Introduction
Colorectal cancer is one of the leading causes of
cancer-associated morbidity and mortality worldwide. Despite recent
advances in surgery and chemoradiotherapy for colorectal cancer,
the prognosis of patients with advanced colorectal cancer remains
poor (1-3). Therefore, a better understanding of
the molecular biology of colorectal cancer progression will provide
clinically applicable biomarkers for the reliable prediction of
cancer progression and the identification of novel therapeutic
targets (3).
Forkhead box A1 (FOXA1) is a transcription factor of
the forkhead box gene superfamily necessary for the binding and
activity of other transcriptional factors to chromatin, and serves
critical roles in the development and differentiation of epithelial
cells in several human organs (4,5).
Studies have shown that the expression of FOXA1 is associated with
the development and progression of various types of cancer
(6-9) and its functions may change according
to specific types of cancer (6-9).
FOXA1 acts as a tumor suppressor in various types of human cancer,
including cancer of the breast, endometrium, bladder, liver and
pancreas (10-17). However, FOXA1 induces aggressive
behavior in lung, esophageal, prostate and thyroid cancer,
implicating an oncogenic role (18-22).
Further molecular mechanistic studies have revealed that FOXA1
promotes tumor progression by recruiting other transcription
factors, and acts as a transcription factor in suppressing tumor
development by directly regulating target gene expression (6-9).
FOXA1 is also known to control the specificity of cancer cell types
due to of the existence of unique FOXA1 targeting in each cancer
cell type (9). However, the
biological functions, underlying mechanisms and clinical
significance of FOXA1 in human colorectal cancer remain to be fully
elucidated. Previously, only one study has reported the oncogenic
role of FOXA1 in human colorectal cancer (23).
The present study aimed to evaluate whether FOXA1
affects the oncogenic biological behavior of human colorectal
cancer cells, to assess the expression of FOXA1 in human colorectal
cancer tissues and to examine its association with
clinicopathological features, including survival rate.
Materials and methods
Cell culture and materials
Human colorectal carcinoma cell lines (HCT116,
CCL-247™; Caco2, HTB-37™; - SW480, CCL-228™; HT29, HTB-38™; DLD1,
CCL-221™; and COLO205, CCL-222™) were obtained from the American
Type Culture Collection (Manassas, VA, USA). The DKO1 (CVCL-9798)
cell line was purchased from the ExPASy (Cellosaurus, Lausanne,
Switzerland). All cell lines, excluding HCT116 cells, were cultured
in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco, Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin, whereas the HCT116 cells were cultured in
McCoy’s 5a Medium (WelGENE, Inc., Daegu, Korea) supplemented with
10% FBS and 1% penicillin/streptomycin at 37°C in a humidified
atmosphere with 5% CO2.
Gene transfection
FOXA1 cDNA was subcloned into the pcDNA6-myc vector
(Invitrogen, Thermo Fisher Scientific, Inc.). The FOXA1
construction was verified by sequencing. FOXA1 small interfering
(si)RNA (GGAGGAGAGAUAAG UUAUA-dTdT) and scrambled siRNA (AllStars
Negative Control siRNA, cat. no. 1027281) were purchased from
Bioneer (Daejeon, Korea) and Qiagen GmbH (Hilden, Germany),
respectively. To transfect siRNA, the SW480 and HCT116 cells were
seeded into 6-well plates at a density of 3×105
cells/well at 37°C and were 40-60% confluent at the time of
transfection. For FOXA1 knockdown, 20 µM of FOXA1 siRNA (FS)
was transfected with 5 µl Lipofectamine™ RNAiMAX reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). To overexpress FOXA1,
0.5 µg of FOXO1-pcDNA6-myc vector (FV) was transfected with
5 µl Lipofectamine™ 2000 reagent (Invitrogen, Thermo Fisher
Scientific, Inc.). Scrambled siRNA (SS) and empty-pcDNA6-myc vector
(EV) were used as a negative control, respectively. Following
incubation for 24 h at 37°C, identification of the expression of
FOXA1 was performed by western blotting and the transfected cells
were used in the following experiments.
Cell proliferation assay
The transfected HCT116 and DLD1 cells were seeded
into a 96-well plate at a density of 1×104 cells/well
and incubated for 24 h at 37°C. Cell viability was determined using
a water-soluble tetrazolium salt (DoGen, Daeillab, Seoul, Korea).
Following application, the absorbance at 450 nm was measured on a
microplate reader (Infinite M200; Tecan, Mannedorf, Switzerland).
Each experiment was performed in triplicate wells and was repeated
at least three times.
Apoptosis analysis
The DLD1 and HCT116 cells at a density of
5×105/well were seeded into a 6-well plate and incubated
for 24 h at 37°C prior to FOXA1 transfection. The transfected cells
were collected and resuspended in 100 µl of binding buffer
(BD Biosciences, San Diego, CA, USA) for 20 min at 4°C. The cells
were incubated with 7-amino-actinomycin D (BD Biosciences) and
Annexin V-APC (BD Biosciences) for 20 min at room temperature. To
analyze the number of apoptotic cells, a FACSCalibur flow cytometer
(BD Biosciences) and WinMDI version 2.9 (The Scripps Research
Institute, San Diego, CA, USA) were used.
Cell cycle analysis
The DLD1 and HCT116 cells at a density of
5×105/well were seeded into a 6-well plate and
transfected with FS and FV for 24 h at 37°C. The transfected cells
were fixed in ice-cold 70% ethanol for 1 h to determine the cell
cycle distribution and then washed twice with PBS. The cells were
incubated in 100 µl of 10 µg/ml ribonuclease A
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 20 min at 37°C.
Following this, 100 µl of 50 µg/ml propidium iodide
(Sigma-Aldrich; Merck KGaA) was added for 30 min at room
temperature in the dark. Cell cycle analysis was performed using a
FACSCalibur flow cytometer (BD Biosciences) and WinMDI version 2.9
(The Scripps Research Institute).
Western blotting
The transfected cells were lysed in RIPA buffer
containing Halt™ Protease inhibitor and Halt™ Phosphatase inhibitor
cocktail (Thermo Fisher Scientific, Inc.) for 30 min in an ice
bath. The protein concentration of the lysate from transfected
cells was measured using the BCA™ protein assay (Thermo Fisher
Scientific, Inc.). The proteins (20 µg per lane) were
separated using 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto a polyvinylidene fluoride
(PVDF) membrane (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The transferred membranes was blocked with 5% bovine serum albumin
(BSA) for 1 h at room temperature and then incubated overnight at
4°C with primary antibodies at 1:1,000 dilution. Antibodies against
the following proteins were used: FOXA1 (cat. no. ab170933, Abcam,
Cambridge, UK), cleaved poly (ADP-ribose) polymerase (PARP, cat.
no. 5625), p53 upregulated modulator of apoptosis (PUMA; cat. no.
4976), Bax (cat. no. 2772), BH3 interacting domain death agonist
(Bid; cat. no. 2002), Bcl-xL (cat. no. 2764), cyclin D1 (cat. no.
2926), p21 (cat. no. 2947), cyclin-dependent kinase 2 (CDK2, cat.
no. 2546), CDK4 (cat. no. 2906), myeloid cell leukemia-1 (Mcl-1;
cat. no. 5453), phos pho-signal transducer and activator of tran
scription-3 (phospho-STAT3, cat. no. 9145), STAT3 (cat. no. 9139)
from Cell Signaling Technology, Inc. (Danvers, MA, USA), and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH, cat. no. FL-335)
from Santa Cruz Biotechnology, Inc.. The membranes were washed four
times with Tris-buffered saline-0.1% Tween-20 (TBS-T) and were
incubated with a horseradish peroxidase-conjugated secondary
antibody (anti-rabbit, cat. no. 7074, anti-mouse, cat. no. 7076,
Cell Signaling, Technology, Inc.) at 1:2,000 dilution for 1 h at
room temperature. Following washing with TBS-T, the protein bands
were developed using an Enhanced Chemiluminescent reagent
(Amersham, GE Healthcare Life Sciences) and were analyzed on the
luminescence image analyzer LAS-4000 (Fujifilm, Tokyo, Japan). The
bands of the immunoblot were quantified using Multi-Gauge software
(ver. 3.0, Fujifilm).
