Introduction
Prostate cancer (PCa) is the most frequently
diagnosed cancer and the second leading cause of cancer-related
death among men in developed countries (1). Most patients with PCa are initially
responsive to androgen deprivation therapy; however, their cancers
eventually become resistant to androgen-deprivation therapy and
progress to castration-resistant prostate cancer (CRPC). Several
novel treatments have recently been developed for patients with
advanced PCa. These treatments, however, cannot completely control
the progression and metastasis of PCa (2). Therefore, more effective treatment
strategies, based on current genomic approaches, are required.
Moreover, because it is difficult to obtain clinical specimens of
CRPC, there are very few reports describing the genomic analysis of
CRPC.
Small non-coding RNAs known as microRNAs (miRNAs)
have been shown to regulate gene expression by repressing
translation or cleaving RNA transcripts in a sequence-dependent
manner (3). At present, a
substantial amount of evidence has suggested that miRNAs are
aberrantly expressed in many human cancers, including PCa, and play
significant roles in human oncogenesis, metastasis, and drug
resistance (4–7). Several reports have shown that miRNAs
regulate ~30–60% (or more) of the protein-coding genes in the human
genome by bioinformatics predictions (7,8).
Thus, normal regulatory mechanisms of RNA networks can be disrupted
by the aberrant expression of tumor-suppressive or oncogenic miRNAs
in cancer cells. We propose that identification of aberrantly
expressed miRNAs in clinical specimens is an important first step
toward elucidating the details of RNA networks in cancer cells.
To identify novel RNA networks in PCa, we have
previously constructed miRNA expression signatures using
androgen-dependent PCa clinical specimens (9,10).
Using this signature, we have identified tumor-suppressive
miRNA-mediated PCa oncogenic pathways (11–16).
More recently, we constructed miRNA expression signatures using
CRPC clinical specimens and reported that the miRNA cluster
miR-221/miR-222 functions as a tumor suppressor in PCa and
CRPC cells (17). Moreover,
miR-223 expression has been shown to be significantly
reduced in PCa and CRPC specimens and to act as a tumor suppressor
by targeting integrin-A3/B1 oncogenic signaling (18). The CRPC expression signature
reported by our laboratory has provided important information
needed to elucidate novel RNA networks in CRPC cells.
In this study, we focused on miR-320a because
downregulation of miR-320a was observed in our two miRNA
signatures for androgen-dependent PCa and CRPC clinical specimens
(9,17). The aim of this study was to
investigate the functional significance of miR-320a and to
identify the molecular targets and pathways mediated by
miR-320a in PCa and CRPC cells. Our data showed that
restoration of mature miR-320a in PC3 and DU145 PCa cells
significantly inhibited cancer cell migration and invasion. Direct
regulation of lysosomal-associated membrane protein 1
(LAMP1) by miR-320a was observed in PCa cells.
Silencing of LAMP1 using specific small interfering RNA
(siRNA; si-LAMP1) significantly inhibited cell
proliferation, migration, and invasion in PCa cells. The discovery
of tumor-suppressive miR-320a-mediated molecular pathways
provides new insights into the potential mechanisms of metastasis
in PCa and suggests novel therapeutic strategies for the treatment
of this disease.
Materials and methods
Clinical PCa specimens, cell lines, and
RNA isolation
Clinical prostate specimens (PCa and normal prostate
tissues) collected by needle biopsy or autopsy were obtained from
patients admitted to Teikyo University Chiba Medical Center
Hospital from 2008 to 2013. Twenty-nine prostate samples [PCa,
n=15; non-cancerous prostate tissues (non-PCa), n=14] were obtained
by transrectal prostate needle biopsy from patients with elevated
serum prostate-specific antigen (PSA) levels, and eight metastatic
PCa samples were obtained from patients who died of CRPC. The
backgrounds of the patients are summarized in Table I. All patients provided written
informed consent for tissue donation for research purposes. The
protocol was approved by the Institutional Review Boards of Chiba
University and Teikyo University.
 | Table IPatient characteristics. |
Table I
Patient characteristics.
| No. | Diagnosis | Age (years) | PSA (ng/ml) | Gleason score | cT | cN | cM |
|---|
| 1 | Non-PCa | 54 | 5.4 | - | - | - | - |
| 2 | Non-PCa | 60 | 5.6 | - | - | - | - |
| 3 | Non-PCa | 67 | 5.9 | - | - | - | - |
| 4 | Non-PCa | 67 | 8.1 | - | - | - | - |
| 5 | Non-PCa | 60 | 14 | - | - | - | - |
| 6 | Non-PCa | 69 | 6 | - | - | - | - |
| 7 | Non-PCa | 56 | 8.4 | - | - | - | - |
| 8 | Non-PCa | 61 | 8.6 | - | - | - | - |
| 9 | Non-PCa | 62 | 35.5 | - | - | - | - |
| 10 | Non-PCa | 57 | 5.2 | - | - | - | - |
| 11 | Non-PCa | 64 | 4.4 | - | - | - | - |
| 12 | Non-PCa | 60 | 5.7 | - | - | - | - |
| 13 | Non-PCa | 63 | 11.4 | - | - | - | - |
| 14 | Non-PCa | 65 | 13.2 | - | - | - | - |
| 15 | PCa | 65 | 277 | 4+5 | 4 | 1 | 1 |
| 16 | PCa | 73 | 478 | 4+3 | 3b | 0 | 1 |
| 17 | PCa | 75 | 1,000 | 4+5 | 4 | 1 | 1 |
| 18 | PCa | 79 | 63.2 | 4+5 | 3b | 1 | 1 |
| 19 | PCa | 69 | 95.6 | 4+4 | 4 | 1 | 1 |
| 20 | PCa | 70 | 248 | 3+4 | 3a | 1 | 0 |
| 21 | PCa | 66 | 36.1 | 4+5 | 3a | 1 | 0 |
| 22 | PCa | 81 | 1,338 | 4+5 | 4 | 0 | 1 |
| 23 | PCa | 72 | 102 | 4+4 | 3a | 0 | 0 |
| 24 | PCa | 65 | 212 | 4+4 | 4 | 1 | 1 |
| 25 | PCa | 81 | 11.4 | 4+4 | 2 | 0 | 0 |
| 26 | PCa | 75 | 22.7 | 4+4 | 3b | 0 | 0 |
| 27 | PCa | 73 | 467 | 4+4 | 3b | 1 | 0 |
| 28 | PCa | 58 | 482 | 4+4 | 3b | 1 | 0 |
| 29 | PCa | 73 | 76.4 | 4+4 | 3a | 0 | 0 |
| 30 | CRPC liver
metastasis | 64 | 4,100 | 4+5 | 3b | 0 | 1b |
| 31 | CRPC lymph node
metastasis | | | | | | |
| 32 | CRPC bone
metastasis | | | | | | |
| 33 | CRPC bone
metastasis | 75 | 4,690 | 4+5 | 3b | 0 | 0 |
| 34 | CRPC lung
metastasis | | | | | | |
| 35 | CRPC liver
metastasis | 78 | 3,850 | 3+5 | 4 | 0 | 1b |
| 36 | CRPC dural
metastasis | | | | | | |
| 37 | CRPC bone
metastasis | | | | | | |
Human prostate cancer cells (PC3 and DU145 cells)
and normal prostate cells (RWPE-1) were obtained from the American
Type Culture Collection (Manassas, VA, USA). PC3 and DU145 cells
were maintained in RPMI-1640 medium supplemented with 10% fetal
bovine serum (FBS) in a humidified atmosphere of 5% CO2
and 95% air at 37°C. RWPE-1 cells were cultured in keratinocyte
serum-free medium containing 5 ng/ml epidermal growth factor and 50
μg/ml bovine pituitary extract.
Total RNA was isolated using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer's
protocol as described previously (14,17,18).
Quantitative real-time reverse
transcription polymerase chain reaction (RT-PCR)
The expression levels of miR-320a were
analyzed by TaqMan quantitative real-time PCR (assay ID: 0000542;
Applied Biosystems, Foster City, CA, USA) and normalized to the
expression of RNU48 (assay ID: 001006; Applied Biosystems).
TaqMan probes and primers for LAMP1 (P/N: Hs00174766_m1;
Applied Biosystems), GAPDH (the internal control; P/N:
Hs02758991_m1; Applied Biosystems) and GUSB (the internal
control; P/N: Hs00939627_m1; Applied Biosystems) were
assay-on-demand gene expression products. The procedure for PCR
quantification was described previously (14,17,18).
Transfection with mature miRNA and
siRNA
The following mature miRNA species were used in this
study: mature miRNA and Pre-miR miRNA Precursor
(hsa-miR-320a; P/N: 4427975; Applied Biosystems). The
following siRNAs were used: Stealth Select RNAi siRNA,
si-LAMP1 (cat no. HSS180593, HSS180594; Invitrogen) and
negative control miRNA/siRNA (P/N: AM17111; Applied Biosystems).
Transfection procedures and transfection efficiencies of miRNA in
PC3 and DU145 cells were described previously (14,17,18).
