Introduction
Lung cancer has been recognized as the most serious
human disease worldwide (1). Based
on the online database GLOBOCAN, lung cancer was the most commonly
diagnosed and severest cancer all over the world in 2013,
accounting for 13% of all the diagnosed cancers and 19.4% of all
cancer-associated mortalities in 184 nations and regions (2). Nearly 85% of all lung cancer cases
are non-small cell lung cancer (NSCLC) cases, including
bronchioloalveolar cancer, large cell cancer, adenocarcinoma and
squamous cell cancer (2). Over 50%
of patients with NSCLC developed metastasis following diagnosis,
with only 14% surviving for at least 5 years (3,4).
Chemotherapy resistance, aggressive tumor growth and distant
metastasis are the primary contributors to NSCLC-associated
mortality worldwide (3,4).
Adenylyl cyclase-associated protein 1 (CAP1) is an
actin monomer-binding protein coded by a gene termed CAP1
(5), which was initially cloned in
budding yeast and is located downstream of RAS (6). The human homolog of CAP1 was first
identified in the early 1990s (7).
CAPs are associated with actin in various organisms ranging from
yeast to mammals (8). In addition,
CAP1 has been demonstrated to serve a crucial role in accelerating
actin filament turnover (9). Based
on previous findings demonstrating the crucial roles of actin
filaments in regulating CAP1 expression and cell migration
(10,11), studies further confirmed the close
association between CAP1 expression and tumor metastasis (12-14).
Previous findings by our group indicated that CAP1 expression is
upregulated in patients with NSCLC compared with that in healthy
individuals (15-17). Furthermore, CAP1 expression was
demonstrated to be upregulated in patients with metastatic NSCLC
compared with that in patients with non-metastatic NSCLC (15,16).
Therefore, CAP1 expression could be used to predict metastasis in
and the prognosis of patients with (18).
RNA inference (i)-mediated gene-silencing in mammals
can effectively inhibit gene expression at transcriptional,
post-transcriptional and translational levels (19). RNAi has been recently employed for
the selective interference of specific gene expression via the
introduction of artificially synthesized small interfering (si)RNAs
(19). However, the entry of
siRNAs into the target cells is barely feasible without a suitable
carrier (20). Nanoparticles (NPs)
have demonstrated good performances in the delivery of siRNAs to
silence key genes and inhibit the progression of disease in animal
models, thereby highlighting their potential application in human
clinical trials (20-22). Additionally,
poly(lactic-co-glycolic acid; PLGA) is an ideal non-toxic and
non-immunostimulatory vehicle for delivering siRNAs (23-25).
Materials and methods
Materials
The following reagents were used in the present
study: Poly(lactide-co-glycolide) Resomer RG502 [PLGA-COOH;
molecular weight (MW), 20,000; Jinan Daigang Biomaterial Co., Ltd.,
Jinan, China], A549 and H1299 cells (Cell Bank of the Chinese
Academy of Sciences), RPMI-1640 medium, fetal bovine serum (both
Gibco; Thermo Fisher Scientific, Inc.), Cell Counting Kit (CCK)-8
(Dojindo Molecular Technologies, Inc.), amine-poly(ethylene
glycol)-carboxymethyl (NH2-PEG-COOH; MW, 3,400) was
purchased from Seebio Biotech (Shanghai) Co., Ltd. Triethylamine
and dichloromethane were purchased from Sinopharm Chemical Reagent
Co., Ltd.. All chemical reagents were analytical grade or above.
Nude mice were obtained from the Experimental Animal Center of
Shanghai Tenth People's Hospital of Tongji University (Shanghai,
China).
Synthesis of PLGA-PEG macromolecule
Triethylamine and dichloromethane were dried with
calcium hydride before use as described previously (26). Carboxylate-functionalized copolymer
PLGA-PEG was synthesized by the conjugation of PLGA-COOH and
NH2-PEG-COOH. Briefly, 200 mg PLGA-COOH, 2.9 mg
1-(3-Dimethylaminopropyl)-3-ethyl-carbodiimide hydrochloride
(Sigma-Aldrich; Merck KGaA), 1.8 mg N-Hydroxysuccinimide (NHS;
Sinopharm Chemical Reagent Co., Ltd.) and 1 mg
4-dimethylaminopyridine (Sino-pharm Chemical Reagent Co., Ltd.)
were dissolved in 10 ml dichloromethane, and stirred for 24 h at
room temperature to convert PLGA-COOH to PLGA-NHS. Then, 34 mg
NH2-PEG-COOH and 8.6 µl triethylamine were added
into the solution. After further stirring for 8 h at room
temperature, dichloromethane was removed through the use of a
rotary evaporator and 5 ml N,N-Dimethylformamide (Sino-pharm
Chemical Reagent Co., Ltd.) was added to dissolve PLGA-PEG. Then,
the solution was added to a dialysis bag (width, 44 mm; MW cut-off,
8,000-14,000); unreacted PEG and other small molecules were removed
by dialyzing for 3 days, and then PLGA-PEG was freeze-dried for
24-48 h.
Preparation of PLGA/siCAP1 NPs
PLGA/siCAP1 NPs were prepared by the revised solvent
emulsion/evaporation method to obtain capsules with a polymeric
shell that encapsulates siCAP1 (Shanghai GenePharma Co., Ltd.,
Shanghai, China) as previously described (27). The sequence for the siCAP1 was
5′-GGAGCCAGCUGUACUUGAATT-3′ and reverse,
5′-UUCAAGUACAGCUGGCUCCTT-3′. The sequences of negative control
siRNA were: 5′-UUCUCCGAACGUGU-CACGUTT-3′ (sense) and
5′-ACGUGACACGUUCGGAGA ATT-3′ (antisense).
Briefly, 10 mg PLGA-PEG was dissolved in a solvent
mixture containing 200 µl dichloromethane, 200 µl 15%
(w/v) Span80 dichloromethane (Shenzhen Sheng Xin Automation
Equipment Co., Ltd., Shenzhen, China) along with 16 µg
siCAP1. Following a thorough mixing, the resulting mixture was
emulsified using T 10 basic ULTRA-TURRAX® (IKA England
Ltd.) to produce a pre-emulsion with a final volume of 2 ml. A
15-ml centrifuge tube was used to emulsify the solution on ice for
1 min. Subsequently, an ultrasonic homogenizer/probe sonicator with
a vibrating metallic tip (model no. JY 92-IIN; tip diameter, 3 mm;
Ningbo Scientz Biotechnology Co., Ltd.) was used to sonicate the
emulsion on ice for 3 sec, followed by a 10 sec of pause; the
process was repeated several times for 3 min each under 50% power
(300 W). After an additional 10 min of stirring (cycle, 1 min of
stirring and a 1-min pause) using T 10 basic ULTRA-TURRAX on ice,
the organic solvents were subjected to 3 h of evaporation with
magnetic stirring at room temperature (27-30).
To contrast the PLGA-PEG/siCAP1 NPs (PLGA/siCAP1), a PLGA/negative
control siRNA (Ctrl; Shanghai GenePharma Co., Ltd.), a PLGA
emulsion alone and PLGA-fluorescein isothiocyanate (FITC; Shanghai
GenePharma Co., Ltd.) were prepared using the same process. In
addition, PLGA mixed with PLGA-FITC [1:1 (w/w)] was used to prepare
the emulsion for in vivo bio-distribution imaging.
siCAP1 binding and gel retardation
assay
Increasing amounts of PLGA (w/w) were complexed to a
fixed amount of siCAP1 (500 ng) in OptiMEM I reduced serum culture
medium (Gibco; Thermo Fisher Scientific, Inc.) for 20 min at room
temperature in order to determine the amount of PLGA needed to
fully self-assemble with siCAP1. Then, the complexes were
electrophoresed and analyzed. Samples of PLGA/siCAP1 (500 ng/lane)
were loaded in a 4% agarose gel. The bands on the gel were observed
using the UV imaging system (model no. TA-9403; Tianan United
Technology Co.).
Characterization of nanoparticles
The sizes of the prepared particles were measured
using a Malvern Zetasizer Nano ZS (Malvern Instruments Ltd.) based
on dynamic light scattering. The morphologies of the probes were
monitored by transmission electron microscopy on a JEOL-2100F
instrument (JEOL) under 200 kV of accelerating voltage.