Transwell invasion assay
The transfected cells were plated in the upper well
of Transwell filter chambers (8.0-µm pore size; Costar,
Cambridge, MA, USA) coated with 1% gelatin. Fibronectin (10
µg/ml, Sigma-Aldrich; Merck KGaA) was added as a
chemoattractant to 0.2% BSA (Sigma-Aldrich; Merck KGaA) containing
medium in the lower chamber. Following overnight incubation at
37°C, the cells invaded to the lower surface of the upper chamber
were fixed with 70% ethanol for 1 min at room temperature and
stained with Hemacolor® Rapid staining solution (Merck
KGaA) following the manufacturer’s protocol. Under a light
microscope (magnification, ×200, Olympus BX51, Olympus, Tokyo,
Japan), the stained upper chambers were placed on the corner
squares of a hemocytometer and images were captured. Subsequently,
the stained cells were counted in five selected fields (each 0.25
mm2, four smaller squares). The results are expressed as
the mean ± SE of the number of cells/field of three individual
experiments.
Patients and tissue samples
Formalin-fixed, paraffin-embedded tissue blocks were
obtained from 403 patients who underwent surgery for colorectal
cancer at Chonnam National University Hwasun Hospital (Jeonnam,
Korea) between July 2004 and June 2006. The tissue blocks were
selected by viewing original pathologic slides and selecting blocks
that showed the junction between normal colorectal epithelium and
tumor tissue. Patient characteristics, including sex, age at the
time of surgery, histologic grade and stage, were obtained by
examining medical records and contacting pathologists and
physicians when necessary. No patient had received anticancer
therapy prior to surgery. The tumors were staged in accordance with
the American Joint Committee on Cancer staging system (24). Overall survival was measured from
the time of surgery until follow-up on December 31, 2013. The
present study was approved by the Institutional Review Board of the
Chonnam National University Hwasun Hospital (Jeonnam, Korea;
CNUHH-2017-163). This was a retrospective study and written
informed consent was obtained from each participant prior to tissue
acquisition at the time of hospitalization. All participants
provided written consent for their information to be stored in the
hospital database and used for research.
Immunohistochemistry and evaluation of
the expression of FOXA1
The paraffin-embedded tissue samples
(4-µm-thick tissue sections) were deparaffinized using
xylene and rehydrated with ethanol at graded concentrations
(100-60%). The samples were then boiled in citrate buffer (pH 6.0,
Dako, Agilent Technologies, Inc., Carpentaria, CA, USA) for 10 min
in a pressure boiler for antigen retrieval. Endogenous peroxidase
activity was blocked using peroxidase-blocking solution (Dako,
Agilent Technologies, Inc.) for 10 min at room temperature, and
non-specific reactivity was blocked with Dako® Protein
Block Serum-Free solution (Dako, Agilent Technologies, Inc.) for 30
min at room temperature. The samples were incubated with primary
anti-FOXA1 (1:200 dilution; cat. no. ab170933, Abcam) antibodies
for 1 h at room temperature. Bound antibody was visualized with the
DakoReal™ Envision HRP/DAB detection system (Dako, Agilent
Technologies, Inc.). Nuclear counterstaining was performed with
Mayer’s hematoxylin solution (Sigma-Aldrich, Merck KGaA). The
stained tissues were observed and images were captured under a
light microscope (Olympus BX51). Immunohistochemical staining was
assessed by two independent pathologists blinded to the patient
clinical outcome data. FOXA1 staining intensity was scored as
follows: 0, no staining; 1, weak; 2, moderate; 3, strong. The
percentage of the stained area was scored as 0 for the absence of
positive staining in tumor cells, 1 for positive staining in
<10% of the tumor cells, 2 for positive staining in 10-50% of
the tumor cells, and 3 for positive staining in >50% of the
tumor cells. The final score index was obtained by multiplying the
intensity and area percentage scores. The final scores for the 403
tumor samples ranged between 0.0 and 12.0 with a mean of 6.0.
Samples with a total score of ≥6 were designated as positive for
FOXA1 expression, whereas those with a total score of <6 were
designated as negative for FOXA1 expression.
Assessment of apoptosis and tumor cell
proliferation
Tumor cell apoptosis was determined using the
terminal deoxynucleotidyl transferase-mediated dUTP nick-end
labeling (TUNEL) system (Promega, Madison, MA, USA) according to
the manufacturer’s instructions. Briefly, the tissues were
deparaffinized, rehydrated through a graded alcohol series
(100-60%) and incubated in permeabilization solution. Labeling was
performed by adding the terminal deoxynucleotide transferase enzyme
reaction mix to tissue sections mounted on slides. Following
washing with TBS-T, the slides were incubated with the enzyme
substrate 3,3-diaminobenzidine (DAB) for color development, which
was used to localize the labeled cells. The apoptotic index (AI)
was calculated as the number of TUNEL-positive cells per 1,000
tumor cell nuclei. Proliferation of the tumor cells was visualized
by immunohistochemical staining with Ki-67 (1:500 dilution, cat.
no. ab833, Abcam) antibody. The samples were incubated with primary
anti-ki-67 diluent for 1 h at room temperature. Bound antibody was
visualized with the DakoReal™ Envision HRP/DAB detection system
(Dako, Agilent Technologies, Inc.). Nuclear counterstaining was
performed with Mayer’s hematoxylin solution (Sigma-Aldrich, Merck
KGaA). The stained tissues were observed and images were captured
under a light microscope (Olympus BX51). Nuclei stained with Ki-67
antibody were considered positive. The Ki-67 labeling index (KI)
was defined as the number of Ki-67-positive nuclei per 1,000 tumor
cell nuclei.
Statistical analysis
Statistical analyses were conducted using the
Statistical Package for Social Sciences software version 20.0 (IBM
Corp., Armonk, NY, USA). A χ2 test was used to analyze
the association between the expression of FOXA1 and
clinicopathological parameters. The survival rates were calculated
using the Kaplan-Meier method, and the statistical significance of
differences was examined using the log-rank test. The Student’s
t-test was used for the analysis of association between the
expression of FOXA1 and the apoptosis and proliferation of cells in
human colorectal cancer. Experimental differences between the FOXA1
knockdown or overexpression group and control group were tested
with the Student’s t-test. Each experiment was repeated at least
three times. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression of FOXA1 in various human
colorectal cancer cell lines
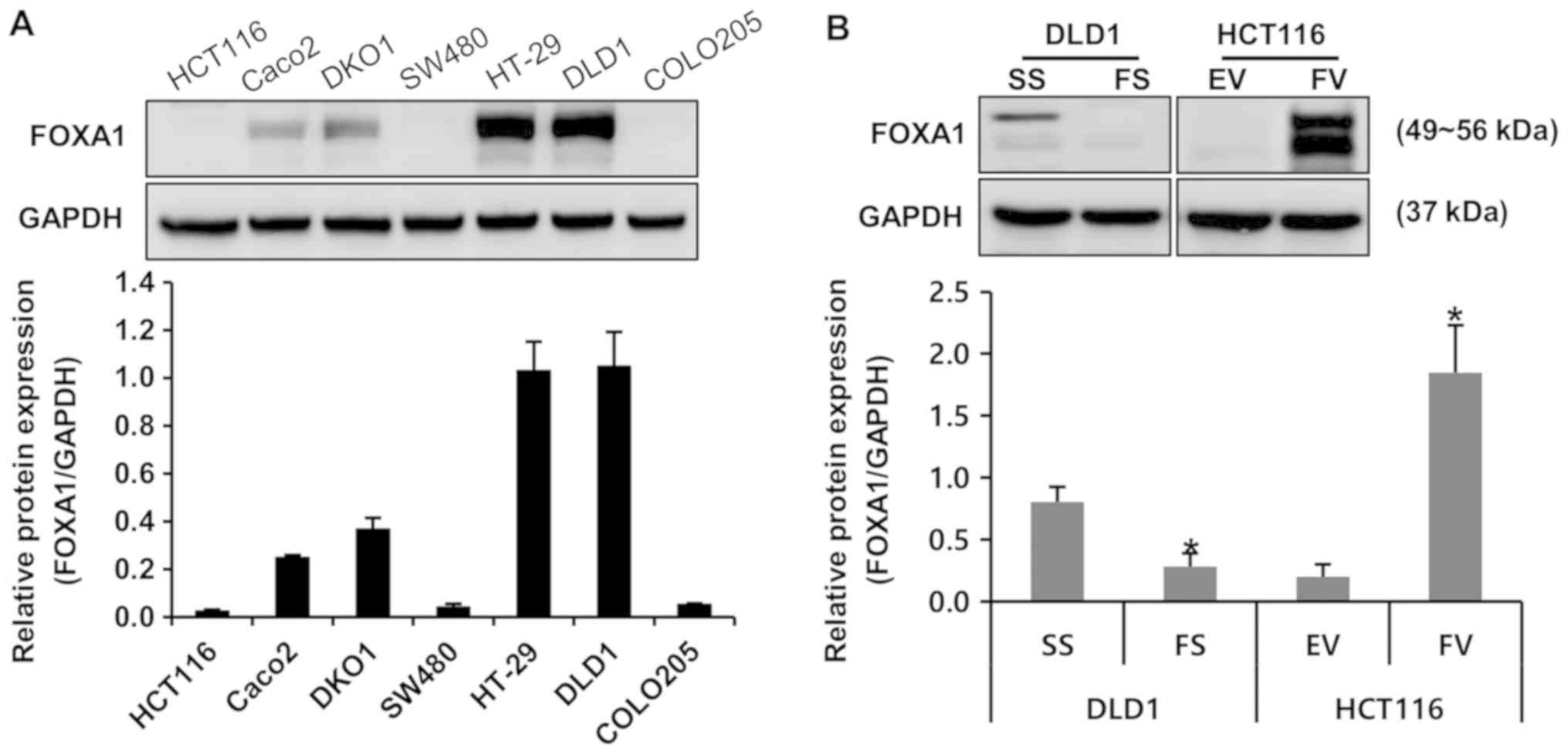
To investigate the protein expression of FOXA1 in
colorectal cancer cells, various human colorectal cancer cell
lines, including HCT116, DLD1, DKO1, SW480, Caco2, COLO205 and
HT-29, were subjected to western blot analysis. Among these cells,
the protein expression of FOXA1 was the highest in DLD1 cells and
the lowest in HCT116 cells (Fig.
