Introduction
The transcription factor p53 is activated in
response to various stresses, including DNA damage, nutrient
deprivation, oncogene activation and hypoxia (1). p53 is a well-established tumor
suppressor and 'guardian of the genome' that induces cell cycle
arrest and apoptosis by activating downstream target genes
(2). However, p53 is mutated in
around half of all human cancers (3), leading to loss of its tumor
suppressor activity and, in certain cases, gain-of-function
activities (4) that promote cell
proliferation, tumor development, and drug resistance (5,6).
Thus, mutant p53 has become an important target for the development
of anticancer treatments. A number of small molecule targeted to
mutant p53 have been described, most of which promote the
proteasomal or autophagic degradation of mutant p53 or restoration
of wild-type p53 function. The current study identified
andrographolide (ANDRO), a labdane diterpenoid isolated from the
Chinese herb Andrographis paniculata, as a mutant p53
suppressor from a small-molecule screen, and investigated the
effects of ANDRO on human cancer cell lines harboring mutant or
wild-type p53. ANDRO suppressed the activity of mutant p53 and the
growth of cancer cells via induction of the heat shock protein
(Hsp) 70, resulting in enhanced proteasomal degradation of mutant
p53. ANDRO also effectively reduced the growth of mutant p53 tumors
in a mouse xenograft model.
Materials and methods
Cell lines and culture conditions
All cell lines were obtained from RIKEN BioResource
Center (Tsukuba, Japan). PANC-1 (human pancreatic cancer) and
HCT116 (human colorectal cancer) were maintained in Dulbecco's
modified Eagle's medium (DMEM; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). HuCCT1 (human bile duct cancer) and MKN45
(human gastric cancer) were maintained in RPMI-1640 medium
(Sigma-Aldrich; Merck KGaA). DMEM and RPMI-1640 media were
supplemented with 10% fetal bovine serum (Sigma-Aldrich; Merck
KGaA) and 100 µg/ml kanamycin (Meiji Seika Pharma Co., Ltd.,
Tokyo, Japan). The cell lines were maintained at 37°C in a
humidified atmosphere containing 20% O2 and 5%
CO2.
Chemicals
ANDRO, acacetin, berberine, honokiol, pentamidine,
3-(5′-hydroxymethyl-2′-furyl)-1-benzyl indazole (YC-1), and
2-mercaptoethanol (2-ME) were purchased from Sigma-Aldrich (Merck
KGaA). Noscapine and irinotecan (CPT-11) were obtained from Tokyo
Chemical Industry Co., Ltd. (Tokyo, Japan). The compounds were
dissolved in dimethyl sulfoxide (DMSO) and stored at -20°C prior to
use. For experiments, the compounds were diluted in medium to the
appropriate doses immediately prior to use. These compounds were
used as indicated or at the following concentrations: ANDRO, 100
µM; acacetin, 100 µM; berbeline, 50 µM;
honokiol, 50 µM; noscapine, 100 µM; pentamigine, 200
µM; YC-1, 100 µM; 2-ME, 30 µM.
Immunoblotting
Cells were plated in 60 mm dishes and allowed to
grow to 60–70% confluence. The cells were then treated with the
appropriate reagents for 18 h and lysed in lysis buffer consisting
of 150 mM NaCl, 50 mM Tris-HCl (pH 7.6), protease inhibitor
cocktail mix (Roche Diagnostic GmbH, Mannheim, Germany) and
phenylmethylsulfonyl fluoride. Proteins (20–37.5 µg/well)
were separated by SDS-polyacrylamide gel electrophoresis on 10%
gels and electroblotted onto Immobilon membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Membranes were blocked with
5% non-fat milk in 150 mM NaCl, 25 mM Tris pH 7.5 and 0.1% Tween-20
(TBS-T) and probed overnight at 4°C with primary antibodies diluted
1:500–1:1,000 in the same buffer. Membranes were washed with TBS-T
twice for 5 min each and then incubated with secondary antibody
(1:1,000 dilution; goat anti-rabbit and goat anti-mouse
IgG-horseradish peroxidase; cat nos. sc-2004 and sc-2005; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) for 30 min at room
temperature. The blots were developed with enhanced
chemiluminescence reagents (Amersham ECL Prime; GE Healthcare,
Chicago, IL, USA) and analyzed with a Fujifilm LAS-3000 imaging
system (Fujifilm Corporation, Tokyo, Japan). The primary antibodies
were specific for: p53 (1:1,000 dilution; clone DO-1; cat no.
sc-126), p21 (1:500 dilution; clone C-19; cat. no. sc-397), E3
ubiquitin-protein ligase Mdm2 (1:1,000 dilution; MDM2; clone SMP14;
cat. no. sc-965), E3 ubiquitin-protein ligase CHIP (1:500 dilution;
CHIP; clone I-16; cat. no. sc-33264) signal transducer and
activator of transcription-3 (STAT-3; 1:1,000 dilution; clone C-20;
cat. no. sc-482) and Hsp40 (1:1,000 dilution; clone C-20; cat. no.
sc1800), all from Santa Cruz Biotechnology, Inc.; Bcl-2-binding
component 3 (PUMA; 1:1,000 dilution; cat. no. 7467), caspase-3
(1:1,000 dilution; cat. no. 9662) and Hsp70 (1:1,000 dilution; cat.
no. 4873), all Cell Signaling Technology, Inc. (Danvers, MA, USA);
phorbol-12-myristate-13-acetate-induced protein 1 (NOXA; 1:1,000
dilution; cat. no. 114C307; Calbiochem; Merck KGaA); Hsp90 (1:1,000
dilution; cat. no. AC88; Enzo Life Sciences, Inc., Farmingdale, NY,
USA); cyclin D1 (1:1,000 dilution; cat. no. ab134175; Abcam,
Cambridge, UK) and β-actin (1:1,000 dilution; cat. no. A2228;
Sigma-Aldrich; Merck KGaA).
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted using a Qiagen RNA
extraction kit, converted to cDNA (37°C for 15 min for RT, 98°C for
5 min to terminate the reaction and 12°C hold) using an iScript
cDNA Synthesis kit (Bio-Rad Laboratories, Inc.), and analyzed by
qPCR. The gene-specific primer sequences were: p53 forward,
5′-GCCCAACAACACCAGCTCCT-3 and reverse, 5′-CCTGGGCATCCTTGAGTTCC-3′;
p21 forward, 5′-GGCGGCAGACCAGCATGACAGATT-3′ and reverse,
5′-GCAGGGGGCGGCCAGGGTAT-3′; PUMA forward, 5′-GACCTCAACGCACAGTAC
GAG-3′ and reverse, 5′-AGGAGTCCCATGATGAGATTGT-3′; NOXA forward,
5′-ATTACCGCTGGCCTACTGTG-3′ and reverse, 5′-GTGCTGAGTTGGCACTGAAA-3′;
heat shock transcription factor 1 forward,
5′-GCCTTCCTGACCAAGCTGT-3′ and reverse, 5′-AAGTACTTGGGCAGCACCTC-3′;
Hsp40 forward, 5′-GGCTTCACCAACGTGAACTT-3′ and reverse,
5′-CGCTTGTGGGAGATTTTCAT-3′; Hsp70 forward,
5′-CAAGATCACCATCACCAACG-3′ and reverse, 5′-TCGTCCTCCGCTTTGTACTT-3′;
and Hsp90 forward, 5′-GCAGAAATTGCCCAACTCAT-3′ and reverse,
5′-AAGGGTCTGTCAGGCTCTCA-3′. qPCR was performed using a CFX Connect™
Real-Time PCR Detection System with PrimePCR™ SYBR®
Green Assay reagents (both from Bio-Rad Laboratories, Inc.)
according to the manufacturer's instructions. After performing a
denaturation step at 95°C for 3 min, PCR amplification was
conducted with 45 cycles of 15 sec of denaturation at 95°C, 5 sec
of annealing at 60°C and 10 sec of extension at 72°C. Expression of
the gene of interest was normalized to β-actin forward,
ACTCTTCCAGCCTTCCTTCC and reverse, GACAGCACTGTGTTGGCGTA, mRNA levels
by 2−ΔΔCq method (7).
