou JM, Wang LR, Zhang KZ, Zhang QB, Ao JY, et al: MicroRNA-26a
inhibits angiogenesis by down-regulating VEGFA through the
PIK3C2α/Akt/HIF-1α pathway in hepatocellular carcinoma. PLoS One.
8:e779572013. View Article : Google Scholar
|
|
11
|
Sun S, Sun P, Wang C and Sun T:
Downregulation of microRNA-155 accelerates cell growth and invasion
by targeting c-myc in human gastric carcinoma cells. Oncol Rep.
32:951–956. 2014.PubMed/NCBI
|
|
12
|
Li T, Yang J, Lv X, Liu K, Gao C, Xing Y
and Xi T: miR-155 regulates the proliferation and cell cycle of
colorectal carcinoma cells by targeting E2F2. Biotechnol Lett.
36:1743–1752. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang F, Zhou J, Zhang Y, Wang Y, Cheng L,
Bai Y and Ma H: The value of microRNA-155 as a prognostic factor
for survival in non-small cell lung cancer: A meta-analysis. PLoS
One. 10:e01368892015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bertoli G, Cava C and Castiglioni I:
MicroRNAs: New biomarkers for diagnosis, prognosis, therapy
prediction and therapeutic tools for breast cancer. Theranostics.
5:1122–1143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang X, Zhang Y, Liu X, Fang A, Wang J,
Yang Y, Wang L, Du L and Wang C: Direct quantitative detection for
cell-free miR-155 in urine: A potential role in diagnosis and
prognosis for non-muscle invasive bladder cancer. Oncotarget.
7:3255–3266. 2016.
|
|
16
|
Slezak-Prochazka I, Kluiver J, de Jong D,
Smigielska-Czepiel K, Kortman G, Winkle M, Rutgers B, Koerts J,
Visser L, Diepstra A, et al: Inhibition of the miR-155 target NIAM
phenocopies the growth promoting effect of miR-155 in B-cell
lymphoma. Oncotarget. 7:2391–2400. 2016.
|
|
17
|
Baba O, Hasegawa S, Nagai H, Uchida F,
Yamatoji M, Kanno NI, Yamagata K, Sakai S, Yanagawa T and Bukawa H:
MicroRNA-155-5p is associated with oral squamous cell carcinoma
metastasis and poor prognosis. J Oral Pathol Med. Aug 26–2015.(Epub
ahead of print). PubMed/NCBI
|
|
18
|
Han S, Yang S, Cai Z, Pan D, Li Z, Huang
Z, Zhang P, Zhu H, Lei L and Wang W: Anti-Warburg effect of
rosmarinic acid via miR-155 in gastric cancer cells. Drug Des Devel
Ther. 9:2695–2703. 2015.PubMed/NCBI
|
|
19
|
Shen R, Wang Y, Wang CX, Yin M, Liu HL,
Chen JP, Han JQ and Wang WB: MiRNA-155 mediates TAM resistance by
modulating SOCS6-STAT3 signalling pathway in breast cancer. Am J
Transl Res. 7:2115–2126. 2015.PubMed/NCBI
|
|
20
|
Hou YQ, Yao Y, Bao YL, Song ZB, Yang C,
Gao XL, Zhang WJ, Sun LG, Yu CL, Huang YX, et al: Juglanthraquinone
C induces intracellular ROS increase and apoptosis by activating
the Akt/Foxo signal pathway in HCC cells. Oxid Med Cell Longev.
2016:49416232016. View Article : Google Scholar
|
|
21
|
Ewald F, Nörz D, Grottke A, Bach J,
Herzberger C, Hofmann BT, Nashan B and Jücker M: Vertical targeting
of AKT and mTOR as well as dual targeting of AKT and MEK signaling
is synergistic in hepatocellular carcinoma. J Cancer. 6:1195–1205.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu X, Liao W, Yuan Q, Ou Y and Huang J:
TTK activates Akt and promotes proliferation and migration of
hepatocellular carcinoma cells. Oncotarget. 6:34309–34320.
2015.PubMed/NCBI
|
|
23
|
Murakami Y, Yasuda T, Saigo K, Urashima T,
Toyoda H, Okanoue T and Shimotohno K: Comprehensive analysis of
microRNA expression patterns in hepatocellular carcinoma and
non-tumorous tissues. Oncogene. 25:2537–2545. 2006. View Article : Google Scholar
|
|
24
|
Nakagawa R, Leyland R, Meyer-Hermann M, Lu
D, Turner M, Arbore G, Phan TG, Brink R and Vigorito E:
MicroRNA-155 controls affinity-based selection by protecting
c-MYC+ B cells from apoptosis. J Clin Invest.
126:377–388. 2016. View
Article : Google Scholar
|
|
25
|
Lerner C, Wemmert S, Bochen F, Kulas P,
Linxweiler M, Hasenfus A, Heinzelmann J, Leidinger P, Backes C,
Meese E, et al: Characterization of miR-146a and miR-155 in blood,
tissue and cell lines of head and neck squamous cell carcinoma
patients and their impact on cell proliferation and migration. J
Cancer Res Clin Oncol. Nov 30–2015.(Epub ahead of print).
PubMed/NCBI
|
|
26
|
Hou L, Chen J, Zheng Y and Wu C: Critical
role of miR-155/FoxO1/ROS axis in the regulation of non-small cell
lung carcinomas. Tumour Biol. Nov 9–2015.(Epub ahead of print).
|
|
27
|
You J, Yang H, Lai Y, Simon L, Au J and
Burkart AL: ARID2, p110α, p53, and β-catenin protein expression in
hepatocellular carcinoma and clinicopathologic implications. Hum
Pathol. 46:1068–1077. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Raab JR, Resnick S and Magnuson T:
Genome-wide transcriptional regulation mediated by biochemically
distinct SWI/SNF complexes. PLoS Genet. 11:e10057482015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu P, Wu D, You Y, Sun J, Lu L, Tan J and
Bie P: miR-208-3p promotes hepatocellular carcinoma cell
proliferation and invasion through regulating ARID2 expression. Exp
Cell Res. 336:232–241. 2015. View Article : Google Scholar : PubMed/NCBI
|