1A). FS or FV were used to modulate the endogenous protein
expression of FOXA1 in DLD1 and HCT116 cells, respectively. The
protein expression of FOXA1 was specifically decreased at the
protein level by transfection of FS in DLD1 cells and was increased
by the transfection of FV in HCT116 cells (Fig. 1B).
Impact of FOXA1 on the invasion of human
colorectal cancer cells
The number of invading FS-transfected DLD1 cells was
significantly decreased, compared with those in the SS-transfected
cells (P<0.05). By contrast, the FV-transfected HCT116 cells
exhibited a significantly increased number of invading cells
compared with the EV-transfected cells (P<0.05) (Fig. 2).
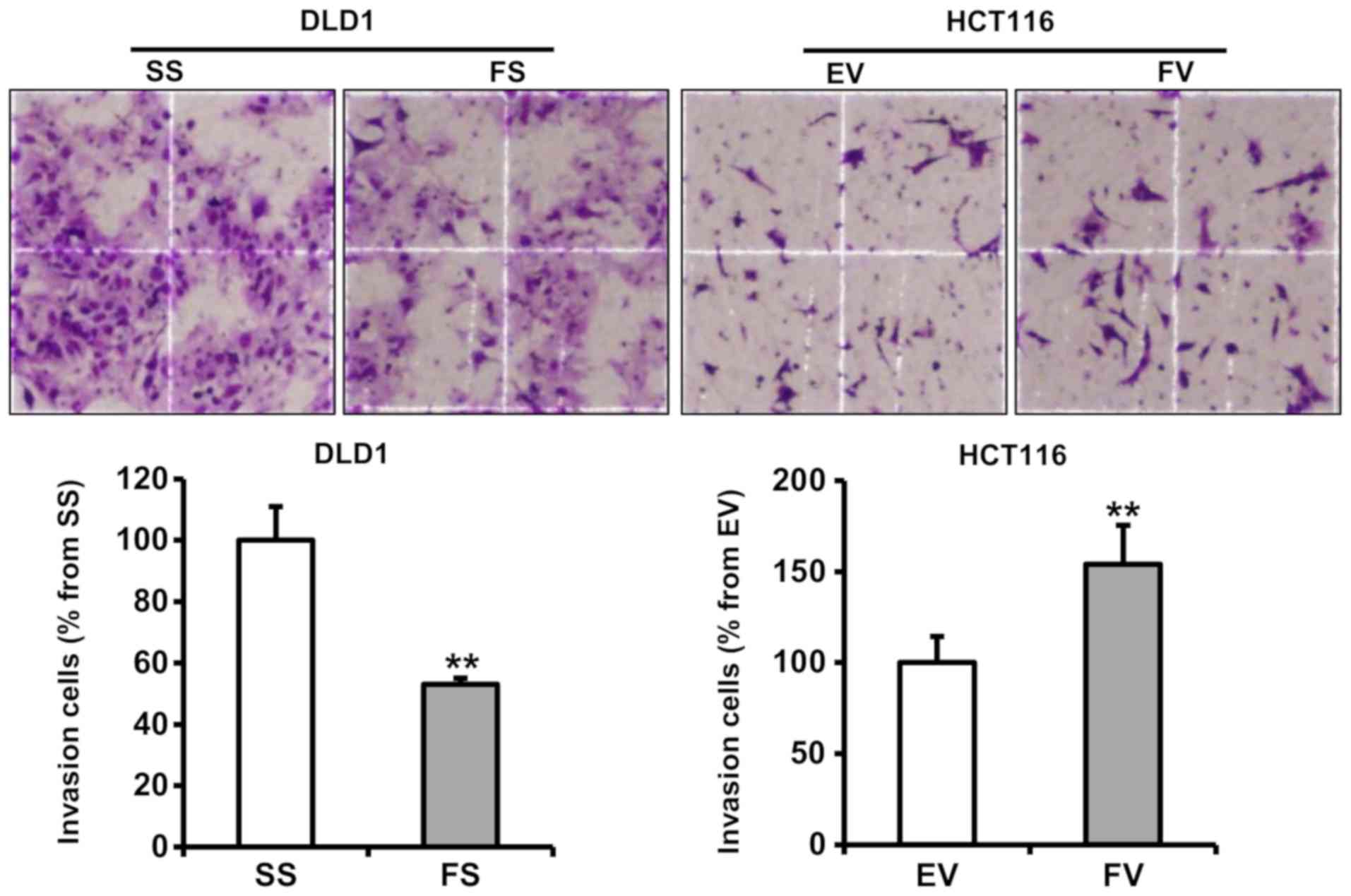 | Figure 2Impact of FOXA1 on the invasion of
human colorectal cancer cells. An invasion assay using cells
transfected with pcDNA6-myc or siRNA was performed. Stained
invading cells were counted under a light microscope
(magnification, ×200, four smaller squares = 0.25 mm2) and are
represented as a graph between groups. The number of FS-transfected
cells that invaded was significantly lower than that of invading
SS-transfected cells (mean ± SE, n=6; **P<0.01, compared to
respective control). FOXA1, forkhead box protein A1; siRNA, small
interfering RNA; SS, scrambled siRNA; FS, FOXA1 siRNA; EV,
empty-pcDNA6-myc vector; FV, pcDNA6-myc-FOXA1 vector. |
Impact of FOXA1 on human colorectal
cancer cell proliferation
To determine the potential effects of FOXA1 on cell
proliferation, cells were subjected to a cell proliferation assay 2
days after transfection with FS or FV. Proliferating cells, as
determined by absorbance, were decreased significantly in the
FS-transfected cells, compared with the SS-transfected cells in
DLD1 cells (P<0.05). By contrast, the FV-transfected HCT116
cells exhibited significantly increased proliferation, compared
with the EV-transfected cells (P<0.05) (Fig. 3).
