Introduction
Acute lymphoblastic leukaemia (ALL) is a malignant
clonal proliferation of lymphoid progenitor cells (1). There are two main immunological types
of ALL; pre-B cell and pre-T cell. ALL is the most common type of
childhood cancer (2). It
represents 75% of childhood leukaemia whereas it is the least
common type of adult leukaemia. The American Cancer Society
estimates that 3,000 children and adolescents will be diagnosed
with ALL this year (ACS website). Advances in the understanding of
the molecular genetics and pathogenesis of the disease together
with the development of novel treatment strategies have contributed
to pediatric cure rates of up to 90% (1,3).
However, some patients elude this due to presentation of poor
prognostic factors and acquired drug resistance. Furthermore, the
modern cure rates for adult ALL have yet to exceed 40% and lag far
behind those observed in childhood ALL (1). In order to maintain current remission
rates in children and to improve the moderate cure rates for adults
there is a need to develop novel chemotherapeutic agents for the
treatment of ALL.
Death receptors (DRs) are members of the tumour
necrosis factor (TNF) receptor superfamily. These receptors are
capable of activating the cells extrinsic apoptotic pathways in
response to extracellular death signals (4). Eight human DRs have been identified
to date (5). Two DRs (DR4 and DR5)
are preferentially expressed on cancer cells (6,7).
Ligands targeting these death receptors and/or molecules that
induce expression of these receptors can be exploited to
selectively induce cell death in cancer cells whilst sparing normal
healthy cells (6). The TNF family
member, tumor necrosis factor-related apoptosis-inducing ligand
(TRAIL), is a potent ligand for DR4 and DR5 (8). Non-functional decoy TRAIL receptors
(DcR1 and DcR2) are expressed on normal cells protecting them from
the TRAIL death signal. Hence, TRAIL can selectively induce
apoptosis in a vast array of tumour-derived cell lines and
consequently inhibit tumour growth in various animal models
(9). This discovery instigated the
rapid development of therapeutic agents including recombinant human
TRAIL and agonistic monoclonal antibodies against DR4 and DR5
(10,11). However, despite the initial appeal
of TRAIL as a selective anticancer agent many cancers including
hematological malignancies are resistant to TRAIL monotherapy,
thus, limiting its therapeutic potential (12). Conversely, TRAIL was shown to
induce the survival and proliferation in a subset of TRAIL
resistant ALL cells (13).
Combination therapies offer a solution to overcoming resistance to
TRAIL monotherapy (14,15). Early clinical trials using
monoclonal agonistic antibodies which target DR4 with established
chemotherapeutics such as paclitaxel and carboplatin were well
tolerated and demonstrated clinical benefit (16). Further studies are required to
identify agents that sensitise tumour cells to TRAIL together with
extensive biochemical studies to identify signalling pathways
involved in synergistic combinations.
Our group recently demonstrated that
pyrrolo-1,5-benzoxazepine-15 (PBOX-15) increased DR5 expression in
multiple myeloma cells and sensitised these cells to TRAIL-induced
cell death (17). The PBOXs are a
novel group of tubulin targeting agents (18) which potently induce apoptosis in a
wide spectrum of cancer cells including those originating from the
four main types of leukaemia (19–22)
and those exhibiting multi-drug resistance (23). The anti-leukaemic potential of the
PBOXs extended to ex vivo CML and CLL patient samples
including those derived from poor prognostic subgroups and those
resistant to current first line therapies (20,24).
Furthermore, PBOX-6, a potent representative member of the PBOXs,
significantly reduced the growth of CML cells in vivo whilst
exhibiting no adverse effects (24). Moreover, the PBOXs are selective
anticancer agents and display no toxicity towards normal peripheral
blood cells or bone marrow cells at concentrations that are toxic
to leukaemia cells (20,21). Hence, the PBOXs represent an ideal
chemotherapeutic to combine with TRAIL for the treatment of ALL.
Herein, we present novel findings demonstrating the potential of
the PBOXs as single agents and in combination with TRAIL for the
treatment of ALL. Several key signalling pathways mediating
synergistic combinations are identified.
Materials and methods
Unless otherwise stated, chemicals were obtained
from Sigma-Aldrich (Poole, UK) and tissue culture vessels were
sourced from Greiner Bio-One GmbH (Frickenhausen, Germany).
Cell culture
Acute lymphoblastic leukaemia cell lines, Jurkat (T
cell), Nalm-6 and Reh (B cell precursor) were purchased from DSMZ
(Braunschweig, Germany) and CEM (T cell) were originally obtained
from the American Type Culture Collection (ATCC; Manassas, VA,
USA). All cells were maintained in Roswell Park Memorial Institute
(RPMI)-1640 medium enhanced with GlutaMAX-I and supplemented with
10% fetal bovine serum (FBS), 50 units/ml penicillin and 50 μg/ml
streptomycin (all from Gibco-Invitrogen, Carlsbad, CA, USA). Cells
were maintained at densities between 0.5–1.5×106
cells/ml (Jurkat), 0.2–2×106 cells/ml (CEM) or
0.5–4×106 cells/ml (Nalm-6 and Reh) in a humidified
incubator at 37°C in 5% CO2.
Reagents
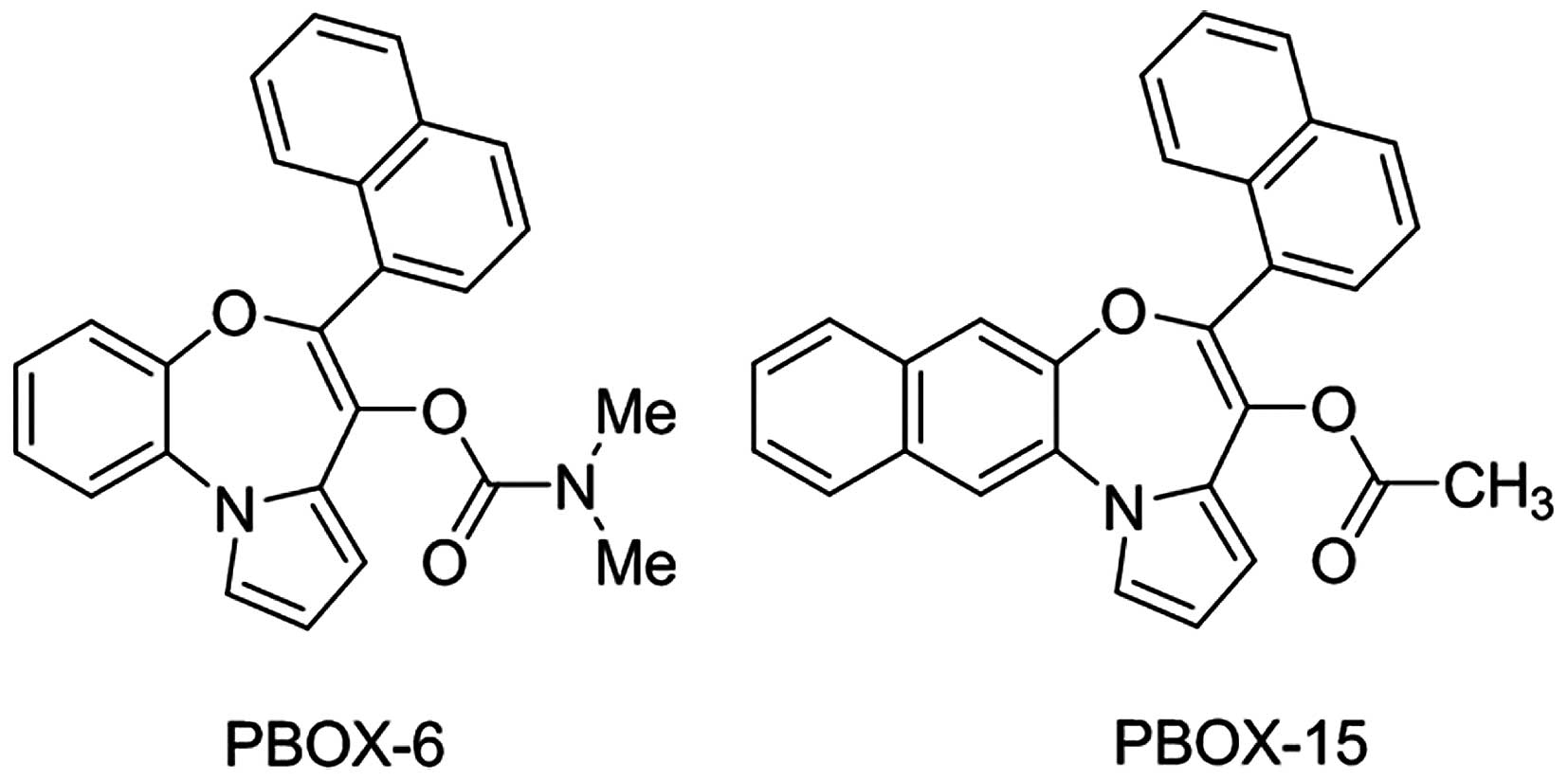
The pyrrolo-1,5-benzoxazepine compounds,
7-[(N,N-dimethylcarbamoyl)oxy]-6-(naphth-1-yl)pyrrolo[2,1-d][1,5]benzoxazepine
(PBOX-6) and 4-acetoxy-5-(1-naphthyl)
naphtho[2,3-b]pyrrolo[2,1-d][1,4]oxazepine (PBOX-15)
were synthesised following the procedures described in the study by
McGee et al (25). The
compounds were dissolved in ethanol and stored at −20°C. Their
chemical structure is shown in Fig.
1. Recombinant human TRAIL (amino acids 114–281) was purchased
from Merck Millipore (Nottingham, UK) in a buffer containing 500 mM
NaCl, 10 mM Na2HPO4, 2.7 mM KCl, 2 mM
KH2PO4, 1 mM DTT, ≤10% glycerol. The TRAIL
was aliquoted as supplied (1.2 mg/ml) and stored at −70°C. A
DR5-selective TRAIL variant, D269H/E195R, was generated as
previously described (26,27). D269H/E195R was diluted to a
concentration of 0.5 mg/ml in a buffer containing 200 mM NaPi (pH
7.4), 150 mM NaCl, 10% glycerol, 1 M DTT and 20 mM
ZnSO4. Aliquots were then stored at −70°C. Monoclonal
antibodies capable of neutralising DR5 were purchased from Alexis
(Enzo Life Sciences, Exeter, UK). Caspase inhibitors, z-IETD-fmk
(caspase-8), z-LEHD-fmk (caspase-9) and z-VAD-fmk (general caspase
inhibitor), all purchased from Merck Biosciences Ltd. (Nottingham,
UK), were dissolved in DMSO and aliquoted prior to storage at
−20°C. The phosphoinositide 3-kinase (PI3K) inhibitor, LY294002,
was also dissolved in DMSO and stored at −20°C.
Cell proliferation
Cell proliferation was monitored using AlamarBlue™
dye (BioSource, Invitrogen, Carlsbad, CA, USA) which changes to a
fluorescent state in the reduced environment of living cells. ALL
cells were seeded onto 96-well plates and then treated with a range
of concentrations of PBOX-6 or PBOX-15 for 72 h. AlamarBlue™ [final
concentration 10% (v/v)] was added and incubated at 37°C.
Fluorescence was measured at an excitation wavelength of 544 nm and
an emission wavelength of 590 nm using a SpectraMax Gemini
spectrofluorometric plate reader (Molecular Devices, Sunnyvale, CA,
USA). The results were expressed as the percentage cell viability
relative to vehicle-treated control cells (100%). Dose-response
curves were plotted and IC50 values (concentration of
drug resulting in 50% reduction in cell viability) were obtained
using Prism GraphPad 4.
