Introduction
Metastasis is the most life-threatening
characteristic of cancer; it is a multi-step process that causes
cancer cells to spread throughout the body (1). Malignant tumors containing both
neoplastic cells and non-neoplastic cellular components possess the
ability to metastasize due to interactions between the cellular
elements of the tumor with the extracellular matrix (ECM) (2,3).
Numerous growth factors and their receptors, in addition to
angiogenic factors, are involved in the process of cancer
metastasis (4).
Breast cancer is the most common malignant tumor in
women worldwide, and recurrence and metastasis remain the leading
causes of mortality in patients with this disease. However, there
are numerous treatment modalities currently available (5), which demonstrates the need for novel
therapeutic targets and agents for the treatment and prognosis of
breast cancer.
Hydrogen sulfide (H2S), as the third
gasotransmitter signaling molecule alongside nitric oxide (NO) and
carbon monoxide (CO), functions as a potent angiogenic factor
(6,7) and is involved in a number of other
physiological processes (8-11). A
previous study also revealed endogenously produced H2S
in the angiogenic process (12).
The pharmacological inhibition and silencing of the enzymes
involved in H2S synthesis attenuates the angiogenic
properties of endothelial cells, including proliferation, migration
and formation of a tube-like structure network. By contrast, the
enhanced production of H2S by substrate supplementation
or the overexpression of H2S-producing enzymes leads to
enhanced angiogenic responses in cultured endothelial cells and
upregulates the expression of vascular endothelial growth factor
(VEGF), a key angiogenic factor.
Cystathionine-γ-lyase (CSE), one of key enzymes of
the trans-sulfuration pathway, catalyzes endogenous H2S
production (13). Studies have
demonstrated that endogenous H2S produced by CSE can
promote the proliferation of human cancer cells (14,15).
A previous study revealed that high expression levels of CSE
promoted the progression of breast cancer in association with the
STAT3 signaling pathway (16).
Therefore, the CSE/H2S system serves an important role
in the progression of cancer.
It is already well established that tumor metastasis
is closely associated with tumor angiogenesis. Based on the
function of endogenous H2S promoting angiogenesis and
the role of the CSE/H2S system in tumors, it is possible
that the CSE/H2S system may be involved in the
metastasis of a tumor. Therefore, the aim of the present study was
to elucidate the roles and regulatory mechanisms of CSE in the
metastasis of breast cancer cells. In addition, the effects and
mechanisms of novel CSE inhibitors against breast cancer metastasis
were investigated. The present study reveals novel avenues for
investigations into useful therapeutic targets and agents for the
treatment of breast cancer.
Materials and methods
Patient samples, animals and cell
lines
A total of 30 breast cancer tissues and adjacent
non-tumor tissues were obtained from patients at Huaihe Hospital
(Kaifeng, China). All patients (age range, 37-70 years; no smoking
history available) had a non-specific type of invasive breast
cancer (10 cases of grade II and 20 cases of grade III breast
cancer; 13 cases of non-lymph node metastasis; and 17 cases of
lymph node metastasis). Balb/c nude mice (specific pathogen-free;
female; 4-6 weeks) weighing 18-22 g were supplied by Beijing
Weitong Lihua Experimental Animal Technology Co., Ltd. The mice
were allowed free access to food and water, and were housed in an
environment with constant temperature (24±1°C), humidity (50-70%),
ventilation rate (10-20 times/h), noise (<40 db) and working
illumination (250 lx) under a 12-h light/dark cycle. Drinking
water, food and experimental supplies were sterilized and
disinfected. The present study was approved by the Ethics Committee
at the Medical School, Henan University (Kaifeng, China). Early
metastatic breast cancer cells (MDA-MB-231), non-metastatic breast
cancer cells (MCF7) and mammary epithelial cells (Hs578Bst) were
obtained from the American Type Culture Collection, and cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (Zeta-life) in a 37°C incubator with 5%
CO2.
Immunohistochemistry
Immunohistochemical staining of the surgical
specimens from patients with breast cancer was performed in serial
sections (average section area, 1×1 cm) of formalin-fixed,
paraffin-embedded tissues (0.2 cm-thick). Following
deparaffinization, the slides were placed in 0.01 M citrate salt
solution (Epitope Retrieval Solution) and heated in a microwave
oven for 7 min. Following cooling and washing with PBS, endogenous
peroxidase was blocked with 30% H2O2 for 10
min and incubated with 5% BSA (Sigma-Aldrich; Merck KGaA) to block
non-specific binding of antibodies. The slides were then incubated
with CSE primary antibody (cat. no. 60234-1-Ig; 1:100; ProteinTech
Group, Inc.) at 4°C overnight, followed by incubation with
biotin-conjugated secondary antibody (cat. no. SA00004-1; 1:1,000;
ProteinTech Group, Inc.) and streptavidin horseradish peroxidase
for 10 min each. Antigen-antibody complexes were visualized in
3,3′-diamino-benzidine, and cells were subsequently stained with
hematoxylin and dehydrated, and images were captured with a laser
confocal microscope (LSM710; Carl Zeiss AG). Unless otherwise
specified, all incubation steps were performed at room
temperature.
Western blot analysis
Proteins were extracted from human breast cancer and
paracancerous tissues, the MCF7 and MDA-MB-231 human breast cancer
cells, CSE-knockdown MDA-MB-231 cells [MDA-MB-231 cells transfected
with CSE small interfering RNA (siRNA) for 48 h, or treated with
10, 20 and 30 μM I157172 for 24 h, with CSE-knockdown
MDA-MB-231 cells transfected with the CSE plasmid for 48 h] and
normal mammary epithelial cells (Hs578Bst) using
radioimmunoprecipitation assay buffer [50 mM Tris-HCl (pH 8.0), 150
mM sodium chloride, 1.0% NP-40, 0.5% sodium deoxycholate and 0.1%
SDS] supplemented with 10 μg/ml phenylmethylsulfonyl
fluoride (Sigma-Aldrich; Merck KGaA). The samples were then
centrifuged at 12,000 × g for 10 min at 4°C, and protein
concentration was determined using the Bicinchoninic Acid Protein
Quantitative kit (Solarbio Science & Technology Co., Ltd.). The
protein samples (40 μg) were separated by SDS-PAGE (10% gel)
and transferred onto polyvinylidene difluoride membranes (EMD
Millipore) at 70 mA for 2 h at 4°C. The membrane was then blocked
in 5% fat-free milk for 2 h at room temperature, and probed with
specific primary antibodies against CSE, matrix metalloproteinase
(MMP)-2, MMP-9, VEGF, phosphatidylinositol 3-kinase (PI3K), protein
kinase B (Akt), phosphorylated (p)Akt, focal adhesion kinase (FAK),
paxillin, rat sarcoma (Ras), rapidly accelerated fibrosarcoma
(Raf), extracellular signal-regulated protein kinases (ERK)1/2, and
pERK1/2 at 4°C overnight. Following incubation with the secondary
antibody for 2 h at room temperature, the proteins were visualized
using an EasyBlot Enhanced Chemiluminescence kit (Sangon Biotech
Co., Ltd.) and detected using a FluorChem Q Multifluor system
(ProteinSimple). GAPDH was used as a loading control. The primary
antibodies were as follows: CTH (CSE) mouse monoclonal antibody
(1:100; cat. no. sc-365382; lot no. LL2619) from Santa Cruz
Biotechnology, Inc.; MMP-2 rabbit polyclonal antibody (1:1,000;
cat. no. 4022), MMP-9 rabbit polyclonal antibody (1:1,000; cat. no.