All experiments were performed in triplicate.
Cell viability assay
Cell viability was determined using the
Sulforhodamine B-based In Vitro Toxicology Assay kit
(Sigma-Aldrich; Merck KGaA). Cells were seeded in 6-well plates and
allowed to reach 60–70% confluency. Cells were then treated with
ANDRO and incubated for a further 24 h. Cell staining and
quantification of viability were performed according to the
manufacturer's instructions. All experiments were performed in
duplicate.
Immunoprecipitation
HuCCT1 and PANC-1 cells were seeded into 150 mm
dishes. After 24 h, the cells were treated with ANDRO at 100
µM and incubated for 18 h. Cells were then lysed in lysis
buffer as described above. The samples were centrifuged at 16,000 ×
g for 15 min at 4°C, and the clarified cell lysates were removed
and incubated overnight at 4°C with 15 µl Protein G
plus/Protein A-agarose (cat. no. sc-2003) and 1 µg anti-p53
antibody (clone DO-1; cat no. sc-126) (both from Santa Cruz
Biotechnology, Inc.). The beads were then washed three times with
lysis buffer. Aliquots of cell lysates (input) or
immunoprecipitates were resolved by SDS-PAGE on 10% gels,
transferred to nitrocellulose membranes, and probed with specific
primary antibodies as described above.
Flow cytometry
Cell cycle analysis was performed by flow cytometry
after staining of DNA with propidium iodide (PI). Cells were
treated with DMSO or ANDRO at the indicated concentrations for 18
h, harvested by trypsinization, washed with PBS, and fixed in 70%
ethanol overnight at 4°C. The cells were centrifuged, washed twice
with PBS, resuspended in 300 µl PBS containing 200
µg/ml of RNase (Wako Pure Chemical Industries, Ltd., Osaka,
Japan), and incubated for 30 min at 37°C. PI (50 µg/ml) was
then added and the cells were incubated in the dark for 30 min at
4°C. Finally, the cells were analyzed using a FACSVerse flow
cytometer (BD Biosciences, San Jose, CA, USA) and FlowJo 10.2
software (FlowJo LLC, Ashland, OR, USA).
RNA interference
Small interfering RNAs (siRNAs) were obtained from
Santa Cruz Biotechnology, Inc. (siControl, cat. no. sc-37007;
siHsp70, cat. no. sc-29352) and transfected into cells using
Lipofectamine RNAiMax (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol.
Animal experiments
An animal experimental protocol was reviewed and
approved by the Animal Care Committee of Saga University (Saga,
Japan; permission no. 27-039-0). A total of 60 female BALB/c mice
(4–6 week-old) were purchased from Kyudo Co., Ltd. (Saga, Japan).
The animals were maintained in an animal facility in a 12/12 h
light/dark cycle in a temperature (20°C) and humidity-controlled
environment. Food and water were freely available. HuCCT1
(5×106) and MKN45 (7×106) cells were injected
subcutaneously into the flanks of nude mice (n=6/group). Tumor size
was monitored using a vernier caliper and the volume was calculated
from the length (L) and width (W) using the formula: volume = L ×
W2 × π/6. Treatment was initiated when the tumor volume
reached 50 mm3. The mice were randomly divided into
three groups as follows: i) Untreated control (DMSO in PBS); ii)
ANDRO 10 mg/kg; and iii) CPT-11 10 mg/kg. All treatments were
administered three times per week by intraperitoneal (i.p.)
injection. Body weights were also measured three times per week.
After 11 days of treatment, the mice were euthanized. The average
weight of the mouse at the time of purchase and sacrifice was
18.02±0.47 and 17.69±0.56 g, respectively.
Statistical analysis
Data are expressed as the mean ± standard deviation,
and analyses were performed using JMP Pro 12 software (SAS
Institute, Inc., Cary, NC, USA). Differences between mean values
were evaluated using two-way analysis of variance followed by
Tukey's test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Identification of andrographolide as a
mutant p53 suppressor and inducer of p53 target genes
The pancreatic cancer cell line PANC-1, which
harbors mutant p53 R273H, was incubated for 18 h with various small
molecules to investigate their effects on the expression of p53.
The compounds were tested in pilot experiments and the
concentrations used were equivalent to the 50% inhibitory
concentrations (i.e., induced the death of ~50% of PANC-1 cells in
18 h). Among the compounds tested, only ANDRO reduced the
expression of mutant p53 protein (Fig.
1A). The structure of ANDRO is presented in Fig 1B. To examine the expression level of
p53 in with different p53 mutation statuses, HuCCT1 (bile duct
carcinoma; mutant p53 R175H), HCT116 (colon cancer; wild-type p53),
and MKN45 (gastric cancer; wild-type p53) cells were also used.
Following treatment of the cells with ANDRO for 18 h, HuCCT1 cells
also showed reduced mutant p53 protein levels, whereas expression
of wild-type p53 in HCT116 and MKN45 cells was increased (Fig. 1C). The expression of the
p53-regulated proteins; p21, which is an inhibitor of
cyclin-dependent kinase activity and regulates cell cycle
progression, and NOXA, which is pro-apoptotic protein, were also
examined. Notably, ANDRO treatment also increased the level of
these proteins in cells carrying mutant or wild-type p53 (Fig. 1C). The protein level of mutant p53
was semi-quantified by measuring relative band intensity. The
results demonstrated that the expression of mutant p53 protein was
decreased by ANDRO in a concentration-dependent manner (Fig. 1D). Finally, mRNA levels of p21,
PUMA and NOXA were also increased by ANDRO in a time-dependent
manner in the mutant p53-expressing cells HuCCT1 and PANC-1.
However, the mRNA level of mutant p53 showed no response to ANDRO
(Fig. 1E).
