Introduction
Gastric cancer (GC) currently ranks as the fourth
most frequently diagnosed cancer and the second leading cause of
mortality from malignant diseases worldwide (1). According to statistics, >950 000
new GC cases are diagnosed every year, and an estimated 723,100
mortalities occurred in 2012 (1,2). By
the time of diagnosis, the majority of patients present with
advanced GC, with a 5-year survival rate of <30% (3,4).
Pre-operative chemotherapy has been successfully used in the
treatment of locally advanced GC, as it may increase the
possibility of complete resection through shrinking the tumor
(5). Based on clinical data,
patients with GC that exhibit a positive response to neo-adjuvant
chemotherapy can obtain a better prognosis than non-responders
(5). However, different patients
exhibit distinct therapeutic responses due to various genetic and
epigenetic alterations that are involved in gastric tumorigenesis
(4,6). Therefore, a comprehensive
understanding of the molecular variables that affect the GC disease
pathway may contribute to identifying novel therapeutic targets or
prognostic biomarkers that may help identify patients that are
likely to benefit from chemotherapy.
Psoriasin, also termed S100A7, belongs to the S100
multi-genic family of calcium-binding EF-hand proteins and was
initially identified in 1991 as a highly expressed protein in
psoriatic keratinocytes (7,8).
Psoriasin, a low molecular weight protein (11.4 kDa), is localized
in the cytoplasm, cell nucleus or secreted outside of the cell in
certain cases (9,10). Dysregulated expression of psoriasin
has been observed in various types of cancer, including oral
squamous cell carcinoma (11),
breast cancer (10,12), lung cancer (13), bladder cancer (14), skin cancer (15), head and neck cancer (16), prostate cancer (17) and cervical cancer (18). Functionally, psoriasin usually has
malignant functions in cancer; however, it can also exhibit tumor
suppressive effects in certain types of oral carcinoma and breast
cancer by decreasing β-catenin activity (11,12).
Subsequent studies have suggested that intracellular and secreted
forms of psoriasin protein have an important role in tumor
development by regulating cell proliferation, metastasis,
migration, invasion, apoptosis and angiogenesis (17–19).
Mechanistically, several signaling pathways have been demonstrated
to have a correlation with psoriasin, including Hippo, receptor for
advanced glycation endproducts (RAGE), extracellular
signal-regulated kinase (ERK), β-catenin and nuclear factor (NF)-κB
pathways. The downregulation of psoriasin in lung cancer suppresses
cell growth and invasive potential by regulating NF-κB activity and
the expression of matrix metalloproteinases and vascular
endothelial growth factor (19).
Similarly, psoriasin promotes cell survival by binding to c-Jun
activation domain-binding protein1 and enhancing the
phosphorylation of NF-κB and protein kinase B in breast cancer
(20). Another study demonstrated
that psoriasin binds to RAGE and activates ERK signaling to
increase migration and invasion in cervical cancer (18).
However, the role of psoriasin in GC and its
underlying mechanisms remain unclear. To the best of our knowledge,
the present study is the first to characterize the expression of
psoriasin in patients with GC and investigate the role of psoriasin
in GC pathogenesis. Psoriasin was markedly upregulated in patients
with GC and high expression of psoriasin was associated with poor
survival. Furthermore, psoriasin positively regulated ERK signaling
and promoted proliferation, migration and invasion. In addition,
overexpression of psoriasin decreased the sensitivity of GC cells
to cisplatin. Taken together, the findings suggested that psoriasin
has a critical role in the pathogenesis of GC and may be a novel
therapeutic target or a potential diagnostic marker for the patient
response to pre-operative chemotherapy in GC.
Materials and methods
Cell line and cell culture
The human AGS (no. 89090402) and HGC-27 (no.
94042256) GC cell lines were purchased from the European Collection
of Animal Cell Cultures (ECACC; Salisbury, UK). The cells were
cultured in Dulbecco's modified Eagle's medium (DMEM)-F12 medium
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), supplemented with
10% fetal calf serum (FCS; Sigma-Aldrich; Merck KGaA) and 1%
antibiotics (penicillin and streptomycin; Sigma-Aldrich; Merck
KGaA) at 37°C in a humidified 5% CO2 atmosphere.
Patients and human GC tissue
Two cohorts were used in the present study, namely a
generic collection and a collection with chemosensitive
information. The first cohort had a total of 321 GC tissues and 173
adjacent normal tissues were collected from patients with GC that
underwent surgical resection, and some were treated with
preoperative radiotherapy or chemotherapy at the Peking University
Cancer Hospital (Beijing, China). Tissue and data were obtained
between September 2003 and December 2007. The mean age of the
patients in the cohort was 59.8 years with male/female ratio of
229:92 (Table I). The other cohort
comprised of 87 patients with GC that had received chemotherapy
with oxaliplatin/5-fluorouracil/leucovorin prior to surgery at the
Peking University Cancer Hospital between December 2001 and
November 2006. All of the tissues were stored immediately following
surgery at −80°C until use. Clinicopathological and follow-up
information was obtained from patient data. GC staging was
determined according to the 2010 tumor-node-metastasis (TNM)
classification recommended by the American Joint Committee on
Cancer (AJCC 7th edition) (21).
This study was approved by Peking University Cancer Hospital
Research Ethics Committee (ethics no. 2006021) and written consent
was obtained from all patients involved.
 | Table IAssociation between psoriasin mRNA
expression and clinicopathological features in gastric cancer
tissues. |
Table I
Association between psoriasin mRNA
expression and clinicopathological features in gastric cancer
tissues.
| Variable | n | Psoriasin
expression (median) | P-value |
|---|
| Tissue | | | <0.001 |
| Tumor | 321 | 12.00 | |
| Adjacent
normal | 173 | 0.04 | |
| Sex | | | 0.573 |
| Male | 229 | 9.00 | |
| Female | 92 | 13.00 | |
| Depth of
invasion | | | 0.011 |
| T1–2 | 42 | 1.00 | |
| T3–4 | 279 | 15.00 | |
| Lymph node
status | | | <0.001 |
| N0 | 71 | 0.03 | |
| N1+2+3 | 250 | 16.00 | |
| Distant
metastasis | | | <0.001 |
| M0 | 280 | 4.00 | |
| M1 | 41 | 384.00 | |
| TNM staging | | | <0.001 |
| I+II | 85 | 1.00 | |
| III+IV | 236 | 17.00 | |
|
Differentiation | | | |
| High | 1 | 0.15 | |
| High-medium | 21 | 20.00 | |
| Medium | 67 | 15.00 | 0.629a |
| Medium-low | 87 | 13.00 | 0.732a |
| Low | 145 | 3.00 | 0.453a |
| Clinical
outcome | | | <0.001 |
| Alive | 134 | 1.00 | |
| Died | 187 | 25.00 | |
RNA isolation, reverse transcription,
conventional polymerase chain reaction (PCR) and quantitative PCR
(qPCR)
Total RNA was extracted from cells or homogenized
tissues using TRI reagent (Sigma-Aldrich; Merck KGaA) following the
manufacturer's instructions, and subsequently reverse transcribed
into cDNA using Promega Reverse Transcription kit (Promega
Corporation, Madison, WI, USA). The reverse transcription
conditions were annealing at 25°C for 5 min, extension at 42°C for
60 min, and inactivation at 70°C for 15. PCR was then performed
using GoTaq Green MasterMix (Promega Corporation) under the cycling
conditions as follows: 95°C for 2 min, followed by 40 cycles of
95°C for 30 sec, 55°C for 30 sec and 72°C for 30 sec, and a final
extension of 5 min at 72°C. The PCR products were visualized on
1–2% agarose gels stained with SYBR Safe (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The primer sequences used for
PCR in this study are listed in Table
II.
| Table IIPrimers sequences. |
Table II
Primers sequences.
