Introduction
Cancers are a group of diseases characterized by
uncontrolled cell growth and spread. Primary liver cancer,
especially hepatocellular carcinoma (HCC), is the fifth most common
malignancy with more than 500,000 new cases diagnosed every year,
and the third leading cause of cancer death with a
mortality-to-incidence ratio exceeding 0.9 in the world (1,2). The
rate of HCC is annually increasing worldwide between 3 and 9%, and
the incidence of HCC is particularly higher in Southeast Asia and
sub-Saharan Africa due to the higher frequency of chronic viral
hepatitis (3). Generally, HCC is
associated with dietary aflatoxin B1 intake or heavy alcohol
consumption, and alternative causes of hepatic cirrhosis with a
persistent hepatitis B virus or C virus infection, which is a 3.2
kb, partial dsDNA, non-cytopathic virus, and the most important
etiologic factor for malignant HCC (4). Until now, although surgical
resection, orthotropic liver transplantation, and radiofrequency
ablation have shown excellent results in the treatment of early
stage liver cancer, there is no curative therapy for patients with
advanced HCC (2). Therefore, HCC
remains a serious global problem, and more effective prevention,
diagnosis and treatment strategies are urgently needed.
Apoptosis is linked to cell cycle arrest, and the
blockade of the cell cycle is regarded as effective for eliminating
cancer cells. In recent years, many chemotherapeutic agents have
been shown to impart anti-proliferative effects via arrest of cell
division at certain checkpoints in the cell cycle. The concept of
cell cycle-mediated apoptosis has gained increasing attention as
this pathway may provide minimal opportunity for acquired drug
resistance, decreased mutagenesis and reduced toxicity (5,6).
These observations suggest new approaches could alter uncontrolled
cancer cell growth by modulating cell cycle regulators causing cell
cycle arrest and could be useful in prevention and/or intervention
in human cancer (5).
Herbal medicines play an increasing role in primary
health care systems among the population as synthetic anticancer
remedies are beyond the reach of the common man due to the high
cost and toxic side effects. Many plant-derived drugs have a vital
role in cancer therapy which execute their therapeutic effects by
inhibiting cancer activating enzymes and hormones, stimulating DNA
repair mechanism, promoting production of protective enzymes
inducing antioxidant action, killing tumors via programmed cell
death and enhancing immunity (7).
Cyperus amuricus (C. amuricus) is a monocotyledonous and
multipurpose medicinal herb which belongs to Cyperaceae
family. It is a perennial sedge with slender and scaly creeping
rhizomes, fibrous base and arises singly from the triquetrous
tubers with dense tuft of 10 to 60-cm. This delicate grass, growing
in tropical, subtropical and temperate regions; is widespread in
North America, Japan, Korea and Russia (Far East) (8). A few studies have been recently
commissioned to elucidate the pharmacological activities of C.
amuricus. The dried whole plant of this herb, well-known as
Chinese Amuersuocao or Korean Bangdongsani, has been traditionally
prescribed for exerting astringent, diuretic, diaphoretic,
desiccant and cordial properties in folk medicine (9). Infusion of this grass has been
commonly credited with treating wounds, tumors, piles and other
intestinal problems in Bangladesh (10). Previous phytochemical investigation
on C. amuricus revealed the presence of three antioxidant
phenolic components, including 3,4-dimethoxy benzoic acid,
4-hydroxybenzoic acid, and piceatannol which exhibited powerful
free radical scavenging, especially against DPPH and superoxide
anions (11). The steam
distillation of C. amuricus also showed 57.3% inhibitory
activity of pancreatic lipase in vitro (12). However, there is no scientific
report regarding the potential of C. amuricus in the
prevention and treatment of cancer. Hence, the present study was
conducted to elucidate the underlying mechanisms of the extract of
C. amuricus-induced antiproliferation, cell cycle arrest,
and apoptosis in HCC Hep3B cells in vitro, so as to supply
scientific rationales for using C. amuricus as a new
promising chemopreventive and/or chemotherapeutic agent against
liver cancer.
Materials and methods
Cell culture and reagents
Hep3B (hepatocellular carcinoma), A549 (human lung
adenocarcinoma) and HaCaT (non-cancerous human keratinocyte) cells
were purchased from the American Tissue Culture Collection (ATCC;
Manassas, VA, USA). Cells were cultured in Dulbecco's modified
Eagle's medium (DMEM) (for Hep3B and HaCaT), RPMI-1640 (for A549)
(HyClone Laboratories, Logan, UT, USA) medium supplemented with 10%
heat inactivated fetal bovine serum (FBS; HyClone Laboratories) and
1% penicillin-streptomycin (PAA Laboratories GmbH, Pasching,
Austria) at 37°C and 5% CO2. The methyl alcohol extract
from the whole plant of C. amuricus (distribution number:
010–032) was obtained from the Korea Plant Extract Bank (KPEB,
Cheongju, Korea) with the purity of ≥99.9%, HPLC.
Cell viability assay
Exponential phases of Hep3B, A549 and HaCaT cells
(1×104 cells/well) were seeded on 96-well plates (SPL
Life Sciences, Gyeonggi, Korea) in triplicate. Following overnight
incubation, cells were treated with various concentrations (50,
100, 150 and 200 μg/ml) of an extracted fraction of C.
amuricus and incubated for 24 h. After the treatment, 10 μl of
EZ-Cytox Cell Viability Assay Solution WST-1® (Daeil Lab
Service, Seoul, Korea) was added to each well and incubated for an
additional 3 h. The absorbance of the reaction was measured using
an ELISA reader (Molecular Devices, Sunnyvale, CA, USA) at 460 nm
and cell viability was calculated. The cytotoxic activity of the
extract was expressed as an IC50 value, which is the
concentration of the extract that caused 50% cell death. The
extract of C. amuricus with an IC50 value ≤150
μg/ml was considered active on Hep3B cells. Dimethyl sulfoxide
(DMSO; Sigma-Aldrich, St. Louis, MO, USA) was used to dilute the
extract and the final concentration of DMSO in each well was not in
excess of 0.05% (v/v). No adverse effect due to the presence of
DMSO was observed.
Treatment of Z-VAD-fmk
Hep3B cells were divided into four groups to compare
caspase-dependent and caspase-independent cell death as follows:
non-treated, Z-VAD-fmk (pan-caspase inhibitor; Sigma-Aldrich),
C. amuricus, and C. amuricus with Z-VAD-fmk groups.
After 24 h, the cells were transferred to a fresh medium containing
no agent (control group), Z-VAD-fmk (50 μM Z-VAD-fmk), C.
amuricus (150 μg/ml C. amuricus), or a combination (150
μg/ml C. amuricus and 50 μM Z-VAD-fmk). After treatment for
24 h, 10 μl of WST-1 solution was added to each well, further
incubated for 3 h, and then cell viability was measured at 460 nm
using an ELISA reader.