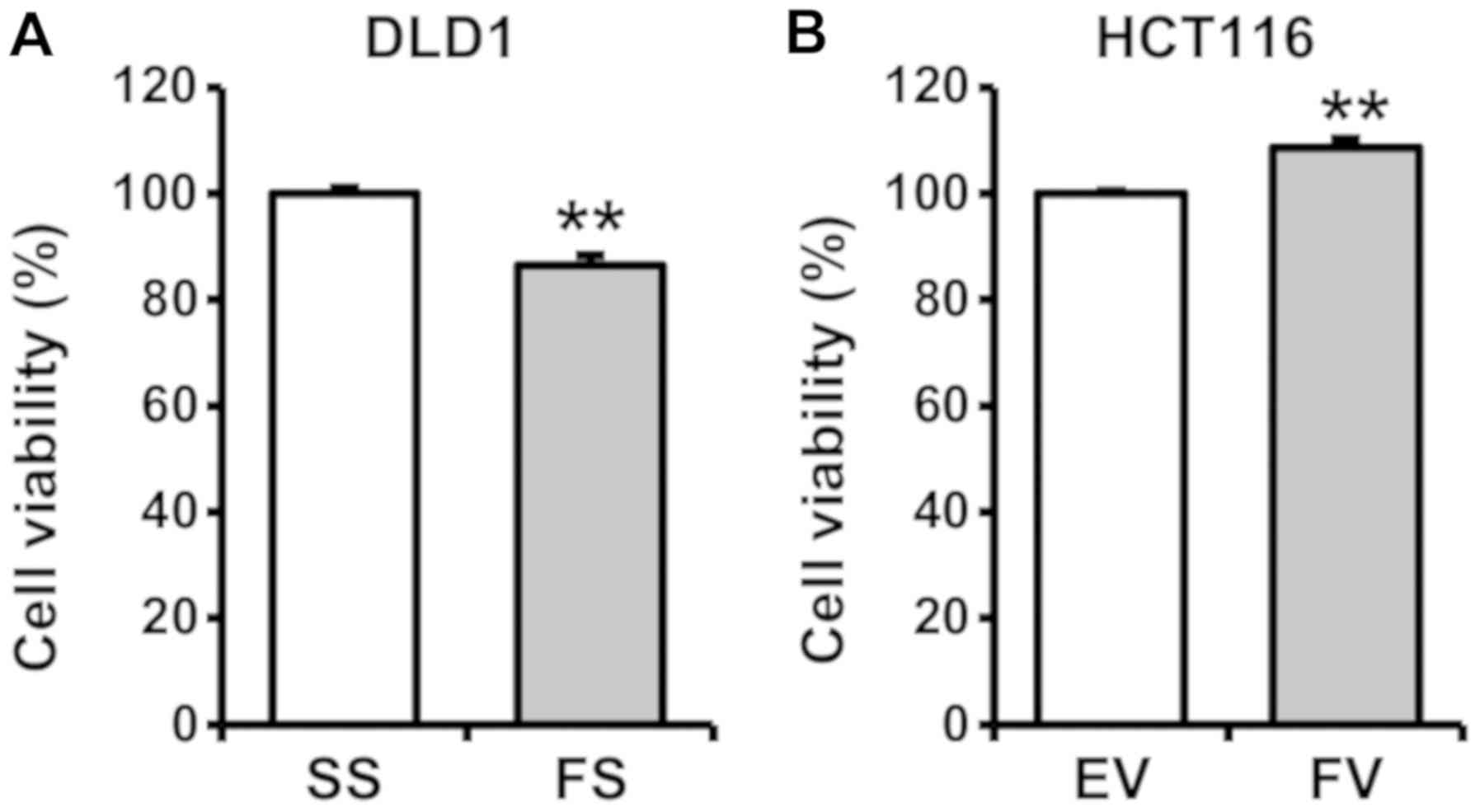 | Figure 3Impact of FOXA1 on the proliferation
of human colorectal cancer cells. To determine the potential
effects of FOXA1 on cell proliferation, cell viability was measured
using the water-soluble tetrazolium salt assay. A cell
proliferation assay was performed 2 days after transfection with
pcDNA6-myc or siRNA. (A) Proliferating cells, as determined by
absorbance, decreased significantly in the FS-transfected cells,
compared with those in the SS-transfected DLD1 cells
(**P<0.05). (B) FV-transfected HCT116 cells exhibited
significantly increased proliferation compared with the
EV-transfected cells (**P<0.05). FOXA1, fork-head box
protein A1; siRNA, small interfering RNA; SS, scrambled siRNA; FS,
FOXA1 siRNA; EV, empty-pcDNA6-myc vector; FV, pcDNA6-myc-FOXA1
vector. |
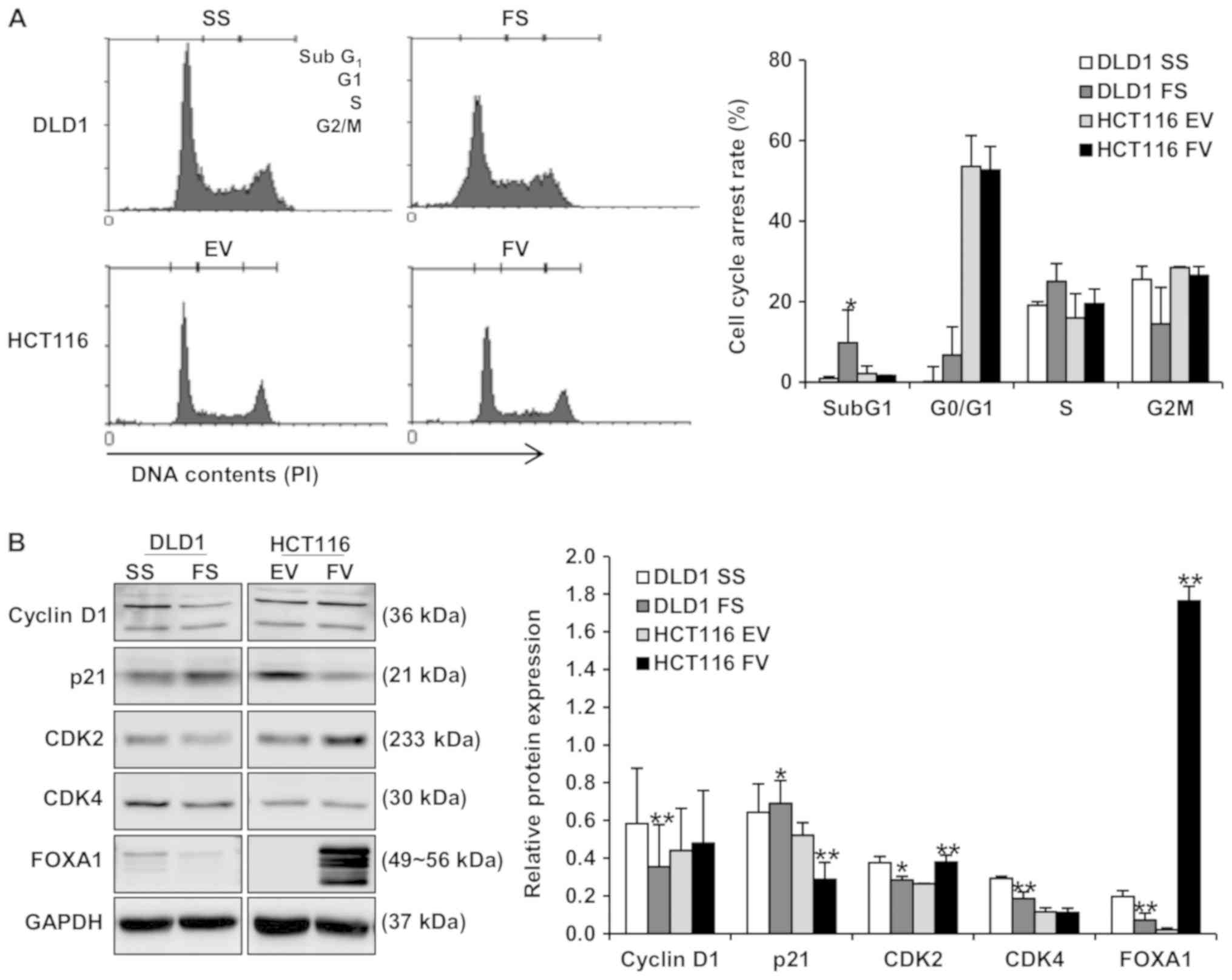
Impact of FOXA1 on apoptosis and cell
cycle distribution in human colorectal cancer cells
Flow cytometric analyses were performed to evaluate
the impact of FOXA1 on apoptosis and cell cycle distribution. The
apoptotic rate of cells transfected with FS was significantly
increased, compared with those transfected with SS in DLD1 cells
(8.65±4.9, vs. 12.69±5.0%; P<0.01). In addition, the apoptotic
rate was decreased in HCT116 cells following the overexpression of
FOXA1 (15.14±2.3, vs. 13.89±0.2%; Fig.
4A). To determine the activation of caspases during the
knockdown and overexpression of FOXA1, caspase-specific activities
were detected. The expression of cleaved PARP was increased in DLD1
cells following the knockdown of FOXA1, and was decreased in HCT116
cells following the overexpression of FOXA1 (Fig. 4B). Whether the impact of FOXA1 on
apoptosis is associated with the modulation of apoptosis regulatory
proteins was further examined. As shown in Fig. 4B, FOXA1 knockdown led to an
increase in the pro-apoptotic protein, PUMA. By contrast, the
overexpression of FOXA1 led to a decrease in PUMA and an increase
in anti-apoptotic proteins Bid and Mcl-1. The effects of FOXA1 on
cell cycle distribution and the regulators involved in cell cycle
distribution were evaluated in human colorectal cancer cells. The
overexpression of FOXA1 inhibited cell cycle arrest in the subG1
phase of HCT116 cells, and its knockdown induced cell cycle arrest
in DLD1 cells (Fig. 5A). As shown
in Fig. 5B, positive regulators of
the cell cycle, including cyclin D1, CDK2 and CDK4, exhibited
significantly decreased protein levels, whereas the negative
regulator of CDKI-p21 was significantly increased by FOXA1
knockdown in DLD1 cells. The protein levels of cyclin D1 and CDK2
were significantly increased and the protein level of p21 was
significantly decreased by the overexpression of FOXA1 in HCT116
cells (Fig. 5B).