| Name | Forward | Reverse |
|---|
| Psoriasin |
ATGAGCAACACTCAAGCTG |
ACTGGCTGCCCCCGGAACA |
| Psoriasin
(qPCR) |
TGTGACAAAAAGGGCACAAA | ACTGAACCTGACCGTACACCCAGCAAGGACAGAAACTC |
| Snail |
CGCTCTTTCCTCGTCAG |
GTTGCAGTATTTGCAGTTGA |
| Slug |
CTCTCCTCTTTCCGGATACT |
AGCAGTTTTTGCACTGGTAT |
| Vimentin |
GATGCTTCAGAGAGAGGAAG |
CTCTTCGTGGAGTTTCTTCA |
| E-cadherin |
CGAGAGCTACACGTTCAC |
GGGAAAAATAGGCTGTCCTT |
| N-cadherin |
CAACGACGGGTTAGTCAC |
ATTGGGGTCTGGAGTTTC |
| Zeb-1 |
CTTGTGATTTGTGTGACAAGA |
ATGCCTTTTTACAGATTCCA |
| MMP-2 |
TTTGATGACGATGAGCTATG |
TGCAGCTCTCATATTTGTTG |
| MMP-3 |
TCATTTTGGCCATCTCTTCC |
GTGCCCATATTGTGCCTTCT |
| MMP-9 |
AACTACGACCGGGACAAG |
ATTCACGTCGTCCTTATGC |
| ERK1 |
ACACGCAGTTGCAGTACA |
CCACATACTCCGTCAGGA |
| ERK2 |
CCAACCTCTCGTACATCG |
GTCAGGAACCCTGTGTGA |
| c-Fos |
CAGACTACGAGGCGTCATCC |
TCTGCGGGTGAGTGGTAGTA |
| c-Jun |
AAGATCCTGAAACAGAGCAT |
GCTGGACTGGATTATCAGG |
| Cyclin D1 |
CGGTGTCCTACTTCAAATGT |
ACCTCCTCCTCCTCCTCT |
| Cyclin D2 |
GGAGAAGCTGTCTCTGATCC |
GGGTACATGGCAAACTTAAA |
| GAPDH |
CGCTGCTTTTAACTCTGGTA |
GACTGTGGTCAGAGTCCTT |
| GAPDH (qPCR) |
CTGAGTACGTCGTCGTGGAGTC | ACTGAACCTGACCGTACACAGAGATGATGACCCTTTTG |
| Actin (qPCR) |
GGACCTGACTGACTACCTCA | ACTGAACCTGACCGTACAAGCTTCTCCTTAATGTCACG |
qPCR was run on the StepOne Plus Real-Time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.), using
Precision FAST 2X qPCR MasterMix (Primerdesign Ltd., Chandler's
Ford, UK) in a final volume of 10 µl. cDNA samples from GC
cells and two cohorts of GC tissues were examined for psoriasin
transcript expression with standard and negative controls. Reaction
conditions were as follows: 94°C for 10 min, followed by 100 cycles
of 94°C for 10 sec, 55°C for 30 sec and 72°C for 10 sec. GAPDH or
actin was used as an internal control. The primer sequences used
for qPCR are listed in Table II.
Primers for qPCR were designed to the specific target using the
Beacon Designer software (version 2; Premier Biosoft International,
Palo Alto, CA, USA). An additional sequence, known as the Z
sequence (5′-ACTGAACCTGACCGTACA-3′), which acts as a template for
the amplifluor uniprimer probe, was added with the reverse primers.
Expression of the target sequence was qualified in conjunction with
a range of known dilution series of podoplanin gene transcript,
which are used to generate a standard curve to enable the
calculation of transcript copy.
Construction of psoriasin overexpression
vector in GC cell lines
Full-length human psoriasin coding sequence was
amplified from human prostate tissues with appropriate primers
(Table I). The synthesized product
was cloned into the pEF6/V5-His-TOPO plasmid vector (K961020;
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The overexpression constructs and
control plasmids (25 µg pre transfection) were then
transfected into 2.5×106 AGS and HGC-27 cells using a
Gene Pulser Xcell electroporation system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). After 2 weeks of selection using 6
µg/ml blasticidin (Melford Laboratories, Ltd., Ipswich, UK),
stable transfectants were obtained and verified for psoriasin
overexpression. Then cells were cultured in DMEM with a lower
concentration of blasticidin (0.6 µg/ml) to maintain plasmid
expression.
Treatment with ERK inhibitor and
cisplatin
AGS and HGC-27 were cultured in serum-free medium
for 4–6 h, and then treated with 1 µM ERK inhibitor (FR
180204; Tocris Bioscience, Bristol, UK) or 16.6 µM cisplatin
(Tocris Bioscience) for 24 h, followed by PCR, western blotting or
flow cytometry analysis.
Western blotting
AGS and HGC-27 were isolated by pre-chilled lysis
buffer (50 mM Tris-Cl, pH 7.5, 1% Nonidet P-40, 0.5% sodium
deoxycholate, 0.05% SDS, 1 mM EDTA, 150 mM NaCl) containing
protease inhibitor cocktail (aprotinin and leupeptin) and
phosphorylation inhibitor (sodium vanadate) and qualified with a
DCTM protein assay kit (Bio-Rad Laboratories, Inc.). Protein
extract of each sample (30 µg) was separated by 10% or 12%
SDS-PAGE and transferred onto a 0.45 µm
polyvinylidenedifluoride membrane (EMD Millipore, Billerica, MA,
USA). The membranes were blocked with 5% non-fat dried milk in PBST
solution (0.05% Tween-20 in PBS) for 1 h at room temperature. The
blocked membranes were then incubated in a diluted primary
antibodies (1:500) overnight as 4°C, washed in PBST and incubated
with a diluted specific horseradish peroxidase (HRP)-conjugated
secondary antibody (1:2,000) for 1 h at room temperature. The
following primary antibodies were used in the study:
Anti-S100A7/psoriasin (clone 47C1068; catalog no.; Novus
Biologicals, Ltd., Cambridge, UK); phosphor (p)-ERK (Tyr 204; cat.
no. sc-7976), ERK1/2 (clone C-9; cat. no. sc-514302), p21 (cat. no.
sc-6246), p53 (clone DO-1; cat. no. sc-126), vimentin (cat. no.
sc-66002), E-cadherin (cat. no. sc-1500), N-cadherin (cat. no.
sc-7939), zinc finger E-box binding homeobox 1 (Zeb-1; cat. no.
sc-81428) and GAPDH (cat. no. sc-32233; all Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The HRP secondary antibodies
(cat. no. A5278, anti-mouse IgG; cat. no. A0545, anti-rabbit IgG)
were obtained from Sigma-Aldrich (Merck, KGaA). Detection was
performed using an EZ-ECL chemiluminescence kit (Biological
Industries USA, Inc., Cromwell, CT, USA). Immunoreactive bands were
visualized and quantified by densitometry using the Syngene G:BOX
chemiluminescence imaging system and Gene Tools 4.03 (Syngene
Europe, Cambridge, UK).
Cell proliferation assay
Briefly, AGS and HGC-27 cells were seeded into
96-well plate (2×103 cells/well) in 6 replicates
followed by incubation for 0, 1, 2, 3, 4 and 5 days. Following
fixation in 4% formalin for 30 min at room temperature, the cells
were stained with 0.5% (w/v) crystal violet (Sigma-Aldrich) for 15
min at room temperature. Following washing, the cells were then
extracted with 10% (v/v) acetic acid and cell density was
determined by measuring the absorbance at 540 nm using a
spectrophotometer (Elx800; BioTek Instruments, Inc., Winooski, VT,
USA).
Cell viability assay
To evaluate the cytotoxicity of cisplatin, AGS and
HGC-27 cells were plated into 96-well plate (cells/well) in 6
replicates. After 24-h incubation, the cells were exposed to serial
concentrations of cisplatin (0, 3.125, 6.25, 12.5, 25 and 50
µM, respectively) for 72 h. Cells viability was assessed by
staining with 0.5% (w/v) crystal violet for 15 min at room
temperature.
Cell colony assay
AGS and HGC-27 were plated into 6-well plate at the
density of 400 cells/well and incubated at 37°C and at an
atmosphere of 5% CO2 for 2 weeks. The cell colonies were
fixed in 4% formalin for 30 min and stained with 0.5% (w/v) crystal
violet for 15 min. After images were taken with a camera, the
colonies were dissolved in 10% (v/v) acetic acid, and the
absorbance was measured at 540 nm using a spectrophotometer
(Elx800; BioTek Instruments, Inc.).
