1. Introduction
In order to individualize treatment of cancer
patients, there is a growing need to characterize different cancer
subgroups. More knowledge on cancer subtypes could improve both
prognosis and treatment.
The protein Annexin A2 has been investigated as a
prognostic marker because of its widespread presentation in several
cancer forms. This study takes an overview of Annexin A2 and its
presentation in various cancers including its prognostic values and
potential as therapeutic target.
2. Background
Annexin A2 is part of the Annexin family consisting
of up to 160 unique Annexin proteins (1). There are two criteria for being an
Annexin protein. The first criterion is the ability to bind
negatively charged phospholipid in a calcium-dependent manner.
Second is the structural containment of an Annexin repeat, a
segment of 70 amino acid residues. Annexin proteins comprise four
or eight Annexin repeats and an α-helix disc (1). These folds allow Annexin to move
intracellularly between lipofobic cytosol and lipophilic membrane
compartment in a calcium-dependent manner (2).
Annexin proteins consist of three domains: a
divergent NH2-terminal, a C-terminal and a preserved
domain making the core of the protein (1,3). For
Annexin A2 the NH2-terminal acts as a binding site for S100A10 and
tissue plasminogen activator (t-Pa). The core binds to calcium and
the cell membrane. The C-terminal contains the binding site for
F-actin (4), heparin (5) and plasminogen (6). The substrates for Annexin A2 reveal
its function as an intercellular transport protein, interactor in
cell division and migration and main interactor in plasmin
production.
Intracellular Annexin A2
Free cytoplasmic Annexin A2 exists as a 36-kDa
protein (1). Intracellularly,
Annexin A2 is involved in exocytosis (7), endocytosis (7,8) and
membrane trafficking through lipid micro-domains (9). Knockdown of the Annexin A2 gene,
ANXA2, has been shown to diminish DNA synthesis and cell
proliferation, suggesting that Annexin A2 is a factor in cell
division (10). Furthermore, an
interaction between Annexin A2 and CD44 has been shown to be
essential for the formation of lipid rafts that interact with the
cellular cytoskeleton (11). A
complex of Annexin A2 and S100A10 directly binds F-actin at
cholesterol-rich membrane passages, thereby interacting with the
cytoskeleton (1). It has been
shown that ANXA2 interacts in p53-mediated apoptosis (12) and prevents radiation-induced
apoptosis, the latter by activating pro-survival signals such as
nuclear factor κB (13). Annexin
A2 also inhibits phospholipase A2 (PLA2) in an endogen manner,
acting to inhibit PlA2 induced inflammation (14).
Extracellular and membrane bound Annexin
A2
By binding directly or indirectly to
phosphatidylserine on cells marked for apoptosis, Annexin A2
attends in the engulfment of cells (15). Membrane bound Annexin A2
contributes to fibrinolysis and has anticoagulation effects and
involves binding to t-Pa and S100A10, hereby facilitating plasmin
production (16,17). Furthermore, Annexin A2 seems to
impact neo-angiogenesis which may explain its effect on solid
tumours (17).
Interactions of Annexin A2
S100A10
S100A10 is part of the S100 protein family being
calcium binding proteins of EF-hand type. The S100 family consist
of 25 distinct isoforms weighing from 9 to 13 kDa (18). Twenty-two out of 25 S100 genes are
located in the 1q21 chromosome region which is prone to genomic
rearrangement. This indication of an unstable region supports how
S100 proteins may be a relevant focus in cancer development
(18).
The intracellular function of S100A10 includes
calcium homeostasis, cell cycle regulation, phosphorylation, cell
growth, migration and interactions with cytoskeleton components and
regulation of transcriptional factors (18).
The extracellular function of S100 proteins is
comparable to a cytokine-like behaviour by binding to cell surface
receptors (18). S100A10 is seen
to play a critical role in angiogenesis in vivo, suggesting
its role in endothelial cell function (18).
S100A10 is unique in its way of being locked in a
permanently open conformation (19). The binding of Annexin A2 is
accommodated in the free hydrophobic space between Helix III and IV
of the S100A10 dimer (19). In the
cell membrane, Annexin A2 combines with S100A10 forming a 94 kDa
heterotetramer of two Annexin A2 units and two 11-kDa S100A10
proteins. The Annexin A2-S100A10 heterotetramer is a key
plasminogen receptor that on the cell surface mediates the
formation of plasmin. S100A10 furthermore enhance the sensitivity
of Annexin A2 to calcium, interfering with the calcium level needed
to conduct Annexin A2 function (20). S100A10-Annexin A2 interaction seems
to play a role in the cell-to-cell adhesion of breast cancer cells
and multiple types of endothelium. Interaction has been shown by
investigating the protein expression of Annexin A2 and S100A10 in
tissue from breast cancer patients (21).
The plasminogen/plasmin system is interaction
between coagulation factors and enzymes. Plasminogen is the
inactive form of plasmin found in plasma and extra cellular matrix
(ECM). Plasminogen is cleaved by plasminogen activators (tPa)
through the hydrolysis of the Arg561-Val562 peptide bond to yield
the serine protease, plasmin (22). Plasmin is an enzyme cleaving fibrin
in the ECM by direct binding or by activating other proteases
(23). Altogether
plasminogen/plasmin/fibrin are important regulators of proteolysis
of ECM, fibrin clot degradation, macrophage migration, tissue
remodelling, invasion and angiogenesis (22). An overproduction of plasmin in the
tumour microenvironment enhance the degradation of ECM, hereby
facilitating tumour invasion (22). Annexin A2 has independent binding
sites for both tPa and plasminogen. By assembling these proteins on
the cell surface, Annexin A2 accelerates the production of plasmin
(22). The binding site between
Annexin A2 and tPa are mechanically blocked by lipoprotein(a) and
homocysteine which both are arteriothrombotic agents (2). Annexin A2-S100A10 regulates 50–90% of
the plasmin generation in several types of normal cells and cancer
cells (24).
Human epididymis protein 4 (HE4)
The binding site of HE4 to Annexin A2 is located
after the 26th amino acid at the N-terminus (5). HE4 and Annexin A2 activates the
extracellular signal-regulated kinase/mitogen-activated protein
kinase (ERK/MAPK) and focal adhesion kinase (FAK) pathways
(25). The ERK/MAPK signalling
pathway influences proliferation, migration and apoptosis (26). FAK is a regulator of cell
signalling within the tumour microenvironment and controls cell
movement, invasion and survival (27).
Angiostatin (AS)
Angiostatin is a 38-kDa internal fragment of
plasminogen that interacts through the lysine binding domain on the
C-terminus of Annexin A2 on the cell surface (28). This is also the binding site for
plasminogen (22). By using AS to
compete with plasminogen binding, ~40% of the binding to bovine
arterial endothelium cells were blocked (29).
Human procathepepsin
Procathepepsin B is an enzyme hosted in lysosomes.
Procathepepsin B can degrade ECM proteins such as laminin,
fibronectin and collagen IV, hereby facilitating invasion of the
tumours cells (30).
Immunohistochemistry (IHC) showed recombinant procathepsin to
interact with the Annexin A2-S100A10 heterotetramer (31). Likewise, it has been shown that
procathepepsin and the Annexin A2 heterotetramer co-localize in two
lines of human cancer cells, one of epithelial origin and one of
mesenchymal origin. Procathepsin B can alone or in interaction with
Annexin A2 activate other proteolytic proteins such as
urokinase-type plasminogen activator and collagenase. These
proteins also degrade ECM and facilitate invasion (30).
Stromal derived factor 1
(SDF1/CXCL-12)
Multiple myeloma, breast and prostate cancer are
known to metastasize to the bone marrow (32) causing distinct pain and sometimes
neural complications for the patients. In vivo and in
vitro studies have showed how Annexin A2 and CXCL-12
co-localize in the bone marrow. Studies using whole bone marrow
cells and investigating the migration of hematopoietic stem cells
(HSC) demonstrated a synergistic effect of CXCL-12 bound to Annexin
A2 (33). Expression of Annexin A2
in bone marrow stromal cells (BMSC) significantly increased the
binding of prostate cancer (PC) cells to BMSC (34). Furthermore, studies demonstrated
how CXCL-21 expression induced the migration of PC cells towards
BMSC (34). Multiple myeloma cells
also express Annexin A2. It was demonstrated how this Annexin A2
expression facilitated the adhesion of osteoblasts and stromal
cells in the bone marrow (35).
Functional regulation of Annexin
A2
The N-terminus of Annexin A2 is regulated by
phosphorylation. Annexin A2 and S100A10 form their heterotetramer
on the cell surface in response to changes in intracellular (ICL)
calcium concentration (2). This
presentation on the surface can be provoked by heat induced stress
of the cell. By using small interfering RNAs (siRNA), targeting
S100A10, it was demonstrated that by reducing S100A10 expression,
the heat stimulated translocation of Annexin A2 was markedly
reduced (2). This shows how
S100A10 is essential to the translocation of Annexin A2 caused by
mild cellular stress (2).
Phosphorylation of Annexin A2 at residue 23 by
src-like tyrosine kinases plays a role in the translocation of
Annexin A2 to the cell surface (2). By using tyrosine kinase inhibitors,
the heat stress induced expression of Annexin A2 on the cell
surface was completely blocked demonstrating how tyrosine kinase
phosphorylation of tyrosine 23 in Annexin A2 is essential to the
translocation to the cell surface (2). Furthermore, protein kinase C (PKC)
combined with calcium phosphorylates Annexin A2. This can be
inhibited by Annexin A5 by interaction with PKC and diminishing its
effect thereby reducing Annexin A2 translocation. Annexin A5 might
have a therapeutic function in preventing Annexin A2s adverse
function in cancer development.