3852), Akt rabbit monoclonal antibody (1:1,000; cat no. 4685), pAkt
rabbit monoclonal antibody (1:1,000; cat no. 4060), Ras rabbit
monoclonal antibody (1:1,000; cat no. 3965), p44/42 MAPK (Erk1/2)
rabbit monoclonal antibody (1:1,000; cat. no. 4695) and pErk1/2
rabbit monoclonal antibody (1:1,000; cat. no. 4376) from Cell
Signaling Technology, Inc.; VEGF rabbit polyclonal antibody
(1:1,000; cat. no. 19003-1-AP), PI3Kp110(β) rabbit polyclonal
antibody (1:1,000; cat. no. 20584-1-AP), FAK rabbit polyclonal
antibody (1:1,000; cat. no. 12636-1-AP), paxillin rabbit polyclonal
antibody (cat. no. 22172-1-AP) and RAF rabbit polyclonal antibody
(cat. no. 551140-1-AP) from ProteinTech Group, Inc.; and GAPDH
mouse monoclonal antibody (1:1,000; cat. no. AG019) from Beyotime
Institute of Biotechnology. The secondary antibodies were as
follows: Horseradish peroxidase-conjugated goat anti-mouse
(1:10,000; cat. no. SA00001-1) and horseradish
peroxidase-conjugated goat anti-rabbit (1:10,000; cat. no.
SA00001-2) from ProteinTech Group, Inc. The quantitative
differences were determined using Image-J2x software (Rawak
Software, Inc.).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from the human breast cancer
and adjacent non-tumor tissues, human breast cancer cells and
mammary epithelial cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), and genomic DNA elimination and cDNA
synthesis were performed using PrimeScript™ RT Reagent kit (RR047A;
Takara). qPCR was performed using SYBR Premix Ex Taq II, and the
PikoReal™ Real-Time PCR system (Thermo Fisher Scientific, Inc.) was
used to measure mRNA expression. The reaction conditions were as
follows: Initial template degeneration, 95°C for 30 sec; followed
by 40 cycles at 95°C for 5 sec, 55°C for 30 sec and 72°C for 30
sec; and a final step at 60°C for 30 sec. The reactions for each
sample-primer set were performed in triplicate. Relative
quantification analysis was performed using the comparative
2−ΔΔCq method (17).
All data were normalized to the internal control GAPDH. The primer
sequences were as follows: CSE, forward, 5′-CCCATCTCACTGTCCACCAC-3′
and reverse, 5′-GTGCTGCCACTGCTTTTTCA-3′; length, 115 bp; GAPDH,
forward, 5′-CTCTGCTCCTCCTGTTCGAC-3′ and reverse,
5′-ACCAAATCCGTTGACTCCGA-3′; length, 109 bp.
siRNA and plasmid transfection, and
construction of stable CSE-knockdown MDA-MB-231 cells
For knockdown in the MDA-MB-231 cells, MDA-MB-231
cells were cultured in 6-well plates until they reached 30-40% cell
confluence. The cells were transfected with scrambled siRNA (Sc
siRNA) or specific siRNA against human CSE using Lipofectamine™
2000 (both from Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C
for 48 h. The medium was replaced at 6 h post-transfection, and the
silencing efficiency was determined via western blotting at 48 h
post-transfection. The primer sequences for the CSE-specific siRNA
were as follows: Forward, 5′-GGUUUAG CAGCCACUGUAAdTdT-3′ and
reverse, 5′-UUACAGUGGCU GCUAAACCdTdT-3′; the primers were designed
to target the open reading frame region of CSE mRNA.
For overexpression in MDA-MB-231 cells, the
MDA-MB-231 cells were cultured in 6-well plates until they reached
80-90% confluence. The cells were then transfected with pCMV-EGFP
vector and pCMV-EGFP-hCSE (GeneChem Co., Ltd.) using Lipofectamine
2000, according to the manufacturer's protocol. The cells were
exposed to fresh medium at 6 h post-transfection and the
overexpression efficiency was determined via western blot analysis
at 48 h post-transfection.
For the construction of stable CSE-knockdown
MDA-MB-231 cells, the MDA-MB-231 cells (5×104) were
seeded into 12-well plates. After 24 h, 500 μl of the medium
was replaced with medium containing 50 μl lentiviral
packaging CSE short hairpin (sh)RNA
(hU6-MCS-CBh-gcGFP-IRES-puromycin-CSE shRNA (1×107
TU/ml). After another 10 h, all the medium was removed and replaced
with normal medium. After 72 h, puromycin was added to screen and
obtain the stable CSE-knockdown MDA-MB-231 cells.
Establishment of a human breast cancer
xenograft tumor model in nude mice and analysis of the effect of
CSE on breast cancer metastasis in vivo
A total of 19 BALB/c nude mice (female; age range,
4-6 weeks) were divided into three groups: Blank group (three
BALB/c nude mice); CSE expression group (eight BALB/c nude mice);
and CSE knockdown group (eight BALB/c nude mice). MDA-MB-231 cells
(2×106) and MDA-MB-231 cells stably transfected with CSE
shRNA (2×106) were injected into the nude mice of two
groups (CSE expression group and CSE knockdown group, respectively)
through the tail vein. Physiological saline was injected into nude
mice of the blank group. After 4 weeks, the nude mice were
sacrificed and the lung tissues were dissected. Hematoxylin and
eosin (H&E) staining of the lung tissues was then performed to
further analyze the effect of the expression of CSE on breast
cancer metastasis. H&E staining of lung tissues of nude mice
was performed in serial sections (average section area, 1×1 cm) of
formalin-fixed, paraffin-embedded tissues (0.2 cm-thick). Following
deparaffinization, the slides were first dipped in the updated
xylene for 15 min (twice) and then placed in the updated 100%
ethanol for 5 min (twice), followed by soaking in 95% ethanol, 85%
ethanol, 75% ethanol and double distilled water for 5 min each. The
sections were then stained with hematoxylin for 1 min,
differentiated with 1% hydrochloric acid-ethanol for 10 sec, and
washed with water for 20 sec, followed by staining with eosin for
30 sec. Finally, the sections were dehydrated with double distilled
water, 75% ethanol, 85% ethanol, 95% ethanol and anhydrous ethanol,
and images were captured with a microscope (BX43F; Olympus
Corporation).