Cell-matrix adhesion assay
A total of 2×104 cells were plated in
each well of a black 96-well plate previously coated with Matrigel
(10 µg/well; Corning Incorporated, Corning, NY, USA) in 6
replicates. After incubation for 1 h, the cells were washed once
with PBS and then incubated in cell dissociation solution (100
µl/well; Sigma-Aldrich; Merck KGaA) for 30 min containing
0.002 µg/µl calcein AM (eBioscience; Thermo Fisher
Scientific, Inc.). Fluorescence of the cell suspension was read
using a GloMax®-Multi Detection System (Promega
Corporation) at 495 nm for excitation and 519 nm for emission.
In vitro invasion assay
Cells (5×104) were loaded into an 8
µm-pore ThinCertTM 24-well plate insert (Greiner Bio-One
International GmbH, Kremsmünster, Austria) coated with 100
µg Matrigel (each sample in triplicate). The upper chamber
was provided with serum-free medium, and the lower chamber was
filled with DMEM with 10% FCS. Following incubation for 24 h, the
underside of the insert was incubated in 350 µl dissociation
solution containing 1 µM calcein AM for 1 h. The cell
suspension was then aliquoted into a black 96-well plate (100
µl/well) and read using the GloMax®-Multi
Detection System at 495 nm for excitation and 519 nm for
emission.
In vitro cell migration assay and the
effect of ERK inhibitor on cell mobility
Electric cell-substrate impedance sensing (ECIS)
instruments (Applied Biophysics, Inc., Troy, NY, USA) were used to
monitor cell migration by detecting impedance variation based on
the current flow change on the gold electrodes of a 96-well array
(Applied Biophysics Inc.) (22).
There are two parameters, resistance and capacitance, derived from
current and voltage measurements. Among them, resistance,
consisting of para- and trans-cellular current flow, is better to
represent the quality and function of cell barrier (23). Briefly, 6×104 cells
diluted in 200 µl DMEM with or without ERK inhibitor (1
µM) were seeded into each ECIS 96-plate well, and the cells
were wounded by applying electric current (3,000 µA, 60 kHz)
when confluent monolayers were formed. The array was placed into a
CO2 incubator, which was connected to the ECIS Model
9600 Controller, and the impedance data, which indicate cellular
migration, were collected continuously for 6 h. In this study, the
normalized resistance measured at 4,000 Hz was chosen to evaluate
cell function due to its high sensitivity to the current flow
variation caused by the change of cell behavior as described in the
literature (24).
Flow cytometric analysis
For protein expression analysis, AGS and HGC-27 were
fixed with IC Fixation Buffer (eBioscience; Thermo Fisher
Scientific, Inc.) at room temperature for 30 min, and then
permeabilized in ice-cold 100% methanol at 4°C for 30 min.
Following blocking with PBS containing 0.1% Tween-20 (v/v) and 1%
bovine serum albumin (w/v) (Sigma-Aldrich; Merck KGaA), the cells
were incubated with a cleaved poly(ADP-ribose) polymerase 1 (PARP1)
antibody (1:40; cat. no. 44-698G; eBioscience; Thermo Fisher
Scientific, Inc.) overnight at 4°C. Mouse IgG2b K isotype antibody
(eBioscience; Thermo Fisher Scientific, Inc.) was used as a
control. The samples were then washed by PBS twice and resuspended
in PBS containing 2 mM EDTA and 5% FCS for flow cytometry.
For cell cycle analysis, the cells were fixed with
70% ethanol for 15 min on ice, and stained with FxCycle™ PI/RNase
staining solution (Thermo Fisher Scientific, Inc.) for 30 min. For
apoptosis analysis, the cells were stained using an Annexin V
Apoptosis Detection kit (eBioscience; Thermo Fisher Scientific,
Inc.) following the manufacturer's instructions. Flow cytometry was
performed using a Canto II flow cytometer (BD Bioscience, San Jose,
CA, USA), and the data were analyzed using FCS Express version 4
Research Edition (De Novo Software, Glendale, CA, USA).
Statistical analysis
The results are presented as data from three
independent experiments and expressed as the mean ± standard
deviation. The cumulative survival curves were generated using
Kaplan-Meier plots and analyzed using log-rank test. Kruskal-Wallis
test was used for comparisons of three or more groups followed by
Tukey Honest Significant Difference as a post hoc test, and
Mann-Whitney U test was used for comparisons between two groups.
Statistical analysis was performed using SPSS v18 (SPSS, Inc.,
Chicago, IL, USA) and GraphPad Prism 6.0 (GraphPad Software, La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of psoriasin transcripts and
its correlation with clinicopathological parameters in GC
tissues
To determine the clinical relevance of psoriasin in
GC progression, the transcript levels of psoriasin in GC (n=321)
and the adjacent normal specimens (n=173) were examined by RT-qPCR.
As presented in in Table II,
psoriasin was higher in GC tissues than in adjacent normal tissues
(P<0.001), and a higher level of psoriasin was observed in
advanced stages of GC (TNM III+IV) compared with early stages (TNM
I+II; P<0.001; Table II).
Notably, higher expression of psoriasin was detected in tumors with
deeper infiltration (T3–4 vs. T1-2; P=0.011), lymph node
involvement (P<0.001) and distance metastasis (P<0.001,
Table II). Furthermore, psoriasin
exhibited higher expression in patients with GC that had died prior
to the end of follow-up than in those who remained alive at the end
of the follow-up period (P<0.001; Table II). Kaplan-Meier analysis and
log-rank test revealed that patients with GC with higher expression
of psoriasin had worse overall survival (P<0.001; Fig. 1A) and disease-free survival
(P<0.001; Fig. 1B), compared
with those with lower expression.
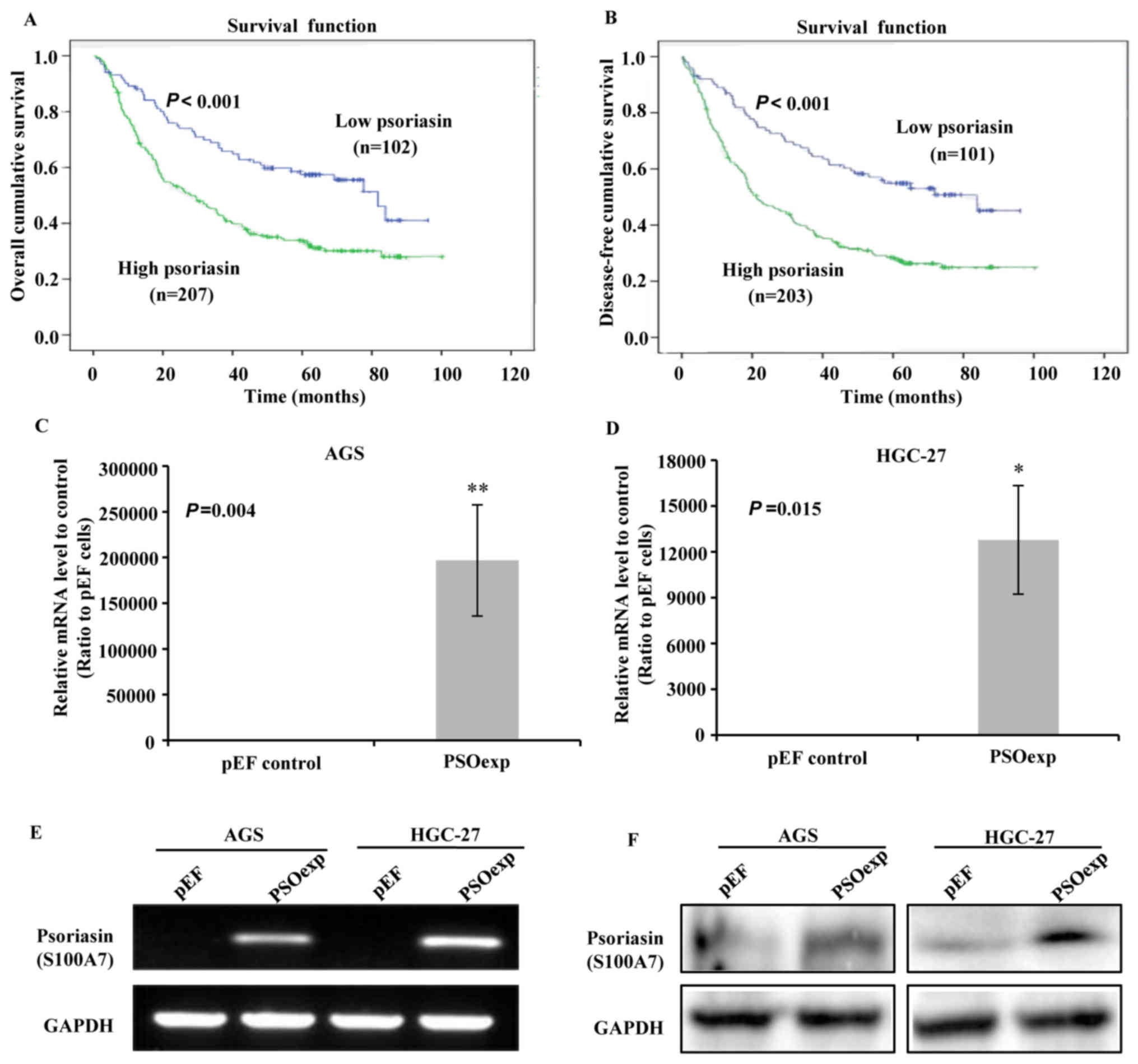 | Figure 1Survival curves of patients with GC,
and the validation of psoriasin overexpression in GC cell lines.