Cell cytotoxicity assays
A549 and H1299 cells were seeded into 96-well plates
at 5,000 cells/well. The cells were incubated in 100 µl
Roswell Park Memorial Institute (RPMI)-1640 culture medium
containing 1% antibiotics (penicillin-streptomycin; Gibco; Thermo
Fisher Scientific, Inc.) and 10% Foundation™ fetal bovine serum
(FBS; Gemini Bio-Products, Inc.) for 24 h at 37°C in a humidified
atmosphere containing 5% CO2. Then, cells were incubated
in RPMI-1640 culture medium containing 600 µg PLGA/siCAP1,
PLGA/Ctrl, PLGA or left untreated in a triple gas incubator for 4 h
at 37°C. The supernatant was removed after 24 and 48 h, and cell
viability was determined by performing the CCK-8 assay. The
released formazan product was detected with an Epoch Microplate
Spectrometer (BioTek Instruments, Inc.) at an absorbance wavelength
of 490 nM.
siRNA transfection
A549 cells were untransfected or transfected with
PLGA/siCAP1, PLGA, siCAP1 and PLGA/Ctrl in 6-well plates. Cells
were seeded into 6-well culture plates (5×104) and the
following day, the cells were transfected. In the PLGA/siCAP1
group, cells were transfected with PLGA/siCAP1 concentrations
[100:1-800:1 (w/w)] with siCAP1 at 50 nM. Cells in the PLGA/Ctrl
group were transfected with PLGA/Ctrl with Ctrl at 50 nM, while
those in the siCAP1 group were transfected with 50 nM siCAP1 using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Cells
in the PLGA group were transfected with 50 nM PLGA. After 4-6 h of
transfection at 37°C, the medium was replaced with fresh RPMI-1640
medium and after a 24-h transfection at 37°C, the cells were washed
three times in warm PBS and harvested; total RNA or protein was
then extracted. Cells incubated in culture media were used as the
untreated group.
Western blot analysis
Transfected cells were washed with PBS, harvested
and lysed in radioimmunoprecipitation assay buffer with a protease
inhibitor cocktail (Roche Applied Science) to isolate total cell
proteins. Then, the concentration of protein samples was determined
with a Pierce BCA Protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Equal amounts of protein (30 µg/lane)
were separated by SDS-PAGE on a 10% gel and then transferred to
nitrocellulose membrane (EMD Millipore). Following blocking with 5%
non-fat milk for 1 h at room temperature, the membranes were
incubated overnight at 4°C with primary antibodies against GAPDH
(polyclonal, rabbit anti-human IgG; cat. no. ab9485) and CAP1
(rabbit anti-human IgG; cat. no. ab155079; both 1:1,000; Abcam).
After washing the membrane with 1X PBS containing 0.05% Tween-20
three times, it were incubated at room temperature for 1 h with
either horseradish peroxidase-conjugated goat anti-rabbit (cat. no.
ZB-2301) or goat anti-mouse (cat. no. ZB-2305; both 1:2,000;
OriGene Technologies, Inc.) secondary antibodies. The
immunoreactive signals for CAP1 were visualized using the enhanced
chemiluminescence system from Amersham (GE Healthcare) and
subjected to densitometric analyses using ImageJ software (version
1.51; National Institutes of Health). Relative protein expression
levels were normalized to GAPDH.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
CAP1 mRNA expression in transfected A549 cells was
determined by RT-qPCR. Total RNA was isolated from freshly cultured
cells ground in liquid nitrogen was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and 1 µg of
total RNA was reverse transcribed into cDNA using a QuantiNova
Reverse Transcription kit (Qiagen, Inc.). cDNA templates were then
mixed with SYBR Green PCR Master Mix (Toyobo Life Science) and the
corresponding primers; qPCR was subsequently performed on the ABI
7500 Real-time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were set to 95°C
for 5 min, followed by 40 cycles of 95°C for 10 sec and 60°C for 30
sec. The subsequent dissociation stage was set to 95°C for 15 sec,
60°C for 30 sec and 95°C for 15 sec. Primers were synthesized by
Shanghai GenePharma Co., Ltd. The reference gene, GAPDH, was
amplified using the following primers: forward, 5′-ACT CGC TGC TTG
CTG GTC-3′ and reverse, 5′-ATG GGT GCC AAC AAA TCG-3′. The test
gene, CAP1, was amplified using the following primers: forward,
5′-GAT TTG TTG GCA CCC ATC TC-3′ and reverse, 3′-GCC TGG ATA CTT
TCG CTG AC-5′. Data analysis was conducted using the
2−ΔΔCq method (31) and
all RT-qPCR reactions were performed in triplicate.
NSCLC mouse model
The NSCLC mouse model was established through the
seeding of A549 cells (5×106) under the skins of the
right limbs of 20 five-week-old male nude mice (16-20 g) as
previously described (21). All
mice were kept under specific pathogen-free conditions, at room
temperature (20-26°C) with a humidity level of 50-60% and a 12-h
light/dark cycle. Mice received 5 g food and 100 ml water per 100 g
body weight per day. Mice were anesthetized with isoflurane mixed
with O2. Induction of anesthesia required 2% isoflurane
using Rodent Anesthesia Machine (Shanghai Yuyan Instruments Co.,
Ltd.). The skin of the mice were disinfected with 70% alcohol. A549
cells (5×106) were suspended in 30 µl RPMI-1640
medium. A 29-gauge needle connected to a 0.5-ml syringe containing
the A549 cells was directly injected under the skin to a depth ~3
mm. Following confirming their stabilization, the mice were
returned to their cages. After 14 days, tumors under the skin were
confirmed and the growth was measured every 3 days.
To determine whether PLGA could deliver a sufficient
amount of siRNAs to the developing tumors, mice were administered
with PLGA/siCAP1 [600:1 (w/w); 1 µg siCAP1] via the tail
vein. The mice injected with fluorescent siCAP1s alone or PBS were
used as controls. A total of 4 h after the injections, ex
vivo fluorescent intensities were measured to evaluate the
delivery performances, then mice were sacrificed and dissected. The
tumor tissues were fixed, cut into slices and stained. Tumor
sections were fixed with 10% formalin at room temperature for 24 h,
dehydrated with different concentrations of ethanol (70, 85, 95 and
100%) and xylene, embedded in paraffin and sliced into
4-µm-thick sections. Following deparaffinization in xylene,
and rehydration in 100, 95, 85 and 70% ethanol and distilled water,
these sections were stained with hematoxylin for 5 min at room
temperature and eosin for 2 min at room temperature. They were then
dehydrated with 70, 85, 95 and 100% ethanol and xylene, and covered
with a coverslip and mounting medium (Sangon Biotech Co., Ltd.).
Then they were observed under an inverted light microscope (Nikon
Eclipse TS 100; Nikon Corporation) at a magnification of ×200.
FITC-labeled PLGA/siCAP1 was kept in refrigerator of 4°C and in the
dark. All experimental protocols were evaluated and approved by the
Institutional Review Board for Animal Research at Shanghai Tenth
People's Hospital (Shanghai, China).
Transwell assay
For the cell migration assays, chambers with the
upper and lower wells were used for cell incubation. Transfected
A549 cells (3×104) in serum-free RPMI-1640 medium were
seeded in the upper chambers, which were equipped with 8-µm
Transwell membranes (Corning Incorporated, Corning, NY, USA), and
500 µl RPMI-1640 medium supplemented with 1% antibiotics and
10% FBS was added to the lower chambers. After 24 h of incubation
at a constant temperature of 37°C, cells were removed from the
upper surface using a cotton tip and the chambers were washed
thrice with 1X PBS. Next, the cells that migrated to lower surface
of the chamber were fixed with 95% ethanol for 10 min at room
temperature, stained with 0.1% crystal violet solution for 10 min
at room temperature, washed thrice in PBS and subsequently
air-dried. Then, a light microscope was used to photograph and
count these cells in five random fields per well (magnification,
×100). Cells were counted using ImageJ software (version 1.51).
Wound healing assay
Cell migration was evaluated by conducting Transwell
assays and wound healing assays. A549 cells were cultured in 6-well
plates until reaching 95% confluence. Cells were scratched using a
sterile 10-µl pipette tip. Then, wound healing was observed
and photographed after 0 and 24 h using a fluorescent microscope at
a magnification of ×200. The gap width was quantified with ImageJ
software (version 1.51).
Tumor metastasis model
A total of 20 nude mice were randomly assigned into
the following four equal groups: Untreated (the blank control
group, mice injected with A549 cells), PLGA/Ctrl [the negative
control group, mice injected with PLGA/Ctrl-treated A549 cells],
PLGA (the control group, mice injected with PLGA-treated A549
cells) and PLGA/siCAP1 (the gene silencing group, mice injected
with PLGA/siCAP1-treated A549 cells). Each mouse was
intraperitoneally injected with 5×106 cells and observed
daily. All mice were sacrificed after 14 days and the tumor weights
were measured. The nude mice were sacrificed and their abdominal
cavities were opened to observe the growth of tumors.