Method
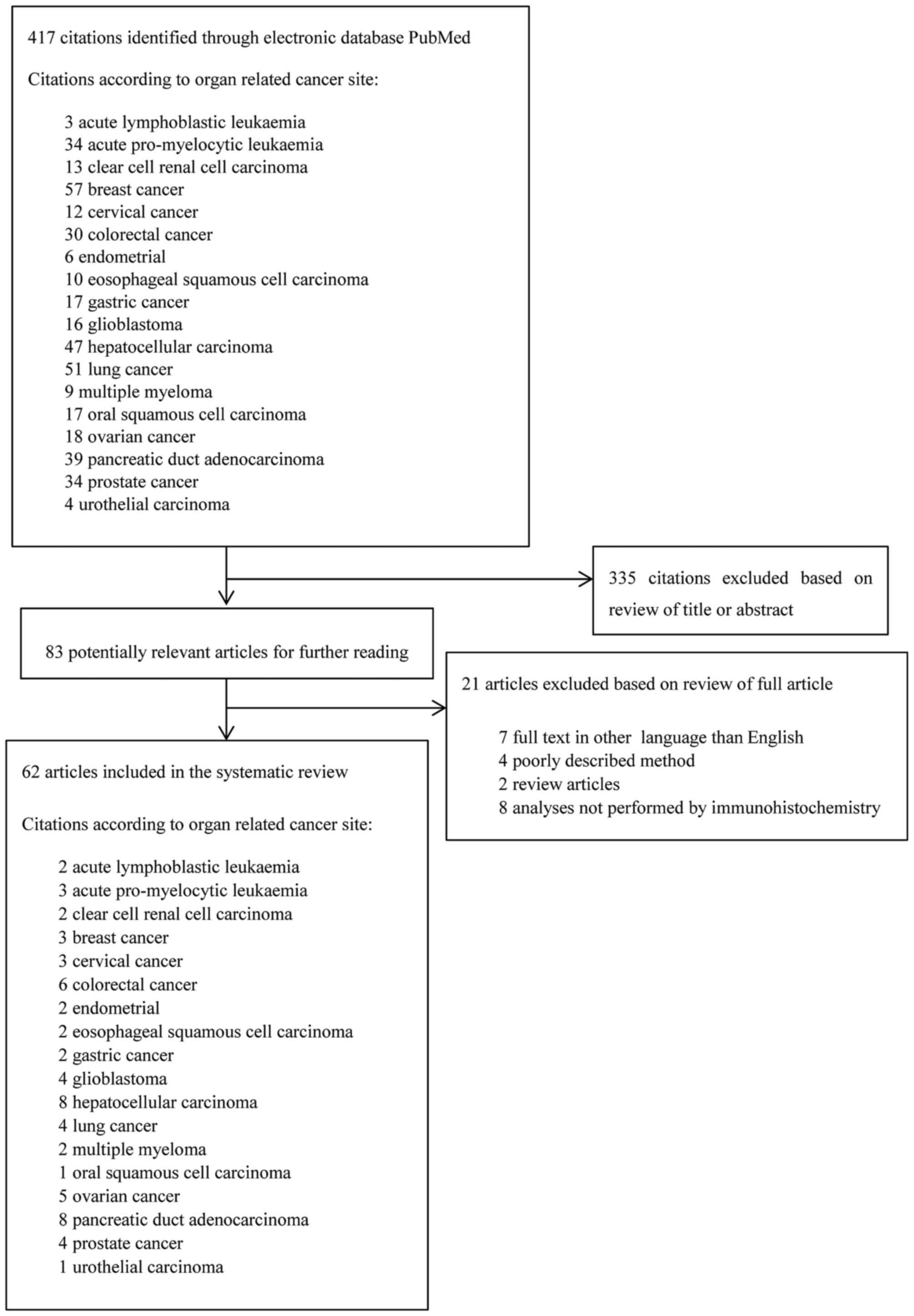
Initial search in PubMed database was made February
6, 2017 (Fig. 1). The following
search criteria 'the organ related cancer site' and 'Annexin A2'
was used. The search was combined with a screening for literature
in reference sections of relevant studies. For preliminary
screening of the articles, all titles and abstracts were read. A
total of 62 studies were included in the review and included
studies were published from 1990–2016.
3. Overview of the publications
Annexin A2 is overexpressed in clear cell renal cell
carcinoma, breast-, cervical-, colorectal-, endometrial-, gastric
cancer, hepatocellular carcinoma, lung- and ovarian cancer,
pancreatic duct adenocarcinoma, glioblastoma and urothelial
carcinoma, acute lymphoblastic leukaemia, acute promyelocytic
leukaemia and multiple myeloma (36–86).
Downregulation of Annexin A2 is reported in
oesophageal squamous cell carcinoma (88,89).
Both upregulation and downregulation of Annexin A2
have been suggested as prognostic markers for patients diagnosed
with oral squamous cell carcinoma (90) and prostate cancer (34,91–93).
In Table I an overview of the
relationship between Annexin A2 and different cancers is
presented.
 | Table ISummary of the tissue protein
expression of Annexin A2 in various cancer forms. |
Table I
Summary of the tissue protein
expression of Annexin A2 in various cancer forms.
| Cancer | Expression | Clinical
manifestation | Therapeutic
interest | Refs. |
|---|
| Haematological
cancers | Acute lymphoblastic
leukaemia | Upregulated | Resistant to
glucocorticoid
Binding and retention of ALL cells in the bone marrow | Combining Annexin
A2 inhibitors with dexamethasone and vincristine increased the
effect.
Using agents to disrupt ANXA2-P11 interaction reduced amount of ALL
cells in blood, spleens and bone marrow. | (36,37) |
| Acute promyelocytic
leukaemia | Upregulated | Haemorrhagic
complications | Treatment with ATRA
or siRNA targeting Annexin A2 could resolve hyperfibrinolysis in
APL. | (38–40) |
| Multiple
myeloma | Upregulated |
Proliferation
Adhesion to osteoblast and stromal cells
Invasive potential | siRNA targeting
Annexin A2 could interfere with the malignant properties of MM
cells | (35,73) |
| Urological
cancers | Clear cell renal
cell carcinoma | Upregulated | Higher
TNM-stage
Histological grade
Infiltration of the renal capsule
Metastatic potential
Shortened 5-year survival rate | | (41,42) |
| Urothelial
cancer | Upregulated | Depth of
invasion
Lymph node metastasis
Distant metastasis
Survival rate | | (86) |
| Prostate
cancer | Downregulated | Gleason score
5-7
Tumour stage
Recurrence
Lymph node metastasis
Distant metastasis
Survival rate | | (90–92) |
| Prostate
cancer | Upregulated | Gleason score
8
Homing and adhesion to bone-marrow
Resistance to chemotherapy | | (34,90) |
| Breast cancer | Breast cancer | Upregulated | Herceptin
resistance
Neo-angiogenesis
Migration and invasion
Proliferation
Metastasis | Treatment with
siRNA targeting Annexin A2 could diminish Herceptin resistance and
cell proliferation Anti-Annexin A2 antibodies disrupt
neo-angiogenesis | (29,43,44) |
| Gynaecological
cancers | Cervical
cancer | Upregulated | Chemotherapy
resistance
Advanced cancer stage
Decreased progression free-survival
HPV16 internalisation | Treatment with
anti-Annexin A2 antibodies or Annexin A2 ligands might decrease
cervical cancer caused by HPV16 | (45–47) |
| Endometrial
cancer | Upregulated | Higher histological
grade
Higher FIGO stage
Depth of invasion
Lymph node metastasis
Distant metastasis
Overall survival rate | | (54,55) |
| Ovarian cancer | Upregulated | Poorly
differentiated tumours
Histological grade
Ascites
Malignant tumour cells in peritoneal fluid
FIGO-stage
Invasion and migration
Metastasis
Disease progression
Death caused by OC
Reduced PFS
Shortened overall survival rate | siRNA targeting
Annexin A2 decrease motility of OC cells Treatment with Annexin A2
neutralizing antibodies reduced tumour burden in vivo and
could possibly be transmitted to human OC patients
RNA nanoparticle harbouring Annexin A2 could be used to deliver
doxorubicin directly into OC cells overcoming chemotherapy
resistance | (22,74–77) |
| Gastroenterological
cancers | Colorectal
cancer | Upregulated | Tumour
size
Higher TNM stage
Growth factor mediating
Poor prognosis
Recurrence | | (48–53) |
| Colorectal
cancer | Decreased in
serum | Tumour
size
TNM-stage
Tumour invasion
Lymph node metastasis
Distant metastasis | | (53) |
| Esophagael squamous
cell carcinoma | Downregulated | Lymph node
metastasis
Depth of invasion
Poor differentiation | | (87,88) |
| Gastroenterological
cancers | Gastric cancer | Upregulated | Tumour
size
Histological type
Depth of invasion
Vessel invasion
Lymph node metastasis
Distant metastasis
TNM-stage
Shortened 5-year survival rate | | (56,57) |
| Hepatocellular
carcinoma | Upregulated | Tumour
size
Intra- and extrahepatic metastasis
Portal vein thrombosis
TNM-stage
Metastasis
Poor prognosis
Shortened 5-year survival rate
Progression free-survival | By facilitating
degradation of Annexin A2, UBAP2 might be of therapeutic interest
in countering the adverse effect of Annexin A2 among HCC
patients. | (61–68) |
| Oral squamous cell
carcinoma | Upregulated | Tumour
size
Tumour recurrence | | (89) |
| Oral squamous cell
carcinoma | Downregulated | Histological
grade | | (89) |
| Pancreatic duct
adenocarcinoma | Upregulated | Cell motility and
viability
Histological grade
Poor prognosis
Distant metastasis
Shortened progression free-survival
Shortened overall survival rate
resistance to adjuvant chemotherapy with gemcitabine | Anti-Annexin A2
antibodies, microRNA-206 and shRNA harbouring Annexin A2 has shown
to decrease the adverse outcomes from PDA patients. | (78–85) |
| Neurological
cancers | Glioblastoma | Upregulated |
Neo-angiogenesis
Tumour stages
Three year survival | | (58–60,95) |
| Lung cancer | Non-small cell lung
cancer | Upregulated | Tumour
diameter
Pathological grade
pN-status
pT-status
Pleural invasion
Advanced clinical stage
Shortened overall survival | Annexin A2 is
expressed in NSCLC cell lines resistant to chemotherapy. Targeting
Annexin A2 might diminish multi-drug resistant tumours.
