Introduction
Laryngeal carcinoma is one of the most common
malignant tumors among head and neck carcinomas, and >95% of
laryngeal carcinomas are of the squamous cell type [laryngeal
squamous cell carcinoma (LSCC)]. In 2018, there were ~177,422 new
cases and ~94,771 LSCC-related deaths worldwide (1). The incidence of LSCC is increasing
annually worldwide. Despite major advances in LSCC diagnosis and
treatment, the prognosis of patients with advanced LSCC remains
dismal. Treatment failure is most commonly attributed to recurrence
and metastasis (2). Approximately
60% of patients with LSCC are already at an advanced stage (III or
IV) at the time of treatment (3).
Therefore, the identification of specific molecular biomarkers for
early diagnosis and effective targeted therapy is crucial for the
early detection, treatment and follow-up of LSCC.
With the development of high-throughput sequencing
technologies, only <2% of the transcripts were found to encode
proteins, whereas the vast majority were transcribed into
non-coding RNAs, including microRNAs (miRNAs or miRs), small
interfering RNAs (siRNAs) and 1ong non-coding RNAs (lncRNAs)
(4). lncRNAs are a class of
non-coding RNAs with a length of >200 nucleotides which,
although they cannot encode proteins themselves, regulate gene
expression at the transcriptional, as well as the
post-transcriptional level. Furthermore, lncRNAs are involved in a
number of cellular functions, such as chromatin remodeling, RNA
decay, epigenetic regulation and chromatin modification (5,6), and
are closely associated with tumor formation, invasion and
metastasis (7,8). However, the role of lncRNAs in LSCC
has not yet been fully determined. Certain lncRNAs, such as
H19 (9), HOTAIR
(10,11), NEAT1 (12) and TUG1 (13), have been shown to be upregulated in
laryngeal cancer cells and tissues, and may promote cancer by
participating in various biological processes. The differential
expression of lncRNAs was detected by microarray assays on four
pairs of LSCC and adjacent normal tissues. The lncRNA,
miR-155 host gene (MIR155HG), was found to be
upregulated in LSCC tissues. By searching biological information
websites (UCSC, http://genome.ucsc.edu/), miR-155-5p was found
to be located in the third exon of MIR155HG. In recent
years, the inter-relationship between miRNAs and lncRNAs has become
a main focus of research. lncRNAs can form the precursors of miRNAs
through intracellular shear action, and certain genes can be
transcribed to lncRNAs and miRNAs at the same time (14). For example, lncRNA
MIR100HG-derived miR-100 and miR-125b have
been shown to participate in the resistance of colorectal cancer to
cetuximab through Wnt/β-catenin signaling (15). MIR31HG, the host gene of
miR-31, has been shown to contribute to the pathogenesis of
Hirschsprung’s disease through the
MIR31HG-miR-31/31*-ITIH5/PIK3CG pathway
(16). MIR155HG-derived
miR-155-5p and miR-155-3p have been shown to suppress
the transcription and translation of protocadherin (PCDH)9
and PCDH7, thereby promoting glioma cell migration and
invasion (17). However, the roles
of miR-155-5p and MIR155HG and their interaction in
laryngeal cancer have not yet been fully elucidated.
Epithelial-to-mesenchymal transition (EMT) is
associated with distant metastasis and tumor dissemination.
Multiple growth factors and cytokines may induce EMT, and
transforming growth factor (TGF)-β is a key factor in
the induction of EMT (18). EMT
has been reported to be involved in the development of LSCC.
Non-coding RNAs, such as miR-203 (19), miR-205 and miR-375
(20), may regulate the
progression of LSCC by regulating EMT. The coding gene enhancer of
zeste homolog 2 (EZH2) may also promote the invasion and
migration of LSCC cells through EMT (21). However, whether lncRNAs are
involved in the EMT of laryngeal cancer cells remains unclear.
Therefore, the aim of the present study was to
detect the expression of MIR155HG and its exon miRNA,
miR-155-5p, in LSCC, and to explore the potential functional
roles and downstream regulatory mechanisms of the
MIR155HG/miR-155 axis in the development and
progression of LSCC, as well as its role in TGF-β-induced
EMT. The aim of this study was also to determine whether
MIR155HG plays a carcinogenic role in LSCC, and whether it
may be used as a biomarker and as a target in novel therapeutic
strategies for patients with LSCC.
Materials and methods
Patients and tissue specimens
LSCC tissue samples and adjacent normal tissues were
collected from 45 patients with LSCC at the Otorhinolaryngology
Head and Neck Surgery Biobank of Hebei Medical University
(Shijiazhuang, China) from October, 2016 to March, 2018. Informed
consent was obtained from all patients, none of whom had received
chemotherapy or radiotherapy prior to surgery. The use of human
tissues specimens was approved by and carried out according to the
guidelines of the Ethics Committee of the Second Hospital of Hebei
Medical University (Shijiazhuang, China). One part of the tissue
specimens was placed into RNAlater solution (CoWin Biosciences,
Beijing, China) and stored at -80°C for RNA extraction. The other
part of the tissue specimens was fixed in 10% neutral formaldehyde
solution, and paraffin blocks were routinely prepared and
preserved. Tumor and normal adjacent tissues were confirmed by
routine pathological diagnosis. Agilent SBC Human (4*180K) lncRNA
Microarray (ID: 74348) was used to test the transcript expression
profiles in 4 pairs of LSCC and normal tissues. The
clinicopathological characteristics of the 45 paired specimens are
presented in Table SI.
Cell culture
Three human LSCC cell lines (TU686, TU177 and
AMC-HN-8) and 293T cells were purchased from BNBIO (Beijing, China)
and preserved at the Otorhinolaryngology Head and Neck Surgery
Biobank of Hebei Medical University. The TU686 and TU177 cells were
cultured in RPMI-1640 medium (Gibco/Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), supplemented with 10% fetal bovine serum (FBS;
Gibco/Thermo Fisher Scientific, Inc.). The AMC-HN-8 and 293T cells
were cultured in Dulbecco’s modified Eagle’s medium (Gibco/Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS. The TU177 cells
were treated with 10 ng/ml recombinant TGF-β (R&D
Systems, Inc., Minneapolis, MN, USA) for 7 days and the medium was
replenished every 2 days. All the cells were cultured at 37°C in a
humidified 5% CO2 incubator (Thermo Fisher Scientific,
Inc.).
RNA extraction and reverse tran
scription-quantitative polymerase chain reaction (RT-qPCR)
assay
Total RNA was extracted from the tissues and cells
using the the Eastep®Super Total RNA Extraction kit
(Promega, Madison, WI, USA), and the RNA integrity was evaluated by
1% agarose gel electrophoresis (containing DEPC; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). cDNA was synthesized using
the Transcriptor First Strand cDNA Synthesis kit (Roche, Basel,
Switzerland) following the manufacturer’s protocol. RT-qPCR was
performed with GoTap®qPCRMaster Mix (Promega) according
to the manufacturer’s instructions using CFX96™ Real-Time PCR
Detection Systems. The reaction conditions were as follows
(two-step method): One cycle of pre-denaturation for 2 min at 95°C,
followed by 40 cycles of 15 sec at 95°C for denaturation, and for
annealing to extension, selecting the most suitable annealing
temperature according to different primers for 60 sec. The relative
expression levels were estimated with the 2-ΔΔCq method.
Each specimen was tested 3 times. The specific primer sequences are
listed in Table SII.
Cell cytoplasm and nuclear fraction
isolation
Subcellular fractionation was performed with the
PARIS™ Kit Protein and RNA Isolation System (Invitrogen/Thermo
Fisher Scientific, Inc.) from LSCC cell lines according to the
manufacturer’s instructions.
Cell transfection
The 4 shRNAs specifically targeting MIR155HG
and the control sh-NC were synthesized by Vigene Biosciences
(Shandong, China). The pcDNA3.1-MIR155HG vector was
synthesized by Sangon Biotech (Beijing, China).
hsa-miRNA-155-5p mimic/inhibitor/negative controls were
purchased from GenePharma (Shanghai, China) (Table SII). The cells were grown on
6-well plates and transfected using Lipofectamine 2000
(Invitrogen/Thermo Fisher Scientific, Inc.) when they reached 80%
confluence, according to the manufacturer’s instructions. At 48 h
following transfection, the cells were collected for RT-qPCR
verification and further experiments.
Cell proliferation assay
Cell proliferation was detected by MTS assay. Cells
were seeded in 96-well plates at 2×103 per well
following transfection for 24 h. Cell proliferation was evaluated
using the CellTiter96®AQueousOne Solution Cell
Proliferation Assay kit (Promega) according to the manufacturer’s
instructions. MTS reagent (20 μl) was added to 100 μl
culture medium after seeding for 0, 24, 48, 72 and 96 h, and
incubated in CO2 incubator at 37°C for 2.5 h each time.
The absorbance at 490 nm was measured using a Spark®
multimode microplate reader (Tecan, Männedorf, Switzerland).
Colony formation assay
The cells were seeded in 6-well plates at a density
of 2×103 per well following transfection for 24 h. After
12-14 days, the cells were washed twice with phosphate-buffered
saline, fixed with paraformaldehyde and stained with 0.5% crystal
violet solution at room temperature for 20 min. (Generay, Shanghai,
China). The number of colonies (≥50 cells) was counted under a
microscope (Olympus, Tokyo, Japan).