Determination of DNA content
Following treatment, cells were harvested by
centrifugation at 800 × g for 10 min. Cell pellets were resuspended
in 200 ml PBS and fixed by a drop-wise addition of 2 ml of ice-cold
70% (v/v) ethanol/PBS while gently vortexing. Following overnight
fixation at −20°C the cells were again centrifuged to remove the
ethanol and resuspended in PBS supplemented with 0.5 mg/ml RNase A
and 0.15 mg/ml propidium iodide (PI). Cells were incubated in the
dark at 37°C for 30 min. Fluorescence from PI was measured on a
linear scale using a FACSCalibur flow cytometer (Becton-Dickinson,
San Jose, CA, USA). Data collections (10,000 events per sample)
were gated to exclude cell debris and cell aggregates. Fluorescence
(from PI) was proportional to the amount of DNA present in each
entity and therefore indicated the stage of the cell cycle. Cells
in G0/G1 were diploid (2N DNA content), cells
in the S phase had DNA contents between 2N and 4N, cells in
G2/M were tetraploid (4N DNA content), while apoptotic
cells were hypoploid and contained <2N DNA. All data was
recorded and analysed using the CellQuest software
(Becton-Dickinson).
Analysis of protein expression,
phosphorylation and cleavage by western blotting
Cells were harvested in a RIPA buffer consisting of
50 mM Tris-HCl (pH 8.0), 150 mM sodium chloride, 1.0% (v/v) Igepal
CA-630 (NP-40), 0.5% (w/v) sodium deoxycholate, and 0.1% (w/v)
sodium dodecyl sulphate (SDS) supplemented with protease and
phosphatase inhibitor cocktails from Sigma-Aldrich. The samples
were then diluted in 3× sample buffer [187.5 mM Tris-HCl (pH 6.8),
6% (w/v) SDS, 30% glycerol, 150 mM DTT, 0.03% w/v bromophenol
blue]. For the detection of PARP, cells were harvested in whole
cell lysis buffer containing 62.5 mM Tris (pH 6.8), 2% (w/v) SDS,
10% (v/v) glycerol, 0.00125% (w/v) bromophenol blue and 50 mM DTT
and the samples sonicated briefly. All extracts were denatured at
65°C for 10 min before separation of proteins on a polyacrylamide
gel and transfer to PVDF membrane. The PVDF transfers were probed
overnight at 4°C with primary antibodies and then incubated with
horseradish peroxidase-conjugated anti-mouse or anti rabbit
secondary antibodies (Promega, Madison, WI, USA). Protein
expression was visualised by enhanced chemiluminescence. All
primary antibodies were supplied by Cell Signaling Technology Inc.
(Beverly, MA, USA) except for those generated against PARP, Bcl-2,
pro-caspase-3, c-FLIP, β-tubulin and GAPDH which were obtained from
Merck Biosciences and Bcl-x, and BubR1 from BD Biosciences
(Bedford, MA, USA).
Measurement of mitochondrial membrane
potential
Mitochondrial transmembrane potential was measured
as a function of drug treatment using the potentiometric dye
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine
iodide (JC-1) (Molecular Probes, Invitrogen, Germany). Following
treatment, the cells were washed with PBS and stained with JC-1 (5
μM) in PBS for 30 min at 37°C. Cells were washed with PBS,
resuspended in ice-cooled PBS and immediately assessed for red and
green fluorescence by flow cytometry (FACSCalibur flow cytometer;
Becton-Dickinson). Excitation at 488 nm and emission filters of 535
and 595 nm were used to quantify the population of mitochondria
with green (JC-1 monomers) and red (JC-1 aggregates) fluorescence,
respectively. The red-to-green fluorescence ratio for individual
cells was calculated using the CellQuest software
(Becton-Dickinson).
Analysis of TRAIL receptor
expression
Cells were washed twice in 1% (w/v) BSA/PBS then
incubated for 45 min on ice with monoclonal antibodies to DR4, DR5,
DcR1, DcR2 or isotype control antibodies (Enzo Life Sciences) in
BSA/PBS. The cells were again washed twice in BSA/PBS then
incubated in darkness for a further 45 min in anti-mouse IgG-FITC
(DakoCytomation, Glostrup, Denmark) in BSA/PBS. Cells were washed,
resuspended in PBS and TRAIL receptor expression detected by flow
cytometry (FACSCalibur; Becton-Dickinson). Results were then
analysed using the FlowJo software (Tree Star Inc., San Carlos, CA,
USA).
Statistical analysis
Results were presented as mean ± SEM. The
statistical analysis of experimental data was performed using the
computer program Prism GraphPad 4. P-values were determined using
either a two-tailed Student's paired t-test or one-way ANOVA
(Bonferroni's test). A value of P<0.05 was considered to be
significant.
Analysis of drug interactions
Drug interactions were determined by median dose
effect analysis using the Calcusyn software (Biosoft, Cambridge,
UK). This method is based on the drug effect equation of Chou and
Talalay and can determine the degree of synergism or antagonism
between two compounds by generating a combination index (CI) value.
CI values of <1, =1 and >1 indicate synergism, an additive
effect and antagonism, respectively.
Results
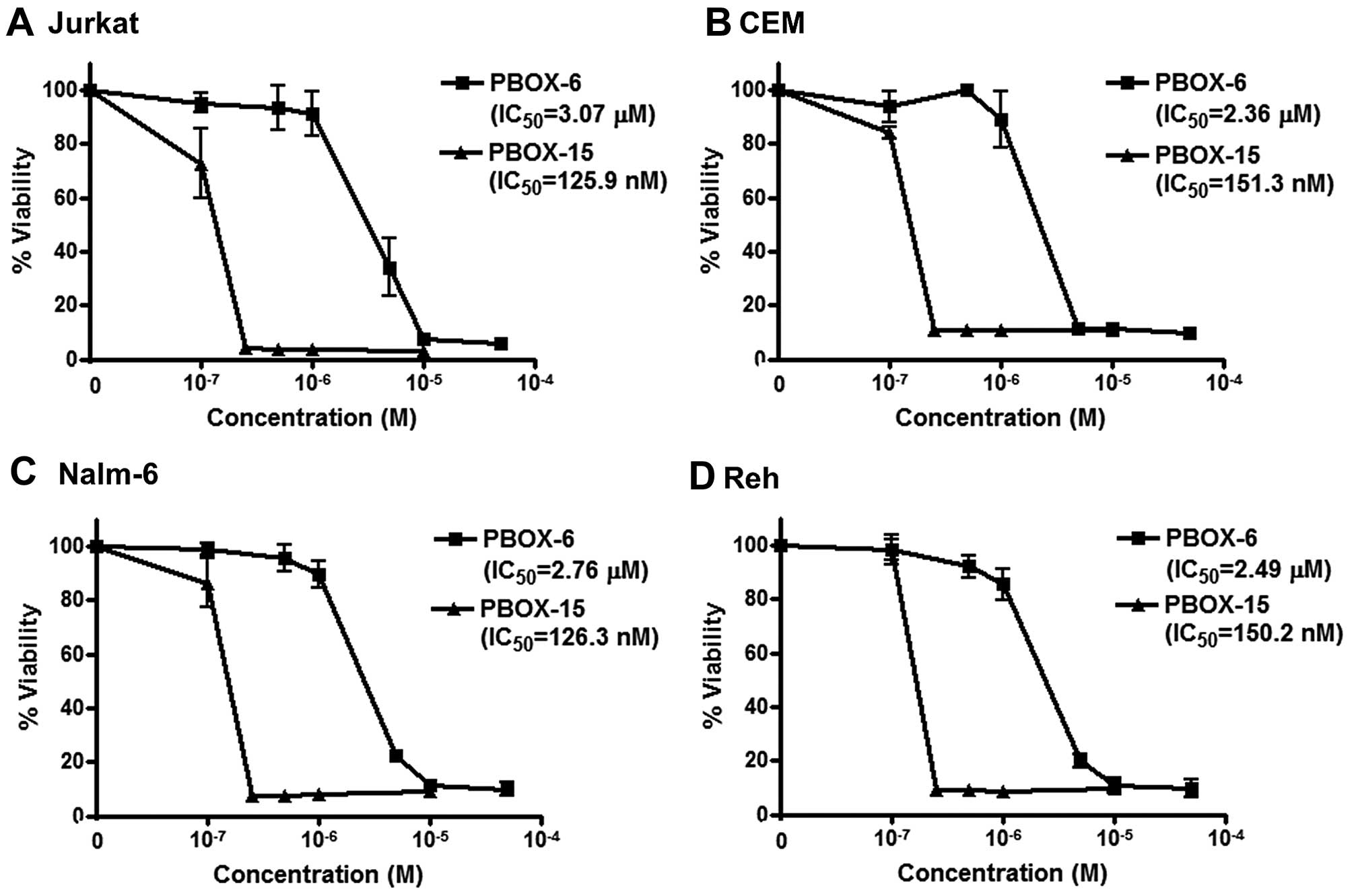
Effect of PBOX compounds on viability of
ALL cells
Jurkat, CEM, Nalm-6 and Reh cells were treated for
72 h with a range of concentrations of PBOX-6 and PBOX-15. Cell
viabilities were measured by AlamarBlue assay (Fig. 2). PBOX-6 and PBOX-15 reduced
viability of both T cell ALL cells (Jurkat and CEM) and B cell
precursor ALL cells (Nalm-6 and Reh) in a dose-dependent manner.
IC50 values (concentration of drug resulting in 50%
reduction in cell viability) for PBOX-6 were 3.1, 2.4, 2.8 and 2.5
μM in Jurkat, CEM, Nalm-6 and Reh cells, respectively. Similarly,
IC50 values for PBOX-15 were 0.13, 0.15, 0.13 and 0.15
μM in Jurkat, CEM, Nalm-6 and Reh cells, respectively. As PBOX-15
was found to be the more potent compound, it was chosen for further
evaluation in combination with TRAIL.
Effect of PBOX-15 alone and in
combination with TRAIL on cell cycle arrest and apoptosis in ALL
cells
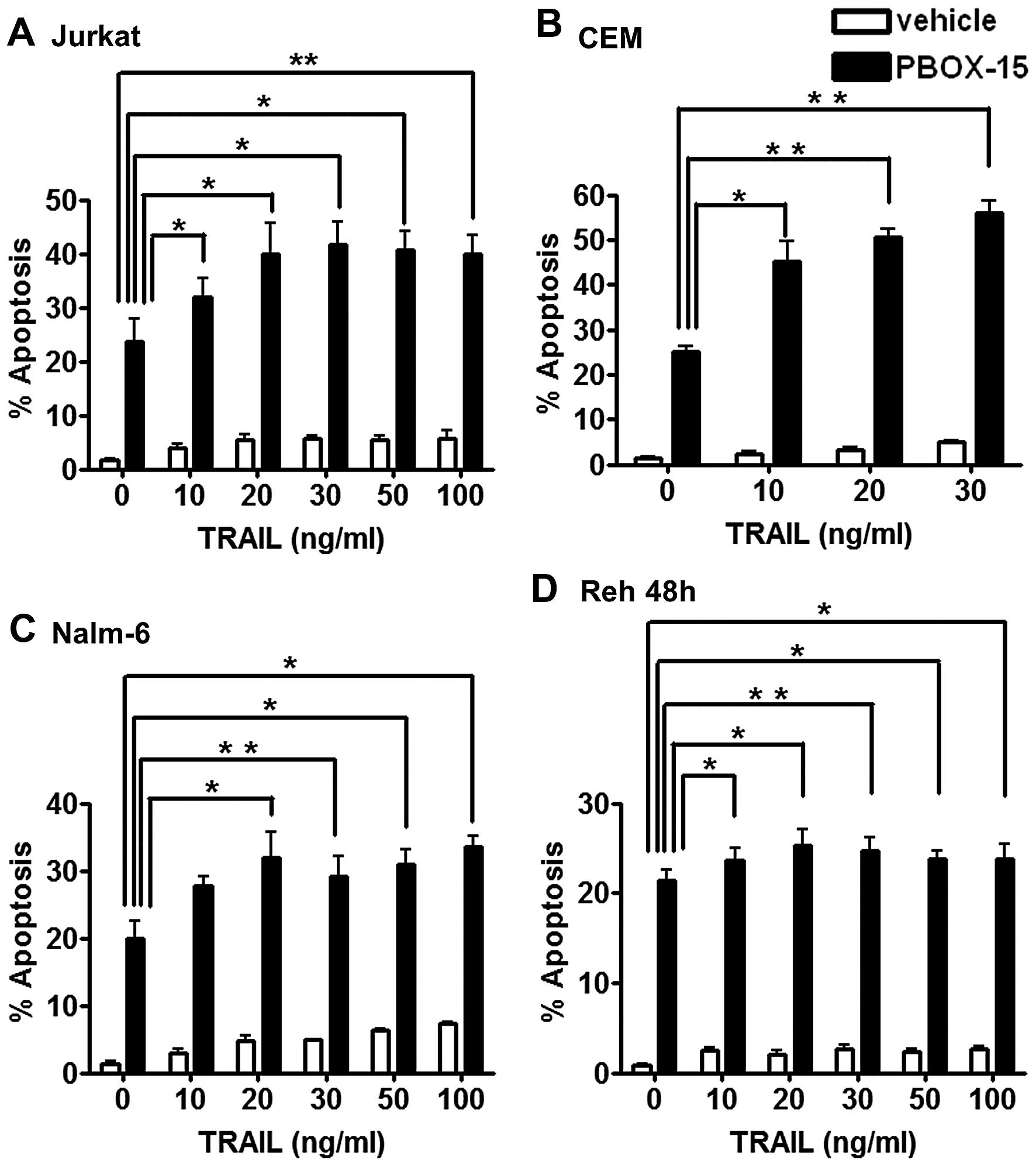
To investigate if PBOX-15 enhanced the apoptotic
efficacy of TRAIL in ALL cells, we treated Jurkat, CEM, Nalm-6 and
Reh cells with either vehicle (0.2% EtOH) or PBOX-15 (1 μM) alone
or in combination with either TRAIL (10–100 ng/ml) or TRAIL buffer
(control, 0 ng/ml) for 24 h (or 48 h for Reh cells). Apoptosis was
quantified by flow cytometric analysis as the number of cells with
hypoploid DNA content (<2N DNA) (Fig. 3). In all four cell lines, treatment
with TRAIL alone, even at the highest concentrations tested,
resulted in low levels of apoptosis <7% compared to ~2% with
buffer and vehicle alone. PBOX-15 alone resulted in between 20–25%
apoptosis depending on the cell type. The combination of TRAIL and
PBOX-15 resulted in statistically significant increases in
apoptosis compared to PBOX alone or TRAIL alone in all four cell
lines.
