Introduction
With approximately 1.8 million new diagnosed cases
in 2012, lung cancer is one of the most common types of cancer,
accounting for the most frequent cancer-related deaths (1.6
million) worldwide (1).
Approximately 80-85% of these lung cancer patients suffer from the
subtype, non-small cell lung cancer (NSCLC). The reasons for the
high mortality rate of this cancer type are not only the late
detection of the disease, but also due to the limited treatment
options. Mutations within the epidermal growth factor
receptor (EGFR) occur frequently in NSCLC. Often this
mutation leads to a constitutive activation of the receptor and
increases downstream signalling cascades, such as the
mitogen-activated protein kinase kinase (MEK)/extracellular-signal
regulated kinases (ERK), phosphoinositide 3-kinase (PI3K)/protein
kinase B (AKT) and signal transducer and activator (STAT) pathway.
Consequently, proliferation and angiogenesis, as well as metastasis
are increased and apoptosis is decreased (2), which promotes tumour growth.
Glycodelin [gene name, progesterone associated
endometrial protein (PAEP)] is a protein initially
described in reproduction. The four isoforms glycodelin A, S
(3,4), F and C (5), differ in their glycosylation
patterns. Each of them has different biological functions, such as
influencing the capacitation, acrosome reaction, sperm-oocyte
binding and immune system suppression during the establishment of
pregnancy (6,7). In the female reproduction system,
glycodelin synthesis can be increased by progesterone (8), human chorionic gonadotropin (hCG)
(9,10) and relaxin (11).
Additionally, PAEP/glycodelin is known to be
expressed not only in female-specific tumours, such as breast
(12), endometrial (13), ovarian (14) and cervical cancer (15), but also in biphasic synovia
sarcomas (16), melanoma (17), malignant pleural mesothelioma
(18) and lung cancer (19,20).
It has been shown that PAEP/glycodelin expression is
associated with both a better and worse prognosis in these
different cancer types (20-23).
However, in general, glycodelin seems to promote tumour malignancy
by influencing proliferation, differentiation, invasion,
angiogenesis and the immune system (24). In a previous study, we identified
glycodelin as a potent follow-up biomarker for NSCLC, since
increased levels of glycodelin were associated with recurrence and
metastatic disease in patients' sera (20). Furthermore, an altered cell
structure, a reduced migration and the upregulation of immune
system-regulating ligands was observed in NSCLC cell lines in which
PAEP expression was silenced (20). These results suggest that NSCLC
tumours secrete glycodelin to overcome immune surveillance.
Therefore, glycodelin may be a potential target to weaken immune
system defence of NSCLC tumours. Pathways regulating
PAEP/glycodelin expression in cancer might provide some
insight in this respect. However, they are, particularly in lung
cancer, mostly unknown. A few inducers and components of pathways,
which are commonly altered in cancer, have already been described
to influence glycodelin expression. In a myelogenous leukaemia cell
line, PAEP/glycodelin expression was shown to be stimulated
by phorbol 12-myristate 13 acetate (PMA) (25). Lysophosphatidic acid (LPA) has also
been shown to elevate PAEP/glycodelin expression in
cervical, endometrial, ovarian cancer and myelogenous leukaemia
cell lines (26). Since PMA and
LPA are known as protein kinase C (PKC) activators (27,28),
it might be of interest to determine whether PAEP/glycodelin
expression is influenced by this kinase. Furthermore, the
inhibition of MEK1/2 and protein kinase A has been described to
attenuate phytoestrogen-induced glycodelin expression (29). Other previous studies have
postulated that the activation of PAEP expression is
associated with the transcription factors GATA-binding protein 3
(GATA3), the microphthalmia-associated transcriptions factor (MITF)
and the specific protein1 (Sp1) (30-32).
Only the Krüppel-like transcription factor11 (KLF11) has been
assumed to suppress glycodelin expression (33).
Based on these facts, the aim of this study was to
gain insight into the so far unknown underlying mechanisms
regulating PAEP/glycodelin expression in NSCLC. Therefore,
we analysed the effects of various pathway inducers and their
downstream signalling cascades, all of which are known as major
regulators in cancer, on PAEP/glycodelin expression in two
NSCLC cell lines. Moreover, the in vitro data were validated
in NSCLC patient tissue.
Materials and methods
Tissue sample collection
Tissue samples from patients with NSCLC were
provided by the Lung Biobank Heidelberg, a member of the accredited
Tissue Bank of the National Centre for Tumour Diseases (NCT)
Heidelberg, the BioMaterialBank Heidelberg and the Biobank Platform
of the German Centre for Lung Research (DZL). Written informed
consent was obtained from all participants and/or their legal
guardian/s before the use of the tissue for research purpose. The
study was approved by the local Ethics Committee of the University
of Heidelberg (no. 270/2001) and all experiments were performed in
accordance with relevant guidelines and regulations. A total of 179
patients with NSCLC, who underwent surgical resection at the
Thoraxklinik Heidelberg, were included. Tumour tissue, as well as
the corresponding healthy lung parenchyma, with a distance of >5
cm from the tumour, was used. A pathologist made the diagnosis in
compliance with the World Health Organization (WHO) classification
for lung cancer from 2004 (34).
Tumours were staged according to the 7th edition of the Union for
International Cancer Control's (UICC) tumour, node and metastasis
(35). Following surgical
resection, tissues were snap-frozen in liquid nitrogen within 30
min and stored at −80°C until subsequent processing.
Cell culture
The H1975 lung adenocarcinoma (ADC) cell line was
purchased from American Type Culture Collection (CRL-5908; ATCC,
Manassas, VA, USA) and authenticated by DNA profiling using 8
different and highly polymorphic short tandem repeat (STR)
(Leibniz-Institut DSMZ, Braunschweig, Germany). The 2106T cells
were generated from a human lung squamous cell carcinoma (SQCC) and
characterised as previously described (36). Both cell lines were maintained in
DMEM/Ham's F-12 (Thermo Fisher Scientific, Carlsbad, CA, USA)
supplemented with 1% GlutaMAXTM 100x (Thermo Fisher Scientific) and
10% foetal calf serum (FCS; Thermo Fisher Scientific).
siRNA-mediated gene depletion
The H1975 and 2106T cells were seeded into a 12-well
plate at an initial density of 4×104 cells per well. The
following day, the cells were transfected with small interfering
ribonucleic acids (siRNAs; Qiagen, Hilden, Germany) targeting JUNB
(JUNB_3: acagactcgattcatattgaa; JUNB_4: aaacacgcacttagtctctaa;
JUNB_5: cccgacgaccaccatcagcta), NF-κB1 (NFκB1_7:
tacctggtgcctctagtgaaa; NFκB1_8: tcagttggtcacaaatggaaa; NFκB1_10:
gacgccatctatgacagtaaa) and STAT3 (STAT3_3: ctggtcttaactctgattgta;
STAT3_4: cacctttgagaccgaggtgta; STAT3_7: cagcctctctgcagaattcaa;
STAT3_8: caggctggtaatttatataat) using Lipofectamine™ RNAiMax
(Thermo Fisher Scientific) according to the manufacturer's
instructions. Therefore, a pool of 3 to 4 different siRNAs, as well
as the particular single siRNAs were used. AllStars negative
control siRNA (Qiagen) served as a non-silencing control. The
siRNAs were applied at a final concentration of 10 nM. At 72 h
following transfection, the cells were processed for total RNA
isolation or western blot analysis.
Applying signalling pathway
modulators
Both cell lines were seeded into a 12-well plate at
1.6×105 cells per well. The following day, the cells
were serum-starved for approximately 16 h. For determining
PAEP/glycodelin expression, the cells were subsequently
treated with distinct pathway inducers or modulators for 24 h,
solely the PKC activator Bryostatin (Santa Cruz Biotechnology,
Heidelberg, Germany) was used for 1 h. To detect phosphorylated
pathway components or transcription factors, the cells were treated
only for 30 min to 1 h. Following the different treatments, the
cells were prepared for total ribonucleic acid (RNA) isolation or
western blot analysis. The following signalling pathway inducers
and modulators were used: LPA (Santa Cruz Biotechnology), PMA
(Cayman Chemical, Ann Arbor, MI, USA), epidermal growth factor
(EGF; Biomol, Hamburg, Germany), heparin-binding (HB)-EGF (Biomol),
TGF-β1, -2, -3 (WuXi Biosciences, San Diego, CA, USA), Bryostatin1
(Selleckchem, Houston, TX, USA), GF109203X (StressMarq Biosciences,
Victoria, BC, Canada), MK-2206 (Selleckchem) as well as RO5126766
(Selleckchem).
Cell lysis and western blot analysis
The cells were lysed with 2X sodium dodecyl sulfate
(SDS) sample buffer (0.13 M Tris HCL pH 6.8, 10% glycerol, 4% SDS,
1.4 mM β-mercaptoethanol and 1 % pyronine) and boiled for 5 min at
95°C. After separating the samples on a SDS-polyacrylamide gel (10
or 15%, self-made), they were transferred to a nitrocellulose
membrane. According to the manufacturer's instructions, the
membrane was incubated wih the primary antibodies overnight at 4°C
and subsequent incubation with the secondary antibody was performed
for 1 h at room temperature. The following primary antibodies were
used: β-actin (1:10,000, AC-15; anti-mouse; cat. no. A5441;
Sigma-Aldrich, Steinheim, Germany), glycodelin 1:300 (N20;
anti-goat; cat. no. sc-12289; Santa Cruz Biotechnology), Smad2/3
1:1,500 (anti-mouse; cat. no. 610843; BD Biosciences, Franklin
Lakes, NJ, USA), AKT (pan; 1:2,000; C67E7; anti-rabbit; cat. no.