Cell proliferation, migration, and
invasion assays
PC3 and DU145 cells were transfected with 10 nM
miRNAs or siRNAs by reverse transfection. Cell proliferation was
determined by XTT assay using a Cell Proliferation Kit II (Roche
Applied Sciences, Tokyo, Japan). Cell migration activity was
analyzed using uncoated Transwell polycarbonate membrane filters.
Cell invasion was evaluated using modified Boyden chambers
containing Transwell-precoated Matrigel membrane filter inserts.
These assays were performed as described previously (14,17,18).
Selection of putative target genes
regulated by miR-320a in PCa cells
To identify miR-320a target genes, we used
in silico analysis and genome-wide gene expression analysis.
First, we used TargetScan Release 7.0 (http://www.targetscan.org/) to search for genes
containing the miR-320a seed sequence in the 3′-untranslated
region (UTR). Next, to identify upregulated genes in clinical PCa
specimens, we analyzed a publicly available gene expression dataset
in the GEO database (GEO accession no. GSE29079). Finally, we
attempted to identify miR-320a target genes using
miR-320a-transfected PC3 cells. A SurePrint G3 Human GE 60K
v3 microarray (Agilent Technologies, Santa Clara, CA, USA) was used
for genome-wide expression analysis of miR-320a
transfectants compared with mock-transfected PC3 cells (GEO
accession no. GSE77790). As a result, genes fulfilling the three
following conditions were listed: containing putative
miR-320a target sites, upregulated in PCa clinical
specimens, and downregulated by miR-320a restoration.
Immunohistochemical staining and
scoring
A tissue microarray containing a total of 71
prostate samples, 51 PCa specimens, 10 prostatic intraepithelial
neoplasia (PIN) samples, and 10 normal prostate samples was
obtained from Provitro (Berlin, Germany; cat. no. 4012209, lot no.
146.1P.020212.27). Table III
shows the characteristics of patients included in the tissue
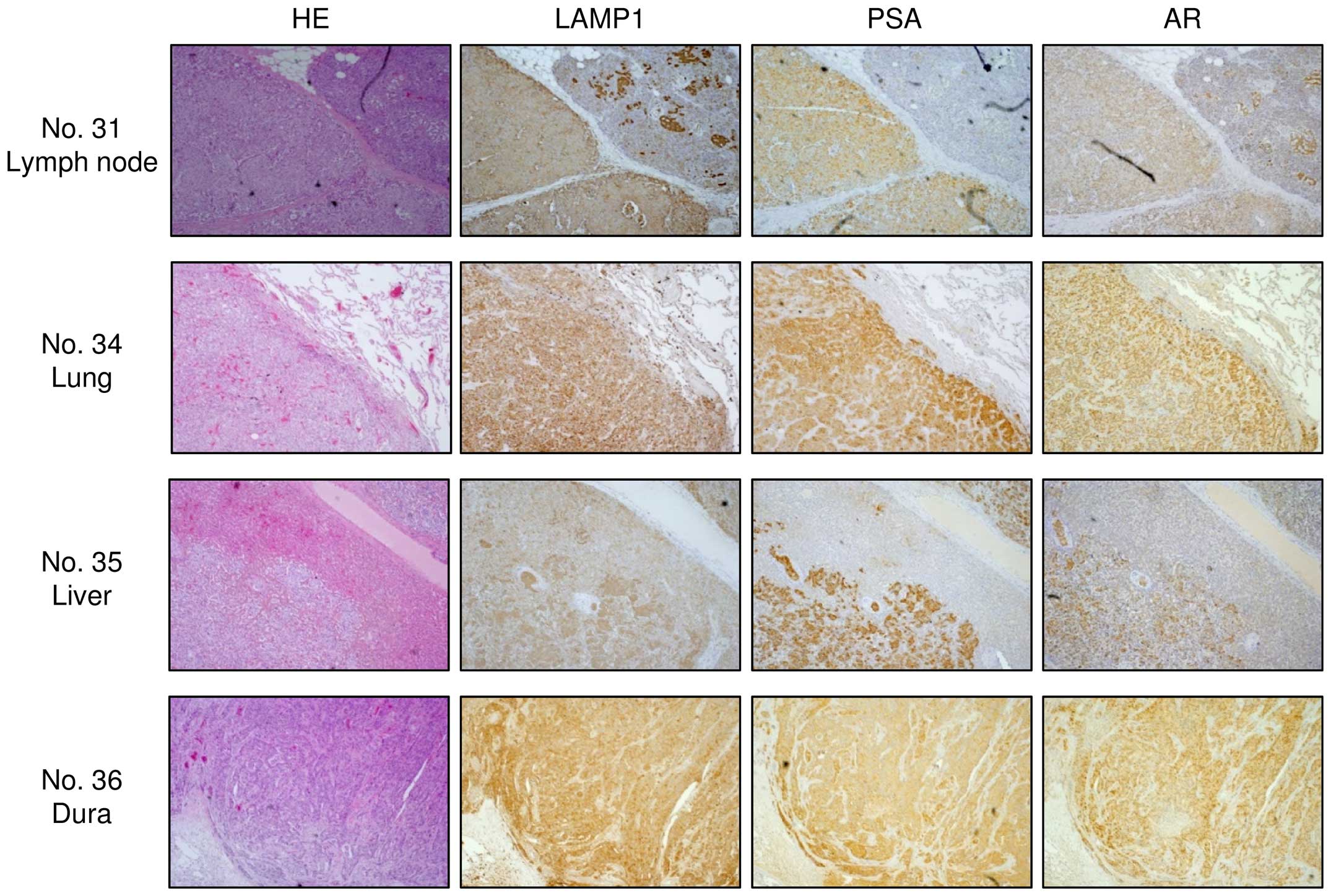
microarray. Four specimens were used as CRPC tissues (Table I, nos. 31 and 34–36). Tissue
specimens were immunostained following the manufacturer's protocol
with the Ultra-Vision Detection System (Thermo Scientific, Fremont,
CA, USA). Primary rabbit polyclonal antibodies against androgen
receptor (AR; 1:50, ab9474; Abcam, Cambridge, UK), antibodies
against PSA (1:500, HPA000764; Sigma-Aldrich, St. Louis, MO, USA),
and antibodies against LAMP1 (1:1,000; #9091; Cell Signaling
Technology, Danvers, MA, USA) were used for immunochemistry. The
slides were treated with biotinylated goat antibodies (Histofine
SAB-PO kit; Nichirei, Tokyo, Japan). The IHC score was the sum of
the score of the weighted intensity and extension of cancerous
area. The intensity of staining was graded from 0 to 3 as follows:
0, negative staining; 1, mild staining; 2, moderate staining; and
3, intense staining. The area of staining in the cancerous area was
graded from 0 to 3 as follows: 0, no staining of cells in any
microscopic field; 1, <30% of malignant cells stained; 2, 30–60%
of malignant cells stained; and 3, >60% of malignant cells
stained. The procedure was carried out as described previously
(14,17,18).
| Table IIICharacteristics of patients included
in the tissue microarray analysis. |
Table III
Characteristics of patients included
in the tissue microarray analysis.