Intraperitoneal tumor formation in the internal organs, including
the peritoneum, heart, liver, spleen, lung, kidney, intestine,
uterus, ovary, diaphragm and omentum, were evaluated in terms of
location, the number of tumors, and size of the tumors in the nude
mice. The number of tumors with a diameter of >5 mm was defined
as the tumor number. The number of internal organs with tumor
formation was defined as the tumor position. The tumor formation
rate was calculated as follows: Tumor formation rate = (number of
nude mice with a tumor diameter >5 mm / total number of nude
mice in the experimental group) x 100%.
Hematoxylin and eosin (H&E) and
immunohisto-chemical (IHC) assay
Tumor sections were fixed with 10% formalin at room
temperature for 24 h and dehydrated with 70, 85, 95 and 100%
ethanol and xylene. Then they were embedded in paraffin and sliced
into 4-µm-thick sections. Following deparaffinization in
xylene and rehydration 100, 95, 85 and 70% ethanol and distilled
water. These sections were stained with hematoxylin for 5 min at
room temperature and eosin for 2 min at room temperature,
dehydrated with 70, 85, 95 and 100% ethanol and xylene, and covered
with a coverslip and mounting medium (Sangon Biotech Co., Ltd.).
The H&E sections were observed under an inverted light
microscope (Nikon Eclipse TS 100; Nikon Corporation) at a
magnification of ×200 to identify any cancer nests.
For IHC, tissues were fixed in 10% formalin at room
temperature for 24 h and dehydrated with 70, 85, 95 and 100%
ethanol and xylene. Then they were embedded in paraffin and sliced
into 4-µm-thick sections. Deparaffinization was performed in
xylene and rehydration in 100, 95, 85 and 70% ethanol and distilled
water. Then, the sections were soaked in 0.3%
H2O2 to block the activities of endogenous
peroxidases. After washing the sections with 10 mmol/l citrate
buffer (pH 6.0), they were autoclaved for 20 min at 121°C for
antigen retrieval. Then, the sections were washed with PBS (pH 7.2)
and subsequently incubated with 10% goat serum (Sangon Biotech Co.,
Ltd.) at room temperature for 1 h to prevent the occurrence of
non-specific reactions. The sections were incubated overnight with
anti-CAP1 antibodies (1:500) at a constant temperature of 4°C. The
monoclonal mouse anti-CAP1 antibodies were constructed and provided
by Professor Jeffrey Field, Department of Systems Pharmacology and
Translational Therapeutics, Perelman School of Medicine, University
of Pennsylvania School of Medicine (Philadelphia, PA, USA). All
sections were processed using the peroxidase-anti-peroxidase method
(Dako; Agilent Technologies GmbH) (32). The sections were washed with PBS,
then incubated in 3,3′-diaminobenzidine tetrahydrochloride hydrate
in 0.05 mol/l Tris buffer (pH 7.6) containing 0.03%
H2O2 to observe the reaction of peroxidases.
Next, the sections were washed with sterile water, counterstained
with hematoxylin, dehydrated and added to a cover slip. The
counterstained sections were then observed under an inverted light
microscope (Nikon Eclipse TS 100) at a magnification of ×200. The
sections were observed in ≥10 random high-power fields and ≥400
cells in each field were counted.
Statistical analysis
Each experiment was conducted at least in
triplicate. All experimental data are presented as mean ± standard
deviation from at least three independent experiments. One-way
analysis of variance with Dunnett's post hoc test was used to
analyse the differences among the groups. Statistical analysis was
carried out using the SPSS 23.0 software package (IBM Corp.).
P≤0.05 indicated that the difference between groups was
statistically significant.
Results and Discussion
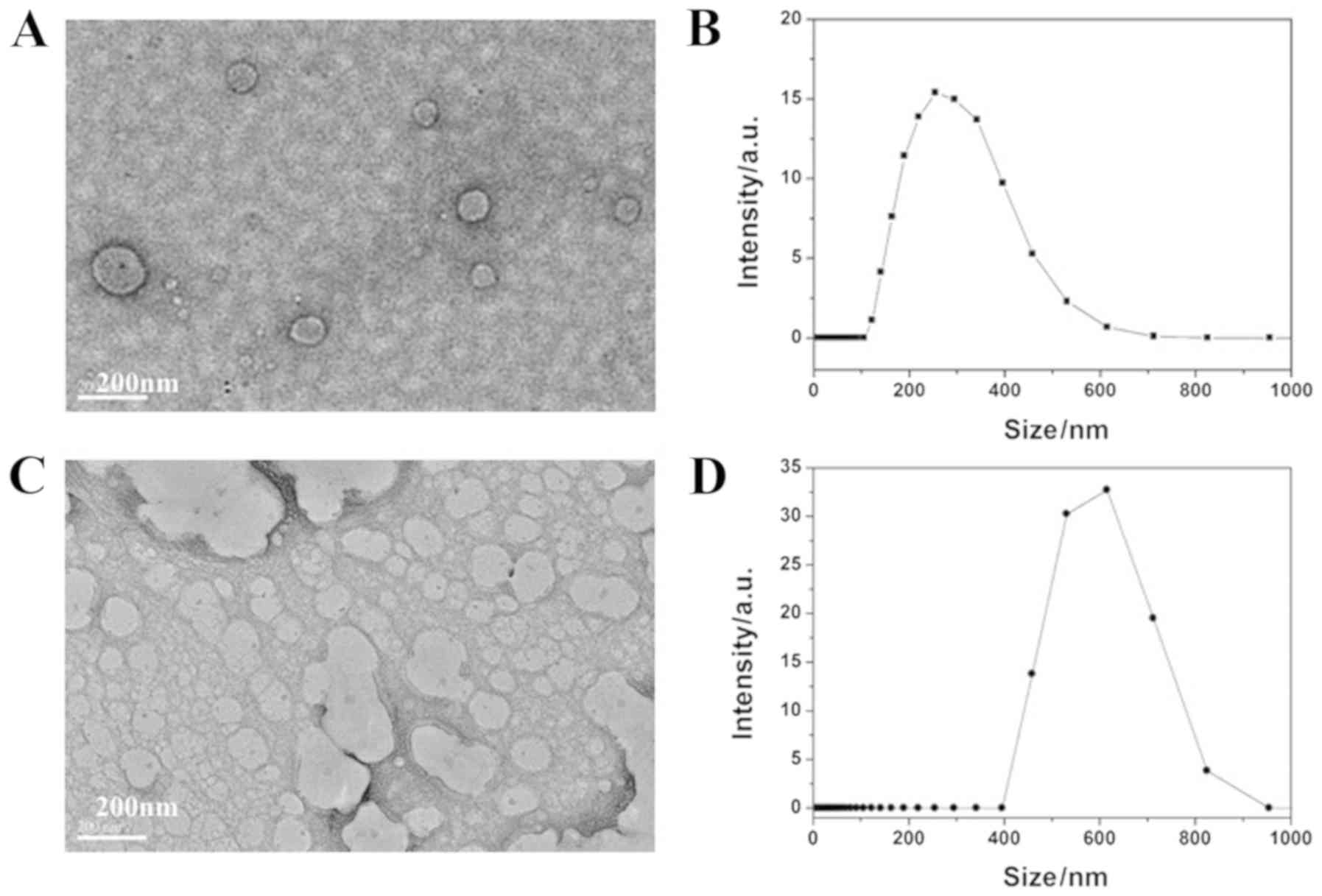
PLGA/siCAP1 NPs are larger than PLGA NPs
and reduce CAP1 mRNA expression
The morphology and structure of PLGA/siCAP1 emulsion
were characterized. The average hydrodynamic size of the PLGA
emulsion in deionized water was ~200 nm (Fig. 1A and B). The average hydrodynamic
size of the PLGA/siCAP1 emulsion in deionized water was ~600 nm
(Fig. 1C and D). These results
indicate that the addition of siCAP1 increased the emulsion size
and that siCAP1 was successfully encapsulated into the PLGA
polymers. The expected products were successfully obtained and were
further analyzed to investigate their effects on A549 cells.