4691), JUNB (1:2,500; C37F9; anti-rabbit; cat. no. 3753), NF-κB
p105/p50 (1:2,000; anti-rabbit; cat. no. 3035), p44/42 MAPK
(Erk1/2; 137F5; 1:2,000; anti-rabbit; cat. no. 4695), phospho-AKT
(Thr308; 1:1,000; D25E6; anti-rabbit; cat. no. 13038)
XP®, phospho-JUNB (Thr102/Thr104; 1:2,500; D3C6;
anti-rabbit; cat. no. 8053), phospho-NF-κB p105 (Ser933; 1:1,000;
18E6; anti-rabbit; cat. no. 4806), phospho-p44/42 MAPK (Erk1/2;
Thr202/Tyr204; 1:2,000; anti-rabbit; cat. no. 9101), phospho-PKC
substrate motif [(R/KXpSX(R/K)] MultiMabTM (1:750; anti-rabbit;
cat. no. 6967), phospho-Smad2 (Ser465/467; 1:1,000; 138D4;
anti-rabbit; cat. no. 3108), phospho-STAT3 (Ser727; 1:1,000; 6E4;
anti-mouse; cat. no. 9136) and STAT3 (1:1,500; 124H6; anti-mouse;
cat. no. 9139) (all from Cell Signalling Technology®,
Danvers, MA, USA). Horseradish peroxidase (HRP)-coupled secondary
antibodies (peroxidase-linked goat IgG (cat. no. A5420);
peroxidase-linked mouse IgG (cat. no. A4416); peroxidase-linked
rabbit IgG (cat. no. A6154); all 1:5,000) were purchased from
Sigma-Aldrich. The brightness and contrast of the western blot
images were adjusted using the software adobe photoshop elements13
(Adobe Systems, San Jose, CA, USA).
Total RNA isolation and cDNA
synthesis
For RNA isolation from patient tumour tissue, a
tumour content of ≥50% was the minimum prerequisite. A total of
10-15 tumour cryosections (10-15 µM) from each patient were
sliced and the first, as well as the last section of a series were
stained with haematoxylin and eosin (H&E). A lung pathologist
determined the proportion of viable tumour cells, stromal cells,
healthy lung cells and necrotic areas. Total RNA was isolated from
patient tissue using an AllPrep DNA/RNA/miRNA Universal kit
(Qiagen) according to the manufacturer's instructoins. An RNeasy
Mini kit (Qiagen) was applied to isolate RNA from the cell lines.
Afterwards, the quality of total RNA was assessed by utilizing an
Agilent 2100 Bioanalyser and an Agilent RNA 6000 Nano kit (Agilent
Technologies, Boeblingen, Germany). With the Transcriptor First
Strand cDNA Synthesis kit (Roche, Basel, Switzerland) the total RNA
was transcribed to complementary desoxyribonucleic acid (cDNA) and
used for quantitative polymerase chain reaction (qPCR). A complete
description of the procedure is provided elsewhere (20).
qPCR
A total of 5 ng of the cDNA was utilized for qPCR
with the LightCycler480® (Roche) in a 384-well plate
format according to the Minimum Information for Publication of qPCR
Experiments (MIQE)-guidelines (37). In the context of a Universal
ProbeLibrary (UPL) assay (Roche), gene-specific primers (TIB
Molbiol, Berlin, Germany) were combined with the primaQuant 2X qPCR
Probe-MasterMix (Steinbrenner Laborsysteme, Wiesenbach, Germany).
Threshold cycle (Ct)-values were evaluated with the
LightCycler480® software release 1.5 and the 2nd
derivative maximum method (Roche). The complete procedure is
described elsewhere (20). The
following primers and UPLs were used: ESD forward (UPL #50),
tcagtctgcttcagaacatgg and ESD reverse (UPL #50),
cctttaatattgcagccacga; JUNB forward (UPL #32), caaggtgaagacgctcaagg
and JUNB reverse (UPL #32), tcatgaccttctgtttgagctg; NF-κB forward
(UPL #49), cctggaaccacgcctcta and NF-κB reverse (UPL #49),
ggctcatatggtttcccattta; PAEP forward (UPL #77),
cctgtttctctgcctacagga and PAEP reverse (UPL #77),
cgtcctccaccaggactct; RPS forward (UPL #46), cttccacaggaggcctacac
and RPS reverse (UPL #46), cgcaaaatatgctggaacttt; and STAT3 forward
(UPL #17), gagcagagatgtgggaatgg and STAT3 (UPL #17) reverse,
cggtctcaaaggtgatcagg.
Microarray gene expression profiling
Isolated RNA from the cells and patient tissue was
further processed with the GeneChip™ 3' IVT PLUS Reagent kit
(Thermo Fisher Scientific) and the GeneChip™ Human Genome U133 Plus
2.0 Array (Thermo Fisher Scientific) according to the
manufacturer's instructions. For gene expression profiling of the
patient tissues, we selected samples with the highest or lowest
PAEP expression (40 ADCs and 30 SQCCs), which was determined
by qPCR analyses in our previous study (20). The raw data were normalized using
the software Expression Console™ (Thermo Fisher Scientific)
[Algorithm: robust multi-array average (RMA)] and analysed by
Transcriptome Analysis Console™ 3.0 (Thermo Fisher Scientific). For
further evaluation with the software Ingenuity pathway analysis
(IPA; IPA-42012434; Qiagen) (upstream regulator analysis),
only genes with an expression fold-change <−1.5 or >1.5 were
considered. A detailed description of the IPA analysis is available
on the manufacturer's homepage (https://www.qiagenbioinformatics.com/products/features/).
The microarray data, which were part of this study, are available
as described below.
Statistical analysis
Expression differences investigated by qPCR between
the control and pathway inducer/modulator-treated cells or control
siRNA- and siRNA-transfected cells were evaluated utilizing the
software GraphPad Prism 5 (GraphPad Software, La Jolla, CA, USA).
Statistical significance was calculated using one-way ANOVA with
Dunnett's test for multiple comparisons or an unpaired t-test for
single comparisons. A P-value <0.05 was considered to indicate a
statistically significant difference, since this study had an
explorative character.
Results
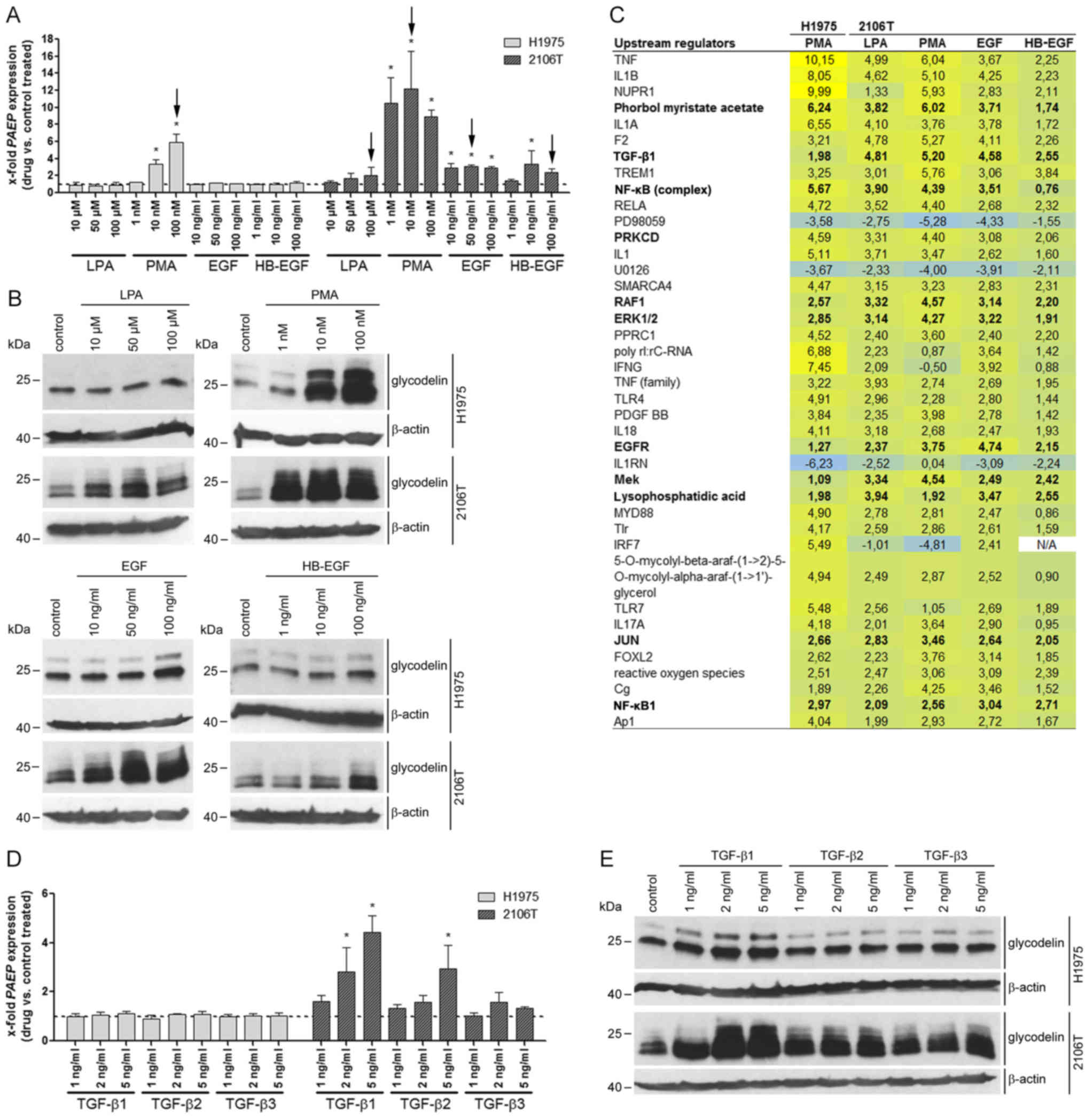
The pathway inducers LPA, PMA, EGF,
HB-EGF and TGF-β increase the expression of PAEP/glycodelin
In a recent study of ours, we screened different
lung ADC and lung SQCC cell lines regarding their glycodelin
expression (20). Among these
histological subtypes, H1975 [containing the EGFR mutations
(T790M and L858R), as well as the PIK3CA mutation (G118D)] and
2106T were the only cell lines that secreted glycodelin. In NSCLC,
various mutations activate different pathways, such as the MEK/ERK,
PI3K/AKT and/or STAT signalling cascades. This is also the case in
H1975 cells due to their EGFR and PIK3CA mutations.