| No. | Diagnosis | Age (years) | Gleason score | Stage pT | Stage pN |
|---|
| 1 | PCa | 64 | 4+3 | 3b | 0 |
| 2 | PCa | 67 | 3+4 | 2b | 0 |
| 3 | PCa | 58 | 3+4 | 2b | 0 |
| 4 | PCa | 63 | 7 | 3b | 0 |
| 5 | PCa | 65 | 3+3 | 2b | 0 |
| 6 | PCa | 61 | 4+4 | 3b | x |
| 7 | PCa | 62 | 3+4 | 2b | x |
| 8 | PCa | 66 | 4+4 | 2b | x |
| 9 | PCa | 61 | 3+4 | 3a | x |
| 10 | PCa | 74 | 4+3 | 2b | x |
| 11 | PCa | 54 | 3+4 | 2c | x |
| 12 | PCa | 68 | 3+4 | 3a | 0 |
| 13 | PCa | 58 | 3+4 | 3a | 0 |
| 14 | PCa | 65 | 3+3 | 2a | 0 |
| 15 | PCa | 77 | 3+4 | 4 | 0 |
| 16 | PCa | 58 | 3+4 | 3a | 0 |
| 17 | PCa | 50 | 4+3 | 2b | 0 |
| 18 | PCa | 53 | 3+3 | 2b | 0 |
| 19 | PCa | 59 | 4+5 | 3a | 0 |
| 20 | PCa | 70 | 2+3 | 2b | 0 |
| 21 | PCa | 65 | 5+4 | 3a | 0 |
| 22 | PCa | 57 | 3+5 | 2b | 0 |
| 23 | PCa | 68 | 4+4 | 2b | 0 |
| 24 | PCa | 58 | 3+3 | 2b | 0 |
| 25 | PCa | 63 | 3+4 | 2b | 0 |
| 26 | PCa | 56 | 3+4 | 2b | 0 |
| 27 | PCa | 63 | 5+3 | 3a | 0 |
| 28 | PCa | 64 | 3+5 | 3a | 0 |
| 29 | PCa | 60 | 3+4 | 2b | 0 |
| 30 | PCa | 60 | 3+3 | 3a | 0 |
| 31 | PCa | 57 | 3+2 | 2b | 0 |
| 32 | PCa | 50 | 3+3 | 2a | 0 |
| 33 | PCa | 68 | 3+3 | 3a | 0 |
| 34 | PCa | 65 | 3+4 | 3b | 1 |
| 35 | PCa | 69 | 5+5 | 3a | 1 |
| 36 | PCa | 51 | 2+3 | 2b | 0 |
| 37 | PCa | 62 | 3+3 | 3a | 0 |
| 38 | PCa | 61 | 3+4 | 3a | 0 |
| 39 | PCa | 53 | 4+4 | 3b | 1 |
| 40 | PCa | 56 | 4+3 | 2b | 0 |
| 41 | PCa | 59 | 2+3 | 2b | 0 |
| 42 | PCa | 61 | 3+4 | 2b | 0 |
| 43 | PCa | 62 | 3+4 | 3b | 1 |
| 44 | PCa | 66 | 3+3 | 3a | 0 |
| 45 | PCa | 62 | 3+3 | 2b | 0 |
| 46 | PCa | 56 | 3+3 | 2b | 0 |
| 47 | PCa | 58 | 3+3 | 3a | 0 |
| 48 | PCa | 66 | 5+4 | 3a | 0 |
| 49 | PCa | 55 | 3+4 | 3a | 0 |
| 50 | PCa | 67 | 2+3 | 2b | 0 |
| 51 | PCa | 61 | 3+5 | 2b | 0 |
| 52 | PIN | 59 | - | - | - |
| 53 | PIN | 58 | - | - | - |
| 54 | PIN | 62 | - | - | - |
| 55 | PIN | 51 | - | - | - |
| 56 | PIN | 58 | - | - | - |
| 57 | PIN | 68 | - | - | - |
| 58 | PIN | 64 | - | - | - |
| 59 | PIN | 56 | - | - | - |
| 60 | PIN | 61 | - | - | - |
| 61 | PIN | 51 | - | - | - |
| 62 | Normal | 70 | - | - | - |
| 63 | Normal | 63 | - | - | - |
| 64 | Normal | 62 | - | - | - |
| 65 | Normal | 81 | - | - | - |
| 66 | Normal | 67 | - | - | - |
| 67 | Normal | 76 | - | - | - |
| 68 | Normal | 66 | - | - | - |
| 69 | Normal | 69 | - | - | - |
| 70 | Normal | 63 | - | - | - |
| 71 | Normal | 71 | - | - | - |
Western blotting
Cells were harvested 72 or 96 h after transfection,
and lysates were prepared. Cell lysates (20 μg protein) were
separated on Mini-PROTEAN TGX gels (Bio-Rad, Hercules, CA, USA) and
transferred to PVDF membranes. Immunoblotting was carried out with
rabbit anti-LAMP1 antibodies (1:2,000; #9091; Cell Signaling
Technology); anti-GAPDH antibodies (1:4,000; ab8245; Abcam) were
used as an internal loading control. Membranes were washed and
incubated with anti-rabbit IgG horseradish peroxidase (HRP)-linked
antibodies (7074; Cell Signaling Technology). Complexes were
visualized with Clarity Western ECL Substrate (Bio-Rad). The
procedure was performed as described previously (14,17,18).
Plasmid construction and dual-luciferase
reporter assays
The partial wild-type sequence of the LAMP1
3′-UTR or that with deletion of the miR-320a target site was
inserted between the XhoI-PmeI restriction sites in
the 3′-UTR of the hRluc gene in the psiDHECK-2 vector
(C8021; Promega, Madison, WI, USA). The procedure for the
dual-luciferase reporter assay was described previously (14,17,18).
Statistical analysis
The relationships between 2 groups and the numerical
values obtained by RT-PCR were analyzed using Mann-Whitney U tests.
The relationships among more than three variables and numerical
values were analyzed using Bonferroni-adjusted Mann-Whitney U
tests. All analyses were performed using Expert StatView (version
5; SAS Institute Inc., Cary, NC, USA).
Results
Expression levels of miR-320a in PCa
specimens and cell lines
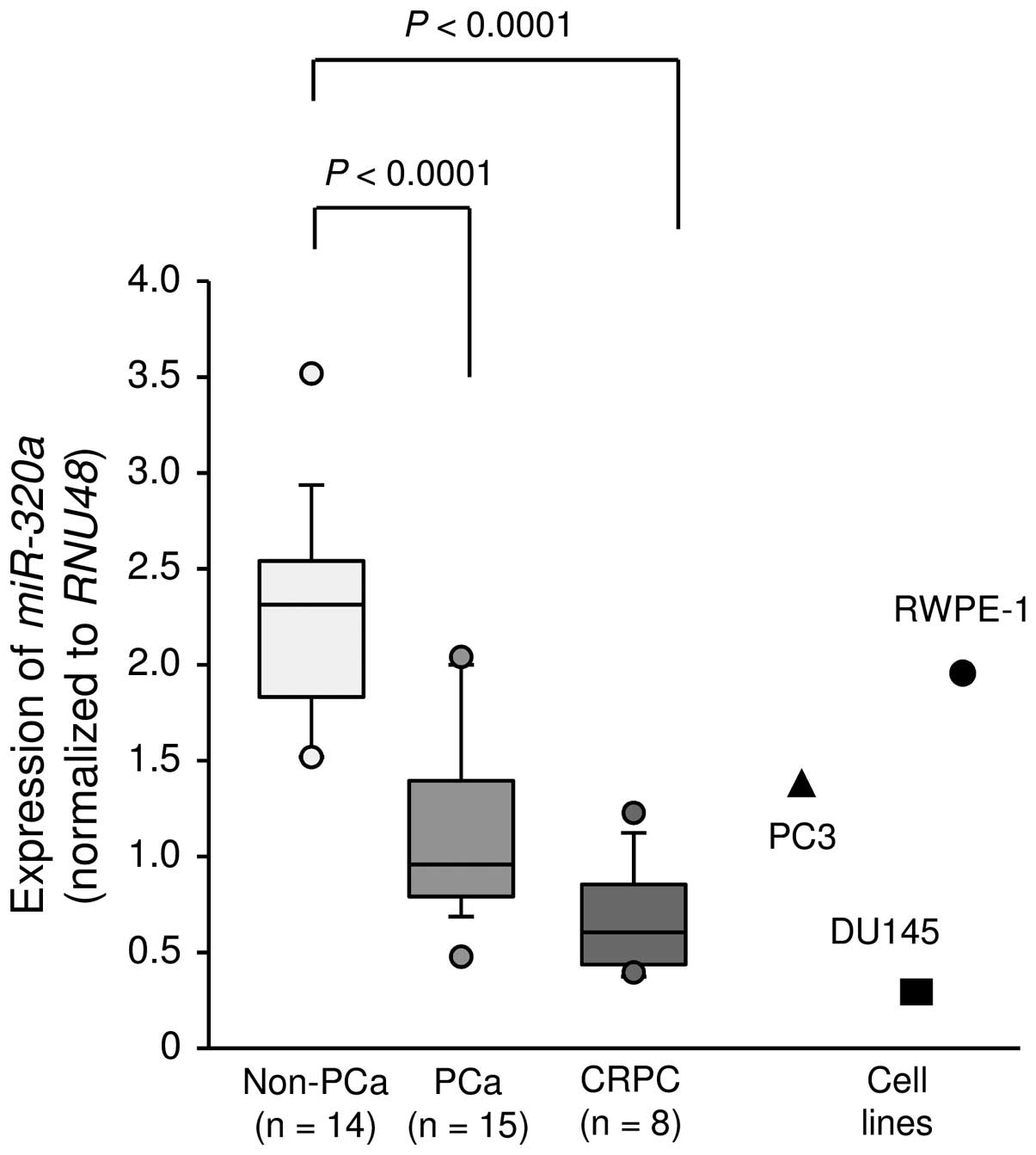
First, we evaluated the expression of
miR-320a in clinical prostate specimens and cell lines. The
median PSA level of patients with normal prostate tissues was 7.05
ng/ml (range, 4.4–35.5 ng/ml), and that of patients with PCa was
212 ng/ml (range, 11.4–1338 ng/ml). Of the patients with
PCa, only 4 patients had organ-confined disease (Table I). The expression levels of
miR-320a were significantly lower in cancer tissues than in
non-cancerous tissues (P<0.0001; Fig. 1). In PC3 and DU145 cells,
expression levels of miR-320a were relatively low compared
with those in clinical specimen and were lower than those in normal
prostate cells (RWPE-1 cells; Fig.
1).
