Introduction
Neuroblastoma (NB) is a malignant paediatric tumor
of the sympathetic nervous system that presents as a highly
heterogeneous disease, ranging from spontaneous regression to high
risk of fatality (1–3). Although the overall survival of
neuroblastoma has increased greatly with the advances of diagnostic
methods and therapeutic treatment over the recent years, the cure
rate and life quality for high-risk neuroblastoma have only
improved marginally when given aggressive conventional treatment
(4,5). Due to issues like early osseous and
(or) bone marrow metastasis and minimal residual disease (MRD), the
effect of surgical operation in advanced neuroblastoma patients is
limited. Therefore, combination chemotherapy, especially the
molecular targeted agent-based combination, remains the dominant
strategy in the field of neuroblastoma treatment and research.
Numerous pieces of evidence show that activation of
PI3K/Akt/mTOR pathway correlates with poor prognosis and
chemotherapy resistance in neuroblastoma (6–8).
NVP-BEZ235 is a potent novel dual PI3K and mTOR kinase inhibitor
which has shown great inhibitory efficacy on colorectal, breast,
non-small cell lung carcinoma, renal cancer, and sarcoma (9–13).
This compound as a single agent has also been investigated in
neuroblastoma, where it was shown ineffective in MYCN-non amplified
neuroblastoma, which led us to speculate that NVP-BEZ235 might be
better suited as a part of combination therapy (14). Oridonin is an ent-kaurane
diterpenoid extracted from the plant Rabdosia rubescens
which has been used as antimicrobial, anti-inflammation and
antitumor agent in traditional Chinese medicine for thousands of
years (15,16). As regards its antitumor activity,
previous studies have demonstrated that oridonin as a supplement
may potentiate the therapeutic effects of anticancer drugs
gemcitabine and imatinib in the treatment of pancreatic cancer and
Ph+ acute lymphoblastic leukemia separately (17,18).
However, it is currently unknown whether oridonin can strengthen
the effects of NVP-BEZ235 in neuroblastoma treatment.
In this study, we investigated the effect of
combined treatment with NVP-BEZ235 and oridonin on the growth of
human neuroblastoma cells in culture and in a preclinical mouse
model. The data indicated that oridonin significantly enhanced the
inhibitory effect of NVP-BEZ235 on neuroblastoma, leading to
synergistic cell apoptosis and inhibition of NB xenograft tumor
growth.
Materials and methods
Reagent
NVP-BEZ235 was purchased from Novartis (East
Hanover, NJ, USA) and dissolved in dimethylsulfoxide (DMSO) to a
stock concentration of 1 mmol/l. Oridonin (purity >98%) was
purchased from Chengdu Must Bio-Technology Co., Ltd. (Chengdu,
Sichuan, China) and dissolved in DMSO to a stock concentration of
10 mmol/l. The stock solutions were wrapped in foil and maintained
at −20°C.
Cell lines and cell culture
Neuroblastoma cell lines (SHSY-5Y, SK-N-MC) were
obtained from the Shanghai Institutes for Biological Sciences
(Shanghai, China) and were cultured in monolayer culture in
RPMI-1640 medium (Gibco-BRL, Long Island, NY, USA) supplemented
with 10% fetal bovine serum (FBS) (Gibco-BRL) and 1%
penicillin-streptomycin (Gibco-BRL), under standard culture
conditions (37°C and 5% CO2). Cells in the logarithmic
phase of growth were used in all experiments.
Cell Counting Kit-8 assay
The cell proliferation was assessed by Cell Counting
Kit-8 reagent (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). SHSY-5Y and SK-N-MC cells were seeded at a
density of 5×103 cells/200 μl/well in 96-well culture
plates and allowed to settle overnight. Then the wells were treated
with NVP-BEZ235 and/or oridonin for 12 or 24 h. Finally, the
cultured wells were treated with 20 μl/well of CCK-8 solution and
incubated for 2 h and the optical density of wells was measured at
450 nm using a microplate reader (Bio-Rad, Hercules, CA, USA).
Flow cytometric analysis of cell cycle
and apoptosis
Neuroblastoma cells were seeded onto 6-well plate at
a density of 1×105 per well. After 24-h incubation, they
were treated with NVP-BEZ235 and/or oridonin for 24 h and
collected. For cell cycle analysis, cells were washed and fixed
with ice-cold 75% (v/v) ethanol at −20°C for 2 h, then after
washing twice with PBS, the cells were suspended with 500 μl PI
solution (PI 50 μg/ml and RNase A 100 μg/ml). The samples were
analyzed by FACScan flow cytometer (BD, Franklin Lakes, NJ, USA).
FITC Annexin V Apoptosis Detection Kit I (BD Pharmingen, San Jose,
CA, USA) was used to quantify drug-induced cell apoptosis following
the manufacturer's guidelines.
Acridine orange/ethidium bromide (AO/EB)
fluorescence
Neuroblastoma cells were cultivated in a 24-well
plate exposed to NVP-BEZ235 and/or oridonin for 24 h in a
humidified incubator (37°C, 5% CO2). After removing the
culture solution and washing with PBS, the cells were stained with
200 μl mixture of (100 μg/ml) acridine orange (AO) and (100 μg/ml)
ethidium bromide (EB) (Sigma, St. Louis, MO, USA) with the 1:1 AO
to EB, and then incubated the plate for 3 min in the incubator. The
cells were visualized by a fluorescence microscope (Nikon Eclipse
Ti, Japan).
Lentiviral transduction
shRNA pLKO.1 lentiviral constructs were obtained
from Hanbio (Shanghai, China). Sequences are as follows: Beclin-1
#1, 5′-CCC GTG GAA TGG AAT GAG ATT-3′; Beclin-1 #2, 5′-GCT TGG GTG
TCC TCA CAA TTT-3′. Scrambled, 5′-GTG GAC TCT TGA AAG TAC TAT-3′.
The pLKO.1 vectors and package plasmids (pLP1-gag/pol, pLP2-Rev,
pLP-VSVG) (Life Technologies, Carlsbad, CA, USA) were
co-transfected into packaging 293T cells using Lipofectamine-3000
(Life Technologies). After 48 h, the viral supernatants were
collected and used to infect SH-SY5Y cells. Stable clones
expressing the shRNAs were obtained via 5 μg/ml puromycin
dihydrochloride (Santa Cruz Biotech Inc.; sc-205821) selection.
Western blot analysis
Cells were lysed with SDS lysis buffer containing
phosphatase and protease inhibitor cocktail (Roche, Mannheim,
Germany) and the protein concentration was assayed with Bio-Rad
Protein Assay kit (Bio-Rad). Cell lysates with equal protein
content were separated by SDS-polyacrylamide gel electrophoresis
and transferred to PVDF membranes (Bio-Rad). After 3 washes, the
membranes were blocked with 5% non-fat milk in Tris-buffered saline
containing 0.1% Tween-20 (TBST, pH 7.4) for 1 h at room
temperature, followed by incubation overnight at 4°C with primary
antibodies to cleaved caspase-3, cleaved PARP, cyclin D1, cyclin
E1, p62, LC3B and Beclin-1 (all from Cell Signaling Technology, MA,
USA). After washing 3 times, the membranes were incubated for 1 h
at room temperature with species-specific HRP-conjugated secondary
antibodies (Cell Signaling Technology). Immunoreactive bands were
visualized using Immobilon Western chemiluminescent HRP substrate
(Millipore Co., Billerica, MA, USA). Each experiment was performed
at least 3 times independently. As a loading control, the GAPDH
contents in the samples were also immunoblotted using GAPDH
horseradish peroxidase-conjugated antibody (Cell Signaling
Technology).