MTS assay
The MDA-MB-231 cells were seeded into 96-well plates
at a density of 1×106/ml, and treated with CSE siRNA,
the CSE overexpression plasmid, and 0, 5, 10, 20, 30 and 40
μM I157172 at 37°C for 48 h, respectively. Cell viability
was evaluated by determining the number of cells with MTS
(Sigma-Aldrich; Merck KGaA), according to the manufacturer's
protocol. The assay was performed in triplicate for three
independent experiments.
5-ethynyl-2′-deoxyuridine (EdU)
assay
The EdU assay was used to examine the effects of the
novel CSE inhibitor I157172 on cell proliferation. Briefly, the
MDA-MB-231 cells were cultured in 96-well plates at a density of
1×104/well and then exposed to 0, 10, 20 and 30
μM I157172 (Compound Handing B.V.) for 24 h at 37°C.
Subsequently, the cells were incubated with 50 μm EdU for 2
h at room temperature, fixed with 4% formaldehyde for 30 min at
room temperature, incubated with glycine (2 mg/ml) for 5 min and
treated with 0.5% Triton X-100 for 10 min to permeabilize the
cells. Following washing with PBS for 5 min, the cells were
incubated with 1X Apollo® 567 (Guangzhou Ribobio Co., Ltd.) for 30
min and treated twice with 0.5% Triton X-100. The DNA was stained
with Hoechst 33342 for 30 min and visualized using fluorescence
microscopy. Five groups of cells in the images were randomly
selected.
Scratch assay
A scratch assay was used to determine cell
migration. The MDA-MB-231 cells were seeded into a 6-well plate and
were scraped with a 10-μl pipette tip once they reached ~90%
confluence to generate a wound, following which they were rinsed
twice with PBS. Subsequently, the cells were transfected with CSE
siRNA or the CSE overexpression plasmid and were cultured in medium
containing 5% FBS for 24 h at 37°C. The distance of wound closure
was measured at the beginning of the experiment and after 24 h
using an inverted microscope (IX53; Olympus Corporation). The assay
was performed in triplicate.
Transwell assay
A Transwell assay was performed to examine cell
migration and invasion as previously described (18). Briefly, for the invasion assay,
prior to cell seeding, 24-well Transwell chambers (8-μm pore
size; Corning Incorporated) were coated with Matrigel matrix (BD
Biosciences). Following this, 5×104 cells were suspended
in 200 μl DMEM (with 1% FBS) and seeded into the upper
chambers, with DMEM with 15% FBS placed in the lower chambers. For
the migration assay, 2.5×104 cells were suspended in 200
μl DMEM (with 1% FBS) and seeded into the upper chambers,
with DMEM with 15% FBS placed in the lower chambers. After 24 h,
the cells in the upper chambers were treated with 0, 10, 20 and 30
μM I157172 or were transfected with CSE siRNA or the CSE
overexpression plasmid for 24 h at 37°C, and the cells on the upper
surface of the membrane were then removed. The migratory and
invasive cells attaching to the lower surface of the membrane were
fixed with 4% polyoxymethylene for 30 min and stained with 0.1%
crystal violet for 15-20 min at room temperature. Images of the
stained cells were captured under an inverted microscope (IX53;
Olympus Corporation). Subsequently, the cells were dissolved in 10%
acetic acid and the optical density at 570 nm was measured using a
microplate spectrofluorometer (EnSpire; PerkinElmer, Inc.)
Determination of H2S
production
The methylene blue method was used to determine
H2S release to assess the effect of I157172 on the
production of H2S in MDA-MB-231 cells. Briefly, the
MDA-MB-231 cells (1×106/ml) were seeded into a 6-well
tissue culture plate. After 24 h, the cells were treated with 0,
10, 20 and 30 μM I157172, with 2 mM L-cysteine and 0.5 mM
pyridoxal phosphate for 24 h at 37°C. Concurrently, 1% (w/v) zinc
acetate (500 μl) was added to filter papers adhered to the
lid of the 6-well tissue culture plate for 24 h, in order to absorb
H2S. The filter papers were then placed into tubes
containing 0.2% (w/v)
N,N-dimethyl-p-phenylenediamine-dihydrochloride dye (500
μl), 10% (w/v) and incubated for 20 min at room temperature.
The absorbance at 670 nm was subsequently measured. The production
of H2S was determined using a standard curve of NaHS
(0-1 mM; R2, 0.9997) and presented as nmol/min-1 per
1×106 cells. The assay was performed in triplicate.