Transwell migration and invasion
assays
For migration assays, non-Matrigel-coated chambers
(Corning Costar, Corning, NY, USA) with 8-μm pore membranes
were used. A total of 1×105 cells per well were seeded
into the upper chamber and 650 μl medium (10% FBS) was added
to the lower chamber. Following incubation in CO2
incubator (37°C, 5% CO2) (Thermo Fisher Scientific,
Inc.) for 24 h, the residual cells on the upper surface of the
membrane were removed, the membrane was fixed in 4%
paraformaldehyde for 20 min, stained with 0.5% crystal violet
solution for 20 min and examined under a microscope (CKX53;
Olympus, Tokyo, Japan) in 5 randomly selected fields. For invasion
assays, the upper chamber was pre-coated with 50 μl 1X
Matrigel® Basement Membrane Matrix (Corning Costar); the
remaining steps were as described above for the migration assay.
For the Transwell assays of TGF-β-treated and untreated
cells, 5×104 cells were seeded into the upper
chamber.
Tumor xenograft model
A total of 16 male nude (BALB/c-nude) mice (age, 5
weeks; weight, 20-25 g) were obtained from Beijing Vital River
Laboratory Animal Technology Co., Beijing, China; animal permit
no.: [SCXY (Jing) 2016-0006]. The maintenance conditions for the
mice were as follows: Temperature, 23-27°C; humidity, 40-60%;
ventilation, 15 times/h; 12 h light/12 h dark cycle. The mice were
fed standard laboratory food and water. The health and maintenance
condition of the mice were monitored every 2 days. The mice were
randomly divided into 2 groups (pcDNA3.1, and pcDNA3.1-MIR155HG;
n=8 per group). The TU177 cells (5×106) stably
transfected with pcDNA 3.1 and pcDNA3.1-MIR155HG in 100
μl PBS were subcutaneously injected into the right flanks of
the mice. Tumor volume was measured using a caliper at indicated
time points and calculated using the following formula: V = 0.5 ×
length x width2. After 4 weeks, the mice were sacrificed
by cervical dislocation and the tumors were removed and weighed.
The maximum diameter of a single tumor found was 17 mm and no mouse
developed multiple tumors. All mice were in well body condition
throughout the experiment. All animal experiments were performed at
the Experimental Animal Center of the Fourth Hospital of Hebei
Medical University according to the NIH guidelines, and were
approved by the Institutional Animal Care and Use Committee of the
Fourth Hospital of Hebei Medical University.
Prediction of miR-155-5p target
genes
The potential downstream targets of
miR-155-5p were predicted using TargetScan (http://www.targetscan.org/vert_72/) and miRwark
(http://mirwalk.umm.uni-heidelberg.de/). SOX10
was predicted by the two databases and considered a potential
target of miR-155-5p. The 3′-UTR of SOX10 mRNA
containing the intact miRNA-155-5p recognition sequences was
PCR-amplified and subcloned into the Nhel and Xbal
sites of the pmirGLO vector (Youbio, Changsha, China). Mutation
sites were promoted using the Site-Directed Mutagenesis kit (New
England Biolabs, Inc., Ipswich, MA, USA). Mutation primers were
designed in NEBase changer (http://nebasechanger.neb.com/) and synthesized by
Sangon Biotech (Table SII).
Western blot analysis
Cell protein lysates were prepared using RIPA buffer
(Solarbio, Beijing, China) followed by the addition of protease
inhibitor cocktail (Promega) and PMSF (Solarbio), according to the
manufacturers’ instructions. The protein concentration was measured
using the BCA Protein Assay kit (Generay). Proteins were separated
by 10% SDS-PAGE and then transferred to PVDF membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The membranes were
incubated with 5% non-fat milk (Becton-Dickinson and Company,
Suzhou, China) for 2 h at room temperature and incubated overnight
at 4°C with rabbit-anti-human SOX10 (molecular weight: 60
kDa) (1:2,000; cat. no. DF8009; Affinity, Cincinnati, OH, USA) and
rabbit-anti-human GAPDH (molecular weight: 37 kDa) (1:5,000;
cat. no. 10494-1-AP; Protein Tech Group, Inc., Chicago, IL, USA)
antibodies. Subsequently, the membranes were incubated at room
temperature with IgG (H+L) HRP (1:10,000; cat. no. RS0002; Ruiying
Bio, Suzhou, China) for 1 h. Proteins were visualized with an
enhanced ECL reagent (Vazyme, Nanjing, China) by ChemiDoc™ XRS+
(Bio-Rad Laboratories, Inc.).
Dual luciferase reporter assay
The TU177 and 293T cells were seeded at a density of
3×104 cells per well in 24-well plates. After 24 h, the
cells were co-transfected with pmirGLO-SOX10-WT or
pmirGLO-SOX10-MUT reporter plasmids, and miR-155-5p mimic or
inhibitor or NC. Following transfection for 48 h, the relative
luciferase activity was measured using the Dual-Luciferase Reporter
Assay System (Promega) according to the manufacturer’s protocol.
The luciferase activity of each well was normalized to
Renilla luciferase activity.
Statistical analysis
The results were analyzed using SPSS version 21.0
(IBM Corp., Armonk, NY, USA) and GraphPad Prism version 7 (GraphPad
Software, Inc., La Jolla, CA, USA). The differences between 2
groups were analyzed with the Student’s t-test. Differences between
>2 groups were determined by one-way ANOVA followed by Tukey’s
post hoc test. Differences between the growth of different groups,
two-way ANOVA was performed and followed by Tukey’s post hoc test.
Pearson’s correlation analysis was used to analyze the correlation
between the expression of MIR155HG and that of
miR-155-5p, and between the expression of miR-155-5p
and SOX10. Each experiment was performed at least 3 times.
All data are presented as the means ± standard deviation. P<0.05
was considered to indicate a statistically significant
difference.
Results
MIR155HG is upregulated in LSCC and is
associated with advanced clinicopathological characteristics
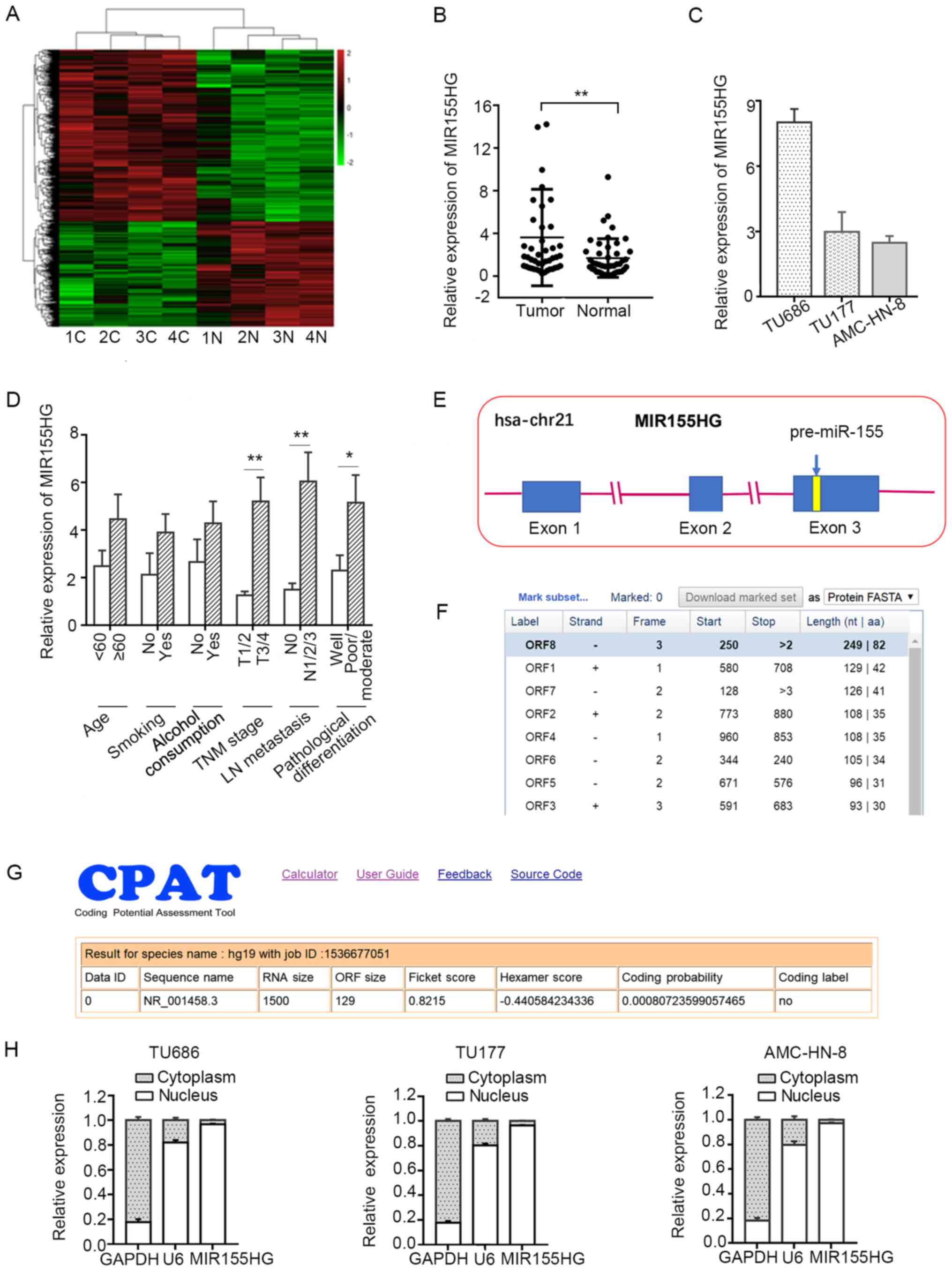
To determine the potential role of lncRNAs in LSCC,
microarray analysis was used to compare lncRNA expression levels
between LSCC tissues and corresponding normal tissues. There were
3,073 differentially expressed lncRNAs in total (fold change ≥2,
P<0.05), of which 1,967 were upregulated and 1,106 were
downregulated in LSCC tissues compared with corresponding normal
tissues (Fig. 1A). A miRNA host
gene, MIR155HG, which was found to be upregulated in the
microarray analysis, was selected for further analysis. To
investigate the functional role of MIR155HG in LSCC, the
expression of MIR155HG was initially analyzed in 45 pairs of
LSCC tissues and corresponding adjacent non-tumor tissues by
RT-qPCR. The expression level of MIR155HG was found to be
significantly increased in the LSCC tissues compared with the
adjacent normal tissues (Fig. 1B).