Neuroblastoma xenografts and
treatments
Animal experiments were approved by the
Institutional Animal Care and Use Committee (IACUC) of Shanghai
Jiao Tong University. Five- to six-week old female athymic (nu/nu)
mice were housed under pathogen-free conditions in micro-isolator
cages with laboratory chow and water ad libitum. SHSY-5Y
cells at 5×106 suspended in 100 μl PBS were injected
s.c. into the flank region of nude mice. When tumors were
measurable and reached an average volume of 100 mm3, the
mice were randomized into four groups for the following treatments:
vehicle control, NVP-BEZ235 (20 mg/kg/day, i.p.), oridonin (10
mg/kg/day; i.p.), and their combination. Tumor volumes were
measured using caliper measurements once every 5 days and
calculated with the formula V= π (length × width × height)/6. Mice
were sacrificed after 4 weeks of interventions and tumor tissues
were excised and fixed in 10% buffered formalin and embedded in
paraffin.
Histopathology and
immunohistochemistry
Paraffin-embedded blocks of all tumor samples were
cut at 4 μm and each sample was stained with hematoxylin and eosin
(H&E) for histologic analysis. For immunohistochemical
detection of Ki-67, sections were deparaffinized in xylene and
ethanol and pretreated with a citrate buffer solution (0.01 mol/l
citric acid and 0.01 mol/l sodium citrate, pH 6.0) in a microwave
oven at 750 W for two cycles of 10 min each. Endogenous peroxidase
activity was blocked by treatment with 3% hydrogen peroxide in
methanol for 10 min at room temperature. After a blocking step with
normal goat serum (Vector, Burlingame, CA, USA), the anti-Ki-67
antibodies (Cell Signaling Technology) were applied overnight in a
moist chamber at 4°C. The following day, tissue sections were
incubated with a secondary biotinylated anti-rabbit antibody and
with an avidin-biotin-peroxidase complex (Vector). The final
reaction product was revealed by exposure to 0.03% diaminobenzidine
(Sigma) and nuclei were counterstained with hematoxylin.
Appropriate negative controls for the immunostaining were prepared
by omitting the primary antibody step and substituting it with
nonimmune rabbit serum. All samples were analyzed by scoring
staining intensity.
TUNEL assay
To observe the degree of apoptosis, TUNEL assay was
performed as per the manufacturer's protocol (In Situ Cell Death
Detection kit, POD, Roche). All samples were analyzed by scoring
staining intensity.
Statistical analyses
Each sample was analyzed in triplicate, and
experiments were repeated three times. In all figures, error bars
are standard deviations. Statistical analyses were performed by
Microsoft Office Excel 2003 (Microsoft, Albuquerque, NM, USA) and
Statisca ver. 10 (StatSoft, Tulsa, OK, USA). Differences between
mean values were evaluated by the unpaired t-test. Differences were
considered statistically significant at P<0.05.
Results
NVP-BEZ235 inhibits proliferation but
does not induce apoptosis in NB
To determine the effect of the dual kinase inhibitor
NVP-BEZ235 on the proliferation of human neuroblastoma cells,
SHSY-5Y and SK-N-MC were treated with varying concentrations of
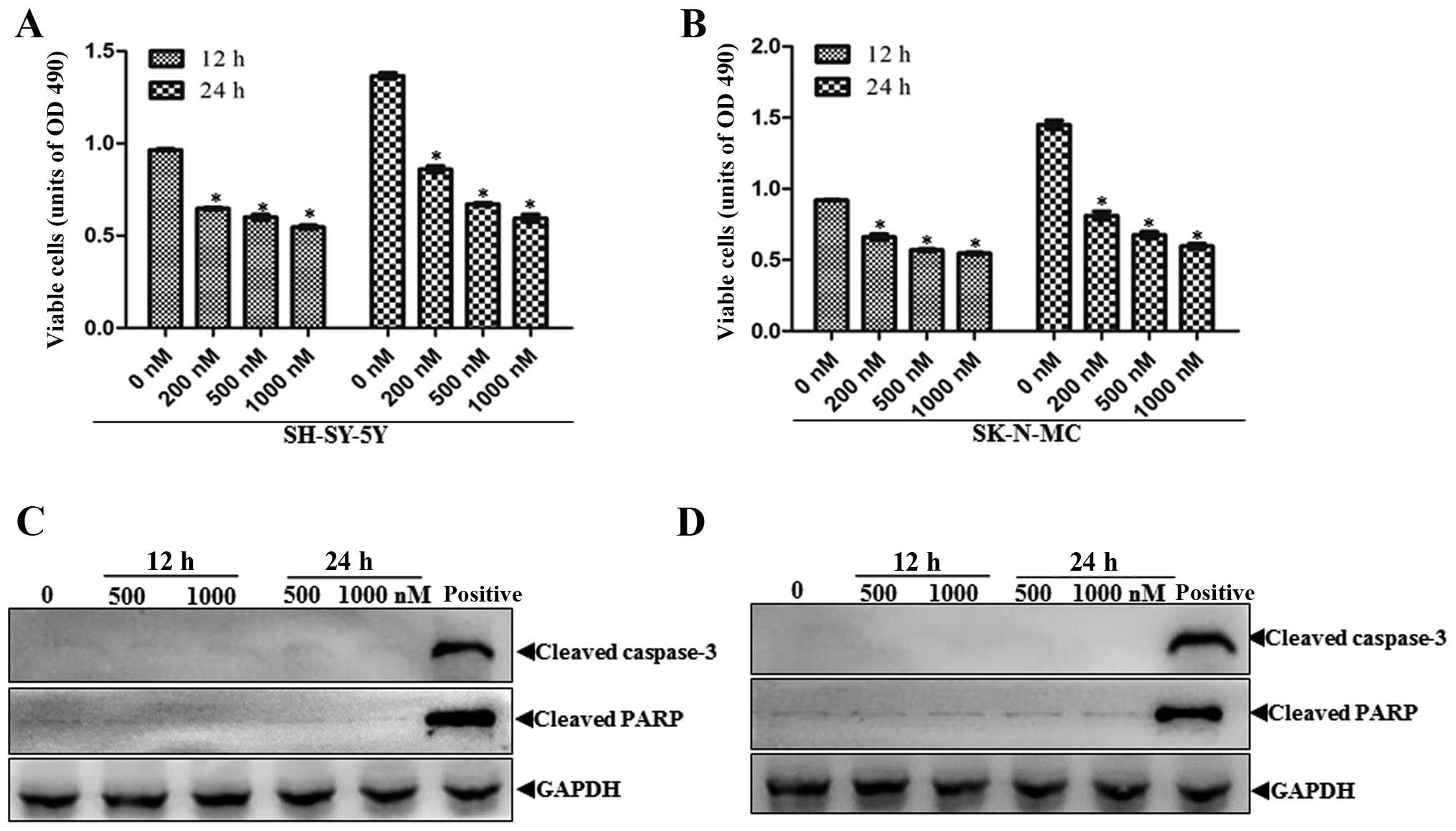
NVP-BEZ235 (0, 200, 500 and 1,000 nM) for 12 and 24 h and then cell
viability was measured by CCK-8 assay. As shown in Fig. 1A and B, NVP-BEZ235 causes a time-
and dose-dependent inhibition of cell proliferation in both SHSY-5Y
and SK-N-MC cells. Based on the above results, we further
investigated whether NVP-BEZ235-induced proliferation inhibition is
associated with apoptosis. We detected the apoptotic relative
proteins caspase-3 and PARP cleavage in NVP-BEZ235 treated
neuroblastoma cells. In contrast with the antiproliferative effects
of NVP-BEZ235, it exerted little toxic effect, as indicated by the
absence of cleaved caspase-3 and PARP in both treated cell lines
(Fig. 1C and D).