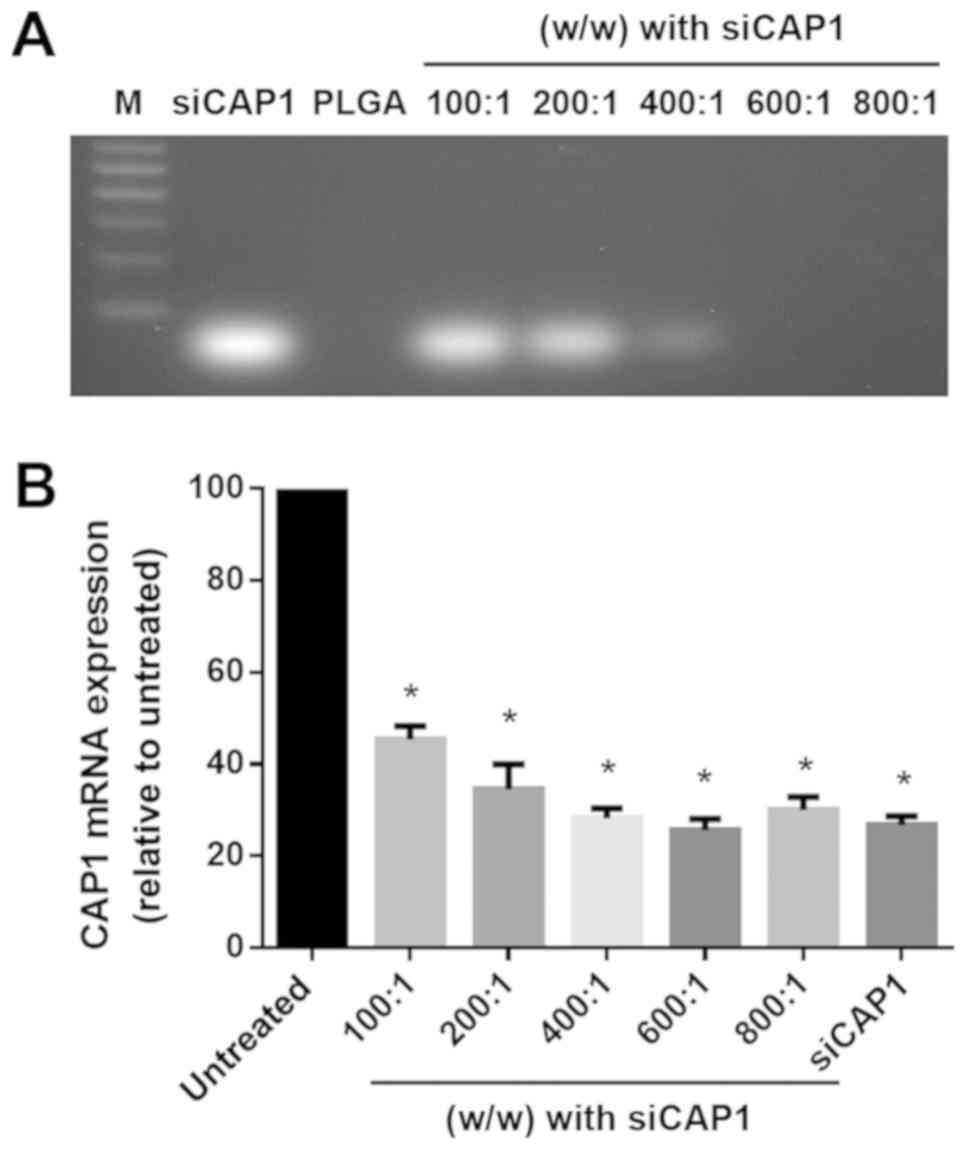
Agarose gel electrophoresis was conducted to
determine the amount of PLGAs needed for full self-assembly with
siRNAs mediated by electrostatic interactions. The migration of
non-complexed siRNAs was observed at the bottom of the gel as
presented in Fig. 2A. Increasing
the amount of PLGA complexed to siCAP1 [100:1-800:1 (w/w)] was
demonstrated to prevent the migration of the non-complexed siCAP1.
These results indicated that increasing the amount of PLGAs to the
range of 600:1-800:1 (w/w) promoted the full complex formation of
the PLGAs with the siCAP1 (Fig.
2A). As presented in Fig. 2B,
an increasing mass of PLGA [100:1-800:1 (w/w); 45.40±1.664,
34.53±3.133, 28.23±1.220, 25.60±1.389, 30.07±1.602, 26.73±1.132,
respectively; P<0.001] combined with a constant mass of siCAP1
(50 nM) was used to transfect A549 cells to investigate whether or
not the varying ratios of PLGA influenced gene silencing. Cells in
the control group were treated with culture medium, while other
A549 cells were treated with siCAP1 complexed to PLGA or
Lipofectamine 2000 (L2K). PLGA/siCAP1 in each selected ratio
[100:1-800:1 (w/w)] was demonstrated to effectively silence CAP1.
PLGA/siCAP1 in each selected ratio [100:1-800:1 (w/w)] and was
demonstrated to effectively silence CAP1 expression in A549 cells
as CAP1 mRNA expression was significantly lower in the PLGA/siCAP1
and siCAP1 complexed to L2K groups compared with the control group
(all P<0.01; Fig. 2B). When the
ratio was adjusted to the range of 600:1-800:1 (w/w), the siCAP1
significantly inhibited CAP1 expression by over 70% when compared
with the control group. Based on these findings, PLGA/siCAP1 at the
ratio of 600:1 (w/w) was demonstrated to be optimal for siRNA
delivery in NSCLC cells.
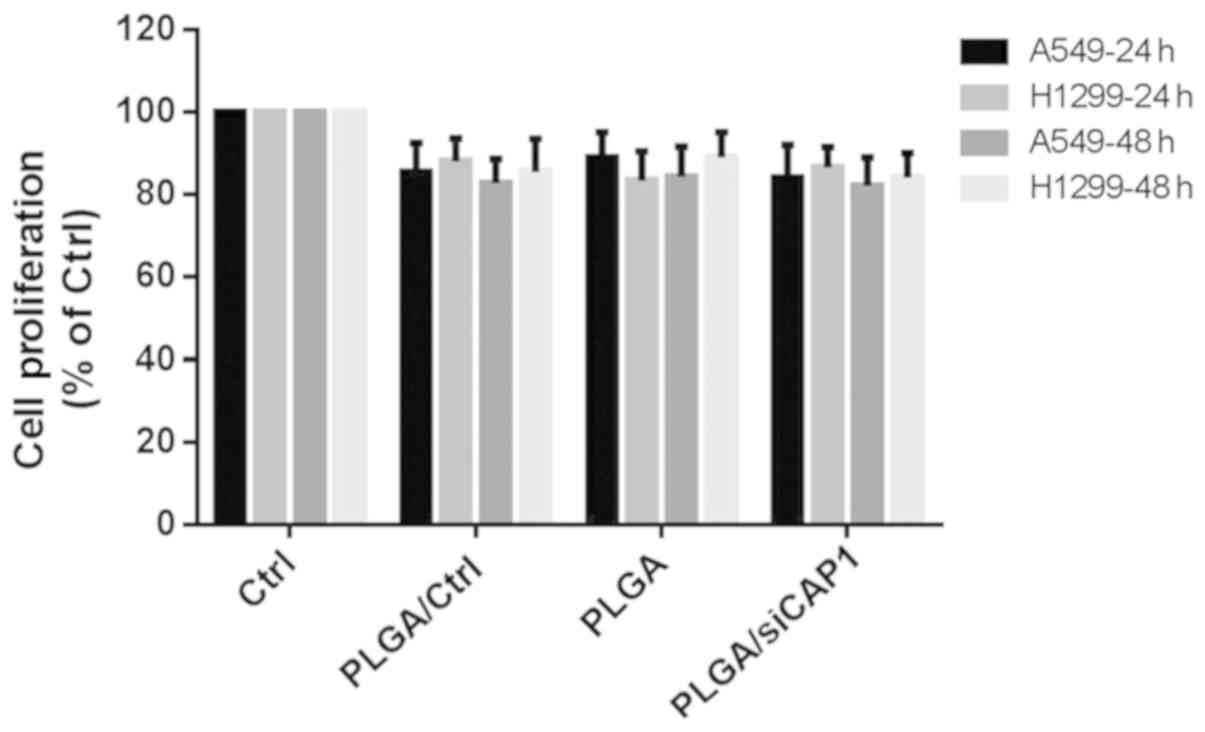
PLGA/siCAP1 NPs have little effect on
cell cytotoxicity
CCK-8 assays were conducted to evaluate the
cytotoxicity of PLGA and PLGA/siCAP1 at the ratio of 600:1 w/w).
Fig. 3 demonstrates the effects of
PLGA NPs on the viability of A549 and H1299 after 24 h and 48 h of
treatment. A small, non-significant decrease in cell viability was
observed following the addition of the PLGA/siCAP1 emulsion.
Approximately 85% of all H1299 and A459 cells remained viable after
24 h and 48 h of incubation, indicating that the PLGA/siCAP1
emulsion exhibits good biocompatibility and minimal influence on
cell viability. Additionally, A549 and H1299 cells treated with
PLGA and PLGA/Ctrl exhibited slightly reduced cell viability, which
was attributed the mild cytotoxic effects of PLGA.