Statistical analysis
Statistical analyses were performed using SPSS
software (version 17.0; SPSS, Inc.). Data are expressed as the mean
± standard deviation. Differences between groups were analyzed
using one-way analysis of variance followed by the Bonferroni post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
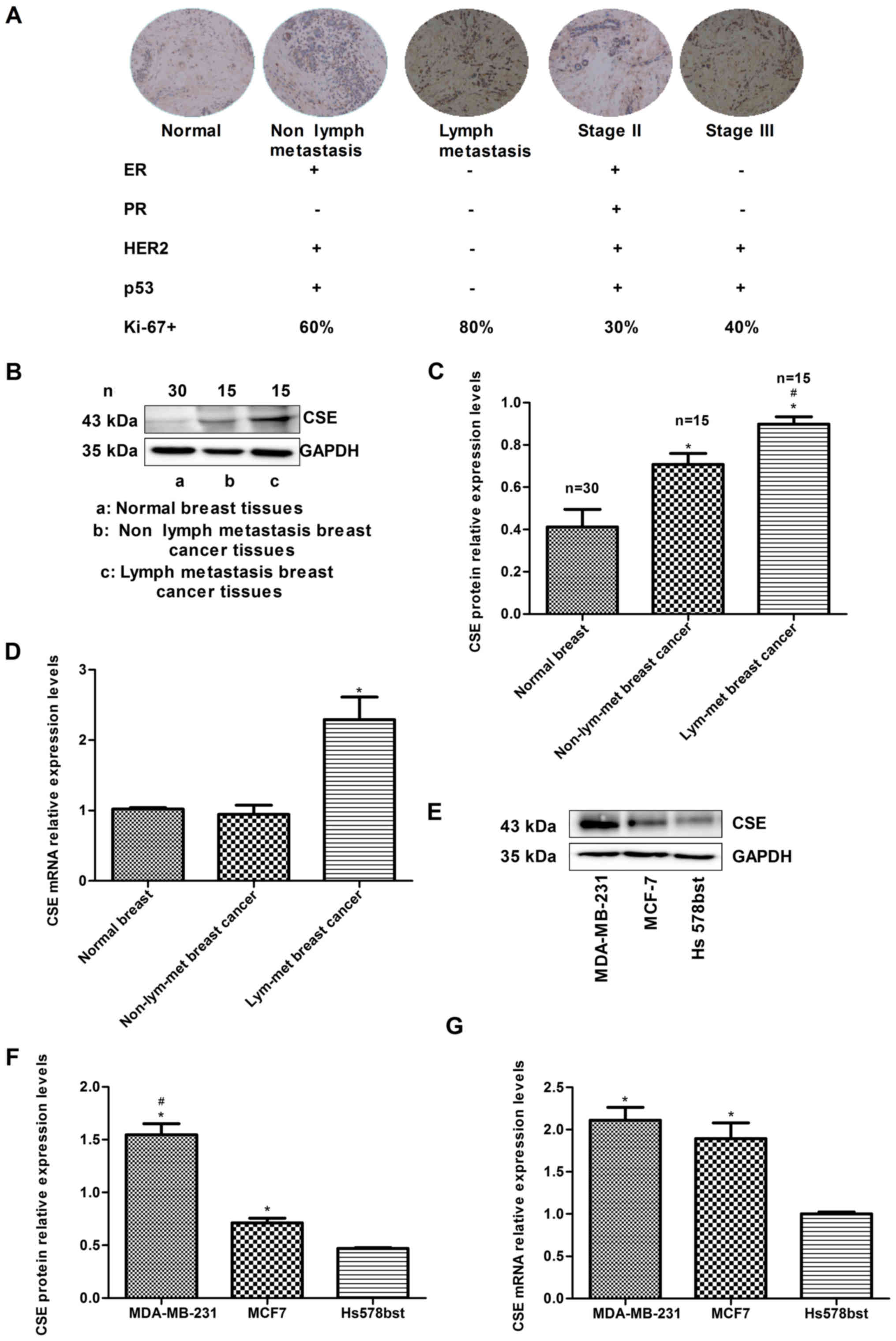
Metastasis of human breast cancer is
associated with the increased expression of CSE
To investigate the association between the
expression of CSE and breast cancer metastasis, the expression
levels of CSE in human breast cancer samples were compared with
different histological grades and lymph node metastasis. The
immunohistochemistry data indicated that the expression of CSE in
samples from patients with breast cancer exhibiting lymph node
metastasis was higher than that in samples without lymph node
metastasis. In addition, the level of CSE in grade III breast
cancer tissues was higher than that in grade II tissues (Fig. 1A). The expression levels of CSE and
the status of breast cancer molecular classification markers were
also analyzed, including estrogen receptor (ER), progesterone
receptor (PR), human epidermal growth factor receptor 2 (HER2), p53
and Ki-67. The results demonstrated that high expression levels of
CSE were negatively associated with an ER+,
PR+ and HER2+ status, and positively
associated with an ER−/PR−/HER2−
(triple negative) status, which is a molecular subtype with a high
risk of recurrence and metastasis (Fig. 1A). The western blotting and RT-qPCR
results also revealed that the levels of CSE in samples from
patients with breast cancer exhibiting lymph node metastasis were
higher than those in the samples with no lymph node metastasis
(Fig. 1B-D). In addition, it was
observed that the mRNA and protein expression levels of CSE in
early metastatic MDA-MB-231 breast cancer cells were higher than
those in non-metastatic MCF7 breast cancer cells (Fig. 1E-G). These data indicated that the
increased expression of CSE may act as a promoting factor in breast
cancer invasion and metastasis and have a relatively unfavorable
prognostic effect during chemotherapy.
Expression of CSE promotes the growth,
migration and invasion of MDA-MB-231 breast cancer cells
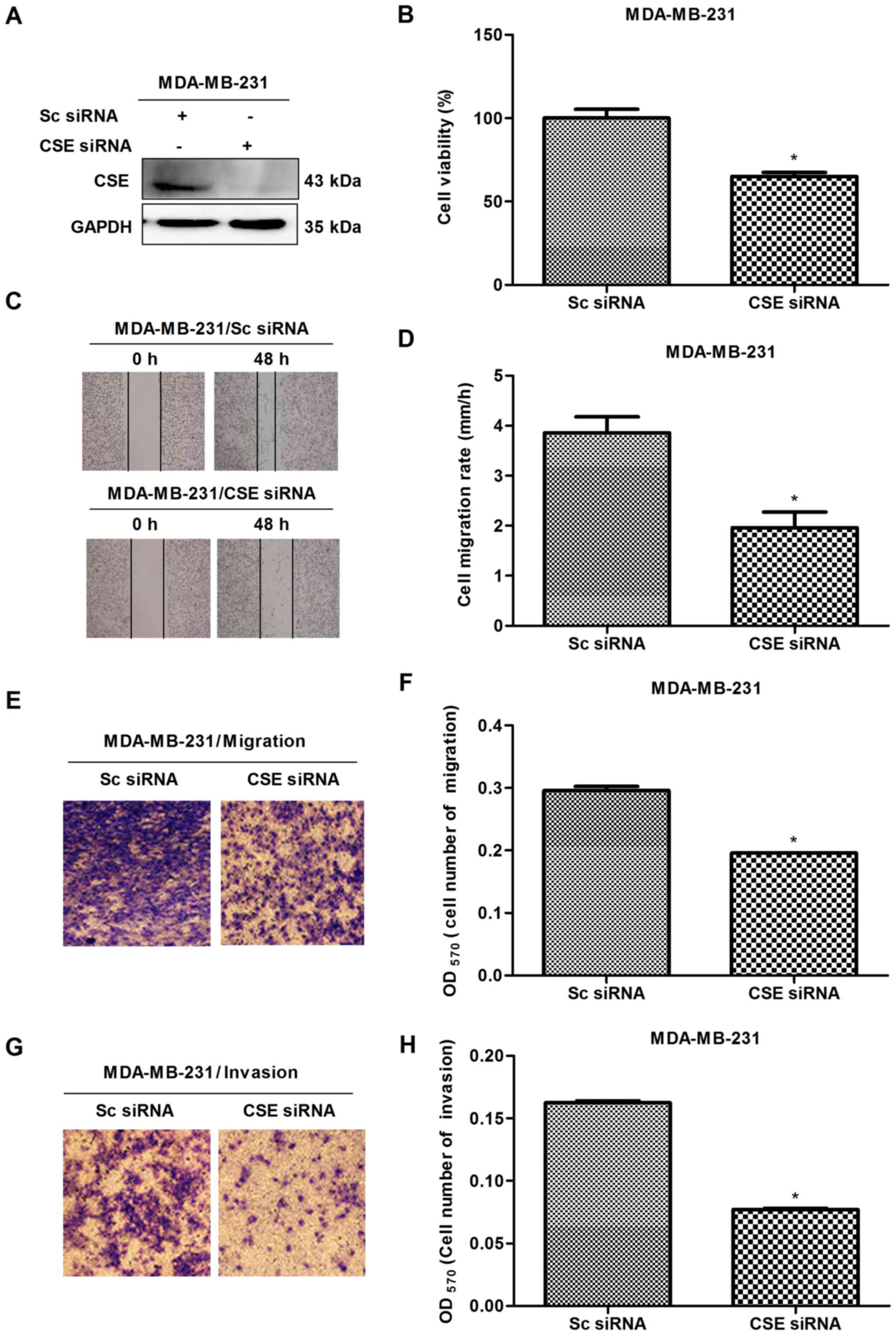
In order to investigate the potential role of CSE in
breast cancer metastasis, CSE was first knocked down with
CSE-specific siRNA in early metastatic MDA-MB-231 breast cancer
cells, following which MTS, scratch and Transwell assays were
performed. The knockdown of CSE was confirmed via western blot
analysis (Fig. 2A). The MTS
results revealed that the knockdown of CSE caused the growth
inhibition of MDA-MB-231 cells (Fig.