By using the H1975 and 2106T cells in the following experiments, we
covered a rather representative range of mutation associated
activated, as well as unaffected pathways in NSCLC.
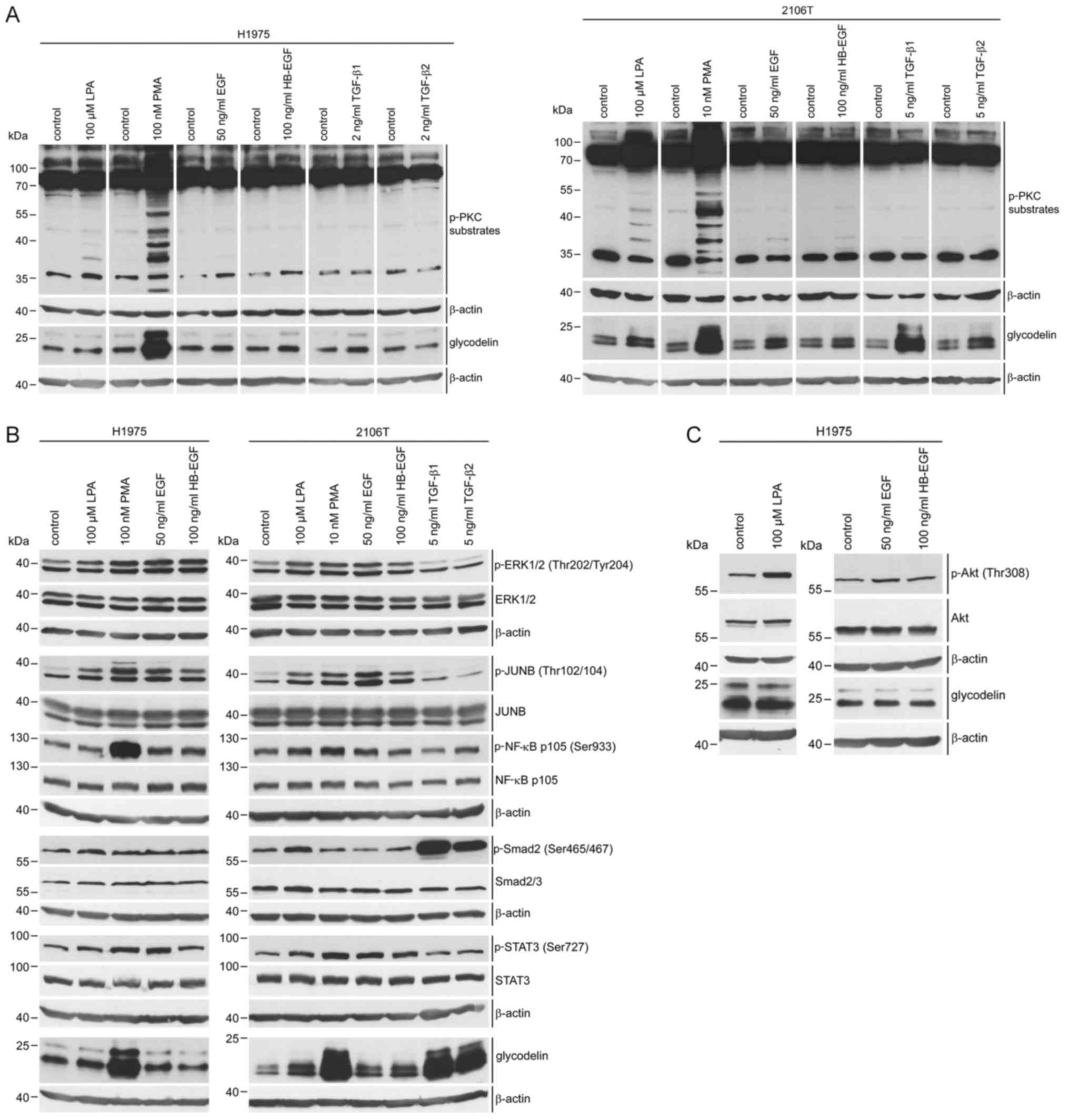
First, the effects of several pathway inducers on
PAEP/glycodelin expression were examined by qPCR and western
blot analysis in the H1975 and 2106T cells (Fig. 1A and B). Additionally, to the
already described PAEP/glycodelin expression stimulators LPA
and PMA (25,26), we also selected EGF, as well as
HB-EGF. Both are ligands of the EGFR, which plays a crucial role in
NSCLC (2). PMA increased
PAEP/glycodelin expression in the H1975 cells, whereas in
the 2106T cells, all of the inducers caused higher relative protein
levels. Herein, the most prominent effect was demonstrated by PMA
treatment (Fig. 1A and B).
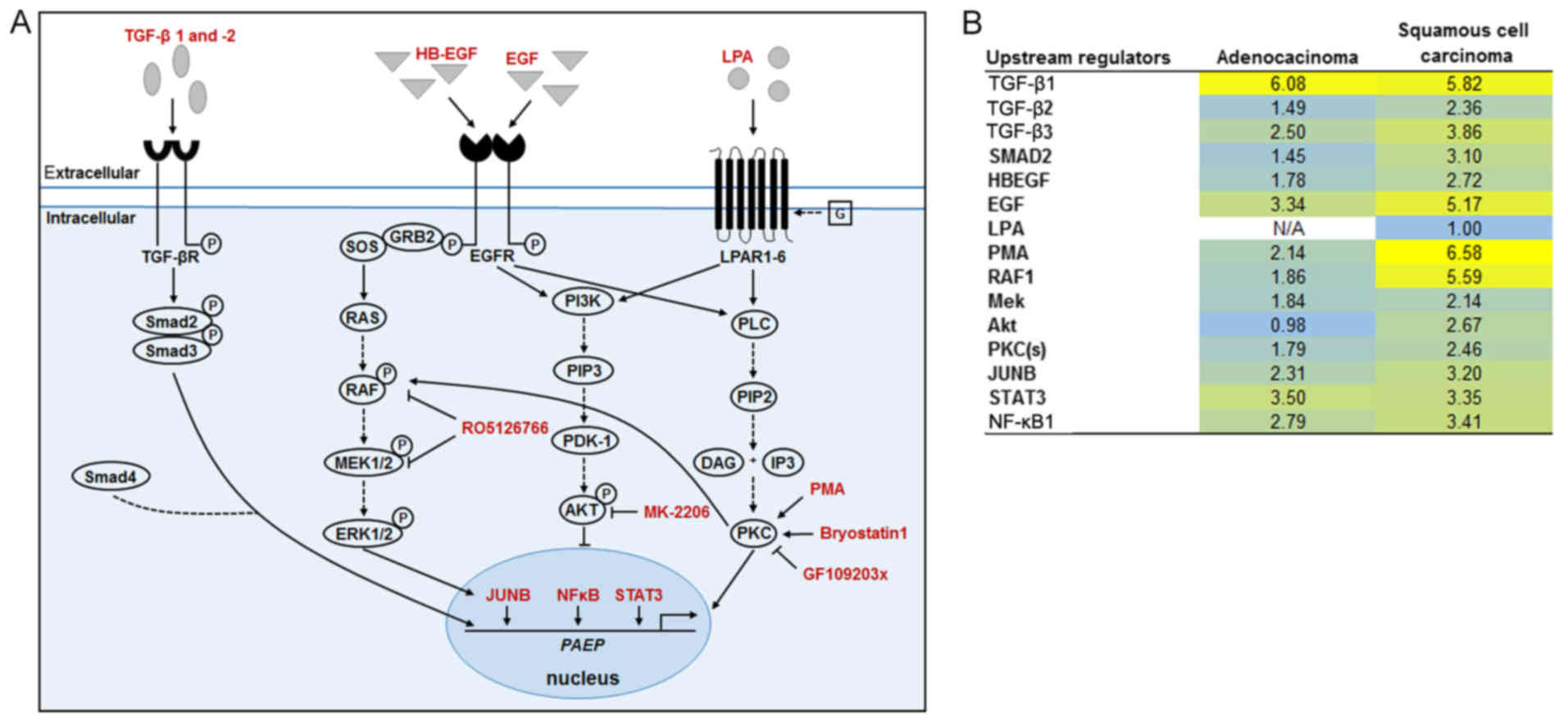
 | Figure 1PAEP/glycodelin expression is
stimulated by the pathway inducers, LPA, PMA, EGF, HB-EGF and TGF-β
in NSCLC cells. (A, B, D and E) Overnight serum-starved H1975 and
2106T cells were treated with the indicated pathway inducers for 24
h. (A and D) PAEP expression following pathway induction
compared to the controls is shown from 3 independent experiments.
