Introduction
As a high risk cancer, esophageal cancer possesses
several adverse characteristics including difficult early
diagnosis, rapid tumor growth and metastasis, relatively low
response to drug treatment and poor 5-years survival. The major
type of esophageal cancer in Asian countries is esophageal squamous
cell carcinoma (ESCC), while esophageal adenocarcinoma occupies
predominant cancer type in Western countries. Although great
efforts have been made to explore the mechanisms of epigenetic or
genetic modifications occurred during esophageal tumorigenesis, the
molecular biology of esophageal cancer is still too complicated to
understand throughly. Research on esophageal squamous cancer
indicated that activation of oncogenes and impaired signaling
pathways, such as c-Myc, VEGF and TGF-β (1), actively participated in the
development of esophageal cancer, and correspondingly molecular
treatments targeted to abnormal signaling pathways were widely
designed and tested.
Bone morphogenetic proteins (BMP), as the members of
transforming growth factor-β (TGF-β) superfamily, were initially
described for osteoinductive potentiality. During human
development, BMPs also take part in many biological activities
including cell proliferation, migration, apoptosis and
differentiation (2). As secretory
proteins, BMPs exert functions by coupling with receptors. There
are two groups of BMP receptors, named type I and II receptors of
serine/threonine kinase. Type I receptors contain activin
receptor-like kinase-2, -3 and -6 (ALK-2, ALK3 and ALK-6), while
type II receptors include BMPRII, ActRIIA and ActRIIB. By binding
to type II receptor and subsequently activated type I receptor,
BMPs transduce signaling via phosphorylating SMAD1/5/8 proteins,
which further coordinate with co-SMAD4 protein to activate target
downstream genes. Except their indispensable roles in normal
development, BMPs and their receptors were extensively found
abnormally expressed in carcinomas, such as prostate, breast and
ovarian cancer (3–5). Studies of those cancers indicated
that malfunctions of BMP signaling pathways were more or less
responsible for cancer progress, including proliferation, invasion
and metastasis and protected cells from death. BMP2 and BMP4 are
different from BMP6 and BMP7 based on their structural sequence
similarity. BMP2 and BMP4 share 92% similar amino acid sequence,
while BMP6 and BMP7 can be categorized as a subgroup (6,7).
BMP2 was found related with metastasis in gastric cancer and
melanoma (8,9), proliferation in prostate and lung
cancer (10,11), while BMP4 was reported to induce
epithelial-to-mesenchymal transition (EMT) in ovarian and
pancreatic cancer (12,13). BMP6 showed its roles in promoting
E-cadherin expression and potential function in breast cancer
metastasis (3). BMP7 exhibited its
contradictory roles in promoting EMT in melanoma and related with
metastasis in breast cancer or inhibiting TGF-β-induced EMT in
esophageal adenocarcinoma (1,14,15).
SMAD-mediated BMP signaling pathway predominates in most BMP
involved activities, but other signaling pathways have also been
reported to be employed or cross-linked with BMPs, such as
PI3K/AKT, MAPK/ERK/p38, WNT/β-catenin and TGF-β (16–18).
The flexible usage of BMP receptors and comprehensive signaling
pathways may explain the conflicting results of BMP functions in
cancers. Recently, Yuen et al (19) found BMP6 expression widely in
clinical esophageal cancer samples and strong BMP6 expression
associated with faint BMP inhibitor Noggin expression was
correlated with shorter survival. BMP7 expression was also reported
by Megumi et al (20) to
predict poor prognosis of esophageal squamous cells. However, the
roles of BMPs in esophageal cancer and underlying mechanisms need
to be elucidated for further understanding the value of BMPs in
esophageal cancer therapy.
In the present study, we detected BMPs and their
receptor expression in ESCC cell lines and explored whether
overexpression of BMP2, 4, 6 and 7 would affect ESCC cell
proliferation, invasion and metastasis. Distinct influence on
invasion and metastasis exhibited by BMP2/4 and BMP6/7 were
attributed to different signal pathways. BMP6/7 promoted ECA-109
cell invasion and metastasis through ALK2 and BMPRII-mediated
classical SMAD1/5/8-dependent pathways while BMP2/4 reduced
invasive and migratory properties of ECA-109 without effective
activation of SMAD1/5/8 pathways but simultaneously upregulated
phosphorylated P-38, ERK and JNK. Classical BMP signal pathway
interference by DN-ALK2 or DN-BMPRII significantly downregulated
ECA-109 cell growth, survival, invasion and metastasis. Clinical
ESCC patient samples showed different expression pattern of BMP6,
ALK2 and BMPRII, but tumors with triple-positive expression of the
proteins had deeper invasion. Taken together, this study indicates
that BMP6/7 promotes invasion and metastasis of esophageal squamous
cancer through ALK2-SMAD pathways, which can serve as potential
targets for esophageal squamous cancer therapy.
Materials and methods
Cell culture and materials
Esophageal squamous cancer cell lines ECA-109,
KYSE150 and KYSE180 were maintained in RPMI-1640 medium (HyClone
Laboratories, Inc., Logan, UT, USA) supplemented with 10% fetal
bovine serum (FBS; HyClone Laboratories). HEK293 cells used for
adenovirus amplification and HCT-116 cells used for
adenovirus-expression protein condition medium were cultured in
Dulbecco's modified Eagle's medium (DMEM) with 10% FBS. All cells
were maintained with 1% penicillin/streptomycin in 5%
CO2 at 37°C. Adenovirus expressing BMP2, BMP4, BMP6,
BMP7 and Noggin or dominant-negative mutants of ALK2 and BMPRII
were generously offered by professor Tong-Chuan He (Chicago
University, Chicago, IL, USA). Ad-GFP served as negative control
for the adenovirus involved experiments. Viral titers were
determined by infecting 293 cells with gradient dilutions of
adenovirus before use. Adenovirus of pfu up to 108 were
applied, respectively.
RNA extraction and PCR
Total RNA from esophageal cancer cells were obtained
by TRIzol reagent (Invitrogen) and chloroform-isopropanol
extraction according to the protocol provided by the manufacturer.
RNA quantity and integrity were evaluated by NanoDrop 1000
spectrophoto-meter (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and denaturing gel electrophoresis. Two nanogram of total RNA
was reverse transcribed and synthesized into cDNA by PrimeScript
kit (Takara). Primers used to detect different gene expression and
PCR conditions are listed in Table
I with GAPDH used as internal control. The analysis of PCR
products was performed on Quantity One software (Bio-Rad
Laboratories, Hercules, CA, USA).
 | Table IPCR primers for BMPs and BMP
receptors. |
Table I
PCR primers for BMPs and BMP
receptors.