2B). The scratch and Transwell assays revealed that CSE
knockdown decreased the migration and invasion rates of MDA-MB-231
cells (Fig. 2C-H). To further
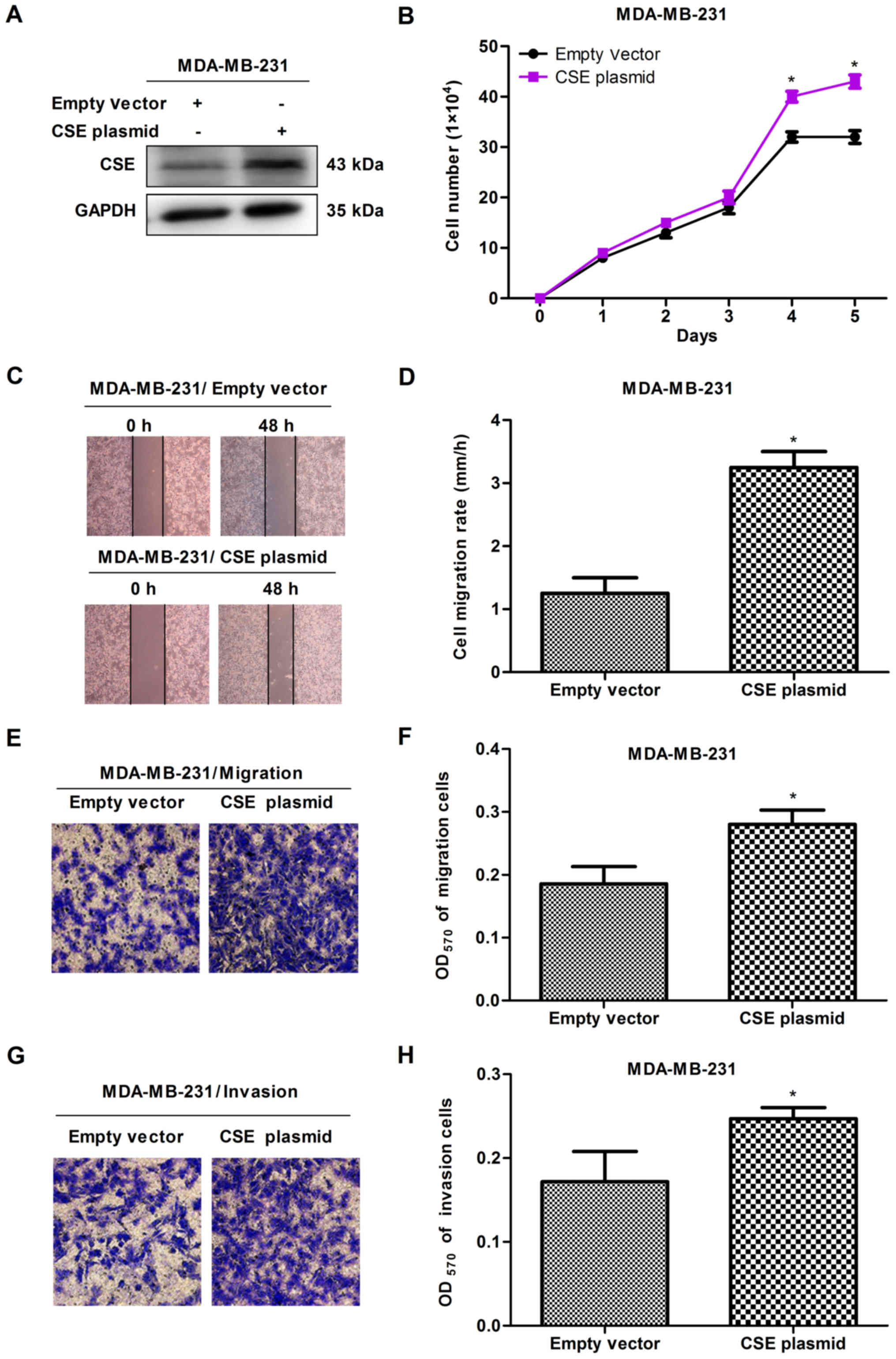
confirm the promoting effects of CSE in breast cancer metastasis, a
gain-of-function cell model was constructed by transfecting a CSE
expression plasmid into MDA-MB-231 cells. The expression of
exogenous CSE was confirmed via western blot analysis (Fig. 3A). The MTS, scratch and Transwell
assays revealed that the overexpression of CSE promoted cell
proliferation (Fig. 3B), migration
and invasion (Fig. 3C-H), compared
with the vector control. These observations suggested that a high
expression of CSE promoted the growth and metastasis of breast
cancer cells.