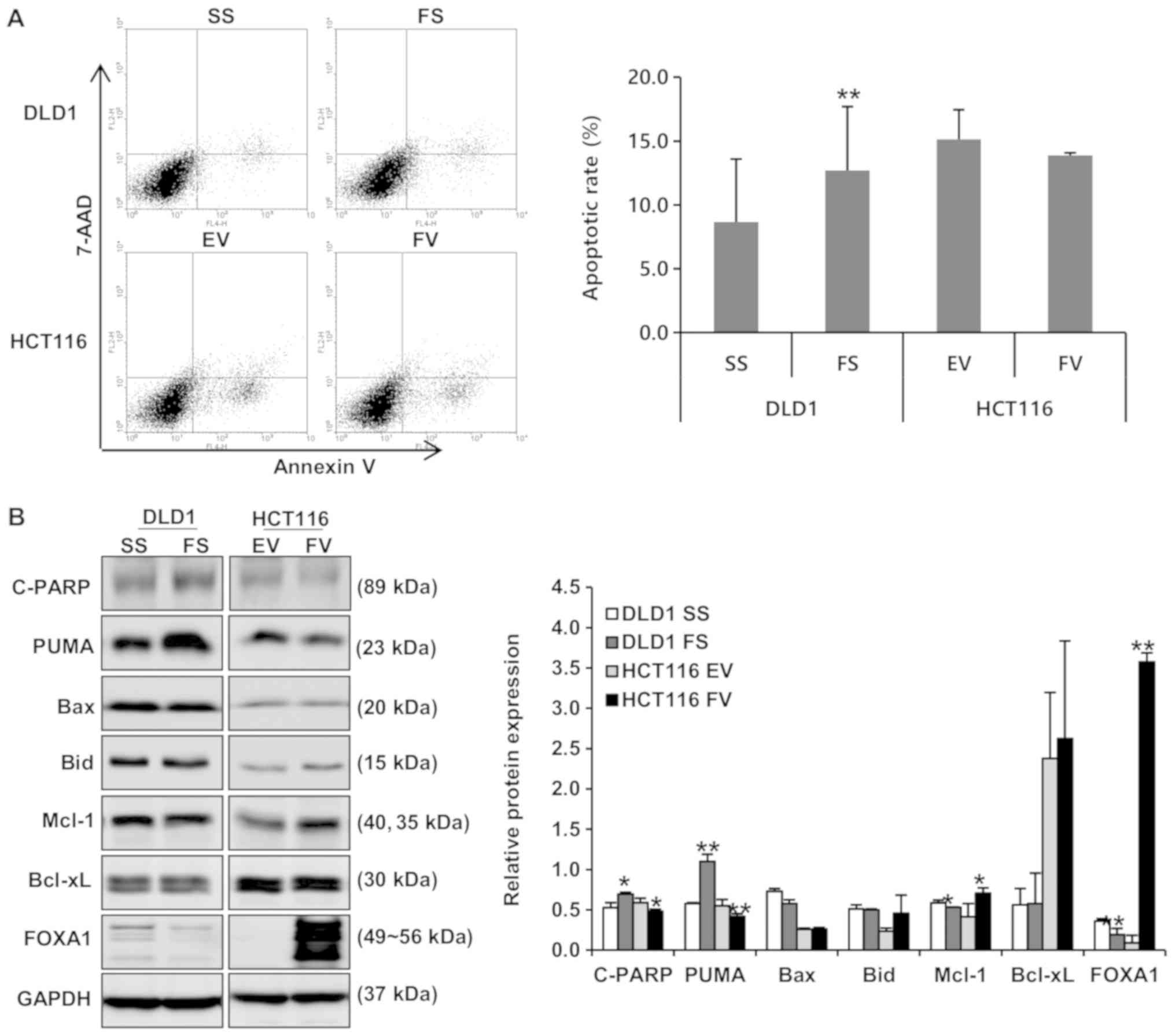 | Figure 4Impact of FOXA1 on apoptosis in human
colorectal cancer cells. Flow cytometric analyses and western
blotting were performed to evaluate the impact of FOXA1 on
apoptosis. (A) Proportion of apoptotic cells was decreased in
FV-transfected cells and was increased in FS-transfected cells
(**P<0.01). (B) Expression levels of cleaved PARP and
PUMA were decreased by the overexpression of FOXA1 and increased by
FOXA1 knockdown. Expression levels of Bid and Mcl-1 were increased
by the overexpression of FOXA1 (*P<0.05 and
**P<0.01, compared to respective control). Bands of
the immunoblot were quantified using Multi-Gauge software (ver.
3.0). FOXA1, forkhead box protein A1; siRNA, small interfering RNA;
SS, scrambled siRNA; FS, FOXA1 siRNA; EV, empty-pcDNA6-myc vector;
FV, pcDNA6-myc-FOXA1 vector, C-PARP; cleaved poly (ADP-ribose)
polymerase, PUMA; p53-up-regulated modulator of apoptosis; Bid, BH3
interacting domain death agonist; Mcl-1; myeloid cell
leukemia-1. |
Impact of FOXA1 on the oncogenic
signaling pathway in human colorectal cancer cells
To examine whether FOXA1 activates oncogenic
signaling pathways in human colorectal cancer cells, the present
study determined the phosphorylation level of STAT3 signaling
protein using western blotting. It was found that the
phosphorylation of STAT3 was decreased by FOXA1 knockdown in DLD1
cells. By contrast, the phosphorylation of STAT3 was increased by
the overexpression of FOXA1 in HCT116 cells (Fig. 6).
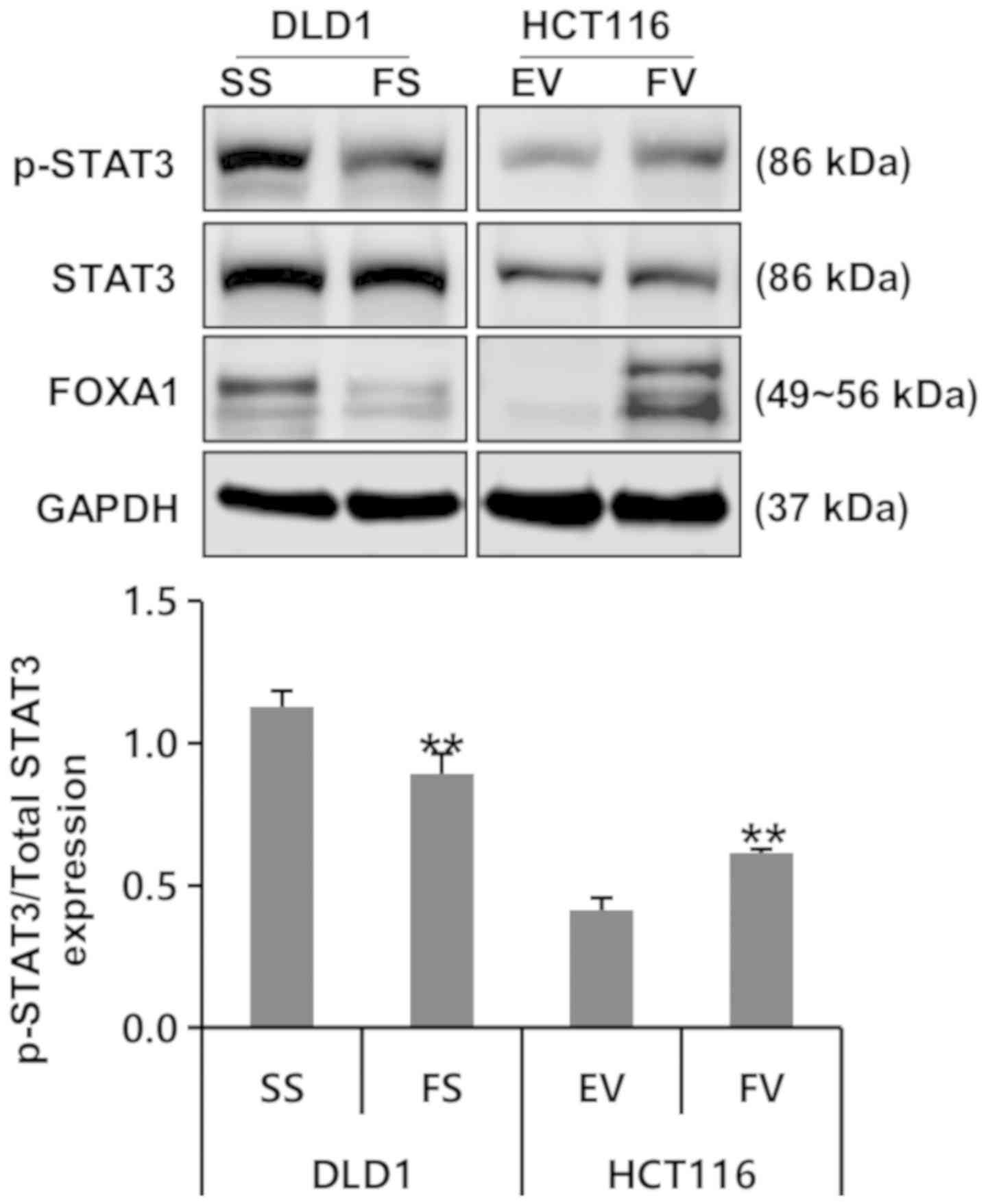 | Figure 6Impact of FOXA1 on oncogenic
signaling pathways in human colorectal cancer cells.