The high-low groups of patients were divided the median transcript
values of psoriasin, guided by clinical stages (TNM staging). (A)
Association between the transcript levels of psoriasin and overall
survival. Low psoriasin group (median expression, 1), n=102; high
psoriasin expression (median expression, 25), n=207, P<0.001.
(B) The correlation between the transcript levels of psoriasin and
disease-free survival. Low psoriasin group (median expression, 1),
n=101; high psoriasin expression (median expression, 25), n=203.
P<0.001. Reverse transcription-quantitative PCR analyzing mRNA
expression of psoriasin in (C) AGS and (D) HGC-27 cell line, which
was normal-ized with the reference gene GAPDH. The data are
presented as a ratio to pEF control. (E) Confirmation of psoriasin
overexpression in AGS and HGC-27 cells using conventional PCR. (F)
Western blot analysis of the level of psoriasin expression in AGS
and HGC-27 cell lines using an anti-S100A7/psoriasin antibody
(clone 47C1068). *P<0.05, **P<0.01 vs.
pEF control. PCR, polymerase chain reaction; pEF, cells transfected
with the empty plasmid vector; PSOexp, cells transfected with
psoriasin plasmid. |
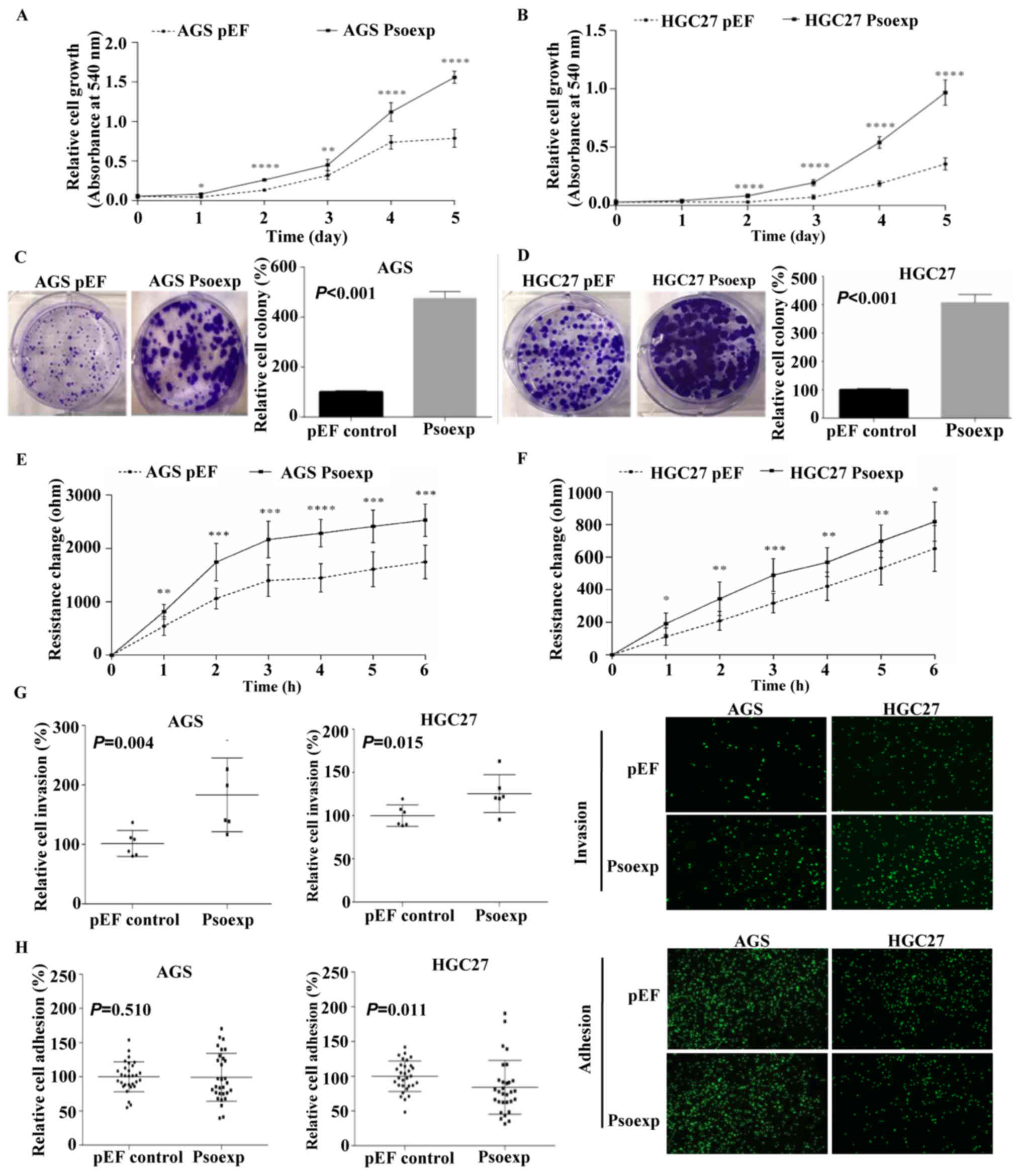
Psoriasin overexpression promotes cell
proliferation, invasion and migration in AGS and HGC-27, and
decreases adhesion in HGC-27 cells
AGS and HGC27 are two well-characterized gastric
cancer cell lines (25,26). As demonstrated in Fig. 1, the gene expression of psoriasin
is low in these two GC cell lines, as indicated by RT-qPCR
(Fig. 1C and D) and conventional
PCR (Fig. 1E). Western blotting
also demonstrated that the protein levels of psoriasin in AGS and
HGC27 were low (Fig. 1F). Attempts
were made to subculture other GC cell lines, such as NUGC-1,
MKN-45, MKN7 and MKN74. Unfortunately, they were either floating
(NUGC-1, MKN-45) or was difficult expand during the establishment
of the stable cells lines (MKN7 and MKN74). Therefore, AGS and
HGC-27 were to establish stable psoriasin-overexpressed cell lines
(AGS PSOexp and HGC-27 PSOexp) to determine the role of psoriasin
in the tumorigenesis and progression of GC. The mRNA and protein
levels of psoriasin was markedly increased in AGS PSOexp and HGC-27
PSOexp cells compared with the cells that were transfected with the
vector alone (AGS pEF and HGC-27 pEF) as verified by RT-qPCR
(Fig. 1C and D), PCR (Fig. 1E) and western blotting (Fig. 1F).
There was a significant increase in cell
proliferation in psoriasin-overexpressed cells compared with the
controls, which was assessed by crystal violet assay (P<0.001;
Fig. 2A and B). Similarly, cell
colony formation was also enhanced in the psoriasin-overexpressed
cells (Fig. 2C and D). However,
there was no effect on the cell cycle profile following psoriasin
overexpression (data not shown).
As psoriasin expression was positively associated
with lymph node involvement and distant metastasis (Table II), the migratory and invasive
abilities of GC cells were evaluated in vitro. The Matrigel
invasion assay and ECIS assay demonstrated that the overexpression
of psoriasin dramatically enhanced migration (Fig. 2E and F) and invasion (Fig. 2G) in GC cell lines compared with
the vector controls. Furthermore, upregulation of psoriasin
significantly decreased cell-matrix adhesion in HGC-27 cells, but
not in AGS cells (Fig. 2H).