NVP-BEZ235 induces G0/G1 cell cycle
arrest in NB
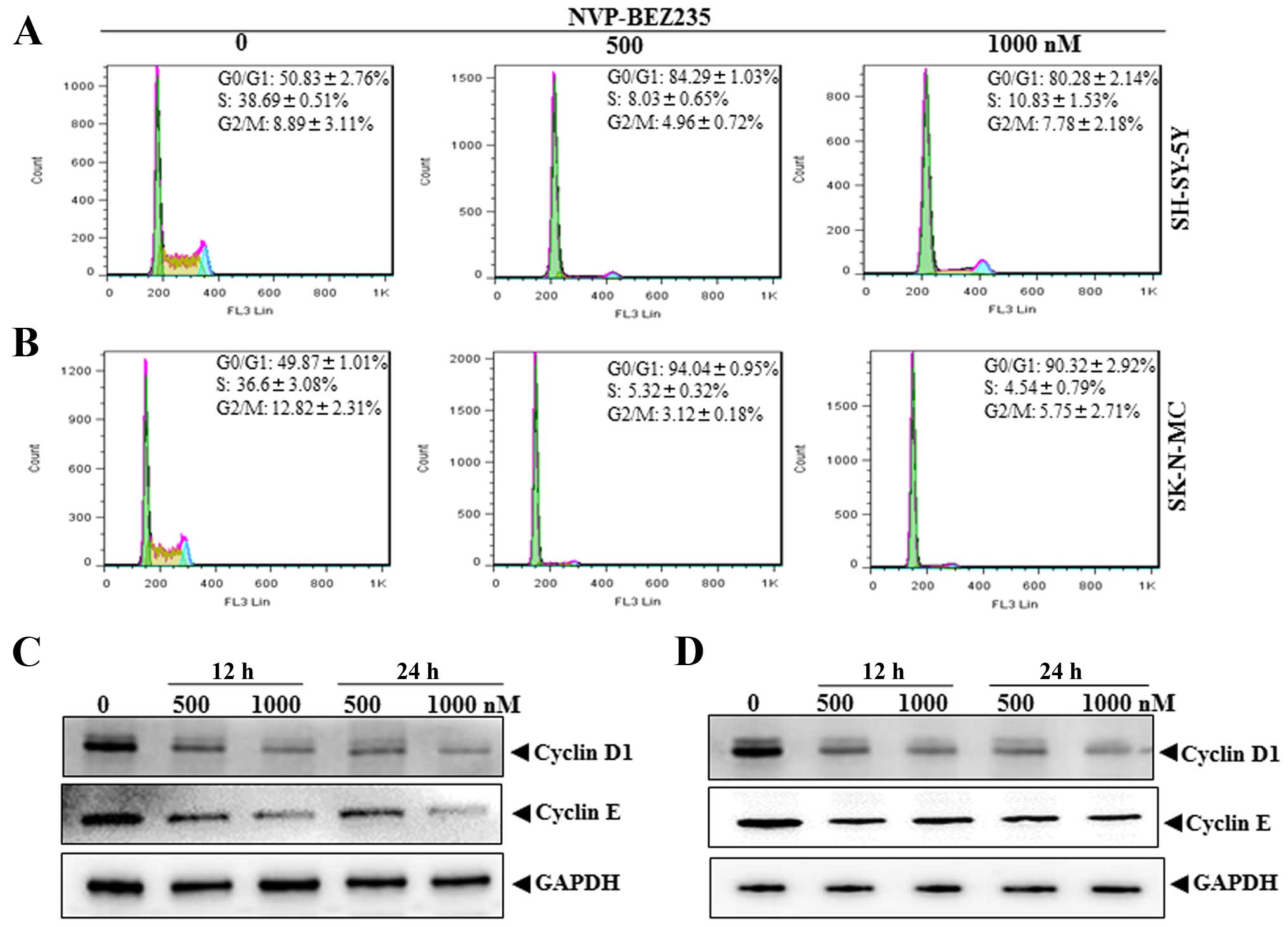
As apoptosis could not have accounted for the potent
inhibitory effect of NVP-BEZ235 on NB cell growth, we next analyzed
the effect of NVP-BEZ235 on cell cycle progression in neuroblastoma
cells by flow cytometry using PI staining of DNA content. Exposed
to 500 and 1,000 nM NVP-BEZ235 for 24 h, there was a significant
increase in the percentage of SHSY-5Y cells in the G0/G1 phase
(84.29±1.03% 500 nM, 80.28±2.14% 1,000 nM) compared with the
control group (50.83±2.76%) (Fig.
2A). The significant increase of G0/G1 phase cells was also
detected in SK-N-MC cells (Fig.
2B). To further confirm NVP-BEZ235-induced G1 cell cycle arrest
in neuroblastoma cells, we examined the expression of endogenous
cyclins 12 and 24 h after NVP-BEZ235 treatment at 500 and 1,000 nM
in SHSY-5Y and SK-N-MC cells. Cyclin D1 and cyclin E1 are
well-known G1-phase cyclins, governing the G1→S phase progression
and their inhibition results in G1 phase arrest. As shown in
Fig. 2C and D, NVP-BEZ235
treatment caused a remarkable reduction in cyclin D1 and cyclin E1
in both cell lines. Based on the above results, we concluded that
the dual PI3K/mTOR inhibitor NVP-BEZ235 inhibits NB cell
proliferation through the induction of cell cycle arrest but not
apoptosis.