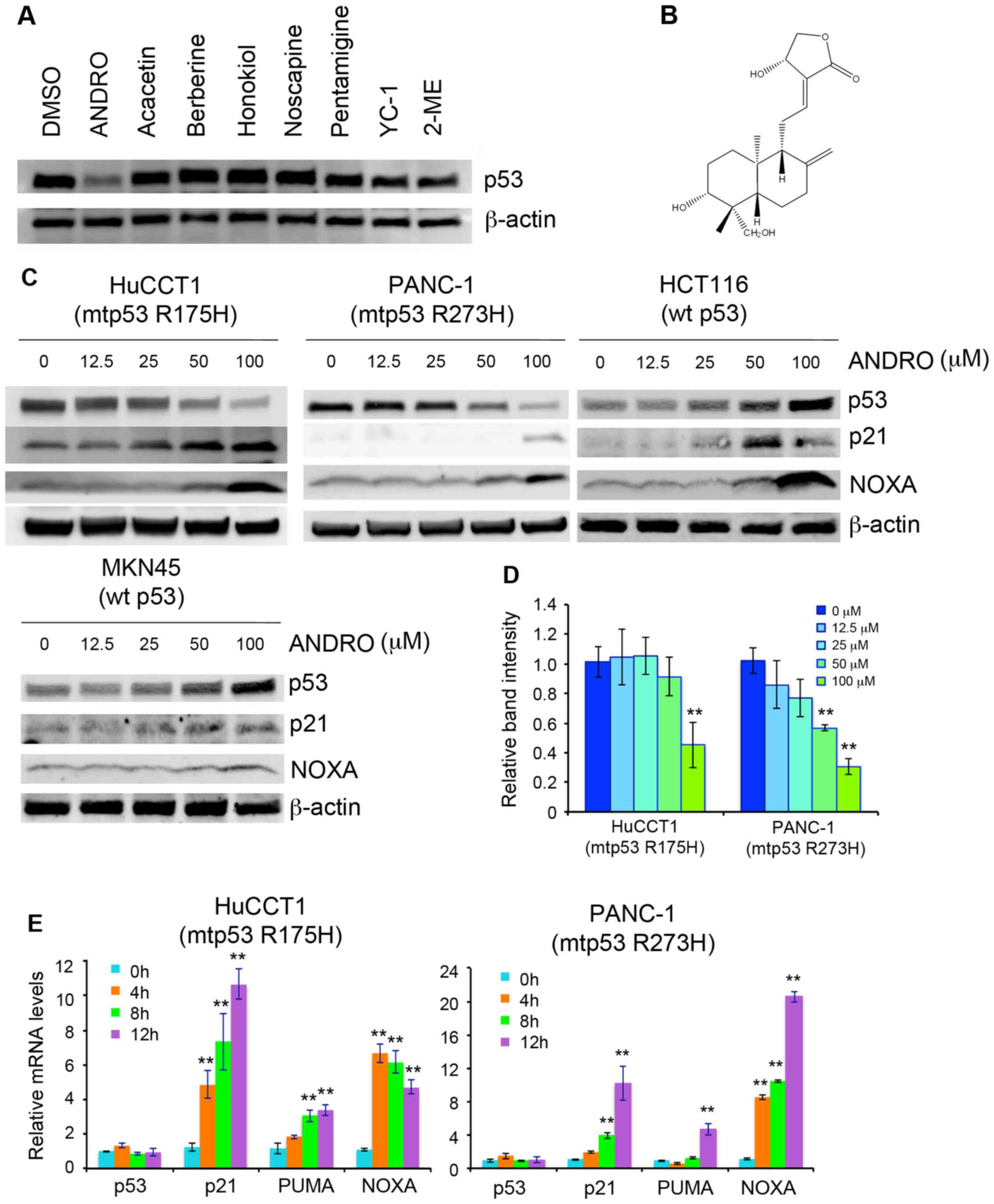 | Figure 1Identification of ANDRO as a
suppressor of mutant p53 and inducer of p53 target gene expression.
(A) Expression of mutant p53 in PANC-1 cells treated with various
small molecules. (B) Chemical structure of ANDRO. (C) Immunoblot
analysis of p53 target gene expression in cells treated with ANDRO
for 18 h. (D) The intensity of p53 band in three independent
experiments was determined by densitometric scanning using ImageJ
software and expressed relative to the control. (E) Reverse
transcription-quantitative polymerase chain reaction analysis of
p53, p21, PUMA, and NOXA mRNA levels after ANDRO treatment. Values
are presented as the mean ± standard deviation of three independent
experiments performed in triplicate. **P<0.01 vs. 0
h. DMSO, dimethyl sulfoxide; ANDRO, andrographolide; YC-1,
3-(5′-hydroxymethyl-2′-furyl)-1-benzyl indazole; 2-ME,
2-mercaptoethanol; mtp53, mutant p53; wt p53, wild-type p53; NOXA,
phorbol-12-myristate-13-acetate-induced protein 1. |
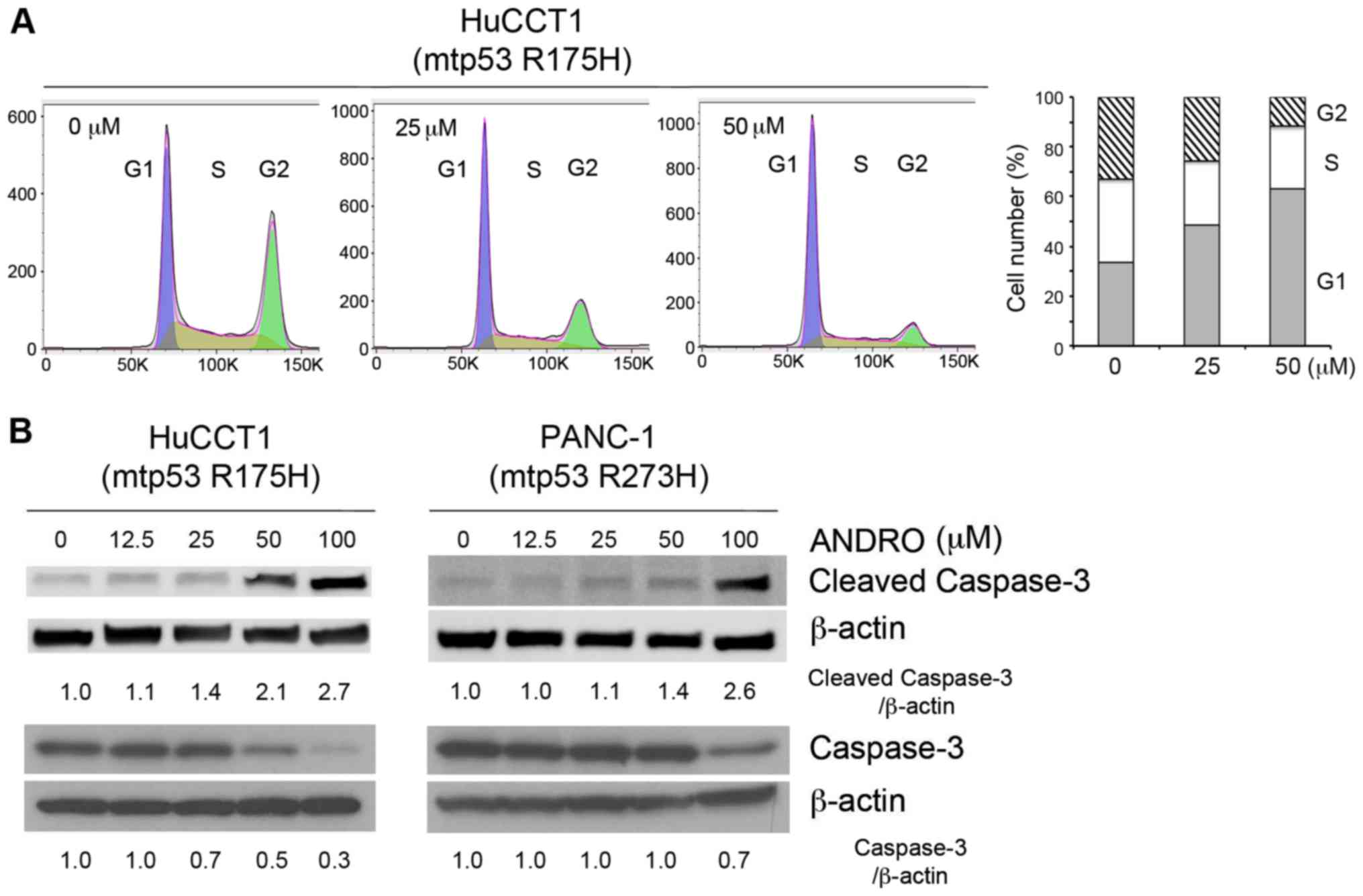
Anticancer effects of ANDRO
To determine whether ANDRO affects cell cycle
progression and/or apoptosis, the DNA profiles of PI-stained cells
were analyzed by flow cytometry or the expression of cleaved
caspase-3, a marker of apoptosis, was determined by immunoblotting
(Fig. 2A and B). In the flow
cytometric assay, treatment with ANDRO for 24 h induced arrest of
HuCCT1 cells in the G1 phase (Fig.