Phosphorylation of STAT3 was decreased by FOXA1 knockdown. By
contrast, phosphorylation of STAT3 was increased by the
overexpression of FOXA1 (**P<0.01, compared to
respective control). Bands of the immunoblot were quantified using
Multi-Gauge software (ver. 3.0). FOXA1, forkhead box protein A1;
siRNA, small interfering RNA; SS, scrambled siRNA; FS, FOXA1 siRNA;
EV, empty-pcDNA6–myc vector; FV, pcDNA6-myc-FOXA1 vector, STAT3;
signal transducers and activators of transcription 3; p-,
phosphorylated. |
Association between FOXA1 and
clinicopathological parameters of human colorectal cancer
To examine the prognostic role of FOXA1 in the
progression of human colorectal cancer, the present study
investigated the expression of FOXA1 immunohistochemically in
formalin-fixed, paraffin-embedded tissue sections obtained from 403
patients with colorectal cancer, and results were associated with
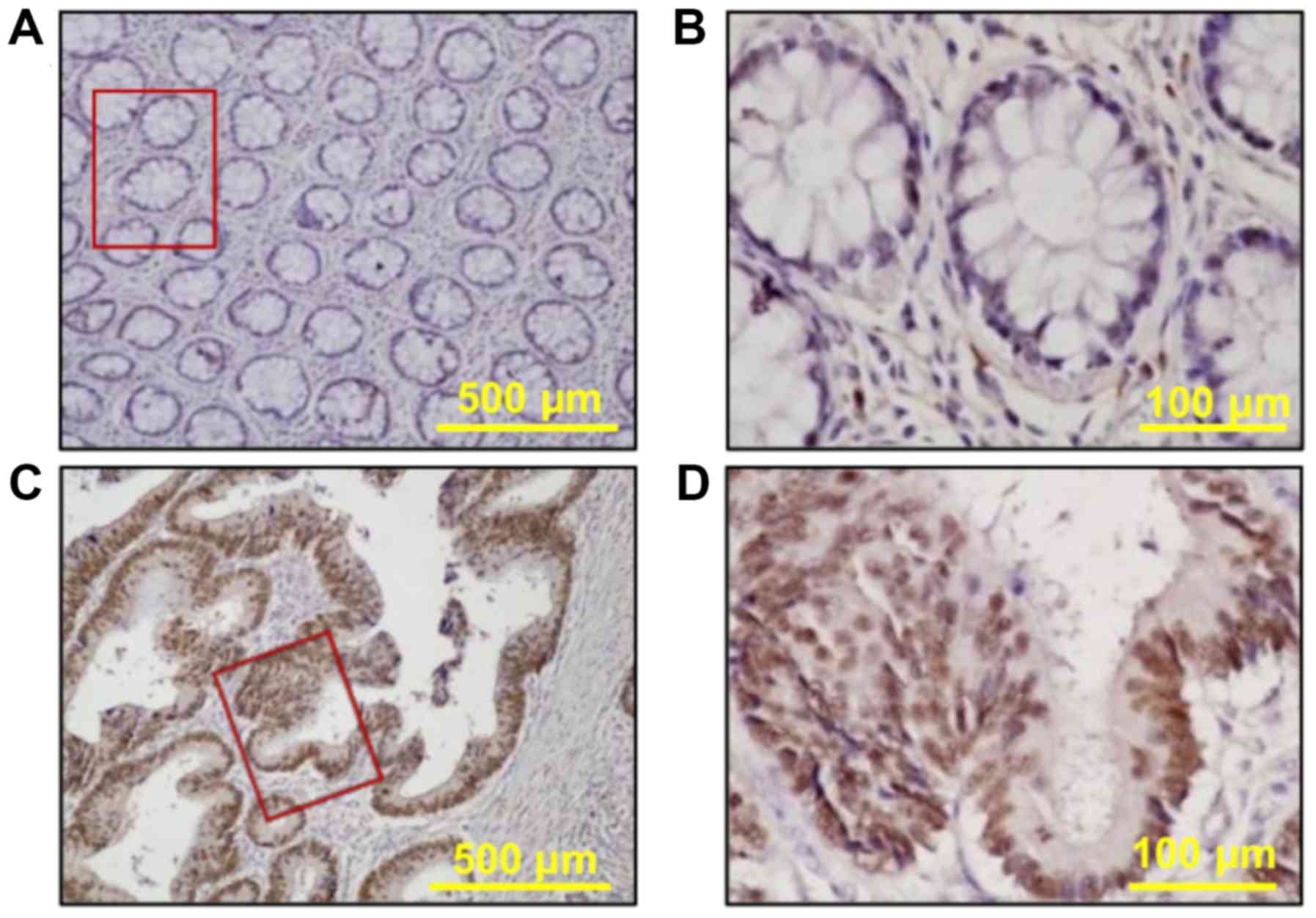
the clinicopathological data of these patients. FOXA1 protein
staining was either absent or weak in the normal colorectal mucosa
(Fig. 7A and B). The
immunohistochemical staining of FOXA1 protein was predominantly
identified in the nucleus of colorectal cancer cells and was not
detected in the tumor stroma (Fig. 7C
and D). The survival rates of patients with colorectal cancer
and the association between the expression of FOXA1 and
clinicopathological parameters in these patients were analyzed. It
was observed that the expression of FOXA1 was significantly
associated with differentiation, cancer stage, lymphovascular
invasion, depth of invasion and lymph node metastasis (P=0.006,
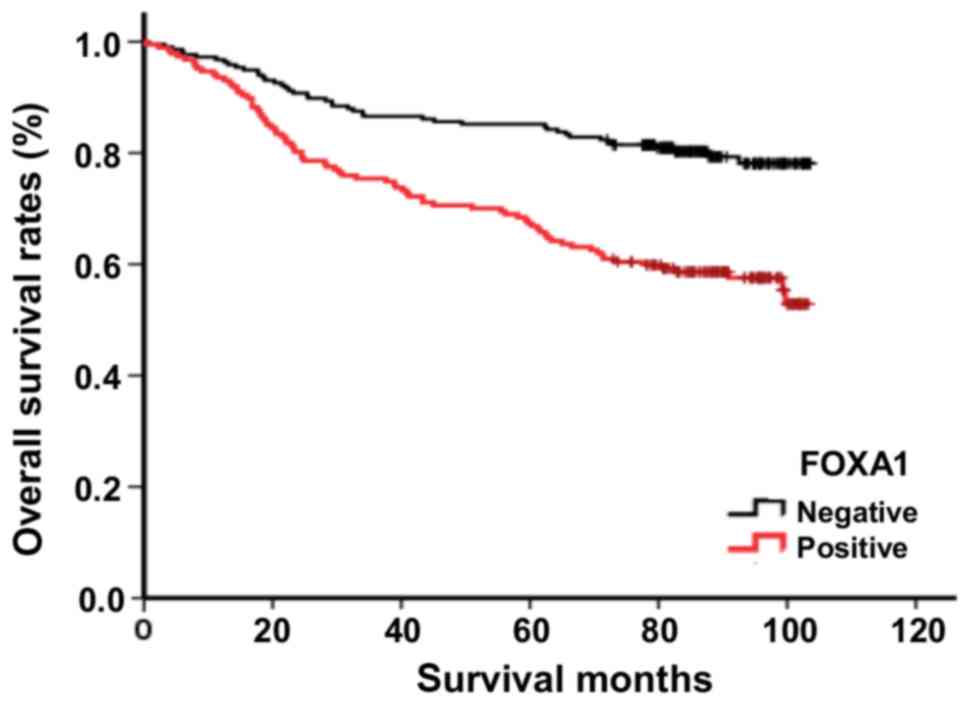
0.001, 0.034, 0.009 and 0.002, respectively; Table I). In addition, the overall
survival rates of patients with FOXA1-positive tumors were
significantly lower than those of patients with FOXA1-negative
tumors (P<0.001; Fig. 8).