Co-treatment with NVPBEZ235 and oridonin
induces enhanced antiproliferation and autophagy in NB
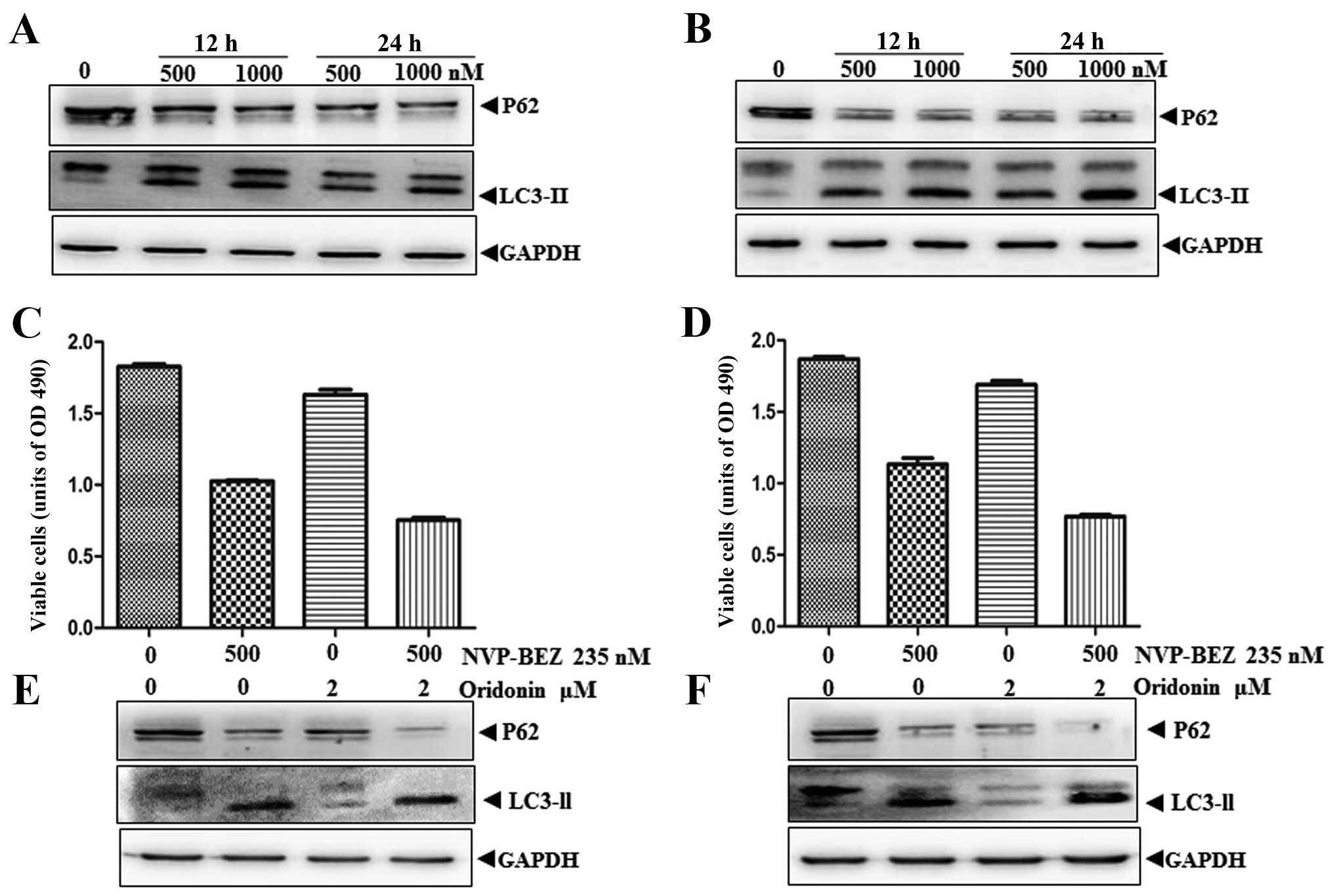
It is reported that enhanced autophagy induced by
combination treatment can lead to programmed cell death and
autophagic cell death has been proven to be effective in cases of
solid tumors (19,20). Given that NVP-BEZ235 alone can
activate autophagy in neuroblastoma cells (Fig. 3A and B), we selected another
autophagy related compound, oridonin, trying to combine with
NVP-BEZ235 to improve its therapeutic efficacy. We first
investigated the anti-proliferative effect of the combination
treatment. The results showed that cell growth in both cell lines
was markedly decreased following 24 h NVPBEZ235 and oridonin
co-treatment when compared with either single agent alone (Fig. 3C and D). Then, we examined the
autophagy activity of SHSY-5Y and SK-N-MC after the combination
treatment through immunoblot analysis. As shown in Fig. 3E and F, there was substantially
more LC3-II conversion after combination treatment compared with
NVP-BEZ235 or oridonin treatment alone in both cell lines. In
addition, co-treatment with NVP-BEZ235 and oridonin led to a
significant decrease in the levels of p62 in the tested cells. This
indicated that the combination of NVP-BEZ235 and oridonin can
synergistically induce enhanced autophagy.
Co-treatment with NVPBEZ235 and oridonin
induces apoptosis in NB
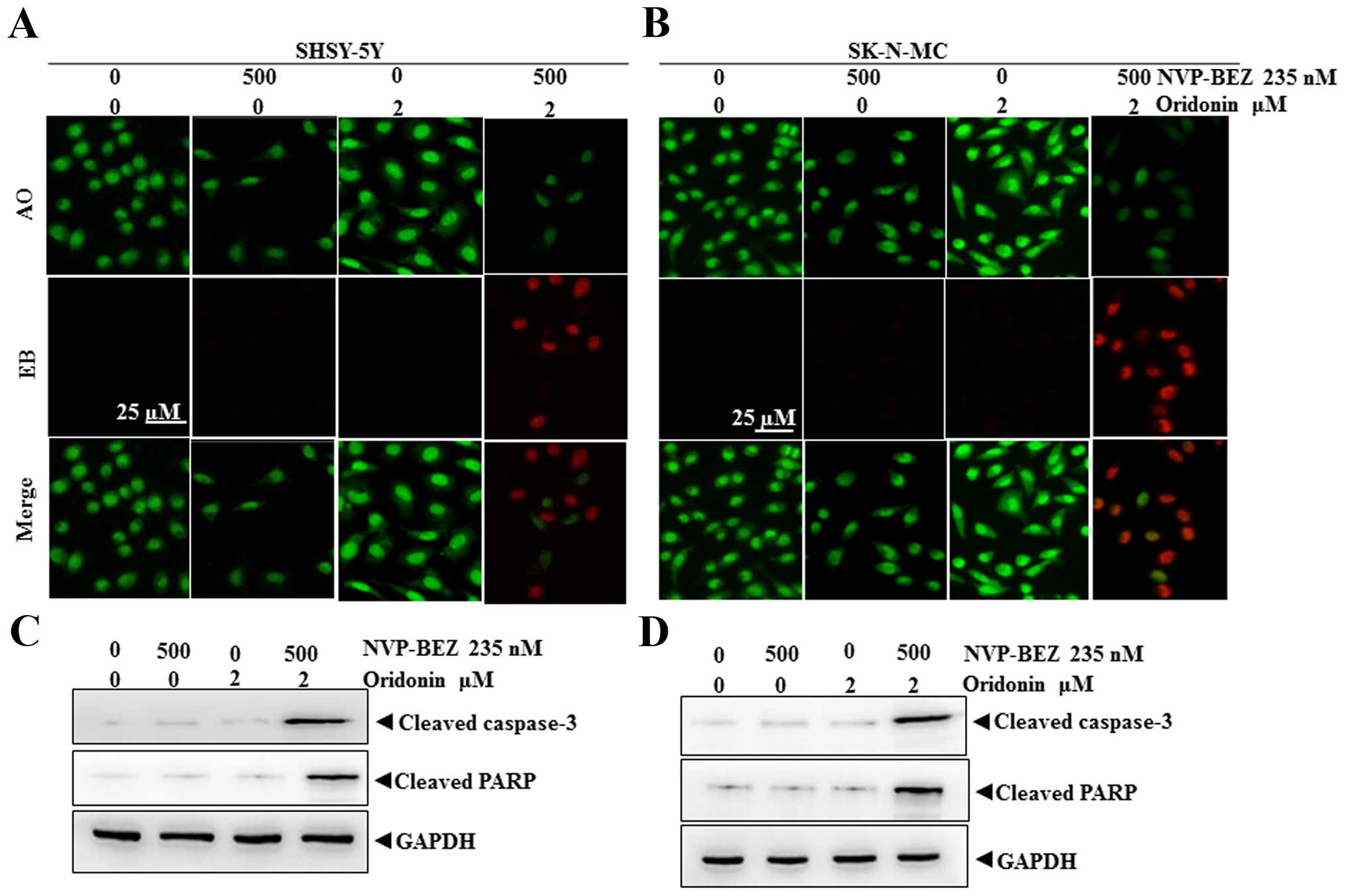
In order to examine whether the combination of
NVP-BEZ235 and oridonin could synergistically induce apoptosis of
NB, the treated neuroblastoma cells were stained with AO/EB. AO was
able to infiltrate into the viable cells, and the nuclei were
stained a bright green color. For the integrity of the cell plasma
membrane, EB was unable to infiltrate into the cells which remained
alive or were at early stage of apoptosis, while the late apoptotic
cells or dead cells had EB inside and the nuclei were stained a
bright red color. Fig. 4A and B
show various morphologies of NB cells stained with AO/EB. After 500