2A). In addition, ANDRO enhanced the expression of cleaved
caspase-3 (17 kDa) and decreased the expression of caspase-3 (35
kDa) in HuCCT1 and PANC-1 cells in a dose-dependent manner
(Fig. 2B). To verify the
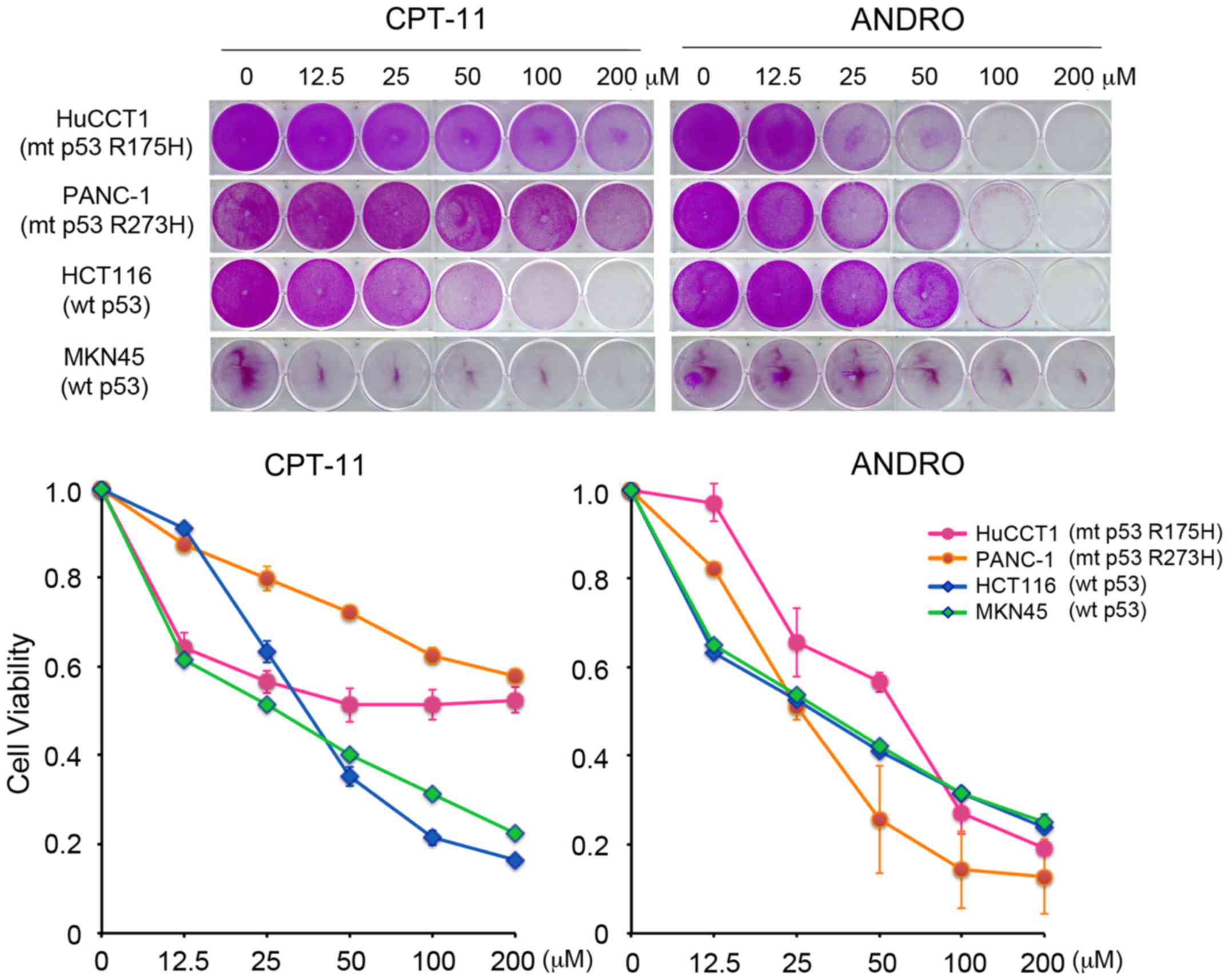
anticancer effect of ANDRO, cell viability was analyzed with a
Sulforhodamine B assay. The effects of ANDRO were compared with
that of the chemotherapeutic drug CPT-11 (irinotecan), which
induces apoptosis in a wild-type p53-dependent manner. As presented
in Fig. 3, CPT-11 effectively
reduced the viability of cell lines harboring wild-type p53,
however, it had only a limited effect on cell lines harboring
mutant p53. By contrast, ANDRO was more cytotoxic towards cells
expressing mutant p53 than those expressing wild-type p53 (Fig. 3).
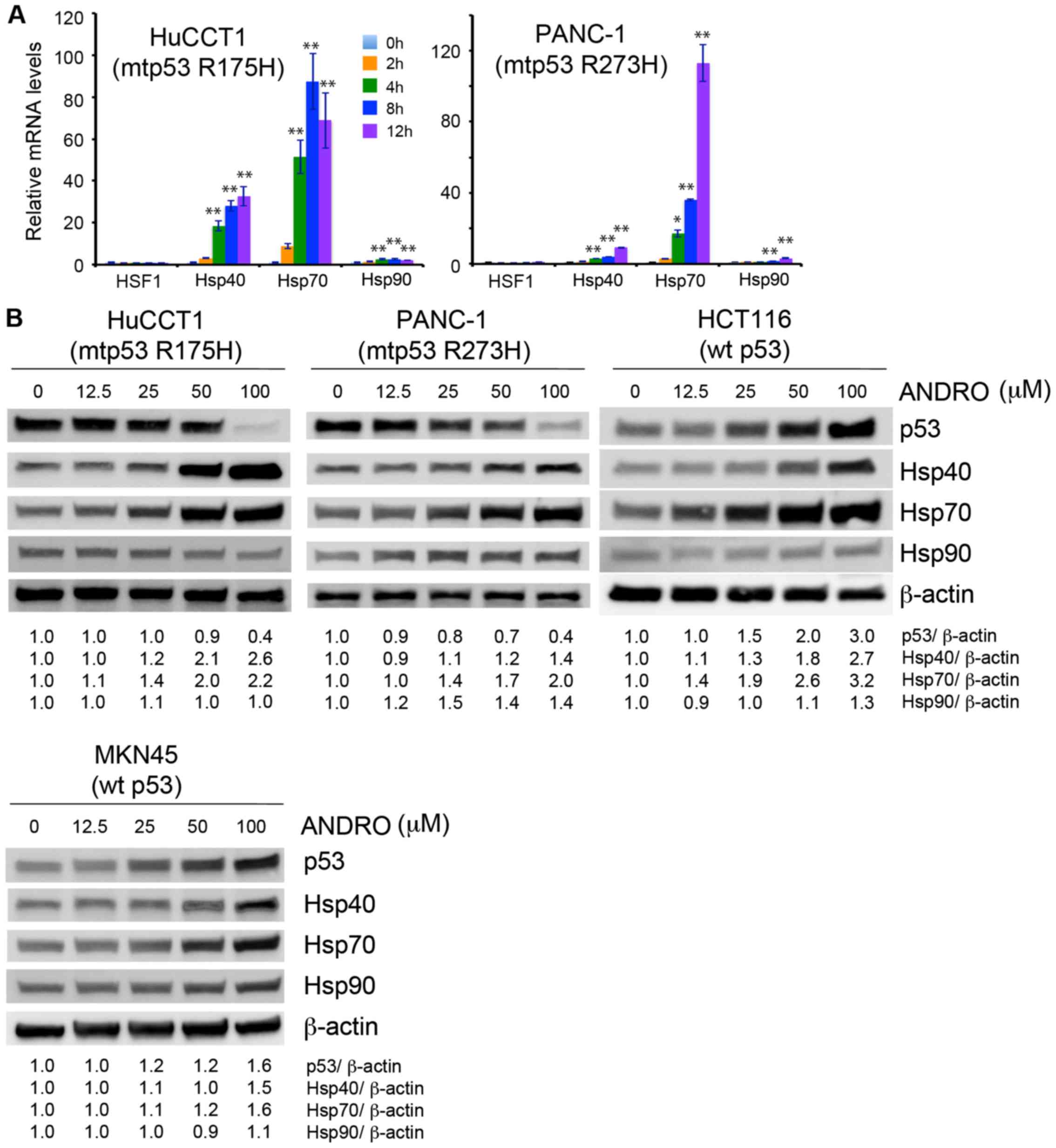
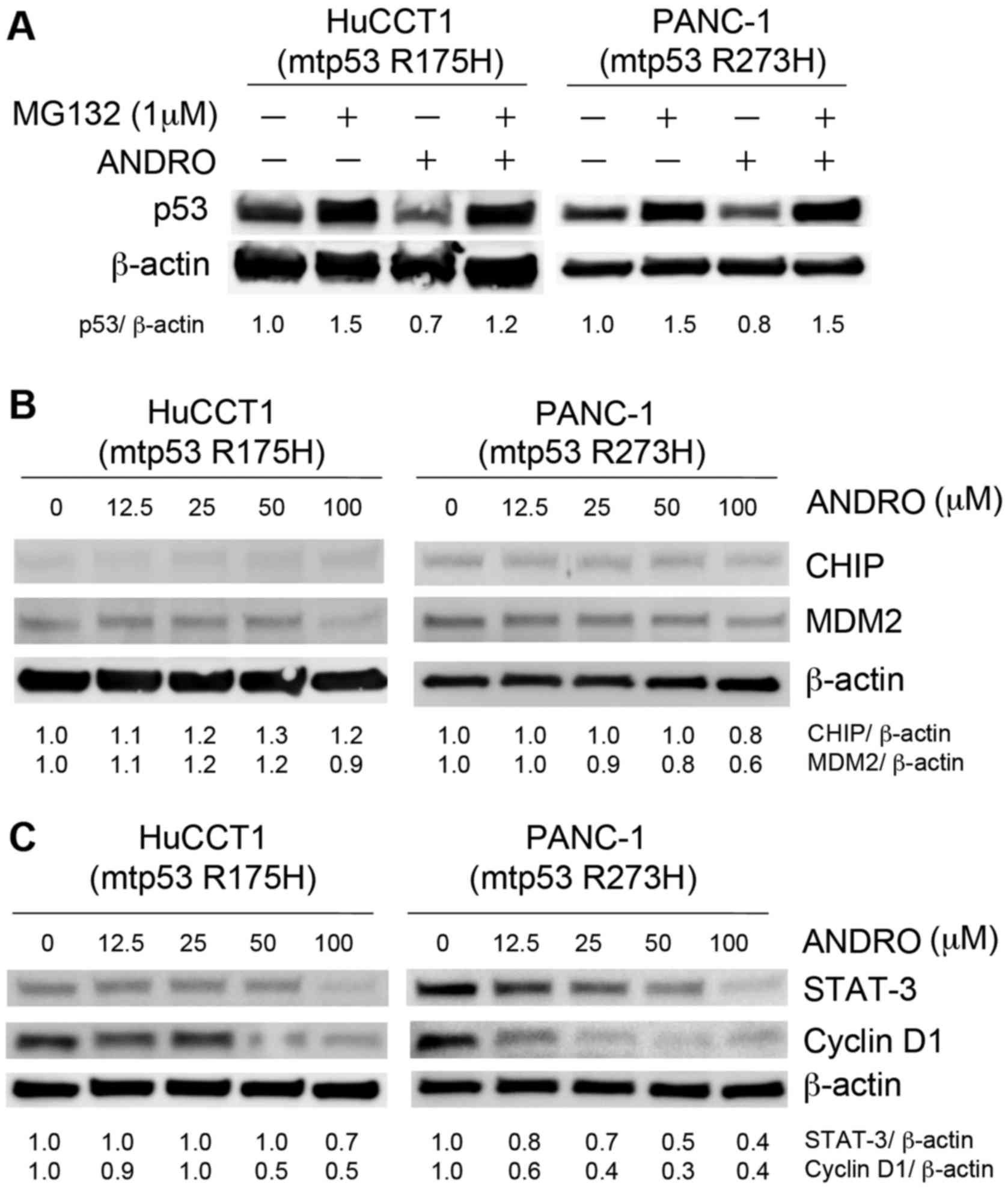
ANDRO promotes degradation of mutant p53
via induction of Hsp70
To understand how ANDRO reduces the expression of
mutant p53, it was hypothesized that ANDRO may destabilize and/or
promote the degradation of the mutant protein. The heat-shock
proteins are known to directly bind to p53 and regulate its
conformation and stability by functioning as chaperones or