Together, these data suggest that psoriasin may function as an
inducer of cell migration and invasion in GC.
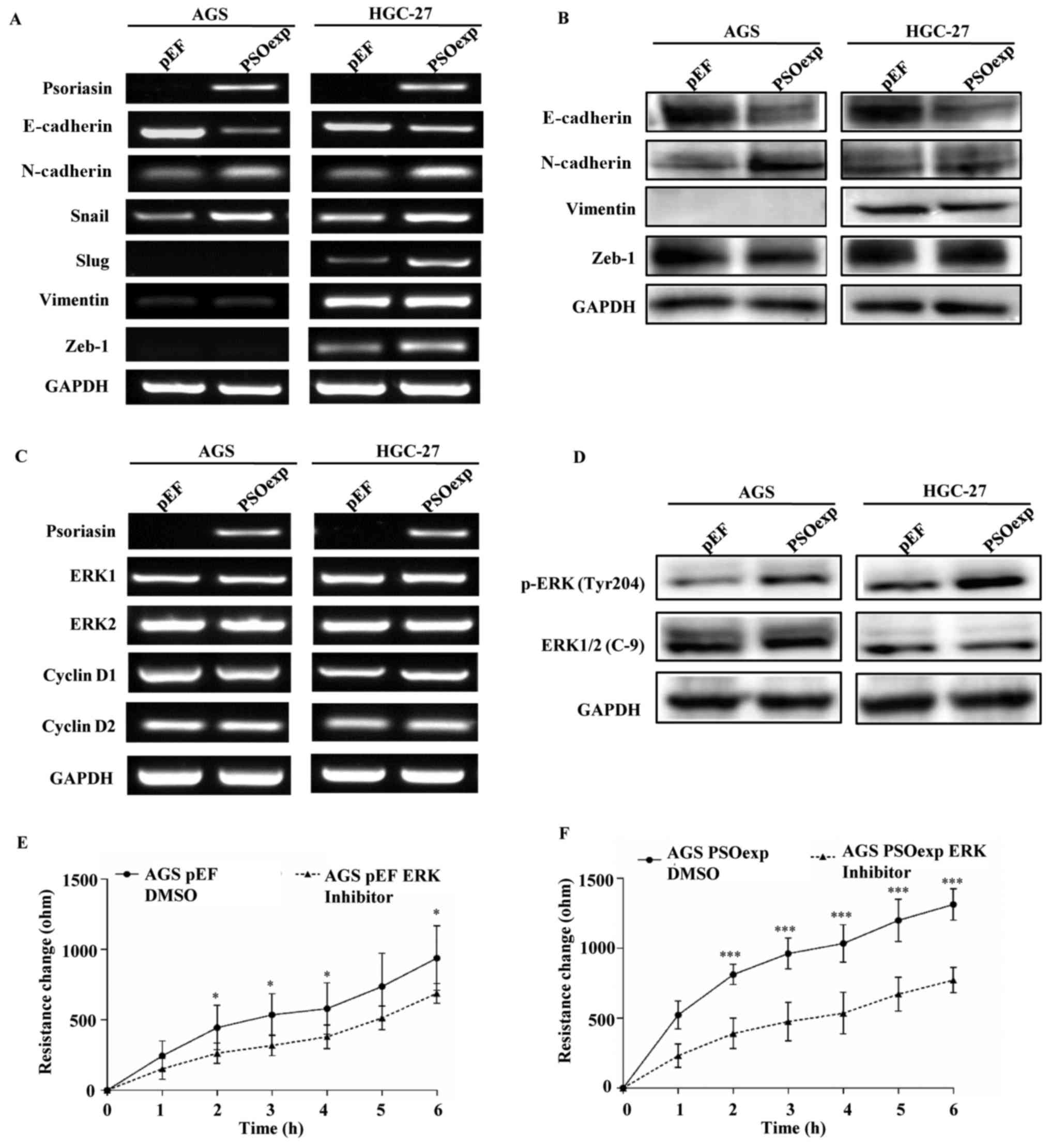
Expression of epithelial-mesenchymal
transition (EMT) markers in GC cells
Several EMT-associated molecules have been
implicated in the development and progression of GC by altering the
malignant properties of cancer cells (14,27).
Therefore, the expression of epithelial markers, mesenchymal
markers and EMT transcription factors were evaluated following
overexpression of psoriasin by PCR and western blotting. The
results demonstrated that the levels of N-cadherin and Snail
expression were increased, while E-cadherin expression was
decreased in AGS PSOexp and HGC-27 PSOexp cells compared with the
empty vector controls. In addition, the upregulation of psoriasin
promoted Slug and Zeb-1 expression levels in HGC-27 cells compared
with the control cells, whereas the vimentin level was unchanged in
both cell lines (Fig. 3A and B).
Collectively, the activation of psoriasin may alter the patterns of
EMT in GC cells, promoting cell invasion and migration.
Psoriasin enhances cell migration via
activating ERK signaling in GC cells
The ERK signaling cascade is one of the key
signaling pathways that regulates cell proliferation, invasion and
migration by controlling multiple nuclear and cytoplasmic targets
in cancer (28). In the present
study, the effect of psoriasin on the ERK signaling pathway was
investigated in GC. The upregulation of psoriasin significantly
promoted the level of phosphorylated ERK (Tyr 204) protein, without
altering the total ERK1/2 mRNA and protein levels in AGS and HGC-27
cells, compared with the control cells (Fig. 3C and D).
By contrast, there was no difference in the levels
of cyclin D1 and cyclin D2 mRNA between the control cells and the
cells that overexpressed psoriasin (Fig. 3C). Therefore, the effect of
psoriasin on the viability of GC cells was unlikely due to the
regulation of these regulatory proteins of cyclin-dependent
kinases.
Treatment with an ATP-competitive ERK inhibitor
suppressed the migratory ability of AGS cells (PSOexp cells and
control cells), particularly in AGS PSOexp cells (Fig. 3E and F). Together, these results
suggest that psoriasin may enhance cell migration at least
partially via activation of ERK. Further study is required to
determine whether ERK is a direct substrate of psoriasin.