The expression levels of MIR155HG in 3 LSCC cell lines
(TU686, TU177 and AMC-HN-8) are shown in Fig. 1C. The association between
MIR155HG expression and different clinicopathological
characteristics in LSCC was further analyzed. High expression
levels of MIR155HG were found to be closely associated with
poor differentiation, lymph node metastasis and a more advanced TNM
stage. However, there was no observed association between the
expression of MIR155HG and age, alcohol consumption or
smoking (Fig. 1D).
MIR155HG is located on chromosome 21q21.2,
and contains 3 exons and 2 introns. miR-155-5p is located
within the third exon of MIR155HG (Fig. 1E). The coding potential of
MIR155HG was analyzed by using several prediction software
programs, and the results revealed that the open reading frame
(ORF) of MIR155HG was very short, containing only 82 amino
acids (https://www.ncbi.nlm.nih.gov/orffinder/), and had no
coding potential (http://lilab.research.bcm.edu/cpat/) (Fig. 1F and G). These results indicated
that MIR155HG is a non-coding RNA. Furthermore,
MIR155HG was found to be predominantly located in the
nucleus of LSCC cells by subcellular fractionation assay (Fig. 1H).
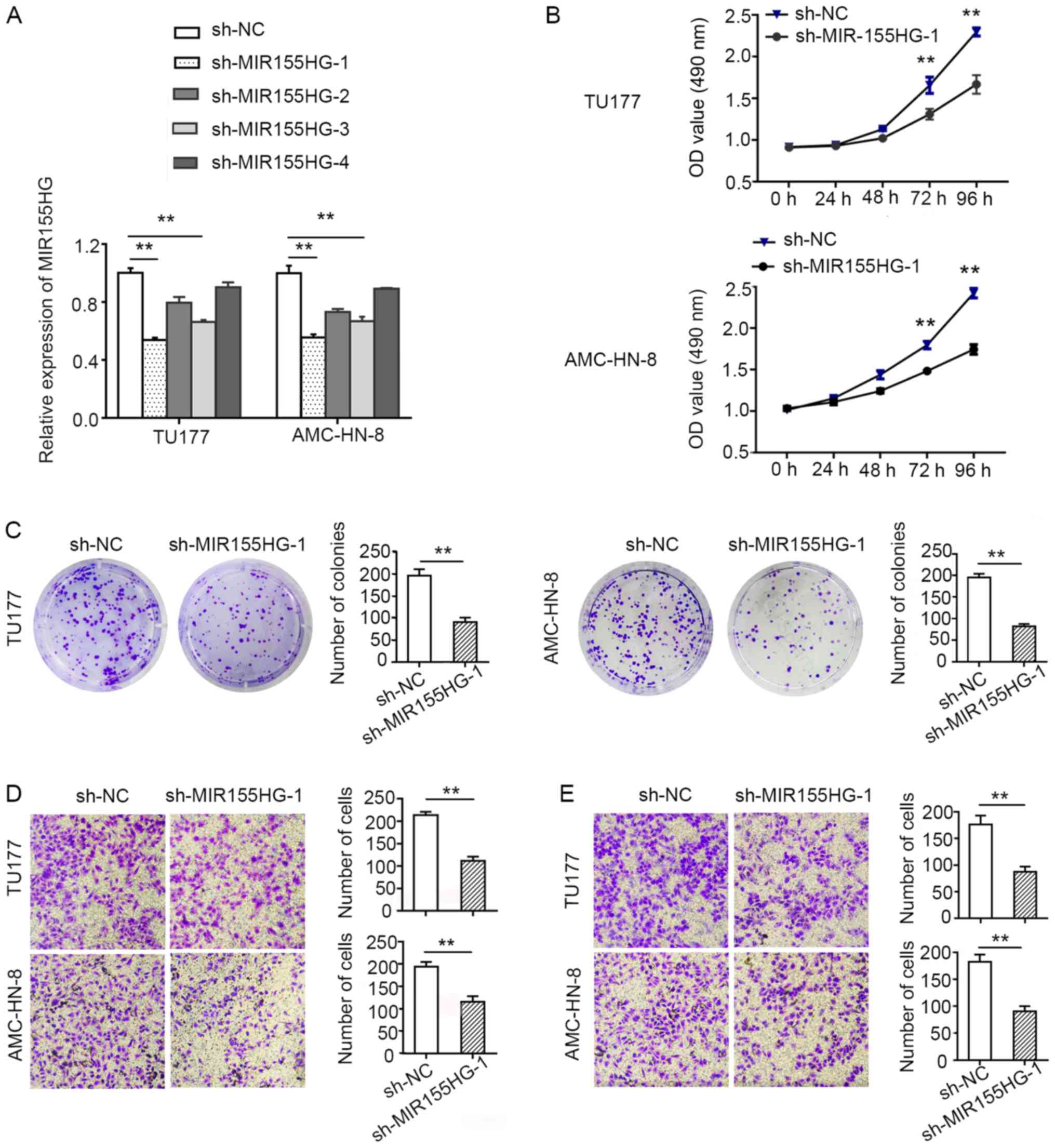
Knockdown of MIR155HG suppresses LSCC
cell proliferation, migration and invasion
To explore the biological function of
MIR155HG, shRNAs against MIR155HG
(sh-MIR155HG) were transfected into the LSCC cells to knock
down endogenous MIR155HG expression. Due to the poor
transfection efficiency, the TU686 cells, which had a relatively
higher expression level of MIR155HG, were not selected for
the in vitro assay. The TU177 and AMC-HN-8 cells, with a
relatively higher transfection efficiency, were used as model cells
for the follow-up experiments. sh-MIR155HG-1, which exhibited the
most evident knockdown efficacy, was selected for use in further
experiments (Fig. 2A).
MTS and colony formation assays were used to measure
cell proliferation. MIR155HG knockdown suppressed TU177 and
AMC-HN-8 cell proliferation (Fig.
2B). Consistent with the results of MTS assay, the results of
colony formation assay also revealed that the downregulation of
MIR155HG suppressed the colony-forming ability of the TU177
and AMC-HN-8 cells (Fig. 2C).
These data provide evidence to suggest the tumor growth-promoting
role of MIR155HG in vitro.
In order to examine the effects of MIR155HG
on the malignant biological properties of LSCC cells, the cell
migratory and invasive abilities were evaluated using Transwell
Matrigel migration and invasion assays. The migration and invasion
capacities of the TU177 and AMC-HN-8 cells were found to be
markedly decreased when MIR155HG was knocked down (Fig. 2D and E). Taken together, these data
demonstrated that MIR155HG suppressed the proliferation,
migration and invasion of LSCC cells.