| Primer | Sequence | Temperature (°C) | Product length
(bp) |
|---|
| ALK1 |
GGCTCCCTCTACGACTTTCT | 57 | 137 |
|
TGGGCAATGGCTGGTTT | | |
| ALK2 |
AGGATTACAAGCCACCG | 52 | 163 |
|
TACCAGCATTCTTTCATTAG | | |
| ALK3 |
TCTTGGAGGAGTCGTAA | 50 | 181 |
|
GTAAATGTATAGCTGAGGC | | |
| ALK6 |
AAATGTGGGCACCAAGAAAGA | 55 | 171 |
|
ACAGGCAACCCAGAGTCATC | | |
| BMP2 |
CCTACATGCTAGACCTGTATCGCA | 52 | 362 |
|
CACCAACCTGGTGTCCAAAAGT | | |
| BMP4 |
CAGCACTGGTCTTGAGTATCCTGA | 52 | 307 |
|
CGTGTCCAGTAGTCGTGTGATGA | | |
| BMP6 |
CGACAACAGAGTCGTAATCG | 52 | 195 |
|
GCATTCTCCATCACAGTAATTG | | |
| BMP7 |
ACGCTTCGACAATGAGACGTTC | 52 | 572 |
|
TGGCGTTCATGTAGGAGTTCAG | | |
| BMPRII |
AAATAGCCTGGCAGTGAG | 53 | 196 |
|
ATGTGACAGGTTGCGTTC | | |
| ActRIIA |
GAAGATGAGGCCCACCATTA | 52 | 179 |
|
GACAGAGGTCACCAGGGAAA | | |
| ActRIIB |
AAACCTGCCATATCTCAC | 52 | 199 |
|
GCACCCTCTAATACCTCT | | |
| ID1 |
CGGTCTCATTTCTTCTCG | 57 | 182 |
|
TCGGTCTTGTTCTCCCTC | | |
| GAPDH |
CAGCGACACCCACTCCTC | 52 | 120 |
|
TGAGGTCCACCACCCTGT | | |
Western blot analysis
After different infection of Ad-BMPs for 24 h, ESCC
cells were serum-starved for 24 h and RIPA cell lysis buffer
(Biyuntian Biotechnology Co., Ltd., Shanghai, China) supplemented
with 1 mmol PMSF was added. After centrifuged at 1,3000 × g, at 4°C
for 30 min, protein concentration was determined by NanoDrop 1000
spectrophotometer (Thermo Fisher Scientific). A total of 50
µg protein samples were loaded and resolved by 10% SDS-PAGE.
Then protein bands were transferred onto polyvinylidene difluoride
membranes (PVDF; Millipore, Billerica, MA, USA). Primary antibody
was diluted in TBST buffer (dilution as 1:1,000) and applied on the
membrane to incubate at 4°C overnight. Horseradish
peroxidase-conjugated secondary antibody (Zhongshan Golden Bridge
Biotechnology Co., Ltd., Beijing, China) was incubated with PVDF
membrane for 1 h at 37°C next day after rinsing by TBST buffer.
Then the membrane was extensively washed by TBST and protein bands
visualized using chemiluminescence reagent (Pierce) in ChemiDoc™
XRS+ System (Bio-Rad Laboratories). Primary anti-p-Smad1/5/8
(1/5-S453/465 and 8-S462/428), anti-p-Smad1 (S463/465),
anti-T-Smad1, anti-p-JNK (T183/185), anti-JNK and anti-p-P38
(T188/Y182) were purchased from Cell Singaling Technology (Danvers,
MA, USA). Internal control anti-β-actin was purchased from Santa
Cruz Biotechnology (Santa Cruz, CA, USA).
Cell proliferation assay
Cell proliferation was performed by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT;
Promega, Madison, WI, USA) colorimetric assay. Briefly, esophageal
cancer cells were infected by different adenovirus for 24 h and
then seeded into 96-well plate at 5,000 cells/well. After cells
tightly attached, MTT reagent was added to detect cell viability as
0 h time-point or at 24-h interval. For each well, 20 µl 5
mg/ml MTT reagent was added and incubated at 37°C for 4 h. DMSO was
used to dissolve formazan precipitation at the end of incubation
and then optical density of each well was measured at 490 nm by a
plate reader (Sunrise Remote; Tecan Group Ltd., Männedorf,
Switzerland). Each sample contained at least 5-wells and
experiments were performed three times.
Invasion and migration assay
To determine ESCC cell invasion and migration,
Transwell migration chambers (Corning Incorporated, Corning, NY,
USA) with 8 µm pores were used. After adenovirus infection
for 24 h, cells were trypsinized and seeded at 5×105 in
duplicate in the upper chamber of Transwell with or without
Matrigel (Invitrogen). Lower chamber of 24-well plate contained 20%
FBS. After allowing cell migration for 24 h, the filters of
Transwell were dried, fixed and stained by hematoxylin and eosin
(H&E) or crystal violet. Ten random high power fields with
evenly distributing cells were chosen to count invaded cells. All
experiments were performed at least three times.
Transfection and SMAD-responsive alkaline
phosphatase activity detection
Alkaline phosphatase reporter plasmid pSEAP2
(Clontech Laboratories, Inc., Mountain View, CA, USA) containing
two BMP response elements (BRE) was constructed previously, named
as pSEAP2-BRE. pSEAP2-BRE was transfected into ECA-109 cells using
Lipofectamine 2000 reagent (Invitrogen) with conditional medium of
GFP or BMPs produced by the HEK293 cells. Supernatant of cells was
extracted after transfection for 24 h and alkaline phosphatase
activity was detected by the SEAP chemiluminescence.
Colony formation assay
After ECA-109 infection with AdDN-ALK2 or
AdDN-BMPRII for 24 h, low density cells were seeded in 12-well
plate by 100 cells/well. At least three wells were employed for
each group. ECA-109 and ECA-109 infected by AdGFP served as
control. Then cells were incubated in 37°C for a week. The plate
was washed gently by phosphate-buffered saline (PBS) to remove dead
cells and then stained by crystal violet.
ESCC tumor samples
ESCC patients enrolled in this study were the same
cohort as a previous report (21).
All samples, including tumor tissues and non-cancerous tissues,
were collected and kept in liquid nitrogen until RNA extraction.
The patient clinical histopathological profiles were also collected
and classified according to the differentiation status, TMN stage
and tumor depth, which contain shallow tumor invasion within the
mucosa and deep submucosal invasion.
Statistical analysis
Statistical analysis was performed using the SPSS
19.0 (IBM Corp., Armonk, NY, USA). The significance of the
differences in RNA and protein expression between control group and
BMPs groups was analyzed by independent sample t-test. Clinical
histopathological parameters between the two groups were performed
by the Fisher's exact test or the χ2 test. P<0.05 was
considered statistically significant in all tests.