Sh-RNA plasmids toward Annexin A2 might inhibit the effect of
Annexin A2 in development of advanced clinical stage tumours of
NSCLC. | (69-72) |
Haematological cancers
Acute lymphoblastic leukaemia
(ALL)
In patients with ALL, elevated levels of ANXA2 and
increased amounts of phosphorylated Annexin A2 relate to resistance
to glucocorticoid treatment (36).
Annexin A2 is phosphorylated by Src-kinase, the reaction is
facilitated by S100A10 (37).
Elevated Annexin A2, S100A10 and Src-kinase activity may predict
drug resistance in ALL patients (36,37).
By inhibiting the Src-kinase, the ALL cells were sensitised to
glucocorticoid treatment suggesting that Src-kinase inhibitors
might be a supplement to treatment of glucocorticoid resistant ALL
patients (36). Treatment of ALL
cells with anti-Annexin A2 antibody and knockdown of S100A10
abrogate ALL adhesion to osteoblasts (37). This inhibition of ALL cell adhesion
to osteoblasts indicates how Annexin A2 and S100A10 influence the
binding and retention of ALL cells to the bone marrow (37). Additionally, long-term engraftment
assays from mice showed reduced percentage of ALL cells in blood,
spleen and bone marrow if treated with agents that disrupts the
Annexin A2-S100A10 interaction (37). Furthermore, using mouse monoclonal
antibodies against Annexin A2 increased the effect of treatment of
ALL with dexamethasone and vincristine by disruption of the binding
between ALL cells and osteoblasts (37).
Acute promyelocytic leukaemia
(APL)
Overexpression of Annexin A2 in APL cells is thought
to be the mechanism behind haemorrhagic complications of APL
patients (38,39). The t(15;17) translocation positive
APL cells express Annexin A2 in a greater manner than other
leukaemia cells (38). Annexin A2
facilitates the combining of t-Pa and plasminogen on the cell
surface. APL cells with t(15;17) translocation had twice as
effective t-Pa dependent plasmin generation. Annexin A2
overexpression might be the mechanism for haemorrhagic
complications of APL patients (38,39).
By treating APL cells with siRNA targeting Annexin A2, a decreased
t-Pa mediated plasmin generation have been shown (39). Furthermore, APL cells treated with
all-trans retinoic acid (ATRA) showed downregulation of
Annexin A2. These findings indicate that treatment with siRNA
targeting Annexin A2 or ATRA could resolve hyperfibrinolysis in APL
(39,40).
Multiple myeloma (MM)
Annexin A2 is expressed in MM cells in 8/8 patients
(35). Annexin A2 has been shown
to stimulate proliferation of MM cells and to support adhesion of
MM cells to osteoblasts and stromal cells (35). Other studies, suggest that siRNA
silencing of Annexin A2 can induce apoptosis in MM cell lines.
Furthermore, by silencing Annexin A2 the invasive potential of MM
cells were significantly diminished (74). This makes siRNA targeting Annexin
A2 as a potential therapeutic focus to induce apoptosis of MM cells
and interfere with the invasive potential (74).
Urological cancers
Clear cell renal cell carcinoma
(ccRCC)
Annexin A2 is expressed mainly in the membrane of
ccRCC and the amount of Annexin A2 in ccRCC was higher compared to
normal tissue (41). In primary
ccRCC tumours, the expression of Annexin A2 was positively
associated with a higher TNM-stage (P<0.05) (41,42),
histological grade (P<0.05) (41), infiltration of the renal capsule
(P<0.01) (41) and metastatic
potential (P<0.01) (41,42).
Furthermore, Annexin A2 overexpression was significantly correlated
with shortened 5-year survival rate of ccRCC patients compared to
patients with lower expression of Annexin A2 (P<0.01) (41).
Urothelial cancer
Higher Annexin A2 expression is reported in
urothelial carcinoma (55%, 175/315) compared to overexpression in
17.5% (11/63) of the non-tumour mucosa samples (P<0.01)
(87). The expression of Annexin
A2 was associated with the depth of invasion, lymph node metastasis
and distant metastasis (P<0.05). Furthermore, Annexin A2
expression is a significant independent prognostic factor for
survival among urothelial carcinoma patients (P=0.012) (87).
Prostate cancer (PC)
Annexin A2 is localized primarily in the membrane
and faintly in the cytoplasm on PC cells (91). The expression level of Annexin A2
was significantly lower in the PC cases when compared to patients
with benign prostate hyperplasia (P<0.01) (92,93).
Lower Annexin A2 expression was negatively related to Gleason score
5–7, tumour stage, recurrence, lymph node metastasis and distant
metastasis P<0.01 (92).
Additionally, survival rate was significantly correlated to a
downregulation of Annexin A2 expression (P<0.01) (92).
Annexin A2 expression seems to play a critical role
in the homing and adhesion of PC cells to the bone marrow (34). Furthermore, it is demonstrated how
Annexin A2 in bone marrow stromal cells could play a role in the
resistance of PC cells to chemotherapy (34). Although low expression of Annexin
A2 correlated to Gleason score 5–7, a strong and diffuse staining
of Annexin A2 was seen in PC biopsies indicating an association
between Annexin A2 and the most severe PC subtypes (91).
Breast cancer
No expression of Annexin A2 is found in normal or
hyperplastic ductal epithelial cells of the human mammarian tissue.
On the contrary, protein expression of Annexin A2 is found in
breast cancer and ductal carcinoma in situ (CIS) (43). Correspondingly, Annexin A2 has been
shown to be upregulated in HER-2 negative and herceptin resistant
breast cancer cells (44). Annexin
A2s ability to stimulate the production of plasmin combined with
the functional role of plasmin, indicates the possible role of
Annexin A2 in angiogenesis and metastasis of breast cancer cells
(43,94). Annexin A2 has been shown to
maintain constitutive activation of the EGFR-pathway leading to
cell proliferation, migration and viability (94). Annexin A2 downregulation by siRNA
increased apoptosis and decreased cell viability and migration by
inhibiting the Annexin A2 induced, constitutively active
EGFR-pathway (44). Furthermore,
it was shown that anti-Annexin A2 antibodies inhibited
neo-angiogenesis by inducing apoptotic cell death of endothelial
cells (94). This suggests that
siRNA against Annexin A2 could be of therapeutic value in HER-2
negative, herceptin-resistant cancer cells.
Gynaecological cancers
Cervical cancer
Abnormal expression of Annexin A2 and S100A proteins
has been reported to induce resistance to cisplatin-based
chemotherapy among cervical cancer patients. IHC analysis showed
increased Annexin A2 expression in cervical tumour stromal cells
after chemotherapy treatment. In addition to this, Annexin A2
tumour expression was significantly higher in the group of tumours
not responding to chemotherapy treatment, indicating that Annexin
A2 upregulation may play a role in resistance to chemotherapy.
Furthermore, Annexin A2 expression in stromal cells of cervical
cancer patient is an independent prognostic factor for decreased
progression free-survival (46,47).
Annexin A2 was shown to be positively correlated with advanced
cancer (47) indicating how
expression of Annexin A2 relates to higher cancer stages.
Human papilloviruses (HPV) are sexually transmitted
viruses that causally associate with the development of cervical
cancers. The most common, HPV16, is an obligatory intracellular
virus that must gain entry into host cells to survive (48). This HPV16 internalisation has been
demonstrated to be partly facilitated by the Annexin A2-S100A10
heterotetramer. By inhibiting Annexin A2 in an endogenous manner or
with anti-Annexin A2 antibodies, the HPV16 internalisation was
significantly decreased.