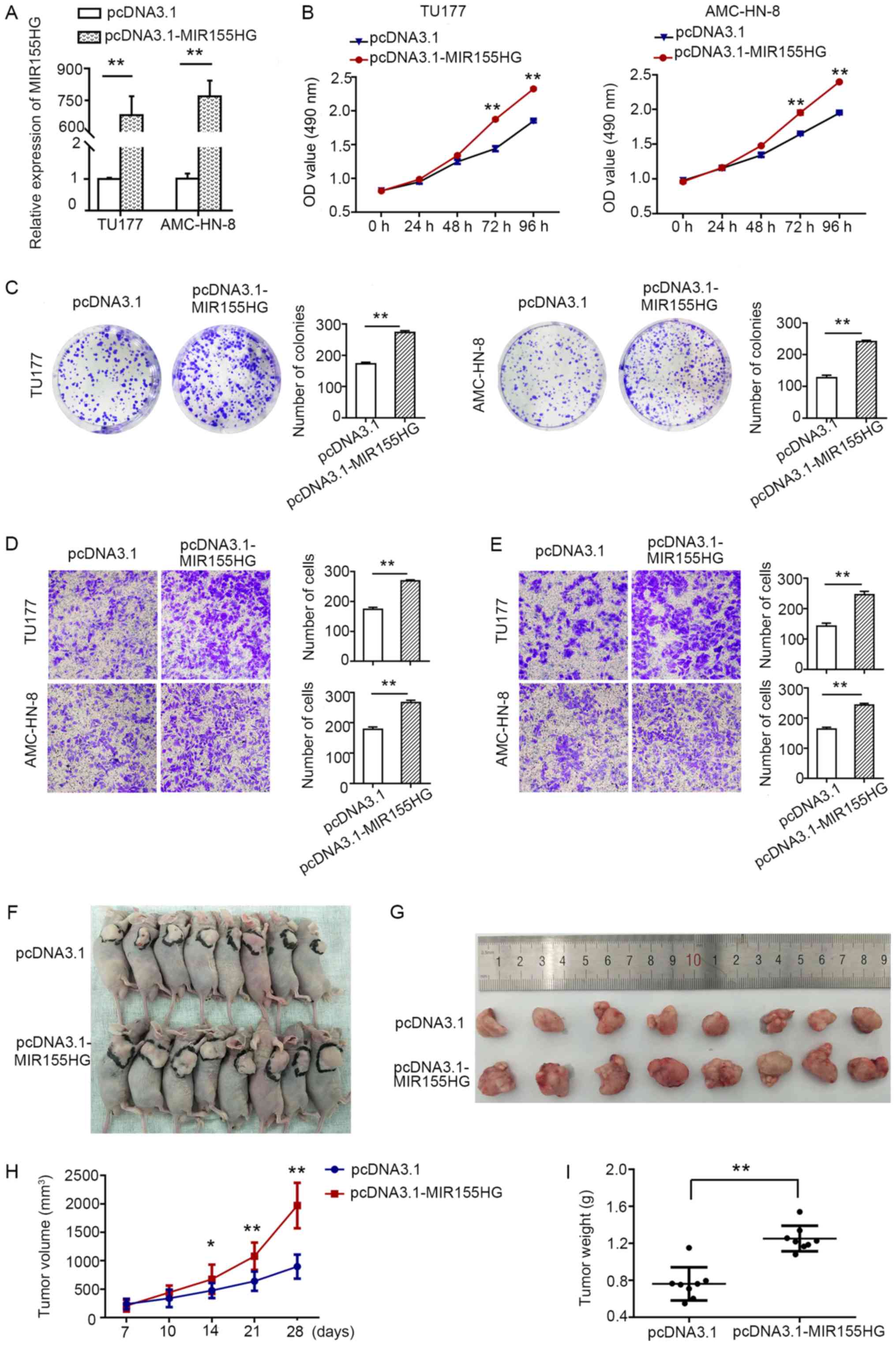
Overexpression of MIR155HG promotes LSCC
cell proliferation, migration and invasion in vitro and promotes
tumorigenesis in vivo
Subsequently, in order to assess the biological
effects of MIR155HG overexpression, the
pcDNA3.1-MIR155HG expression plasmid was transfected into
the TU177 and AMC-HN-8 cells with relatively low expression levels
of MIR155HG. pcDNA3.1 was used as a control. The relative
expression level of MIR155HG in the
pcDNA3.1-MIR155HG-transfected TU177 and AMC-HN-8 cells was
markedly increased (Fig. 3A). MTS
(Fig. 3B) and colony formation
assays (Fig. 3C) demonstrated that
MIR155HG upregulation promoted TU177 and AMC-HN-8 cell
proliferation. Moreover, MIR155HG overexpression enhanced
the migratory and invasive capacity of both cell lines (Fig. 3D and E). All these results
indicated that the overexpression of MIR155HG increased the
tumorigenicity of LSCC cells. To further verify the results of the
in vitro experiments, animal experiments were conducted. A
TU177 cell line stably transfected with pcDNA3.1 and
pcDNA3.1-MIR155HG was constructed, and 5×106
cells were subcutaneously injected into nude mice. The results
demonstrated that the overexpression of MIR155HG
significantly promoted tumor growth compared with the pcDNA3.1
group (Fig. 3F and G), whereas the
volume and weight of the tumors were higher in the MIR155HG
overexpression group (Fig. 3H and
I).
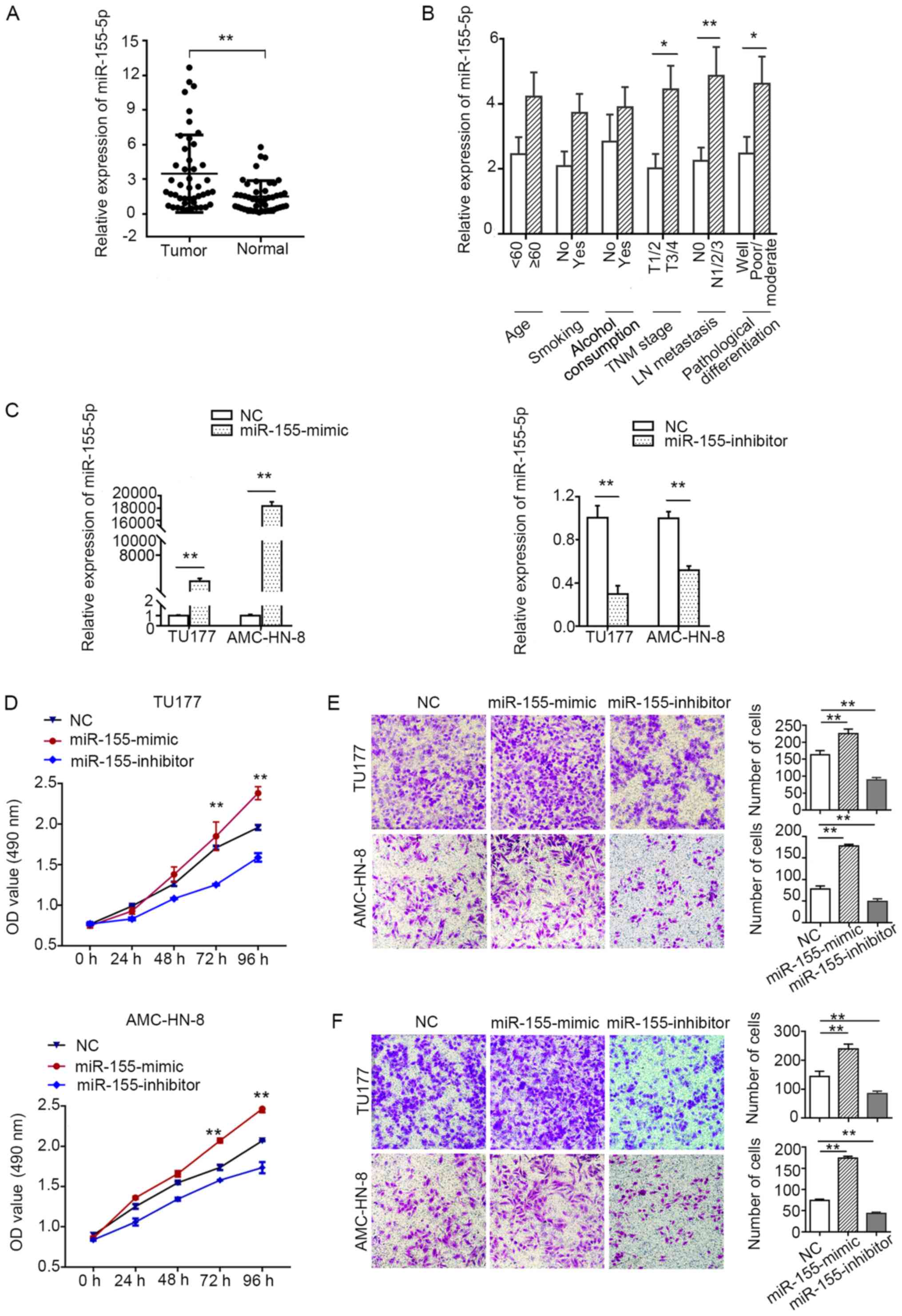
miR-155-5p is upregulated in LSCC and
promotes cancer cell proliferation, migration and invasion
As MIR155HG is the host gene of
miR-155-5p, we further investigated whether
miR-155-5p exerts the same effects as MIR155HG on
LSCC. The expression of miR-155-5p was found to be
significantly increased in the LSCC tissues compared with the
adjacent normal tissues (Fig. 4A).
The high expression level of miR-155-5p was found to be
closely associated with the TNM stage, lymph node metastasis and
poor differentiation. However, there was no association between the
expression of miR-155-5p and age, alcohol consumption, or
smoking (Fig. 4B).
To further determine whether miR-155-5p is
associated with the progression of LSCC, miR-155-5p mimics
and inhibitor were transfected into the TU177 and AMC-HN-8 cells
(Fig. 4C). As shown in Fig. 4D-F, the overexpression of
miR-155-5p markedly promoted the proliferation, migration
and invasion of TU177 and AMC-HN-8 cells, whereas the
downregulation of miR-155-5p markedly inhibited cell
proliferation, migration and invasion. Collectively, these data
suggest that miR-155-5p acts as an oncogenic miRNA by
promoting LSCC cell proliferation, migration and invasion.