Expression of CSE promotes human breast
cancer metastasis in nude mice
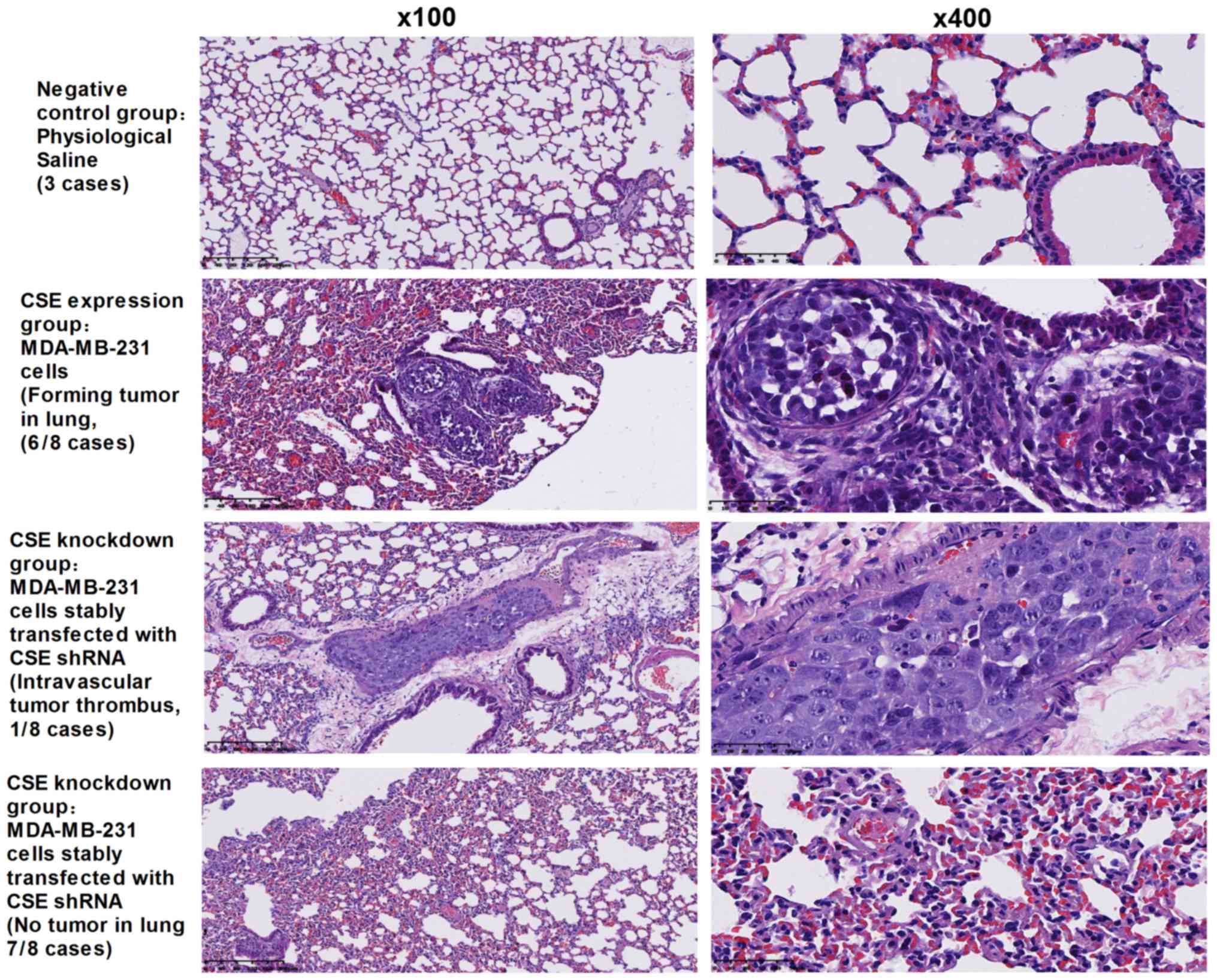
Following demonstration of the promotion of CSE on
cancer cell metastasis in vitro, whether CSE can promote
tumor metastasis in vivo was investigated. A xenograft tumor
model was used to assess the metastasis of MDA-MB-231 human breast
cancer cells with a high expression of CSE and CSE shRNA stable
transfectants of MDA-MB-231 cells in nude mice. As presented in
Fig. 4, the rate of lung
metastasis was 75% in nude mice receiving MDA-MB-231 cells
expressing CSE, whereas the knockdown of CSE in MDA-MB-231 cells
resulted in a significant decrease in the rate of lung metastases
(12.5%) in nude mice and only led to intravascular tumor thrombus.
Collectively, these data indicate that CSE may possess a
significant effect on promoting tumor metastasis in breast
cancer.
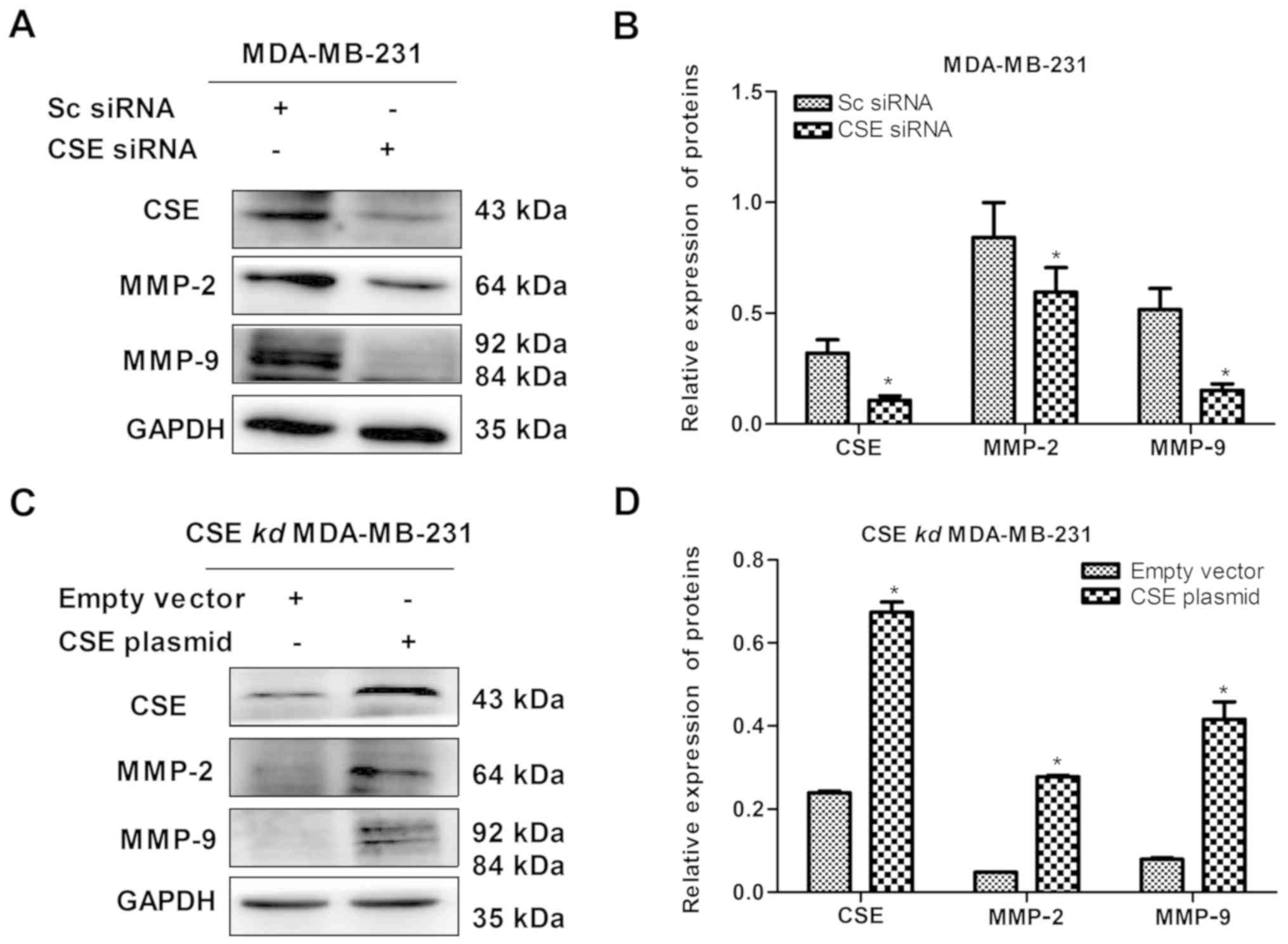
Expression of CSE increases the levels of
MMP-2 and MMP-9 in MDA-MB-231 cells
Following confirmation of the role of CSE in
promoting breast cancer metastasis, the reason why CSE promotes
breast cancer metastasis was investigated. MMPs secreted by cancer
cells can degrade the basement membrane and consequently promote
tumor cells to enter the blood circulation, so as to facilitate the
diffusion and metastasis to the distal end. Therefore, the effect
of the level of CSE on the expression of MMP-2 and MMP-9 in
MDA-MB-231 cells was detected. As presented in Fig. 5A and B, the knockdown of CSE
distinctly decreased the levels of MMP-2 and MMP-9 in MDA-MB-231
cells, whereas the upregulation of CSE increased the levels of