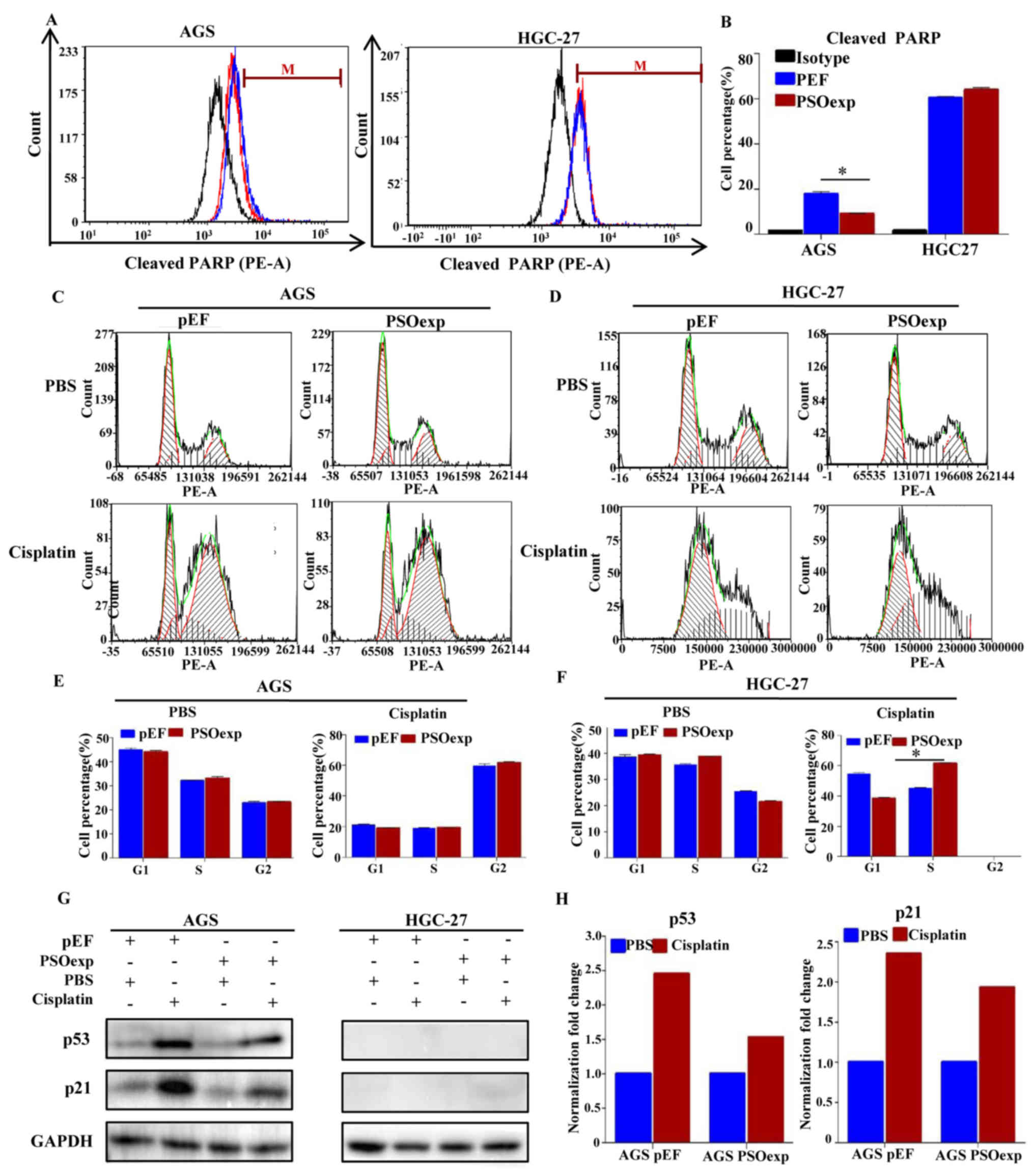
Psoriasin overexpression increases
resistance to cisplatin in GC cells
To determine the effect of psoriasin on cisplatin
resistance in GC, the gene expression of psoriasin was analyzed in
specimens from patients with GC that received pre-operative
chemotherapy (n=87). The results indicated that levels of psoriasin
expression were higher in patients that exhibited no response to
neoadjuvant chemotherapy (P=0.023; Table III). Flow cytometry analysis of
cleaved PARP was performed to evaluate the levels of apoptosis in
the stable cell lines that were treated with cisplatin. As
presented in Fig. 4A and B, in the
presence of cisplatin, the less AGS PSOexp cells expressed cleaved
PARP protein (8.95%) than the AGS pEF cells (16.87%), suggesting an
increase in resistance to cisplatin. However, such increase was not
observed in HGC-27 cells, as the cleaved PARP expression was not
markedly altered between the HGC-27 PSOexp cell line (63.03%) and
the HGC-27 pEF cell line (59.38%; Fig.
4A and B). The cell cycle analysis indicated that in response
to the cisplatin treatment, a higher portion of AGS cells was
arrested in the G2/M phase compared with PBS treatment in AGS pEF
cells (58.68% vs. 22.51%) and AGS PSOexp cells (61.04% vs. 23.55%).
However, there was no difference in cell cycle profile between the
AGS PSOexp and AGS pEF cell lines in the presence of cisplatin. By
contrast, the S-phase population of the HGC-27 PSOexp cells was
increased compared with the pEF control cells in the presence of
cisplatin (44.68% in HGC-27 pEF cells and 60.84% in HGC-27 PSOexp
cells; Fig. 4C–F). Furthermore,
psoriasin overexpression in AGS cells inhibited the
cisplatin-induced increase in the levels of p53 and p21 proteins
compared with the cells transfected with the backbone pEF plasmid
(Fig. 4G and H). However, the cell
viability assay showed that although psoriasin overexpression
improved survival rates in the presence of particular doses of
cisplatin (e.g. 6.25 µM) in transfected AGS and HGC-27 cells
compared with the pEF controls (P<0.05), the overall
dose-dependent cytotoxic effect of cisplatin on the cell lines was
similar to the control cell lines (data not shown). Therefore, the
upregulation of psoriasin in GC cells may desensitize cells to
cisplatin by promoting the proportion of cells in the S-phase or
inhibiting apoptosis, instead of a direct effect via cellular
cytotoxicity.
| Table IIIQuantitative gene expression analysis
of psoriasin in patients with gastric cancer receiving
chemotherapy. |
Table III
Quantitative gene expression analysis
of psoriasin in patients with gastric cancer receiving
chemotherapy.
| Variable | n | Psoriasin
expression (median) | P-value |
|---|
| Tissue | | | 0.005 |
| Tumor | 87 | 0.40 | |
| Adjacent
normal | 83 | 1.09 | |
| Sex | | | 0.816 |
| Male | 63 | 0.41 | |
| Female | 24 | 0.33 | |
|
Differentiation | | | |
|
Differentiated | 20 | 0.53 | |
|
Undifferentiated | 59 | 0.41 | 0.500a |
| Others | 8 | 0.24 | 0.102a |
| Tumor size
(mm) | | | 0.202 |
| >50 | 9 | 0.41 | |
| <50 | 78 | 0.29 | |
| Depth of
invasion | | | 0.116 |
| T1–2 | 9 | 0.19 | |
| T3–4 | 78 | 0.41 | |
| Lymph node
status | | | 0.891 |
| Negative | 16 | 0.34 | |
| Positive | 71 | 0.41 | |
| TNM staging | | | 0.113 |
| I+II | 9 | 0.34 | |
| III+IV | 78 | 0.48 | |
| Response to
neoadjuvant chemotherapy | | | 0.023 |
| Response | 30 | 0.24 | |
| No response | 57 | 0.50 | |
Discussion
Psoriasin is dysregulated in multiple solid tumors
and performs multiple roles in a variety of pathophysiological
processes, which are closely associated with tumorigenesis and
cancer progression. However, knowledge on the expression patterns
and biological function of psoriasin in the progression of GC
remains limited. In the present study, there was an increased level
of psoriasin mRNA expression in GC tissues compared with adjacent
normal tissues. Furthermore, this increase in psoriasin expression
was also associated with lymph node involvement in patients,
distant metastasis and advanced stages of the disease. These
findings are consistent with the previous studies on cervical
cancer and breast cancer (18,29).
Additionally, Kaplan-Meier plot analysis indicated that higher
psoriasin expression led to worse overall survival and disease-free
survival, which was supported by the outcomes of meta-analysis from
the Kaplan-Meier Plotter (gastric cancer) database (http://kmplot.com/ analysis; data not shown). The
association between psoriasin expression and clinicopathological
characteristics indicates that psoriasin has a role in the
development and progression of GC.
To further clarify the roles of psoriasin in gastric
carcinogenesis and progression, the effects of upregulating
psoriasin on cell function were evaluated. In this study,
overexpression of psoriasin in AGS and HGC-27 GC cell lines
promoted cell proliferation, migration and invasion, and inhibited
adhesion in the HGC-27 cell line. A recent study reported that
psoriasin may promote cyclin activation and cell cycle progression
in pancreatic cancer cells (30).
However, in the present study psoriasin had no effect on cell cycle
distribution and gene expression of cell cycle regulators.
EMT is a biological process that allows polarized
epithelial cells to undergo multiple changes in order to acquire a
mesenchymal cell phenotype, therefore enhancing cell migration,
invasion and metastasis (31).
Loss of E-cadherin and increased expression of N-cadherin are
hallmarks of EMT, which promote the interaction of the cell with
endothelial and stromal components (27,32,33).
Notably, the present data demonstrated that psoriasin drives the
upregulation of N-cadherin, Snail, Slug and Zeb-1 expression and
loss of E-cadherin expression in GC cells. Snail, Slug and Zeb-1,
which are important transcription factors, trigger EMT by
coordinating the induction of mesenchymal biomarkers and the
repression of epithelial biomarkers (27). Collectively, these results suggest
that psoriasin may be a potent inducer of EMT, which regulates cell
invasion and migration in GC. However, the precise mechanism by
which this occurs in GC requires further investigation.
ERK signaling, one of the most important
intracellular pathways, has multiple functions in cancer cells,
including roles in proliferation, cell cycle, EMT, migration,
invasion, repression of apoptosis and induction of drug resistance
(34–36). In the present study, migration was
notably increased by ERK activation induced by psoriasin
overexpression. Furthermore, the migration of AGS cells induced by
psoriasin overexpression was inhibited by treatment with ERK
inhibitor. Therefore, psoriasin may promote EMT by activating the
ERK pathway, leading to enhanced migration and invasion of GC
cells. However, the specific mechanism by which psoriasin affects
cell migration and invasion requires further investigation.