Overexpression of miR-155-5p reverses the
inhibitory effects of MIR155HG knockdown on LSCC cell
proliferation, migration and invasion
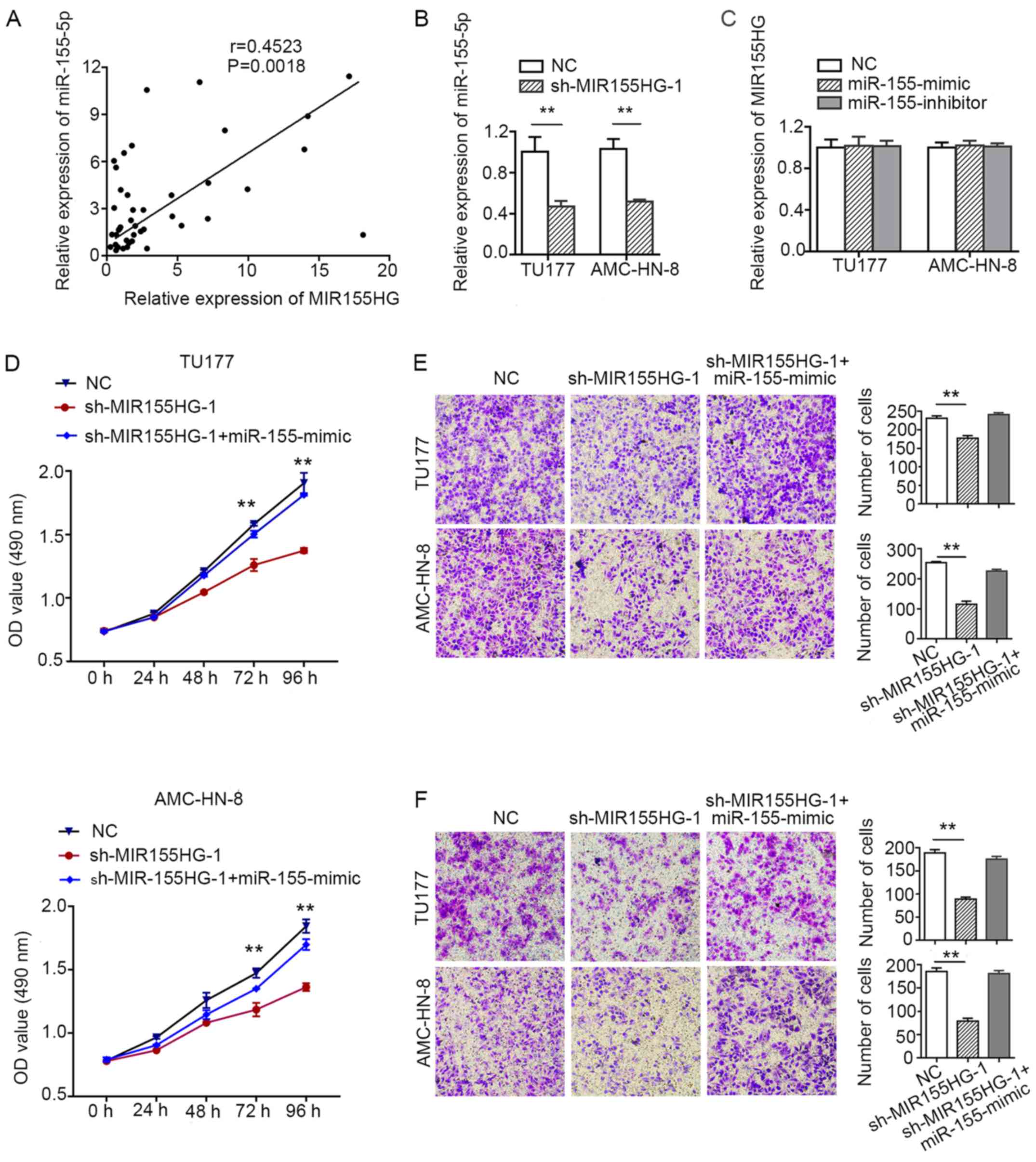
The expression level of miR-155-5p was found
to highly correlate with the expression of MIR155HG
(Fig. 5A). These findings
suggested that miR-155-5p and its host gene,
MIR155HG, may be co-expressed in LSCC. The downregulation of
MIR155HG reduced the expression levels of miR-155-5p
in the TU177 and AMC-HN-8 cells (Fig.
5B). However, the expression of MIR155HG was not
affected by miR-155-5p knockdown or overexpression in LSCC
cells (Fig. 5C). Overexpression of
miR-155-5p may abrogate the inhibitory effect of
MIR155HG knockdown on cell proliferation, migration and
invasion (Fig. 5D-F). These
results indicated that miR-155-5p may act synergistically
with MIR155HG to promote LSCC progression.
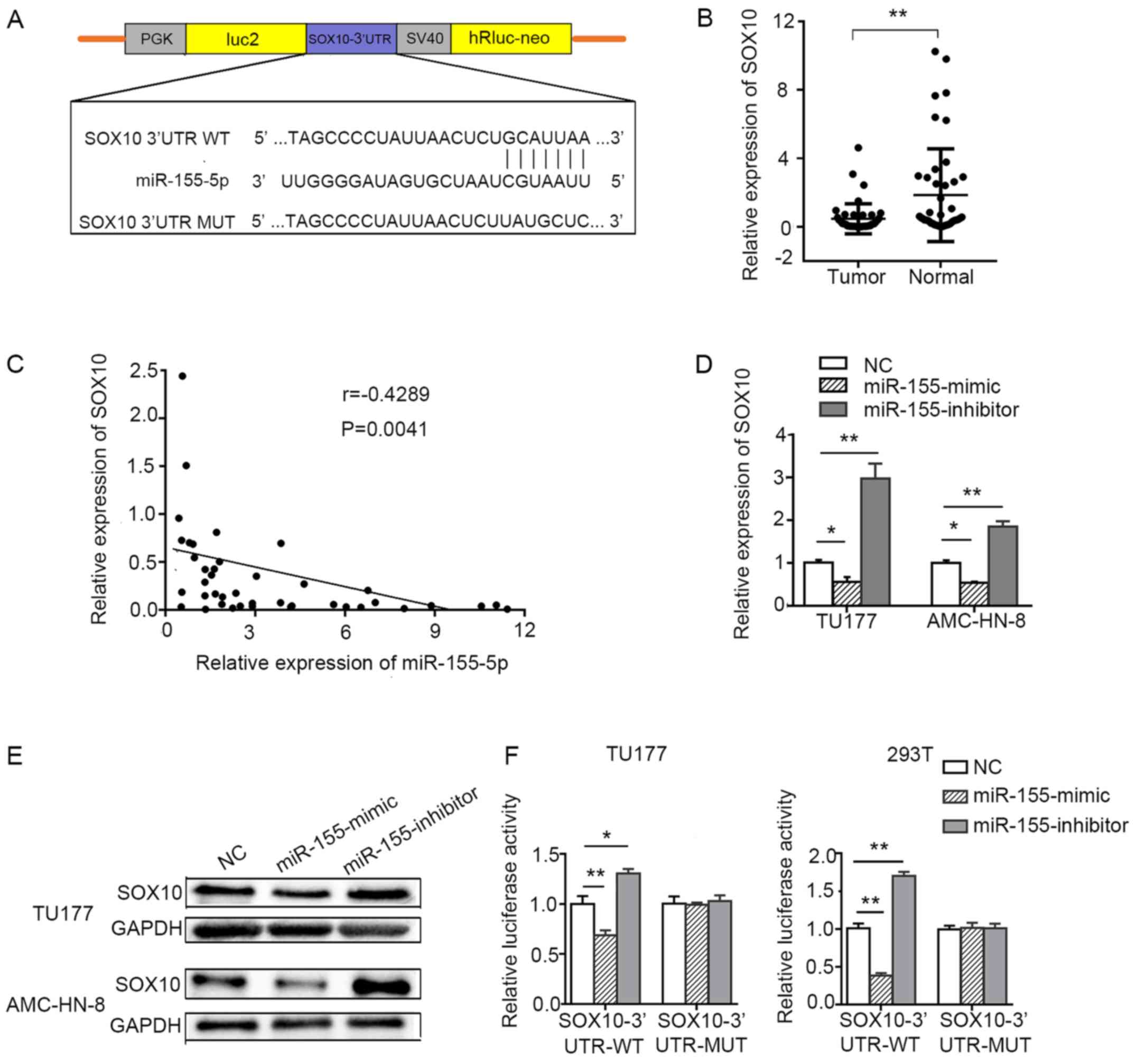
miR-155-5p directly targets SOX10 in LSCC
cells
To further explore the mechanisms underlying the
effects of miR-155-5p on the prolif eration, migration and
invasion of LSCC cells, bioinformatics tools, such as TargetScan
and miRwalk, were used to search for potential target genes of
miR-155-5p. Among these target genes, we focused on
SOX10, a tumor suppressor gene that is involved in
suppressing tumorigenicity (22)
and EMT (23). TargetScan
prediction revealed that the 3′UTR of SOX10 contains a
conserved binding site for miR-155-5p (Fig. 6A). The expression level of
SOX10 was markedly decreased in the LSCC tissues compared
with the corresponding adjacent non-tumor tissues (Fig. 6B). The expression level of
miR-155-5p negatively correlated with the expression of
SOX10 in LSCC (Fig. 6C).
The downregulation of miR-155-5p by transfection with
miR-155-5p inhibitor markedly increased the transcriptional
level and protein expression of SOX10, while the
overexpression of miR-155-5p significantly decreased the
transcriptional level and protein expression of SOX10 in
LSCC cell lines (Fig. 6D and E).