co-chaperones (8). Therefore,
whether ANDRO affected the expression of these heat-shock proteins
was examined. Indeed, HuCCT1 and PANC-1 cells treated with ANDRO
exhibited increased expression of Hsp40 and Hsp70 mRNA levels
(Fig. 4A), and all cell lines,
regardless of their p53 status, exhibited increased levels of Hsp40
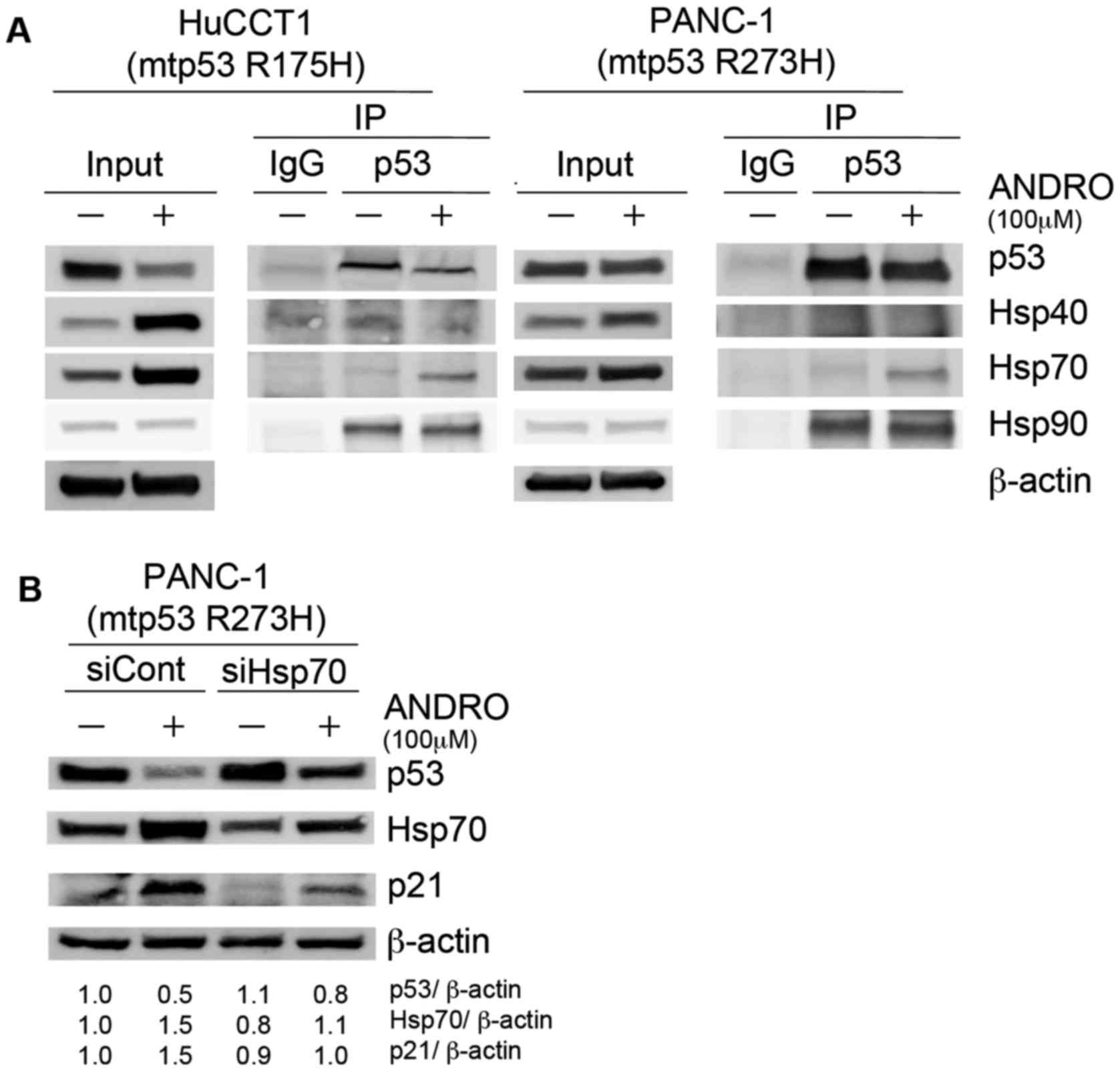
and Hsp70 protein following ANDRO treatment (Fig. 4B). To determine whether ANDRO
increased the binding of Hsp40, Hsp70, or Hsp90 to mutant p53, p53
was immunoprecipitated from control and ANDRO-treated cells and the
presence of the heat-shock proteins were probed for by
immunoblotting. More Hsp70 was bound to mutant p53 in ANDRO-treated
than vehicle-treated HuCCT1 and PANC-1 cells (Fig. 5A), suggesting that Hsp70 may be
involved in reducing mutant p53 levels. Consistent with this,
siRNA-mediated knockdown of Hsp70 reduced the ANDRO-induced
suppression of mutant p53 levels and concomitantly increased the
expression of p21 in PANC-1 cells (Fig. 5B).
ANDRO induces proteasomal degradation of
mutant p53
As ANDRO promotes the association of mutant p53 with
Hsp70, it was subsequently determined whether the enhanced
association influenced the stability/degradation of p53. For this,