DAPI staining
Hep3B cells were treated with the concentration of
100, 150 and 200 μg/ml of the C. amuricus extract for 24 h.
The cells were rinsed once with phosphate-buffered saline (PBS)
buffer (135 mM sodium chloride, 2.7 mM potassium chloride, 4.3 mM
sodium phosphate, 1.4 mM potassium dihydrogen phosphate) and
stained by addition of 1 μg/ml DAPI solution (Roche Applied
Science, Indianapolis, IN, USA) in methanol (Sigma-Aldrich). After
incubation in the dark at 37°C for 20 min, cells were washed twice
with PBS buffer, and then fixed with 4% formaldehyde (Junsei,
Tokyo, Japan) for 15 min. The nuclear morphology of the cells was
observed under a Laser Scanning Confocal microscope (Carl Zeiss LSM
700; Carl Zeiss, Oberkochen, Germany).
DNA fragmentation
For detecting genomic DNA fragmentation, the treated
Hep3B cells were washed with ice-cold PBS buffer and harvested. The
collected cells were handled by following the protocol of
DNeasy® Blood and Tissue kit (Qiagen GmbH, Hilden,
Germany). The isolated DNA was separated in a 1.5% agarose gel
(Life Technologies Inc., Grand Island, NY, USA) and visualized by
ethidium bromide staining (Sigma-Aldrich) under a UV
transilluminator (Vilber Lourmat, Marne-la-Vallee, France).
Cell cycle analysis
Briefly, Hep3B cells were harvested by
trypsinization and fixed with 70% ethanol overnight at 4°C. Then,
the cells were resuspended in PBS buffer containing 0.2 mg/ml RNase
and incubated for 1 h at 37°C. The cells were stained with 40 μg/ml
propidium iodide (Sigma-Aldrich) at room temperature for 30 min in
the dark. The cell cycle was analyzed based on DNA contents using a
flow cytometer (Becton-Dickinson, Mountain View, CA, USA).
Protein extraction and western blot
analysis
Hep3B cells were washed once with PBS buffer and
then lysed by the addition of lysis buffer [(50 mM Tris-Cl (pH
7.5), 150 mM NaCl, 1 mM DTT, 0.5% NP-40, 1% Triton X-100, 1%
Deoxycholate, 0.1% SDS and proteinase inhibitors (PMSF, EDTA,
aprotinin, leupeptin and prostatin A)] (Intron Biotechnology,
Gyeonggi, Korea). After 30 min on ice, lysates were collected and
clarified by centrifugation at 14,000 rpm for 20 min at 4°C.
Cytosolic fractions were prepared using NE-PER® Nuclear
and Cytoplasmic Extraction reagents (Thermo Fisher Scientific,
Rockford, IL, USA) according to the manufacturer's protocol.
Aliquots of whole cell lysates or cytosolic fractions were
subjected to 10% SDS-PAGE and then transferred to a nitrocellulose
membrane (Pall Corp., Pensacola, FL, USA). The membranes were
blocked with 5% skim milk in PBST (PBS buffer and 0.5% Tween-20).
After blocking non-specific sites, the membranes were probed with
primary antibodies (Cell Signaling Technology, Inc., Danvers, MA,
USA) and then washed in PBST three times, followed by incubation
for 1 h with horseradish peroxidase-conjugated anti-rabbit IgG or
anti-mouse IgG as second antibodies (Cell Signaling Technology).
The blots were then washed in PBST and visualized by an enhanced
chemiluminescent (ECL) detection solution (Pierce, Rockford, IL,
USA). Anti-Bim was obtained from Santa Cruz Biotechnology (Santa
Cruz, CA, USA). All other antibodies were purchased from Cell
Signaling Technology.
Statistical analysis
Determinations were performed in triplicate and the
results are presented as mean ± SD. Differences between groups were
analyzed by one or two way ANOVA, followed by Fisher's protected
least-significant difference. P<0.05 was considered as
statistical significance (*P<0.05;
**P<0.01 in the figures).
Results
Effects of C. amuricus on cell
growth
In order to understand the cytotoxicity of the
extract, Hep3B, A549 and HaCaT cells were exposed to the methanol
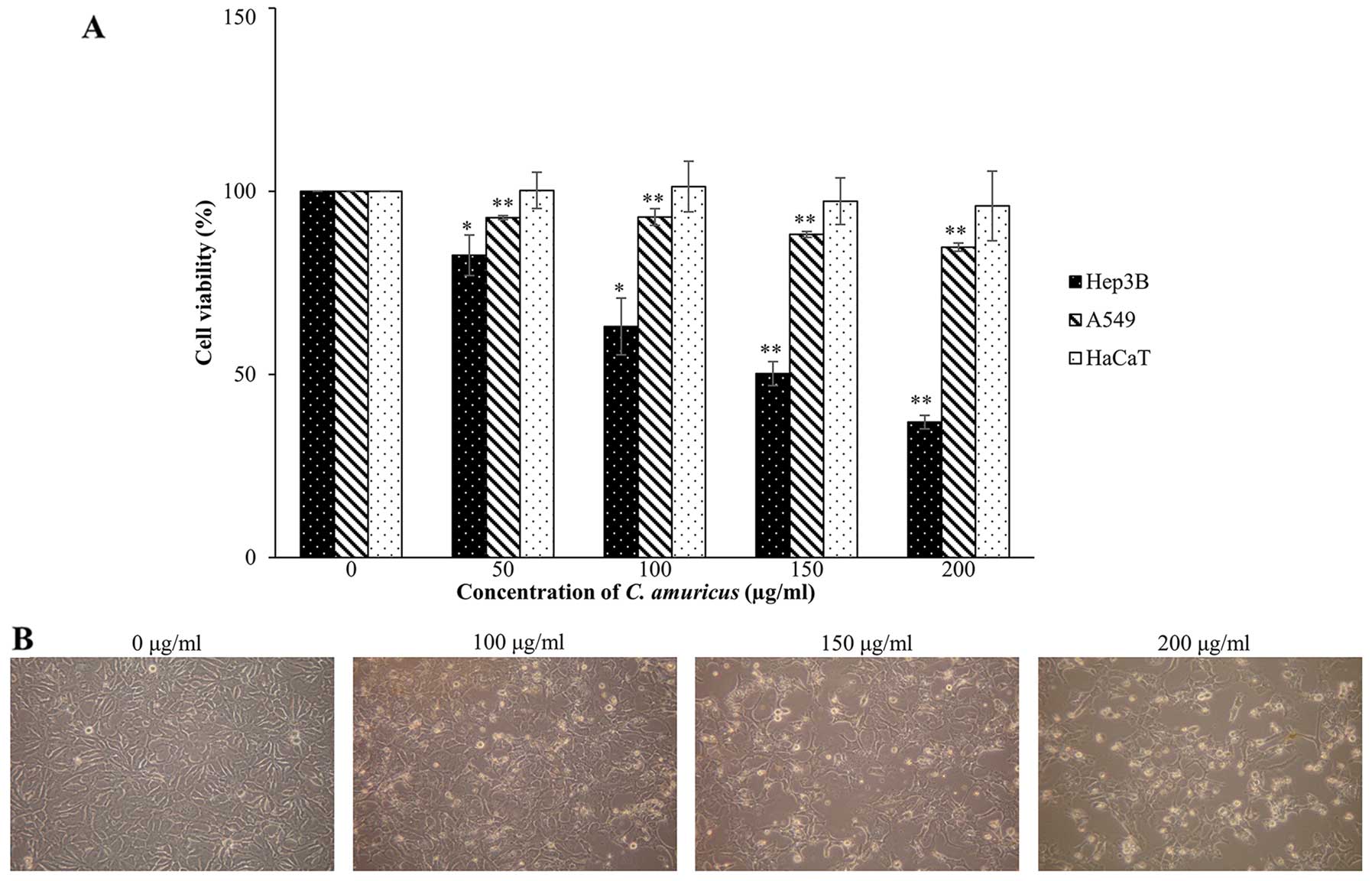
extract of C. amuricus by cell viability assay. As shown in
Fig. 1A, the C. amuricus
extract did not have significant effects on either A549 or HaCaT
cell viability while the extract (over 100 μg/ml) caused remarkable
growth inhibition and a marked decrease in cell viability only in
Hep3B cells. Cell death reached 50% with 150 μg/ml
(IC50) and 63% with 200 μg/ml of the C. amuricus
treatment for 24 h, respectively, and those concentrations were
used in subsequent experiments to investigate the mechanism of cell
death. Additionally, morphological changes of Hep3B cells treated
with or without the extract of C. amuricus for 24 h were
visualized under the inverted microscope (magnification, ×100).