Results
BMPs and their receptor expression in
ESCC cell lines
To evaluate the functions of BMP signaling pathways
in ESCC, RT-PCR analysis to detect mRNA transcripts for BMP ligands
(BMP2, 4, 6 and 7), type I receptors (ALK1, 2, 3 and 6) and type II
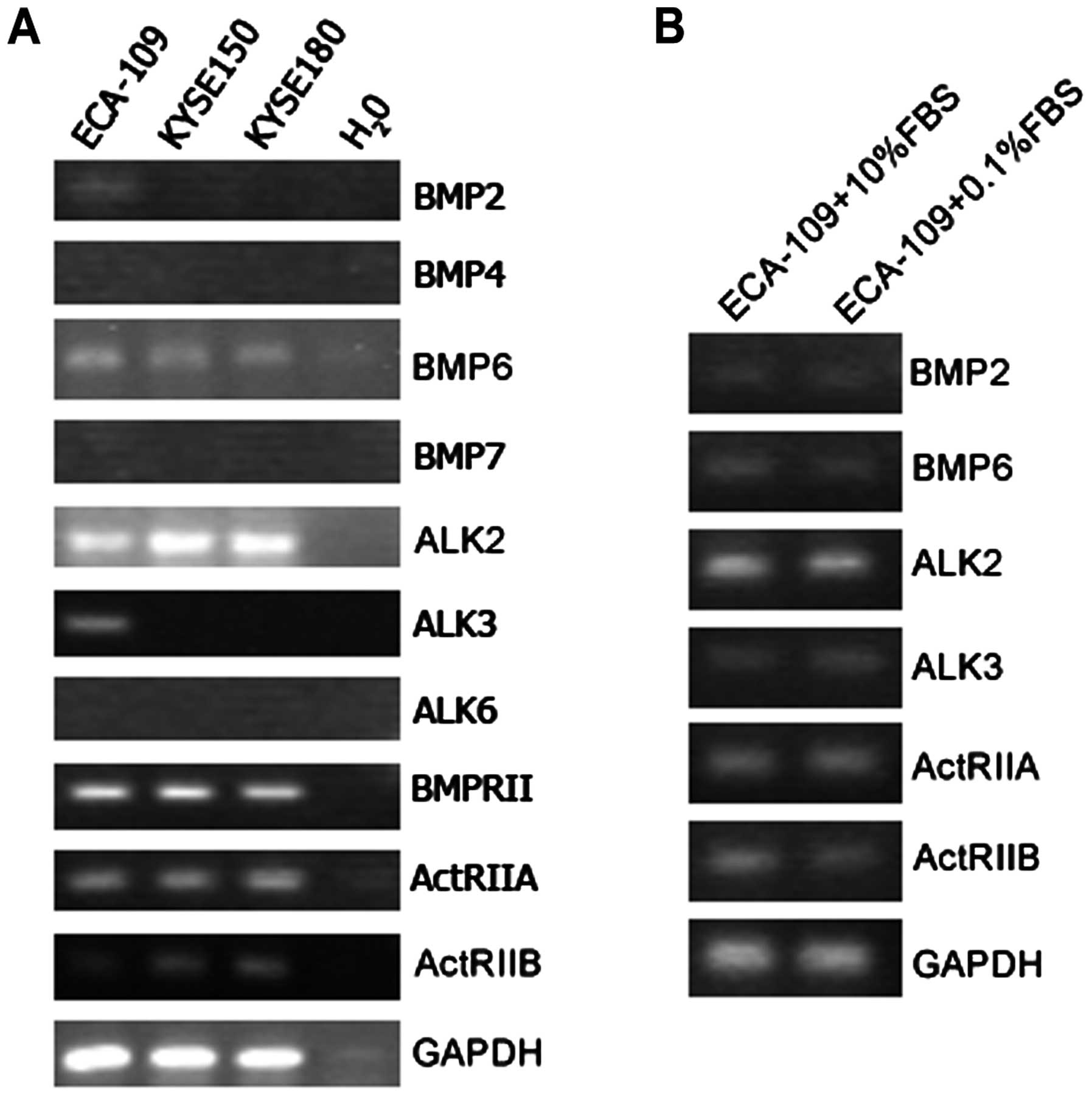
receptors (BMPRII, ActRIIA and ActRIIB) was performed. As shown in
Fig. 1A, transcripts of ALK2,
BMPRII, ActRIIA and ActRIIB were positive in all three ESCC cell
lines, ECA-109, KYSE150 and KYSE180. None of them expressed type I
receptor ALK6. BMP6 expression was detected in all these cell
lines, consistent with the previous report of Yuen et al
(19) that BMP6 can be detected in
most of esophageal squamous cells and clinical esophageal cancer
tissues. We also found weak expression of BMP2 in ECA-109 cells.
However, BMP4 and BMP7 showed negative expression in three ESCC
cell lines.
In some reports, serum concentration is seemingly an
important factor which greatly affects BMPs expression and their
functions. To address this question, we carried out RT-PCR to
detect BMPs and their receptors expression levels under the
condition of 10 or 0.1% FBS supplement for 36 h. No detectable
difference was observed in the expression of BMP2, BMP6, ALK2,
ALK3, BMPRII, ActRIIA and ActRIIB in ECA-109 cells (Fig. 1B).
Overexpression of BMPs has no effect on
cell proliferation but differentially affect invasion and
metastasis
Expression of BMP receptors in esophageal cancer
cell lines lays the foundation that BMP signaling pathway may be
potentially viable in ESCC cells. Therefore, different BMPs were
overexpressed in ECA-109 cells, respectively, to evaluate their
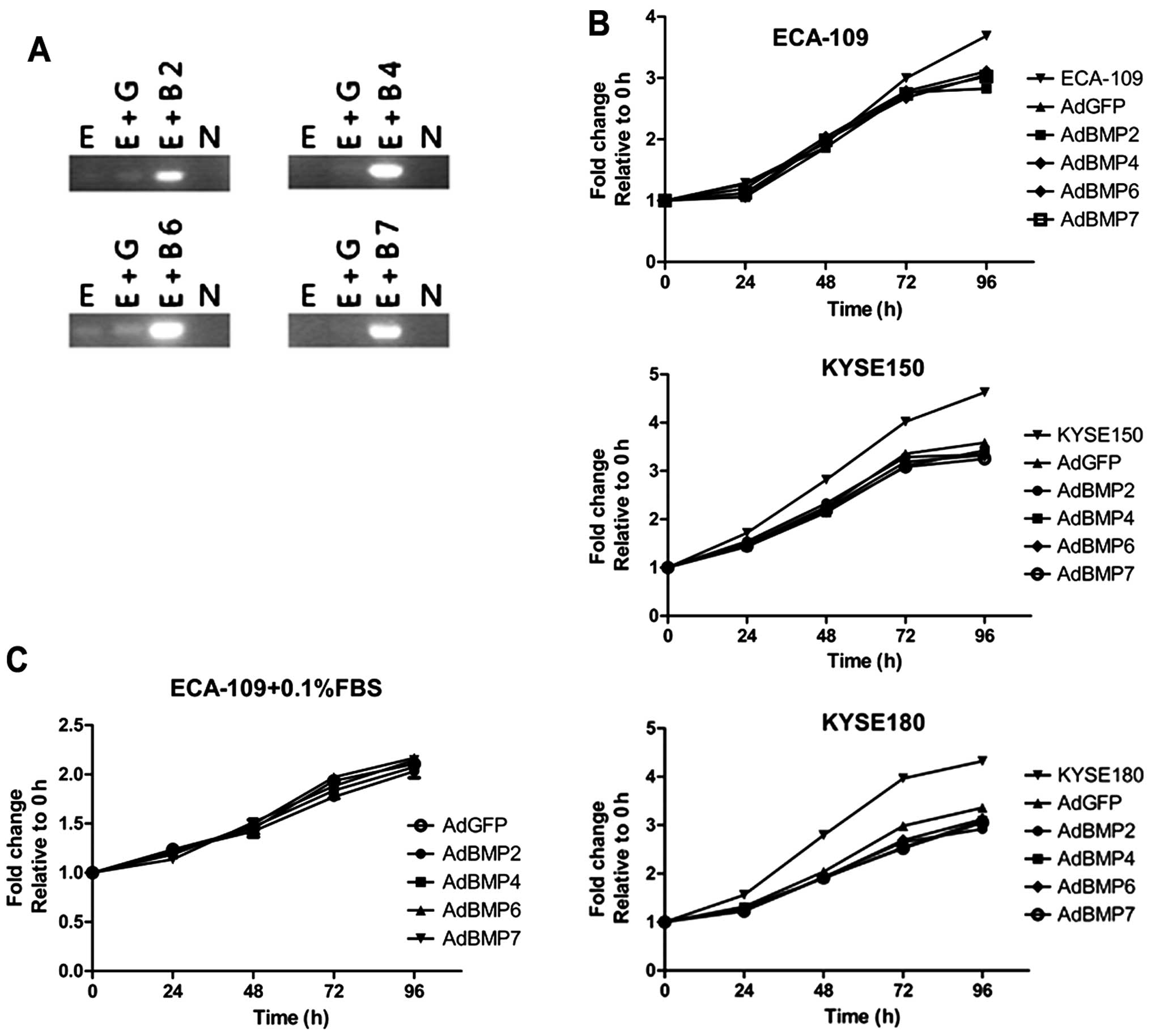
impacts on cells proliferation (Fig.