Endometrial cancer
Annexin A2 is expressed in both membrane and the
cytoplasm of endometrial cancer cells in 95.2% of the endometrial
carcinomas compared to 55.6% of the normal endometrium (P<0.05)
(55). In vitro studies
suggest Annexin A2 may play a role in the promotion of metastasis
in that endometrial cancer. Knockdown of Annexin A2 resulted in the
absence of lung and hematogenous metastasis (56), implying Annexin A2 to play a role
in the development of distant metastasis among patients with
endometrial cancer. For 91.7% (22/24) of endometrial carcinoma
patients in stage III–IV, a high expression of Annexin A2 was
found. The expression was significantly higher than for patients in
stage I–II with 55% (33/60) (P<0.05). Overexpression of Annexin
A2 is correlated with shorter overall survival (P<0.05)
(55). Together, this suggests
that Annexin A2 could be a potential therapeutic focus in order to
avoid spread of endometrial tumours and to predict recurrence and
overall survival.
Ovarian cancer (OC)
Annexin A2 is expressed in the membrane (77%) and
the cytoplasm (82.6%) of serous OC cells as well as the surrounding
stromal cells (58.5%) (75).
Annexin A2 seems to play a role in regulating cell proliferation of
OC cell lines (76). A significant
increase in Annexin A2 expression in FIGO-stage IV compared to
stage II and III is reported (P=0.001 and P=0.005, respectively)
(75). Annexin A2 expression is
related to histological grade (P=0.002) (76). Furthermore, high expression of
Annexin A2 was significantly related to presence of ascites
(P<0.001) and malignant tumour cells in peritoneal fluid
(P<0.001) (76). Downregulation
of Annexin A2 in OC cells significantly reduced the ability of
invasion and migration (P<0.05). By analysing metastasis from OC
patients it was revealed that there was a high number of lung
metastatic nodules in the high Annexin A2 expression group, whereas
almost no lung metastasis were found in the low expression group
(25). High stromal expression is
significantly associated with reduced progression-free survival
(PFS) (P=0.014) and reduced overall survival (OS) (75). Patients with high stromal Annexin
A2 had a 1.8-fold increased risk of disease progression (P=0.0014)
and a 1.6-fold increased risk of disease related death (P=0.046)
(75). Combined with S100A10,
Annexin A2 expression predicts adverse outcomes for OC patients.
For patients with high expression of stromal Annexin A2 and
cytoplasmic S100A10 the 5-year survival rate was 11.1% compared to
50% for the patients with low stromal Annexin A2 and cytoplasmic
S100A10 (75).
siRNA targeting Annexin A2 significantly decreased
motility (P=0.0069) and invasion (P=0.0047) in OC cell lines. In
vivo studies in mice showed how treatment with Annexin A2
neutralizing antibodies significantly reduced the tumour burden
(77). This makes siRNA targeting
Annexin A2 and Annexin A2 neutralizing antibodies a potential
therapeutic focus of OC treatment. Another study showed that
RNA-nanoparticles harbouring Annexin A2 can be used to deliver
doxorubicin into the OC cells. Knowledge of this mechanism may help
overcome chemotherapy resistant OC and to minimize the adverse
effect of chemotherapy to healthy tissue (78).
Gastroenterological cancers
Colorectal cancer (CRC)
Annexin A2 is highly expressed in CRC cell lines,
both on mRNA level and as protein (49). The expression of Annexin A2 is
shown to induce significant changes on the microstructure of the
cells (50). Upregulated Annexin
A2 promotes proliferation, migration and invasion of CRC cells
in vitro caused by the changes in microstructure (49). In studies investigating Annexin A2
using IHC, high expression of Annexin A2 was significantly
correlated with tumour size (P=0.03), poorly differentiated tumours
(P=0.01), depth of invasion (P=0.02) and TNM-stage (P=0.02)
(51). Annexin A2 was shown to be
an independent factor for poor prognosis in patients with CRC
(51). Annexin A2 in the cell
membrane is a characteristic for tumours with high invasiveness.
This ability to invade tissue shows how Annexin A2 could affect
lymph node metastasis (52,53).
Annexin A2 has also been shown to be important for the effect of
progastrins and gastrins, hereby partially mediating the effect of
growth factors on colon cancer cells (95). Furthermore, Annexin A2 could be
used to predict recurrence of CRC. The 5-year recurrence rate was
69.4% in the high expression group compared to 35.9% in the low
expression group among patients with stage I–II disease (53). On the other hand, the serum Annexin
A2 level is significantly lower in patients with CRC compared with
healthy controls (P<0.001) (54). Low serum Annexin A2 levels were
related to increased tumour size (P=0.003), higher TNM-stage
(P=0.004), tumour invasion (P=0.005), lymph node metastasis
(P=0.003) and distant metastasis (P=0.005) (54). This makes Annexin A2 levels in
serum of interest to classify colon cancer patients.
Oesophageal squamous cell carcinoma
(ESCC) and oral squamous cell carcinoma (OSCC)
Annexin A2 is downregulated on both mRNA and protein
level in ESCC cells (88,89). Annexin A2 was found in 9.1% (2/22)
of the ESCC samples compared to 90.9% (20/22) of the controls using
qRT-PCR and western blot analysis. The expression of Annexin A2 was
confirmed by IHC. Annexin A2 in ESCC cells is found mainly in the
cell membrane. Low Annexin A2 expression is correlated with lymph
node metastasis (P<0.05), depth of invasion (P<0.05) and poor
differentiation (P<0.05) (88).
Annexin A2 is expressed in the cell membrane of
normal epithelial cells of the oral cavity. Annexin A2 was
expressed in 82% (87/106) of the OSCC cases (90). Annexin A2 is associated with
histological grade (P=0.02). Low expression of Annexin A2 is seen
in poorly differentiated tumours compared to well differentiated
tumours (90). On the contrary, a
higher expression of Annexin A2 correlated to tumour size (P=0.003)
and tumour recurrence (P= 0.04) (90).
There are few studies investigating the expression
of Annexin A2 and its relation to ESCC and OSCC. The studies
reviewed have at low number of participants, 22 and 106,
respectively, and it is therefore difficult to indicate a clear
correlation between the expression of Annexin A2 and ESCC and
OSCC.
Gastric cancer
Annexin A2 is found predominantly on the cell
membrane of gastric cancer cells. Annexin A2 is found in non-tumour
mucosa and in human gastric cancer cases in 19.6 and 40.1% of
patients, respectively (57,58).
The expression of Annexin A2 correlates with tumour size (57), histological type (57), depth of invasion (57), vessel invasion (57), lymph node metastasis, distant
metastasis and TNM-stage (P<0.05) (57,58).
Furthermore, for cancer stages I, II and III, the 5-year survival
rate of the patients with high expression of Annexin A2 were
significantly lower compared with 5-year survival for the patients
with low expression (57).
Hepatocellular carcinoma (HCC)
Annexin A2 expression is localized to the cell
membrane and cytoplasm of HCC cells (62,63).
Annexin A2 expression was found in 73.8% (62/84) of the HCC tissues
compared to 35.6% (21/59) of the benign liver disease (BLD) tissue
(P<0.001) (64). Tumours with a
high expression of Annexin A2 were larger in size compared to low
expression tumours (P=0.016) (63,65).
Annexin A2 expression significantly correlated with intrahepatic
metastasis (P=0.02), portal vein thrombosis (P=0.003) and higher
TNM-stage (P=0.024) (64,66). Survival analysis revealed how the
high expression group had a poorer prognosis and a shortened 5-year
survival rate (63,64). Furthermore, Annexin A2 seems to
regulate actin remodelling thereby facilitating invasion and
metastasis of HCC cells making high expression of Annexin A2 a
marker for metastatic potential of HCC cells (67,68).
By comparing serum levels of Annexin A2 from HCC patients and BLD
patients, it was revealed that Annexin A2 was significantly
elevated even at early stage HCC (P=0.0024 and P=0.0048,
respectively) (69). Monitoring
Annexin A2 expression levels in combination with alfa fetoprotein
may contribute to a higher sensitivity and specificity in the
clinical practice of diagnosing HCC (69). Ubiquitin associated protein 2
(UBAP2) has been shown to make a complex with Annexin A2, marking
it for degradation. This makes UBAP2 a potential therapeutic focus
for patients with HCC to counter the adverse effects of Annexin A2
(63).
Pancreatic duct adenocarcinoma
(PDA)
Annexin A2 is localized in the cytoplasm of normal
pancreatic epithelial cells. In late stage pancreatic
intraepithelial neoplasia (PanIN) and invasive PDA, Annexin A2 is
relocated to the outer luminal surface (79). The expression of Annexin A2 gives
PDA tumour cells the ability to grow into the liver (79). Annexin A2 has been shown to
co-localize with S100A6 on the cell membrane of PDA tumours.
Annexin A2 combined with S100A6 contributes to PDA cell motility
(81). Annexin A2 is also
significantly associated with histopathological grading (P=0.029)
(82). Annexin A2 promotes
secretion of the class 3 Semaphorin Sema3D. It was demonstrated how
Sema3D is enriched in metastatic tumours of PDA. Furthermore,
Sema3D is expressed in primary PDA from patients with a poor
prognosis and patients who died from widely metastatic disease
(79). This indicates how Annexin
A2, through Sema3D, promotes metastasis in human PDA and could
predict a poor prognosis for PDA patients (79). Another mechanism by which Annexin
A2 promotes invasion of PDA cells is mediated by Tyrosine 23
phosphorylation of Annexin A2 (80). Annexin A2 can interact with the p50
subunit of NF-κB independently of calcium. By interfering with
NF-κB Annexin A2 helps to induce cellular viability of PDA
(83). An in vivo study on
mice showed how expression of Annexin A2 significantly correlated
with a shortened survival rate (P=0.001) (80). Attributing to this, patients with
high expression of Annexin A2 showed a significantly shortened PFS
and OS compared to low expression patients (P=0.008 and P=0.033,
respectively) (84). Treatment
with anti-Annexin A2 antibodies prolonged the survival rate of mice
compared with mice treated with isotype control IgG (P=0.02).