Compared with the non-treated cells, most of the C.
amuricus-treated Hep3B cells (100–200 μg/ml of the extract) did
exhibit morphological features of apoptosis, such as cell membrane
blebbing, cell shrinkage, increased cytoplasm granules and
detachment from culture plates (Fig.
1B). The data initially indicate that the extract of C.
amuricus induces selective cytotoxicity in HCC Hep3B cells.
Effects of C. amuricus on the induction
of apoptosis
Further experiments were then carried out to
determine whether the extract of C. amuricus inhibits the
proliferation of Hep3B cells through the induction of apoptosis.
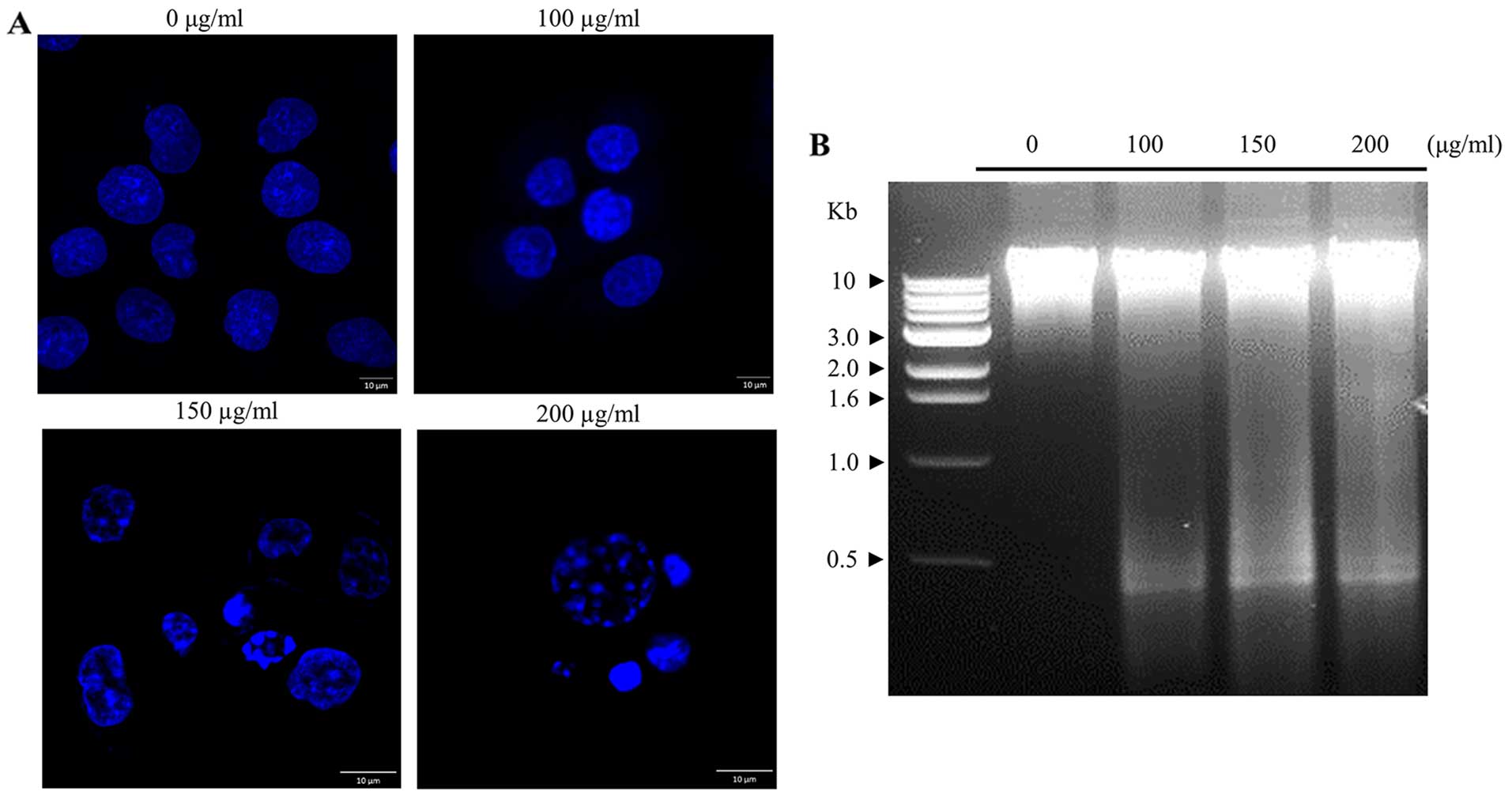
Morphological analysis following DAPI staining was examined to
analyze the phenotypic changes in the cell nucleus. The classical
hallmark of apoptotic cells with chromatin condensation and
apoptotic bodies, was observed in Hep3B cells treated with the
C. amuricus extract in a dose-dependent manner, indicating
that the extract of C. amuricus-stimulated Hep3B cell death
was a typical apoptotic cell death (Fig. 2A). More evidence in support of
apoptosis was performed by DNA fragmentation assay. The biochemical
hallmark of apoptotic cell death, in which cleavage of chromosomal
DNA at internucleosomal fragments or multiples at ~180 bp (13), results in a typical DNA
electrophoresis ladder, was visible during incubation with the
C. amuricus extract (Fig.
2B). On the basis of the above data, the profile for C.
amuricus-caused apoptosis closely correlated with its growth
suppressive effect. Thus, these phenomena demonstrate that the
extract of C. amuricus may be considered as an inducer of
apoptosis in Hep3B cells.