MMP-2 and MMP-9 in the CSE-knockdown MDA-MB-231 cells (Fig. 5C and D). Therefore, it was clear
that increased levels of CSE promoted degradation of the basement
membrane and consequently facilitated the diffusion and metastasis
of breast cancer cells.
Expression of CSE promotes the VEGF
signaling pathways in MDA-MB-231 cells
Angiogenesis is essential for tumor metastasis, and
VEGF, an endothelial cell-specific mitogenic factor, is the most
common tumor angiogenesis-stimulating factor. The effect of the
expression of CSE on the level of VEGF was investigated in
MDA-MB-231 cells. It was demonstrated that the knockdown of CSE
significantly decreased the level of VEGF, whereas the upregulation
of CSE reversed this (Figs. 6A and
B, and 7A and B). In order to
further investigate the effect of CSE on the VEGF downstream
signaling pathway, the expression of key genes were assessed in
MDA-MB-231 cells transfected with CSE siRNA or the CSE
overexpression plasmid. As presented in Fig. 6C-H, CSE knockdown significantly
decrease the levels of key proteins in the PI3K-AKT pathway (PI3K,
Akt and pAkt), FAK-paxillin pathway (FAK and paxillin) and Ras-MAPK
pathway (Ras, Raf, ERK1/2 and pERK1/2). However, it was also
observed that the upregulation of CSE reversed these effects
(Fig. 7C-H). These results suggest
that the expression of CSE promoted the VEGF signaling pathway and
ultimately advanced the metastasis of breast cancer cells.
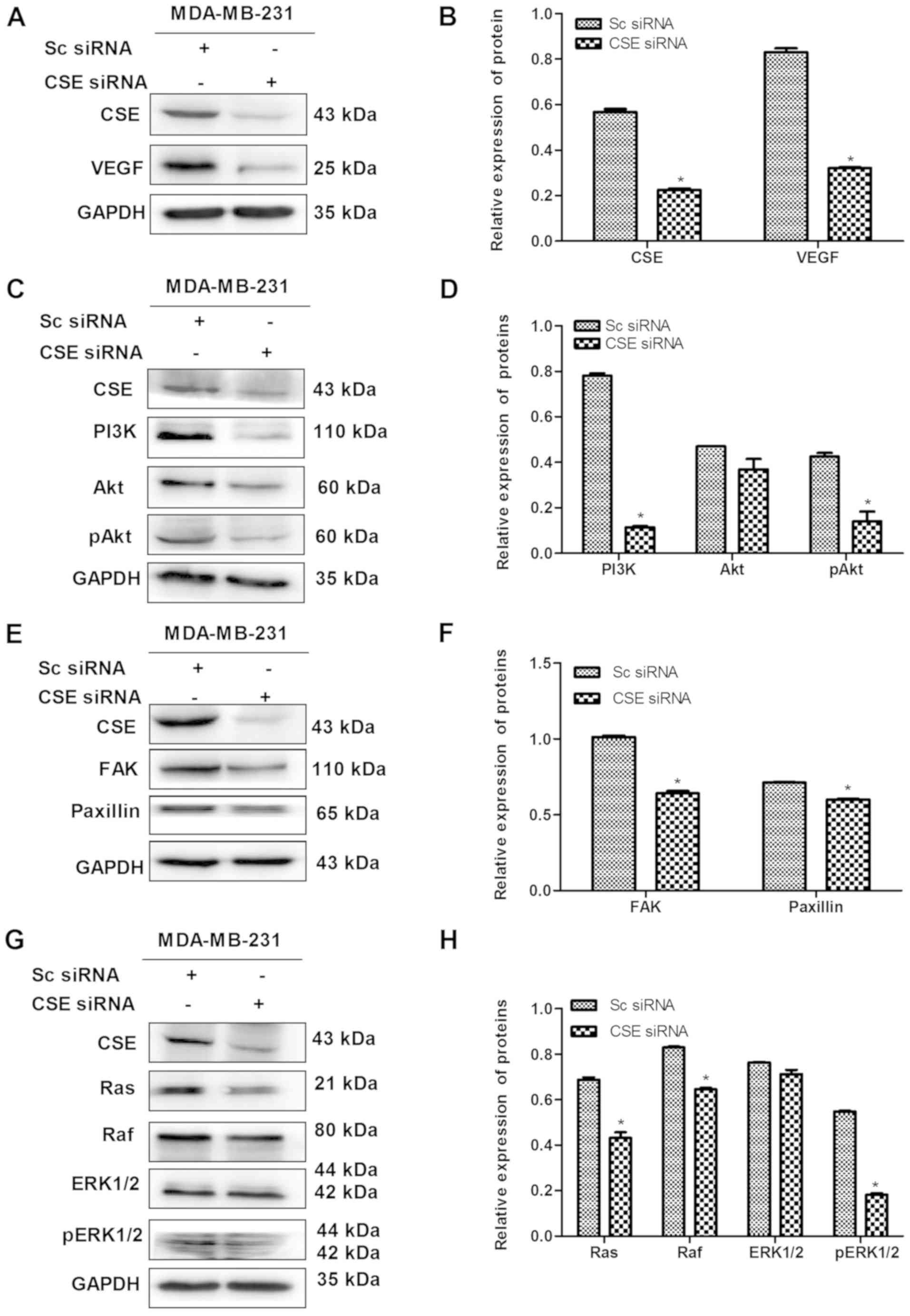 | Figure 6CSE knockdown decreases the
expression of VEGF and its downstream pathway in MDA-MB-231 cells.