Dotted line at 1 represents the expression in the control-treated
cells (mean of the Ct-values and mean SD are shown).
Black arrows mark the samples used in (C). Statistical significance
was defined as *P<0.05 (one-way ANOVA and
Dunnett's test; referring to control-treated cells). (B and E)
Glycodelin expression in cells treated with pathway inducers was
detected by western blot analysis. β-actin served as a loading
control (cropped blots are shown). (C) Cells were treated only with
pathway inducers that stimulated PAEP expression.
Corresponding microarray gene expression profiling data were
evaluated with an upstream regulator analysis by the software
Ingenuity Pathway Analysis (IPA). Shown are the activation z-scores
(significant for >2 or <−2). Upstream regulators are marked
in bold, if they were examined in further experiments. PAEP,
progesterone associated endometrial protein; NSCLC, non-small
cell lung cancer; LPA, lysophosphatidic acid; PMA, phorbol
12-myristate 13 acetate; EGF, epidermal growth factor; HB-EGF,
heparin-binding epidermal growth factor; TGF-β, transforming growth
factor-β. |
Additional pregnancy-associated hormones (relaxin1
and 2, progesterone, and hCG), as well as other hormones
[endothelin-1, prostaglandins (PGE1, PGE2, PGI2, PGF2)], which were
upregulated after PAEP silencing in the NSCLC cell lines in
our previous study, were analysed with regard to their influence on
PAEP/glycodelin expression (20). Exclusively, hCG modulated
PAEP/glycodelin expression in the H1975 and 2106T cells
(data not shown). However, only high hCG concentrations led to an
increased PAEP/glycodelin expression, but also to a
degradation of β-actin. None of the other hormones influenced
glycodelin levels at the RNA or protein level in the investigated
NSCLC cell lines (data not shown). Due to these results, the tested
hormones were excluded from subsequent analyses.
The inducers that elevated PAEP/glycodelin
expression (Fig. 1A, black arrows)
were applied in subsequent microarray analyses to gain insight for
further pathway inducers or downstream pathways influencing
glycodelin expression. Therefore, an upstream regulator analysis
[software: IPA] of differentially expressed genes (<−1.5 or
>1.5) was performed (Fig. 1C).
As observed before, PMA, EGFR and LPA were confirmed as upstream
regulators. Moreover, several other possibly involved upstream
regulators appeared. For the following experiments, TGF-β1, NF-κB
complex, NF-κB1, PRKCD (isoform of PKC), rapidly accelerated
fibrosarcoma (RAF)1, ERK1/2, MEK and JUN were selected, since they
were determined to be involved in signalling pathways associated
with the regulation of glycodelin expression (Fig. 1C) (38-40)
or they were previously directly shown to influence glycodelin
amounts (29).
Since TGF-β is a further possible pathway inducer,
its effect on PAEP/glycodelin expression was consecutively
investigated. Other downstream signalling pathway components and
transcription factors were addressed in subsequent experiments.
TGF-β1, -2 and -3 led to elevated RNA and protein levels in the
2106T, but not in the H1975 cells (Fig. 1D and E). Subsequent experiments
focused on TGF-β1 and -2, since these two isoforms exhibited the
most potent effect.
Downstream signalling pathways involving
PKC, AKT and RAF/MEK are mediators in the regulation of
PAEP/glycodelin expression
There are distinct downstream signalling pathways
that can be activated by the former tested inducers. Therefore, we
investigated the influence of the PKC, PI3K/AKT and MEK/ERK
signalling cascade on PAEP/glycodelin expression by applying
different pathway modulators (Fig.
2A). Activating PKC with bryostatin1 led to an increased
glycodelin expression at the RNA and protein level in both cell
lines (Fig. 2B and C). The PKC
inhibitor GF109203X had a slight inducing or no effect at low
concentrations (1 and 5 µM), but markedly decreased the
PAEP/glycodelin amounts at the highest concentration (10
µM) (Fig. 2D and E). The
inhibition of AKT with MK-2206 (41) led to an increase in
PAEP/glycodelin expression in the H1975, but not in the
2106T cells (Fig. 2F and G). The
PAEP/glycodelin levels were negatively affected by the
RAF/MEK inhibitor, RO5126766 (42), in both cell lines (Fig. 2H and I).
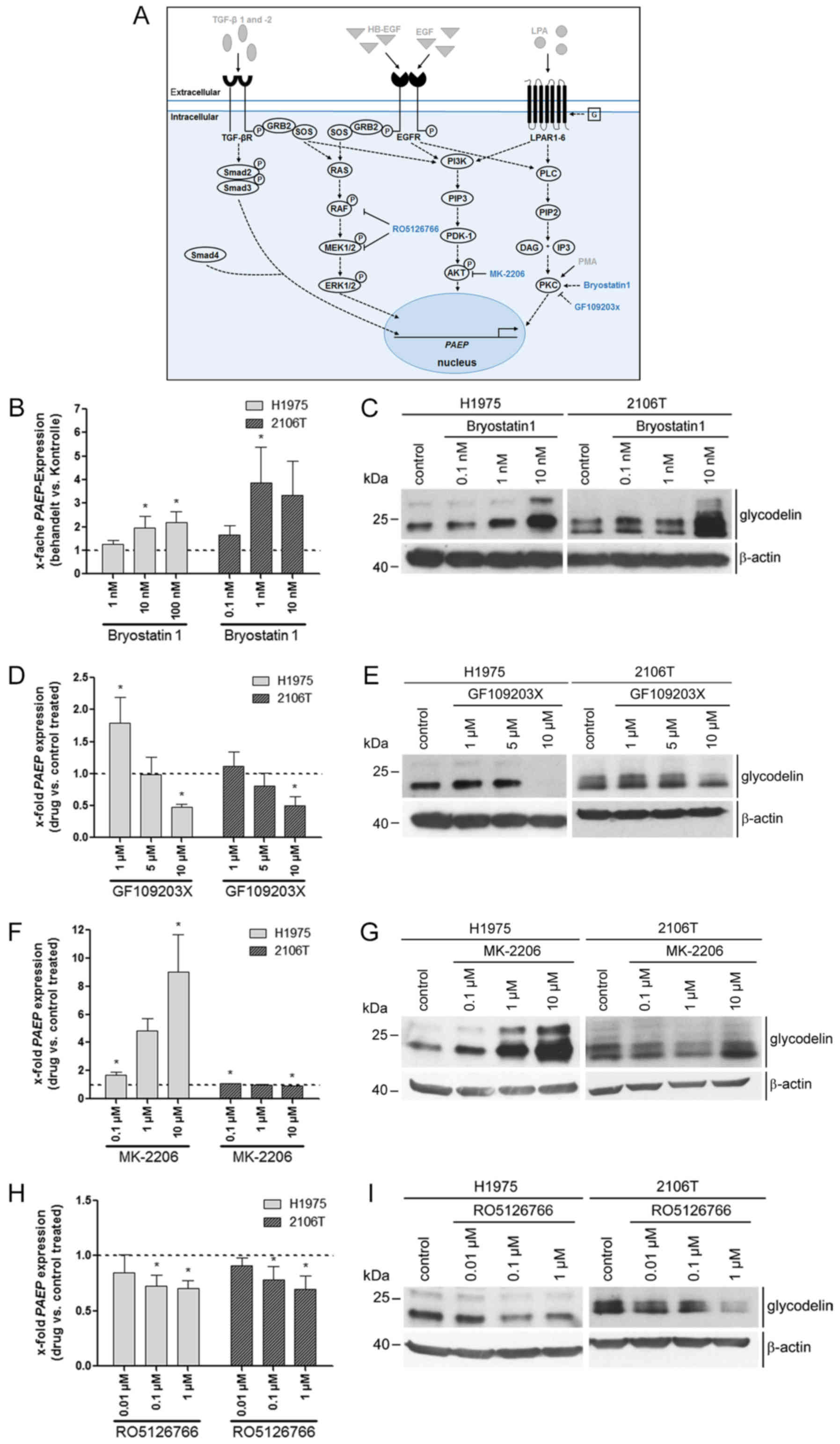 | Figure 2Downstream signalling pathways
including PKC, AKT and RAF/MEK affected PAEP/glycodelin
levels in NSCLC cells. (A) Overview of previously investigated
pathway inducers (grey with drawn down arrows) and components of
signalling pathways that were analysed in the next experiments
(blue). Dotted arrows depict assumed or literature data based
relationships between signalling pathway regulators. (B-I) Two
NSCLC cell lines were serum-starved overnight. The cells were then
cultivated with an activator of PKC (Bryostatin) for 1 h, as well
as with inhibitors of PKC (GF109203X), AKT (MK-2206) and RAF/MEK
(RO5126766) for 24 h. (B, D, F and H) qPCR visualizing PAEP
expression was performed from 3 independent experiments. Dotted
line at 1 represents the expression in control-treated cells (mean
of the Ct-values and mean SD are shown). Statistical
significance was defined as *P<0.05 (one-way ANOVA
and Dunnett's test; referring to control-treated cells). (C, E, G
and I) Western blot analysis detected the glycodelin levels.