To confirm whether the SOX10 mRNA expression is regulated by
miR-155-5p through direct binding to the 3′UTR of
SOX10, a dual luciferase reporter assay was performed in the
TU177 and 293T cells. The luciferase signal was reduced in the
miR-155-5p mimic and pmirGLO-SOX10-3′UTR-wild-type
co-transfected cells compared with the control group. However,
co-transfection of miR-155 mimic and
pmirGLO-SOX10-3′UTR-mutant-type did not reduce the luciferase
signal compared with the control group. The luciferase signal was
increased in the miR-155-5p inhibitor and
pmirGLO-SOX10-3′UTR-wild-type, but not the mutant-type,
co-transfected cells compared with the control group (Fig. 6F).
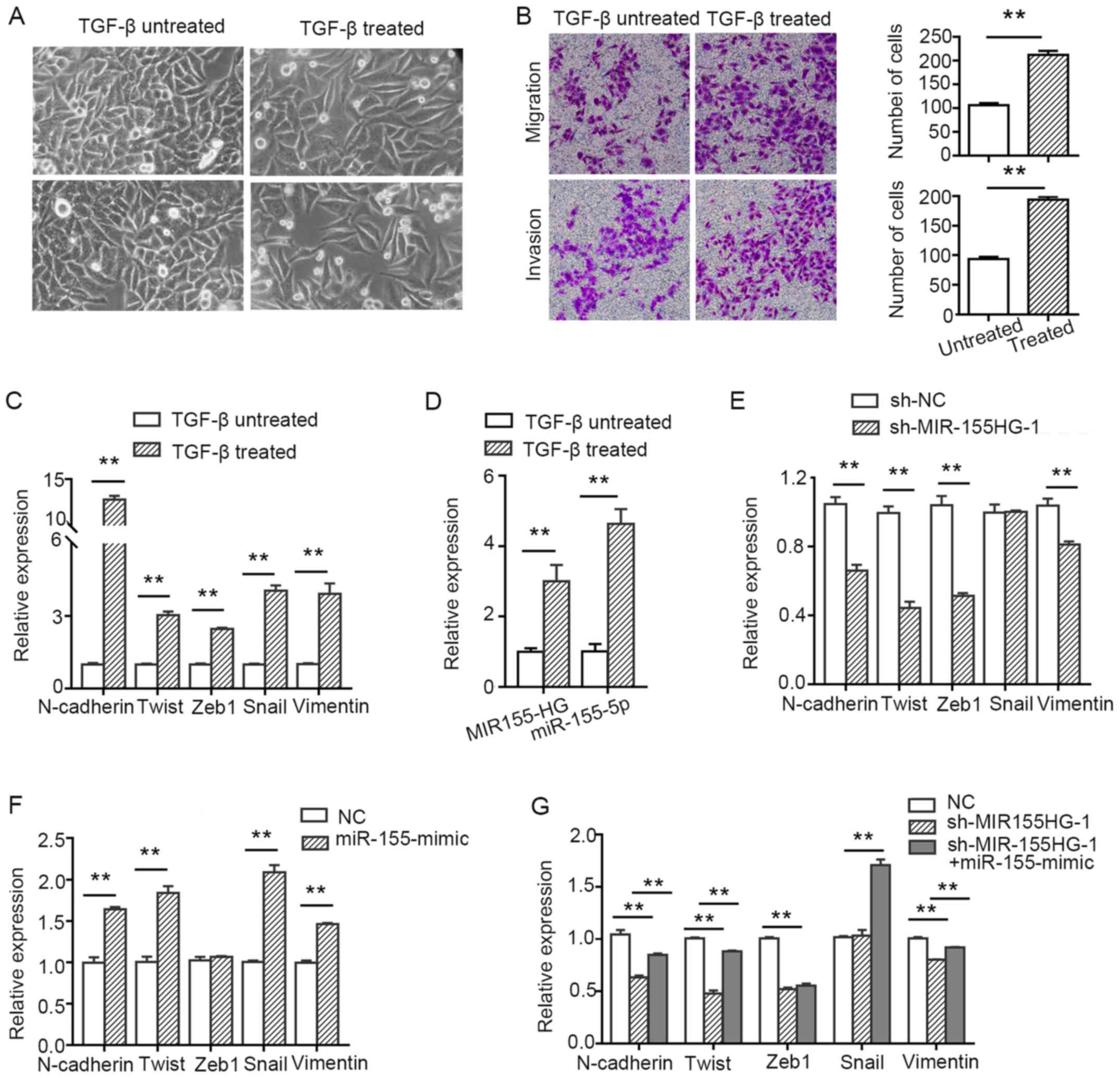
MIR155HG and miR-155-5p are upregulated
in TGF-β-induced TU177 cells and synergistically contribute to the
process of EMT
To investigate whether MIR155HG and
miR-155-5p regulate the migration and invasion of LSCC cells
via EMT, the TU177 cells were treated with 10 ng/ml of TGF-β
for 7 days. The cells treated with TGF-β exhibited a change
in morphology and acquired a spindle-shaped morphology (Fig. 7A). Furthermore, their migratory and
invasive abilities were enhanced compared with the untreated cells
(Fig. 7B). Subsequently, the
molecular markers of EMT were analyzed by RT-qPCR. The levels of
the mesenchymal markers, N-cadherin, Twist, Zeb1, Snail and
vimentin, were found to be upregulated. However, we did not detect
the expression of the epithelial marker, E-cadherin (after using
several common primers of E-cadherin), possibly due to the fact
that the expression abundance was too low to be monitored (Fig. 7C). These results suggested that the
TU177 cells treated with TGF-β exhibited EMT-related
characteristics. Moreover, to further determine whether
MIR155HG and miR-155-5p were involved in
TGF-β-induced EMT, their expression was detected by RT-qPCR
analysis. The results indicated that the levels of MIR155HG
and miR-155-5p were upregulated after the TU177 cells were
treated with TGF-β (Fig.
7D). The knockdown of MIR155HG inhibited the expression
of the mesenchymal markers, N-cadherin, vimentin, Twist and Zeb1,
but did not affect the expression of Snail (Fig. 7E). miR-155-5p overexpression
promoted the expression of the mesenchymal markers, N-cadherin,
vimentin, Twist and Snail, but did not increase the expression of
Zeb1 (Fig. 7F), whereas it
partially reversed the inhibitory effect exerted by MIR155HG
knockdown (Fig. 7G). Taken
together, the above-mentioned results indicate that MIR155HG
and miR-155-5p are EMT-related non-coding RNAs, and they
synergistically promote EMT in LSCC cells.
Discussion
In recent years, non-coding RNAs, particularly
lncRNAs and miRNAs, have been found to play important roles in the
development and progression of tumors, including LSCC. In our
study, the differential expression of lncRNAs was detected by
microarray assays, and MIR155HG was found to be highly
expressed in LSCC tissues. MIR155HG, also referred to as
B-cell integration cluster (BIC), was first identified as a
novel gene that was transcriptionally activated in avian leukosis
virus-induced lymphomas by the insertion of retrovirus integration
sites in the promoter region (24). MIR155HG has also been found
to be upregulated in pediatric Hodgkin’s lymphoma and Burkitt’s
lymphoma (25-27). The transcription of MIR155HG
is regulated by multiple transcription factors, such as MYB,
NF-ĸB and AP-1 (28-30).
A recent study reported that MIR155HG acts as an oncogene in
glioma, and is associated with prognosis and tumor progression
(17). In the present study, the
expression of MIR155HG was found to be upregulated in LSCC
tissues, suggesting that MIR155HG is a potential oncogene in
LSCC. Moreover, the high expression level of MIR155HG was
associated with a poor differentiation, lymph node metastasis and a
higher TNM stage, suggesting that MIR155HG may be of
clinical value in the assessment of invasion and metastasis,
malignant behavior and prognosis of laryngeal carcinoma.
Furthermore, in vitro and in vivo assays verified the
oncogenic role of MIR155HG in LSCC.
It is well known that lncRNAs exert their effects
through a variety of mechanisms, including serving as the host
genes and regulating the expression of miRNAs (14). miR-155 is located in the
third exon of MIR155HG, and has been reported to play
important roles in a number of solid malignancies (31-33).
The overexpression of miR-155 has been found to promote cell
proliferation and migration through targeting TGFβR2 in
gastric cancer (31).
miR-155-5p has also been found to inhibit the migration and
invasion of colorectal cancer cells by targeting CTHRC1
(32). miR-155-5p has been
found to be upregulated in oral squamous cell carcinoma and to be
associated with metastasis, poor prognosis and EMT progression
(34,35). The aberrant expression of
miR-155 has also been previously observed in LSCC (36-38).
miR-155 acts as an oncogene in LSCC, promoting the growth,
migration and invasion of LSCC cells by regulating SOSC1 and
STAT3 (37). Another study
demonstrated that miR-155 was highly expressed in the plasma
and tissues of patients with LSCC (38). However, the correlation between
miR-155 and its host gene, MIR155HG, in LSCC has not
yet been elucidated, and the other target genes of miR-155
require further investigation. The present study demonstrated that
the expression of miR-155-5p was upregulated in LSCC
tissues, and that the overexpression of miR-155-5p
significantly promoted the growth, migration and invasion of LSCC
cells. Furthermore, the expression level of miR-155-5p was
found to be associated with the malignant phenotype of LSCC, which
was consistent with the findings on MIR155HG. Although a
positive correlation between the expression level of
MIR155HG and miR-155-5p was detected, and
MIR155HG may regulate the expression of miR-155-5p,
miR-155-5p did not affect the expression of MIR155HG,
suggesting that the transcriptional activity of miR-155 is
under the control of MIR155HG, which is consistent with the
findings of other studies on Hodgkin’s lymphoma, Burkitt’s lymphoma
and glioma (17,25,26).