 | Table IAssociation between the expression of
FOXA1 and the clinicopathological parameters of human colorectal
cancer. |
Table I
Association between the expression of
FOXA1 and the clinicopathological parameters of human colorectal
cancer.
| Total | FOXA1
| P-value |
|---|
| Negative | Positive |
|---|
| Parameter
(n=403) | | (n=216) | (n=187) | |
| Age (years) | | | | 0.254 |
| <69.6 | 176 | 100 | 76 | |
| ≥69.6 | 227 | 116 | 111 | |
| Sex | | | | 0.900 |
| Male | 247 | 133 | 114 | |
| Female | 156 | 83 | 73 | |
| Tumor size
(cm) | | | | 0.096 |
| <4.9 | 199 | 115 | 84 | |
| ≥4.9 | 204 | 101 | 103 | |
| Histologic
type | | | | 0.006 |
|
Differentiated | 356 | 182 | 174 | |
|
Undifferentiated | 47 | 34 | 13 | |
| Stage | | | | 0.001 |
| I | 63 | 44 | 19 | |
| II | 147 | 86 | 61 | |
| III | 160 | 74 | 86 | |
| IV | 33 | 12 | 21 | |
| Lymphovascular
invasion | | | | 0.034 |
| Negative | 294 | 167 | 127 | |
| Positive | 109 | 49 | 60 | |
| Perineural
invasion | | | | 0.170 |
| Negative | 285 | 159 | 126 | |
| Positive | 118 | 57 | 61 | |
| Depth of
invasion | | | | 0.009 |
| T1/T2 | 83 | 55 | 28 | |
| T3/T4 | 320 | 161 | 159 | |
| Lymph node
metastasis | | | | 0.002 |
| N0 | 218 | 132 | 86 | |
| N1-3 | 185 | 84 | 101 | |
| Distant
metastasis | | | | 0.248 |
| M0 | 349 | 191 | 158 | |
| M1 | 54 | 25 | 29 | |
Association between the expression of
FOXA1 and cell apoptosis and proliferation in human colorectal
cancer
All tumor samples were subjected to a TUNEL assay
and immunohistochemical staining of Ki-67 to determine apoptosis
and cell proliferation in the tumor cells. The AI of the 403 tumor
samples ranged between 0.9 and 19.9, with a mean AI of 8.7±6.3. No
significant association was observed between the expression of
FOXA1 and AI (P=0.152). The KI of the 403 tumor samples ranged
between 32.4 and 97.3, with a mean KI of 62.4±18.5. The mean KI
value of FOXA1-positive tumors was 74.8±14.9, which was
significantly lower than that of FOXA1-negative tumors (P<0.001)
(Table II).
| Table IIAssociation between the expression of
FOXA1 and the apoptosis and proliferation of cells in human
colorectal cancer.. |
Table II
Association between the expression of
FOXA1 and the apoptosis and proliferation of cells in human
colorectal cancer..
| Parameter (mean ±
SD) | Total (n=403) | FOXA1 expression
| P-value |
|---|
| Negative
(n=217) | Positive
(n=187) |
|---|
| AI | 8.7±6.3 | 9.8±7.3 | 7.2±4.6 | 0.152 |
| KI | 62.4±18.5 | 53.2±15.0 | 74.8±14.9 | <0.001 |
Discussion
FOXA1 is a member of the forkhead superfamily of
transcription factors. FOXA1 is an important regulator in the
development, differentiation and metabolism of numerous human
organs (4,5). Furthermore, accumulating evidence has
progressively contributed to our understanding of the critical role
of FOXA1 in human cancer (6-9).
FOXA1 is multifunctional and has been shown to function as a tumor
suppressor gene or an oncogene in various types of human cancer
(10-23). However, the role of FOXA1 in
colorectal cancer lacks support from basic and clinical data.
The regulation of cell migration, invasion and
survival is crucial in maintaining normal cellular homeostasis and
organogenesis of human tissues. Its loss is a major hallmark of
cancer, leading to cancer development and progression (25-27).
The present study first investigated the impact of FOXA1 on the
alteration of phenotypes in human colorectal cancer cells. Among
the human colorectal cancer cells assessed, the protein expression
of FOXA1 was the highest in DLD1 cells and the lowest in HCT116
cells. Therefore, siRNA in DLD1 and the pcDNA6-myc vector in HCT116
cells were used to control the endogenous protein expression of
FOXA1, either through knockdown or overexpression.
In the present study, FOXA1 knockdown suppressed
cell invasion, induced apoptosis and cell cycle arrest and
inhibited cell proliferation in human colorectal cancer cells;
these effects were reversed following the overexpression of FOXA1.
These results suggest that FOXA1 contributes to the alteration of
invasive and oncogenic phenotypes in human colorectal cancer cells.
In addition, previous studies have indicated that FOXA1 has a
potential role in regulating cell migration, invasion and survival
in various human cancer cells as an inhibitor or enhancer (11,15,16,28).
To examine the potential mechanisms involved in the
above effects, the present study examined the effect of FOXA1 on
the stimulation of an oncogenic signaling pathway, involved in cell
migration, invasion and cell survival. STAT3 is a key signaling
protein that is activated by the stimulation of various cytokines,
hormones and growth factors, and elicits diverse biological
outcomes including cell growth, differentiation and survival. STAT3
is phosphorylated on its tyrosine residues via Janus kinases, and
then forms homo- or heterodimers, translocates to the nucleus, and
binds DNA to initiate the transcription of target genes (29-31).
The constitutive activation of STAT3 signaling has been reported in
numerous human cancer types. However, the regulation and biological
consequences of the activation of STAT3 are complex. Aberrant
activation of STAT3 has been found to be associated with either
oncogenic or tumor suppressing functions in various types of human
cancer (32-34). In the present study, the
phosphorylation of STAT3 was decreased by FOXA1 knockdown. By
contrast, the phosphorylation of STAT3 was increased by the
overexpression of FOXA1 in human colorectal cancer cells. These
results suggest that FOXA1 may be associated with activation of
STAT3, which is important for tumor cell survival in human
colorectal cancer.
Subsequently, the expression of FOXA1 was we
evaluated in a well-defined series of human colorectal cancer,
including long-term and complete follow-up, with specific reference
to patient prognosis. It was observed that the expression of FOXA1
was increased in human colorectal cancer tissues compared with that
in normal colorectal mucosa. The expression of FOXA1 was
significantly associated with cell differentiation, lymphovascular
invasion, cancer stage, invasion depth, lymph node metastasis and
poor survival rate. Previously, the overexpression of FOXA1 was
reported as a good prognostic marker in human estrogen
receptor-positive breast cancer (11), endometrial (12) and bladder cancer (14), but a poor prognostic indicator in
human lung (18), thyroid
(19), colorectal (23) and gastric cancer (28). These results suggest that FOXA1 may
be a potential prognostic marker, depending on the specific cancer
type, and may serve as a poor prognostic marker and a promising
therapeutic target in colorectal cancer.
Finally, the present study evaluated the association
between the expression of FOXA1 and cell survival, including
proliferation and apoptosis, in human colorectal cancer tissues to
confirm the results obtained from the in vitro experiments.
It was observed that the mean KI value of FOXA1-positive tumors was
significantly higher than that of FOXA1-negative tumors. Ki-67 is a
nuclear antigen and an established proliferation marker of tumor
cells in various types of human cancer, including colorectal cancer
(35-37). However, no significant association
was observed between the expression of FOXA1 and the AI value.
These in vivo results are in accordance with the conclusion
that FOXA1 serves a crucial role in cell proliferation in human
colorectal cancer cell lines.
In conclusion, FOXA1 is an important mediator of
proliferative and anti-apoptotic activities in human colorectal
cancer cells. FOXA1 was upregulated in human colorectal cancer
tissues and was associated with poor prognosis, suggesting an
oncogenic role of FOXA1 in the development and progression of human
colorectal cancer.