nM NVP-BEZ235 treated, the number of viable cells in both cell
lines was remarkably decreased in comparison with the control,
while the cells cultivated with 2 μM oridonin were similar to the
control cells. More notably, the percentage of apoptotic cells was
significantly increased in cells treated with the combination of
NVP-BEZ235 and oridonin compared with either single agent alone.
The results indicated the cell proliferation of NB was strongly
inhibited by NVP-BEZ235, but it had very slight cell cytotoxicity
compared to the combination groups which commit NBs to apoptosis.
For further confirming the results, we examined the cleavage of
caspase-3 and PARP in the treated cells. As shown in Fig. 4C and D, when SHSY-5Y and SK-N-MC
cells were treated with the combination of NVP-BEZ235 and oridonin
for 24 h, there was a significant increase of the cleaved form of
caspase-3 and PARP compared with either single agent alone,
suggesting that apoptotic pathway was activated in response to the
combination treatment.
Enhanced autophagy is essential for the
induction of apoptosis by the co-treatment with NVP-BEZ235 and
oridonin
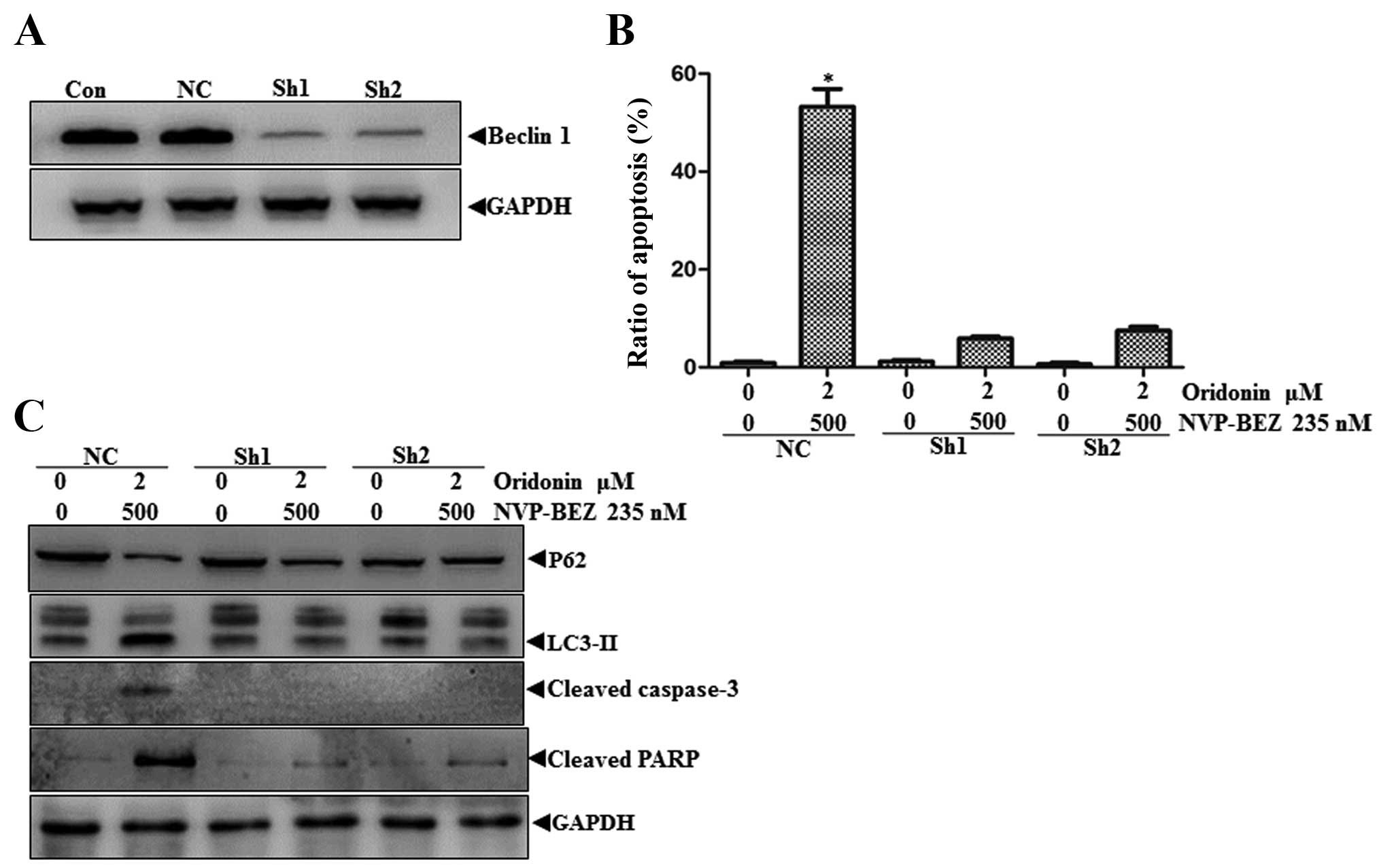
To determine whether the enhanced autophagy is
necessary for cell death induced by the combination treatment, we
used shRNA to reduce the expression of Beclin-1 (Atg6), an integral
component of the autophagic machinery, and examined whether its
loss-of-function impacts on the combination-induced apoptosis. As
expected, the expression level of Beclin-1 in SHSY-5Y cells was
markedly reduced by shRNA treatment (Fig. 5A). To provide another level of
confirmation, we performed flow cytometric analysis of Beclin-1
deficient cells stained with fluorescein isothiocyanate
(FITC)-labeled Annexin V and propidium iodide (PI) to examine the
population level of apoptotic response to the combination
treatment. As shown in Fig. 5B,
reduction of Beclin-1 protein decreased significantly the fraction
of apoptotic cells in response to the co-treatment as quantified by
the percentage of both early and late apoptotic cells.