Furthermore, the upregulation of miR-155-5p reversed the
inhibitory effects of MIR155HG knockdown on cell malignant
biological properties, suggesting that these two non-coding RNAs
synergistically control the same biological processes in LSCC.
SOX10, a member of the SOX family,
has been found to be a target gene of miR-155-5p.
SOX10 plays an important role in the formation of the neural
crest and peripheral nervous system, the maturation and
differentiation of Schwann and oligodendrocyte lineage cells, and
the occurrence and development of tumors (39,40).
In recent years, the bidirectional role of SOX10 in
regulating tumor progression has been gradually revealed. For
example, SOX10 has been reported to inhibit the growth and
metastasis of digestive tumors via inhibiting the Wnt/β-catenin
pathway (22). However,
SOX10 has been found to be highly expressed in
nasopharyngeal carcinoma, whereas SOX10 knockdown has been
shown to markedly inhibit nasopharyngeal carcinoma cell
proliferation, migration, invasion and EMT (23,41).
SOX10 acts as an oncogene in hepatocellular carcinoma by
activating Wnt/β-catenin signaling (42). In the present study, SOX10
was found to be downregulated in LSCC tissues. The overexpression
of miR-155-5p reduced the transcriptional and translational
levels of SOX10, whereas miR-155-5p downregulation
exerted the opposite effects. Moreover, the direct target
association between miR-155-5p and SOX10 was proven
by a dual-luciferase reporter assay. These results indicate that
SOX10 is a target gene of miR-155-5p, and that
miR-155-5p exerts oncogenic effects partly via the
regulation of SOX10.
EMT is an important biological process for
malignant tumor cells, characterized by epithelial cells acquiring
mesenchymal characteristics and, thereby, the ability of migration
and invasion, which is a prerequisite for the initiation of tumor
invasion and metastasis (43).
Numerous studies have demonstrated that lncRNAs induced by
TGF-β play important roles in EMT in different types of
cancer (44-46). lncRNA ATB (lncRNA-activated by
TGF-β), a regulator and mediator of the TGF-β
signaling pathway, has been found to play a key role in inducing
EMT and promoting invasion and metastasis in hepatocellular
carcinoma (44). LINC01186,
a downregulated lncRNA in TGF-β-treated lung cancer cells,
has been shown to be regulated by TGF-β/SMAD3 and to
inhibit the malignant biological behavior of lung cancer by
regulating EMT (46). However, the
involvement of lncRNAs in the process of EMT in LSCC is poorly
understood. For this purpose, in this study, the TU177 cells were
treated with TGF-β for the indicated number of days. The
treated cells exhibited a change in morphology to a spindle-shaped
one, and their migratory and invasive abilities were enhanced
compared with the untreated cells. Moreover, the expression levels
of mesenchymal markers were upregulated. However, we could not
detect the expression of the epithelial marker, E-cadherin,
possibly since the increase in its expression was too low to be
monitored. The expression levels of MIR155HG and
miR-155-5p were also upregulated in the TGF-β-treated
group, which indicated that MIR155HG and miR-155-5p
are TGF-β-induced non-coding RNAs. Consistent with our
conclusions, MIR155HG has been reported to be associated
with mesenchymal transition in glioma (17), and miR-155 is an EMT-related
onco-miRNA (47,48) that has been found to be induced by
stimulation of TGF-β in hepatocellular carcinoma cells
(49). Further experiments
indicated that MIR155HG and miR-155-5p may
synergistically affect the expression of EMT-related markers,
thereby promoting EMT. As stated above, SOX10 contributes to
EMT in nasopharyngeal carcinoma. Therefore, we hypothesized that
SOX10 may promote EMT in LSCC. All the above-mentioned
results indicate that MIR155HG and miR-155-5p are
EMT-related non-coding RNAs, and that MIR155HG may promote
EMT in LSCC cells by regulating the miR-155-5p/SOX10
axis.
In conclusion, the present study is, to the best of
our knowledge, the first to confirm the functional promoting role
of MIR155HG and its co-expression with miR-155-5p in
LSCC tumorigenesis. miR-155-5p acts synergistically with
MIR155HG to promote the progression of LSCC, partly by
regulating the downstream target gene, SOX10. Moreover, the
findings of the present study indicate a novel role of
MIR155HG in TGF-β-induced EMT of LSCC cells by
regulating EMT marker expression through the
miR-155/SOX10 axis. In addition, these findings
indicate a novel mechanism underlying the aggressive biological
behavior of LSCC, and the
MIR155HG/miR-155-5p/SOX10 axis may represent a
promising therapeutic target for patients with LSCC.
Supplementary Materials
Funding
The study was supported by grants from the Key
Program of Hebei Natural Science Foundation (no. H2017206391) and
the Project of Clinical Medical TalentTraining and Basic Project
Research Funded by Government (Hebei finance society (2017) no.
46).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors’ contributions
BW conceived and designed the study, modified the
manuscript; WCu collected and analysed the data, designed and
perform the experiments, drafted the manuscript; WM, LZ, WCh and HC
performed the experiments and were involved in the revise of
manuscript; All authors participant in revising manuscript, and
agree with the final manuscript.
Ethics approval and consent to
participate
The use of human tissues specimen was approved by
and carried out according to the guidelines of the Ethics Committee
of the Second Hospital of Hebei Medical University (Hebei, China)
and written informed consent was obtained from all patients. All
animal experiments were performed at the Experimental Animal Center
of the Fourth Hospital of Hebei Medical University according to the
NIH guidelines, and were approved by the Institutional Animal Care
and Use Committee of the Fourth Hospital of Hebei Medical
University.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
LSCC
|
laryngeal squamous cell carcinoma
|
|
lncRNA
|
long non-coding RNA
|
|
MIR155HG
|
miR-155 host gene
|
|
TGF-β
|
transforming growth factor-β
|
|
EMT
|
epithelial-to-mesenchymal
transition
|
|
SOX10
|
SRY-related-HMG-box 10
|
|
3′UTR
|
3′ untranslated region
|
Acknowledgments
Not applicable.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yu Q, Zhang X, Ji C, Yang H, Gao M, Hong S
and Hu G: Survival analysis of laryngeal carcinoma without
laryngectomy, radiotherapy, or chemotherapy. Eur Arch
Otorhinolaryngol. 269:2103–2109. 2012. View Article : Google Scholar
|
|
3
|
Groome PA, O’Sullivan B, Irish JC,
Rothwell DM, Schulze K, Warde PR, Schneider KM, Mackenzie RG,
Hodson DI, Hammond JA, et al: Management and outcome differences in
supraglottic cancer between Ontario, Canada, and the Surveillance,
Epidemiology, and End Results areas of the United States. J Clin
Oncol. 21:496–505. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kaikkonen MU and Adelman K: Emerging roles
of non-coding RNA transcription. Trends Biochem Sci. 43:654–667.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mou K, Liu B, Ding M, Mu X, Han D, Zhou Y
and Wang LJ: lncRNA-ATB functions as a competing endogenous RNA to
promote YAP1 by sponging miR-590-5p in malignant melanoma. Int J
Oncol. 53:1094–1104. 2018.PubMed/NCBI
|
|
8
|
Li X, Zhao X, Yang B, Li Y, Liu T, Pang L,
Fan Z, Ma W, Liu Z and Li Z: Long non-coding RNA HOXD-AS1 promotes
tumor progression and predicts poor prognosis in colorectal cancer.
Int J Oncol. 53:21–32. 2018.PubMed/NCBI
|
|
9
|
Qu Wu T, He L, Tian G, Li L, Zhou L, Jin
H, Ren Q, Wang J, Wang YJ, et al: Regulation of laryngeal squamous
cell cancer progression by the lncRNA H19/miR-148a-3p/DNMT1 axis.
Oncotarget. 7:11553–11566. 2016.PubMed/NCBI
|
|
10
|
Zheng J, Xiao X, Wu C, Huang J, Zhang Y,
Xie M, Zhang M and Zhou L: The role of long non-coding RNA HOTAIR
in the progression and development of laryngeal squamous cell
carcinoma interacting with EZH2. Acta Otolaryngol. 137:90–98. 2017.
View Article : Google Scholar
|
|
11
|
Li D, Feng J, Wu T, Wang Y, Sun Y, Ren J
and Liu M: Long intergenic noncoding RNA HOTAIR is overexpressed
and regulates PTEN methylation in laryngeal squamous cell
carcinoma. Am J Pathol. 182:64–70. 2013. View Article : Google Scholar
|
|
12
|
Wang P, Wu T, Zhou H, Jin Q, He G, Yu H,
Xuan L, Wang X, Tian L, Sun Y, et al: Long noncoding RNA NEAT1
promotes laryngeal squamous cell cancer through regulating
miR-107/CDK6 pathway. J Exp Clin Cancer Res. 35:222016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Z, Wang X, Cao S, Han X, Wang Z,
Zhao X, Liu X, Li G, Pan X and Lei D: The long noncoding RNA TUG1
promotes laryngeal cancer proliferation and migration. Cell Physiol
Biochem. 49:2511–2520. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dhir A, Dhir S, Proudfoot NJ and Jopling
CL: Microprocessor mediates transcriptional termination of long
noncoding RNA transcripts hosting microRNAs. Nat Struct Mol Biol.