β-actin was used as a loading control (cropped blots are shown).
PAEP, progesterone associated endometrial protein; NSCLC,
non-small cell lung cancer; PKC, protein kinase C. |
The levels of PAEP/glycodelin are
regulated by the transcription factors, JUNB, NF-κB1 and STAT3
Using bioinformatics analysis of the promotor region
of PAEP, we identified a set of transcription factors that
might bind before or within this region and can be activated by at
least one of the signalling pathways tested before. For these
reasons and based on the results of the microarray gene expression
profiling (Fig. 1C), we focused on
the involvement of the jun-B proto-oncogene (JUNB), NF-κB1 and
STAT3 in PAEP/glycodelin expression. The expression of the
transcription factors was silenced by the siRNA transfection of the
H1975 and 2106T cells (Fig. 3A).
All of the individual siRNAs and the pooled siRNAs decreased the
expression of the specific genes and proteins (Fig. 3B-G). In the case of NF-κB1, its
precursor (p105), as well as its active form (p50) were decreased.
The knockdown of the 3 genes led to a decreased PAEP
expression in both cell lines (Fig.
3B-F). Western blot analysis confirmed the RNA data at the
protein level (Fig. 3C, E and G).
Thus, JUNB, NF-κB1 and STAT3 silencing
diminished the glycodelin amounts in the H1975 and 2106T cells.
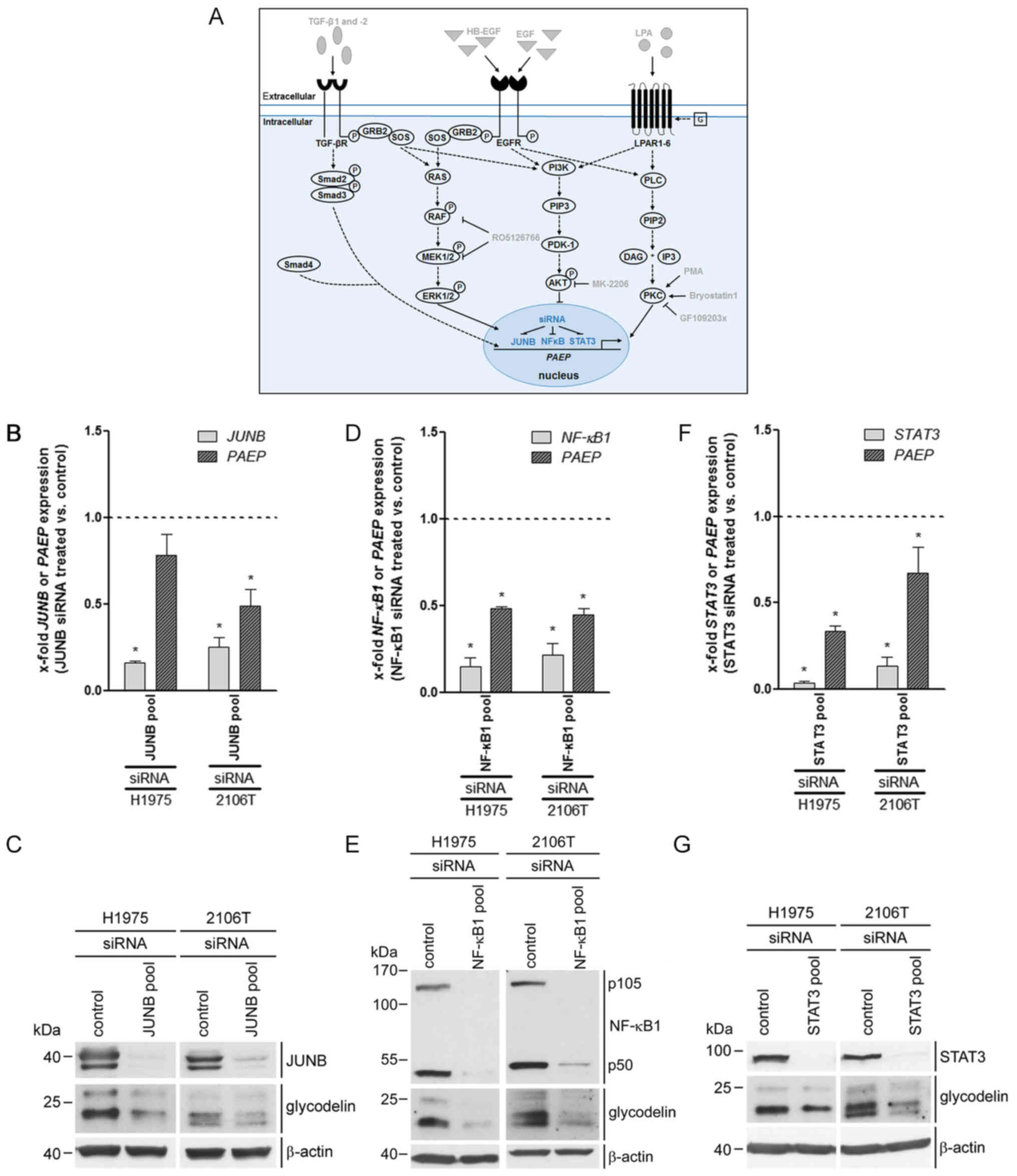 | Figure 3PAEP/glycodelin amounts in
NSCLC cells are influenced by the transcription factors JUNB,
NF-κB1 and STAT3. (A) Overview of previously applied pathway
inducers as well as signalling pathway modulators (grey) and
transcription factors, which were investigated in the next
experiments (blue). Dotted arrows illustrate activation of pathway
components known from the literature (2,27,28,38,43,44,58),
whereas drawn down lines show directly proven influences. (B-G)
H1975 and 2106T cells were transfected with a pool of 3 to 4
different siRNAs targeting JUNB, NF-κB1 and
STAT3 for 72 h. (B, D and F) qPCR analyses determined the
expression of the particular siRNA targeted gene (light grey) and
PAEP (dark grey), which were investigated in 3 independent
experiments. Dotted line at 1 represents the expression in
control-treated cells (mean of the Ct-values and mean SD
are shown). Statistical significance was defined as
*P<0.05 (unpaired t-test; referring to
control-treated cells). (C, E and G) Protein expression of JUNB,
NF-κB1, STAT3, after the silencing of their corresponding genes,
and glycodelin was examined by western blot analysis. β-actin was
used as a loading control (cropped blots are shown). PAEP,
progesterone associated endometrial protein; NSCLC, non-small
cell lung cancer. |
Associations between pathway inducers,
downstream pathways and transcription factors as regards their
effect on glycodelin expression
We further aimed to better understand the crosstalk
between the investigated pathway inducers, signalling pathways and
transcription factors. The H1975 and 2106T cells were treated with
the pathway inducers, LPA, PMA, EGF, HB-EGF, TGF-β1 and -2.
Subsequently, the activity of downstream pathways and transcription
factors was examined by western blot analysis. To detect the
stimulation of signalling pathways, the phosphorylation of only one
representative component was examined. In order to determine the
best point of time suitable for detecting the induction of
phosphorylation, we exemplarily examined substrates phosphorylated
by conventional PKCs at different time periods following treatment
with PMA. The strongest phosphorylation of PKC substrates was
observed between 30 min and 1 h following induction (data not
shown). Consequently, these treatment durations were applied in
further analysis.
PMA was the only pathway inducer that elevated the
expression of glycodelin in the H1975 cells. By contrast, the
glycodelin levels were increased due to each of the investigated
inducers in the 2106T cells (Fig. 4A
and B). LPA and PMA activated PKC in the H1975 cells, while the
other pathway inducers exhibited no effect on PKC activity. In the
2106T cells, PKC was activated by LPA, PMA and to a lower extent
also by EGF and HB-EGF treatment. TGF-β1 and -2 did not influence
PKC activity (Fig. 4A).