2A). Compared with GFP control group, none of the BMP
overexpression changed the proliferation rates of ESCC cells, while
adenovirus-loading effect retarded cell growth relatively (Fig. 2B). Besides, although serum
deprivation (0.1% FBS) slightly depressed ECA-109 proliferation, no
difference was observed among BMP-overexpressing groups and control
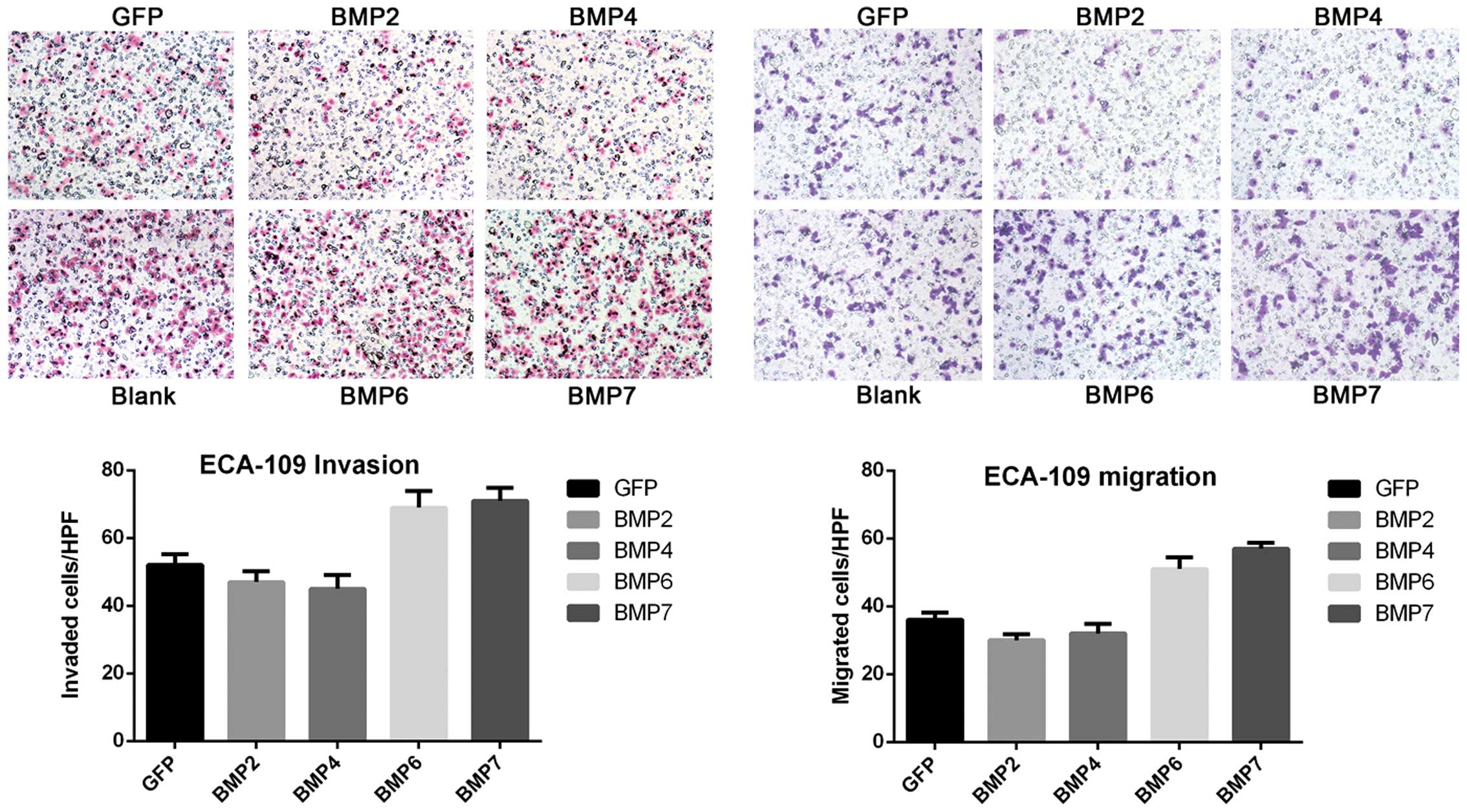
group (Fig. 2C). Many reports have
indicated that BMPs extensively participate in cancer-related
invasion and metastasis. Therefore, the impact of BMPs on invasion
and metastasis of ESCC cells was determined by Transwell chamber
assay. Surprisingly, not only overexpression of BMPs significantly
influenced ESCC cell invasion and metastasis, but different BMPs
showed distinct effects. BMP2/4 expression led to >20% reduction
of invasion and migration, while BMP6/7 overexpression somehow
increased ECA-109 cell invasion and migration by 10–20% (Fig. 3). Similar results were also found
in KYSE150 and KYSE180 cell lines (Table II).
| Table IITranswell assay with or without
Matrigel (mean cell numbers/HPS). |
Table II
Transwell assay with or without
Matrigel (mean cell numbers/HPS).