To determine if the enhancement in apoptosis we
observed between TRAIL and PBOX-15 could be considered synergistic,
we used the CalcuSyn program to perform a median dose analysis of
our flow cytometry results (Table
I). We found that PBOX-15 in combination with the indicated
concentrations of TRAIL (10–100 ng/ml) was considered synergistic
in Jurkat, CEM and Nalm-6 cells as they displayed combination index
(CI) values of <1. The combination was found to be additive in
Reh cells as the CI value was approximately equal to 1 in these
cells (Table I).
 | Table ITRAIL synergistically enhances
PBOX-15-induced apoptosis in ALL cells. |
Table I
TRAIL synergistically enhances
PBOX-15-induced apoptosis in ALL cells.
| TRAIL (ng/ml) | PBOX-15 (μM) | Fraction
affected | CI | Symbol | Description |
|---|
| Jurkat | 10 | 1 | 0.3197 | 0.629 | +++ | Synergism |
| 20 | 1 | 0.4001 | 0.377 | +++ | Synergism |
| 30 | 1 | 0.4163 | 0.341 | +++ | Synergism |
| 50 | 1 | 0.4063 | 0.363 | +++ | Synergism |
| 100 | 1 | 0.3985 | 0.38 | +++ | Synergism |
| CEM | 10 | 1 | 0.4501 | 0.483 | +++ | Synergism |
| 20 | 1 | 0.5043 | 0.448 | +++ | Synergism |
| 30 | 1 | 0.5604 | 0.413 | +++ | Synergism |
| Nalm-6 | 10 | 1 | 0.2717 | 0.743 | ++ | Moderate
synergism |
| 20 | 1 | 0.3185 | 0.502 | +++ | Synergism |
| 30 | 1 | 0.2917 | 0.63 | +++ | Synergism |
| 50 | 1 | 0.3083 | 0.553 | +++ | Synergism |
| 100 | 1 | 0.335 | 0.455 | +++ | Synergism |
| Reh | 10 | 1 | 0.237 | 1.083 | ± | Nearly
additive |
| 20 | 1 | 0.2537 | 0.986 | ± | Additive |
| 30 | 1 | 0.2468 | 1.024 | ± | Nearly
additive |
| 50 | 1 | 0.2385 | 1.073 | ± | Nearly
additive |
| 100 | 1 | 0.2375 | 1.079 | ± | Nearly
additive |
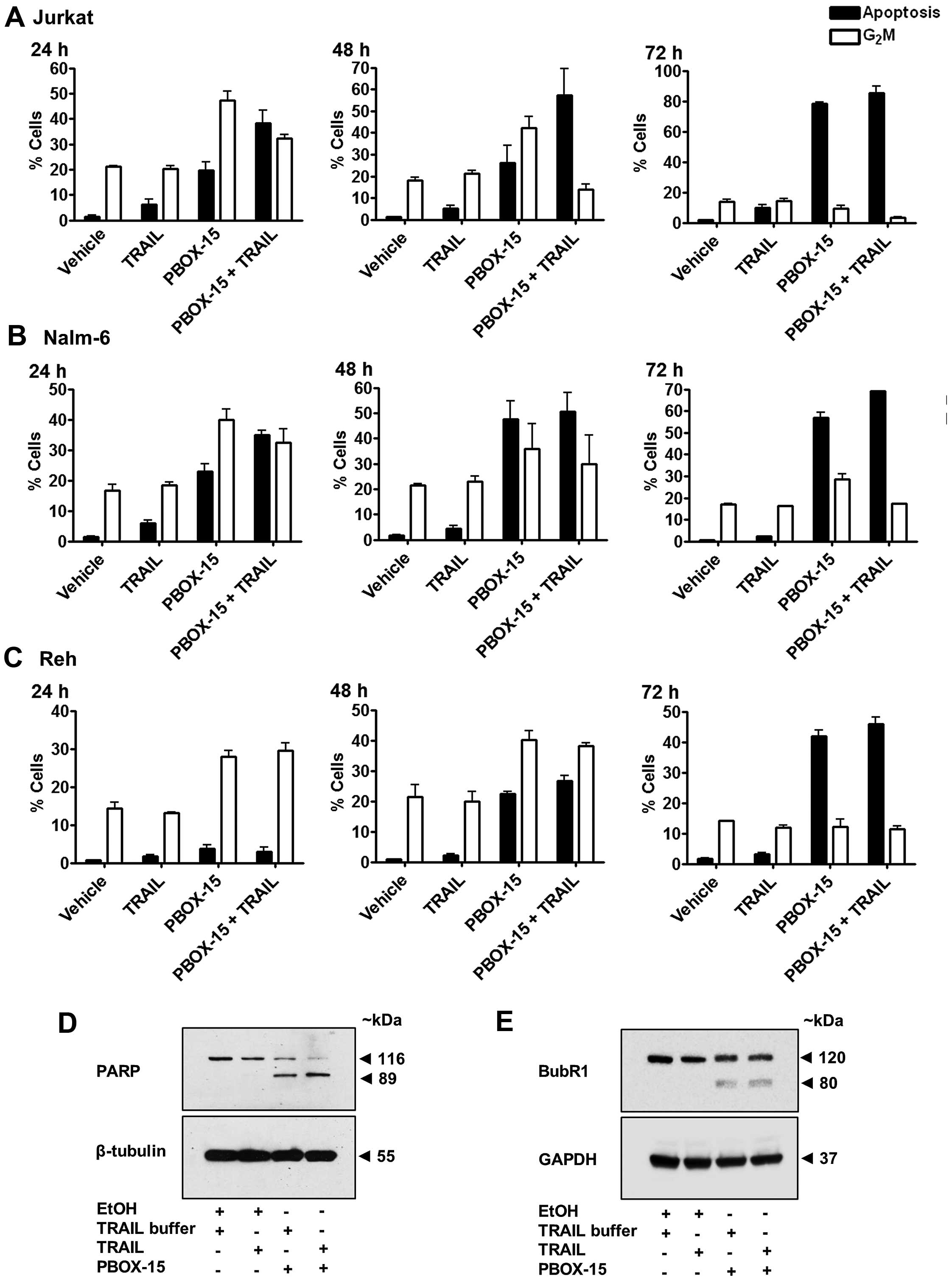
We next examined the relationship between apoptosis
and G2M arrest induced by PBOX-15 alone, TRAIL alone or
the PBOX-15/TRAIL combination in ALL cells between 24–72 h
(Fig. 4A–C). In all three cell
lines, TRAIL induced only low levels of apoptosis even after 72 h
in Jurkat (7.5±0.2%), Nalm-6 (2.4±0.2%) and Reh (3.1±0.6%) compared
to vehicle-treated cells which displayed 1.8±0.2, 0.7±0.1 and
1.7±0.4% apoptosis, respectively. This suggested that ALL cells
exhibit only limited responsiveness to TRAIL alone. As expected,
TRAIL alone did not significantly alter the percentage of cells
undergoing G2/M arrest compared to vehicle-treated
cells. The percentage of cells undergoing PBOX-15-induced apoptosis
increased in a time-dependent manner, 19.8±3.1 to 78.1±0.9% in
Jurkats (Fig. 4A), 22.8±4.6 to
57.0±3.6% in Nalm-6 (Fig. 4B) and
3.8±1.0 to 42.0±2.6% in Reh (Fig.
4C) from 24 to 72 h. Levels of G2/M decreased over
time from 47.4±3.4 to 9.1±2.9% in Jurkats and 39.9±3.5 to 28.5±2.6%
in Nalm-6 from 24 to 72 h. Reh cells initially underwent an
increase in G2M arrest from 27.0±1.6 at 24 h to
40.2±2.9% at 48 h followed by a decrease to 12.3±2.3%
G2M at the later time-point of 72 h, illustrating a
slower time course of PBOX-15-induced response in Reh cells. These
results illustrate that ALL cells undergo G2M arrest
followed subsequently by apoptosis in response to PBOX-15
treatment.
ALL cells treated with the combination followed a
similar pattern of time-dependent increases in apoptosis along with
decreases in G2M as PBOX-15 alone, however, at any
particular time-point, levels of apoptosis were higher and
G2M lower than with PBOX-15 alone. For example, after 24
h, percentage apoptosis was 19.8±3.1% and G2M arrest was
47.4±3.4% in Jurkats treated with PBOX-15 alone but with the
TRAIL/PBOX-15 combination apoptosis increased to 38.1±5.0% and
G2M decreased to 32.2±1.5%. Similarly, in Nalm-6 after
24 h PBOX-15/TRAIL induced 34.9±1.6% apoptosis and 32.5±4.4%
G2M compared to 22.8±4.6% apoptosis and 39.9±3.5%
G2M with PBOX alone. In Reh cells, apoptosis rose from
22.5±1.2 to 26.8±1.6% and G2M decreased from 40.2±2.9 to
38.2±1.0%, after treatment with combination for 48 h compared to
PBOX alone. Therefore, the enhancement of apoptosis induced by the
PBOX-15/TRAIL combination compared to either agent alone appears to
be associated with acceleration in the transition from
G2M arrest to subsequent apoptosis.
To support these findings, we also assessed cleavage
of the DNA repair enzyme PARP and degradation of the mitotic
spindle checkpoint protein BubR1. Cleavage of PARP is one of the
key indicators of apoptosis. Treatment of Jurkat cells for 24 h
with PBOX-15 alone or in combination with TRAIL resulted in
cleavage of PARP, evidenced as a decrease in full-length PARP (116
kDa) and the appearance of a cleaved fragment (89 kDa) (Fig. 4D). In keeping with our flow
cytometric data, the combination resulted in visibly higher amounts
of PARP cleavage compared to the PBOX-15 alone. Although there was
a slight reduction in the expression of full length PARP with TRAIL
alone compared to vehicle treated cells, it was not sufficient to
detect the cleaved fragment. We have previously demonstrated that
the mitotic spindle checkpoint protein BubR1 is phosphorylated
during G2/M arrest followed subsequently by degradation
as arrest progressed to apoptosis following treatment of CML and
prostate cancer cells with PBOX-15 (28,29).