Effects of C. amuricus on cell cycle
distribution
In order to estimate the inhibitory effect of C.
amuricus on Hep3B cell growth, Hep3B cells were exposed to
escalating concentrations of the C. amuricus extract and
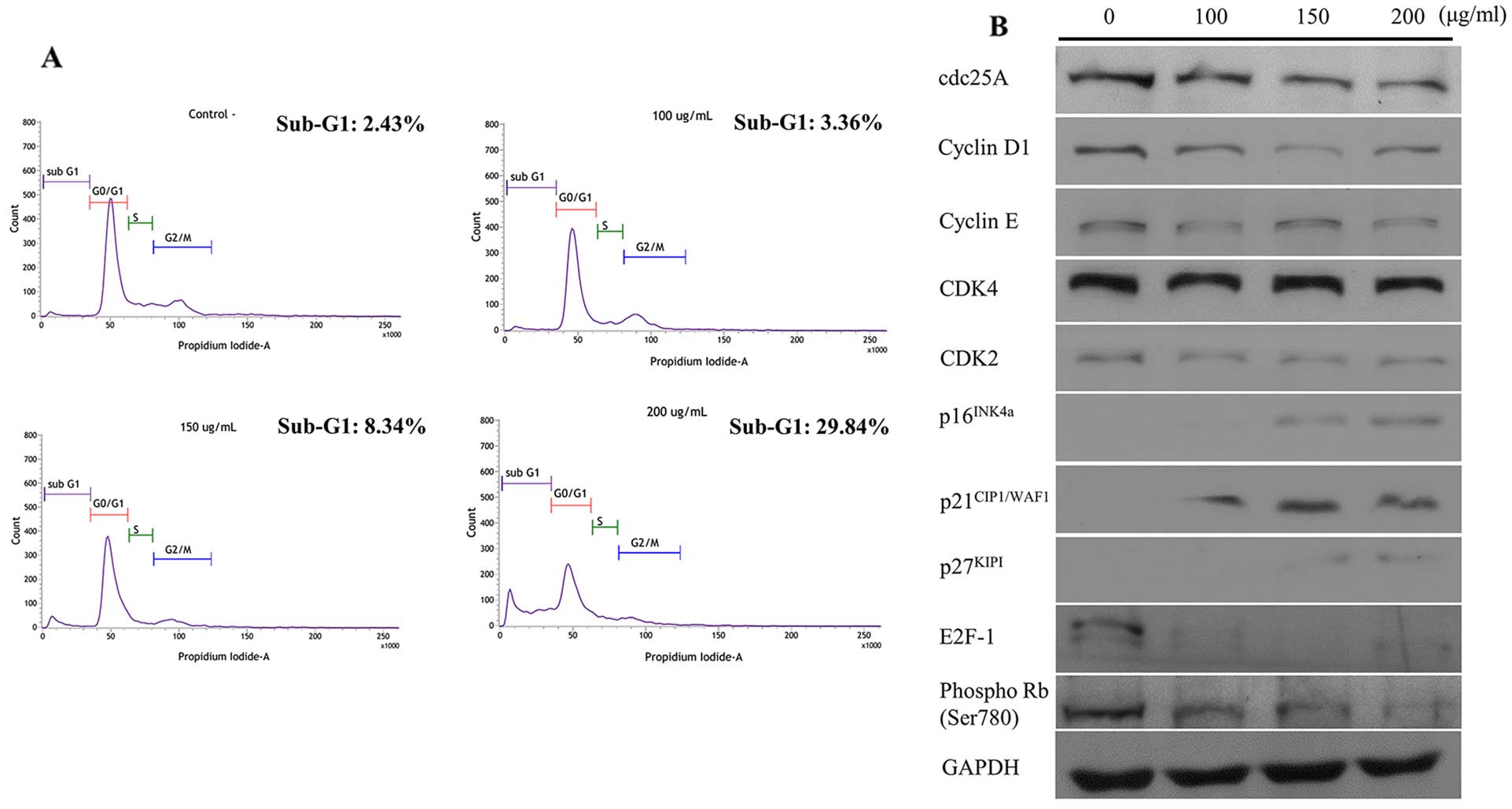
subjected to cell cycle analysis. It could be seen that a 24-h
treatment of the C. amuricus extract to Hep3B cells caused a
definite rise in sub-G1 fractions in a dose-dependent manner. The
relative percentages of cells staying at the sub-G1 phase gradually
increased from 2.43% in the non-treated cells to 3.36, 8.34 and
29.84% in Hep3B cells treated with 100, 150 and 200 μg/ml of the
C. amuricus extract, respectively (Fig. 3A). As the elevated accumulation of
sub-G1 cells exhibits the presence of apoptotic cells, the extract
of C. amuricus-promoted cell death was concomitant with the
growth inhibitory effect. To dissect the biochemical events
controlling the transition of cell cycle phases, G1-related
proteins subsequently were examined by western blot analysis
(Fig. 3B). Downregulation of
cdc25A, cyclin D1 and cyclin E, CDK4 and 2, and upregulation of
p21CIP1/WAF1, p27KIPI and p16INK4a
were noted in the Hep3B cells following C. amuricus
treatment for 24 h. Consistently, the expression of the E2F-1 and
phospho-Rb were lower in the addition of C. amuricus. The
experimental findings imply that the methanol extract of C.
amuricus blocks Hep3B cell cycle progression at the sub-G1
phase.
Effects of C. amuricus on the expression
of apoptosis-related proteins
The possible molecular mechanisms underlying C.
amuricus-induced apoptosis in Hep3B cells were investigated.