| Matrigel | GFP | BMP2 | BMP4 | BMP6 | BMP7 |
|---|
| ECA-109 | − | 52 | 47 | 45 | 69 | 71 |
| + | 36 | 30 | 32 | 51 | 57 |
| KYSE150 | − | 69 | 21 | 20 | 55 | 47 |
| + | 33 | 26 | 29 | 41 | 45 |
| KYSE180 | − | 57 | 49 | 45 | 73 | 66 |
| + | 31 | 24 | 28 | 47 | 45 |
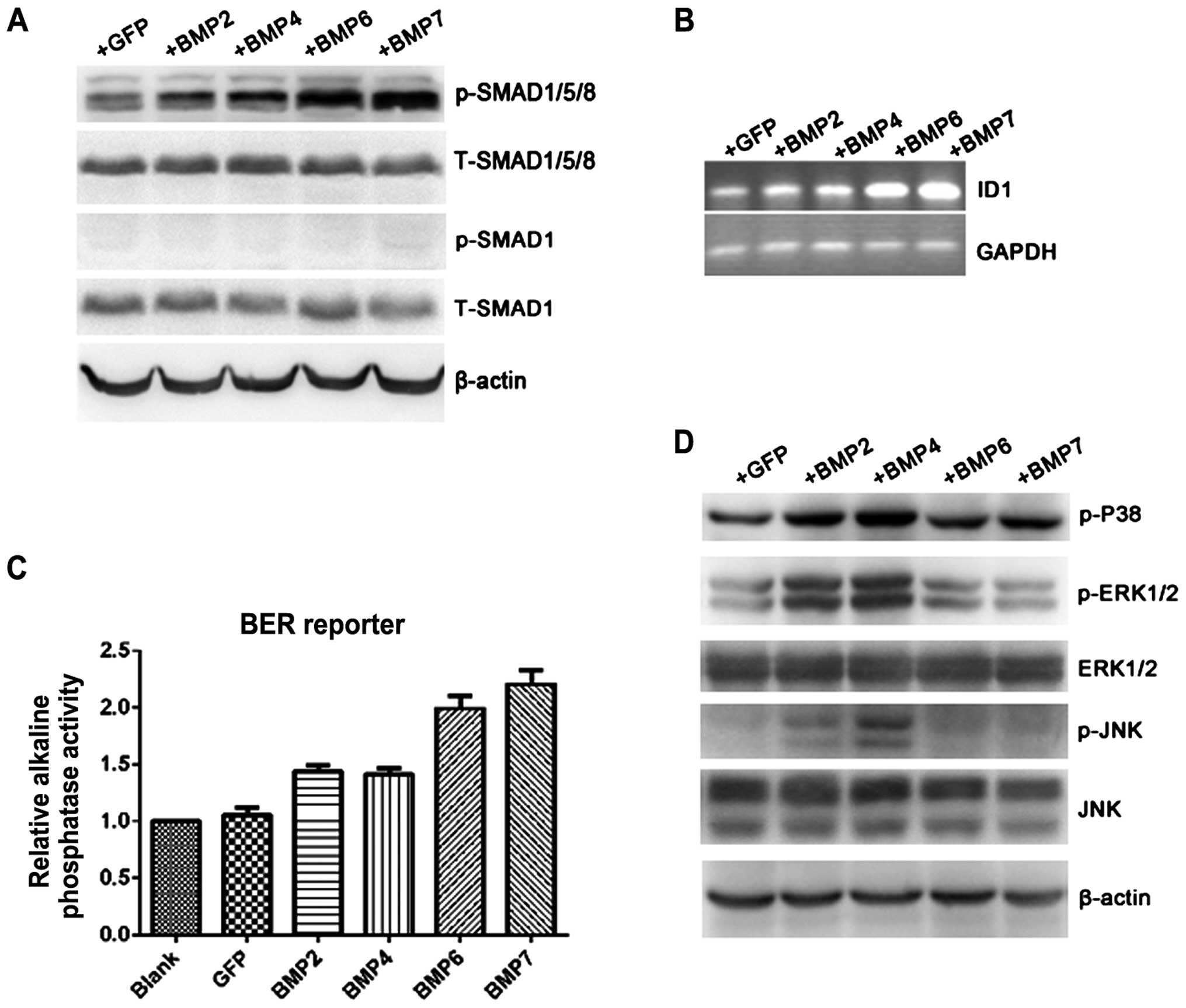
BMPs can differently activate Smad1/5/8
signal pathway
Most of BMPs exert their functions by activating
classical Smad1/5/8-mediated pathway which lead to transcriptional
activation of downstream genes. Total protein was extracted under
different BMP overexpression conditions to detect the activation of
various signal pathways by western blot analysis. Compared with
control, the phosphorylated form Smad1/5/8 was significantly
induced under BMP6 (1.39-fold) and BMP7 (1.56-fold) stimulation
while slight increment can be seen in BMP2 (1.18-fold) and BMP4
(1.22-fold) groups. Total Smad1/5/8 showed equivalent expression in
all groups. There was no detectable expression of p-Smad1 in
ECA-109 cells treated either by Ad-GFP or Ad-BMPs, although total
Smad1 can be clearly and prevalently observed (Fig. 4A). If BMP-Smad1/5/8 signal pathway
is functionally transduced, downstream target genes containing BMP
response element in promoter region can be transcriptionally
activated accordingly. ID1, as a well-known BMP-target gene, was
detected by PCR after BMP overexpression in ECA-109 for 48 h. ID1
mRNA was significantly upregulated by treatment with BMP6
(2.12-fold) or BMP7 (2.17-fold) but slightly increased in BMP2 and
BMP4 overexpressing cells (Fig.
4B). BMP response element (BRE) alkaline phosphatase reporter
was then employed to verify Smad-dependent signaling activation.
Alkaline phosphatase activity was obviously high in BMP6/7-treated
cells (1.9- and 2.2-fold, respectively) compared with control, but
mildly increased in BMP2/4-treated cells (1.38- and 1.4-fold;
Fig. 4C). Those results indicated
that BMP6/7 can significantly activate the downstream signaling
pathway in Smad-dependent manner.
BMP2/4 can activate MAPK and JNK
pathways
In addition to classical SMAD1/5/8 pathway, BMPs
have been reported employing other signaling pathways to exert
different functions in cell biology along with embryo development.
Therefore, we explored whether the phenomena also existed in ESCC
cells. As shown in Fig. 4D,
phosphorylated P-38, ERK1/2 and JNK were obviously upregulated in
BMP2- and BMP4-treated groups, but not in BMP6- and BMP7-treated
groups.
ALK2 and BMPRII participates in
BMP6/7-induced cell invasion and metastasis
Because of sequence homology, BMP6 and BMP7 have
similar preference to employ type I receptor ALK2 and type II
receptor BMPRII for signal transduction. The classical
Smad-mediated pathway activation, and coincidently increased cell
invasion as well as metastatic properties prompted us to detect
whether ALK2 and BMPRII involved in BMP6/7 overexpression-related
events. Adenovirus expressing DN-ALK2 and DN-BMPRII were used,
respectively. As demonstrated in Fig.