Effects of miR-320a restoration on the
proliferation, migration, and invasion of PCa cells
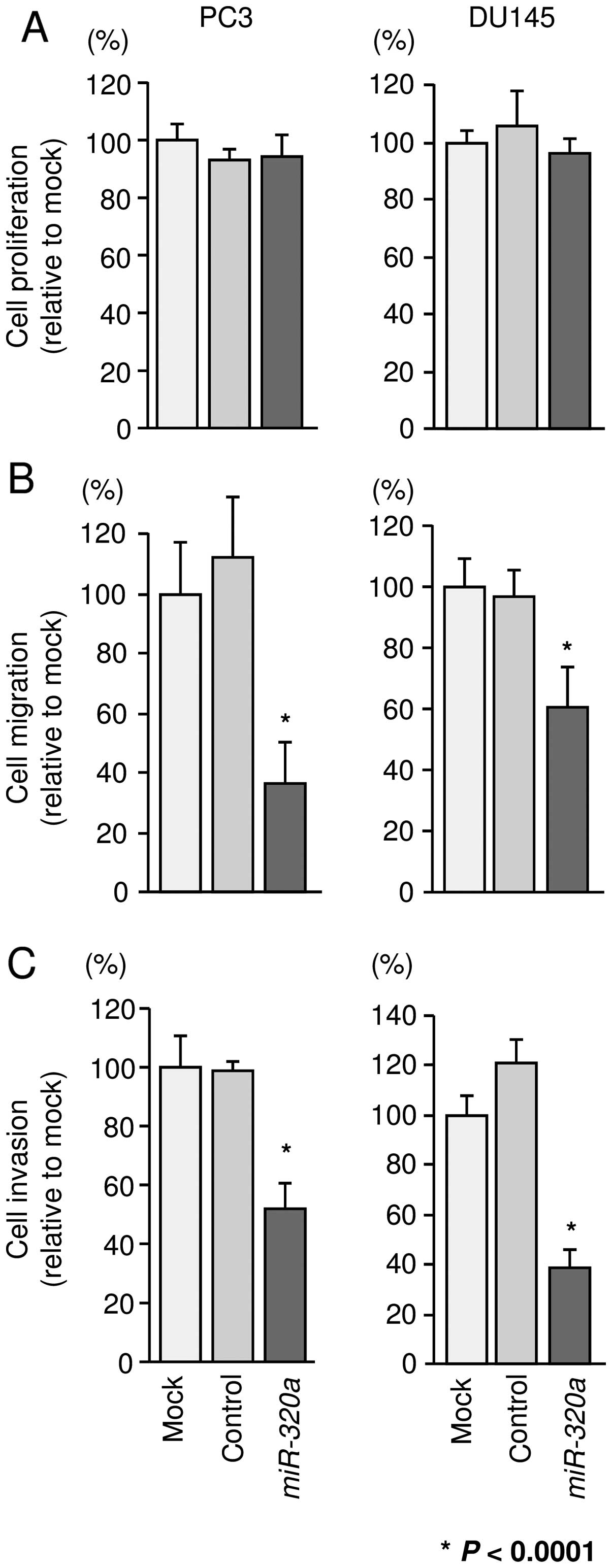
To investigate the functional effects of miR-320a,
we performed gain-of-function studies using miRNA transfection into
PC3 and DU145 cells. XTT assays revealed that cell proliferation
was not inhibited in miR-320a transfectants in comparison
with that in mock- or miR-control-transfected PC3 and DU145 cells
(Fig. 2A). However, cell migration
and invasion activities were significantly inhibited in
miR-320a transfectants in comparison with those in mock- or
miR-control-transfectants (P<0.0001; Fig. 2B and C).
Identification of candidate target genes
of miR-320a in PCa cells
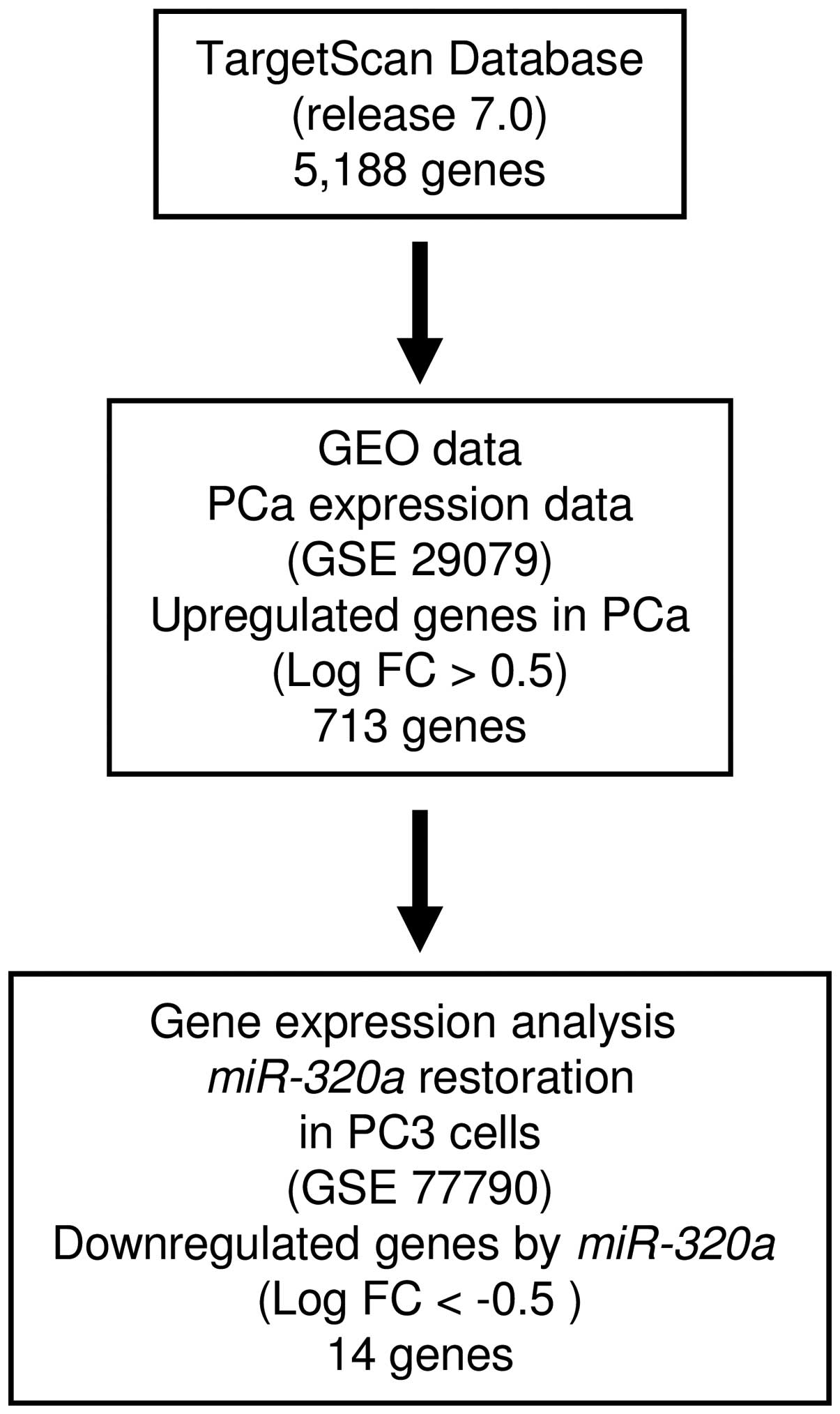
To identify target genes regulated by
miR-320a, we performed a combination of in silico
analysis and genome-wide gene expression analysis. First, we
screened genes having putative 3′-UTR sites matching the seed
sequence of miR-320a. Using the TargetScan online program,
5,188 genes with putative target sites for miR-320a were
selected. Next, to choose upregulated genes in PCa clinical
specimens compared with normal prostatic tissue, the genes were
analyzed with available gene expression data from GEO (accession
no. GSE29079), which compared 48 normal and 47 PCa tissue samples.
As a result, 713 genes upregulated (log2 ratio >0.5)
in PCa tissue were selected. To gain further insight into which
genes were affected by tumor-suppressive miR-320a in PCa, we
carried out genome-wide gene expression analysis using a microarray
comparing miR-320a-transfected and mock-transfected PC3
cells (deposited in the GEO database, accession no. GSE77790).