Funding
This study was supported by research funds from the
Research Institute of Clinical Medicine, Chonnam National
University Hwasun Hospital in 2017 (grant no. HCRI 17912-1),
Republic of Korea, and the National Research Foundation of Korea
grant (grant no. NRF-2017R1A2B4004703) funded by the Korean
government (Ministry of Science, ICT and Future Planning), Republic
of Korea.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
YEJ was involved in the conceptualization of the
study. YLP, SHK, SYP, MWJ, SYH, JHC, DSM, SBC, WSL and HSK were
involved in data curation, were responsible for formal analysis and
provided resources. YLP, SHK and YEJ were involved in the
investigative part of the study. YEJ was involved in the
methodology, was involved in project administration and was
involved in the writing of the manuscript and original draft
preparation. YLP, SHK and YEJ were involved in the writing,
reviewing and editing of the manuscript. All authors read and
approved the final manuscript
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of the Chonnam National University Hwasun Hospital
(Jeonnam, Korea; CNUHH-2017-163). This was a retrospective study
and written informed consent was obtained from each participant
prior to tissue acquisition at the time of hospitalization. All
participants provided written consent for their information to be
stored in the hospital database and used for research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar
|
|
2
|
Choi Y, Sateia HF, Peairs KS and Stewart
RW: Screening for colorectal cancer. Semin Oncol. 44:34–44. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahmed FE: Development of novel diagnostic
and prognostic molecular markers for sporadic colon cancer. Expert
Rev Mol Diagn. 5:337–352. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Katoh M and Katoh M: Human FOX gene family
(Review). Int J Oncol. 25:1495–1500. 2004.PubMed/NCBI
|
|
5
|
Kaestner KH: The FoxA factors in
organogenesis and differentiation. Curr Opin Genet Dev. 20:527–532.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bernardo GM and Keri RA: FOXA1: A
transcription factor with parallel functions in development and
cancer. Biosci Rep. 32:113–130. 2012. View Article : Google Scholar
|
|
7
|
Augello MA, Hickey TE and Knudsen KE:
FOXA1: Master of steroid receptor function in cancer. EMBO J.
30:3885–3894. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Katoh M, Igarashi M, Fukuda H, Nakagama H
and Katoh M: Cancer genetics and genomics of human FOX family
genes. Cancer Lett. 328:198–206. 2013. View Article : Google Scholar
|
|
9
|
Zhang G, Zhao Y, Liu Y, Kao LP, Wang X,
Skerry B and Li Z: FOXA1 defines cancer cell specificity. Sci Adv.
2:e15014732016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shou J, Lai Y, Xu J and Huang J:
Prognostic value of FOXA1 in breast cancer: A systematic review and
meta-analysis. Breast. 27:35–43. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park S, Koh E, Koo JS, Kim SI, Park BW and
Kim KS: Lack of both androgen receptor and forkhead box A1 (FOXA1)
expression is a poor prognostic factor in estrogen
receptor-positive breast cancers. Oncotarget. 8:82940–82955. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abe Y, Ijichi N, Ikeda K, Kayano H,
Horie-Inoue K, Takeda S and Inoue S: Forkhead box transcription
factor, forkhead box A1, shows negative association with lymph node
status in endo-metrial cancer, and represses cell proliferation and
migration of endometrial cancer cells. Cancer Sci. 103:806–812.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamashita H, Amponsa VO, Warrick JI, Zheng
Z, Clark PE, Raman JD, Wu XR, Mendelsohn C and DeGraff DJ: On a FOX
hunt: Functions of FOX transcriptional regulators in bladder
cancer. Nat Rev Urol. 14:98–106. 2017. View Article : Google Scholar
|
|
14
|
Reddy OL, Cates JM, Gellert LL, Crist HS,
Yang Z, Yamashita H, Taylor JA III, Smith JA Jr, Chang SS, Cookson
MS, et al: Loss of FOXA1 drives sexually dimorphic changes in
urothelial differentiation and is an independent predictor of poor
prognosis in bladder cancer. Am J Pathol. 185:1385–1395. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gan HY, Li N, Zhang Q and Feng ZZ:
Silencing FOXA1 gene regulates liver cancer cell apoptosis and cell
proliferation. Eur Rev Med Pharmacol Sci. 22:397–404.
2018.PubMed/NCBI
|
|
16
|
He S, Zhang J, Zhang W, Chen F and Luo R:
FOXA1 inhibits hepatocellular carcinoma progression by suppressing
PIK3R1 expression in male patients. J Exp Clin Cancer Res.
36:1752017. View Article : Google Scholar
|
|
17
|
Song Y, Washington MK and Crawford HC:
Loss of FOXA1/2 is essential for the epithelial-to-mesenchymal
transition in pancreatic cancer. Cancer Res. 70:2115–2125. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Zhang S, Zhu L and Ma S: Role of
transcription factor FOXA1 in non-small cell lung cancer. Mol Med
Rep. 17:509–521. 2018.
|
|
19
|
Li M, Zhang W, Liu C, Shi Y, Tang W, Chen
S, Gu H, Yin J, Zhang Z and Jiang P: Forkhead box A1 (FOXA1)
tagging polymorphisms and esophageal cancer risk in a Chinese
population: A fine-mapping study. Biomarkers. 21:523–529. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tsourlakis MC, Eleftheriadou A, Stender A,
Weigand P, Grupp K, Hube-Magg C, Kluth M, Schroeder C, Steurer S,
Hinsch A, et al: FOXA1 expression is a strong independent predictor
of early PSA recurrence in ERG negative prostate cancers treated by
radical prostatectomy. Carcinogenesis. 38:1180–1187. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takayama K and Inoue S: Transcriptional
network of androgen receptor in prostate cancer progression. Int J
Urol. 20:756–768. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nucera C, Eeckhoute J, Finn S, Carroll JS,
Ligon AH, Priolo C, Fadda G, Toner M, Sheils O, Attard M, et al:
FOXA1 is a potential oncogene in anaplastic thyroid carcinoma. Clin
Cancer Res. 15:3680–3689. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma W, Jiang J, Li M, Wang H, Zhang H, He
X, Huang L and Zhou Q: The clinical significance of forkhead box
protein A1 and its role in colorectal cancer. Mol Med Rep.
14:2625–2631. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Greene FL, Page DL, Fleming ID, Fritz AG,
Balch CM, Haller DG and Morrow M: AJCC cancer staging manual. 6th
edition. Springer-Verlag; New York, NY: 2002, View Article : Google Scholar
|
|
25
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brábek J, Mierke CT, Rösel D, Veselý P and
Fabry B: The role of the tissue microenvironment in the regulation
of cancer cell motility and invasion. Cell Commun Signal. 8:222010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kiechle FL and Zhang X: Apoptosis:
Biochemical aspects and clinical implications. Clin Chim Acta.
326:27–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ren H, Zhang P, Tang Y, Wu M and Zhang W:
Forkhead box protein A1 is a prognostic predictor and promotes
tumor growth of gastric cancer. Onco Targets Ther. 8:3029–3039.
2015.PubMed/NCBI
|
|
29
|
Villarino AV, Kanno Y, Ferdinand JR and
O’Shea JJ: Mechanisms of Jak/STAT signaling in immunity and
disease. J Immunol. 194:21–27. 2015. View Article : Google Scholar :
|
|
30
|
Abroun S, Saki N, Ahmadvand M, Asghari F,
Salari F and Rahim F: STATs: An old Story, yet mesmerizing. Cell J.
17:395–411. 2015.PubMed/NCBI
|
|
31
|
Zundler S and Neurath MF: Integrating
immunologic signaling networks: The JAK/STAT pathway in colitis and
colitis-associated cancer. Vaccines (Basel). 4:E52016. View Article : Google Scholar
|
|
32
|
Guanizo AC, Fernando CD, Garama DJ and
Gough DJ: STAT3: A multifaceted oncoprotein. Growth Factors.
36:1–14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Aigner P, Just V and Stoiber D: STAT3
isoforms: Alternative fates in cancer. Cytokine Jul. 18:2018(Epub
ahead of print). pii:S1043-4666(18)30300-4.2018. View Article : Google Scholar
|
|
34
|
Lai PS, Rosa DA, Magdy Ali A, Gómez-Biagi
RF, Ball DP, Shouksmith AE and Gunning PT: A STAT inhibitor patent
review: Progress since 2011. Expert Opin Ther Pat. 25:1397–1421.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun X and Kaufman PD: Ki-67: More than a
proliferation marker. Chromosoma. 127:175–186. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ragab HM, Samy N, Afify M, El Maksoud NA
and Shaaban HM: Assessment of Ki-67 as a potential biomarker in
patients with breast cancer. J Genet Eng Biotechnol. 16:479–484.
2018. View Article : Google Scholar
|
|
37
|
Hilska M, Collan YU, O Laine VJ, Kössi J,
Hirsimäki P, Laato M and Roberts PJ: The significance of tumor
markers for proliferation and apoptosis in predicting survival in
colorectal cancer. Dis Colon Rectum. 48:2197–2208. 2005. View Article : Google Scholar
|