Furthermore, microRNA-206 has shown the ability to reduce Annexin
A2 plasmin production and thereby inhibiting PDA cell invasion
(86).
Takano et al (84) examined the relevance of Annexin A2
in drug-resistant PDA and found high expression of Annexin A2 as an
independent factor for recurrence in patients undergoing
gemcitabine adjuvant chemotherapy. Knocking down Annexin A2 by
shRNA significantly increased the cytotoxic effect of gemcitabine
by the following downregulation of the NF-κB pathway (83).
These studies of anti-Annexin A2 antibodies,
microRNA-206 and shRNA targeting Annexin A2 altogether indicate
Annexin A2 to be of therapeutic relevance in patients with PDA
(85).
Neurological cancers
Glioblastoma
The majority of human glioblastoma cases expressed
Annexin A2 (59,60). Twenty-five out of 30 cases were
Annexin A2 positive, with 11/30 cases strongly positive, 14/30
cases weakly positive and the remaining 5 cases negative (60). Annexin A2 expression is not
detectable in normal glia cells (96). Furthermore, Annexin A2 is
overexpressed in the cases with a higher grade tumour, pathological
grade II–IV, compared to low grade tumours without infiltration
(61). The 3-year OS rate of the
Annexin A2 positive group was significantly lower than survival of
the Annexin A2 negative group, 31.5 and 51.8%, respectively
(59). Annexin A2 expression was
shown to correlate with vascular endothelial growth factor (VEGF)
and platelet-derived growth factor (PDGF), P<0.05. Annexin A2's
effect on glioblastoma cells through VEGF and PDGF may represent an
important anti-angiogenesis therapeutic target in the treatment of
glioma (60). A significant lower
cell proliferation was shown after knockdown of Annexin A2 by using
a wound closure rate (59).
Knockdown of Annexin A2 reduced the invasiveness of glioma cells
(59).
Lung cancer
Non-small cell lung cancer
(NSCLC)
Annexin A2 is shown in cytoplasm and cell membranes
in lung adenocarcinoma cells lines (70). Annexin A2 expression is
significantly correlated with tumour diameter (P=0.003),
pathological grade (P=0.014), pT status (P<0.001) and pleural
invasion (71). High expression of
Annexin A2 was associated with lymph node metastasis in comparison
with tumours with low Annexin A2 expression (P<0.001) (72). Annexin A2 overexpression correlates
to advanced clinical stage (P<0.001) and patients with high
Annexin A2 expression have shorter survival compared to patients
with low expression (P<0.001) (71,72).
Short hairpin plasmid mediated RNA (shRNA)
interference is a way of post-translation gene silencing (70). A plasmid expressing a shRNA
targeting Annexin A2 could effectively inhibit the expression of
Annexin A2. This implies that the shRNA plasmid against Annexin A2
could be of therapeutic interest to decrease the proliferation and
invasion capability of NSCLC cells (70).
Another study using immunoprecipitation and flow
cytometry showed Annexin A2 expression and phosphorylation in cell
lines with resistance properties to doxorubicin, vinca alkaloids
and epipodophyllotoxins indicating a relation of Annexin A2 to
prediction of multi-drug resistance (73).
4. Discussion
The mechanism for Annexin A2 influence in different
tumour types varies and thus the general role of Annexin A2 in
malignant tumours remains unclear. Table II summarizes the clinical
manifestations correlated to overexpression of Annexin A2 and the
cancers in which they are represent. In general it seems that
upregulation is the predominant phenomenon which can be correlated
to an adverse clinical outcome for the patients.
| Table IISummary of the clinical
manifestations of overexpression of Annexin A2 and the organ
related cancer site. |
Table II
Summary of the clinical
manifestations of overexpression of Annexin A2 and the organ
related cancer site.
| Clinical
manifestation | Organ related
cancer site |
|---|
| Resistance to
treatment | Acute lymphoblastic
leukaemia, breast, cervical, pancreatic duct adenocarcinoma,
prostate cancer |
| Binding to the bone
marrow | Acute lymphoblastic
leukaemia, multiple myeloma, prostate cancer |
| pT-stage | Clear cell renal
cell carcinoma, colorectal, esophageal squamous cell carcinoma
(downregulation), endometrial, gastric, hepatocellular carcinoma,
non-small cell lung, ovarian, oral squamous cell carcinoma,
urothelial cancer |
| pN-stage | Clear cell renal
cell carcinoma, colorectal, esophageal squamous cell carcinoma
(downregulation), endometrial, gastric, non-small cell lung cancer,
ovarian, pancreatic duct adenocarcinoma, prostate (downregulation),
urothelial cancer |
| pM-stage | Clear cell renal
cell carcinoma, breast, colorectal, endometrial, gastric,
hepatocellular carcinoma, ovarian, pancreatic duct adenocarcinoma,
prostate (downregulation), urothelial cancer |
| Histological grade
and type | Clear cell renal
cell carcinoma, cervical, endometrial, gastric, non-small cell
lung, ovarian, oral squamous cell carcinoma (downregulation),
pancreatic duct adenocarcinoma, prostate cancer |
| Shortened
survival | Clear cell renal
cell carcinoma, endometrial, gastric, glioblastoma, hepatocellular
carcinoma, non-small cell lung, ovarian, pancreatic duct
adenocarcinoma, prostate cancer (downregulation) |
Annexin collaborate with different proteins such as
plasminogen, S100A10 and HE4. It might be the complex interaction
between these agents and Annexin A2 that play a part in its
malignant potential. Activation by phosphorylation seems to play a
role in carcinogenesis and to some extent Annexin A2 seems to be
regulated by Annexin A5. This accentuates the need to investigate
the expression patterns of different Annexins within the different
cancer forms. The ongoing regulation and collaboration of Annexin
A2 might be the foundation for its malignant potential and might be
the focus for further investigation on targeting treatment toward
Annexins and plasminogen, S100 proteins and HE4.
Although it is hypothesised how Annexin A2 mediated
activation of proteases leads to tumour cell proliferation and
invasion, the fact that Annexin A2 is downregulated in some
malignant tumours, poses some inconsistencies in the theory of
Annexin A2 relation in carcinogenesis. By making this overview on
Annexin A2 expression in different cancers there were some
contradicting studies on the topic (90–92,97).
For OSCC and prostate cancer, a down-regulation of Annexin A2 was
demonstrated. Differences between demographic data (age, sex and
time of data collection) could influence the outcome of the results
in both studies investigating OSCC and prostate cancer. The
influence is more noticeable when the number of patients is low,
and larger studies to confirm results are needed. In the OSCC study
by Rodrigo Tapia et al (90) the non-malignant tissue from
patients undergoing non-cancerous surgery was investigated. In
another study by Zhong et al (97) the nonmalignant tissue is not taken
from healthy controls but taken from the cancer patients as
biopsies of epithelial tissue 2 cm from the cancer. This difference
could be debated because it is not well known which kind of
non-malignant tissue is best standard for comparison.
Yee et al (91) suggests that Annexin A2 expression
distinguish benign illnesses of the prostate gland, like basal cell
hyperplasia and prostate atrophy, from high grade intraepithelial
neoplasia and prostate cancer. Conflicting data on the Annexin A2
expression in tumours with Gleason score 8–10 has been published.
Ding et al (92) did not
confirm upregulation of Annexin A2 in Gleason score 8–10 tumours.
The contradictive data could be caused by a different scoring of
expression levels in combination with the slightly different
laboratory protocol. Although both studies used the same Annexin A2
mouse antibody different kit lot number may result in staining
differences.
The dichotomic expression of Annexin A2 in
different cancers emphasizes the need to characterize the
individual protein expression profiles according to the different
cancers. That said, it might not be of interest to make a
comparison of Annexin A2 expression among different cancers but
more relevant to have a broader knowledge of Annexin A2 profiling
within the different cancer subtypes.
All the studies enrolled in the present review were
using IHC which gives them some limitations. The staining level,
antibody concentration and cut-off values could be different among
studies. When comparing the staining level of Annexin A2 the
individual pathologist makes the classification of the tumours. No
international scoring system for Annexin A2 exists. In most of the
studies in this review, two independent pathologists made the
scoring resulting in a consensus score. If Annexin A2 should be
used in clinical routine setting then the quality of staining
should be monitored by attending an assurance program.
This review has some limitations. Even though we
sought to run through the literature of Annexin A2 and cancer, not
all human tumour types are included. Secondly, within the cancer
forms it is not discussed on behalf of the individual histological
cancers subtypes. All histological subtypes are included and might
therefore give a less specific correlation. Thirdly, publications
bias cannot be completely ruled out because of the tendency to
include papers with positive results compared to negative results.
This bias is elaborated by discussing the papers giving a negative
result.
Looking at the possibility of implementing IHC
staining for Annexin A2 in the common practice at the hospital, the
actual use of IHC and the possibility of elaborating the IHC panel
are evaluated. IHC is a supplement to the histopathological
description. Today most tumours are already undergoing IHC staining
with other antibodies. Having to stain for Annexin A2 would only be
an extension to the IHC panel that are used today.