Consequently, a total of 14 candidate genes were identified as
target genes of miR-320a (Table II). Of these, we focused on the
LAMP1 gene for further analyses because the functional
significance of LAMP1 in PCa had not been reported. A flow
chart outlining our strategy for identification of candidate target
genes of miR-320a is shown in Fig. 3.
| Table IICandidate target genes regulated by
miR-320a in PCa. |
Table II
Candidate target genes regulated by
miR-320a in PCa.
| Gene symbol | Gene name | PC3 miR-320a
transfectant | No. of conserved
sites | No. of poorly
conserved sites | GEO fold
change |
|---|
| CNOT6 | CCR4-NOT
transcription complex, subunit 6 | −1.4261212 | 1 | 4 | 0.539992 |
| VDAC1 | Voltage-dependent
anion channel 1 | −1.2843819 | 1 | 0 | 0.609059 |
| LAMP1 |
Lysosomal-associated membrane protein
1 | −0.8793476 | 1 | 0 | 0.623045 |
| TPD52 | Tumor protein
D52 | −0.8678817 | 1 | 0 | 0.606609 |
| SERBP1 | SERPINE1 mRNA
binding protein 1 | −0.6758766 | 1 | 0 | 0.709015 |
| HELZ | Helicase with zinc
finger | −0.65864474 | 2 | 1 | 0.675884 |
| MYEF2 | Myelin expression
factor 2 | −0.6576734 | 1 | 1 | 0.597199 |
| FUS | Fused in
sarcoma | −0.6464454 | 1 | 1 | 0.568564 |
| NEDD4L | Neural precursor
cell expressed, developmentally downregulated 4-like | −0.6353359 | 1 | 0 | 1.241135 |
| FAM117B | Family with
sequence similarity 117, member B | −0.62850374 | 1 | 4 | 0.729023 |
| NUFIP2 | Nuclear fragile X
mental retardation protein interacting protein 2 | −0.60588837 | 3 | 1 | 0.775047 |
| ZC3H7B | Zinc finger
CCCH-type containing 7B | −0.5742502 | 2 | 1 | 0.571856 |
| RANBP2 | RAN binding protein
2 | −0.55872875 | 1 | 1 | 0.562369 |
| FBXO28 | F-box protein
28 | −0.52747154 | 1 | 0 | 0.591344 |
LAMP1 was strongly expressed in clinical
PCa/CRPC specimens
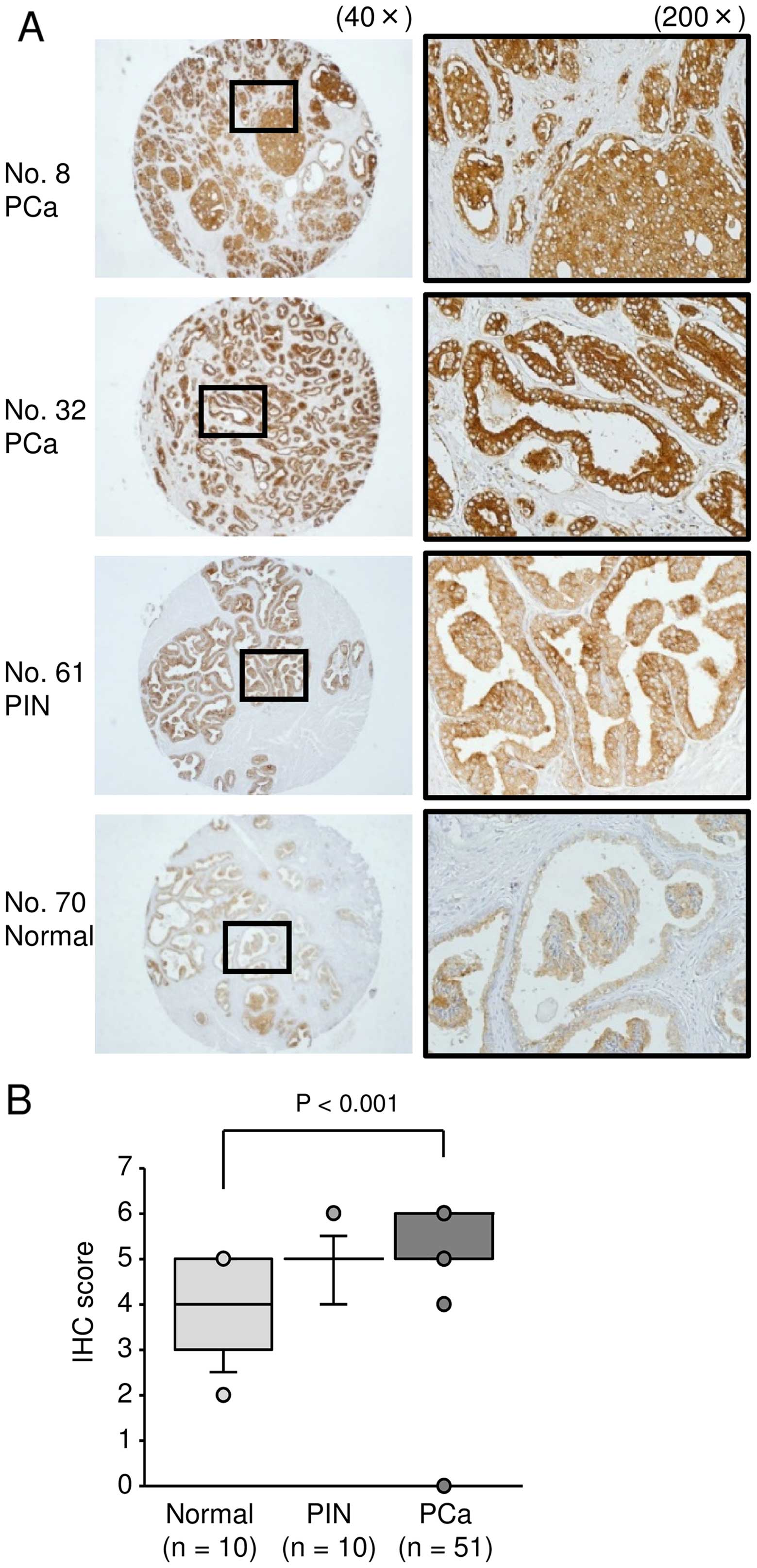
To analyze whether LAMP1 was upregulated in PCa
clinical specimens, we carried out immunohistochemical staining of
PCa, PIN, prostatic hyperplasia (non-PCa), and CRPC tissues. Using
a tissue microarray, we confirmed that LAMP1 expression was
significantly increased in PCa specimens compared with that in
non-PCa specimens (Fig. 4B;
P<0.0001). Representative images of IHC staining for
LAMP1 in the tissue microarray are shown in Fig. 4A. In CRPC specimens, we also found
strong expression of LAMP1 (Fig.
5).
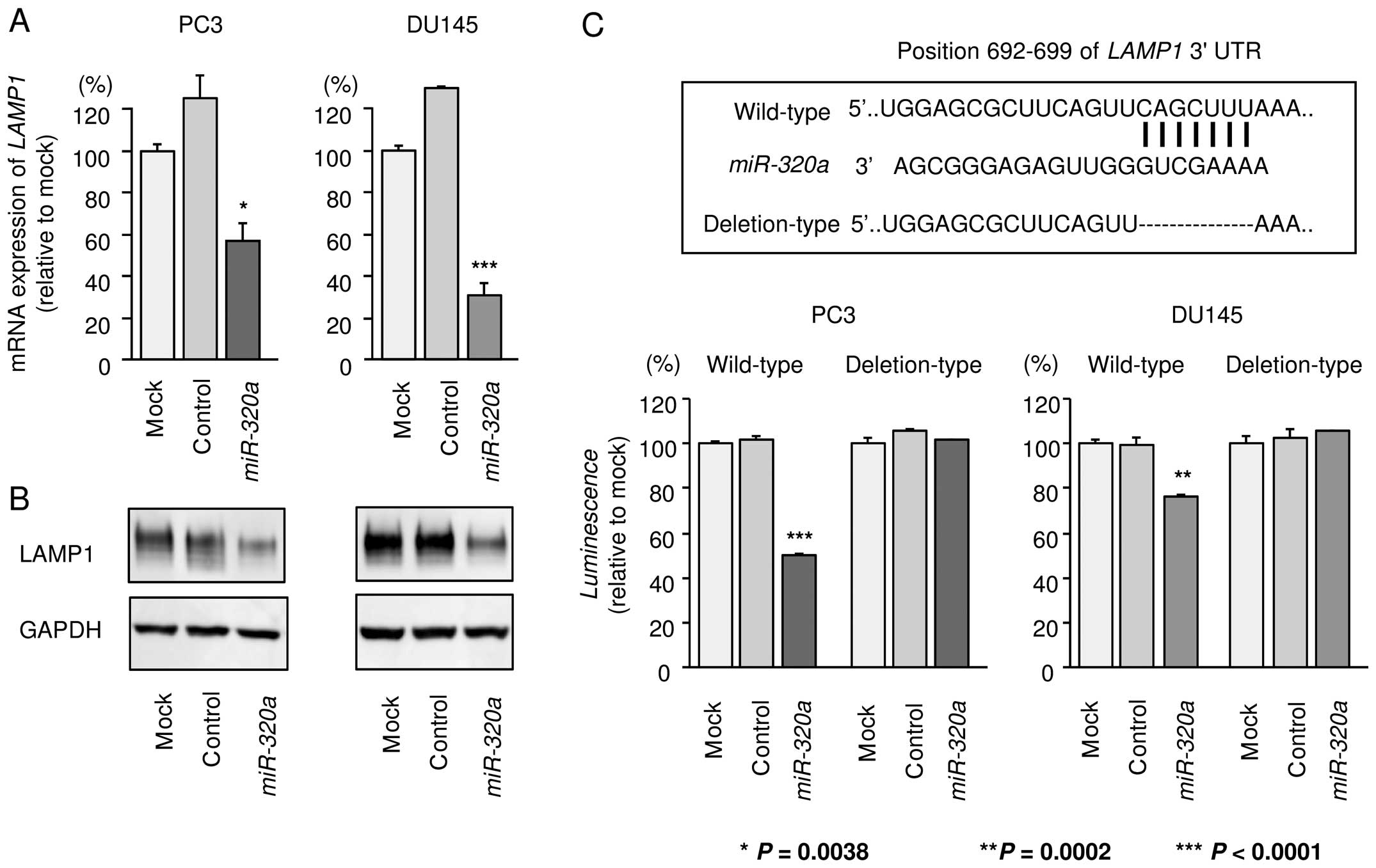
miR-320a directly regulated LAMP1 in PCa
cells
Next, we performed quantitative RT-PCR and western
blotting to analyze whether restoration of miR-320a in PC3
and DU145 cells reduced the expression of LAMP1. As shown in
Fig. 6A and B, the expression
levels of LAMP1 were significantly repressed by miR-320a
transfection in comparison with those in mock- or
miR-control-transfected cells.