ShRNA-mediated knockdown of Beclin-1 led to a reduction of p62
degradation and LC3-II conversion and a significant rescue of cell
death with the evidence of cleaved caspase-3 and PARP absence in
response to the combination treatment of NVP-BEZ235 and oridonin
(Fig. 5C). These data strongly
suggested that autophagy played a key role in the generation of
antineoplastic effects of NVP-BEZ235 co-treatment with oridonin in
neuroblastoma cells.
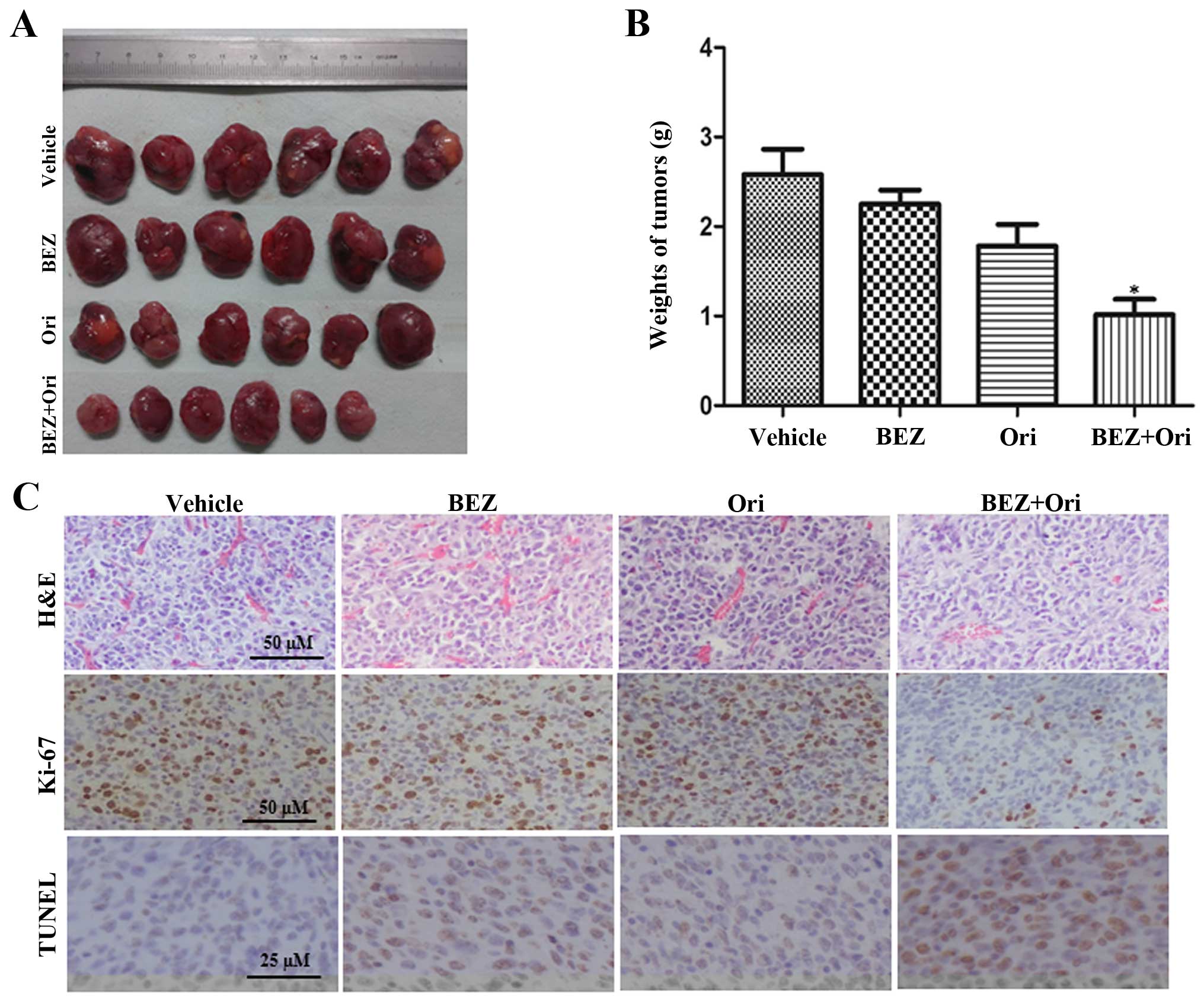
Co-treatment with NVP-BEZ235 and oridonin
induces enhanced antitumor activity in NB xenografts
We next determined whether the synergism also exists
in a preclinical NB xenograft mouse model. We initiated a tumor
with subcutaneous injection of SHSY-5Y cells on the flank of nude
mice. When tumor size reached ~100 mm3, mice were
intraperitoneally injected with vehicle, NVP-BEZ235 (20 mg/kg),
oridonin (10 mg/kg), or their combination daily for 4 weeks and
tumor sizes were measured every fifth day. As shown in Fig. 6A and B, the combination of
NVP-BEZ235 and oridonin had a much more significant effect on
suppression of tumor growth compared to any single drug. In
addition, no significant changes in body weights or daily
activities of tumor-bearing mice were observed after combination
treatment, suggesting that the combination of these two drugs was
well tolerated.
Furthermore, the proliferation and apoptosis index
of tumor tissues were assessed by Ki-67 immunohistochemistry and
TUNEL assay (Fig. 6C). Monotherapy
with NVP-BEZ235 or oridonin only marginally decreased the
proliferation of tumor tissues. While, consistently with the
cytostatic effects induced by the combined treatment, proliferation
of tumor tissues measured by Ki-67 immunostaining was also markedly
reduced in co-treated mice. Quantification of TUNEL staining did
not reveal any significant change in apoptosis between vehicle
control and NVP-BEZ235 or oridonin monotherapy group. However,
there was significantly increased apoptosis of tumor tissues in
combination treated mice as compared to vehicle control.
Discussion
The current chemotherapy strategies for the
treatment of neuroblastoma have focused on intensifying and
alternating combinations of cyclophosphamide, doxorubicin,
vincristine, cisplatin, and etoposide (21,22).
Despite dose-intensive, multiagent induction chemotherapy,
multicenter phase 3 trials suggested that the CR/VGPR rate is still
around 50% (23,24). Given that conventional
chemotherapeutics are nearing their maximum potential with regards
to efficacy and patient tolerance, there is an urgent need to
explore more effective and better-tolerated targeted therapy.