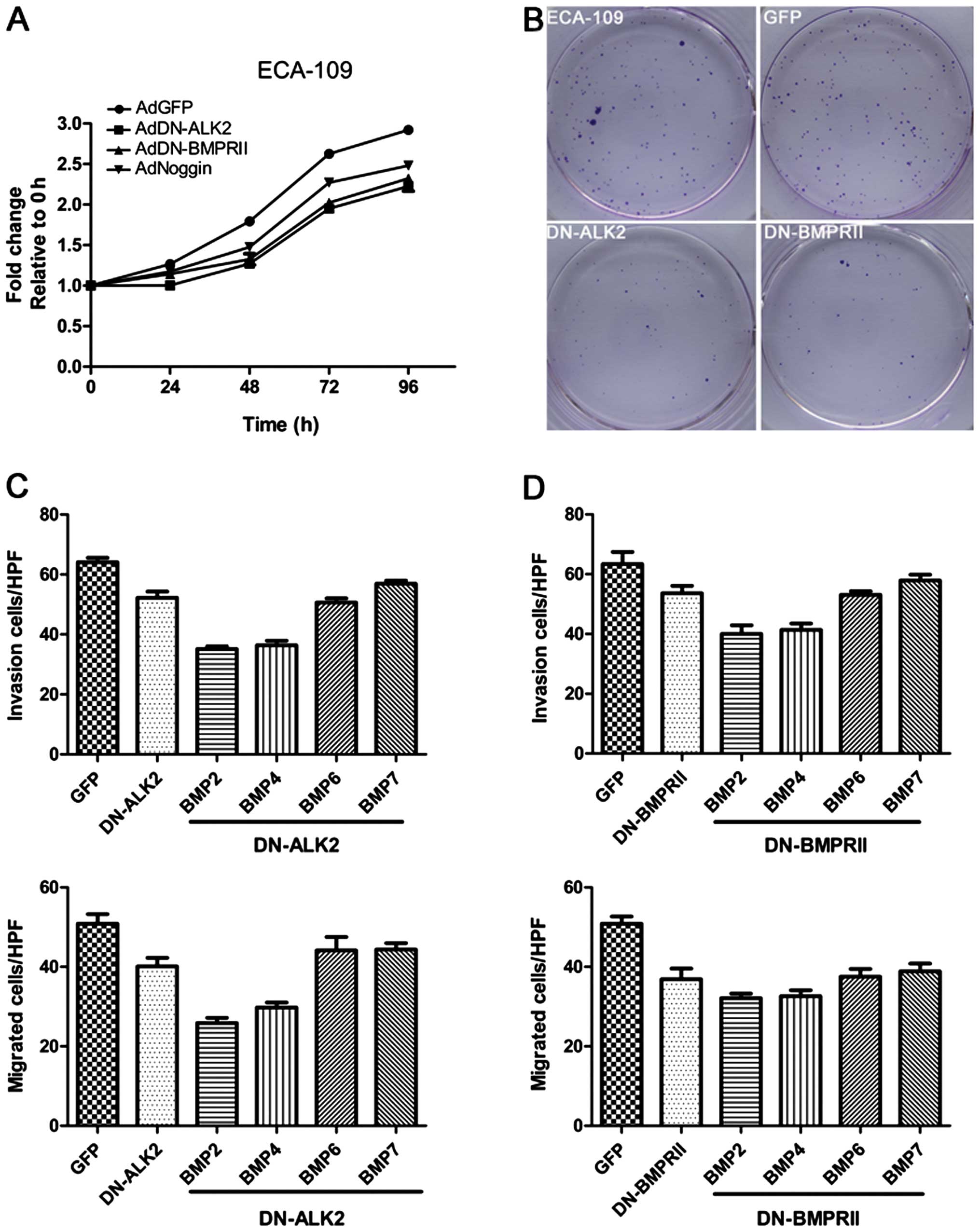
5A, DN-ALK2 or DN-BMPRII supplement by condition medium can
significantly inhibit cell proliferation by ~15–20% in
serum-containing medium (10% FBS). BMP inhibitor Noggin served as
positive control which decreased cell growth by 10%. Colony
survival assay also demonstrated that ECA-109 cells containing
DN-ALK2 or DN-BMPRII exhibited slower growth and less colony
formation (Fig. 5B). Then,
BMP-overexpressing esophageal cancer cell invasion and metastasis
were tested with or without DN-ALK2 or DN-BMPRII interference. As
showed in Fig. 5C and D, relative
to control, DN-ALK2 or DN-BMPRII expressing cells showed markedly
reduced cell number invaded or migrated through Transwell chamber
membranes and further halted the increment of invasive and
migratory properties induced by BMP6/7 overexpression. However,
DN-ALK2 or DN-BMPRII hardly altered the negative effects of BMP2/4
overexpression on invasion and metastasis.
ALK2, BMPRII and BMP6 have different
expression pattern in ESCC patients
We used RT-PCR to detect ALK2, BMPRII and BMP6 mRNA
expression, respectively, in ESCC tumor samples and adjacent
non-cancerous tissues (n=50). Compared to the respective
non-cancerous tissue, BMP6 overexpression was broadly observed in
40 out of 50 samples (90%), while 17 samples showed positive ALK2
(34%) and 15 samples showed BMPRII expression (30%). Only 7 samples
have triple-expression of ALK2, BMPRII and BMP6. Analyzing
expression pattern and clinical histopathological features between
BMP6+ALK2+BMPRII+ group and
BMP6+ALK2-BMPRII− samples indicated that
positive ALK2 and BMPRII expression contributed to deeper tumor
invasion on the background BMP6 expression (Table III), although no effect on tumor
differentiation status and lymphatic invasion was observed.
| Table IIIClinical characteristics and BMP2,
ALK2 and BMPRII expression pattern. |
Table III
Clinical characteristics and BMP2,
ALK2 and BMPRII expression pattern.
| Clinical
histopathological parameters |
BMP6+ALK2+BMPRII+
(n=7) |
BMP6+ALK2−BMPRII−
(n=26) | P-value |
|---|
| Histopathologic
grading | | | 0.244 |
| Well
differentiation | 1 | 11 | |
| Moderate
differentiation | 3 | 9 | |
| Poor
differentiation | 3 | 6 | |
| Lymphatic
invasion | | | 0.080 |
| Negative | 2 | 17 | |
| Positive | 5 | 9 | |
| Tumor depth | | | 0.022a |
| Shallow | 3 | 22 | |
| Deep | 4 | 4 | |
Discussion
BMPs family contains more than twenty identified
members, which are widely involved in multiply biological
processes, especially in bone formation and embryo development. For
decades, the diverse roles of BMPs in tumor cell growth, invasion,
metastasis and angiogenesis have been extensively studied. BMPs and
BMP receptors were found in many cancers, including prostate,
breast, ovarian, melanoma, lung and esophageal cancer (22–24).
Recently, Yuen et al (19)
found that BMP6 expression can be detected in the majority of
esophageal squamous cancer cell lines and clinical esophageal
cancer tissues correlated with tumor progression and prognosis
along with BMP antagonist expression level. In this study, BMP6
expression in all three esophageal squamous cancer cell lines and
BMP type I and type II receptors can also be detected, indicating
that BMP signal would be potentially functional in esophageal
cancer cells. BMP7 expression has also been detected in more than
half of clinical esophageal squamous cell carcinoma samples and
patients with BMP7 expression had deeper invasion and poorer
prognosis (20). We did not find
any BMP7 expression in esophageal squamous carcinoma cell lines and
in clinical ESCC tumors. To investigate the roles BMPs played in
esophageal squamous carcinoma, overexpression of different BMPs by
adenovirus-based gene expressing were carried out in ECA-109 cells.
Although many reports have demonstrated that upregulation of BMPs
can greatly affect cancer cell proliferation such as prostate,
breast and lung cancer, overexpression of BMP2, 4, 6 and 7 in this
study did not affect proliferation rate. One possible reason is
that previous reports always employed recombinant human BMP
proteins at relatively high concentrations. Such as in the study of
Kim et al (25), they found
that exogenous recombinant human BMP2 could decrease ESCC cell
proliferation in vitro and reduce tumor growth volume in
vivo, possibly through Hippo signaling pathways, in which the
concentration of rhBMP2 ranged from 10 nM to 10 µM and 10
µM was used throughout the research. Kokorina et al
(26) also utilized 100 ng/ml
rhBMP2 to test oral carcinoma cell invasive ability. Several
reports showed that the concentration of FBS can greatly affect
tumor cell proliferation or growth suppression under BMP supplement
conditions. According to this, conflicting results were observed
among BMP research on the same tumor type. However, no difference
of growth rate or alteration of BMPs and their receptors expression
can be found cultured with 10 or 0.1% FBS here. The effects of
serum concentration on BMPs in different cancers provided a notion
that the conclusion of the roles of BMPs in cancer should be made
depending on the cancer type itself. Therefore, it is possible that
the influence of serum concentration is too delicate to be
detectable in esophageal squamous carcinoma.
Although the investigation on esophageal squamous
tumor clinical samples showed inconsistent upregulation of BMP6,
ALK2 was prevalently detected in both esophageal cancer tissues
(34%) and in esophageal cancer cell lines. ALK2 expression may
confer a favorable condition for BMP6 binding and promoting
esophageal cancer cell motivation. As indicated in other research
that BMPs exert biological functions by classical SMAD-mediated
pathway, we found that BMP6/7 overexpression significantly
activated Smad1/5/8 signaling pathways and then transcriptionally
upregulated ID1 expression and BRE-driven alkaline phosphatase
reporter activity. Classical Smad-dependent pathway activation has
been implicated involved in BMP-induced invasion and metastasis
(27,28). ID1 transcriptional activation alone
or induced by BMPs was found related with increased cancer
aggressiveness (18,29–31).