Clinical data indicated that patients that
positively responded to pre-operative chemotherapy had a
significantly longer survival time than non-responders (5). Many studies have reported that the
S100 proteins have the ability to regulate the sensitivity of tumor
cells to chemotherapeutics (37–39).
For instance, S100A11 sensitizes cells to cisplatin treatment in
non-small cell lung cancer (40).
However, to date, there has been no investigation into the
association between psoriasin expression and chemotherapy
resistance in tumors. To the best of our knowledge, the present
study was the first to observe that elevated psoriasin expression
in patients with GC was significantly associated with a poor
pathological response to chemotherapy, suggesting that psoriasin
might confer resistance in GC to chemotherapeutic drugs. In
vitro experiments demonstrated that cells with a higher level
of psoriasin expression exhibited less sensitivity to cisplatin
compared with the control cells. The molecular basis of resistance
to chemotherapy is generally complex, involving multiple processes,
including drug metabolism and transport, apoptosis and DNA repair
(41). In order to investigate the
underlying mechanism, the expression cleaved PARP was used as an
indicator of apoptosis and the cell cycle following treatment of
cells with cisplatin. Based on the results, psoriasin may inhibit
the sensitivity of GC cells to cisplatin by ameliorating cell
apoptosis or inducing the S-phase arrest, depending on the
molecular phenotype, epigenetic and genetic features or p53 status
of the cells.
However, there were certain limitations of the
present study. As AGS and HGC-27 cells were available, they were
used to explore the effect of psoriasin overexpression in GC cells.
It may be ideal to find a GC cell line that overexpresses S100A7
and possesses the phenotype of GC, and explore the further
biological functions via psoriasin knockdown. In addition, as AGS
and HGC27 cells have low tumor formation ability in mice, this
study presented the role of psoriasin in GC cells via in
vitro experiments only. In the future, another GC cell line
should be used to determine the role of psoriasin in
vivo.
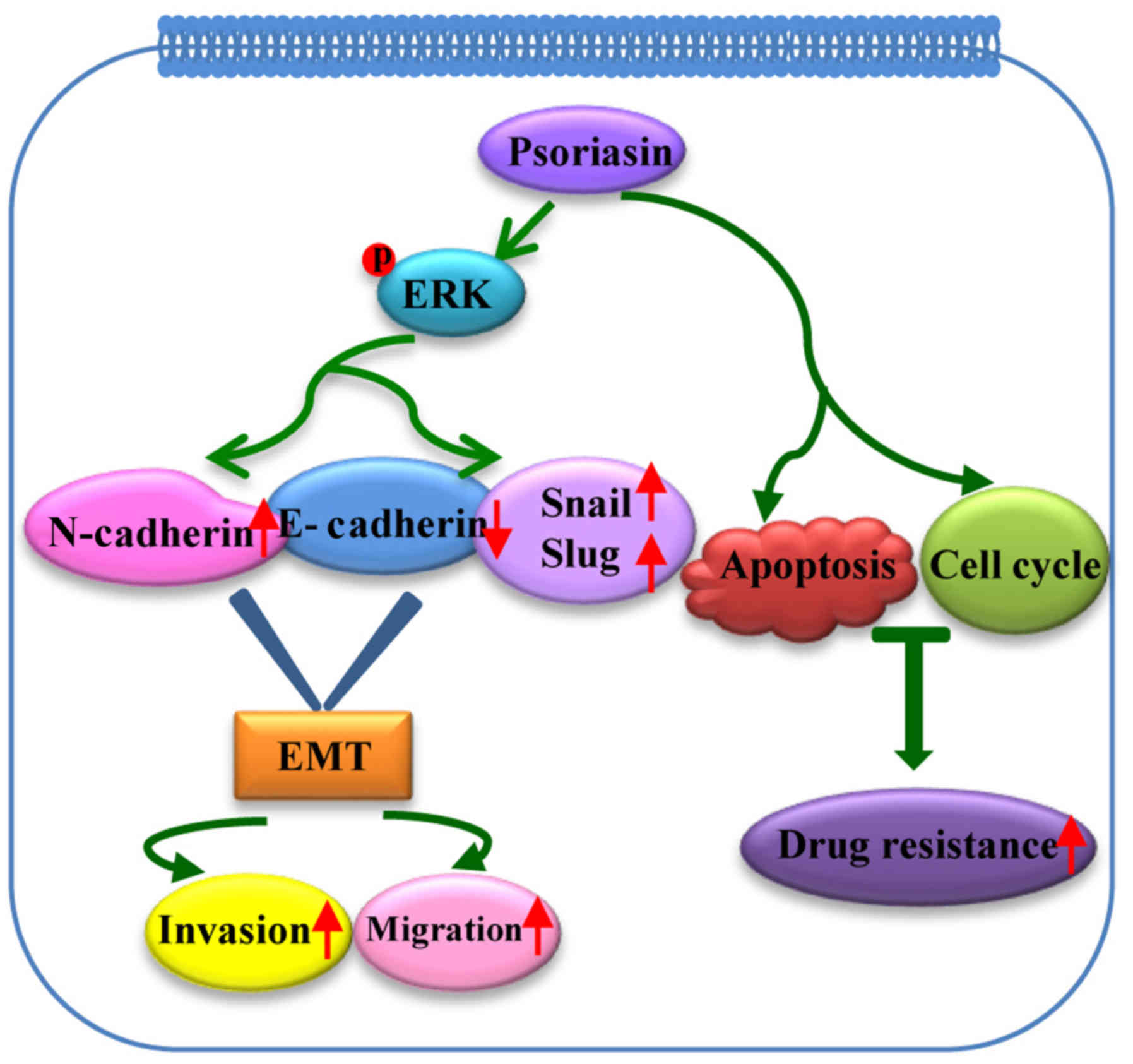
In summary, to our best knowledge, this is the first
report describing the role of psoriasin in GC and chemotherapy
resistance in human tumors. Psoriasin expression is frequently
increased in human GC cell lines and GC tissues, and its expression
is associated with metastasis and a poor survival in patients with
GC. Psoriasin overexpression in GC cells increased cell
proliferation, migration and invasion. Furthermore, the promoting
effects of psoriasin are potentially attributed to a positive
effect on EMT-associated pathways via activation of ERK signaling
(Fig. 5). As psoriasin
overexpression leads to a higher tolerance to cisplatin in GC
cells, patient with GC with a high level of psoriasin may be less
sensitive to cisplatin. Therefore, the findings suggest that
psoriasin has the potential as a therapeutic target in GC and may
be a useful prognostic factor to predict the effectiveness of
pre-operative chemotherapy in patients with GC.
Acknowledgments
The authors would like to thank Mrs. Fiona Ruge
(Cardiff University, Cardiff, UK) for her technical assistance.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Van Cutsem E, Sagaert X, Topal B,
Haustermans K and Prenen H: Gastric cancer. Lancet. 388:2654–2664.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kanat O and O'Neil BH: Metastatic gastric
cancer treatment: A little slow but worthy progress. Med Oncol.
30:4642013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ohtsu A: Chemotherapy for metastatic
gastric cancer: Past, present, and future. J Gastroenterol.