In this study, we observed that NVP-BEZ235 as a
single agent can effectively block NB cell proliferation through
inhibition of cell cycle progression but not activation of
apoptosis, as evidenced by significant G1 phase arrest and the
absence of caspase-3 cleavage. Consistent with this observation,
Manara et al showed that NVP-BEZ235 induced sarcoma stasis,
by arresting cells in G1 phase of the cell cycle, without
remarkable effects on apoptosis (13). It was postulated that autophagy
induced by NVP-BEZ235 was a pro-survival mechanism which rendered
tumor cells capable of anti-apoptosis surviving in the kinase
inhibitor (25–27).
Autophagy is an evolutionarily conserved mechanism
by which cellular material is delivered to lysosomes for
degradation and recycling (28,29).
Although previous reports suggest that the proper quantity of
autophagy promotes malignant cell survival, there is emerging
evidence that excessive autophagy modulates the cell death
machinery in a positive way. Thus, it is conceivable that the
induction of autophagy is cell context-dependent, and the extent of
autophagy dictates the cellular outcome. For instance, there is
evidence that autophagy promotes resistance of breast cancer cells
to the monoclonal anti-HER2 antibody trastuzumab (30). Similarly, hypoxia-induced autophagy
leads to chemoresistance of hepatocellular carcinoma cells
(31,32). On the other hand, there is evidence
that autophagic cell death is critical for the generation of the
effects of As2O3 on acute myelogenous
leukemia cells (33) and
successful induction of prolonged remissions of acute lymphoblastic
leukemia by the combination of vincristine and RAD001 (34), implicating autophagy as a mechanism
by which certain antineoplastic agents generate their antitumor
activities. Hence, enhancement of NVP-BEZ235 induced autophagy may
represent a new chemotherapy strategy for advanced
neuroblastoma.
Therefore, oridonin, a natural compound that has
been demonstrated to induce cell apoptosis and autophagy in several
types of tumor cells, was selected to deal with NB cells together
with NVP-BEZ235. The findings indicated that the combined treatment
of the two drugs led to programmed death of NB cells in a
synergistic manner, dramatically enhancing the antitumor efficacy
on human NB cells in vitro and in vivo. Notably, the
synergistic activation of the apoptotic pathway was accompanied by
significant decreased p62 expression as well as upregulation of
LC3-II. Knockdown of Beclin-1 which plays a key role in the
initiation of autophagy pathway with shRNA led to a significant
rescue of cell death with the evidence of cleaved caspase-3 and
PARP absence and decreased apoptosis rate from flow cytometry in
response to the combination treatment of NVP-BEZ235 and oridonin.
Such enhanced autophagy has also been reported by other studies to
play an important role in synergic therapeutic effects induced by
combination treatment. For instance, Tai et al showed that
combined treatment with Rad001 and propachlor led to excessive
autophagy, and resulted in cell death in prostate cancer cells
(20). In regard to the mechanism
of apoptosis promotion by excessive autophagy we speculated that
apoptosis was promoted through autophagic degradation of negative
regulators that controls the apoptosis machinery. Alternately, we
supposed that autophagy induced by NVP-BEZ235 failed to reach the
threshold to autophagic cell death, but it interrupted tumor
intracellular homeostasis and enhanced the chemosensitivity of
neuroblastoma cells to oridonin treatment. While the challenge
remains to clearly understand the underlying mechanisms by which
enhanced autophagy induced the combination of NVP-BEZ235 with
oridonin to promote programmed cell death of NB.
These results showed that combination treatment of
the two compounds synergize in the induction of NB cell death in
vitro and in vivo, thus providing a novel strategy to
develop combination therapies for advanced neuroblastoma patients
that have failed the currently available therapies.
Acknowledgements
This study was supported in part by the National
Natural Science Foundation of China (81172322 and 81302006),
Shanghai Municipal Education Committee (13ZZ089), Science and
Technology Committee of Shanghai (14401901500) and Science and
Technology Committee of Baoshan District (12-E-2).
Abbreviations:
|
NB
|
neuroblastoma
|
|
MRD
|
minimal residual disease
|
|
PI3K
|
phosphatidyl inositol 3-kinase
|
|
mTOR
|
the mammalian target of rapamycin
|
|
CQ
|
chloroquine
|
|
3-mA
|
3-methyladenine
|
|
shRNA
|
short hairpin RNA
|
|
AO
|
acridine orange
|
|
EB
|
ethidium bromide
|
|
TUNEL
|
TdT-mediated dUTP nick-end
labeling
|
|
FITC
|
fluorescein isothiocyanate
|
|
PI
|
propidium iodide
|
|
CR
|
complete remission
|
|
VGPR
|
very good partial remission
|
References
|
1
|
Berthold F, Boos J, Burdach S, Erttmann R,
Henze G, Hermann J, Klingebiel T, Kremens B, Schilling FH, Schrappe
M, et al: Myeloablative megatherapy with autologous stem-cell
rescue versus oral maintenance chemotherapy as consolidation
treatment in patients with high-risk neuroblastoma: A randomised
controlled trial. Lancet Oncol. 6:649–658. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Park JR, Eggert A and Caron H:
Neuroblastoma: Biology, prognosis, and treatment. Hematol Oncol
Clin North Am. 24:65–86. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Westhoff MA, Faham N, Marx D, Nonnenmacher
L, Jennewein C, Enzenmüller S, Gonzalez P, Fulda S and Debatin KM:
Sequential dosing in chemosensitization: Targeting the
PI3K/Akt/mTOR pathway in neuroblastoma. PLoS One. 8:e831282013.
View Article : Google Scholar
|
|
4
|
Brodeur GM: Neuroblastoma: Biological
insights into a clinical enigma. Nat Rev Cancer. 3:203–216. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maris JM: Recent advances in
neuroblastoma. N Engl J Med. 362:2202–2211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Opel D, Poremba C, Simon T, Debatin KM and
Fulda S: Activation of Akt predicts poor outcome in neuroblastoma.
Cancer Res. 67:735–745. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fulda S: The PI3K/Akt/mTOR pathway as
therapeutic target in neuroblastoma. Curr Cancer Drug Targets.
9:729–737. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Izycka-Swieszewska E, Drozynska E, Rzepko
R, Kobierska-Gulida G, Grajkowska W, Perek D and Balcerska A:
Analysis of PI3K/AKT/mTOR signalling pathway in high risk
neuroblastic tumours. Pol J Pathol. 61:192–198. 2010.
|
|
9
|
Roper J, Richardson MP, Wang WV, Richard
LG, Chen W, Coffee EM, Sinnamon MJ, Lee L, Chen PC, Bronson RT, et
al: The dual PI3K/mTOR inhibitor NVP-BEZ235 induces tumor
regression in a genetically engineered mouse model of PIK3CA
wild-type colorectal cancer. PLoS One. 6:e251322011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen X, Zhao M, Hao M, Sun X, Wang J, Mao
Y, Zu L, Liu J, Shen Y, Wang J, et al: Dual inhibition of PI3K and
mTOR mitigates compensatory AKT activation and improves tamoxifen
response in breast cancer. Mol Cancer Res. 11:1269–1278. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zito CR, Jilaveanu LB, Anagnostou V, Rimm
D, Bepler G, Maira SM, Hackl W, Camp R, Kluger HM and Chao HH:
Multilevel targeting of the phosphatidylinositol-3-kinase pathway
in non-small cell lung cancer cells. PLoS One. 7:e313312012.