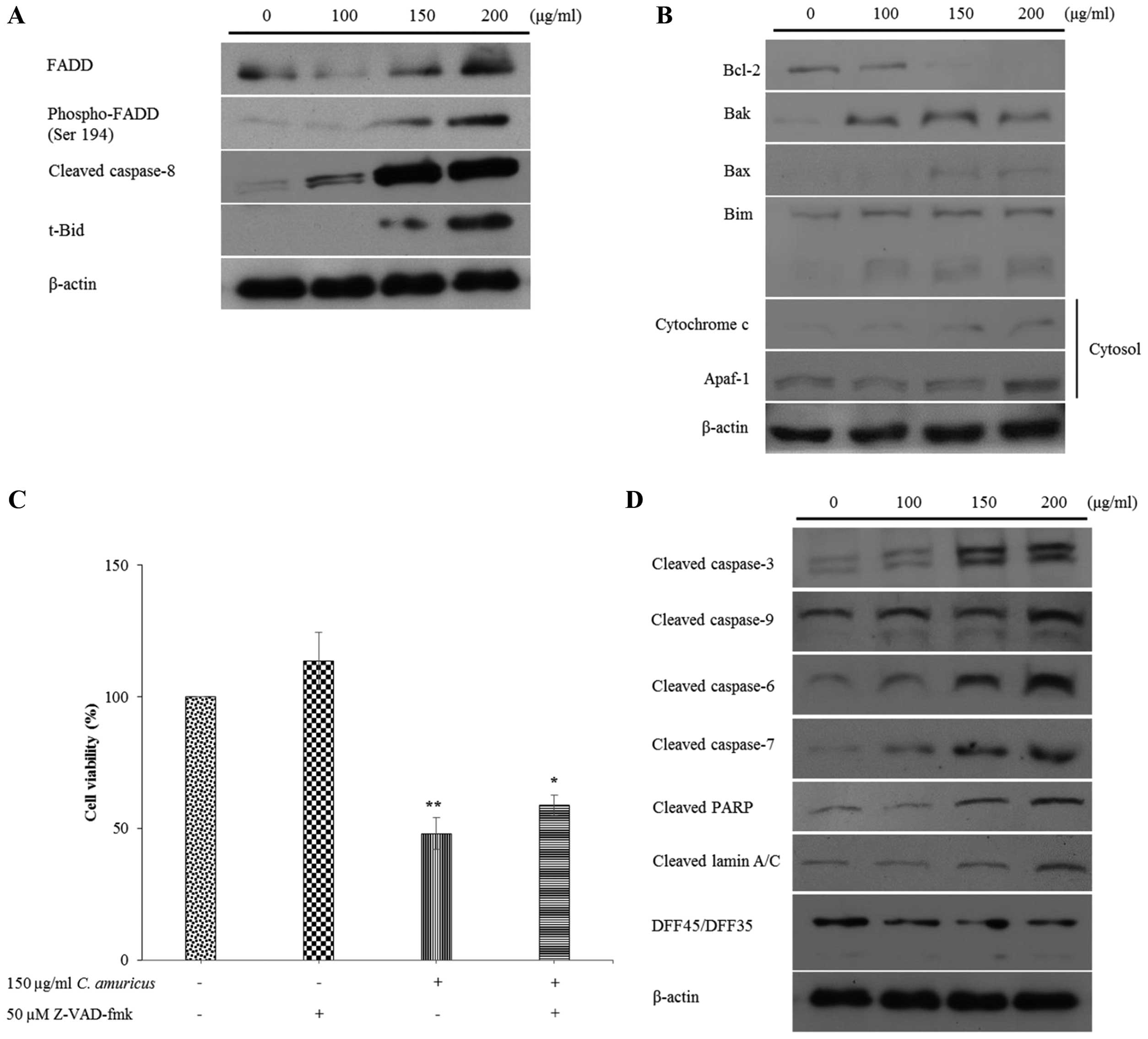
The effects of C. amuricus on the protein expression of
Fas-associated death domain (FADD) and phospho-FADD, cleaved
caspase-8, and truncated Bid (tBid, the active form of Bid) in
Hep3B cells were explored. Western blot analysis revealed that
C. amuricus caused dose-dependent increments in the levels
of FADD and phospho-FADD, cleaved caspase-8 and tBid (Fig. 4A).
The effects of C. amuricus on the
mitochondria-mediated death pathway regulated by several members of
anti- and pro-apoptotic Bcl-2 family, cytosolic cytochrome c
and apoptotic protease activating factor-1 (Apaf-1) were next
examined (Fig. 4B). The expression
of Bcl-2 was clearly decreased, by way of contrast, the protein
levels of Bak, Bax and Bim were notably extended after treatment of
Hep3B cells with the C. amuricus extract. Also, a
dose-dependent advance in the expression of cytosolic cytochrome
c and Apaf-1 was markedly detected in C.
amuricus-treated Hep3B cells.
As a first step in identifying whether caspases were
involved in the extract of C. amuricus-induced apoptosis,
Hep3B cells were treated with the caspase inhibitor Z-VAD-fmk. As
shown in the results, more cells were alive when treated with
Z-VAD-fmk (114%) and the C. amuricus extract plus Z-VAD-fmk
(59%) than treated with the C. amuricus extract alone
(48.1%) (Fig. 4C), suggesting that
the extract of C. amuricus is able to affect the activity of
caspases. In addition, results of western blot analysis presented
the expression of multiple apoptotic proteins, including cleaved
caspase-9, −3, −7 and −6 were distinctly higher in response to the
extract of C. amuricus treatment in a dose-dependent manner.
The activation of caspases often lead to the proteolytic cleavage
of target proteins poly(ADP ribose) polymerase (PARP), lamins and
DNA fragmentation factor DFF45. Under these conditions, cleaved
PARP and cleaved lamin A/C protein were expanded gently, while
DFF45 was reduced effectively by the treatment with increasing
concentrations of the extract of C. amuricus (Fig. 4D).
Discussion
The anticancer activities of natural herbal medicine
were thoroughly investigated long ago. Also, their crucial effects
on the pathophysiology associated with cancers have recently
received special attention, due to many epidemiological reports
proposing that several beneficial effects of herbal extracts
significantly reduce the risk of many cancers with few
side-effects. Although numerous Cyperus species are
popularly used in folk medicine and pharmacies as antioxidants and
anti-inflammatories, scientific evidence of their effects is
essentially required. Therefore, the potential of C.
amuricus as an anticancer preparation was elucidated in the
present study.
The imbalance between cell proliferation and death
is considered to be an important event in cancer progression. Among
the effects of antitumor reagents, apoptosis and growth inhibition
are the most common responses on cancer cells (14). The study demonstrated that the
extract of C. amuricus was significantly toxic towards Hep3B
cells, while essentially non-toxic to A549 and HaCaT cells
(Fig. 1A). Furthermore, apoptosis
is one of the most prevalent pathways through which chemopreventive
and/or chemotherapeutic strategies can inhibit the overall growth
of cancer cells. Apoptosis involves specific morphological and
biochemical changes such as cell shrinkage, chromatin condensation,
membrane blebbing, more floating and DNA fragmentation (13). Hence, induction of apoptosis is
recognized as a useful indicator for cancer treatment and
prevention. Particularly, after 24 h of treatment with C.
amuricus extract, marked morphological changes, including the
obvious destruction of the monolayer, shrinkage and extensive
detachment of cells (Fig. 1B), the
increased nuclear chromatin condensation in DAPI staining (Fig. 2A) as well as evident DNA
fragmentations (Fig. 2B) were
observed, suggesting that C. amuricus-induced cell death
involves an apoptotic mechanism.