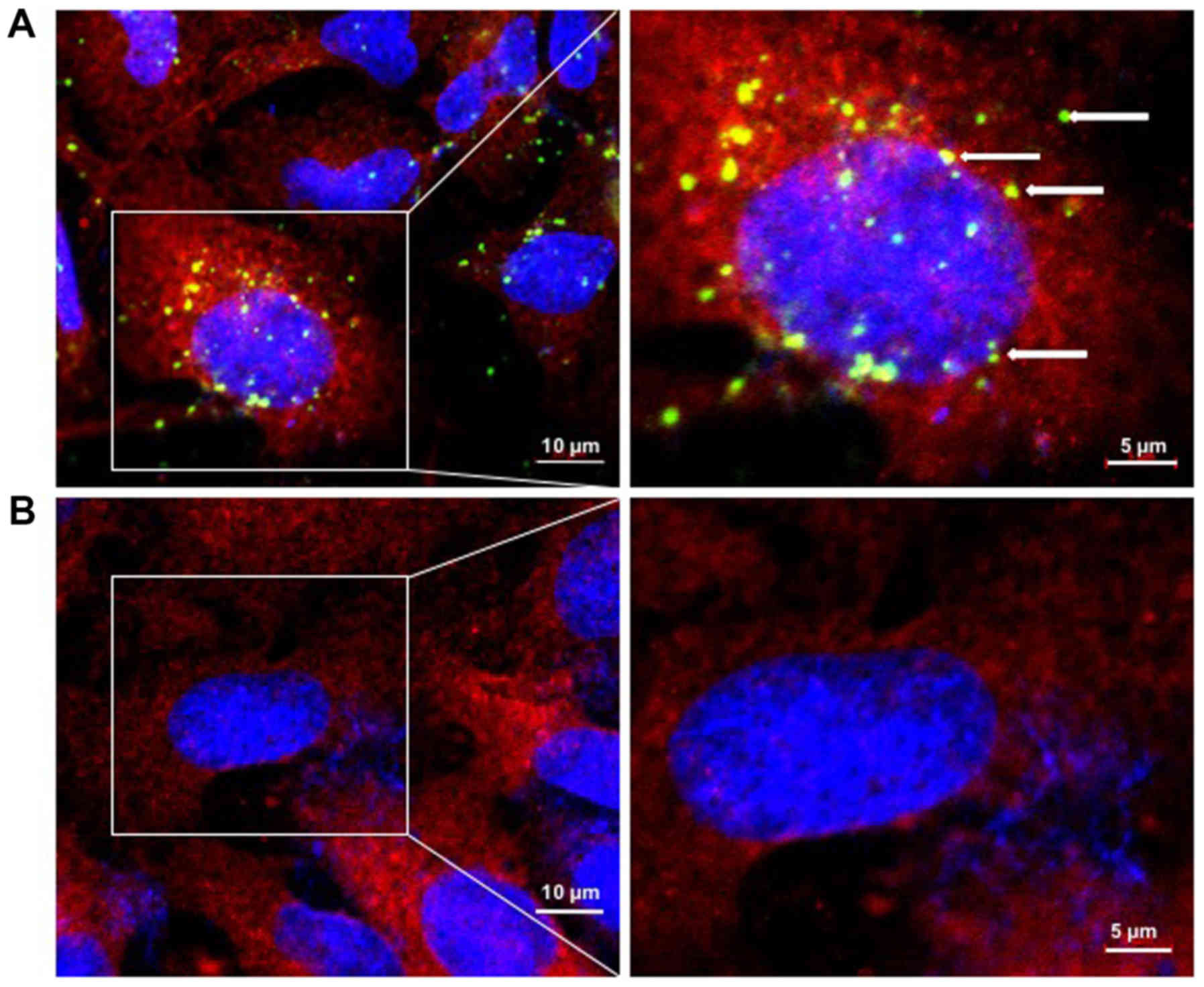
PLGAs deliver siCAP1s to NSCLC cells in
vitro
Previous studies demonstrated the potential use of
siCAP1 as effective delivery vehicles for the therapeutic treatment
of NSCLC (15-17). Furthermore, PLGA was explored as a
delivery system for delivering siRNAs to mice and silencing the
expression of key target genes (33-35).
Inspired by these previous findings, A549 cells were transfected
with PLGA NPs carrying FITC-labeled siCAP1 with the goal of
determining the cellular distribution and uptake of PLGA/siCAP1.
A549 cells transfected with FITC-labeled siCAP1 were used as the
control. After 24 h of transfection, cellular intake of the siCAP1
was examined by confocal microscopy. The results revealed that
siCAP1 is carried by PLGA were efficiently taken up by A549 cells
(Fig. 4A). By contrast, siCAP1s
alone did not effectively enter the cells (Fig. 4B). Taken together, these results
reveal that PLGA NPs can deliver of siCAP1s to NSCLC cells.
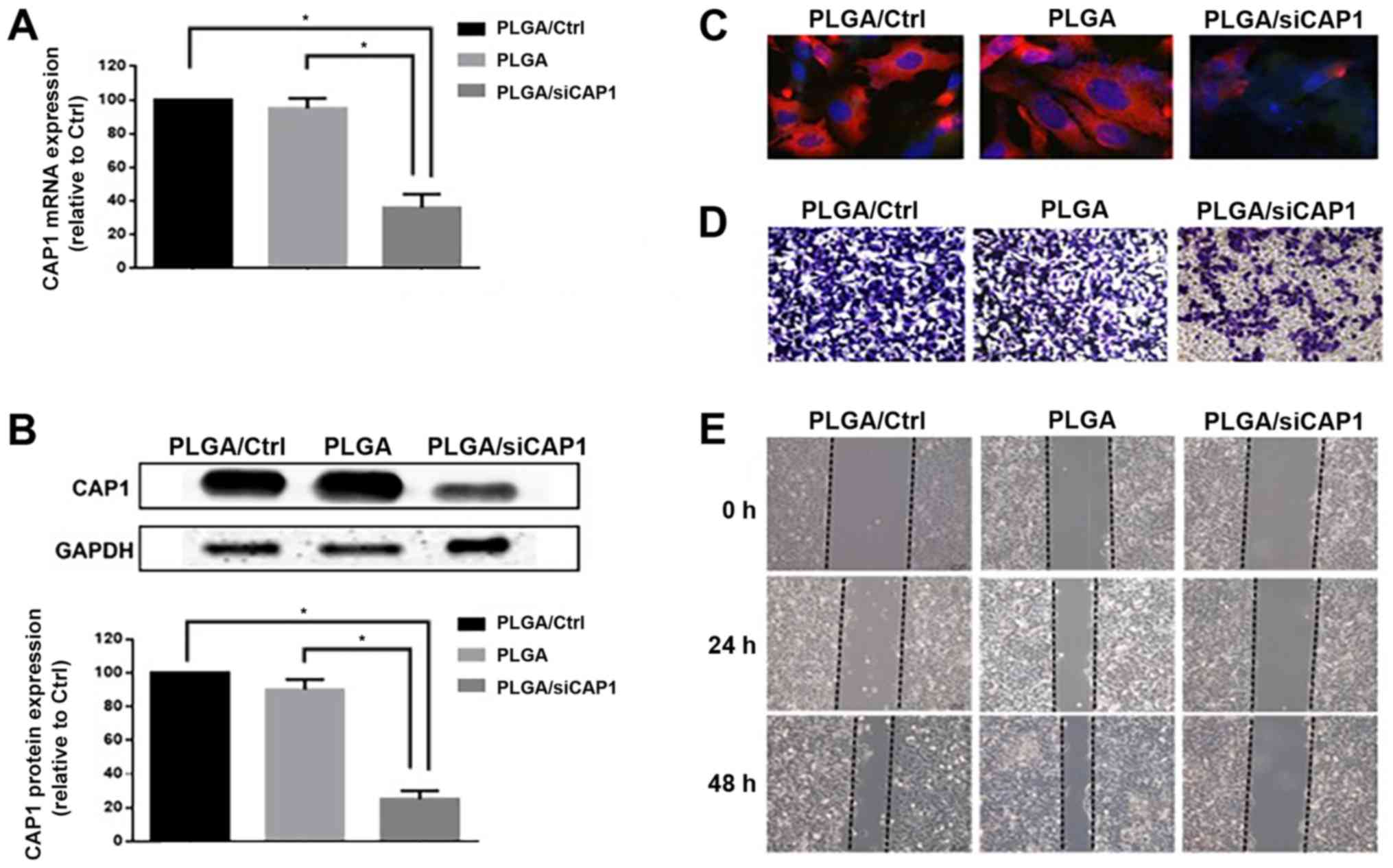
PLGA/siCAP1 silences CAP1 expression and
reduces the motility of NSCLC cells
Yamazaki et al (36) demonstrated that CAP1 overexpression
significantly influenced the aggressiveness of pancreatic cancer.