We now demonstrate evidence of degradation of BubR1 indicative of
apoptotic progression in Jurkat ALL cells treated for 24 h with
PBOX-15 or PBOX-15 in combination with TRAIL (Fig. 4E). The appearance of a cleaved
fragment at ~80 kDa suggests cleavage of BubR1 by caspases.
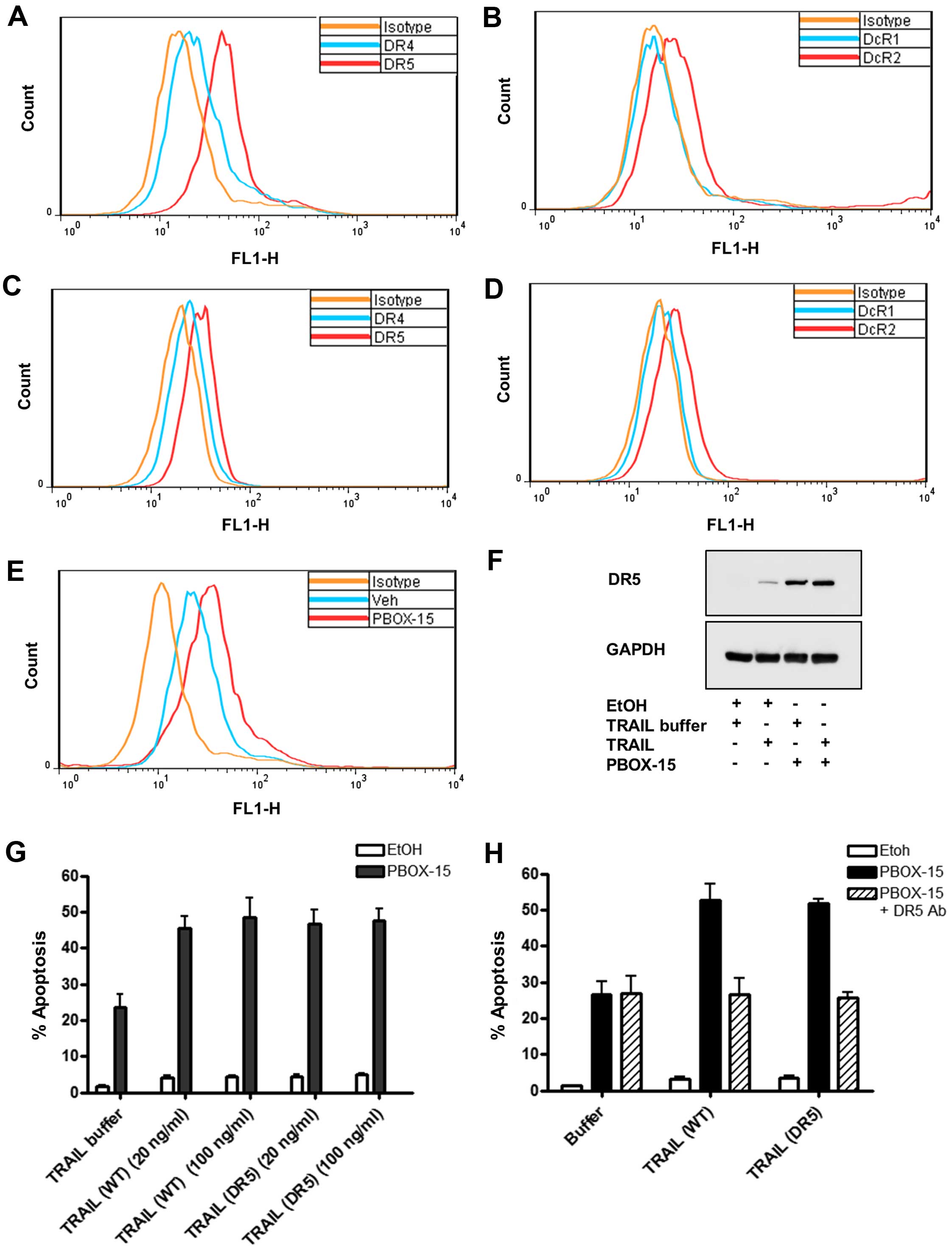
Role of DR5 in PBOX-15/TRAIL-induced
apoptosis in ALL cells
We first sought to confirm death receptor expression
patterns in Jurkat (Fig. 5A and B)
and Nalm-6 (Fig. 5C and D) cells
as representative T cell or B cell precursor ALL lines,
respectively. Cell surface receptor expression was analysed by
immunostaining using flow cytometry. We established that Jurkat
cells predominantly express DR5 (Fig.
5A) identified as a shift to the right of the FL1-H peak
compared to cells stained with the mouse isotype control antibody
(negative control). Cell surface expression of DR4 (Fig. 5A) and DcR2 (Fig. 5B) was also observed in Jurkats but
at lower levels than DR5. Expression of DR4 on Jurkat cells is
somewhat controversial as conflicting reports documenting the DR4
status of Jurkats exist (30).
Expression of DR4 may depend on the strain, the age or the culture
conditions of the Jurkats. Receptor and decoy receptor expression
on Nalm-6 cells (Fig. 5C and D)
followed a similar pattern to Jurkats. Expression of DR5 was not as
pronounced on Nalm-6 as on Jurkat cells.
It has been previously shown that other
microtubule-targeting agents, such as paclitaxel, can increase cell
surface expression of DR5 (31,32).
We next sought to delineate if PBOX-15 enhanced TRAIL-induced
apoptosis by affecting the expression of DR5. We determined by flow
cytometric immunostaining that PBOX-15 increased the cell surface
expression of DR5 on Jurkat cells, evidenced by a shift to the
right in the FL1-H peak compared to EtOH-treated cells (Fig. 5E). This finding was supported by
western blots displaying upregulation in whole cell expression of
DR5 after treatment with PBOX-15 alone or in combination with TRAIL
compared to vehicle controls (Fig.
5F).
To establish the importance of DR5 in
PBOX-15/TRAIL-induced apoptosis, we treated Jurkat cells with PBOX
alone or in combination with TRAIL or a TRAIL variant selective for
DR5 (D269H/E196R). We found that the DR5-selective TRAIL variant
enhanced PBOX-induced apoptosis with a similar efficacy as
wild-type TRAIL (Fig. 5G). To
further delineate the role of DR5 in PBOX-15/TRAIL induced
apoptosis, Jurkat cells were pre-treated with a DR5 neutralising
antibody for 1 h prior to treatment with PBOX-15 and/or TRAIL. The
presence of the DR5 neutralising antibody abrogated the enhancement
of apoptosis usually evident with the PBOX-15 in combination with
either wild-type TRAIL or the DR5-specific TRAIL to the same levels
as PBOX-15 alone (Fig. 5H). This
result indicated the importance of DR5 in PBOX-15 and TRAIL
cooperation.
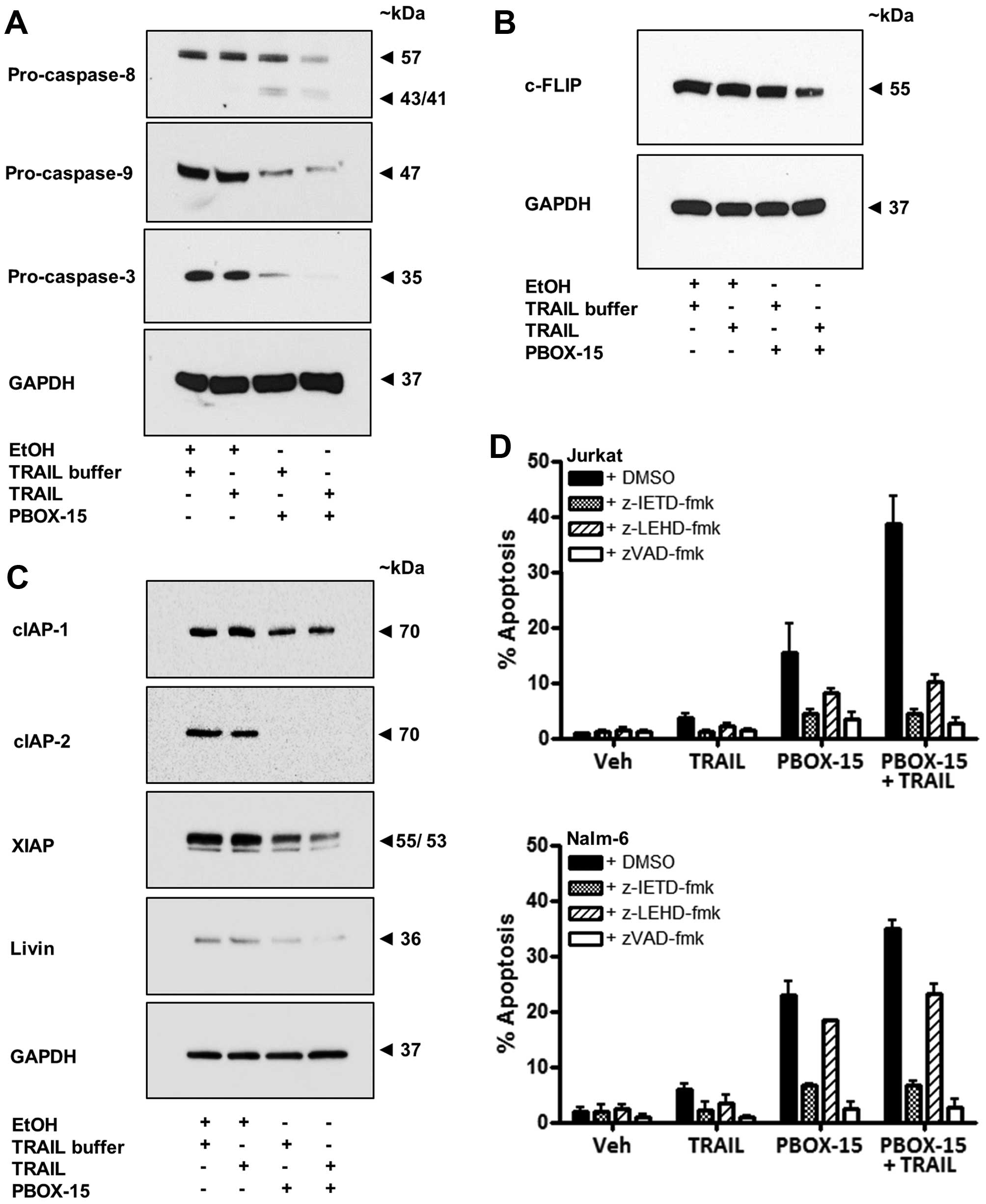
Involvement of caspases in
PBOX/TRAIL-induced apoptosis in ALL cells
To elucidate the apoptotic pathways recruited by
PBOX-15 and TRAIL, alone and in combination, we next investigated
the involvement of upstream initiator caspases, caspase-8 and -9.
We established that treatment of Jurkat cells with the
TRAIL/PBOX-15 combination resulted in processing of both
pro-caspase-8 and pro-caspase-9 indicative of the involvement of
the extrinsic and intrinsic apoptotic pathways, respectively.
Processing was evidenced by western blot analysis as the
disappearance of bands corresponding to the size of the pro-form of
the caspases and in the case of caspase-8, the appearance of
cleaved active fragments (Fig.
6A). Processing of a downstream effector caspase,
pro-caspase-3, was also evident in Jurkat cells treated with
PBOX-15 alone or in combination with TRAIL (Fig. 6A).
Caspase activity is modulated by the presence of
endogenous caspase inhibitors such as cellular FLICE-inhibitory
protein (c-FLIP) (33) or
inhibitor of apoptosis proteins (IAPs) (34). Overexpression of c-FLIP may
contribute to resistance to TRAIL through inhibition of the
signalling cascade initiated by DR5 (12). Treatment of Jurkat cells with
PBOX-15 in combination with TRAIL resulted in downregulation of
c-FLIP protein expression (Fig.