Cancer cells lack normal growth controls, exhibit
loss of cell cycle control, have unlimited reproductive potential
and have growth-signal self-sufficiency (15). Cell cycle machinery depends on the
coordinated activity of protein kinase complexes, each consisting
of a cyclin-dependent kinases (CDKs) and cyclins, which act
consecutively in G1 to initiate S phase and in G2 to initiate
mitosis (16). Progression through
G1 phase requires the activities of cyclin D-dependent CDK4 or 6,
followed by activation of the cyclin E- and cyclin A-dependent
kinase CDK2. The cyclin-CDK complex formed during G1-phase
catalyzes phosphorylation of the dominant inhibitor of G1/S-cell
cycle progression, the retinoblastoma Rb family of tumor suppressor
proteins, thereby blocking their inhibitory activity allowing the
cell to progress into S phase. The Rb family proteins bind to
members of the E2F transcription factor family and block the
E2F-dependent transcription of genes controlling the G1 to S phase
transition and subsequent DNA synthesis. The phosphorylation of Rb
disrupts its association with E2F, thereby reducing the suppression
of E2F target genes and activating the transcription outside G1/S
(17). It is also known that these
cyclin-CDK complexes often bind to CDK inhibitors (CDKIs), to
conduct pivotal functions in cell cycle regulation via the
coordination of internal and external signals that inhibit cell
cycle progression at critical checkpoints. One group is the INK4
(inhibitors of CDK4) family, which has four members
p16INK4a, p15INK4b, p18INK4c and
p19INK4d, all of which share the ability to control the
G1/S transition. The second group of CDKIs includes
p21CIP1/WAF1 and p27KIPI (18). In the present study, cell cycle
analysis exhibited that the extract of C. amuricus has the
ability to induce sub-G1 phase arrest in Hep3B cells (Fig. 3A). Another interesting finding is
that the concomitant occurrence of apoptosis after C.
amuricus treatment is mediated with cell cycle arrest in the
sub-G1 phase. The increase in sub-G1 phase of C.
amuricus-treated Hep3B cells may be due to the induction of S
phase arrest. Western blot analysis further examined proteins
associated with the cell cycle. C. amuricus drastically
decreased the levels of the protein kinase complexes cdc25A, cyclin
D1 and cyclin E, CDK4 and 2 and increased the levels of
p16INK4a, p21CIP1/WAF1 and p27KIPI
proteins in Hep3B cells (Fig. 3B).
C. amuricus also induced a reduction in Rb phosphorylation,
then resulted in a contraction of the E2F-1 protein (Fig. 3B). The data indicated that C.
amuricus elicits cell cycle arrest at sub-G1 phases in Hep3B
cells through the induction of CDKIs and the inhibition of cyclins
and CDKs.
Apoptotic pathways include the extrinsic death
receptor- and intrinsic mitochondria-mediated pathways (19). The extrinsic signaling pathway is
related to the membrane death receptors that belong to the tumor
necrosis factor (TNF) receptor gene superfamily. To date, the fatty
acid synthase ligand/receptor (FasL/FasR) and TNF-α/TNFR1 models
are the best characterized ones. The ligation of FasL to FasR
results in sequential recruitment of adaptor molecular FADD and a
FADD associated procaspase-8 to the death receptor to form a
death-inducing signaling complex (DISC), for execution of cell
death (20). Advanced phospho-FADD
stimulates binding of caspase-8 to FADD, leading to cleavage of
procaspase-8, with the consequent generation of active caspase-8.
Active caspase-8 successively amplifies the apoptotic signal
through either direct activation of downstream executioner
caspase-3 or cleavage of Bid (19). Bid, a BH3-only pro-apoptotic Bcl-2
family protein, is generally considered as a molecular linker
bridging between the death receptor pathway and the mitochondria
pathway (21). Uncleaved Bid is
predominantly localized in the cytosol as an inactive precursor.
Upon cleavage by caspase-8, the activated form of Bid (tBid)
migrates to the mitochondria, where it enhances the permeability of
the mitochondrial membrane, thus conveys extrinsic apoptotic
signals from the cytoplasm to the mitochondria (19). In this study, C. amuricus
may trigger caspase-8 through FADD, and then the activated
caspase-8 cleaved Bid into tBid (Fig.
4A), which was transferred to the mitochondria where it may
contribute to the C. amuricus-induced activation of the
intrinsic apoptotic pathway. Therefore, C. amuricus
medicated cleavage of Bid in Hep3B cells may be an important event
for cross-talk between intrinsic and extrinsic pathways.
The tBid translocates to the mitochondria where it
acts with the pro-apoptotic proteins Bax and Bak to initiate the
release of pro-apoptotic factors from the mitochondria. The action
of Bid and Bax is counteracted by the anti-apoptotic Bcl-2 family
members (Bcl-2 and Bcl-xL) which can inhibit mitochondrial
pro-apoptotic events. The Bcl-2 family of proteins, located on the
outer mitochondrial membrane, is vital in apoptotic modulation by
participating in the formation of the pores and altering the
mitochondrial permeability (22).
The balance between pro-apoptotic (Bid, Bad, Bim, Bax, Bak and
Noxa) and anti-apoptotic (Bcl-2, Bcl-xL and Mcl-1) proteins is
critical for determining the release into the cytoplasm of many
apoptogenic factors (cytochrome c), to influence cell
survival (23). Cytochrome
c is a soluble protein located in the mitochondrial
intermembrane space, which functions as an electron carrier of the
mitochondrial respiration chain between complexes III and IV
(24). Once mitochondria senses
the cell death stimuli, cytochrome c from the mitochondria
can be released into cytosol and binds to Apaf-1, forming the
apoptosome-deoxyadenosine triphosphate-dependent complex, which
activates caspase-9, and then drives the downstream caspase cascade
(caspase-3) and cell death mechanisms. Release of cytochrome
c to the cytoplasm is a key step in the initiation of
mitochondrial-dependent apoptosis (24,25).
Consistent with these findings, C. amuricus downregulated
the levels of the anti-apoptotic proteins Bcl-2 and upregulated the
expression of the pro-apoptotic proteins Bak, Bax and Bim in Hep3B
cells in a dose-dependent manner. C. amuricus treatment in
different doses also promoted the release of cytochrome c
and Apaf-1 into the cytosol, which in turn cleaved and activated
caspase-9 (Fig. 4B). Collectively,
C. amuricus-induced apoptosis of Hep3B cells occurs via the
intrinsic mitochondria-mediated pathway, by regulating the
expression of the Bcl-2 family proteins, by enhancing the
expression of cytosolic cytochrome c, Apaf-1, as well as
triggering the activation of caspase-9 and eventually leading to
apoptosis.