Fig. 4B provides an
overview of the activation patterns of the investigated signalling
molecules. Treatment with LPA, PMA, EGF and HB-EGF resulted in an
increased phosphorylation of ERK, JUNB and slightly also of STAT3
in the H1975 and 2106T cells. NF-κB p105 was mainly activated by
PMA in both cell lines. This indicated that LPA, EGF and HB-EGF
stimulated glycodelin expression inducing pathways in the H1975
cells without having an effect on glycodelin levels. As
demonstrated above, the inhibition of AKT led to an increase in
glycodelin expression in the H1975 cells (Fig. 2F and G). Therefore, we investigated
the influence of LPA, EGF and HB-EGF on AKT activation. This kinase
was strongly phosphorylated following treatment with LPA, whereas
EGF and HB-EGF caused only a weak AKT phosphorylation (Fig. 4C). Stimulation with TGF-β1 and -2
resulted in a marked activation of Smad2 in the 2106T cells,
followed by an upregulation of glycodelin in the 2106T (Fig. 4B), but not in the H1975 cells
(Fig. 1D and E).
Candidates regulating PAEP/glycodelin
expression in cell lines are also activated in NSCLC tissues with a
high PAEP expression
The overview shown in Fig. 5A shows the complex regulatory
mechanism of glycodelin expression in NSCLC cells suggested by the
results of this study. While in the lung SQCC cell line TGF-β
seemed to be a major inducer of PAEP/glycodelin expression,
the regulation in the tested lung ADC cell line was strongly
influenced by AKT. PKC played a major role in the stimulation of
PAEP/glycodelin expression in both cell lines. To validate
the observed influences on PAEP/glycodelin expression in
NSCLC patients, we performed an analysis using the qPCR-derived
patient data of our recent study (20). Patients with the highest or the
lowest PAEP expression in ADC (20 each) and SQCC (15 each) were
selected (Table I) and microarray
gene expression profiling was performed (Fig. 5B). For further analysis of
differentially expressed genes (<−1.5 or> 1.5), the software
IPA was used to identify upstream regulators. Of note, most of the
candidates that led to an elevated expression of PAEP/glycodelin in
the cell lines were also identified as upstream regulators in NSCLC
tissues with a high PAEP expression. According to the in
vitro results, the upstream regulators were more active in SQCC
than in ADC. Only the activity of TGF-β1 and STAT3 was similar in
both NSCLC subtypes (Fig. 5B).
 | Table IPatient and tumour
characteristics. |
Table I
Patient and tumour
characteristics.
| Parameter | Total (n=70) |
|---|
| Age, years | |
| Median
(range) | 65 (40-83) |
| Sex | |
| Male | 51 |
| Female | 19 |
| Histology | |
|
Adenocarcinoma | 40 |
| Squamous cell
carcinoma | 30 |
| Smoking status | |
| Smoker | 23 |
| Non-smoker <6
months | 15 |
| Non-smoker >6
months | 28 |
| Never-smoker | 4 |
| Therapy | |
| OP | 40 |
| OP/RT | 2 |
| OP/CT | 24 |
| OP/RT/CT | 4 |
| P stage | |
| IA | 8 |
| IB | 31 |
| IIA | 0 |
| IIB | 17 |
| IIIA | 14 |
| ECO | |
| 0 | 65 |
| 1 | 4 |
| 2 | 1 |
Discussion
Elucidating the regulation of PAEP/glycodelin
expression could provide insight for the usage of glycodelin as a
therapeutic target to weaken the immune system defence of NSCLC
tumours. After previously demonstrating the cellular functions of
glycodelin (20), in this study,
we analysed the thus far unknown underlying mechanisms regulating
the expression of PAEP/glycodelin in NSCLC cell lines and
NSCLC tissues. At least to the best of our knowledge, for the first
time, we identified involved pathway inducers, intracellular
signalling pathways and transcription factors in this tumour type.
In this context, a complex crosstalk of various signalling pathways
(Fig. 5A) and some differences
between the ADC cell line H1975 and the SQCC cell line 2106T were
observed.
In contrast to many NSCLC tumours, only few lung
carcinoma cell lines, such as H1975 and 2106T, were identified to
express glycodelin (20). Several
studies have already shown the inhibitory effects of glycodelin on
the apoptosis, proliferation and the activity of immune cells
(reviewed in 24). Together with the assumption that NSCLC tumours
secrete glycodelin to circumvent immunosurveillance, this suggests
a major role of the tumour microenvironment. Lacking this
environment in vitro, the majority of the NSCLC cell lines
might stop PAEP/glycodelin expression. Gottschling et
al observed similarities and various differences in the gene
expression patterns of two SQCC cell lines and their corresponding
tissue. In particular, gene clusters associated with immune
response, adhesion, proliferation, differentiation and angiogenesis
were strongly silenced in the cell lines and therefore differed
from their particular tissue. They also assumed the
microenvironment as a reason for this observation (36).
In the H1975 cells, PMA was the only pathway inducer
that stimulated glycodelin expression. By contrast, this effect was
observed after LPA, PMA, EGF, HB-EGF, TGF-β1, -2 and -3 treatment
in the 2106T cells. LPA and PMA have already been postulated to
increase PAEP/glycodelin expression in different cell lines
(25,26). As regards EGF, HB-EGF and TGF-β,
there are no published data available describing an influence of
these inducers on PAEP/glycodelin expression, at least to
the best of our knowledge. In this study, we demonstrated that all
of the inducers activated the PKC and/or the MEK/ERK pathway
downstream, with the exception of TGF-β. In accordance with our
findings, a previous study also postulated an association of MEK
and the expression of glycodelin in other cells (29). The inhibition of both pathways led
to a decreased PAEP/glycodelin expression in the NSCLC cell
lines. Therefore, we hypothesised that stimulating these pathways
with the different pathway inducers should increase
PAEP/glycodelin levels. However, this hypothesis could be
completely confirmed only in the 2106T cells, whereas the
PAEP/glycodelin levels were altered exclusively after PMA
treatment in the H1975 cells. The discrepancy may be explained by
an LPA-, EGF- and HB-EGF-mediated activation of the PI3K/AKT
pathway, which was shown to suppress PAEP/glycodelin
expression in the H1975 cells. This suggests an antagonizing effect
to the PKC- and MEK/ERK pathway-dependent stimulation of
PAEP/glycodelin expression. In this regard, former literature data
have demonstrated a LPA-, EGF- and HB-EGF-dependent PI3K/AKT
pathway stimulation (43,44). Contrary to the results observed
with the H1975 cells, treatment of the 2106T cells with an AKT
inhibitor did not affect PAEP/glycodelin expression.
Currently, the reason for these distinct results is unclear;
however, activating EGFR (T790M, L858R) and PIK3CA
(G118D) mutations in H1975 cells may play a role. Each of these
three mutations has been previously shown to be associated with the
activation of AKT (45-49). This suggests a generally stronger
AKT activity accompanied by a lower glycodelin expression in H1975
cells.
Depending on the particular cell line, treatment
with LPA, PMA, EGF and HB-EGF did not only activate the PKC and
MEK/ERK pathway, but also the transcription factors, JUNB and
STAT3. Therefore, it could be hypothesised that both transcription
factors are involved in these signalling cascades, as already
described in previous studies (39,50-52).
NF-κB1, which has been shown to be associated with both the PKC and
MEK/ERK pathways in the literature (40,53),
was mainly affected by the PMA-stimulated PKC signalling cascade in
our analyses. TGF-β1 and -2, as well as LPA induced the
phosphorylation of Smad2 in the 2106T cells, which is also in
concordance with previous literature data (54,55).
As none of the examined other pathways or transcription factors was
affected by TGF-β, it can be concluded that glycodelin expression
is solely influenced by the canonical TGF-β pathway.
The in vitro derived data were confirmed in
NSCLC patient tissues to a great extent. Most of the analysed
PAEP/glycodelin expression-regulating candidates in the cell
lines were also found as upstream regulators in the patient
tissues. In accordance with the cell line-derived data, a stronger
activation of the upstream regulators was determined in SQCC
compared to ADC, apart from TGF-β1 and STAT3. Analogous to the
results observed with the cell lines, we expected a decreased AKT
activity in ADC tumours with a high PAEP expression.
However, the activation of AKT was slightly increased in these
tumour tissues. As already mentioned, we assume that only an
activated PI3K/AKT pathway, as it was shown for the mutations T790M
(EGFR), L858R (EGFR) and G118D (PIK3CA)
(45-49) of H1975 cells, influences
PAEP/glycodelin expression. The mutational status of the
patients is not known, as molecular alterations are not tested in
operated patients. Probably, an activated PI3K/AKT pathway occurs
only in a few patients. In a previous study activated PI3K/AKT
pathways were shown in 25% of the analysed lung ADCs (56). This might be the reason for the
differences between the in vitro and the in vivo
data.
The pathways that were shown to play a role in the
regulation of PAEP/glycodelin expression are known to be
altered in cancer. Actually, drugs for the treatment of NSCLC
patients are available targeting the MEK/ERK pathway, which
stimulates the PAEP/glycodelin expression. Since 2017, a
combined therapy of a BRAF inhibitor (Dabrafenib) and MEK inhibitor
(Trametinib) is approved for NSCLC patients with a BRAF-V600
mutation. Our results provide evidence that this BRAF/MEK inhibitor
treatment may possibly downregulate glycodelin expression in tumour
cells and hence decrease the postulated immunosuppressive effect.
AKT inhibitors are currently in clinical development for cancer
therapy (57). According to our
results, the treatment of patients with mutational activated AKT
might increase the expression of glycodelin and therefore could
promote immunosurveillance mediated by cancer cells.