43:256–264. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cunningham D, Allum WH, Stenning SP,
Thompson JN, Van de Velde CJ, Nicolson M, Scarffe JH, Lofts FJ,
Falk SJ, Iveson TJ, et al MAGIC Trial Participants: Perioperative
chemotherapy versus surgery alone for resectable gastroesophageal
cancer. N Engl J Med. 355:11–20. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee JH, Kim KM, Cheong JH and Noh SH:
Current management and future strategies of gastric cancer. Yonsei
Med J. 53:248–257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Madsen P, Rasmussen HH, Leffers H, Honoré
B, Dejgaard K, Olsen E, Kiil J, Walbum E, Andersen AH, Basse B, et
al: Molecular cloning, occurrence, and expression of a novel
partially secreted protein 'psoriasin' that is highly up-regulated
in psoriatic skin. J Invest Dermatol. 97:701–712. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Watson PH, Leygue ER and Murphy LC:
Psoriasin (S100A7). Int J Biochem Cell Biol. 30:567–571. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jinquan T, Vorum H, Larsen CG, Madsen P,
Rasmussen HH, Gesser B, Etzerodt M, Honoré B, Celis JE and
Thestrup-Pedersen K: Psoriasin: A novel chemotactic protein. J
Invest Dermatol. 107:5–10. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Enerbäck C, Porter DA, Seth P, Sgroi D,
Gaudet J, Weremowicz S, Morton CC, Schnitt S, Pitts RL, Stampl J,
et al: Psoriasin expression in mammary epithelial cells in vitro
and in vivo. Cancer Res. 62:43–47. 2002.PubMed/NCBI
|
|
11
|
Zhou G, Xie TX, Zhao M, Jasser SA, Younes
MN, Sano D, Lin J, Kupferman ME, Santillan AA, Patel V, et al:
Reciprocal negative regulation between S100A7/psoriasin and
beta-catenin signaling plays an important role in tumor progression
of squamous cell carcinoma of oral cavity. Oncogene. 27:3527–3538.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deol YS, Nasser MW, Yu L, Zou X and Ganju
RK: Tumor-suppressive effects of psoriasin (S100A7) are mediated
through the β-catenin/T cell factor 4 protein pathway in estrogen
receptor-positive breast cancer cells. J Biol Chem.
286:44845–44854. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang H, Zhao Q, Chen Y, Wang Y, Gao S,
Mao Y, Li M, Peng A, He D and Xiao X: Selective expression of
S100A7 in lung squamous cell carcinomas and large cell carcinomas
but not in adenocarcinomas and small cell carcinomas. Thorax.
63:352–359. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang L, Wu RL and Xu AM:
Epithelial-mesenchymal transition in gastric cancer. Am J Transl
Res. 7:2141–2158. 2015.
|
|
15
|
Moubayed N, Weichenthal M, Harder J,
Wandel E, Sticherling M and Gläser R: Psoriasin (S100A7) is
significantly up-regulated in human epithelial skin tumours. J
Cancer Res Clin Oncol. 133:253–261. 2007. View Article : Google Scholar
|
|
16
|
Tripathi SC, Matta A, Kaur J, Grigull J,
Chauhan SS, Thakar A, Shukla NK, Duggal R, DattaGupta S, Ralhan R,
et al: Nuclear S100A7 is associated with poor prognosis in head and
neck cancer. PLoS One. 5:e119392010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ye L, Sun PH, Martin TA, Sanders AJ, Mason
MD and Jiang WG: Psoriasin (S100A7) is a positive regulator of
survival and invasion of prostate cancer cells. Urol Oncol.
31:1576–1583. 2013. View Article : Google Scholar
|
|
18
|
Tian T, Li X, Hua Z, Ma J, Wu X, Liu Z,
Chen H and Cui Z: S100A7 promotes the migration, invasion and
metastasis of human cervical cancer cells through
epithelial-mesenchymal transition. Oncotarget. 8:24964–24977.
2017.PubMed/NCBI
|
|
19
|
Nasser MW, Wani NA, Ahirwar DK, Powell CA,
Ravi J, Elbaz M, Zhao H, Padilla L, Zhang X, Shilo K, et al: RAGE
mediates S100A7-induced breast cancer growth and metastasis by
modulating the tumor microenvironment. Cancer Res. 75:974–985.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Emberley ED, Niu Y, Curtis L, Troup S,
Mandal SK, Myers JN, Gibson SB, Murphy LC and Watson PH: The
S100A7-c-Jun activation domain binding protein 1 pathway enhances
prosurvival pathways in breast cancer. Cancer Res. 65:5696–5702.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: the 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Keese CR, Wegener J, Walker SR and Giaever
I: Electrical wound-healing assay for cells in vitro. Proc Natl
Acad Sci USA. 101:1554–1559. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Szulcek R, Bogaard HJ and van Nieuw
Amerongen GP: Electric cell-substrate impedance sensing for the
quantification of endothelial proliferation, barrier function, and
motility. Journal of visualized experiments J Vis Exp.
85:513002014.
|
|
24
|
Wegener J, Keese CR and Giaever I:
Electric cell-substrate impedance sensing (ECIS) as a noninvasive
means to monitor the kinetics of cell spreading to artificial
surfaces. Exp Cell Res. 259:158–166. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Keates S, Sougioultzis S, Keates AC, Zhao
D, Peek RM Jr, Shaw LM and Kelly CP: cag+ Helicobacter pylori
induce transactivation of the epidermal growth factor receptor in
AGS gastric epithelial cells. J Biol Chem. 276:48127–48134. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Akagi T and Kimoto T: Human cell line
(HGC-27) derived from the metastatic lymph node of gastric cancer.
Acta Med Okayama. 30:215–219. 1976.PubMed/NCBI
|
|
27
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chambard JC, Lefloch R, Pouysségur J and
Lenormand P: ERK implication in cell cycle regulation. Biochim
Biophys Acta. 1773:1299–1310. 2007. View Article : Google Scholar
|
|
29
|
Jiang WG, Watkins G, Douglas-Jones A and
Mansel RE: Psoriasin is aberrantly expressed in human breast cancer
and is related to clinical outcomes. Int J Oncol. 25:81–85.
2004.PubMed/NCBI
|
|
30
|
Liu Y, Bunston C, Hodson N, Resaul J, Sun
PH, Cai S, Chen G, Gu Y, Satherley LK, Bosanquet DC, et al:
Psoriasin promotes invasion, aggregation and survival of pancreatic
cancer cells; association with disease progression. Int J Oncol.
50:1491–1500. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kalluri R and Neilson EG:
Epithelial-mesenchymal transition and its implications for
fibrosis. J Clin Invest. 112:1776–1784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Son H and Moon A: Epithelial-mesenchymal
transition and cell invasion. Toxicol Res. 26:245–252. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Scanlon CS, Van Tubergen EA, Inglehart RC
and D'Silva NJ: Biomarkers of epithelial-mesenchymal transition in
squamous cell carcinoma. J Dent Res. 92:114–121. 2013. View Article : Google Scholar :
|
|
34
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar
|
|
35
|
Steelman LS, Chappell WH, Abrams SL, Kempf
RC, Long J, Laidler P, Mijatovic S, Maksimovic-Ivanic D, Stivala F,
Mazzarino MC, et al: Roles of the Raf/MEK/ERK and
PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity
to therapy-implications for cancer and aging. Aging (Albany NY).
3:192–222. 2011. View Article : Google Scholar
|
|
36
|
Abrams SL, Steelman LS, Shelton JG, Wong
EW, Chappell WH, Bäsecke J, Stivala F, Donia M, Nicoletti F, Libra
M, et al: The Raf/MEK/ERK pathway can govern drug resistance,
apoptosis and sensitivity to targeted therapy. Cell Cycle.
9:1781–1791. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shiota M, Tsunoda T, Song Y, Yokomizo A,
Tada Y, Oda Y and Naito S: Enhanced S100 calcium-binding protein P
expression sensitizes human bladder cancer cells to cisplatin. BJU
Int. 107:1148–1153. 2011. View Article : Google Scholar
|
|
38
|
Acharyya S, Oskarsson T, Vanharanta S,
Malladi S, Kim J, Morris PG, Manova-Todorova K, Leversha M, Hogg N,
Seshan VE, et al: A CXCL1 paracrine network links cancer
chemoresistance and metastasis. Cell. 150:165–178. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Arumugam T, Ramachandran V, Sun D, Peng Z,
Pal A, Maxwell DS, Bornmann WG and Logsdon CD: Designing and
developing S100P inhibitor 5-methyl cromolyn for pancreatic cancer
therapy. Mol Cancer Ther. 12:654–662. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zagryazhskaya A, Surova O, Akbar NS,
Allavena G, Gyuraszova K, Zborovskaya IB, Tchevkina EM and
Zhivotovsky B: Tudor staphylococcal nuclease drives
chemo-resistance of non-small cell lung carcinoma cells by
regulating S100A11. Oncotarget. 6:12156–12173. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dai Z, Huang Y and Sadée W: Growth factor
signaling and resistance to cancer chemotherapy. Curr Top Med Chem.
4:1347–1356. 2004. View Article : Google Scholar : PubMed/NCBI
|