As a downstream product of cytochrome c,
caspases are crucial mediators of the principal factors found in
apoptotic cells. Among them, caspase-3 is a frequently activated
death protease, catalyzing the specific cleavage of many cellular
substrates, most notably PARP. PARP is an enzyme involved in DNA
repair, genome surveillance, and genomic integrity maintenance in
response to environmental stress; thus, cleaved PARP is regarded as
another hallmark of apoptosis (19). The cleavage of PARP induces
separation of DNA-binding motifs in NH2 terminal region of
catalytic domains from 116- to 89- and 24 kDa, respectively, during
drug-induced apoptosis in a variety of cells (26). In the present study, C.
amuricus-induced cell death distinctly reduced when Hep3B cells
were pre-treated with caspase inhibitor Z-VAD-fmk, showing that the
activation of caspases is one of the principal mechanisms by which
C. amuricus extract induces apoptosis (Fig. 4C). The rise of the active (cleaved)
forms of caspase-9, caspase-3, caspase-7 and caspase-6 and cleaved
PARP, was further validated in C. amuricus-treated Hep3B
cells in a dose-dependent manner. In addition, both the caspase-3
activation and the subsequent PARP cleavage decreased the
expression of DFF45 and increased the expression of lamin A/C after
treatment of C. amuricus (Fig.
4D). The DFF45 protein is one of the cleavage targets of
caspase-3. Activation of caspase-3 triggers the cleavage of DFF45
(or its isoform DFF35), inactivates its inhibitory function on
DFF40 and causes nuclear DNA degradation by DFF40, leading to cell
death in C. amuricus exposure at different doses (27). Another marker cleaved by caspase-6
by activating caspase-3 is lamin A/C, functioning in cell cycle
control, DNA replication and nuclear membrane structure and
chromatin organization. During apoptosis, lamin A/C is specifically
cleaved into large (41–50 kDa) and small (28 kDa) fragments
(28), and the cleavage of lamin
A/C results in nuclear dysregulation and cell death. Results
indicated that C. amuricus activates caspase-3 and it
cleaves specific target proteins committing Hep3B cells to
apoptosis. The activation of caspases principally amplifies
apoptotic signaling via two distinct pathways: the extrinsic-death
receptor pathway and the intrinsic mitochondrial pathway. In the
present study, the activation of the intrinsic mitochondrial
pathway is involved in the permeabilization of the outer
mitochondrial membrane with subsequent releases of pro-apoptotic
factors, including cytochrome c, into the cytosol. Cytosolic
cytochrome c alters the conformation of the cytosolic
protein Apaf-1, whereas this protein oligomerizes with inactive
procaspase-9, resulting in the activation of this enzyme.
Additionally, the cleavage and activation of the initiator
caspase-8 and −9 occurred after C. amuricus exposure. This,
in turn, leads to the activation of executioner caspase-3, −6 and
−7, ending with the cleavage of several intracellular polypeptides
(PARP-1, DFF45 and lamin A/C) as well as DNA fragmentation, which
provokes the induction of apoptosis (27).
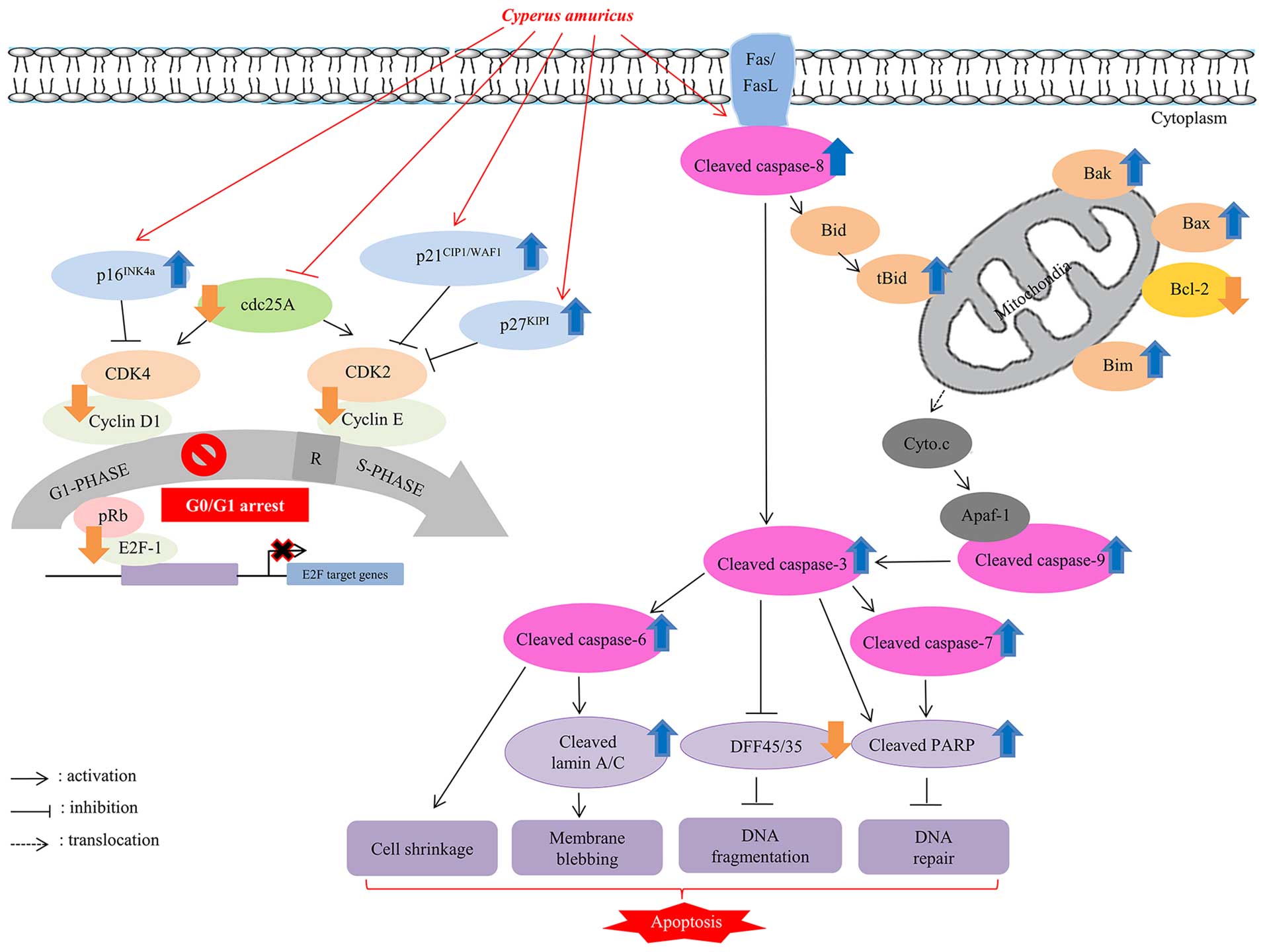
In conclusion, a wide range of anticancer effects of
C. amuricus methanol extract such as cell cycle arrest and
apoptosis on Hep3B cells, a representative p53-null HCC that
contains copies of hepatitis B virus (HBV) genomes in their
chromosomes, and actively secrets HBsAg (29), is illustrated in Fig. 5. This study, therefore, extends the
understanding on the molecular mechanisms underlying the diverse
anticancer activities of C. amuricus on human liver cancer.