cells were incubated with MG132 (1 µM for 18 h), an
inhibitor of proteasomal degradation. Notably, cells incubated with
MG132 and ANDRO exhibited higher mutant p53 levels than those
treated with ANDRO, indicating that ANDRO reduced p53 levels by
promoting its proteasomal degradation (Fig. 6A). MDM2 and CHIP are ubiquitin
ligases known to target mutant p53 for proteasomal degradation
(9). However, neither MDM2 nor
CHIP was induced by treatment of cells with ANDRO; in fact, MDM2
expression was marginally decreased (Fig. 6B), suggesting that other ubiquitin
ligases may be responsible for the enhanced p53 degradation.
Notably, a previous study reported that ANDRO can
induce p21 and apoptosis-associated genes via inactivation of
STAT-3 (10). Indeed, the
expression of STAT-3 and the cell cycle regulatory protein cyclin
D1 were decreased by ANDRO treatment of mutant p53-expressing cells
in a dose-dependent manner (Fig.
6C), suggesting that ANDRO may induce cell cycle arrest and
apoptosis of mutant p53-expressing cells via STAT-3.
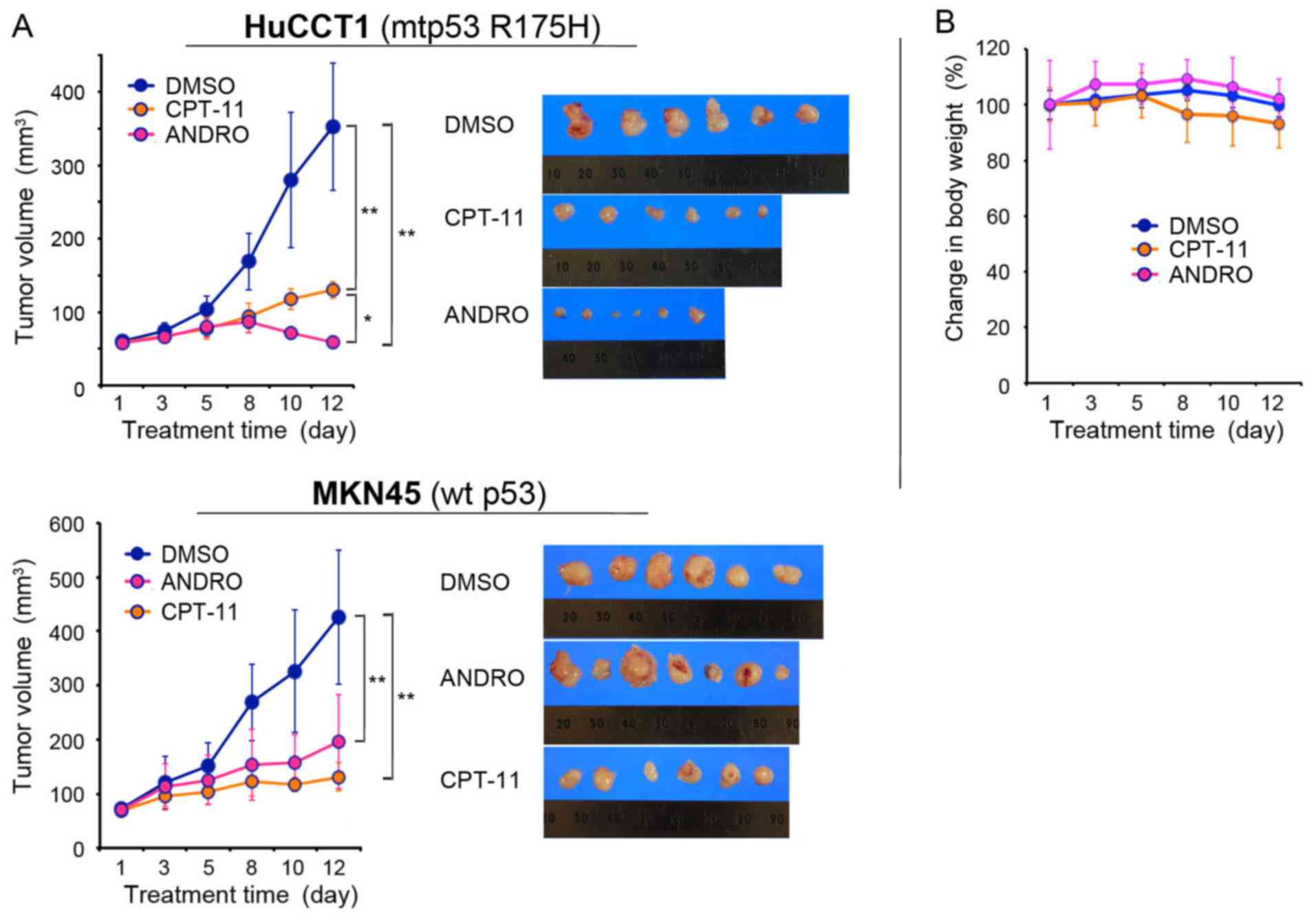
Antitumor effect of ANDRO in a mouse
xenograft model
It was verified that the anticancer effects of ANDRO
were also observed in vivo using a mouse xenograft model.