In addition, other studies confirmed that siCAP1s could silence
CAP1 expression and reduce the motility of NSCLC cells (15,16,37).
Given that PLGA/siCAP1 could deliver siCAP1s to NSCLC cells, the
authors of the current study attempted to determine whether
PLGA/siCAP1 could silence CAP1 expression and reduce the motility
of NSCLC cells; the experiments were conducted using A549 cells
treated with PLGAs and PLGA/siCAP1 NPs as the controls. The results
indicated that 70% of CAP1 gene expression levels were inhibited by
PLGA/siCAP1 after 48 h of transfection compared with the control
groups (PLGA and PLGA/Ctrl; F=114.0; all P<0.01; Fig. 5A). After 72 h of transfection, CAP1
protein expression was significantly downregulated by ~80% compared
with the control groups (F=244.7; all P<0.01; Fig. 5B). Furthermore, immunofluorescence
assays indicated that treatment of A549 cells with PLGA/siCAP1
induced a notable decrease in CAP1 expression (Fig. 5C). In addition, the results of
Transwell migration assays demonstrated that treatment with
PLGA/siCAP1 induced a marked decrease in A549 cell motility
(Fig. 5D). The results of the
wound healing assays revealed that cell migration was significantly
reduced compared with the control groups (F=63.1; all P<0.01;
Figs. 5E and S1A). Additionally, the results of
Transwell assays indicated that cell migration was significantly
reduced compared with the control groups (F=288.3; all P<0.01;
Fig. S1B). These results are
consistent with the hypothesis of the current study that siCAP1 was
capable of suppressing CAP1 expression in A549 cells, which in turn
reduced their cell migration capabilities. Taken together,
silencing CAP1 expression by PLGA/siCAP1 exerts an anti-metastatic
effect on NSCLC cells.
PLGAs deliver siRNAs to NSCLC tumors in
vivo
The nude mouse model was established to evaluate the
potential therapeutic use of PLGA/siCAP1 in vivo. After 7
days of inoculation with tumor cells, tumors could be detected and
measured under the skin. PLGA/siCAP1 group can reduce tumor size as
compared to PLGA and untreated group (Figs. 6A, S2
and S3). To determine whether PLGA could deliver a sufficient
number of siRNAs to the developing tumors, systemic administration
was conducted on the experimental mice with FITC-labeled
PLGA/siCAP1 NPs and controls. The collected tumors were observed by
confocal microscopy, which verified that the fluorescent siCAP1 was
present in the PLGA/siCAP1-treated tumor tissues (Fig. 6B).
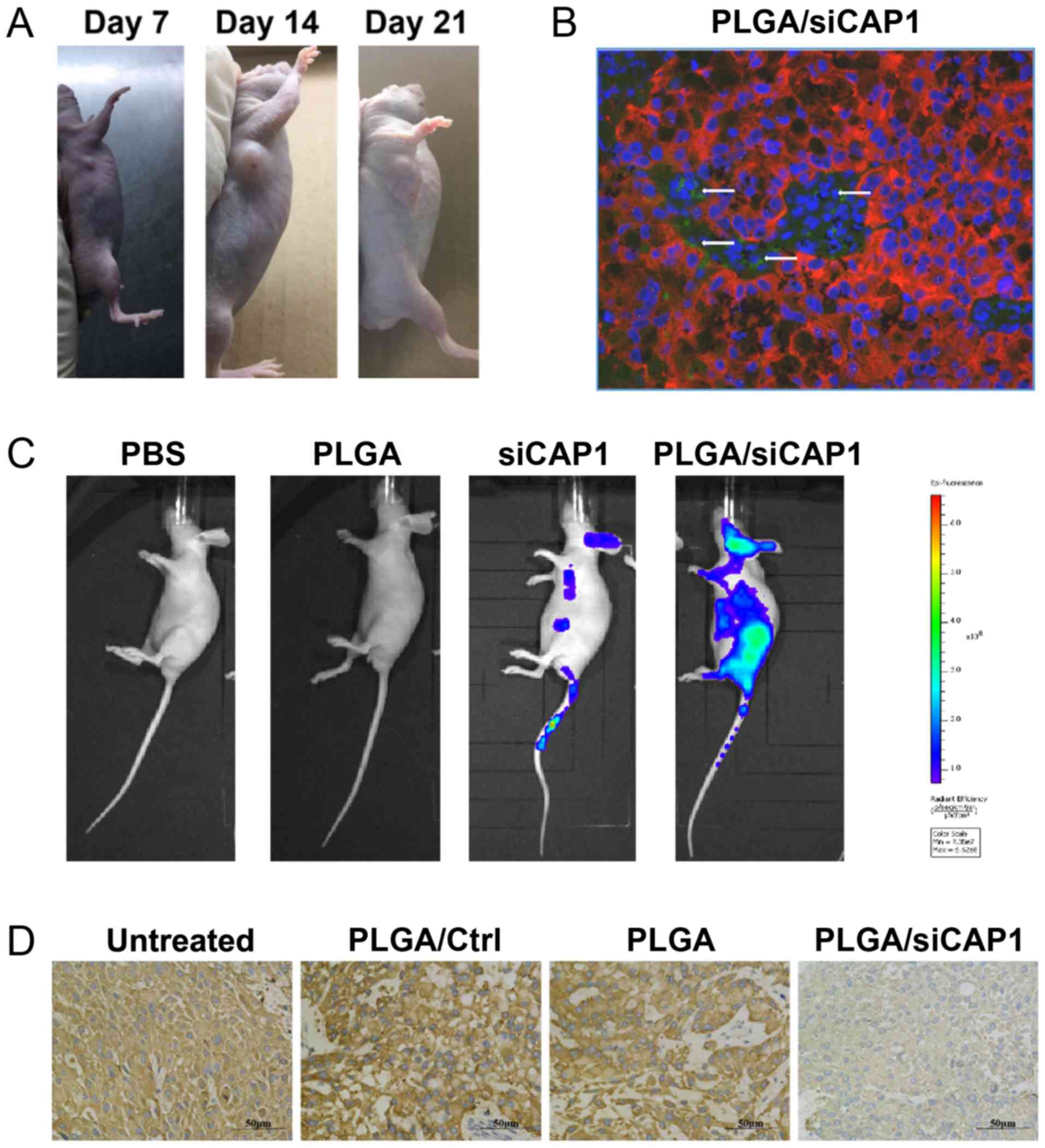 | Figure 6PLGAs deliver siRNAs to NSCLC tumors
in vivo. Mice were injected with PBS, PLGA alone and
FITC-labeled siCAP1, which served as the controls, and with
FITC-labeled PLGA/siCAP1. (A) Images of mice injected with A549
cells. (B) A immunofluorescence image of FITC-labeled PLGA/siCAP1
NPs (green), CAP1 (red) and nuclei (blue) in NSCLC tumor tissues.
The white arrows reveal the location of PLGA/siCAP1 NPs. (C) Ex
vivo fluorescence images of mice after 4 h of treatment. (D)
Immunohistochemical images of CAP1 expression in tumors in mice.
NP, nanoparticle; PLGA, poly(lactic-co-glycolic acid); si, small
interfering RNA; CAP1, adenylyl cyclase-associated protein 1; Ctrl,
control siRNA; NSCLC, non-small cell lung cancer; FITC, fluorescein
isothiocyanate; si, small interfering RNA. |
No fluorescence in the mice that were injected only
with PLGA or PBS was observed, while mice injected with fluorescent
siCAP1 demonstrated weak fluorescence signals, which indicated that
the siCAP1 was eliminated from the body (Fig. 6C). However, mice injected with
PLGA/siCAP1 demonstrated strong fluorescence signals. Furthermore,
following treatment with PLGA/siCAP1, CAP1 expression levels in the
tumor tissues were markedly reduced (Fig. 6D). Collectively, the findings
demonstrated that PLGAs were capable of delivering siCAP1s to NCSLC
tissues grown in vivo similar to human clinical settings and
successfully downregulated CAP1 expression.