Furthermore, interference of type I receptor ALK2 or type II
receptor BMPRII by dominant negative mutants decreased cancer cells
invasion or migration through membrane with or without Matrigel.
Besides, ALK2 interference or BMP inhibitor Noggin supplement also
reduced cell growth and long period clonal survival ability.
Combined with the report by Yuen et al (29) that widely expressed BMP6 and low
level of BMP antagonists were demonstrated in esophageal cancer,
our results suggested that esophageal squamous cancer cell may
acquire favorable cell proliferation partly through a BMP6-ALK2
autocrine manner, similar to BMP9 in ovarian cancer.
Notably, BMP2 and BMP4 overexpression somehow
reduced cell invasion and metastasis. Compared to BMP6 and BMP7,
BMP2 and BMP4 induced less Smad1/5/8 phosphorylation and
correspondingly lower BRE alkaline phosphatase reporter activation
and ID1 mRNA expression. Additionally, DN-ALK2 or DN-BMPRII exerted
much less effect on BMP2/4 related invasive and migratory events
than BMP6/7 exhibited, indicating that BMP-SMAD pathway may not be
effective in BMP2 and BMP4 overexpression cases. This hypothesis in
turn explained why BMP2 and BMP4 did not enhance cell invasiveness
and migration as BMP6 and BMP7 showed. Lavery et al
(32) reported that BMP2/4 and
BMP6/7 utilized different receptors to induce human osteoblastic
differentiation. Whether the similar mechanisms also existing in
cancer biology is unknown, but ALK2 interference did not affect
BMP2/4-induced activities and BMP2/4 preferential receptor ALK6 was
absent. Further investigation on involving signal pathways in
BMP2/4-overexpressed cells showed activation of ERK1/2 and P-38
pathways which were nearly unchanged in BMP6/7-overexpressed cells
compared with control. The utilization of other signaling pathways
by BMPs to exert different functions other than classical SMAD1/5/8
pathways has been addressed in other studies. Jung et al
(33) reported that BMP9 potently
inhibited liver cancer stem cell percentage by phosphorylating P-38
which in turn activated ID3 expression and p21 induction. In blood
vessel endothelium cells, BMP4 induced capillary sprouting relying
on ERK phosphorylation activation (34). Although several reports have
indicated potential crosstalk between BMPs and MAPK signal pathway,
the definite mechanisms is largely unknown. Our reports showed that
MAPK signal pathway may participate in BMP2/4-related biological
activities in esophageal squamous cancer cells. Whether this
phenomenon prevalently exists or is tissue- specific need further
investigation. The discrepancy of BMP2/4 and BMP6/7 effects on ESCC
cells may reflect their different functions in esophageal
development. In normal tissue and organ development, stem cell
differentiation and self-renewal ability are strictly regulated.
The similarities between cancer stem cells and normal stem cells
indicate the complicated tumor traits at differentiation or
dedifferentiation perspective. As shown in the study of Jiang et
al (35), normal adult mouse
esophagus basal progenitor cells resisted BMP4-induced squamous
differentiation by expressing BMP inhibitor follistatin, and human
immortal esophageal progenitor cells can also be induced to
differentiate by BMP4 and proliferation inhibition. BMP4 can also
promote hepatic cancer stem cells to differentiation by activating
ERK1/2 pathway (36). On the
contrary, BMP6 expression in esophageal cancer was negatively
related with tumor differentiation and keratinization (7).
The present study demonstrated that BMP signaling is
potentially activated in esophageal squamous cancer cells and
related with cancer cell growth, invasion and metastasis. BMP6 were
shown prevalently expressed in esophageal squamous cancer tissues
and cell lines and associated with cancer progress and prognosis
(19). BMP6/7 activated
SMAD-dependent classical BMP signaling pathways through type I
receptor ALK2. ALK2 interference diminished cell growth, and
BMP6/7-induced accelerated invasion and metastasis. Therefore,
targeting BMP6/7 signaling pathway in esophageal squamous cancer,
such as ALK2-specific inhibitor application, lay a rational
foundation for esophageal squamous cancer treatment.
References
|
1
|
Rees JR, Onwuegbusi BA, Save VE, Alderson
D and Fitzgerald RC: In vivo and in vitro evidence for transforming
growth factor-beta1-mediated epithelial to mesenchymal transition
in esophageal adenocarcinoma. Cancer Res. 66:9583–9590. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen D, Zhao M and Mundy GR: Bone
morphogenetic proteins. Growth Factors. 22:233–241. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang S, Du J, Wang Z, Yuan W, Qiao Y,
Zhang M, Zhang J, Gao S, Yin J, Sun B, et al: BMP-6 promotes
E-cadherin expression through repressing deltaEF1 in breast cancer
cells. BMC Cancer. 7:2112007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shepherd TG and Nachtigal MW:
Identification of a putative autocrine bone morphogenetic
protein-signaling pathway in human ovarian surface epithelium and
ovarian cancer cells. Endocrinology. 144:3306–3314. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim IY, Lee DH, Ahn HJ, Tokunaga H, Song
W, Devereaux LM, Jin D, Sampath TK and Morton RA: Expression of
bone morphogenetic protein receptors type-IA, -IB and -II
correlates with tumor grade in human prostate cancer tissues.
Cancer Res. 60:2840–2844. 2000.PubMed/NCBI
|
|
6
|
Celeste AJ, Iannazzi JA, Taylor RC, Hewick
RM, Rosen V, Wang EA and Wozney JM: Identification of transforming
growth factor beta family members present in bone-inductive protein
purified from bovine bone. Proc Natl Acad Sci USA. 87:9843–9847.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ebisawa T, Tada K, Kitajima I, Tojo K,
Sampath TK, Kawabata M, Miyazono K and Imamura T: Characterization
of bone morphogenetic protein-6 signaling pathways in osteoblast
differentiation. J Cell Sci. 112:3519–3527. 1999.PubMed/NCBI
|
|
8
|
Park Y, Kim JW, Kim DS, Kim EB, Park SJ,
Park JY, Choi WS, Song JG, Seo HY, Oh SC, et al: The bone
morphogenesis protein-2 (BMP-2) is associated with progression to
metastatic disease in gastric cancer. Cancer Res Treat. 40:127–132.
2008. View Article : Google Scholar
|
|
9
|
Rothhammer T, Poser I, Soncin F, Bataille
F, Moser M and Bosserhoff AK: Bone morphogenic proteins are
overexpressed in malignant melanoma and promote cell invasion and
migration. Cancer Res. 65:448–456. 2005.PubMed/NCBI
|
|
10
|
Langenfeld EM, Calvano SE, Abou-Nukta F,
Lowry SF, Amenta P and Langenfeld J: The mature bone morphogenetic
protein-2 is aberrantly expressed in non-small cell lung carcinomas
and stimulates tumor growth of A549 cells. Carcinogenesis.
24:1445–1454. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ide H, Yoshida T, Matsumoto N, Aoki K,
Osada Y, Sugimura T and Terada M: Growth regulation of human
prostate cancer cells by bone morphogenetic protein-2. Cancer Res.