As the tumour microenvironment is assumed to play a
major role in the regulation of PAEP/glycodelin expression,
future experiments should aim to address this association.
Therefore, we aim to perform co-culture experiments of NSCLC lines
with fibroblasts and immune cells in order to analyse the effect on
PAEP/glycodelin expression. Our group has already postulated
an upregulation of immune system regulating ligands, such as
programmed cell death 1 ligand 1 (PD-L1), in PAEP knockdown
NSCLC cell lines (20). Therefore,
further investigations are required to reason glycodelin as a
target of NSCLC treatments.
In conclusion, in this study, for the first time (at
least to the best of our knowledge), we elucidated regulators of
PAEP/glycodelin expression in NSCLC. Our data point towards
a putative model, in which glycodelin expression is stimulated by
the canonical TGF-β pathway in SQCC cells and the PKC signalling
cascade in both NSCLC cell lines. The PI3K/AKT pathway inhibits
glycodelin expression in ADC cells and an antagonizing role towards
the other investigated signalling cascades is suggested herein.
TGF-β, the PI3K/AKT signalling cascade, as well as the MEK/ERK
pathway, the latter stimulated glycodelin expression, are targets
of NSCLC drugs that are already approved or currently under
investigations. Our results provide evidence that inhibiting these
targets affects the expression of glycodelin and therefore, exerts
an immunosuppressive effect on NSCLC tumours. Furthermore,
understanding the regulation of glycodelin expression may lead to
the development of novel therapeutic approaches to weaken the
immune system defence of NSCLC tumours.
Funding
This study was financially supported in part by the
Federal Ministry of Education and Research (BMBF, Germany) and the
German Centre for Lung Research (DZL, Germany).
Availability of data and materials
The microarray datasets generated and analysed
during the current study are available in the NCBI GEO database
(GSE115458; https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE115458).
Authors' contributions
RW was involved in manuscript writing and the
performance of the experiments. RW, MM, TM, HS and MAS were
involved in the conception and design of the study. RW, MM, TM, MT,
AW, HW, FJFH and MAS were involved in the acquisition of material
and data. RW, MM, TM, MT, HS, AW, HW, FJFH and MAS were involved in
data analysis and interpretation. All authors have read and
approved the manuscript.
Ethics approval and consent to
participate
The study was approved by the local Ethics Committee
of the University of Heidelberg (no. 270/2001).
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Acknowledgments
The authors would like to thank Ms. Elizabeth Chang
Xu, Mr. Martin Fallenbuechel, Ms. Carmen Hoppstock, Ms. Christa
Stolp and Ms. Andrea Bopp for their expert technical assistance.
Furthermore, the authors wish to thank Dr. Dmytro Dvornikov
(Systems Biology of Signal Transduction, DKFZ), who kindly provided
the pSmad2 and Smad2/3 antibodies. The tissue samples were provided
by the Lung Biobank Heidelberg, a member of the accredited Tissue
Bank of the National Centre for Tumour Diseases (NCT) Heidelberg,
the BioMaterialBank Heidelberg and the German Centre for Lung
Research (DZL) in accordance with the regulations of the tissue
bank and the approval of the ethics committee of the University of
Heidelberg.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
2
|
da Cunha Santos G, Shepherd FA and Tsao
MS: EGFR mutations and lung cancer. Annu Rev Pathol. 6:49–69. 2011.
View Article : Google Scholar
|
|
3
|
Morris HR, Dell A, Easton RL, Panico M,
Koistinen H, Koistinen R, Oehninger S, Patankar MS, Seppala M and
Clark GF: Gender-specific glycosylation of human glycodelin affects
its contraceptive activity. J Biol Chem. 271:32159–32167. 1996.
View Article : Google Scholar
|
|
4
|
Dell A, Morris HR, Easton RL, Panico M,
Patankar M, Oehniger S, Koistinen R, Koistinen H, Seppala M and
Clark GF: Structural analysis of the oligosaccharides derived from
glycodelin, a human glycoprotein with potent immunosuppressive and
contraceptive activities. J Biol Chem. 270:24116–24126. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chiu PC, Chung MK, Koistinen R, Koistinen
H, Seppala M, Ho PC, Ng EH, Lee KF and Yeung WS: Cumulus
oophorus-associated glycodelin-C displaces sperm-bound glycodelin-A
and -F and stimulates spermatozoa-zona pellucida binding. J Biol
Chem. 282:5378–5388. 2007. View Article : Google Scholar
|
|
6
|
Seppälä M, Koistinen H, Koistinen R, Chiu
PC and Yeung WS: Glycosylation related actions of glycodelin:
Gamete, cumulus cell, immune cell and clinical associations. Hum
Reprod Update. 13:275–287. 2007. View Article : Google Scholar
|
|
7
|
Okamoto N, Uchida A, Takakura K, Kariya Y,
Kanzaki H, Riittinen L, Koistinen R, Seppälä M and Mori T:
Suppression by human placental protein 14 of natural killer cell
activity. Am J Reprod Immunol. 26:137–142. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Laird SM, Hill CJ, Warren MA, Tuckerman EM
and Li TC: The production of placental protein 14 by human uterine
tubal epithelial cells in culture. Hum Reprod. 10:1346–1351. 1995.
View Article : Google Scholar
|
|
9
|
Fazleabas AT, Donnelly KM, Srinivasan S,
Fortman JD and Miller JB: Modulation of the baboon (Papio anubis)
uterine endometrium by chorionic gonadotrophin during the period of
uterine receptivity. Proc Natl Acad Sci USA. 96:2543–2548. 1999.
View Article : Google Scholar
|
|
10
|
Toth B, Roth K, Kunert-Keil C, Scholz C,
Schulze S, Mylonas I, Friese K and Jeschke U: Glycodelin protein
and mRNA is downregulated in human first trimester abortion and
partially upregulated in mole pregnancy. J Histochem Cytochem.
56:477–485. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tseng L, Zhu HH, Mazella J, Koistinen H
and Seppälä M: Relaxin stimulates glycodelin mRNA and protein
concentrations in human endometrial glandular epithelial cells. Mol
Hum Reprod. 5:372–375. 1999. View Article : Google Scholar
|
|
12
|
Kämäräinen M, Halttunen M, Koistinen R,
von Boguslawsky K, von Smitten K, Andersson LC and Seppälä M:
Expression of glycodelin in human breast and breast cancer. Int J
Cancer. 83:738–742. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hackenberg R, Loos S, Nia AH, Kunzmann R
and Schulz KD: Expression of placental protein 14 by the new
endometrial cancer cell line MFE-280 in vitro and by endometrial
carcinomas in vivo. Anticancer Res. 18(2A): 1153–1158.
1998.PubMed/NCBI
|
|
14
|
Kämäräinen M, Leivo I, Koistinen R,
Julkunen M, Karvonen U, Rutanen EM and Seppälä M: Normal human
ovary and ovarian tumors express glycodelin, a glycoprotein with
immunosuppressive and contraceptive properties. Am J Pathol.
148:1435–1443. 1996.
|
|
15
|
Connor JP, Brudney A, Ferrer K and
Fazleabas AT: Glycodelin-A expression in the uterine cervix.
Gynecol Oncol. 79:216–219. 2000. View Article : Google Scholar
|
|
16
|
Kämäräinen M, Miettinen M, Seppala M, von
Boguslawsky K, Benassi MS, Böhling T and Andersson LC: Epithelial
expression of glycodelin in biphasic synovial sarcomas. Int J
Cancer. 76:487–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren S, Liu S, Howell PM Jr, Zhang G,
Pannell L, Samant R, Shevde-Samant L, Tucker JA, Fodstad O and
Riker AI: Functional characterization of the progestagen-associated
endometrial protein gene in human melanoma. J Cell Mol Med. 14(6B):
1432–1442. 2010. View Article : Google Scholar
|
|
18
|
Schneider MA, Muley T, Kahn NC, Warth A,
Thomas M, Herth FJ, Dienemann H and Meister M: Glycodelin is a
potential novel follow-up biomarker for malignant pleural
mesothelioma. Oncotarget. 7:71285–71297. 2016. View Article : Google Scholar
|
|
19
|
Kunert-Keil C, Steinmüller F, Jeschke U,
Gredes T and Gedrange T: Immunolocalization of glycodelin in human
adenocarcinoma of the lung, squamous cell carcinoma of the lung and
lung metastases of colonic adenocarcinoma. Acta Histochem.
113:798–802. 2011. View Article : Google Scholar
|
|
20
|
Schneider MA, Granzow M, Warth A, Schnabel
PA, Thomas M, Herth FJ, Dienemann H, Muley T and Meister M:
Glycodelin: A New Biomarker with Immunomodulatory Functions in
Non-Small Cell Lung Cancer. Clin Cancer Res. 21:3529–3540. 2015.