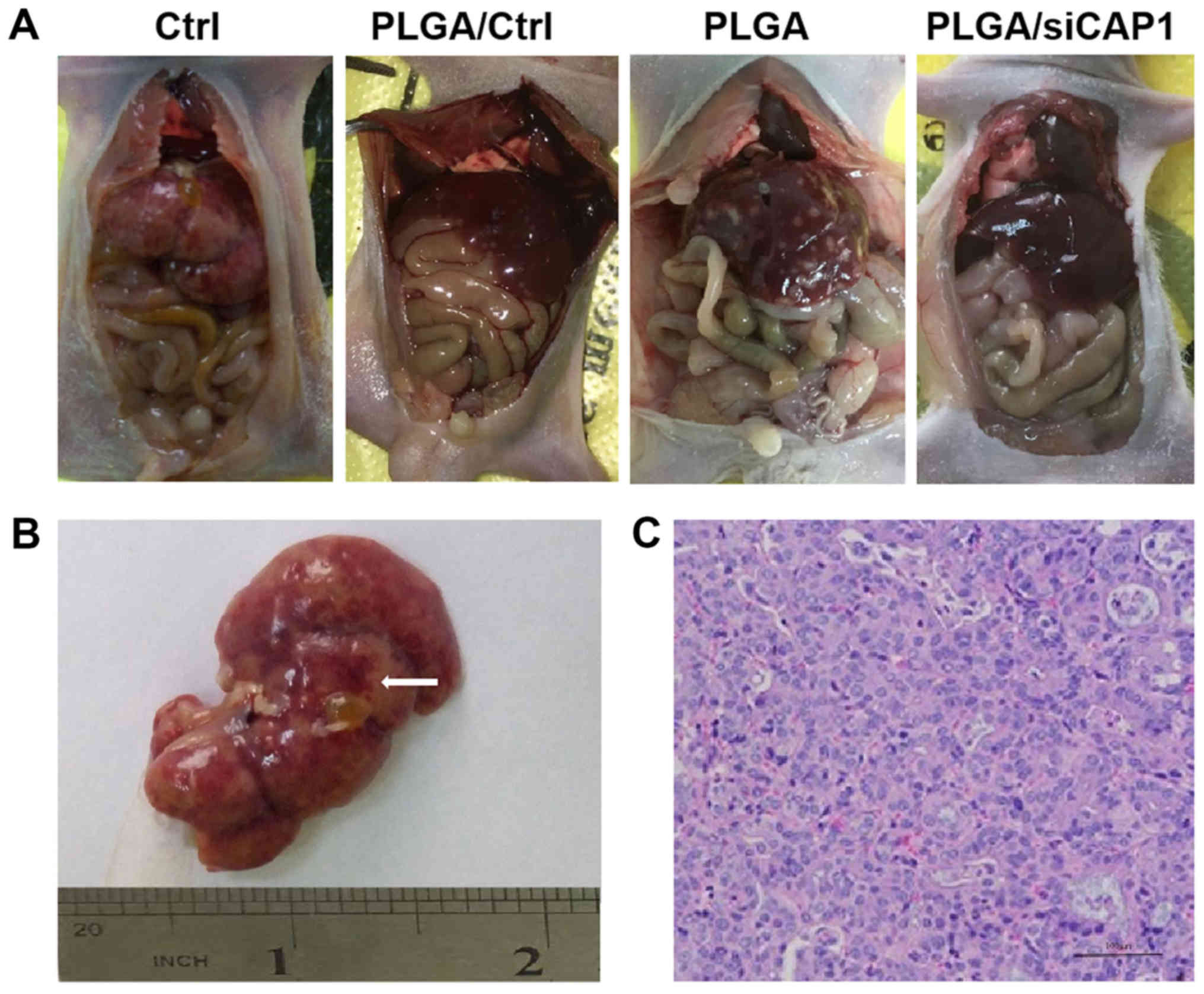
PLGA/siCAP1 reduces NSCLC metastasis in
vivo
Given the high expression of CAP1 in tumors grown
from implanted human NSCLC cells in nude mice, the potential
therapeutic effects of PLGA/siCAP1 on metastatic tumors in
vivo was evaluated. The authors of the current study assessed
the migration and metastatic abilities of NSCLC tumors by observing
tumor growth and inhibition. The four groups of mice experienced
intraperitoneal tumor formation in the internal organs, including
the peritoneum, heart, liver, spleen, lung, kidney, intestine,
uterus, ovary, diaphragm and omentum (Figs. 7 and S4).
The nude mice were sacrificed at 14 days the
inoculation with the different vector and the tumors were
dissected. The tumor formation rate of PLGA/siCAP1 group was
determined to be 60% (3/5) compared with 100% (5/5), 100% (5/5) and
80% (4/5) in the untreated, PLGA/Ctrl and PLGA groups, respectively
(Table I). Although no significant
differences in the overall tumor formation rate among the four
groups was observed, the sizes of the tumors and number of tumors
in the PLGA/siCAP1 group were considerably smaller compared with
those in the three other groups. Significant differences in the
location of the tumors in mice in the PLGA/siCAP1 group were
observed; these tumors were primarily located in the greater
omentum and mesentery, whereas the tumors in the mice in the
untreated, PLGA/Ctrl and PLGA groups were distributed throughout
the omentum, mesentery, liver, renicap-sule and heart (P<0.05;
Fig. 7 and Table I). Furthermore, no significant
differences in the growth of transplanted tumors in mice in the
untreated, PLGA/Ctrl, and PLGA groups were observed.
 | Table IEffects of PLGA/siCAP1 on the growth
of NSCLC tumors in nude mice. |
Table I
Effects of PLGA/siCAP1 on the growth
of NSCLC tumors in nude mice.
| Group | N | Tumor formation
rate | Tumor number | Tumor position |
|---|
| Untreated | 5 | 5 (100) | 7.4±1.8 | 4.8±0.8 |
| PLGA/Ctrl | 5 | 5 (100) | 7.8±1.9 | 4.8±0.8 |
| PLGA | 5 | 4 (80) | 7.2±4.2 | 4.6±2.6 |
| PLGA/siCAP1 | 5 | 3 (60) | 2.3±3.0a,b,c | 1.0±1.4a,b,c |
In the present study, PLGA/siCAP1 NPs were prepared
and used as macromolecule carriers for the targeted delivery of
siRNAs against CAP1 to regulate CAP1 expression in vivo and
in vitro. The results demonstrated that silencing CAP1
expression using PLGA/siCAP1 NPs could markedly inhibit the
migration of A549 cells in vivo and in vitro. To the
best of our knowledge, the current study is the first to
demonstrate the successful delivery of siCAP1s to NSCLC cells using
PLGAs, which reduced the migration of tumor cells in a metastatic
NSCLC mouse model that can mimic the clinical tumor
microenvironment.
The findings of the current study provided a basis
for the development of a novel strategy of drug delivery by
PLGA-mediated delivery of siCAP1s to NSCLC cells in vivo and in
vitro. In addition, PLGA/siCAP1s effectively silenced CAP1
expression in NSCLC cells, and reduced cell motility and metastatic
potential. The current study demonstrated the potential use of
PLGA/siCAP1 for the therapeutic treatment of metastatic NSCLC.
Additionally, the proposed method described in the current study
could be applied to other types of cancers characterized by CAP1
expression.
Supplementary Data
Abbreviations:
|
CAP1
|
adenylyl cyclase-associated protein
1
|
|
NSCLC
|
non-small cell lung cancer
|
|
NP
|
nanoparticles
|
|
siRNA
|
small interfering RNA
|
|
PLGA
|
poly(lactic-polyglycolic acid)
|
|
PLGA/siCAP1
|
CAP1-siRNA in PLGA
|
|
PLGA/Ctrl
|
control-siRNA in PLGA
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
FBS
|
fetal bovine serum
|
|
CCK-8
|
cell counting kit-8
|
Acknowledgments
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant nos. 81472180 and 81802262) and
Shanghai Tenth Hospital's improvement plan for NSFC (grant nos.
04.03.17.032 and 04.01.18.048).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL and SX collected and analyzed the data. YL, MT
and JZ performed the experiments and wrote the manuscript. SX, JZ,
PW, JW and XS designed the study and wrote of the manuscript. CW
and DH conceived the study, directed the project and contributed to
writing of the manuscript. All authors edited and approved the
final version of the manuscript.
Ethics approval and consent to
participate
Animal experiments were approved by the
Institutional Review Board for Animal Research at Shanghai Tenth
People's Hospital (Shanghai, China).
Patient consent for publication
Not applicable.