57:5022–5027. 1997.PubMed/NCBI
|
|
12
|
Thériault BL, Shepherd TG, Mujoomdar ML
and Nachtigal MW: BMP4 induces EMT and Rho GTPase activation in
human ovarian cancer cells. Carcinogenesis. 28:1153–1162. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Milano F, van Baal JW, Buttar NS, Rygiel
AM, de Kort F, DeMars CJ, Rosmolen WD, Bergman JJ, VAn Marle J,
Wang KK, et al: Bone morphogenetic protein 4 expressed in
esophagitis induces a columnar phenotype in esophageal squamous
cells. Gastroenterology. 132:2412–2421. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Na YR, Seok SH, Kim DJ, Han JH, Kim TH,
Jung H, Lee BH and Park JH: Bone morphogenetic protein 7 induces
mesenchymal-to-epithelial transition in melanoma cells, leading to
inhibition of metastasis. Cancer Sci. 100:2218–2225. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Alarmo EL, Korhonen T, Kuukasjärvi T,
Huhtala H, Holli K and Kallioniemi A: Bone morphogenetic protein 7
expression associates with bone metastasis in breast carcinomas.
Ann Oncol. 19:308–314. 2008. View Article : Google Scholar
|
|
16
|
Lim M, Chuong CM and Roy-Burman P: PI3K,
Erk signaling in BMP7-induced epithelial-mesenchymal transition
(EMT) of PC-3 prostate cancer cells in 2- and 3-dimensional
cultures. Horm Cancer. 2:298–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen X, Liao J, Lu Y, Duan X and Sun W:
Activation of the PI3K/Akt pathway mediates bone morphogenetic
protein 2-induced invasion of pancreatic cancer cells Panc-1.
Pathol Oncol Res. 17:257–261. 2011. View Article : Google Scholar
|
|
18
|
Augeri DJ, Langenfeld E, Castle M,
Gilleran JA and Langenfeld J: Inhibition of BMP and of TGFβ
receptors downregulates expression of XIAP and TAK1 leading to lung
cancer cell death. Mol Cancer. 15:272016. View Article : Google Scholar
|
|
19
|
Yuen HF, McCrudden CM, Grills C, Zhang SD,
Huang YH, Chan KK, Chan YP, Wong ML, Law S, Srivastava G, et al:
Combinatorial use of bone morphogenetic protein 6, noggin and SOST
significantly predicts cancer progression. Cancer Sci.
103:1145–1154. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Megumi K, Ishigami S, Uchikado Y, Kita Y,
Okumura H, Matsumoto M, Uenosono Y, Arigami T, Kijima Y, Kitazono
M, et al: Clinicopathological significance of BMP7 expression in
esophageal squamous cell carcinoma. Ann Surg Oncol. 19:2066–2071.
2012. View Article : Google Scholar :
|
|
21
|
Hu M, Liu Q, Song P, Zhan X, Luo M, Liu C,
Yang D, Cai Y, Zhang F, Jiang F, et al: Abnormal expression of the
mitotic checkpoint protein BubR1 contributes to the
anti-microtubule drug resistance of esophageal squamous cell
carcinoma cells. Oncol Rep. 29:185–192. 2013.
|
|
22
|
Pickup MW, Hover LD, Guo Y, Gorska AE,
Chytil A, Novitskiy SV, Moses HL and Owens P: Deletion of the BMP
receptor BMPR1a impairs mammary tumor formation and metastasis.
Oncotarget. 6:22890–22904. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Langenfeld E, Hong CC, Lanke G and
Langenfeld J: Bone morphogenetic protein type I receptor
antagonists decrease growth and induce cell death of lung cancer
cell lines. PLoS One. 8:e612562013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wahdan-Alaswad RS, Song K, Krebs TL, Shola
DT, Gomez JA, Matsuyama S and Danielpour D: Insulin-like growth
factor I suppresses bone morphogenetic protein signaling in
prostate cancer cells by activating mTOR signaling. Cancer Res.
70:9106–9117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim SM, Ye S, Rah SY, Park BH, Wang H, Kim
JR, Kim SH, Jang KY and Lee KB: RhBMP-2 activates hippo signaling
through RASSF1 in esophageal cancer cells. Sci Rep. 6:268212016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kokorina NA, Zakharkin SO, Krebsbach PH
and Nussenbaum B: Treatment effects of rhBMP-2 on invasiveness of
oral carcinoma cell lines. Laryngoscope. 121:1876–1880.
2011.PubMed/NCBI
|
|
27
|
Peng J, Yoshioka Y, Mandai M, Matsumura N,
Baba T, Yamaguchi K, Hamanishi J, Kharma B, Murakami R, Abiko K, et
al: The BMP signaling pathway leads to enhanced proliferation in
serous ovarian cancer-A potential therapeutic target. Mol Carcinog.
55:335–345. 2016. View
Article : Google Scholar
|
|
28
|
Chen JC, Yang ST, Lin CY, Hsu CJ, Tsai CH,
Su JL and Tang CH: BMP-7 enhances cell migration and αvβ3 integrin
expression via a c-Src-dependent pathway in human chondrosarcoma
cells. PLoS One. 9:e1126362014. View Article : Google Scholar
|
|
29
|
Yuen HF, Chan YP, Chan KK, Chu YY, Wong
ML, Law SY, Srivastava G, Wong YC, Wang X and Chan KW: Id-1 and
Id-2 are markers for metastasis and prognosis in oesophageal
squamous cell carcinoma. Br J Cancer. 97:1409–1415. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hao J, Lee R, Chang A, Fan J, Labib C,
Parsa C, Orlando R, Andresen B and Huang Y: DMH1, a small molecule
inhibitor of BMP type i receptors, suppresses growth and invasion
of lung cancer. PLoS One. 9:e907482014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Herrera B, van Dinther M, Ten Dijke P and
Inman GJ: Autocrine bone morphogenetic protein-9 signals through
activin receptor-like kinase-2/Smad1/Smad4 to promote ovarian
cancer cell proliferation. Cancer Res. 69:9254–9262. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lavery K, Swain P, Falb D and
Alaoui-Ismaili MH: BMP-2/4 and BMP-6/7 differentially utilize cell
surface receptors to induce osteoblastic differentiation of human
bone marrow-derived mesenchymal stem cells. J Biol Chem.
283:20948–20958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jung JW, Yoon SM, Kim S, Jeon YH, Yoon BH,
Yang SG, Kim MK, Choe S and Kuo MM: Bone morphogenetic protein-9 is
a potent growth inhibitor of hepatocellular carcinoma and reduces
the liver cancer stem cells population. Oncotarget. Sep
16–2016.Epub ahead of print. View Article : Google Scholar
|
|
34
|
Zhou Q, Heinke J, Vargas A, Winnik S,
Krauss T, Bode C, Patterson C and Moser M: ERK signaling is a
central regulator for BMP-4 dependent capillary sprouting.
Cardiovasc Res. 76:390–399. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jiang M, Ku WY, Zhou Z, Dellon ES, Falk
GW, Nakagawa H, Wang ML, Liu K, Wang J, Katzka DA, et al:
BMP-driven NRF2 activation in esophageal basal cell differentiation
and eosinophilic esophagitis. J Clin Invest. 125:1557–1568. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang L, Sun H, Zhao F, Lu P, Ge C, Li H,
Hou H, Yan M, Chen T, Jiang G, et al: BMP4 administration induces
differentiation of CD133+ hepatic cancer stem cells,
blocking their contributions to hepatocellular carcinoma. Cancer
Res. 72:4276–4285. 2012. View Article : Google Scholar : PubMed/NCBI
|