Next, to determine whether LAMP1 mRNA was
directly regulated by miR-320a, we performed luciferase
reporter assays. We used vectors encoding either the partial
wild-type sequence of the 3′-UTR of LAMP1, including the
predicted miR-320a target site (position 692–699 of the
LAMP1 3′-UTR), or deletion vectors lacking this position. We
found that the luminescence intensities were significantly reduced
by transfection with miR-320a and the vector carrying the
wild-type 3′-UTR of LAMP1, whereas transfection with the
deletion vector blocked the decrease of luminescence in PC3 and
DU145 cells. These data suggested that miR-320a bound
directly to this site in the 3′-UTR of LAMP1 (Fig. 6C).
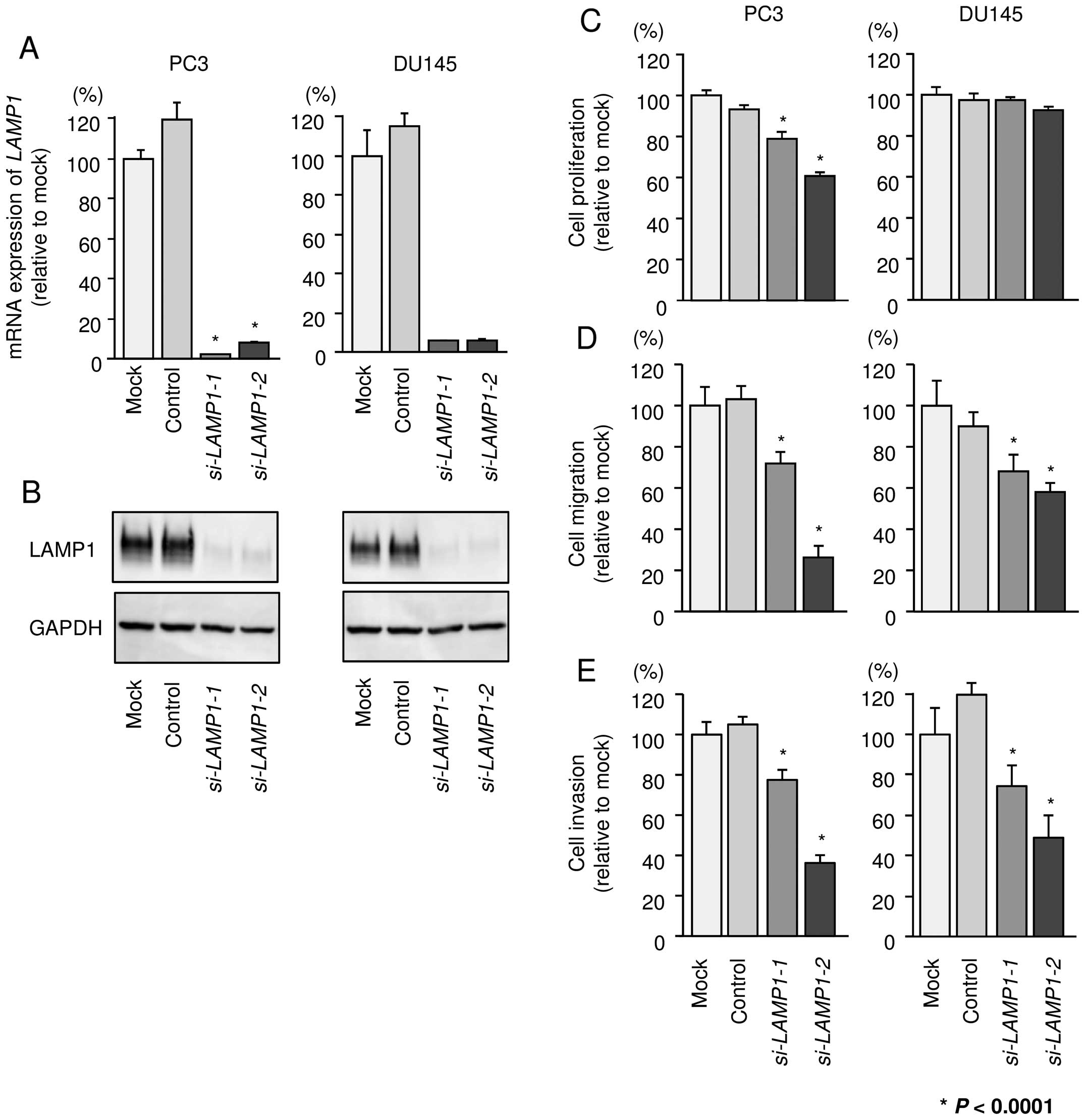
Effects of silencing LAMP1 on cell
proliferation, migration, and invasion in PCa cell lines
To investigate the functional role of LAMP1, we
performed loss-of-function studies using si-LAMP1
transfectants. First, we evaluated the knockdown efficiency of
si-LAMP1 transfection in PC3 and DU145 cells. Real-time PCR
and western blotting indicated that two siRNAs (si-LAMP1-1
and si-LAMP1-2) could effectively reduce the expression of
LAMP1 in PC3 and DU145 cells (Fig. 7A
and B).
Using these two siRNAs, we carried out functional
analyses. XTT assays demonstrated that cell proliferation was
inhibited only in PC3 cells by si-LAMP1 transfection
(Fig. 7C). Cell migration and
invasion activities were significantly inhibited in si-LAMP1
transfectants in both PC3 and DU145 in comparison with mock or
negative control transfectants (Fig.
7D and E).
Downstream genes affected by silencing of
LAMP1 in PCa cells
To further investigate which genes are modulated by
LAMP1 signaling, we performed genome-wide gene expression
analysis using si-LAMP1 in PC3 cells. A SurePrint G3 Human
GE 60K v3 microarray was used for genome-wide expression analysis.
The raw data were submitted to the GEO database (accession no.
GSE77790). In this study, we selected significantly downregulated
genes by si-LAMP1-1 or si-LAMP1-2 transfection
[Log2(si-LAMP1/mock) <-2.0]. LAMP1 was
the most significantly downregulated gene, indicating the
reliability of this array analysis. Genes significantly
downregulated by silencing of LAMP1 are shown in Table IV.
| Table IVGenes downregulated by silencing of
LAMP1 in PC3 cells. |
Table IV
Genes downregulated by silencing of
LAMP1 in PC3 cells.