However, further studies in animal models are needed to validate
the usefulness of this strategy in vivo. It could be
informative to elucidate the precise mechanism and biological
efficacy of this medicinal herb on biological cellular response in
other cancer types to chemo-sensitization.
Acknowledgements
The present study was supported by a research grant
from the Pukyong National University in year 2016.
Abbreviations:
|
Apaf-1
|
apoptotic protease activating
factor-1
|
|
C. amuricus
|
Cyperus amuricus
|
|
CDKIs
|
cyclin-dependent kinases
inhibitors
|
|
CDKs
|
cyclin-dependent kinases
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
|
DFF
|
DNA fragmentation factor
|
|
DISC
|
death-inducing signaling complex
|
|
DMSO
|
dimethyl sulfoxide
|
|
ECL
|
enhanced chemiluminescent
|
|
FADD
|
Fas-associated death domain
|
|
FBS
|
fetal bovine serum
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
HCC
|
hepatocellular carcinoma
|
|
PARP
|
poly(ADP ribose) polymerase
|
|
PBS
|
phosphate-buffered saline
|
|
Rb
|
retinoblastoma proteins
|
|
tBid
|
truncated Bid
|
|
TNF
|
tumor necrosis factor
|
|
WST-1®
|
2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2Htetrazolium,
monosodium salt
|
References
|
1
|
Llovet JM, Burroughs A and Bruix J:
Hepatocellular carcinoma. Lancet. 362:1907–1917. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
3
|
Rampone B, Schiavone B, Martino A, Viviano
C and Confuorto G: Current management strategy of hepatocellular
carcinoma. World J Gastroenterol. 15:3210–3216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Anthony PP: Hepatocellular carcinoma: An
overview. Histopathology. 39:109–118. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sandal T: Molecular aspects of the
mammalian cell cycle and cancer. Oncologist. 7:73–81. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kögel D, Fulda S and Mittelbronn M:
Therapeutic exploitation of apoptosis and autophagy for
glioblastoma. Anticancer Agents Med Chem. 10:438–449. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sofowora A, Ogunbodede E and Onayade A:
The role and place of medicinal plants in the strategies for
disease prevention. Afr J Tradit Complement Altern Med. 10:210–229.
2013.PubMed/NCBI
|
|
8
|
Maximowicz and Mém: Acad. Imp. Sci.
St.-Pétersbourg Divers Savans 9. Prim Fl Amur. 296:1859.
|
|
9
|
Kakarla L, Allu PR, Rama C and Botlagunta
M: A review on biological and chemical properties of Cyperus
species. Res J Pharm Biol Chem Sci. 5:1142–1155. 2014.
|
|
10
|
Rahmatullah M, Ferdausi D, Mollik AH,
Jahan R, Chowdhury MH and Haque WM: A survey of medicinal plants
used by Kavirajes of Chalna area, Khulna district, Bangladesh. Afr
J Tradit Complement Altern Med. 7:91–97. 2009.PubMed/NCBI
|
|
11
|
Lee SI, Choi H, Jeon H, Baek NI, Kim SH,
Kim HJ, Cho CH, Ahn HC, Yang JH, Chae BS, et al: Antioxidant
phenolic components from the whole plant extract of Cyperus
amuricus Max. Korean J Pharmacogn. 39:233–236. 2008.
|
|
12
|
Sharma N, Sharma VK and Seo SY: Screening
of some medicinal plants for anti-lipase activity. J
Ethnopharmacol. 97:453–456. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nicholson DW and Thornberry NA: Caspases:
Killer proteases. Trends Biochem Sci. 22:299–306. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ayed-Boussema I, Bouaziz C, Rjiba K,
Valenti K, Laporte F, Bacha H and Hassen W: The mycotoxin
Zearalenone induces apoptosis in human hepatocytes (HepG2) via
p53-dependent mitochondrial signaling pathway. Toxicol In Vitro.
22:1671–1680. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hartwell LH and Kastan MB: Cell cycle
control and cancer. Science. 266:1821–1828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morgan DO: Cyclin-dependent kinases:
Engines, clocks, and microprocessors. Annu Rev Cell Dev Biol.
13:261–291. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Knudsen ES and Knudsen KE: Tailoring to
RB: Tumour suppressor status and therapeutic response. Nat Rev
Cancer. 8:714–724. 2008. View Article : Google Scholar
|
|
18
|
Li W, Sanki A, Karim RZ, Thompson JF, Soon
Lee C, Zhuang L, McCarthy SW and Scolyer RA: The role of cell cycle
regulatory proteins in the pathogenesis of melanoma. Pathology.
38:287–301. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Ao M, Wu J and Yu L: TNFα and
Fas/FasL pathways are involved in 9-Methoxycamptothecin-induced
apoptosis in cancer cells with oxidative stress and G2/M cell cycle
arrest. Food Chem Toxicol. 55:396–410. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Breckenridge DG and Xue D: Regulation of
mitochondrial membrane permeabilization by BCL-2 family proteins
and caspases. Curr Opin Cell Biol. 16:647–652. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Portt L, Norman G, Clapp C, Greenwood M
and Greenwood MT: Anti-apoptosis and cell survival: A review.
Biochim Biophys Acta. 1813:238–259. 2011. View Article : Google Scholar
|
|
23
|
Leibowitz B and Yu J: Mitochondrial
signaling in cell death via the Bcl-2 family. Cancer Biol Ther.
9:417–422. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Abu-Qare AW and Abou-Donia MB: Biomarkers
of apoptosis: Release of cytochrome c, activation of caspase-3,
induction of 8-hydroxy-2′-deoxyguanosine, increased
3-nitrotyrosine, and alteration of p53 gene. J Toxicol Environ
Health B Crit Rev. 4:313–332. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Green DR: Apoptotic pathways: Ten minutes
to dead. Cell. 121:671–674. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boulares AH, Yakovlev AG, Ivanova V,
Stoica BA, Wang G, Iyer S and Smulson M: Role of poly(ADP-ribose)
polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP
mutant increases rates of apoptosis in transfected cells. J Biol
Chem. 274:22932–22940. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ozören N and El-Deiry WS: Cell surface
death receptor signaling in normal and cancer cells. Semin Cancer
Biol. 13:135–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lazebnik YA, Takahashi A, Moir RD, Goldman
RD, Poirier GG, Kaufmann SH and Earnshaw WC: Studies of the lamin
proteinase reveal multiple parallel biochemical pathways during
apoptotic execution. Proc Natl Acad Sci USA. 92:9042–9046. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Twist EM, Clark HF, Aden DP, Knowles BB
and Plotkin SA: Integration pattern of hepatitis B virus DNA
sequences in human hepatoma cell lines. J Virol. 37:239–243.
1981.PubMed/NCBI
|