Overall there is a limited amount of research on
the topics concerning the Annexin A2 expression and recurrence
rates of different cancers or the possible correlation between
upregulated Annexin A2 and primary chemoresistance. Also there is a
need for more profound research on the pathways and mechanisms of
Annexin A2 in tumour genesis. This should be of interest because of
the potential of Annexin A2 as a therapeutic agent in the treatment
of different cancers. There are only a limited number of studies
concerning the use of anti-Annexin A2 antibodies on human cancers
tissue but the results on mice are promising (98). The ability of siRNAs to interfere
with Annexin A2 expression might be of interest in the treatment of
malignant diseases because of its potential to diminish treatment
resistance and hyperfibrinolytic complications (39,44).
There is a growing need to individualize cancer treatment and it
could be of interest to have more knowledge on the use on
anti-Annexin A2 antibodies and siRNAs on human cancer cells.
6. Conclusion
Overall, the present study reviews the ability of
Annexin A2 expression in various cancer cells to predict adverse
outcome. Expression of Annexin A2 is correlated to advanced stages
and metastatic disease. Annexin A2 is demonstrated to predict
reduced OS, shortened PFS and resistance to present treatment
regimens. Although an overexpression of Annexin A2 is correlated to
adverse outcome of the patient, a lowered expression of Annexin A2
is also correlated to poor prognosis in a few cancer types.
The present review sought to make an outline on
Annexin A2s influence on cancer and make a basis for further
investigation hereby contributing to knowledge of Annexin A2
resulting in consensus of a possible clinical use of this
biomarker.
Acknowledgments
A special thanks to Julie Lilith Hentze at the
Department of Pathology, Molecular Unit, Herlev Hospital,
University of Copenhagen for guidance and discussions during
manuscript processing. The Danish Cancer Society kindly funded a
scholarship for the first author (M.V.C.) as part of a research
Year Project (grant no. R165-A10261-16-S7).
References
|
1
|
Gerke V and Moss SE: Annexins: From
structure to function. Physiol Rev. 82:331–371. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Deora AB, Kreitzer G, Jacovina AT and
Hajjar KA: An annexin 2 phosphorylation switch mediates
11-dependent translocation of annexin 2 to the cell surface. J Biol
Chem. 279:43411–43418. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang CY and Lin CF: Annexin A2: Its
molecular regulation and cellular expression in cancer development.
Dis Markers. 2014:3089762014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Filipenko NR and Waisman DM: The C
terminus of annexin II mediates binding to F-actin. J Biol Chem.
276:5310–5315. 2001. View Article : Google Scholar
|
|
5
|
Kassam G, Manro A, Braat CE, Louie P,
Fitzpatrick SL and Waisman DM: Characterization of the heparin
binding properties of annexin II tetramer. J Biol Chem.
272:15093–15100. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hajjar KA, Jacovina AT and Chacko J: An
endothelial cell receptor for plasminogen/tissue plasminogen
activator. I Identity with annexin II. J Biol Chem.
269:21191–21197. 1994.PubMed/NCBI
|
|
7
|
Sarafian T, Pradel LA, Henry JP, Aunis D
and Bader MF: The participation of annexin II (calpactin I) in
calcium-evoked exocytosis requires protein kinase C. J Cell Biol.
114:1135–1147. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Emans N, Gorvel JP, Walter C, Gerke V,
Kellner R, Griffiths G and Gruenberg J: Annexin II is a major
component of fusogenic endosomal vesicles. J Cell Biol.
120:1357–1369. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Babiychuk EB and Draeger A: Annexins in
cell membrane dynamics. Ca2+-regulated association of
lipid microdomains. J Cell Biol. 150:1113–1124. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chiang Y, Rizzino A, Sibenaller ZA, Wold
MS and Vishwanatha JK: Specific down-regulation of annexin II
expression in human cells interferes with cell proliferation. Mol
Cell Biochem. 199:139–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oliferenko S, Paiha K, Harder T, Gerke V,
Schwärzler C, Schwarz H, Beug H, Günthert U and Huber LA: Analysis
of CD44-containing lipid rafts: Recruitment of annexin II and
stabilization by the actin cytoskeleton. J Cell Biol. 146:843–854.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang Y, Yan CH and Fu SB: The cloning and
expression of apoptosis associated gene ANNEXIN A2 induced by p53
gene. Zhonghua Yi Xue Yi Chuan Xue Za Zhi. 22:661–664. 2005.In
Chinese. PubMed/NCBI
|
|
13
|
Waters KM, Stenoien DL, Sowa MB, von
Neubeck C, Chrisler WB, Tan R, Sontag RL and Weber TJ: Annexin A2
modulates radiation-sensitive transcriptional programming and cell
fate. Radiat Res. 179:53–61. 2013. View Article : Google Scholar
|
|
14
|
Coméra C, Rothhut B and Russo-Marie F:
Identification and characterization of phospholipase A2 inhibitory
proteins in human mononuclear cells. Eur J Biochem. 188:139–146.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fan X, Krahling S, Smith D, Williamson P
and Schlegel RA: Macrophage surface expression of annexins I and II
in the phagocytosis of apoptotic lymphocytes. Mol Biol Cell.
15:2863–2872. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Choi KS, Fitzpatrick SL, Filipenko NR,
Fogg DK, Kassam G, Magliocco AM and Waisman DM: Regulation of
plasmin-dependent fibrin clot lysis by annexin II heterotetramer. J
Biol Chem. 276:25212–25221. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ling Q, Jacovina AT, Deora A, Febbraio M,
Simantov R, Silverstein RL, Hempstead B, Mark WH and Hajjar KA:
Annexin II regulates fibrin homeostasis and neoangiogenesis in
vivo. J Clin Invest. 113:38–48. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen H, Xu C, Jin Q and Liu Z: S100
protein family in human cancer. Am J Cancer res. 4:89–115.
2014.PubMed/NCBI
|
|
19
|
Liu Y, Myrvang HK and Dekker IV: Annexin
A2 complexes with S100 proteins: Structure, function and
pharmacological manipulation. Br J Pharmacol. 172:1664–1676. 2015.
View Article : Google Scholar :
|
|
20
|
Monastyrskaya K, Babiychuk EB, Hostettler
A, Rescher U and Draeger A: Annexins as intracellular calcium
sensors. Cell Calcium. 41:207–219. 2007. View Article : Google Scholar
|
|
21
|
Myrvang HK, Guo X, Li C and Dekker LV:
Protein interactions between surface annexin A2 and S100A10 mediate
adhesion of breast cancer cells to microvascular endothelial cells.
FEBS Lett. 587:3210–3215. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sharma MC and Sharma M: The role of
annexin II in angiogenesis and tumor progression: A potential
therapeutic target. Curr Pharm Des. 13:3568–3575. 2007. View Article : Google Scholar
|
|
23
|
Bolon I, Zhou HM, Charron Y, Wohlwend A
and Vassalli JD: Plasminogen mediates the pathological effects of
urokinase-type plasminogen activator overexpression. Am J Pathol.
164:2299–2304. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Madureira PA, Bharadwaj AG, Bydoun M,
Garant K, O'Connell P, Lee P and Waisman DM: Cell surface protease
activation during RAS transformation: Critical role of the
plasminogen receptor, S100A10. Oncotarget. 7:47720–47737. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhuang H, Tan M, Liu J, Hu Z, Liu D, Gao
J, Zhu L and Lin B: Human epididymis protein 4 in association with
Annexin II promotes invasion and metastasis of ovarian cancer
cells. Mol Cancer. 13:2432014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang M and Huang CZ: Mitogen-activated
protein kinase signaling pathway and invasion and metastasis of
gastric cancer. World J Gastroenterol. 21:11673–11679. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sulzmaier FJ, Jean C and Schlaepfer DD:
FAK in cancer: Mechanistic findings and clinical applications. Nat
Rev Cancer. 14:598–610. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
O'Reilly MS, Holmgren L, Shing Y, Chen C,
Rosenthal RA, Moses M, Lane WS, Cao Y, Sage EH and Folkman J:
Angiostatin: A novel angiogenesis inhibitor that mediates the
suppression of metastases by a lewis lung carcinoma. Cell.
79:315–328. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wahl ML, Kenan DJ, Gonzalez-Gronow M and
Pizzo SV: Angiostatin's molecular mechanism: Aspects of specificity
and regulation elucidated. J Cell Biochem. 96:242–261. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mai J, Waisman DM and Sloane BF: Cell
surface complex of cathepsin B/annexin II tetramer in malignant
progression. Biochim Biophys Acta. 1477:215–230. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mai J, Finley RL Jr, Waisman DM and Sloane
BF: Human procathepsin B interacts with the annexin II tetramer on
the surface of tumor cells. J Biol Chem. 275:12806–12812. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Roodman GD: Genes associate with abnormal
bone cell activity in bone metastasis. Cancer Metastasis Rev.