View Article : Google Scholar
|
|
12
|
Ribback S, Cigliano A, Kroeger N, Pilo MG,
Terracciano L, Burchardt M, Bannasch P, Calvisi DF and Dombrowski
F: PI3K/AKT/mTOR pathway plays a major pathogenetic role in
glycogen accumulation and tumor development in renal distal tubules
of rats and men. Oncotarget. 6:13036–13048. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Manara MC, Nicoletti G, Zambelli D,
Ventura S, Guerzoni C, Landuzzi L, Lollini PL, Maira SM,
García-Echeverría C, Mercuri M, et al: NVP-BEZ235 as a new
therapeutic option for sarcomas. Clin Cancer Res. 16:530–540. 2010.
View Article : Google Scholar
|
|
14
|
Chanthery YH, Gustafson WC, Itsara M,
Persson A, Hackett CS, Grimmer M, Charron E, Yakovenko S, Kim G,
Matthay KK, et al: Paracrine signaling through MYCN enhances
tumor-vascular interactions in neuroblastoma. Sci Transl Med.
4:115ra32012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li CY, Wang EQ, Cheng Y and Bao JK:
Oridonin: An active diterpenoid targeting cell cycle arrest,
apoptotic and autophagic pathways for cancer therapeutics. Int J
Biochem Cell Biol. 43:701–704. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Z, Ouyang L, Peng H and Zhang WZ:
Oridonin: Targeting programmed cell death pathways as an
anti-tumour agent. Cell Prolif. 45:499–507. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu DL, Bu HQ, Jin HM, Zhao JF, Li Y and
Huang H: Enhancement of the effects of gemcitabine against
pancreatic cancer by oridonin via the mitochondrial
caspase-dependent signaling pathway. Mol Med Rep. 10:3027–3034.
2014.PubMed/NCBI
|
|
18
|
Guo Y, Shan Q, Gong Y, Lin J, Yang X and
Zhou R: Oridonin in combination with imatinib exerts synergetic
anti-leukemia effect in Ph+ acute lymphoblastic leukemia
cells in vitro by inhibiting activation of LYN/mTOR signaling
pathway. Cancer Biol Ther. 13:1244–1254. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang Y, Hamed HA, Cruickshanks N, Fisher
PB, Grant S and Dent P: Obatoclax and lapatinib interact to induce
toxic autophagy through NOXA. Mol Pharmacol. 81:527–540. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tai S, Sun Y, Liu N, Ding B, Hsia E, Bhuta
S, Thor RK, Damoiseaux R, Liang C and Huang J: Combination of
Rad001 (everolimus) and propachlor synergistically induces
apoptosis through enhanced autophagy in prostate cancer cells. Mol
Cancer Ther. 11:1320–1331. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fong A and Park JR: High-risk
neuroblastoma: A therapy in evolution. Pediatr Hematol Oncol.
26:539–548. 2009. View Article : Google Scholar
|
|
22
|
National Cancer Institute. Board PDQPTE:
Neuroblastoma Treatment (PDQ(R)): Health Professional Version. PDQ
Cancer Information Summaries. National Cancer Institute (US);
Bethesda (MD): 2002, http://www.cancer.gov/publications/pdq/information-summaries/adult-treatment.
Accessed, January 14, 2016
|
|
23
|
Pearson AD, Pinkerton CR, Lewis IJ, Imeson
J, Ellershaw C and Machin D; European Neuroblastoma Study Group;
Children's Cancer and Leukaemia Group (CCLG formerly United Kingdom
Children's Cancer Study Group). High-dose rapid and standard
induction chemotherapy for patients aged over 1 year with stage 4
neuroblastoma: A randomised trial. Lancet Oncol. 9:247–256. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kreissman SG, Seeger RC, Matthay KK,
London WB, Sposto R, Grupp SA, Haas-Kogan DA, Laquaglia MP, Yu AL,
Diller L, et al: Purged versus non-purged peripheral blood
stem-cell transplantation for high-risk neuroblastoma (COG A3973):
A randomised phase 3 trial. Lancet Oncol. 14:999–1008. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li H, Jin X, Zhang Z, Xing Y and Kong X:
Inhibition of autophagy enhances apoptosis induced by the
PI3K/AKT/mTor inhibitor NVP-BEZ235 in renal cell carcinoma cells.
Cell Biochem Funct. 31:427–433. 2013. View
Article : Google Scholar
|
|
26
|
Echeverry N, Ziltener G, Barbone D, Weder
W, Stahel RA, Broaddus VC and Felley-Bosco E: Inhibition of
autophagy sensitizes malignant pleural mesothelioma cells to dual
PI3K/mTOR inhibitors. Cell Death Dis. 6:e17572015. View Article : Google Scholar :
|
|
27
|
Ji Y, Di W, Yang Q, Lu Z, Cai W and Wu J:
Inhibition of autophagy increases proliferation inhibition and
apoptosis induced by the PI3K/mTOR inhibitor NVP-BEZ235 in breast
cancer cells. Clin Lab. 61:1043–1051. 2015.PubMed/NCBI
|
|
28
|
Jiang X, Overholtzer M and Thompson CB:
Autophagy in cellular metabolism and cancer. J Clin Invest.
125:47–54. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fulda S and Kögel D: Cell death by
autophagy: Emerging molecular mechanisms and implications for
cancer therapy. Oncogene. 34:5105–5113. 2015. View Article : Google Scholar
|
|
30
|
Rodríguez CE, Reidel SI, Bal de Kier Joffé
ED, Jasnis MA and Fiszman GL: Autophagy protects from
trastuzumab-induced cytotoxicity in HER2 overexpressing breast
tumor spheroids. PLoS One. 10:e01379202015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Song J, Qu Z, Guo X, Zhao Q, Zhao X, Gao
L, Sun K, Shen F, Wu M and Wei L: Hypoxia-induced autophagy
contributes to the chemoresistance of hepatocellular carcinoma
cells. Autophagy. 5:1131–1144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Toshima T, Shirabe K, Matsumoto Y, Yoshiya
S, Ikegami T, Yoshizumi T, Soejima Y, Ikeda T and Maehara Y:
Autophagy enhances hepatocellular carcinoma progression by
activation of mitochondrial β-oxidation. J Gastroenterol.
49:907–916. 2014. View Article : Google Scholar
|
|
33
|
Goussetis DJ, Altman JK, Glaser H, McNeer
JL, Tallman MS and Platanias LC: Autophagy is a critical mechanism
for the induction of the antileukemic effects of arsenic trioxide.
J Biol Chem. 285:29989–29997. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Crazzolara R, Cisterne A, Thien M, Hewson
J, Baraz R, Bradstock KF and Bendall LJ: Potentiating effects of
RAD001 (Everolimus) on vincristine therapy in childhood acute
lymphoblastic leukemia. Blood. 113:3297–3306. 2009. View Article : Google Scholar : PubMed/NCBI
|