HuCCT1 cells (mutant p53 R175H) and MKN45 cells (wild-type p53)
were injected subcutaneously into the flanks of nude mice. When the
tumors reached 50 mm3 the mice were treated with vehicle
(DMSO in PBS), ANDRO (10 mg/kg), or CPT-11 (10 mg/kg) administered
via i.p. injection three times per week for 11 days. On day 12, the
tumor volumes of HuCCT1 were: 352.8±86.8 mm3 (range,
256–445.5 mm3; maximum diameter, 10 mm) in the DMSO
group; 130.3±11.5 mm3 (range, 126–147.9 mm3;
maximum diameter, 7 mm) in the CPT-11 group; and 58.8±8.8
mm3 (range, 45.6–75 mm3; maximum diameter, 6
mm) in the ANDRO group (Fig. 7A),
indicating that ANDRO was more effective than CPT-11 in suppressing
the growth of mutant p53-expressing HuCCT1 cells in vivo. By
contrast, CPT-11 was more effective than ANDRO at reducing the
wild-type p53-expressing MKN45 tumor volumes: 425.7±124
mm3 (range, 256–650 mm3; maximum diameter, 11
mm) in the DMSO control group; 131.3±26.5 mm3 (range,
108–171.5 mm3; maximum diameter, 7 mm) in the CPT-11
group; and 195.6±87.1 mm3 (range, 75–294 mm3;
maximum diameter, 10 mm) in the ANDRO group on day 12 (Fig. 7A). Notably, no significant body
weight loss was observed in any of the mouse groups during the
treatment period (Fig. 7B).
Discussion
Several small molecule anticancer drugs have been
reported to target mutant p53. Some of these, such as PRIMA-1
(11), MIRA-1 (12), NSC319726 (13), stictic acid (14) and chetomin (15), function by reactivating mutant p53,
thus re-establishing downstream target gene expression, and
induction of cell cycle arrest and apoptosis. Another group of
small molecules act by inhibiting mutant p53 gain-of-function
activities. These include histone deacetylase inhibitors (16,17),
NSC59984 (18), gambogic acid
(19), disulfiram (20), geldanamycin (21) and spautin-1 (22).
ANDRO was used in the current study as a small
molecule inhibitor of mutant p53. ANDRO, a diterpenoid lactone, is
a naturally occurring compound isolated from the stem and leaves of
a traditional Chinese herb, Andrographis paniculata, and is
commonly used to treat respiratory infections, fever and diarrhea
(23). Traditional Chinese herbs
are gaining increasing attention as a source of anticancer agents,
particularly as they are easy to obtain and have few, if any, side
effects (24). Previous studies
have demonstrated that ANDRO possesses anti-inflammatory (25,26),
anti-asthmatic (27), anti-viral
(28) and hepatoprotective
(29) activities, in addition to
its various anticancer effects (30,31).
However, the effect of ANDRO on mutant p53 has not previously been
investigated.
In the current study, it was demonstrated that ANDRO
induces the degradation of mutant p53 via increased Hsp70
expression. Several recent studies have also observed an
association between heat-shock proteins and mutant p53 in cancer
cells. Paradoxically, although Hsp70 contributes to the stability
of mutant p53 (32), it may also
be associated with the degradation of mutant p53 (33,34).
In the present study, ANDRO increased the expression of Hsp70 and
increased its binding to mutant p53. In addition, siRNA-mediated
knockdown of Hsp70 impaired the ANDRO-mediated reduction in mutant
p53 levels. These results indicated that increased Hsp70 activity
is one mechanism by which ANDRO promotes the degradation of mutant
p53 protein.
Two major pathways are thought to be responsible for
decreasing mutant p53 protein levels; namely, proteasomal and
autophagic degradation (22). The
experiments with MG132 suggest that ANDRO acts via the proteasomal
degradation pathway. Although MDM2 and CHIP, E3 ubiquitin ligases
known to target p53, were not induced by ANDRO, we hypothesize that
another E3 ubiquitin-related protein may mediate the degradation of
mutant p53 in this study. This possibility would be clarified by
mass spectrometric analysis of p53-interacting proteins.
The current study demonstrated that ANDRO has
anticancer effects in vitro and in vivo. Although
p21, PUMA, and NOXA are known to be induced by wild-type p53
protein, it was demonstrated here that they are also induced by
ANDRO treatment of cells harboring mutant p53. One potential
mechanism of the increased p21 levels may be independent of p53.
Zhang et al (35)
previously reported that MDM2 is a negative regulator of p21. The
data of the current study revealed that MDM2 was marginally
decreased following ANDRO treatment of mutant p53 cells. Another
potential mechanism of p21 induction is via STAT-3 inactivation.
Previous study has demonstrated that expression of p21 and
apoptosis-associated proteins are regulated by STAT-3, and mutant
p53 activates these proteins by suppressing the activity of STAT-3
and Akt (10). In the experiments
of the present study, ANDRO inhibited STAT-3, suggesting a
potential mechanism for its induction of p21 and
apoptosis-associated proteins.
In the mouse xenograft model, ANDRO suppressed tumor
growth irrespective of p53 mutational status, but the effect was
greater for mutant p53-expressing cells. As no significant body
weight loss was observed in ANDRO-treated mice, ANDRO may be a
potential treatment for mutant p53-expressing tumors, which
constitute about half of all human cancers. In conclusion, the
findings of the current study demonstrated that ANDRO induces the
degradation of mutant p53 via activation of Hsp70.
Acknowledgments
Not applicable.
Funding
This study was partially supported by Grant-in Aid
for the Young Scientists (B: 16K19938) from Japan Society for the
Promotion of Science and Medical Care Education Research Foundation
(Fukuoka, Japan).
Availability of data and materials
The data sets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
HS and MH conceived and designed the experiments. HS
performed the majority of the experiments; TN and NE performed
certain experiments. HS, TN and MH analyzed the data. HS and MH
contributed to interpretation of the data, and were major
contributors in writing the manuscript. TN, KB, TT and HN
contributed to the research design and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
An animal experimental protocol was reviewed and
approved by the Animal Care Committee of Saga University (Saga,
Japan; permission no. 27-039-0).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Joerger AC and Fersht AR: Structural
biology of the tumor suppressor p53. Annu Rev Biochem. 77:557–582.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lane DP: Cancer. p53, guardian of the
genome. Nature. 358:15–16. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hainaut P and Hollstein M: p53 and human
cancer: The first ten thousand mutations. Adv Cancer Res.
77:81–137. 2000. View Article : Google Scholar
|
|
4
|
Terzian T, Suh YA, Iwakuma T, Post SM,
Neumann M, Lang GA, Van Pelt CS and Lozano G: The inherent
instability of mutant p53 is alleviated by Mdm2 or p16INK4a loss.