6B). A high level of IAP expression in cancer cells also
represents an important anti-apoptotic mechanism and is associated
with chemoresistance. PBOX-15 alone was found to reduce the
expression of cIAP-2 and XIAP and to a lesser extent cIAP-1 and
livin compared to vehicle treated cells (Fig. 6C). A combination of PBOX-15 and
TRAIL led to a greater reduction in expression of XIAP compared to
PBOX-15 alone. It should be noted that endogenous levels of livin
in Jurkat cells tends to be low. These results are of interest as
it has been previously shown that XIAP inhibitors cooperate with
TRAIL to induce apoptosis in ALL cells (14).
To further investigate the role of initiator
caspases in PBOX/TRAIL induced apoptosis we pre-treated Jurkat and
Nalm-6 cells with irreversible cell permeable inhibitors of
caspase-8 (z-IETD-fmk), caspase-9 (z-LEHD-fmk) and a general
broad-range caspase inhibitor (z-VAD-fmk). The broad-range
inhibitor abrogated apoptosis induced by PBOX-15, TRAIL or the
combination in both Jurkat and Nalm-6 cells (Fig. 6D). For example, the PBOX/TRAIL
combination resulted in 38.7±4.9% apoptosis in Jurkats and
34.7±1.6% in Nalm-6 cells. Pre-treatment with z-VAD-fmk reduced
TRAIL/PBOX-15-induced apoptosis to 2.8±0.7% (Jurkat) and 2.7±1.4%
(Nalm-6), levels which are close to background apoptosis evident in
control cells (Jurkat, 0.8±0.1% and Nalm-6, 1.9±0.7%). These
findings confirmed that PBOX/TRAIL-induced apoptosis occurred in a
caspase-dependent manner. The caspase-8 inhibitor reduced
PBOX/TRAIL-induced apoptosis to only 4.5±0.7% in Jurkat or 6.7±0.7%
in Nalm-6 cells, illustrating the importance of caspase-8 and the
extrinsic pathway in the apoptotic response induced by PBOX-15 and
TRAIL in ALL cells. In Jurkat cells, although not quite as
effectively as the caspase-8 inhibitor, the caspase-9 inhibitor
also resulted in a significant reduction in TRAIL/PBOX-15-induced
apoptosis (10.2±1.3%), highlighting the importance of caspase-9 and
the intrinsic apoptotic pathway in Jurkat (type II) cells.
Conversely, in Nalm-6 cells, the caspase-9 inhibitor only reduced
TRAIL/PBOX-15-induced apoptosis to 23.1±1.9%, indicating that
signalling through the intrinsic pathway may not be as important in
B cell precursor Nalm-6 cells as in Jurkat T cells.
Effect of PBOX-15 alone or in combination
with TRAIL on mitochondrial membrane potential (MMP) and the Bcl-2
family in Jurkat ALL cells
We next determined whether exposure of Jurkats to
PBOX-15 alone or in combination with TRAIL can alter MMP using the
potentiometric dye JC-1. A decrease in the red:green fluorescent
ratio of JC-1 stained cells as detected by flow cytometry is
indicative of a loss in MMP and hence, mitochondrial
depolarisation. Both PBOX-15 alone and in combination with TRAIL
significantly increased the number of Jurkat cells with reduced MMP
following treatment for 16, 20 or 24 h (Fig. 7A). For example, after 20-h
treatment with PBOX-15, the number of cells exhibiting
mitochondrial depolarisation was 21.0±3.3% compared to only
3.2±1.0% in the vehicle-treated cells (P<0.001). Furthermore,
exposure to the combination resulted in a statistically significant
increase in the number of cells displaying reduced MMP compared to
either agent alone, such as after 20 h the combination resulted in
28.8±4.1% cells with membrane depolarisation compared to 21.0±3.3%
with PBOX-15 alone (P<0.05). Collectively, these findings
support a role for the involvement of the mitochondrial pathway
during PBOX-TRAIL-induced apoptosis.
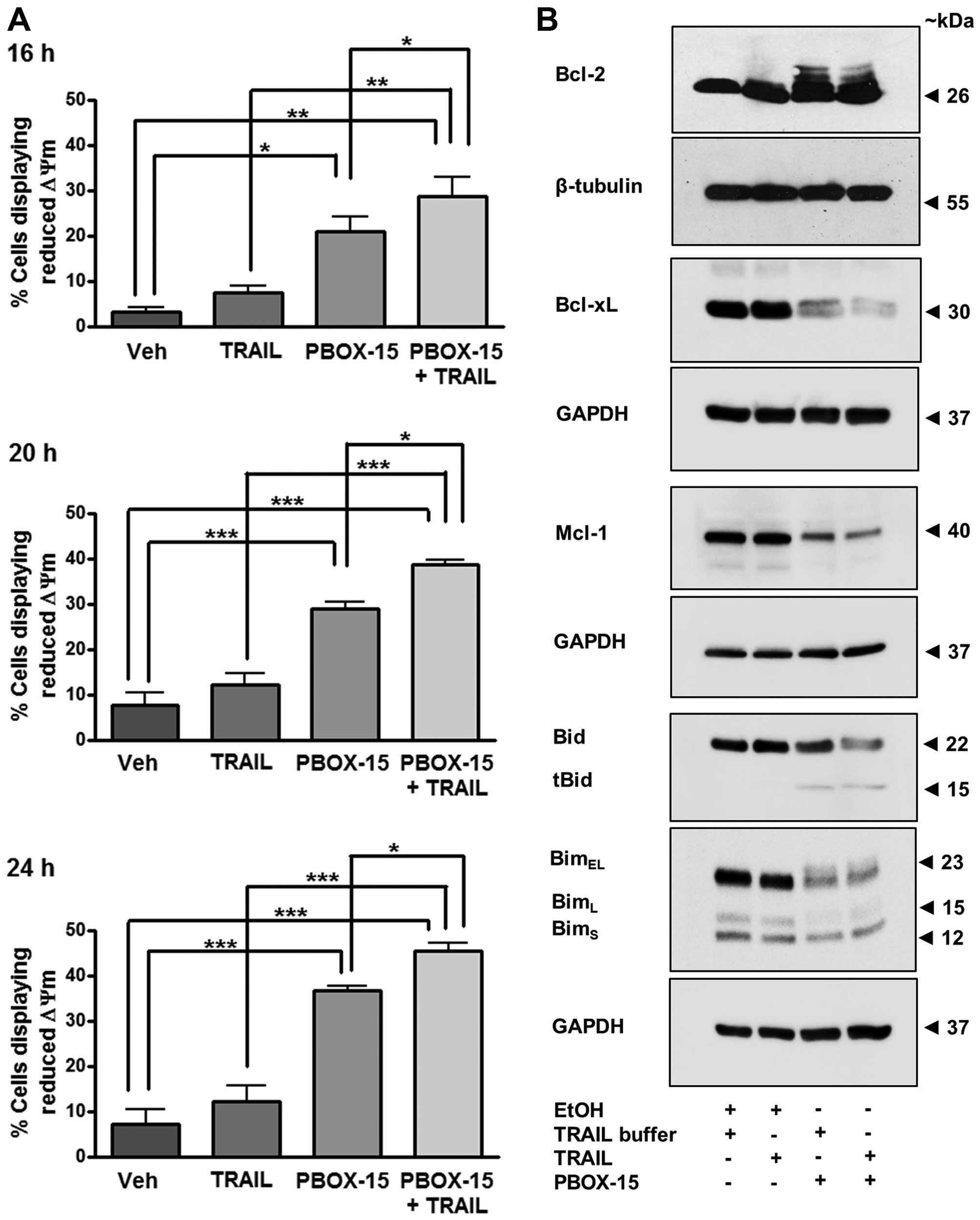 | Figure 7PBOX-15/TRAIL-induced reduction in
mitochondrial membrane potential (ΔΨm) is accompanied by
phosphorylation and/or degradation of antiapoptotic proteins,
Bcl-2, Bcl-xL and Mcl-1, and processing of pro-apoptotic BH3-only
proteins, Bid and Bim, in Jurkat ALL cells. Jurkat cells (500,000
cells/ml) were treated with vehicle [0.2% (v/v) ethanol] or PBOX-15
(1 μM) ± TRAIL (20 ng/ml). (A) Mitochondrial membrane potential was
assessed 16, 20 or 24 h post-treatment by incubation with JC-1 and
analysis by flow cytometry. Values represent the mean ± SEM for
three independent experiments. A P-value of <0.05 was considered
to be statically significant (*P<0.05,
**P<0.01, ***P<0.001). (B) Twenty-four
hours post-treatment, western blotting was used to assess protein
expression of anti-apoptotic proteins, Bcl-2, Bcl-xL and Mcl-1 or
pro-apoptotic BH3-only proteins, Bid and Bim. Expression of
β-tubulin or GAPDH was used as a loading control. Blots are
representative of three independent experiments. |
The Bcl-2 family of proteins are involved in the
control of mitochondrial membrane potential and subsequent
downstream signalling. Overexpression of anti-apoptotic Bcl-2
proteins can confer resistance of cells to inducers of apoptosis
such as TRAIL. Therefore, the ability of a compound to overcome
such resistance can be beneficial therapeutically. We assessed the
effect of PBOX-15 and TRAIL on the antiapoptotic Bcl-2 family
members, Bcl-2, Bcl-xL and Mcl-1 (Fig.
7B). Treatment of Jurkats for 24 h with PBOX-15 alone or in
combination with TRAIL resulted in hyper-phosphorylation of Bcl-2
evidenced by slower migrating bands on a western blot analysis.
Phosphorylation and degradation of Bcl-xL was also evident
following either treatment, as was degradation of Mcl-1. These
results suggest that PBOX-15 treatment may inactivate
anti-apoptotic Bcl-2 proteins in ALL cells.
Pro-apoptotic Bid is activated by its proteolytic
cleavage by caspase-8. Cleaved or truncated Bid (tBid) translocates
to the mitochondria where it can induce cytochrome c
release. Western blotting indicated that treatment of Jurkat cells
with PBOX-15 induced cleavage of inactive precursor Bid (22 kDa)
and the appearance of a band corresponding to the cleaved tBid
fragment (15 kDa) (Fig. 7B). This
result further demonstrates the involvement of crosstalk between
extrinsic pathways and the intrinsic mitochondrial pathway in
PBOX-15/TRAIL-induced ALL cell apoptosis.
Three alternatively spliced isoforms of BH3-only
protein Bim exist and are known as BimEL, BimL and BimS. Although
Bim is a pro-apoptotic protein, decreased expression of BimEL has
been postulated to result in a cleaved form of Bim that displays
increased pro-apoptotic activity in Jurkats cells (35). Herein, we demonstrate that PBOX-15
alone or in combination with TRAIL resulted in decreased expression
of BimEL and BimL (Fig. 7B).