31:569–578. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jung Y, Shiozawa Y, Wang J, Patel LR,
Havens AM, Song J, Krebsbach PH, Roodman GD and Taichman RS:
Annexin-2 is a regulator of stromal cell-derived factor-1/CXCL12
function in the hematopoietic stem cell endosteal niche. Exp
Hematol. 39:151–166.e1. 2011. View Article : Google Scholar
|
|
34
|
Jung Y, Wang J, Lee E, McGee S, Berry JE,
Yumoto K, Dai J, Keller ET, Shiozawa Y and Taichman RS: Annexin
2-CXCL12 interactions regulate metastatic cell targeting and growth
in the bone marrow. Mol Cancer Res. 13:197–207. 2015. View Article : Google Scholar :
|
|
35
|
D'Souza S, Kurihara N, Shiozawa Y, Joseph
J, Taichman R, Galson DL and Roodman GD: Annexin II interactions
with the annexin II receptor enhance multiple myeloma cell adhesion
and growth in the bone marrow microenvironment. Blood.
119:1888–1896. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Spijkers-Hagelstein JA, Mimoso Pinhanços
S, Schneider P, Pieters R and Stam RW: Src kinase-induced
phosphorylation of annexin A2 mediates glucocorticoid resistance in
MLL-rearranged infant acute lymphoblastic leukemia. Leukemia.
27:1063–1071. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gopalakrishnapillai A, Kolb EA, Dhanan P,
Mason RW, Napper A and Barwe SP: Disruption of Annexin II/p11
interaction suppresses leukemia cell Binding, homing and
engraftment, and sensitizes the leukemia cells to chemotherapy.
PLoS One. 10:e01405642015. View Article : Google Scholar
|
|
38
|
Menell JS, Cesarman GM, Jacovina AT,
McLaughlin MA, Lev EA and Hajjar KA: Annexin II and bleeding in
acute promyelocytic leukemia. N Engl J Med. 340:994–1004. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu Y, Wang Z, Jiang M, Dai L, Zhang W, Wu
D and Ruan C: The expression of annexin II and its role in the
fibrinolytic activity in acute promyelocytic leukemia. Leuk Res.
5:879–84. 2011. View Article : Google Scholar
|
|
40
|
Olwill SA, McGlynn H, Gilmore WS and
Alexander HD: All-trans retinoic acid-induced downregulation of
annexin II expression in myeloid leukaemia cell lines is not
confined to acute promyelocytic leukaemia. Br J Haematol.
131:258–264. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang J, Cheng TT, Wang LN, Chen W and Li
X: The expression and clinical significance of Annexin II in
clear-cell renal cell carcinoma. Xi Bao Yu Fen Zi Mian Yi Xue Za
Zhi. 28:59–62. 2012.In Chinese. PubMed/NCBI
|
|
42
|
Ohno Y, Izumi M, Kawamura T, Nishimura T,
Mukai K and Tachibana M: Annexin II represents metastatic potential
in clear-cell renal cell carcinoma. Br J Cancer. 101:287–294. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sharma MR, Koltowski L, Ownbey RT,
Tuszynski GP and Sharma MC: Angiogenesis-associated protein annexin
II in breast cancer: Selective expression in invasive breast cancer
and contribution to tumor invasion and progression. Exp Mol Pathol.
81:146–156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shetty PK, Thamake SI, Biswas S, Johansson
SL and Vishwanatha JK: Reciprocal regulation of annexin A2 and EGFR
with Her-2 in Her-2 negative and herceptin-resistant breast cancer.
PloS One. 7:442992012. View Article : Google Scholar
|
|
45
|
Sharma M, Blackman MR and Sharma MC:
Antibody directed neutralization of annexin II (ANX II) inhibits
neoangiogenesis and human breast tumor growth in a xenograft model.
Exp Mol Pathol. 92:175–184. 2012. View Article : Google Scholar
|
|
46
|
Jin L, Shen Q, Ding S, Jiang W, Jiang L
and Zhu X: Immunohistochemical expression of Annexin A2 and S100A
proteins in patients with bulky stage IB-IIA cervical cancer
treated with neoadjuvant chemotherapy. Gynecol Oncol. 126:140–146.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Choi CH, Chung JY, Chung EJ, Sears JD, Lee
JW, Bae DS and Hewitt SM: Prognostic significance of annexin A2 and
annexin A4 expression in patients with cervical cancer. BMC Cancer.
16:4482016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Woodham AW, Da Silva DM, Skeate JG, Raff
AB, Ambroso MR, Brand HE, Isas JM, Langen R and Kast WM: The
S100A10 subunit of the annexin A2 heterotetramer facilitates
L2-mediated human papillomavirus infection. PloS One. 7:e435192012.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xiu D, Liu L, Qiao F, Yang H, Cui L and
Liu G: Annexin A2 coordinates STAT3 to regulate the invasion and
migration of colorectal cancer cells in vitro. Gastroenterol Res
Pract. 2016:pp. 35214532016, https://doi.org/10.1155/2016/3521453.
View Article : Google Scholar
|
|
50
|
Xing R, He H, He Y, Feng Y, Zhang C, Wu H,
Sun M, Yu X, Liu Y, Song X, et al: ANXA2 remodels the
microstructures of caco2 cells. Cell Mol Biol. 59(59 Suppl):
OL1848–54. 2013.PubMed/NCBI
|
|
51
|
Emoto K, Yamada Y, Sawada H, Fujimoto H,
Ueno M, Takayama T, Kamada K, Naito A, Hirao S and Nakajima Y:
Annexin II overexpression correlates with stromal tenascin-C
overexpression: A prognostic marker in colorectal carcinoma.
Cancer. 92:1419–1426. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tristante E, Martínez CM, Jiménez S, Mora
L, Carballo F, Martínez-Lacaci I and de Torre-Minguela C:
Association of a characteristic membrane pattern of annexin A2 with
high invasiveness and nodal status in colon adenocarcinoma. Transl
Res. 166:196–206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang T, Peng H, Wang J, Yang J, Nice EC,
Xie K and Huang C: Prognostic and diagnostic significance of
annexin A2 in colorectal cancer. Colorectal Dis. 15:e373–e381.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gurluler E, Guner OS, Tumay LV, Turkel
Kucukmetin N, Hizli B and Zorluoglu A: Serum annexin A2 levels in
patients with colon cancer in comparison to healthy controls and in
relation to tumor pathology. Med Sci Monit. 20:1801–1807. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Deng L, Gao Y, Li X, Cai M, Wang H, Zhuang
H, Tan M, Liu S, Hao Y and Lin B: Expression and clinical
significance of annexin A2 and human epididymis protein 4 in
endometrial carcinoma. J Exp Clin Cancer res. 34:962015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Alonso-Alconada L, Santacana M,
Garcia-Sanz P, Muinelo-Romay L, Colas E, Mirantes C, Monge M, Cueva
J, Oliva E, Soslow RA, et al: Annexin-A2 as predictor biomarker of
recurrent disease in endometrial cancer. Int J Cancer.
136:1863–1873. 2015. View Article : Google Scholar
|
|
57
|
Zhang Q, Ye Z, Yang Q, He X, Wang H and
Zhao Z: Upregulated expression of annexin II is a prognostic marker
for patients with gastric cancer. World J Surg Oncol. 10:1032012.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Han Y, Ye J, Dong Y, Xu Z and du Q:
Expression and significance of annexin A2 in patients with gastric
adenocarcinoma and the association with E-cadherin. Exp Ther Med.
10:549–554. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Gao H, Yu B, Yan Y, Shen J, Zhao S, Zhu J,
Qin W and Gao Y: Correlation of expression levels of ANXA2, GAM1
and CALR with glioma grade and prognosis. J Neurosurg. 118:846–853.
2013. View Article : Google Scholar
|
|
60
|
Onishi M, Ichikawa T, Kurozumi K, Inoue S,
Maruo T, Otani Y, Fujii K, Ishida J, Shimazu Y, Yoshida K, et al:
Annexin A2 regulates angiogenesis and invasion phenotypes of
malignant glioma. Brain Tumor Pathol. 32:184–194. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhai H, Acharya S, Gravanis I, Mehmood S,
Seidman RJ, Shroyer KR, Hajjar KA and Tsirka SE: Annexin A2
promotes glioma cell invasion and tumor progression. J Neurosci.
31:14346–14360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mohammad HS, Kurokohchi K, Yoneyama H,
Tokuda M, Morishita A, Jian G, Shi L, Murota M, Tani J, Kato K, et
al: Annexin A2 expression and phosphorylation are up-regulated in
hepatocellular carcinoma. Int J Oncol. 33:1157–1163.
2008.PubMed/NCBI
|
|
63
|
Bai DS, Wu C, Yang LX, Zhang C, Zhang PF,
He YZ, Cai JB, Song ZJ, Dong ZR, Huang XY, et al: UBAP2 negatively
regulates the invasion of hepatocellular carcinoma cell by
ubiquitinating and degradating Annexin A2. Oncotarget.
7:32946–32955. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhang H, Yao M, Wu W, Qiu L, Sai W, Yang
J, Zheng W, Huang J and Yao D: Up-regulation of annexin A2
expression predicates advanced clinicopathological features and
poor prognosis in hepatocellular carcinoma. Tumour Biol.
36:9373–9383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhang HJ, Yao DF, Yao M, Huang H, Wang L,
Yan MJ, Yan XD, Gu X, Wu W and Lu SL: Annexin A2 silencing inhibits
invasion, migration, and tumorigenic potential of hepatoma cells.
World J Gastroenterol. 19:3792–3801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang HJ, Yao DF, Yao M, Huang H, Wu W,
Yan MJ, Yan XD and Chen J: Expression characteristics and
diagnostic value of annexin A2 in hepatocellular carcinoma. World J
Gastroenterol. 18:5897–5904. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Cui HY, Wang SJ, Miao JY, Fu ZG, Feng F,
Wu J, Yang XM, Chen ZN and Jiang JL: CD147 regulates cancer
migration via direct interaction with Annexin A2 and
DOCK3-β-catenin-WAVE2 signaling. Oncotarget. 7:5613–5629. 2016.