Genes Dev. 22:1337–1344. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bossi G, Lapi E, Strano S, Rinaldo C,
Blandino G and Sacchi A: Mutant p53 gain of function: Reduction of
tumor malignancy of human cancer cell lines through abrogation of
mutant p53 expression. Oncogene. 25:304–309. 2006. View Article : Google Scholar
|
|
6
|
Patricia AM and Vousden KH: Mutant p53 in
cancer. Cancer Cell. 25:304–317. 2013.
|
|
7
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
8
|
King FW, Wawrzynow A, Höhfeld J and Zylicz
M: Co-chaperones Bag-1, Hop and Hsp40 regulate Hsc70 and Hsp90
interactions with wild-type or mutant p53. EMBO J. 20:6297–6305.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li D, Marchenko ND, Schulz R, Fischer V,
Velasco-Hernandez T, Talos F and Moll UM: Functional inactivation
of endogenous MDM2 and CHIP by HSP90 causes aberrant stabilization
of mutant p53 in human cancer cells. Mol Cancer Res. 9:577–588.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bao GQ, Shen BY, Pan CP, Zhang YJ, Shi MM
and Peng CH: Andrographolide causes apoptosis via inactivation of
STAT3 and Akt and potentiates antitumor activity of gemcitabine in
pancreatic cancer. Toxicol Lett. 222:23–35. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lambert JM, Gorzov P, Veprintsev DB,
Söderqvist M, Segerbäck D, Bergman J, Fersht AR, Hainaut P, Wiman
KG and Bykov VJ: PRIMA-1 reactivates mutant p53 by covalent binding
to the core domain. Cancer Cell. 15:376–388. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bykov VJ, Issaeva N, Zache N, Shilov A,
Hultcrantz M, Bergman J, Selivanova G and Wiman KG: Reactivation of
mutant p53 and induction of apoptosis in human tumor cells by
maleimide analogs. J Biol Chem. 280:30384–30391. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu X, Vazquez A, Levine AJ and Carpizo DR:
Allele-specific p53 mutant reactivation. Cancer Cell. 21:614–625.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wassman CD, Baronio R, Demir Ö, Wallentine
BD, Chen CK, Hall LV, Salehi F, Lin DW, Chung BP, Hatfield GW, et
al: Computational identification of a transiently open L1/S3 pocket
for reactivation of mutant p53. Nat Commun. 4:14072013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hiraki M, Hwang SY, Cao S, Ramadhar TR,
Byun S, Yoon KW, Lee JH, Chu K, Gurkar AU, Kolev V, et al:
Small-molecule reactivation of mutant p53 to wild-type-like p53
through the p53-Hsp40 regulatory axis. Chem Biol. 22:1206–1216.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li D, Marchenko ND and Moll UM: SAHA shows
preferential cytotoxicity in mutant p53 cancer cells by
destabilizing mutant p53 through inhibition of the HDAC6-Hsp90
chaperone axis. Cell Death Differ. 18:1904–1913. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Blagosklonny MV, Trostel S, Kayastha G,
Demidenko ZN, Vassilev LT, Romanova LY, Bates S and Fojo T:
Depletion of mutant p53 and cytotoxicity of histone deacetylase
inhibitors. Cancer Res. 65:7386–7392. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang S, Zhou L, Hong B, van den Heuvel
AP, Prabhu VV, Warfel NA, Kline CL, Dicker DT, Kopelovich L and
El-Deiry WS: Small-molecule NSC59984 restores p53 pathway signaling
and antitumor effects against colorectal cancer via p73 activation
and degradation of mutant p53. Cancer Res. 75:3842–3852. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang J, Zhao Q, Qi Q, Gu H, Rong J, Mu R,
Zou M, Tao L, You Q and Guo Q: Gambogic acid-induced degradation of
mutant p53 is mediated by proteasome and related to CHIP. J Cell
Biochem. 112:509–519. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paranjpe A and Srivenugopal KS:
Degradation of NF-kappaB, p53 and other regulatory redox-sensitive
proteins by thiolconjugating and -nitrosylating drugs in human
tumor cells. Carcinogenesis. 34:990–1000. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin K, Rockliffe N, Johnson GG,
Sherrington PD and Pettitt AR: Hsp90 inhibition has opposing
effects on wild-type and mutant p53 and induces p21 expression and
cytotoxicity irrespective of p53/ATM status in chronic lymphocytic
leukaemia cells. Oncogene. 27:2445–2455. 2008. View Article : Google Scholar
|
|
22
|
Liu J, Xia H, Kim M, Xu L, Li Y, Zhang L,
Cai Y, Norberg HV, Zhang T, Furuya T, et al: Beclin1 controls the
levels of p53 by regulating the deubiquitination activity of USP10
and USP13. Cell. 147:223–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Akbar S: Andrographis paniculata: A review
of pharmacological activities and clinical effects. Altern Med Rev.
16:66–77. 2011.PubMed/NCBI
|
|
24
|
Xia Q and Mao W: Anti-tumor effects of
traditional Chinese medicine give a promising perspective. J Cancer
Res Ther. 10(Suppl 1): 1–2. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Abu-Ghefreh AA, Canatan H and Ezeamuzie
CI: In vitro and in vivo anti-inflammatory effects of
andrographolide. Int Immunopharmacol. 9:313–318. 2009. View Article : Google Scholar
|
|
26
|
Shao ZJ, Zheng XW, Feng T, Huang J, Chen
J, Wu YY, Zhou LM, Tu WW and Li H: Andrographolide exerted its
antimicrobial effects by upregulation of human β-defensin-2 induced
through p38 MAPK and NF-κB pathway in human lung epithelial cells.
Can J Physiol Pharmacol. 90:647–653. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nguyen VS, Loh XY, Wijaya H, Wang J, Lin
Q, Lam Y, Wong WS and Mok YK: Specificity and inhibitory mechanism
of andrographolide and its analogues as antiasthma agents on NF-κB
p50. J Nat Prod. 78:208–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wintachai P, Kaur P, Lee RC, Ramphan S,
Kuadkitkan A, Wikan N, Ubol S, Roytrakul S, Chu JJ and Smith DR:
Activity of andrographolide against chikungunya virus infection.
Sci Rep. 5:141792015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thingale AD, Shaikh KS, Channekar PR,
Galgatte UC, Chaudhari PD and Bothiraja C: Enhanced
hepatoprotective activity of andrographolide complexed with a
biomaterial. Drug Deliv. 22:117–124. 2015. View Article : Google Scholar
|
|
30
|
Zhang QQ, Ding Y, Lei Y, Qi CL, He XD, Lan
T, Li JC, Gong P, Yang X, Geng JG, et al: Andrographolide suppress
tumor growth by inhibiting TLR4/NF-κB signaling activation in
insulinoma. Int J Biol Sci. 10:404–414. 2014. View Article : Google Scholar :
|
|
31
|
Gao H and Wang J: Andrographolide inhibits
multiple myeloma cells by inhibiting the TLR4/NF-κB signaling
pathway. Mol Med Rep. 13:1827–1832. 2016. View Article : Google Scholar
|
|
32
|
Wiech M, Olszewski MB, Tracz-Gaszewska Z,
Wawrzynow B, Zylicz M and Zylicz A: Molecular mechanism of mutant
p53 stabilization: The role of HSP70 and MDM2. PLoS One.
7:e514262012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim HB, Lee SH, Um JH, Oh WK, Kim DW, Kang
CD and Kim SH: Sensitization of multidrug-resistant human cancer
cells to Hsp90 inhibitors by downregulation of SIRT1. Oncotarget.
6:36202–36218. 2015.PubMed/NCBI
|
|
34
|
Buckley NE, D'Costa Z, Kaminska M and
Mullan PB: S100A2 is a BRCA1/p63 coregulated tumour suppressor gene
with roles in the regulation of mutant p53 stability. Cell Death
Dis. 5:e10702014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Z, Wang H, Li M, Agrawal S, Chen X
and Zhang R: MDM2 is a negative regulator of p21WAF1/CIP1,
independent of p53. J Biol Chem. 279:16000–16006. 2004. View Article : Google Scholar : PubMed/NCBI
|