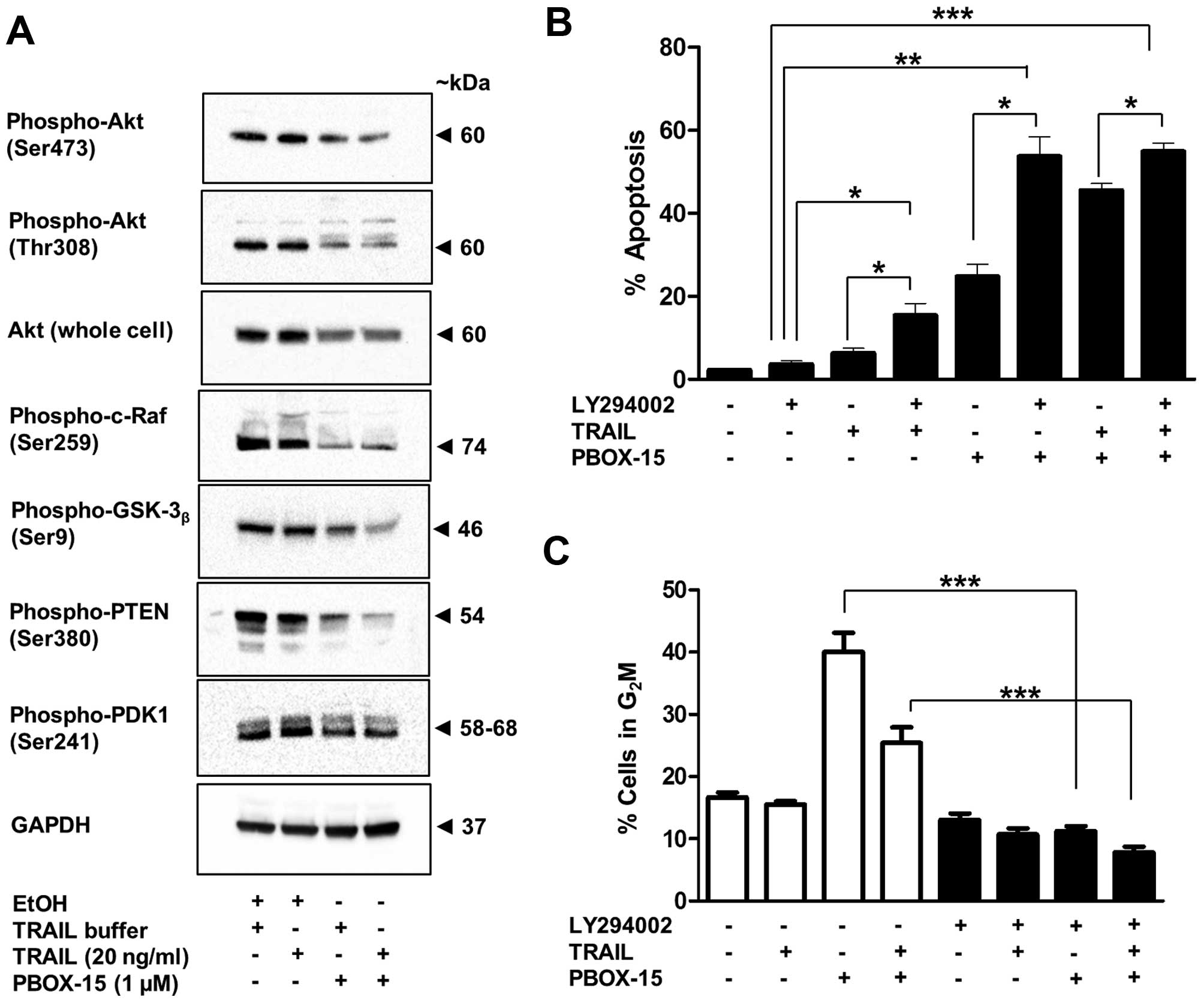
Effect of PBOX-15 alone and in
combination with TRAIL on the PI3K/Akt signalling pathway
The phosphoinositide 3-kinase (PI3K)/Akt
pro-survival signal transduction pathway can confer an aggressive
apoptosis-resistant phenotype to acute leukaemia cells (36). Jurkat ALL cells display high levels
of Akt activation. Treatment of Jurkat cells with PBOX-15 alone and
in combination with TRAIL lead to decreased expression of both
phosphorylated (Ser473 and Thr308) and total cellular Akt (Fig. 8A). Expression of downstream targets
of phospho-Akt, namely, phospho-c-Raf (Ser259) and
phospho-GSK-3β (Ser9) and were also found to be
decreased after treatment (Fig.
8A). Levels of phospho-PDK1 (ser241) were also slightly
decreased (Fig. 8A). This molecule
functions up-stream of Akt and is responsible for its
phosphorylation at Thr308. Notably, expression of the inactive form
of PTEN, Phospho-PTEN (Ser380), was also decreased (Fig. 8A). Dephosphorylated PTEN is a
negative regulator of the Akt/PI3K pathway via dephosphorylation of
PIP3 to PIP2. PIP3 is required for
activation of PDK1 and thus Akt phosphorylation (37).
To understand the role of Akt inactivation in
PBOX-15 and TRAIL-induced apoptosis, Jurkat cells were pretreated
with the PI3K inhibitor LY294002 for 1 h, followed by treatment
with PBOX-15, TRAIL or both for 24 h. Apoptosis was monitored by
flow cytometric analysis of PI stained cells. As shown in Fig. 8B a non-toxic concentration of
LY294002 (10 μM) with a marginally toxic concentration of TRAIL (20
ng/ml: 8%) or modestly active concentration of PBOX-15 (1 μM: ~25%)
at least doubled the amount of apoptosis. Inhibition of the
PI3K/Akt pathway also significantly increased the amount of
apoptosis when both PBOX-15 and TRAIL were combined. This
enhancement of apoptosis was associated with an inhibition of
PBOX-15-induced G2M arrest (Fig. 8C). Taken together with the Akt
western blot experiments, these findings suggest that inactivation
of Akt plays a central role in both PBOX-15 and TRAIL-induced
apoptosis in ALL cells.
Discussion
TRAIL is an established tumour targeting agent able
to selectively eradicate cancer cells by activating a signalling
pathway adapted by the innate immune system (38). However, the therapeutic potential
of TRAIL is limited by resistance mechanisms. Furthermore, recent
studies demonstrated TRAIL-dependent tumour promoting activities
(13). Combination therapies can
frequently amplify the actions of anticancer agents and widen the
therapeutic window. Extensive research into the survival pathways
mediating cellular resistance to TRAIL has identified several
targets for strategic combinational therapies. In this study, we
investigated the therapeutic potential in combining TRAIL with
another tumour selective agent, PBOX-15 in overcoming common
mediators of TRAIL resistance.
The preliminary determinant of TRAIL-sensitivity is
the expression levels of TRAIL receptors on tumor cells. Low
expression of agonist DR4 and DR5 receptors has been shown to limit
TRAIL-induced apoptosis in many cancers including ALL (39–42).
Mutations within the ligand binding or death domain of agonist
TRAIL receptors can also impede the apoptotic potential of TRAIL
(41,43). Furthermore, nuclear localization of
DR5 mediated by importin-β1 can significantly reduce TRAIL-mediated
cell death in cancer cells (44).
It is postulated that high expression of decoy antagonistic
receptors on tumour cell surfaces may compete with agonistic
receptors and reduce the overall apoptotic potential of TRAIL
(45). TRAIL resistance mediated
by agonist receptor expression levels can be overcome by combining
TRAIL with agents that induce the expression of DR4 and DR5 on
tumour cells. The chemotherapeutics etoposide, doxorubicin and
paclitaxel (32,46,47),
the antitumour antibiotic Bleomycin (48), and the histone deacetylase
inhibitor MS-275 (49) have been
shown amongst others to upregulate DR4 and DR5 and thus sensitise
various cancer cells to TRAIL-induced apoptosis. Similarly, our
group demonstrated that novel mictotubulin-targeting agent PBOX-15
synergistically enhanced TRAIL-induced apoptosis in myeloma cells
via upregulation of DR5 (17).
Herein, we report that PBOX-15 synergistically enhanced the
apoptotic potential of TRAIL in T-cell and B-cell precursor
ALL-derived cell lines. PBOX-15 enhanced DR5-selective
variant-induced apoptosis in Jurkat ALL cells with the same
magnitude as wild-type and upregulated the expression of DR5.
Binding of TRAIL to DR4/5 leads to assembly of the death inducing
signaling complex (DISC). Caspase-8 is recruited to DISC where it
is activated and triggers apoptosis via the extrinsic pathway. In
the present study, we demonstrate that caspase-8 is activated
during PBOX-15/TRAIL apoptotic synergy in ALL cells. Furthermore,
the specific caspase-8 inhibitor (z-IETD-fmk) significantly
inhibited apoptosis. These findings support a role for caspase-8 in
PBOX-15-mediated enhancement of TRAIL-induced apoptosis in ALL and
complement previous findings demonstrating caspase-8-dependent
PBOX-15/TRAIL apoptotic synergy in multiple myeloma cells. Taken
together, these findings suggest that PBOX-15 co-operates with
TRAIL by upregulation of DR5 in cancer cells promoting
TRAIL-induced apoptosis via the extrinsic pathway.
Activation of the intrinsic apoptotic pathway was
also evident by activation of caspase-9, MIM depolarisation and Bid
cleavage in response to PBOX-15/TRAIL combinations. Given that the
specific caspase-9 inhibitor (z-LEHD-fmk) only partially inhibited
PBOX-15 alone and PBOX-15/TRAIL-induced apoptosis it may be
inferred that the extrinsic apoptotic pathway is dominant in ALL
cells in response to these agents. There is also the possibility of
cross-talk between the extrinsic and intrinsic pathways initiated
by PBOX-15 alone and when combined with TRAIL. Caspase-8 has been
shown to cleave Bid with subsequent translocation of truncated Bid
to the mitochondria where it interacts with Bax and Bak promoting
cytochrome c release (50).
This cascade of events was recently demonstrated following death
receptor activation by TRAIL linking both intrinsic and extrinsic
apoptotic pathways (51). We
hypothesise that following exposure to PBOX-15/TRAIL the extrinsic
pathway is activated which in turn activates caspase-8 which then
cleaves Bid thus activating the intrinsic pathway and amplifying
the initial apoptotic signal following death receptor
activation.
Several studies have demonstrated some critical cell
survival proteins such as cellular FLICE (FADD-like
IL-1β-converting enzyme)-inhibitory protein (c-FLIP), members of
the Bcl-2 family of anti-apoptotic proteins and inhibitor of
apoptosis proteins (IAPs) in TRAIL resistance. In more detail,
c-FLIP is an anti-apoptotic regulator that inhibits TRAIL-induced
apoptosis in malignant cells (52), and c-FLIP binds to FADD and/or
caspase-8 or -10 and DR5 preventing DISC formation and subsequent
activation of the caspase cascade (53). High expression of c-FLIP correlates
with TRAIL resistance in cancer cells (49,54),
and c-FLIP-mediated TRAIL resistance can be overcome by combining
with c-FLIP targeting chemotherapeutics (55,56)
or c-FLIP siRNA (57). In the
present study, we demonstrate for the first time that PBOX-15 can
downregulate c-FLIP, resulting in increased TRAIL-induced apoptosis
in ALL cells.
The anti-apoptotic protein Mcl-1 is a member of the
Bcl-2 family and plays an important role in the survival of
malignant hematopoietic cells (58). Several pharmacological strategies
targeting the anti-apoptotic proteins Bcl-2 or Bcl-xL are currently
undergoing clinical trials for the treatment of hematological
malignancies (59). We recently
demonstrated that the PBOXs can induce both Bcl-2-dependent
(60) and -independent (20) apoptosis in leukaemia cells. The
PBOXs also phosphorylate and inactive Bcl-xL (29,60).
The effects of the PBOXs on the Bcl-2 family of anti-apoptotic
proteins were amplified when combined with established
chemotherapeutics such as imatinib mesylate (21), carboplatin (61) or the autophagy inhibitor
bafilomycin-a1 (62). In this
study, we demonstrate that PBOX-15 induces the phosphorylation and
inactivation of Bcl-2 and Bcl-xL and also led to a decrease in
Mcl-1 protein levels. Taken together, these results demonstrate the
potential therapeutic efficacy of the PBOX's in targeting the
anti-apoptotic members of the Bcl-2 family and sensitising cancer
cells to various chemotherapeutics and cytokines.
We also report novel findings demonstrating
downregulation of XIAP, cIAP-1 and cIAP-2 in the PBOX-15-induced
apoptotic pathway. Expression of XIAP and cIAP-2 is associated with
resistance to TRAIL in prostate cancer cells (63). Genetic knockdown of IAP expression
restored TRAIL sensitivity in these cells. Similarly, small
molecule IAP antagonists were shown to sensitise various cancer
cells to TRAIL-induced apoptosis (64). Several studies suggest therapeutic
benefit in combining agents targeting IAP expression in combination
with TRAIL (65). Importantly,
small molecule XIAP inhibitors were shown to co-operate with TRAIL
to induce apoptosis in pediatric ALL (14). Accordingly, the levels of XIAP
decreased further when PBOX-15 combined with TRAIL in ALL cells
confirming that agents targeting XIAP can co-operate with TRAIL to
induce apoptosis in ALL. This finding supports other independent
results demonstrating a requirement for cIAP-1 and cIAP-2
downregulation in the signalling pathway mediating TRAIL/Tyrphostin
(a selective Janus kinase 2 inhibitor) apoptotic synergy in ALL
cells (66).