22:319–327. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao Lu Y, Liu X, Li Q, Graves-Deal C, Cao
R, Singh Z, Franklin B, Wang JL, Hu JH, et al: lncRNA
MIR100HG-derived miR-100 and miR-125b mediate cetuximab resistance
via Wnt/beta-catenin signaling. Nat Med. 23:1331–1341. 2017.
View Article : Google Scholar
|
|
16
|
Cai P, Li H, Huo W, Zhu H, Xu C, Zang R,
Lv W, Xia Y and Tang W: Aberrant expression of LncRNA-MIR31HG
regulates cell migration and proliferation by affecting miR-31 and
miR-31* in Hirschsprung’s disease. J Cell Biochem. 119:8195–8203.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Wu X, Yu Y, Nie T, Hu E, Wu Q, Zhi W,
Jiang T, Wang K, Lu XX, et al: Blocking MIR155HG/miR-155 axis
inhibits mesenchymal transition in glioma. Neuro Oncol.
19:1195–1205. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Miyazono K, Ehata S and Koinuma D:
Tumor-promoting functions of transforming growth factor-β in
progression of cancer. Ups J Med Sci. 117:143–152. 2012. View Article : Google Scholar :
|
|
19
|
Tian L, Li M, Ge J, Guo Y, Sun Y, Liu M
and Xiao H: MiR-203 is downregulated in laryngeal squamous cell
carcinoma and can suppress proliferation and induce apoptosis of
tumours. Tumour Biol. 35:5953–5963. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang B, Lv K, Chen W, Zhao J, Luo J, Wu J,
Li Z, Qin H, Wong TS, Yang W, et al: miR-375 and miR-205 regulate
the invasion and migration of laryngeal squamous cell carcinoma
synergistically via AKT-mediated EMT. Biomed Res Int.
2016:96527892016. View Article : Google Scholar
|
|
21
|
Luo H, Jiang Y, Ma S, Chang H, Yi C, Cao
H, Gao Y, Guo H, Hou J, Yan J, et al: EZH2 promotes invasion and
metastasis of laryngeal squamous cells carcinoma via
epithelial-mesen-chymal transition through H3K27me3. Biochem
Biophys Res Commun. 479:253–259. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tong X, Li L, Li X, Heng L, Zhong L, Su X,
Rong R, Hu S, Liu W, Jia B, et al: SOX10, a novel
HMG-box-containing tumor suppressor, inhibits growth and metastasis
of digestive cancers by suppressing the Wnt/β-catenin pathway.
Oncotarget. 5:10571–10583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
He P and Jin X: SOX10 induces
epithelial-mesenchymal transition and contributes to nasopharyngeal
carcinoma progression. Biochem Cell Biol. 96:326–331. 2018.
View Article : Google Scholar
|
|
24
|
Tam W, Ben-Yehuda D and Hayward WS: bic, a
novel gene activated by proviral insertions in avian leukosis
virus-induced lymphomas, is likely to function through its
noncoding RNA. Mol Cell Biol. 17:1490–1502. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kluiver J, Poppema S, de Jong D, Blokzijl
T, Harms G, Jacobs S, Kroesen BJ and van den Berg A: BIC and
miR-155 are highly expressed in Hodgkin, primary mediastinal and
diffuse large B cell lymphomas. J Pathol. 207:243–249. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Metzler M, Wilda M, Busch K, Viehmann S
and Borkhardt A: High expression of precursor microRNA-155/BIC RNA
in children with Burkitt lymphoma. Genes Chromosomes Cancer.
39:167–169. 2004. View Article : Google Scholar
|
|
27
|
van den Berg A, Kroesen BJ, Kooistra K, de
Jong D, Briggs J, Blokzijl T, Jacobs S, Kluiver J, Diepstra A,
Maggio E, et al: High expression of B-cell receptor inducible gene
BIC in all subtypes of Hodgkin lymphoma. Genes Chromosomes Cancer.
37:20–28. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thompson RC, Vardinogiannis I and Gilmore
TD: Identification of an NF-κB p50/p65-responsive site in the human
MIR155HG promoter. BMC Mol Biol. 14:242013. View Article : Google Scholar
|
|
29
|
Vargova K, Curik N, Burda P, Basova P,
Kulvait V, Pospisil V, Savvulidi F, Kokavec J, Necas E, Berkova A,
et al: MYB transcriptionally regulates the miR-155 host gene in
chronic lymphocytic leukemia. Blood. 117:3816–3825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Elton TS, Selemon H, Elton SM and
Parinandi NL: Regulation of the MIR155 host gene in physiological
and pathological processes. Gene. 532:1–12. 2013. View Article : Google Scholar
|
|
31
|
Qu Y, Zhang H and Sun W: MicroRNA-155
promotes gastric cancer growth and invasion by negatively
regulating transforming growth factor-beta receptor 2. 109:618–628.
2018.
|
|
32
|
Liu J, Chen Z, Xiang J and Gu X:
MicroRNA-155 acts as a tumor suppressor in colorectal cancer by
targeting CTHRC1 in vitro. Oncol Lett. 15:5561–5568.
2018.PubMed/NCBI
|
|
33
|
Lin J, Chen Y, Liu L, Shen A and Zheng W:
MicroRNA-155-5p suppresses the migration and invasion of lung
adenocarcinoma A549 cells by targeting Smad2. Oncol Lett.
16:2444–2452. 2018.PubMed/NCBI
|
|
34
|
Baba O, Hasegawa S, Nagai H, Uchida F,
Yamatoji M, Kanno NI, Yamagata K, Sakai S, Yanagawa T and Bukawa H:
MicroRNA-155-5p is associated with oral squamous cell carcinoma
metastasis and poor prognosis. J Oral Pathol Med. 45:248–55. 2016.
View Article : Google Scholar
|
|
35
|
Kim H, Yang JM, Ahn SH, Jeong WJ, Chung JH
and Paik JH: Potential oncogenic role and prognostic implication of
MicroRNA-155-5p in oral squamous cell carcinoma. Anticancer Res.
38:5193–5200. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao X, Zhang W and Ji W: YB-1 promotes
laryngeal squamous cell carcinoma progression by inducing miR-155
expression via c-Myb. Future Oncol. 14:1579–1589. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao XD, Zhang W, Liang HJ and Ji WY:
Overexpression of miR - 155 promotes proliferation and invasion of
human laryngeal squamous cell carcinoma via targeting SOCS1 and
STAT3. PLoS One. 8:e563952013. View Article : Google Scholar
|
|
38
|
Wang JL, Wang X, Yang D and Shi WJ: The
Expression of MicroRNA-155 in plasma and tissue is matched in human
laryngeal squamous cell carcinoma. Yonsei Med J. 57:298–305. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hong CS and Saint-Jeannet JP: Sox proteins
and neural crest development. Semin Cell Dev Biol. 16:694–703.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mokhtarzadeh Khanghahi A, Satarian L, Deng
W, Baharvand H and Javan M: In vivo conversion of astrocytes into
oligodendrocyte lineage cells with transcription factor Sox10;
Promise for myelin repair in multiple sclerosis. PLoS One.
13:e02037852018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao Y, Liu ZG, Tang J, Zou RF, Chen XY,
Jiang GM, Qiu YF and Wang H: High expression of Sox10 correlates
with tumor aggressiveness and poor prognosis in human
nasopharyngeal carcinoma. OncoTargets Ther. 9:1671–1677. 2016.
View Article : Google Scholar
|
|
42
|
Zhou D, Bai F, Zhang X, Hu M, Zhao G, Zhao
Z and Liu R: SOX10 is a novel oncogene in hepatocellular carcinoma
through Wnt/β-catenin/TCF4 cascade. Tumour Biol. 35:9935–9940.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: Acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepato-cellular carcinoma. Cancer Cell. 25:666–681. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lu Z, Li Y, Che Y, Huang J, Sun S, Mao S,
Lei Y, Li N, Sun N and He J: The TGFβ-induced lncRNA TBILA promotes
non-small cell lung cancer progression in vitro and in vivo via
cis-regulating HGAL and activating S100A7/JAB1 signaling. Cancer
Lett. 432:156–168. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hao Y, Yang X, Zhang D, Luo J and Chen R:
Long noncoding RNA LINC01186, regulated by TGF-β/SMAD3, inhibits
migration and invasion through Epithelial-Mesenchymal-Transition in
lung cancer. Gene. 608:1–12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Johansson J, Berg T, Kurzejamska E, Pang
MF, Tabor V, Jansson M, Roswall P, Pietras K, Sund M, Religa P, et
al: MiR-155-mediated loss of C/EBPβ shifts the TGF-β response from
growth inhibition to epithelial-mesenchymal transition, invasion
and metastasis in breast cancer. Oncogene. 32:5614–5624. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li DP, Fan J, Wu YJ, Xie YF, Zha JM and
Zhou XM: MiR-155 up-regulated by TGF-β promotes
epithelial-mesenchymal transition, invasion and metastasis of human
hepatocellular carcinoma cells in vitro. Am J Transl Res.
9:2956–2965. 2017.
|
|
49
|
Kong X, Liu F and Gao J: MiR-155 promotes
epithelial-mesenchymal transition in hepatocellular carcinoma cells
through the activation of PI3K/SGK3/β-catenin signaling pathways.
Oncotarget. 7:66051–66060. 2016. View Article : Google Scholar : PubMed/NCBI
|