| Gene symbol | Gene name |
Log2(si-LAMP1/mock) |
|---|
| LAMP1 |
Lysosomal-associated membrane protein
1 | −5.447 |
| ANLN | Anillin, actin
binding protein | −2.699 |
| KIF2C | Kinesin family
member 2C | −2.610 |
| KIF14 | Kinesin family
member 14 | −2.595 |
| AXL | AXL receptor
tyrosine kinase | −2.594 |
| CCNA2 | Cyclin A2 | −2.581 |
| SPC25 | SPC25, NDC80
kinetochore complex component | −2.543 |
| NEK2 | NIMA-related kinase
2 | −2.507 |
| ESCO2 | Establishment of
sister chromatid cohesion N-acetyltransferase 2 | −2.478 |
| MYL10 | Myosin, light chain
10, regulatory | −2.464 |
| HMGB2 | High mobility group
box 2 | −2.440 |
|
HIST2H3A | Histone cluster 2,
H3a | −2.432 |
| MZT1 | Mitotic spindle
organizing protein 1 | −2.416 |
| CCNE2 | Cyclin E2 | −2.404 |
| NUF2 | NUF2, NDC80
kinetochore complex component | −2.403 |
| CENPF | Centromere protein
F, 350/400 kDa | −2.378 |
| RRM2 | Ribonucleotide
reductase M2 | −2.377 |
| CDK1 | Cyclin-dependent
kinase 1 | −2.319 |
| SKA1 | Spindle and
kinetochore associated complex subunit 1 | −2.301 |
| PBK | PDZ binding
kinase | −2.293 |
| TOP2A | Topoisomerase (DNA)
IIα 170 kDa | −2.274 |
| CASC5 | Cancer
susceptibility candidate 5 | −2.233 |
|
lnc-AKR1C2-3 | lnc-AKR1C2-3:2 | −2.224 |
| NUSAP1 | Nucleolar and
spindle associated protein 1 | −2.212 |
| CEP55 | Centrosomal protein
55 kDa | −2.201 |
| UTS2 | Urotensin 2 | −2.188 |
| SKA3 | Spindle and
kinetochore associated complex subunit 3 | −2.185 |
|
HIST1H3B | Histone cluster 1,
H3b | −2.184 |
| G3BP2 | GTPase activating
protein (SH3 domain) binding protein 2 | −2.174 |
| SPC248 | SPC24, NDC80
kinetochore complex component | −2.170 |
| CENPA | Centromere protein
A | −2.135 |
| GNB4 | Guanine nucleotide
binding protein (G protein), β polypeptide 4 | −2.131 |
| MKI67 | Marker of
proliferation Ki-67 | −2.119 |
| TK1 | Thymidine kinase 1,
soluble | −2.118 |
| MKI67 | Marker of
proliferation Ki-67 | −2.115 |
| GLUD2 | Glutamate
dehydrogenase 2 | −2.102 |
| TYMS | Thymidylate
synthetase | −2.099 |
| E2F8 | E2F transcription
factor 8 | −2.087 |
|
LOC255187 | Uncharacterized
LOC255187 | −2.076 |
| KIF4A | Kinesin family
member 4A | −2.073 |
| CREG2 | Cellular repressor
of E1A-stimulated genes 2 | −2.073 |
| FAM83D | Family with
sequence similarity 83, member D | −2.068 |
|
HIST1H1D | Histone cluster 1,
H1d | −2.063 |
| DTL | Denticleless E3
ubiquitin protein ligase homolog (Drosophila) | −2.062 |
| FAM72D | Family with
sequence similarity 72, member D | −2.054 |
| KIF15 | Kinesin family
member 15 | −2.042 |
| ANXA1 | Annexin A1 | −2.022 |
| FAM72A | Family with
sequence similarity 72, member A | −2.020 |
| NMU | Neuromedin U | −2.019 |
| BIRC5 | Baculoviral IAP
repeat containing 5 | −2.019 |
| BUB1 | BUB1 mitotic
checkpoint serine/threonine kinase | −2.016 |
| E2F2 | E2F transcription
factor 2 | −2.015 |
| RAB31 | RAB31, member RAS
oncogene family | −2.010 |
Discussion
Metastatic PCa initially responds to
androgen-deprivation therapy; however, the cancer cells gradually
acquire resistance to first-line androgen-deprivation therapy and
consequently progress to CRPC. There are no effective treatments
for preventing progression of metastasis in patients with CRPC, and
this condition significantly affects the survival rates of men with
advanced PCa (2,19). Therefore, developing a deeper
understanding of the molecular mechanisms of metastatic pathways in
advanced PCa will facilitate the development of novel treatment
options for the disease. Recently, many studies have shown that
aberrantly expressed miRNAs disrupt normally regulated RNA networks
in cancer cells, and these events trigger cancer development,
progression, and metastasis (4,10).
Based on this background, we have identified tumor-suppressive
miRNAs that specifically block cancer cell migration and invasion
in PCa cells (9,11–18,20).
Our recent studies indicated that the
miR-221/miR-222 cluster is significantly downregulated in
PCa and CRPC specimens and suppresses cancer cell aggressiveness by
targeting Ecm29, a scaffold protein that links the 26S
proteasome to motor proteins (17). Moreover, we showed that
tumor-suppressive miR-223 inhibits cancer cell migration and
invasion by targeting ITGA3 and ITGB1 (18). Interestingly, knockdown of
ITGB1 significantly inhibits downstream oncogenic signaling
pathways, including the focal adhesion kinase (FAK), AKT, and
extracellular signal-regulated kinase (ERK) pathways (18). Thus, suppression of ITGA3/ITGB1
signaling in PCa cells may have applications in the development of
novel therapies for PCa.
In this study, we focused on miR-320a because
our miRNA signatures of hormone-naïve PCa and CRPC showed that
miR-320a was significantly downregulated in cancer tissues
(9,17). We validated the low expression of
miR-320a in hormone-naïve PCa and CRPC clinical specimens.
Restoration of miR-320a significantly inhibited cancer cell
migration and invasion in both PC3 and DU145 cell lines, suggesting
that miR-320a acted as a tumor-suppressive miRNA in PCa
cells.
The anticancer effects of miR-320a have been
reported in PCa and various other types of cancers (21–23).
For example, overexpression of miR-320 inhibits β-catenin
expression and the expression of cancer stem cell markers, such as
CD44, Oct-4, and CD133, in PCa cells (21). β-catenin is a multi-functional
protein that regulates cell adhesion and functions as a
transcriptional coactivator by interacting with several
transcriptional factors (21,22,24).
Accumulating evidence has shown that β-catenin colocalizes with AR,
and these events are more frequently observed in CRPC than in
hormone-naïve PCa (25).
Therefore, β-catenin/AR signaling may be potential therapeutic
target for the management of CRPC. Another study demonstrated that
miR-320a expression is reduced in primary salivary adenoid
cystic carcinoma (SACC) with metastasis compared with that in SACC
without metastasis (26).
Overexpression of miR-320a inhibits the adhesion, invasion,
and migration of SACC cells through regulation of ITGB3
(26). Chronic myeloid leukemia
(CML) is a myeloproliferative disease involving the BCR/ABL fusion
protein. Interestingly, a recent study showed that miR-320a
expression is reduced in K562 cells and CML cancer stem cells.
Additionally, restoration of miR-320a inhibits K562 cell
migration, invasion, and proliferation and promotes apoptosis by
targeting the BCR/ABL oncogene (27). These studies indicated that
continuous analysis of tumor-suppressive miR-320a-regulated
oncogenic pathways may provide insights into novel RNA networks in
cancer cells.
To better understand PCa progression and metastasis,
we identified miR-320a target genes using a combination of
in silico and genome-wide gene expression analyses. Recent
miRNA studies in our laboratory have utilized this strategy to
identify novel molecular targets and pathways regulated by
tumor-suppressive miRNAs in several cancers, including PCa
(17,18). A total of 14 putative target genes
of miR-320a were identified in this study. Among these
genes, we focused on the LAMP1 gene because few reports have
described the role of this target in PCa. Here, we found that
downregulation of LAMP1 inhibited the migration and invasion
of cancer cells. Additionally, overexpression of LAMP1 was observed
in hormone-naïve PCa and CRPC clinical specimens.
LAMP1 is a heavily glycosylated lysosomal membrane
protein that is expressed mainly in the endosome-lysosomal membrane
of cells (28,29). High expression of LAMP1 on the
plasma membrane has been observed in several metastatic cell lines,
including human melanoma, colon carcinoma, fibrosarcoma, and
myelomonocytic leukemia cell lines (30–33).
Previous studies have indicated that cell surface glycosylation in
cancer cells differs from that in normal cells (30). Additionally, the expression of β1,6
branched N-oligosaccharides on the cell surface in several human
cancers has been shown to be correlated with the malignant
potential of the cells (34).
LAMP1 acts as a major carrier of β1,6 branched N-oligosaccharides
in B16 melanoma cells (31). A
recent study also showed that LAMP1 expression on the cell surface
enhances lung metastasis by providing ligands for galectin-3
(Gal-3), a member of the multifunctional β-galactoside binding
lectin family (31). Gal-3 has
high affinity for β-1,6-N-acetylglucosamine branched glycans. This
interaction mediates binding of the lectin to many glycoproteins in
the cell membrane, such as cadherin, integrins, and growth factor
receptors, and these events may enhance and promote cancer cell
growth and metastasis (35).
Several studies have demonstrated that Gal-3 plays pivotal roles in
cancer progression and aggressiveness by regulating cell
proliferation, apoptosis, invasion, and metastasis (35,36).
Moreover, overexpression of Gal-3 promotes PCa cell progression,
and its heterogeneous expression may be associated with different
PCa subtypes (37). These studies
indicate that cell surface expression of LAMP1 may affect Gal-3
activation and mediate multiple stages of cancer progression and
metastasis.
In this study, we identified LAMP1-mediated cancer
pathways by using genome-wide gene expression analysis of
si-LAMP1-transfected cells. Our data showed that several
genes known to contribute to cancer cell aggressiveness, including
ANLN, AXL, CCNA2 and CENPF (16,38–40),
were involved in LAMP1 downstream pathways. The identification of
these novel molecular targets mediated by the miR-320a/LAMP1
axis may lead to a better understanding of PCa progression and
metastasis.
In conclusion, downregulation of miR-320a
was observed in PCa and CRPC clinical specimens, and this miRNA was
shown to function as a tumor-suppressive miRNA in PCa cells.
LAMP1 was directly regulated by tumor-suppressive
miR-320a and contributed to cancer cell aggressiveness. The
identification of novel molecular pathways regulated by the
miR-320a/LAMP1 axis may lead to a better
understanding of PCa progression and metastasis.
Acknowledgements
This study was supported by the KAKENHI, grant
numbers (C) 15K10801, (C) 15K20070, (C) 15K20071, and (B)
25293333.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sridhar SS, Freedland SJ, Gleave ME,
Higano C, Mulders P, Parker C, Sartor O and Saad F:
Castration-resistant prostate cancer: From new pathophysiology to
new treatment. Eur Urol. 65:289–299. 2014. View Article : Google Scholar
|
|
3
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hobert O: Gene regulation by transcription
factors and microRNAs. Science. 319:1785–1786. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
9
|
Fuse M, Kojima S, Enokida H, Chiyomaru T,
Yoshino H, Nohata N, Kinoshita T, Sakamoto S, Naya Y, Nakagawa M,
et al: Tumor suppressive microRNAs (miR-222 and miR-31) regulate
molecular pathways based on microRNA expression signature in
prostate cancer. J Hum Genet. 57:691–699. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goto Y, Kurozumi A, Enokida H, Ichikawa T
and Seki N: Functional significance of aberrantly expressed
microRNAs in prostate cancer. Int J Urol. 22:242–252. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Goto Y, Kojima S, Nishikawa R, Enokida H,
Chiyomaru T, Kinoshita T, Nakagawa M, Naya Y, Ichikawa T and Seki
N: The microRNA-23b/27b/24-1 cluster is a disease progression
marker and tumor suppressor in prostate cancer. Oncotarget.