Our data also demonstrate for the first time that
PBOX-15 reduces the levels of whole cell and phosphorylated Akt
suggesting that reduction of Akt may facilitate PBOX-induced
apoptosis. Akt is a serine/threonine kinase and a target of
phosphatidylinositol 3-kinase/PI3K. Akt plays a pivotal role in
proliferation and survival and is frequently altered in human
cancers (67). PBOX-15/TRAIL
combination also showed decreased Akt expression and consequently a
decrease in the phosphorylation status of downstream targets c-Raf
and GSK-3β. Adult ALL cells display constitutive
hyperactivation of the PI3K/Akt pathway highlighting this system as
a novel therapeutic target for the treatment of ALL (68). Subsequent studies with the PI3K
inhibitor LY294002 demonstrated increased sensitivity to TRAIL and
PBOX-15 alone and in combination suggesting inhibition of the
PI3K/Akt signalling sensitises ALL cells to apoptotic effects of
TRAIL and PBOX-15. These findings support recent results
demonstrating therapeutic benefits by inhibition of the
PI3K/Akt/mTOR pathway in patients with ALL (69).
In conclusion, we demonstrate that PBOX-15-mediated
sensitisation of ALL cells to TRAIL-induced apoptosis can be
attributed to activation of the extrinsic apoptotic pathway, cross
activation of the intrinsic apoptotic pathway and targeting several
key cell survival pathways. As similar events have been reported
with other MTAs it seems likely that microtubular destabilisation
by PBOX-15 plays an important role in the mechanism underlying
these effects. Further characterisation of the mechanism of
PBOX-15-induced upregulation of DR5 and downregulation of survival
pathways would allow the strategic development of improved
treatment regimens for PBOX-15/TRAIL combinations. We provide
preliminary data to support the combination of TRAIL/PBOX-15 with
the PI3K inhibitor LY294002 to further enhance apoptotic outcomes
in pediatric ALL. Results presented herein will contribute to
identifying optimum use of this cytokine in the clinic.
Acknowledgements
The funding for the present study was provided by
The Children's Leukaemia Research Project, Ireland. We acknowledge
the assistance of the technical staff of the School of Biochemistry
and Immunology, Biomedical Sciences Institute, Trinity College
Dublin, Ireland, in particular, Mr. Barry Moran of the Flow
Cytometry Facility.
Abbreviations:
|
ALL
|
acute lymphoblastic leukaemia
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
cFLIP
|
cellular FLICE (FADD-like
IL-1β-converting enzyme)-inhibitory protein
|
|
CI
|
combination index
|
|
CLL
|
chronic lymphocytic leukemia
|
|
CML
|
chronic myeloid leukemia
|
|
DISC
|
death inducing signaling complex
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DR
|
death receptor
|
|
FBS
|
fetal bovine serum
|
|
GAPDH
|
glyceraldehyde 3-phosphate
dehydrogenase
|
|
GSK-3β
|
glycogen synthase kinase 3β
|
|
IAP
|
inhibitor of apoptosis protein
|
|
JC-1
|
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolyl-carbocyanine
iodide
|
|
LY-294002
|
2-(4-Morpholinyl)-8-phenyl-1(4H)-benzopyran-4-one hydrochloride
|
|
Mcl-1
|
myeloid cell leukemia 1
|
|
MIM
|
mitochondrial inner membrane
|
|
MMP
|
mitochondrial membrane potential
|
|
MTAs
|
microtubular-targeting agents
|
|
mTOR
|
mammalian target of rapamycin
|
|
PARP
|
poly (ADP-ribose) polymerase
|
|
PBOX
|
pyrrolo-1,5-benzoxazepine
|
|
PBOX-15
|
4-acetoxy-5-(1-naphthyl)naphtho[2,3-b]pyrrolo[2,1-d][1,4]oxazepine
|
|
PBOX-6
|
7-[(N,N-dimethylcarbamoyl)oxy]-6-(naphth-1-yl)pyrrolo[2,1-d]
[1,5]benzoxazepine
|
|
PDK1
|
phosphoinositide-dependent protein
kinase 1
|
|
PI
|
propidium iodide
|
|
PI3K
|
phosphoinositide-3-kinase
|
|
PIP3
|
phos phatidylinositol
(3,4,5)-trisphosphate
|
|
PTEN
|
phosphatase and tensin homologue
deleted on chromosome ten
|
|
RPMI
|
Roswell Park Memorial Institute
|
|
TNF
|
tumour necrosis factor
|
|
TRAIL
|
tumour necrosis factor-related
apoptosis-inducing ligand
|
|
XIAP
|
X-linked inhibitor of apoptosis
protein
|
References
|
1
|
Inaba H, Greaves M and Mullighan CG: Acute
lymphoblastic leukaemia. Lancet. 381:1943–1955. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Banihashem A, Ghasemi A, Ghaemi N, Moazzen
N and Amirabadi A: Prevalence of transient hyperglycemia and
diabetes mellitus in pediatric patients with acute leukemia. Iran J
Ped Hematol Oncol. 4:5–10. 2014.PubMed/NCBI
|
|
3
|
Woo JS, Alberti MO and Tirado CA:
Childhood B-acute lymphoblastic leukemia: A genetic update. Exp
Hematol Oncol. 3:162014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guicciardi ME and Gores GJ: Life and death
by death receptors. FASEB J. 23:1625–1637. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
French LE and Tschopp J: Protein-based
therapeutic approaches targeting death receptors. Cell Death
Differ. 10:117–123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prasad S, Kim JH, Gupta SC and Aggarwal
BB: Targeting death receptors for TRAIL by agents designed by
Mother Nature. Trends Pharmacol Sci. 35:520–536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Daniels RA, Turley H, Kimberley FC, Liu
XS, Mongkolsapaya J, Ch'En P, Xu XN, Jin BQ, Pezzella F and
Screaton GR: Expression of TRAIL and TRAIL receptors in normal and
malignant tissues. Cell Res. 15:430–438. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang S and El-Deiry WS: TRAIL and
apoptosis induction by TNF-family death receptors. Oncogene.
22:8628–8633. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stuckey DW and Shah K: TRAIL on trial:
Preclinical advances in cancer therapy. Trends Mol Med. 19:685–694.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bellail AC, Qi L, Mulligan P, Chhabra V
and Hao C: TRAIL agonists on clinical trials for cancer therapy:
The promises and the challenges. Rev Recent Clin Trials. 4:34–41.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mahalingam D, Szegezdi E, Keane M, de Jong
S and Samali A: TRAIL receptor signalling and modulation: Are we on
the right TRAIL? Cancer Treat Rev. 35:280–288. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ashkenazi A, Holland P and Eckhardt SG:
Ligand-based targeting of apoptosis in cancer: The potential of
recombinant human apoptosis ligand 2/Tumor necrosis factor-related
apoptosis-inducing ligand (rhApo2L/TRAIL). J Clin Oncol.
26:3621–3630. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ehrhardt H, Fulda S, Schmid I, Hiscott J,
Debatin KM and Jeremias I: TRAIL induced survival and proliferation
in cancer cells resistant towards TRAIL-induced apoptosis mediated
by NF-kappaB. Oncogene. 22:3842–3852. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fakler M, Loeder S, Vogler M, Schneider K,
Jeremias I, Debatin KM and Fulda S: Small molecule XIAP inhibitors
cooperate with TRAIL to induce apoptosis in childhood acute
leukemia cells and overcome Bcl-2-mediated resistance. Blood.
113:1710–1722. 2009. View Article : Google Scholar
|
|
15
|
Hellwig CT and Rehm M: TRAIL signaling and
synergy mechanisms used in TRAIL-based combination therapies. Mol
Cancer Ther. 11:3–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leong S, Cohen RB, Gustafson DL, Langer
CJ, Camidge DR, Padavic K, Gore L, Smith M, Chow LQ, von Mehren M,
et al: Mapatumumab, an antibody targeting TRAIL-R1, in combination
with paclitaxel and carboplatin in patients with advanced solid
malignancies: Results of a phase I and pharmacokinetic study. J
Clin Oncol. 27:4413–4421. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maginn EN, Browne PV, Hayden P,
Vandenberghe E, MacDonagh B, Evans P, Goodyer M, Tewari P, Campiani
G, Butini S, et al: PBOX-15, a novel microtubule targeting agent,
induces apoptosis, upregulates death receptors, and potentiates
TRAIL-mediated apoptosis in multiple myeloma cells. Br J Cancer.
104:281–289. 2011. View Article : Google Scholar :
|
|
18
|
Mulligan JM, Greene LM, Cloonan S, Mc Gee
MM, Onnis V, Campiani G, Fattorusso C, Lawler M, Williams DC and
Zisterer DM: Identification of tubulin as the molecular target of
proapoptotic pyrrolo-1,5-benzoxazepines. Mol Pharmacol. 70:60–70.
2006.PubMed/NCBI
|
|
19
|
Lysaght J, Verma NK, Maginn EN, Ryan JM,
Campiani G, Zisterer DM, Williams DC, Browne PV, Lawler MP and
McElligott AM: The microtubule targeting agent PBOX-15 inhibits
integrin-mediated cell adhesion and induces apoptosis in acute
lymphoblastic leukaemia cells. Int J Oncol. 42:239–246. 2013.
|
|
20
|
McElligott AM, Maginn EN, Greene LM,
McGuckin S, Hayat A, Browne PV, Butini S, Campiani G, Catherwood
MA, Vandenberghe E, et al: The novel tubulin-targeting agent
pyrrolo-1,5-benzoxazepine-15 induces apoptosis in poor prognostic
subgroups of chronic lymphocytic leukemia. Cancer Res.
69:8366–8375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Greene LM, Kelly L, Onnis V, Campiani G,
Lawler M, Williams DC and Zisterer DM: STI-571 (imatinib mesylate)
enhances the apoptotic efficacy of pyrrolo-1,5-benzoxazepine-6, a
novel microtubule-targeting agent, in both STI-571-sensitive and
-resistant Bcr-Abl-positive human chronic myeloid leukemia cells. J
Pharmacol Exp Ther. 321:288–297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zisterer DM, McGee MM, Campiani G, Ramunno
A, Fattorusso C, Nacci V, Lawler M and Williams DC:
Pyrrolo-1,5-benzoxazepines: A new class of apoptotic agents.
Biochem Soc Trans. 29:704–706. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nathwani SM, Butler S, Fayne D, McGovern
NN, Sarkadi B, Meegan MJ, Lloyd DG, Campiani G, Lawler M, Williams
DC, et al: Novel microtubule-targeting agents,
pyrrolo-1,5-benzoxazepines, induce apoptosis in
multi-drug-resistant cancer cells. Cancer Chemother Pharmacol.
66:585–596. 2010. View Article : Google Scholar
|
|
24
|
Bright SA, McElligott AM, O'Connell JW,
O'Connor L, Carroll P, Campiani G, Deininger MW, Conneally E,
Lawler M, Williams DC, et al: Novel pyrrolo-1,5-benzoxazepine
compounds display significant activity against resistant chronic
myeloid leukaemia cells in vitro, in ex vivo patient samples and in
vivo. Br J Cancer. 102:1474–1482. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mc Gee MM, Gemma S, Butini S, Ramunno A,
Zisterer DM, Fattorusso C, Catalanotti B, Kukreja G, Fiorini I,
Pisano C, et al: Pyrrolo[1,5]benzoxa(thia)zepines as a new class of
potent apoptotic agents. Biological studies and identification of
an intracellular location of their drug target. J Med Chem.
48:4367–4377. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
van der Sloot AM, Mullally MM,
Fernandez-Ballester G, Serrano L and Quax WJ: Stabilization of
TRAIL, an all-beta-sheet multimeric protein, using computational
redesign. Protein Eng Des Sel. 17:673–680. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van der Sloot AM, Tur V, Szegezdi E,
Mullally MM, Cool RH, Samali A, Serrano L and Quax WJ: Designed
tumor necrosis factor-related apoptosis-inducing ligand variants
initiating apoptosis exclusively via the DR5 receptor. Proc Natl
Acad Sci USA. 103:8634–8639. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Greene LM, Campiani G, Lawler M, Williams
DC and Zisterer DM: BubR1 is required for a sustained mitotic
spindle checkpoint arrest in human cancer cells treated with
tubulin-targeting pyrrolo-1,5-benzoxazepines. Mol Pharmacol.