View Article : Google Scholar
|
|
68
|
Dong Z, Yao M, Zhang H, Wang L, Huang H,
Yan M, Wu W and Yao D: Inhibition of Annexin A2 gene transcription
is a promising molecular target for hepatoma cell proliferation and
metastasis. Oncol Lett. 7:28–34. 2014.
|
|
69
|
Sun Y, Gao G, Cai J, Wang Y, Qu X, He L,
Liu F, Zhang Y, Lin K, Ma S, et al: Annexin A2 is a discriminative
serological candidate in early hepatocellular carcinoma.
Carcinogenesis. 34:595–604. 2013. View Article : Google Scholar :
|
|
70
|
Wang YX, Lv H, Li ZX, Li C and Wu XY:
Effect of shRNA mediated down-regulation of Annexin A2 on
biological behavior of human lung adencarcinoma cells A549. Pathol
Oncol Res. 18:183–190. 2012. View Article : Google Scholar
|
|
71
|
Jia JW, Li KL, Wu JX and Guo SL: Clinical
significance of annexin II expression in human non-small cell lung
cancer. Tumour Biol. 34:1767–1771. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Luo CH, Liu QQ, Zhang PF, Li MY, Chen ZC
and Liu YF: Prognostic significance of annexin II expression in
non-small cell lung cancer. Clin Transl Oncol. 15:938–946. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Cole SP, Pinkoski MJ, Bhardwaj G and
Deeley RG: Elevated expression of annexin II (lipocortin II, 36) in
a multidrug resistant small cell lung cancer cell line. Br J
Cancer. 65:498–502. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Bao H, Jiang M, Zhu M, Sheng F, Ruan J and
Ruan C: Overexpression of Annexin II affects the proliferation,
apoptosis, invasion and production of proangiogenic factors in
multiple myeloma. Int J Hematol. 90:177–185. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lokman NA, Pyragius CE, Ruszkiewicz A,
Oehler MK and Ricciardelli C: Annexin A2 and S100A10 are
independent predictors of serous ovarian cancer outcome. Transl
Res. 171:83–95.e2. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Deng Y, Chen C, Hua M, Xi Q, Liu R, Yang
S, Liu J, Zhong J, Tang M, Lu S, et al: Annexin A2 plays a critical
role in epithelial ovarian cancer. Arch Gynecol Obstet.
292:175–182. 2015. View Article : Google Scholar
|
|
77
|
Lokman NA, Elder AS, Ween MP, Pyragius CE,
Hoffmann P, Oehler MK and Ricciardelli C: Annexin A2 is regulated
by ovarian cancer-peritoneal cell interactions and promotes
metastasis. Oncotarget. 4:1199–1211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Pi F, Zhang H, Li H, Thiviyanathan V,
Gorenstein DG, Sood AK and Guo P: RNA nanoparticles harboring
annexin A2 aptamer can target ovarian cancer for tumor-specific
doxorubicin delivery. Nanomedicine. 13:1183–1193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Foley K, Rucki AA, Xiao Q, Zhou D, Leubner
A, Mo G, Kleponis J, Wu AA, Sharma R, Jiang Q, et al: Semaphorin 3D
autocrine signaling mediates the metastatic role of annexin A2 in
pancreatic cancer. Sci Signal. 8:ra77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zheng L, Foley K, Huang L, Leubner A, Mo
G, Olino K, Edil BH, Mizuma M, Sharma R, Le DT, et al: Tyrosine 23
phosphorylation-dependent cell-surface localization of annexin A2
is required for invasion and metastases of pancreatic cancer. PloS
One. 6:e193902011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Nedjadi T, Kitteringham N, Campbell F,
Jenkins RE, Park BK, Navarro P, Ashcroft F, Tepikin A, Neoptolemos
JP and Costello E: S100A6 binds to annexin 2 in pancreatic cancer
cells and promotes pancreatic cancer cell motility. Br J Cancer.
101:1145–1154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Huang YK, Liu H, Wang XZ and Zhu S:
Annexin A2 and CD105 expression in pancreatic ductal adenocarcinoma
is associated with tumor recurrence and prognosis. Asian Pac J
Cancer Prev. 15:9921–9926. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Jung H, Kim JS, Kim WK, Oh KJ, Kim JM, Lee
HJ, Han BS, Kim DS, Seo YS, Lee SC, et al: Intracellular annexin A2
regulates NF-κB signaling by binding to the 50 subunit:
Implications for gemcitabine resistance in pancreatic cancer. Cell
Death Dis. 6:e16062015. View Article : Google Scholar
|
|
84
|
Takano S, Togawa A, Yoshitomi H, Shida T,
Kimura F, Shimizu H, Yoshidome H, Ohtsuka M, Kato A, Tomonaga T, et
al: Annexin II overexpression predicts rapid recurrence after
surgery in pancreatic cancer patients undergoing
gemcitabine-adjuvant chemotherapy. Ann Surg Oncol. 15:3157–3168.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zheng L and Jaffee EM: Annexin A2 is a new
antigenic target for pancreatic cancer immunotherapy.
OncoImmunology. 1:112–114. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Keklikoglou I, Hosaka K, Bender C, Bott A,
Koerner C, Mitra D, Will R, Woerner A, Muenstermann E, Wilhelm H,
et al: MicroRNA-206 functions as a pleiotropic modulator of cell
proliferation, invasion and lymphangiogenesis in pancreatic
adenocarcinoma by targeting ANXA2 and KrAs genes. Oncogene.
34:4867–4878. 2015. View Article : Google Scholar :
|
|
87
|
Zhang Q, Zhao Z, Ma Y, Wang H, Ma J, He X
and Zhang D: Combined expression of S100A4 and Annexin A2 predicts
disease progression and overall survival in patients with
urothelial carcinoma. Urol Oncol. 32:798–805. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Feng JG, Liu Q, Qin X, Geng YH, Zheng ST,
Liu T, Sheyhidin I and Lu XM: Clinicopathological pattern and
Annexin A2 and CDC42 status in patients presenting with
differentiation and lymphnode metastasis of esophageal squamous
cell carcinomas. Mol Biol Rep. 39:1267–1274. 2012. View Article : Google Scholar
|
|
89
|
Li X, Zheng S, Liu Q, Liu T, Liang M, Gao
X, Lu M, Sheyhidin I and Lu X: Under-expression of annexin A2 is
associated with Kazakh's esophageal squamous cell carcinoma. Mol
Carcinog. 54:779–788. 2015. View Article : Google Scholar
|
|
90
|
Rodrigo Tapia JP, Pena Alonso E,
García-Pedrero JM, Florentino Fresno M, Suárez Nieto C, Owen Morgan
R and Fernández MP: Annexin A2 expression in head and neck squamous
cell carcinoma. Acta Otorrinolaringol Esp. 58:257–262. 2007.In
Spanish. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yee DS, Narula N, Ramzy I, Boker J,
Ahlering TE, Skarecky DW and Ornstein DK: Reduced annexin II
protein expression in high-grade prostatic intraepithelial
neoplasia and prostate cancer. Arch Pathol Lab Med. 131:902–908.
2007.PubMed/NCBI
|
|
92
|
Ding T, Yang L, Wang Y, Yuan J, Chen T and
Cai X: Downregulation of annexin II in prostate cancer is
associated with Gleason score, recurrence, metastasis and poor
prognosis. Mol Med Rep. 3:781–787. 2010.
|
|
93
|
Smitherman AB, Mohler JL, Maygarden SJ and
Ornstein DK: Expression of annexin I, II and VII proteins in
androgen stimulated and recurrent prostate cancer. J Urol.
171:916–920. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Sharma M, Blackman MR and Sharma MC:
Antibody-directed neutralization of annexin II (ANX II) inhibits
neoangiogenesis and human breast tumor growth in a xenograft model.
Exp Mol Pathol. 92:175–184. 2012. View Article : Google Scholar
|
|
95
|
Singh P, Wu H, Clark C and Owlia A:
Annexin II binds progastrin and gastrin-like peptides, and mediates
growth factor effects of autocrine and exogenous gastrins on colon
cancer and intestinal epithelial cells. Oncogene. 26:425–440. 2007.
View Article : Google Scholar
|
|
96
|
Reeves SA, Chavez-Kappel C, Davis R,
Rosenblum M and Israel MA: Developmental regulation of annexin II
(Lipocortin 2) in human brain and expression in high grade glioma.
Cancer Res. 52:6871–6876. 1992.PubMed/NCBI
|
|
97
|
Zhong LP, Wei KJ, Yang X, Zhang L, Zhou
XJ, Pan HY, Li J, Chen WT and Zhang ZY: Increased expression of
Annexin A2 in oral squamous cell carcinoma. Arch Oral Biol.
54:17–25. 2009. View Article : Google Scholar
|
|
98
|
Sharma MC, Tuszynski GP, Blackman MR and
Sharma M: Long-term efficacy and downstream mechanism of
anti-annexinA2 monoclonal antibody (anti-ANX A2 mAb) in a
pre-clinical model of aggressive human breast cancer. Cancer Lett.
373:27–35. 2016. View Article : Google Scholar : PubMed/NCBI
|