View Article : Google Scholar
|
|
21
|
Lenhard M, Heublein S, Kunert-Keil C,
Vrekoussis T, Lomba I, Ditsch N, Mayr D, Friese K and Jeschke U:
Immunosuppressive Glycodelin A is an independent marker for poor
prognosis in endometrial cancer. BMC Cancer. 13:6162013. View Article : Google Scholar
|
|
22
|
Scholz C, Heublein S, Lenhard M, Friese K,
Mayr D and Jeschke U: Glycodelin A is a prognostic marker to
predict poor outcome in advanced stage ovarian cancer patients. BMC
Res Notes. 5:5512012. View Article : Google Scholar
|
|
23
|
Mandelin E, Lassus H, Seppälä M, Leminen
A, Gustafsson JA, Cheng G, Bützow R and Koistinen R: Glycodelin in
ovarian serous carcinoma: Association with differentiation and
survival. Cancer Res. 63:6258–6264. 2003.
|
|
24
|
Cui J, Liu Y and Wang X: The Roles of
Glycodelin in Cancer Development and Progression. Front Immunol.
8:16852017. View Article : Google Scholar
|
|
25
|
Morrow DM, Xiong N, Getty RR, Ratajczak
MZ, Morgan D, Seppala M, Riittinen L, Gewirtz AM and Tykocinski ML:
Hematopoietic placental protein 14. An immunosuppressive factor in
cells of the megakaryocytic lineage. Am J Pathol. 145:1485–1495.
1994.
|
|
26
|
Ramachandran S, Ramaswamy S, Cho Ch and
Parthasarathy S: Lysophosphatidic acid induces glycodelin gene
expression in cancer cells. Cancer Lett. 177:197–202. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bazzi MD and Nelsestuen GL: Differences in
the effects of phorbol esters and diacylglycerols on protein kinase
C. Biochemistry. 28:9317–9323. 1989. View Article : Google Scholar
|
|
28
|
Sando JJ and Chertihin OI: Activation of
protein kinase C by lysophosphatidic acid: Dependence on
composition of phospholipid vesicles. Biochem J. 317:583–588. 1996.
View Article : Google Scholar
|
|
29
|
Xu JW, Yasui N, Ikeda K, Pan WJ, Watanabe
J, Shiotani M, Yanaihara A, Miki T and Yamori Y: Isoflavones
regulate secretion of leukemia inhibitory factor and transforming
growth factor {beta} and expression of glycodelin in human
endometrial epithelial cells. J Endocrinol. 196:425–433. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dydensborg AB, Rose AA, Wilson BJ, Grote
D, Paquet M, Giguère V, Siegel PM and Bouchard M: GATA3 inhibits
breast cancer growth and pulmonary breast cancer metastasis.
Oncogene. 28:2634–2642. 2009. View Article : Google Scholar
|
|
31
|
Ren S, Howell PM Jr, Han Y, Wang J, Liu M,
Wang Y, Quan G, Du W, Fang L and Riker AI: Overexpression of the
progestagen-associated endometrial protein gene is associated with
microphthalmia-associated transcription factor in human melanoma.
Ochsner J. 11:212–219. 2011.
|
|
32
|
Gao J, Mazella J, Seppala M and Tseng L:
Ligand activated hPR modulates the glycodelin promoter activity
through the Sp1 sites in human endometrial adenocarcinoma cells.
Mol Cell Endocrinol. 176:97–102. 2001. View Article : Google Scholar
|
|
33
|
Tabbaa ZM, Zheng Y and Daftary GS: KLF11
epigenetically regulates glycodelin-A, a marker of endometrial
biology via histone-modifying chromatin mechanisms. Reprod Sci.
21:319–328. 2014. View Article : Google Scholar
|
|
34
|
Beasley MB, Brambilla E and Travis WD: The
2004 World Health Organization classification of lung tumors. Semin
Roentgenol. 40:90–97. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wittekind C: 2010 TNM system: On the 7th
edition of TNM classification of malignant tumors. Pathologe.
31:331–332. 2010.In German. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gottschling S, Jauch A, Kuner R, Herpel E,
Mueller-Decker K, Schnabel PA, Xu EC, Muley T, Sültmann H, Bender
C, et al: Establishment and comparative characterization of novel
squamous cell non-small cell lung cancer cell lines and their
corresponding tumor tissue. Lung Cancer. 75:45–57. 2012. View Article : Google Scholar
|
|
37
|
Bustin SA, Benes V, Garson JA, Hellemans
J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL,
et al: The MIQE guidelines: Minimum information for publication of
quantitative real-time PCR experiments. Clin Chem. 55:611–622.
2009. View Article : Google Scholar
|
|
38
|
Zhang YE: Non-Smad pathways in TGF-beta
signaling. Cell Res. 19:128–139. 2009. View Article : Google Scholar
|
|
39
|
Hodge C, Liao J, Stofega M, Guan K,
Carter-Su C and Schwartz J: Growth hormone stimulates
phosphorylation and activation of elk-1 and expression of c-fos,
egr-1, and junB through activation of extracellular
signal-regulated kinases 1-2. J Biol Chem. 273:31327–31336. 1998.
View Article : Google Scholar
|
|
40
|
Dhawan P and Richmond A: A novel NF-kappa
B-inducing kinase-MAPK signaling pathway up-regulates NF-kappa B
activity in melanoma cells. J Biol Chem. 277:7920–7928. 2002.
View Article : Google Scholar
|
|
41
|
Holland WS, Chinn DC, Lara PN Jr, Gandara
DR and Mack PC: Effects of AKT inhibition on HGF-mediated erlotinib
resistance in non-small cell lung cancer cell lines. J Cancer Res
Clin Oncol. 141:615–626. 2015. View Article : Google Scholar :
|
|
42
|
Wada M, Horinaka M, Yamazaki T, Katoh N
and Sakai T: The dual RAF/MEK inhibitor CH5126766/RO5126766 may be
a potential therapy for RAS-mutated tumor cells. PLoS One.
9:e1132172014. View Article : Google Scholar
|
|
43
|
Jorissen RN, Walker F, Pouliot N, Garrett
TP, Ward CW and Burgess AW: Epidermal growth factor receptor:
Mechanisms of activation and signalling. Exp Cell Res. 284:31–53.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Riaz A, Huang Y and Johansson S:
G-Protein-Coupled Lysophosphatidic Acid Receptors and Their
Regulation of AKT Signaling. Int J Mol Sci. 17:2152016. View Article : Google Scholar
|
|
45
|
Wee P and Wang Z: Epidermal Growth Factor
Receptor Cell Proliferation Signaling Pathways. Cancers (Basel).
9:92017.
|
|
46
|
Sordella R, Bell DW, Haber DA and
Settleman J: Gefitinib-sensitizing EGFR mutations in lung cancer
activate anti-apoptotic pathways. Science. 305:1163–1167. 2004.
View Article : Google Scholar
|
|
47
|
Tracy S, Mukohara T, Hansen M, Meyerson M,
Johnson BE and Jänne PA: Gefitinib induces apoptosis in the
EGFRL858R non-small-cell lung cancer cell line H3255. Cancer Res.
64:7241–7244. 2004. View Article : Google Scholar
|
|
48
|
Burke JE, Perisic O, Masson GR, Vadas O
and Williams RL: Oncogenic mutations mimic and enhance dynamic
events in the natural activation of phosphoinositide 3-kinase p110α
(PIK3CA). Proc Natl Acad Sci USA. 109:15259–15264. 2012. View Article : Google Scholar
|
|
49
|
Orloff MS, He X, Peterson C, Chen F, Chen
JL, Mester JL and Eng C: Germline PIK3CA and AKT1 mutations in
Cowden and Cowden-like syndromes. Am J Hum Genet. 92:76–80. 2013.
View Article : Google Scholar :
|
|
50
|
de Groot RP, Auwerx J, Karperien M, Staels
B and Kruijer W: Activation of junB by PKC and PKA signal
transduction through a novel cis-acting element. Nucleic Acids Res.
19:775–781. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jain N, Zhang T, Kee WH, Li W and Cao X:
Protein kinase C delta associates with and phosphorylates Stat3 in
an interleukin-6-dependent manner. J Biol Chem. 274:24392–24400.
1999. View Article : Google Scholar
|
|
52
|
Levy DE and Lee CK: What does Stat3 do. J
Clin Invest. 109:1143–1148. 2002. View Article : Google Scholar
|
|
53
|
Ghosh S and Baltimore D: Activation in
vitro of NF-kappa B by phosphorylation of its inhibitor I kappa B.
Nature. 344:678–682. 1990. View Article : Google Scholar
|
|
54
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar
|
|
55
|
Jeon ES, Moon HJ, Lee MJ, Song HY, Kim YM,
Cho M, Suh DS, Yoon MS, Chang CL, Jung JS, et al: Cancer-derived
lysophosphatidic acid stimulates differentiation of human
mesenchymal stem cells to myofibroblast-like cells. Stem Cells.
26:789–797. 2008. View Article : Google Scholar
|
|
56
|
Collisson EA, Campbell JD, Brooks AN,
Berger AH, Lee W, Chmielecki J, Beer DG, Cope L, Creighton CJ,
Danilova L, et al: Cancer Genome Atlas Research Network:
Comprehensive molecular profiling of lung adenocarcinoma. Nature.
511:543–550. 2014. View Article : Google Scholar
|
|
57
|
Brown JS and Banerji U: Maximising the
potential of AKT inhibitors as anti-cancer treatments. Pharmacol
Ther. 172:101–115. 2017. View Article : Google Scholar
|
|
58
|
Gazdar AF: Epidermal growth factor
receptor inhibition in lung cancer: The evolving role of
individualized therapy. Cancer Metastasis Rev. 29:37–48. 2010.
View Article : Google Scholar
|