73:419–430. 2008. View Article : Google Scholar
|
|
29
|
Nathwani SM, Cloonan SM, Stronach M,
Campiani G, Lawler M, Williams DC and Zisterer DM: Novel
microtubule-targeting agents, pyrrolo-1,5-benzoxazepines, induce
cell cycle arrest and apoptosis in prostate cancer cells. Oncol
Rep. 24:1499–1507. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sung B, Park B, Yadav VR and Aggarwal BB:
Celastrol, a triterpene, enhances TRAIL-induced apoptosis through
the down-regulation of cell survival proteins and up-regulation of
death receptors. J Biol Chem. 285:11498–11507. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nimmanapalli R, Perkins CL, Orlando M,
O'Bryan E, Nguyen D and Bhalla KN: Pretreatment with paclitaxel
enhances apo-2 ligand/tumor necrosis factor-related
apoptosis-inducing ligand-induced apoptosis of prostate cancer
cells by inducing death receptors 4 and 5 protein levels. Cancer
Res. 61:759–763. 2001.PubMed/NCBI
|
|
32
|
Hunter TB, Manimala NJ, Luddy KA, Catlin T
and Antonia SJ: Paclitaxel and TRAIL synergize to kill
paclitaxel-resistant small cell lung cancer cells through a
caspase-independent mechanism mediated through AIF. Anticancer Res.
31:3193–3204. 2011.PubMed/NCBI
|
|
33
|
Irmler M, Thome M, Hahne M, Schneider P,
Hofmann K, Steiner V, Bodmer JL, Schröter M, Burns K, Mattmann C,
et al: Inhibition of death receptor signals by cellular FLIP.
Nature. 388:190–195. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fulda S: Inhibitor of apoptosis (IAP)
proteins in hematological malignancies: Molecular mechanisms and
therapeutic opportunities. Leukemia. 28:1414–1422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen D and Zhou Q: Caspase cleavage of
BimEL triggers a positive feedback amplification of apoptotic
signaling. Proc Natl Acad Sci USA. 101:1235–1240. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tabellini G, Tazzari PL, Bortul R,
Evangelisti C, Billi AM, Grafone T, Martinelli G, Baccarani M and
Martelli AM: Phosphoinositide 3-kinase/Akt inhibition increases
arsenic trioxide-induced apoptosis of acute promyelocytic and
T-cell leukaemias. Br J Haematol. 130:716–725. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Franke TF: PI3K/Akt: Getting it right
matters. Oncogene. 27:6473–6488. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mérino D, Lalaoui N, Morizot A, Solary E
and Micheau O: TRAIL in cancer therapy: Present and future
challenges. Expert Opin Ther Targets. 11:1299–1314. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akahane K, Inukai T, Zhang X, Hirose K,
Kuroda I, Goi K, Honna H, Kagami K, Nakazawa S, Endo K, et al:
Resistance of T-cell acute lymphoblastic leukemia to tumor necrosis
factor--related apoptosis-inducing ligand-mediated apoptosis. Exp
Hematol. 38:885–895. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zheng T, Fu JJ, Hu L, Qiu F, Hu M, Zhu JJ,
Hua ZC and Wang H: Nanoarchitectured electrochemical cytosensors
for selective detection of leukemia cells and quantitative
evaluation of death receptor expression on cell surfaces. Anal
Chem. 85:5609–5616. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim K, Fisher MJ, Xu SQ and el-Deiry WS:
Molecular determinants of response to TRAIL in killing of normal
and cancer cells. Clin Cancer Res. 6:335–346. 2000.PubMed/NCBI
|
|
42
|
Zhang Y and Zhang B: TRAIL resistance of
breast cancer cells is associated with constitutive endocytosis of
death receptors 4 and 5. Mol Cancer Res. 6:1861–1871. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
McDonald ER III, Chui PC, Martelli PF,
Dicker DT and El-Deiry WS: Death domain mutagenesis of KILLER/DR5
reveals residues critical for apoptotic signaling. J Biol Chem.
276:14939–14945. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kojima Y, Nakayama M, Nishina T, Nakano H,
Koyanagi M, Takeda K, Okumura K and Yagita H: Importin β1
protein-mediated nuclear localization of death receptor 5 (DR5)
limits DR5/tumor necrosis factor (TNF)-related apoptosis-inducing
ligand (TRAIL)-induced cell death of human tumor cells. J Biol
Chem. 286:43383–43393. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sheridan JP, Marsters SA, Pitti RM, Gurney
A, Skubatch M, Baldwin D, Ramakrishnan L, Gray CL, Baker K, Wood
WI, et al: Control of TRAIL-induced apoptosis by a family of
signaling and decoy receptors. Science. 277:818–821. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tong HX, Lu CW, Wang QS and Ma LY:
Combination of IFNγ and chemotherapeutic agents increase TRAIL
sensitivity of neuroblastoma cell lines. Eur J Pediatr Surg.
21:304–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kim HR, Lee MW, Kim DS, Jo HY, Lee SH,
Chueh HW, Jung HL, Yoo KH, Sung KW and Koo HH: Etoposide sensitizes
neuroblastoma cells expressing caspase 8 to TRAIL. Cell Biol Int
Rep 2010. 19:e000172012.PubMed/NCBI
|
|
48
|
Timur M, Cort A, Ozdemir E, Sarikcioglu
SB, Sanlioglu S, Sanlioglu AD and Ozben T: Bleomycin induced
sensitivity to TRAIL/Apo-2L-mediated apoptosis in human
seminomatous testicular cancer cells is correlated with
upregulation of death receptors. Anticancer Agents Med Chem.
15:99–106. 2015. View Article : Google Scholar
|
|
49
|
Venza I, Visalli M, Oteri R, Teti D and
Venza M: Class I-specific histone deacetylase inhibitor MS-275
overrides TRAIL-resistance in melanoma cells by downregulating
c-FLIP. Int Immunopharmacol. 21:439–446. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Green DR: Apoptotic pathways: Paper wraps
stone blunts scissors. Cell. 102:1–4. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Schug ZT, Gonzalvez F, Houtkooper RH, Vaz
FM and Gottlieb E: BID is cleaved by caspase-8 within a native
complex on the mitochondrial membrane. Cell Death Differ.
18:538–548. 2011. View Article : Google Scholar :
|
|
52
|
Safa AR: c-FLIP, a master anti-apoptotic
regulator. Exp Oncol. 34:176–184. 2012.PubMed/NCBI
|
|
53
|
Safa AR: Roles of c-FLIP in apoptosis,
necroptosis, and autophagy. J Carcinog Mutagen (Suppl).
6:0032013.
|
|
54
|
Zang F, Wei X, Leng X, Yu M and Sun B:
C-FLIP(L) contributes to TRAIL resistance in HER2-positive breast
cancer. Biochem Biophys Res Commun. 450:267–273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ding L, Yuan C, Wei F, Wang G, Zhang J,
Bellail AC, Zhang Z, Olson JJ and Hao C: Cisplatin restores TRAIL
apoptotic pathway in glioblastoma-derived stem cells through
up-regulation of DR5 and down-regulation of c-FLIP. Cancer Invest.
29:511–520. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ding J, Polier G, Köhler R, Giaisi M,
Krammer PH and Li-Weber M: Wogonin and related natural flavones
overcome tumor necrosis factor-related apoptosis-inducing ligand
(TRAIL) protein resistance of tumors by down-regulation of c-FLIP
protein and up-regulation of TRAIL receptor 2 expression. J Biol
Chem. 287:641–649. 2012. View Article : Google Scholar :
|
|
57
|
Li LC, Jayaram S, Ganesh L, Qian L,
Rotmensch J, Maker AV and Prabhakar BS: Knockdown of MADD and
c-FLIP overcomes resistance to TRAIL-induced apoptosis in ovarian
cancer cells. Am J Obstet Gynecol. 205:362.e12–362.e25. 2011.
View Article : Google Scholar
|
|
58
|
Bose P and Grant S: Mcl-1 as a therapeutic
target in acute myelogenous leukemia (AML). Leuk Res Rep. 2:12–14.
2013.PubMed/NCBI
|
|
59
|
Scarfò L and Ghia P: Reprogramming cell
death: BCL2 family inhibition in hematological malignancies.
Immunol Lett. 155:36–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
McGee MM, Greene LM, Ledwidge S, Campiani
G, Nacci V, Lawler M, Williams DC and Zisterer DM: Selective
induction of apoptosis by the pyrrolo-1,5-benzoxazepine
7-[[dimethylcarbamoyl] oxy]-6-(2-naphthyl)pyrrolo-[2,1-d]
(1,5)-benzoxazepine (PBOX-6) in Leukemia cells occurs via the c-Jun
NH2-terminal kinase-dependent phosphorylation and inactivation of
Bcl-2 and Bcl-XL. J Pharmacol Exp Ther. 310:1084–1095. 2004.
View Article : Google Scholar
|
|
61
|
Lennon JC, Bright SA, Carroll E, Butini S,
Campiani G, O'Meara A, Williams DC and Zisterer DM: The novel
pyrrolo-1,5-benzoxazepine, PBOX-6, synergistically enhances the
apoptotic effects of carboplatin in drug sensitive and multidrug
resistant neuroblastoma cells. Biochem Pharmacol. 87:611–624. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Greene LM, Nolan DP, Regan-Komito D,
Campiani G, Williams DC and Zisterer DM: Inhibition of late-stage
autophagy synergistically enhances
pyrrolo-1,5-benzoxazepine-6-induced apoptotic cell death in human
colon cancer cells. Int J Oncol. 43:927–935. 2013.PubMed/NCBI
|
|
63
|
Gill C, Dowling C, O'Neill AJ and Watson
RW: Effects of cIAP-1, cIAP-2 and XIAP triple knockdown on prostate
cancer cell susceptibility to apoptosis, cell survival and
proliferation. Mol Cancer. 8:392009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Finlay D, Vamos M, González-López M,
Ardecky RJ, Ganji SR, Yuan H, Su Y, Cooley TR, Hauser CT, Welsh K,
et al: Small-molecule IAP antagonists sensitize cancer cells to
TRAIL-induced apoptosis: Roles of XIAP and cIAPs. Mol Cancer Ther.
13:5–15. 2014. View Article : Google Scholar :
|
|
65
|
Guicciardi ME, Mott JL, Bronk SF, Kurita
S, Fingas CD and Gores GJ: Cellular inhibitor of apoptosis 1
(cIAP-1) degradation by caspase 8 during TNF-related
apoptosis-inducing ligand (TRAIL)-induced apoptosis. Exp Cell Res.
317:107–116. 2011. View Article : Google Scholar
|
|
66
|
Lanuti P, Bertagnolo V, Pierdomenico L,
Bascelli A, Santavenere E, Alinari L, Capitani S, Miscia S and
Marchisio M: Enhancement of TRAIL cytotoxicity by AG-490 in human
ALL cells is characterized by downregulation of cIAP-1 and cIAP-2
through inhibition of Jak2/Stat3. Cell Res. 19:1079–1089. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: Its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gomes AM, Soares MV, Ribeiro P, Caldas J,
Póvoa V, Martins LR, Melão A, Serra-Caetano A, de Sousa AB, Lacerda
JF, et al: Adult B-cell acute lymphoblastic leukemia cells display
decreased PTEN activity and constitutive hyperactivation of
PI3K/Akt pathway despite high PTEN protein levels. Haematologica.
99:1062–1068. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Badura S, Tesanovic T, Pfeifer H, Wystub
S, Nijmeijer BA, Liebermann M, Falkenburg JH, Ruthardt M and
Ottmann OG: Differential effects of selective inhibitors targeting
the PI3K/AKT/mTOR pathway in acute lymphoblastic leukemia. PLoS
One. 8:e800702013. View Article : Google Scholar : PubMed/NCBI
|