5:7748–7759. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nishikawa R, Goto Y, Kojima S, Enokida H,
Chiyomaru T, Kinoshita T, Sakamoto S, Fuse M, Nakagawa M, Naya Y,
et al: Tumor-suppressive microRNA-29s inhibit cancer cell migration
and invasion via targeting LAMC1 in prostate cancer. Int J Oncol.
45:401–410. 2014.PubMed/NCBI
|
|
13
|
Nishikawa R, Goto Y, Sakamoto S, Chiyomaru
T, Enokida H, Kojima S, Kinoshita T, Yamamoto N, Nakagawa M, Naya
Y, et al: Tumor-suppressive microRNA-218 inhibits cancer cell
migration and invasion via targeting of LASP1 in prostate cancer.
Cancer Sci. 105:802–811. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kato M, Goto Y, Matsushita R, Kurozumi A,
Fukumoto I, Nishikawa R, Sakamoto S, Enokida H, Nakagawa M,
Ichikawa T, et al: MicroRNA-26a/b directly regulate La-related
protein 1 and inhibit cancer cell invasion in prostate cancer. Int
J Oncol. 47:710–718. 2015.PubMed/NCBI
|
|
15
|
Kojima S, Chiyomaru T, Kawakami K, Yoshino
H, Enokida H, Nohata N, Fuse M, Ichikawa T, Naya Y, Nakagawa M, et
al: Tumour suppressors miR-1 and miR-133a target the oncogenic
function of purine nucleoside phosphorylase (PNP) in prostate
cancer. Br J Cancer. 106:405–413. 2012. View Article : Google Scholar :
|
|
16
|
Nishikawa R, Goto Y, Kurozumi A,
Matsushita R, Enokida H, Kojima S, Naya Y, Nakagawa M, Ichikawa T
and Seki N: MicroRNA-205 inhibits cancer cell migration and
invasion via modulation of centromere protein F regulating pathways
in prostate cancer. Int J Urol. 22:867–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goto Y, Kojima S, Nishikawa R, Kurozumi A,
Kato M, Enokida H, Matsushita R, Yamazaki K, Ishida Y, Nakagawa M,
et al: MicroRNA expression signature of castration-resistant
prostate cancer: The microRNA-221/222 cluster functions as a tumour
suppressor and disease progression marker. Br J Cancer.
113:1055–1065. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kurozumi A, Goto Y, Matsushita R, Fukumoto
I, Kato M, Nishikawa R, Sakamoto S, Enokida H, Nakagawa M, Ichikawa
T, et al: Tumor-suppressive microRNA-223 inhibits cancer cell
migration and invasion by targeting ITGA3/ITGB1 signaling in
prostate cancer. Cancer Sci. 107:84–94. 2016. View Article : Google Scholar
|
|
19
|
Sturge J, Caley MP and Waxman J: Bone
metastasis in prostate cancer: Emerging therapeutic strategies. Nat
Rev Clin Oncol. 8:357–368. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Goto Y, Nishikawa R, Kojima S, Chiyomaru
T, Enokida H, Inoguchi S, Kinoshita T, Fuse M, Sakamoto S, Nakagawa
M, et al: Tumour-suppressive microRNA-224 inhibits cancer cell
migration and invasion via targeting oncogenic TPD52 in prostate
cancer. FEBS Lett. 588:1973–1982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hsieh IS, Chang KC, Tsai YT, Ke JY, Lu PJ,
Lee KH, Yeh SD, Hong TM and Chen YL: MicroRNA-320 suppresses the
stem cell-like characteristics of prostate cancer cells by
downregulating the Wnt/beta-catenin signaling pathway.
Carcinogenesis. 34:530–538. 2013. View Article : Google Scholar
|
|
22
|
Sun JY, Huang Y, Li JP, Zhang X, Wang L,
Meng YL, Yan B, Bian YQ, Zhao J, Wang WZ, et al: MicroRNA-320a
suppresses human colon cancer cell proliferation by directly
targeting β-catenin. Biochem Biophys Res Commun. 420:787–792. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lü M, Ding K, Zhang G, Yin M, Yao G, Tian
H, Lian J, Liu L, Liang M, Zhu T, et al: MicroRNA-320a sensitizes
tamoxifen-resistant breast cancer cells to tamoxifen by targeting
ARPP-19 and ERRγ. Sci Rep. 5:87352015. View Article : Google Scholar
|
|
24
|
Hoffmans R, Städeli R and Basler K:
Pygopus and legless provide essential transcriptional coactivator
functions to armadillo/beta-catenin. Curr Biol. 15:1207–1211. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yokoyama NN, Shao S, Hoang BH, Mercola D
and Zi X: Wnt signaling in castration-resistant prostate cancer:
Implications for therapy. Am J Clin Exp Urol. 2:27–44.
2014.PubMed/NCBI
|
|
26
|
Sun L, Liu B, Lin Z, Yao Y, Chen Y, Li Y,
Chen J, Yu D, Tang Z, Wang B, et al: MiR-320a acts as a prognostic
factor and Inhibits metastasis of salivary adenoid cystic carcinoma
by targeting ITGB3. Mol Cancer. 14:962015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xishan Z, Ziying L, Jing D and Gang L:
MicroRNA-320a acts as a tumor suppressor by targeting BCR/ABL
oncogene in chronic myeloid leukemia. Sci Rep. 5:124602015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jensen SS, Aaberg-Jessen C, Christensen KG
and Kristensen B: Expression of the lysosomal-associated membrane
protein-1 (LAMP-1) in astrocytomas. Int J Clin Exp Pathol.
6:1294–1305. 2013.PubMed/NCBI
|
|
29
|
Parkinson-Lawrence EJ, Dean CJ, Chang M,
Hopwood JJ, Meikle PJ and Brooks DA: Immunochemical analysis of
CD107a (LAMP-1). Cell Immunol. 236:161–166. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Furuta K, Ikeda M, Nakayama Y, Nakamura K,
Tanaka M, Hamasaki N, Himeno M, Hamilton SR and August JT:
Expression of lysosome-associated membrane proteins in human
colorectal neoplasms and inflammatory diseases. Am J Pathol.
159:449–455. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Agarwal AK, Gude RP and Kalraiya RD:
Regulation of melanoma metastasis to lungs by cell surface lysosome
associated membrane protein-1 (LAMP1) via galectin-3. Biochem
Biophys Res Commun. 449:332–337. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sarafian V, Jadot M, Foidart JM, Letesson
JJ, Vanden Brûle F, Castronovo V, Wattiaux R and Coninck SW:
Expression of Lamp-1 and Lamp-2 and their interactions with
galectin-3 in human tumor cells. Int J Cancer. 75:105–111. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mane SM, Marzella L, Bainton DF, Holt VK,
Cha Y, Hildreth JE and August JT: Purification and characterization
of human lysosomal membrane glycoproteins. Arch Biochem Biophys.
268:360–378. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Krishnan V, Bane SM, Kawle PD, Naresh KN
and Kalraiya RD: Altered melanoma cell surface glycosylation
mediates organ specific adhesion and metastasis via lectin
receptors on the lung vascular endothelium. Clin Exp Metastasis.
22:11–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fortuna-Costa A, Gomes AM, Kozlowski EO,
Stelling MP and Pavão MS: Extracellular galectin-3 in tumor
progression and metastasis. Front Oncol. 4:1382014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ahmed H and AlSadek DM: Galectin-3 as a
potential target to prevent cancer metastasis. Clin Med Insights
Oncol. 9:113–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Y, Nangia-Makker P, Tait L, Balan V,
Hogan V, Pienta KJ and Raz A: Regulation of prostate cancer
progression by galectin-3. Am J Pathol. 174:1515–1523. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dachineni R, Ai G, Kumar DR, Sadhu SS,
Tummala H and Bhat GJ: Cyclin A2 and CDK2 as novel targets of
aspirin and salicylic acid: A potential role in cancer prevention.
Mol Cancer Res. 14:241–252. 2016. View Article : Google Scholar
|
|
39
|
Scaltriti M, Elkabets M and Baselga J:
Molecular pathways: AXL, a membrane receptor mediator of resistance
to therapy. Clin Cancer Res. 22:1313–1317. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhou W, Wang Z, Shen N, Pi W, Jiang W,
Huang J, Hu Y, Li X and Sun L: Knockdown of ANLN by lentivirus
inhibits cell growth and migration in human breast cancer. Mol Cell
Biochem. 398:11–19. 2015. View Article : Google Scholar
|