Conflicts of interest
The authors declare that they have no conflict of
interest.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ramalingam SS, Owonikoko TK and Khuri FR:
Lung cancer: New biological insights and recent therapeutic
advances. CA Cancer J Clin. 61:91–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aisner DL and Marshall CB: Molecular
pathology of non-small cell lung cancer: A practical guide. Am J
Clin Pathol. 138:332–346. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lozano R, Naghavi M, Foreman K, Lim S,
Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, et
al: Global and regional mortality from 235 causes of death for 20
age groups in 1990 and 2010: A systematic analysis for the Global
Burden of Disease Study 2010. Lancet. 380:2095–2128. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fedor-Chaiken M, Deschenes RJ and Broach
JR: SRV2, a gene required for RAS activation of adenylate cyclase
in yeast. Cell. 61:329–340. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Field J, Vojtek A, Ballester R, Bolger G,
Colicelli J, Ferguson K, Gerst J, Kataoka T, Michaeli T, Powers S,
et al: Cloning and characterization of CAP, the S. cerevisiae gene
encoding the 70 kd adenylyl cyclase-associated protein. Cell.
61:319–327. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matviw H, Yu G and Young D: Identification
of a human cDNA encoding a protein that is structurally and
functionally related to the yeast adenylyl cyclase-associated CAP
proteins. Mol Cell Biol. 12:5033–5040. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Freeman NL, Chen Z, Horenstein J, Weber A
and Field J: An actin monomer binding activity localizes to the
carboxyl-terminal half of the Saccharomyces cerevisiae
cyclase-associated protein. J Biol Chem. 270:5680–5685. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moriyama K and Yahara I: Human CAP1 is a
key factor in the recycling of cofilin and actin for rapid actin
turnover. J Cell Sci. 115:1591–1601. 2002.PubMed/NCBI
|
|
10
|
Hubberstey AV and Mottillo EP:
Cyclase-associated proteins: CAPacity for linking signal
transduction and actin polymerization. FASEB J. 16:487–499. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Loisel TP, Boujemaa R, Pantaloni D and
Carlier MF: Reconstitution of actin-based motility of Listeria and
Shigella using pure proteins. Nature. 401:613–616. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu XF, Ni QC, Chen JP, Xu JF, Jiang Y,
Yang SY, Ma J, Gu XL, Wang H and Wang YY: Knocking down the
expression of adenylate cyclase-associated protein 1 inhibits the
proliferation and migration of breast cancer cells. Exp Mol Pathol.
96:188–194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fan YC, Cui CC, Zhu YS, Zhang L, Shi M, Yu
JS, Bai J and Zheng JN: Overexpression of CAP1 and its significance
in tumor cell proliferation, migration and invasion in glioma.
Oncol Rep. 36:1619–1625. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hua M, Yan S, Deng Y, Xi Q, Liu R, Yang S,
Liu J, Tang C, Wang Y and Zhong J: CAP1 is overexpressed in human
epithelial ovarian cancer and promotes cell proliferation. Int J
Mol Med. 35:941–949. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xie SS, Tan M, Lin HY, Xu L, Shen CX, Yuan
Q, Song XL and Wang CH: Overexpression of adenylate
cyclase-associated protein 1 may predict brain metastasis in
non-small cell lung cancer. Oncol Rep. 33:363–371. 2015. View Article : Google Scholar
|
|
16
|
Xie S, Shen C, Tan M, Li M, Song X and
Wang C: Systematic analysis of gene expression alterations and
clinical outcomes of adenylate cyclase-associated protein in
cancer. Oncotarget. 8:27216–27239. 2017.PubMed/NCBI
|
|
17
|
Tan M, Song X, Zhang G, Peng A, Li X, Li
M, Liu Y and Wang C: Overexpression of adenylate cyclase-associated
protein 1 is associated with metastasis of lung cancer. Oncol Rep.
30:1639–1644. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rana TM: Illuminating the silence:
Understanding the structure and function of small RNAs. Nat Rev Mol
Cell Biol. 8:23–36. 2007. View Article : Google Scholar
|
|
19
|
Coelho T, Adams D, Silva A, Lozeron P,
Hawkins PN, Mant T, Perez J, Chiesa J, Warrington S, Tranter E, et
al: Safety and efficacy of RNAi therapy for transthyretin
amyloidosis. N Engl J Med. 369:819–829. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Davis ME, Zuckerman JE, Choi CH, Seligson
D, Tolcher A, Alabi CA, Yen Y, Heidel JD and Ribas A: Evidence of
RNAi in humans from systemically administered siRNA via targeted
nanoparticles. Nature. 464:1067–1070. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tabernero J, Shapiro GI, LoRusso PM,
Cervantes A, Schwartz GK, Weiss GJ, Paz-Ares L, Cho DC, Infante JR,
Alsina M, et al: First-in-humans trial of an RNA interference
therapeutic targeting VEGF and KSP in cancer patients with liver
involvement. Cancer Discov. 3:406–417. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi L, Shao W and Shi D; QiLingfeng; Shao W
and Shi D: JAM-2 siRNA intracellular delivery and real-time imaging
by proton-sponge coated quantum dots. J Mater Chem B Mater Biol
Med. 1:654–660. 2013. View Article : Google Scholar
|
|
23
|
Su J, Baigude H, McCarroll J and Rana TM:
Silencing microRNA by interfering nanoparticles in mice. Nucleic
Acids Res. 39:e382011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sezlev Bilecen D, Rodriguez-Cabello JC,
Uludag H and Hasirci V: Construction of a PLGA based, targeted
siRNA delivery system for treatment of osteoporosis. J Biomater Sci
Polym Ed. 28:1859–1873. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pisani E, Tsapis N, Paris J, Nicolas V,
Cattel L and Fattal E: Polymeric nano/microcapsules of liquid
perfluorocarbons for ultrasonic imaging: Physical characterization.
Langmuir. 22:4397–4402. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Patnaik S, Sharma AK, Garg BS, Gandhi RP
and Gupta KC: Photoisomerization of azobenzene moiety in
crosslinking. Int J Pharm. 342:184–193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cheng J, Teply BA, Sherifi I, Sung J,
Luther G, Gu FX, Levy-Nissenbaum E, Radovic-Moreno AF, Langer R and
Farokhzad OC: Formulation of functionalized PLGA-PEG nanoparticles
for in vivo targeted drug delivery. Biomaterials. 28:869–876. 2007.
View Article : Google Scholar
|
|
28
|
Díaz-López R, Tsapis N, Santin M, Bridal
SL, Nicolas V, Jaillard D, Libong D, Chaminade P, Marsaud V,
Vauthier C, et al: The performance of PEGylated nanocapsules of
perfluorooctyl bromide as an ultrasound contrast agent.
Biomaterials. 31:1723–1731. 2010. View Article : Google Scholar
|
|
29
|
Boyer C, Teo J, Phillips P, Erlich RB,
Sagnella S, Sharbeen G, Dwarte T, Duong HT, Goldstein D, Davis TP,
et al: Effective delivery of siRNA into cancer cells and tumors
using well-defined biodegradable cationic star polymers. Mol Pharm.
10:2435–2444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Byrne FL, Yang L, Phillips PA, Hansford
LM, Fletcher JI, Ormandy CJ, McCarroll JA and Kavallaris M:
RNAi-mediated stathmin suppression reduces lung metastasis in an
orthotopic neuroblastoma mouse model. Oncogene. 33:882–890. 2014.
View Article : Google Scholar
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
32
|
van der Weij JP, van der Veen CJ, de Vries
E and Cats A: The use of a peroxidase-anti-peroxidase complex for
the visualization of monoclonal antibodies on the ultrastructural
level. Clin Exp Immunol. 54:819–826. 1983.PubMed/NCBI
|
|
33
|
McCarroll JA, Gan PP, Liu M and Kavallaris
M: betaIII-tubulin is a multifunctional protein involved in drug
sensitivity and tumorigenesis in non-small cell lung cancer. Cancer
Res. 70:4995–5003. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
De Rosa G and Salzano G: PLGA microspheres
encapsulating siRNA. Methods Mol Biol. 1218:43–51. 2015. View Article : Google Scholar
|
|
35
|
Zhao Y, Zheng C, Zhang L, Chen Y, Ye Y and
Zhao M: Knockdown of STAT3 expression in SKOV3 cells by
biodegradable siRNA-PLGA/CSO conjugate micelles. Colloids Surf B
Biointerfaces. 127:155–163. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yamazaki K, Takamura M, Masugi Y, Mori T,
Du W, Hibi T, Hiraoka N, Ohta T, Ohki M, Hirohashi S, et al:
Adenylate cyclase-associated protein 1 overexpressed in pancreatic
cancers is involved in cancer cell motility. Lab Invest.
89:425–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Frede A, Neuhaus B, Klopfleisch R, Walker
C, Buer J, Müller W, Epple M and Westendorf AM: Colonic gene
silencing using siRNA-loaded calcium phosphate/PLGA nanoparticles
ameliorates intestinal inflammation in vivo. J Control Release.
222:86–96. 2016. View Article : Google Scholar
|