(A) Confirmation of the knockdown of CSE by CSE siRNA and (B)
quantification showing the effect of CSE knockdown on the
expression of VEGF. (C) Blot and (D) graph showing the effect of
CSE knockdown on the PI3K pathway. (E) Blot and (F) graph showing
the effect of CSE knockdown on the FAK/paxillin pathway. (G) Blot
and (H) graph showing effect of CSE knockdown on the Ras/Raf/ERK1/2
pathway. *P<0.05 vs. Sc siRNA group. CSE,
cystathionine-γ-lyase; Sc, scrambled; siRNA, small interfering RNA;
VEGF, vascular endothelial growth factor; PI3K,
phosphatidylinositol 3-kinase/protein kinase B; Akt, protein kinase
B; pAkt, phosphorylated Akt; FAK, focal adhesion kinase; Ras, rat
sarcoma protein; Raf, rapidly accelerated fibrosarcoma; ERK,
extracellular signal-regulated protein kinase; pERK, phosphorylated
ERK. |
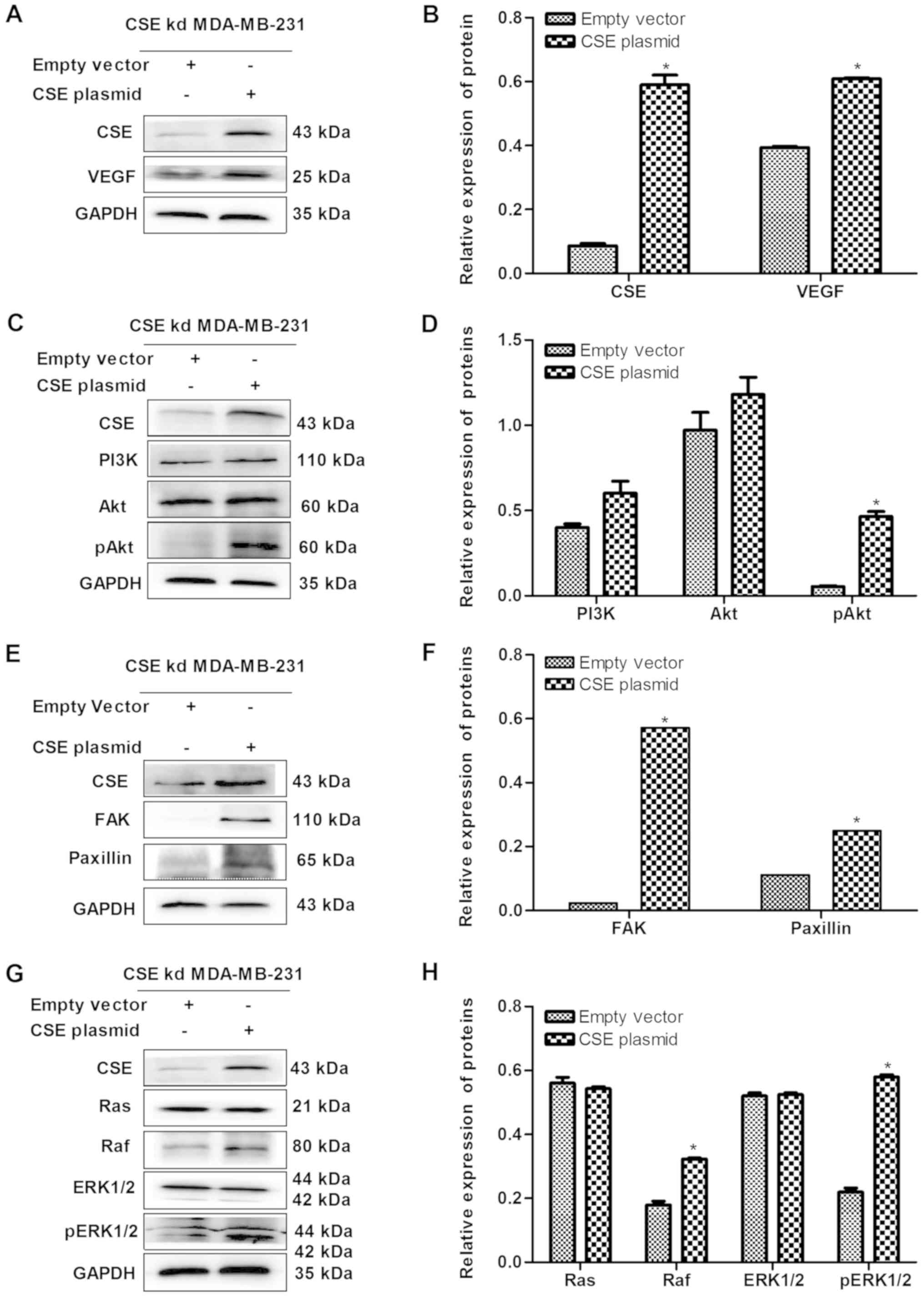 | Figure 7Upregulation of CSE increases the
expression levels of VEGF and its downstream pathway in
CSE-knockdown MDA-MB-231 cells. (A) Blot and (B) graph confirming
of the upregulation of CSE caused by the CSE plasmid and the effect
on VEGF. (C) Blot and (D) graph showing the effect of the
upregulation of CSE on the PI3K/Akt pathway. (E) Blot and (F) graph
showing the effect of the upregulation of CSE on the FAK/paxillin
pathway. (G) Blot and (H) graph showing the effect of the
upregulation of CSE on the Ras/Raf/ERK1/2 pathway.
*P<0.05 vs. empty vector group. CSE,
cystathionine-γ-lyase; VEGF, vascular endothelial growth factor;
PI3K, phosphatidylinositol 3-kinase/protein kinase B; Akt, protein
kinase B; pAkt, phosphorylated Akt; FAK, focal adhesion kinase;
Ras, rat sarcoma; Raf, rapidly accelerated fibrosarcoma; ERK,
extracellular signal-regulated protein kinase; pERK, phosphorylated
ERK; kd, knockdown. |
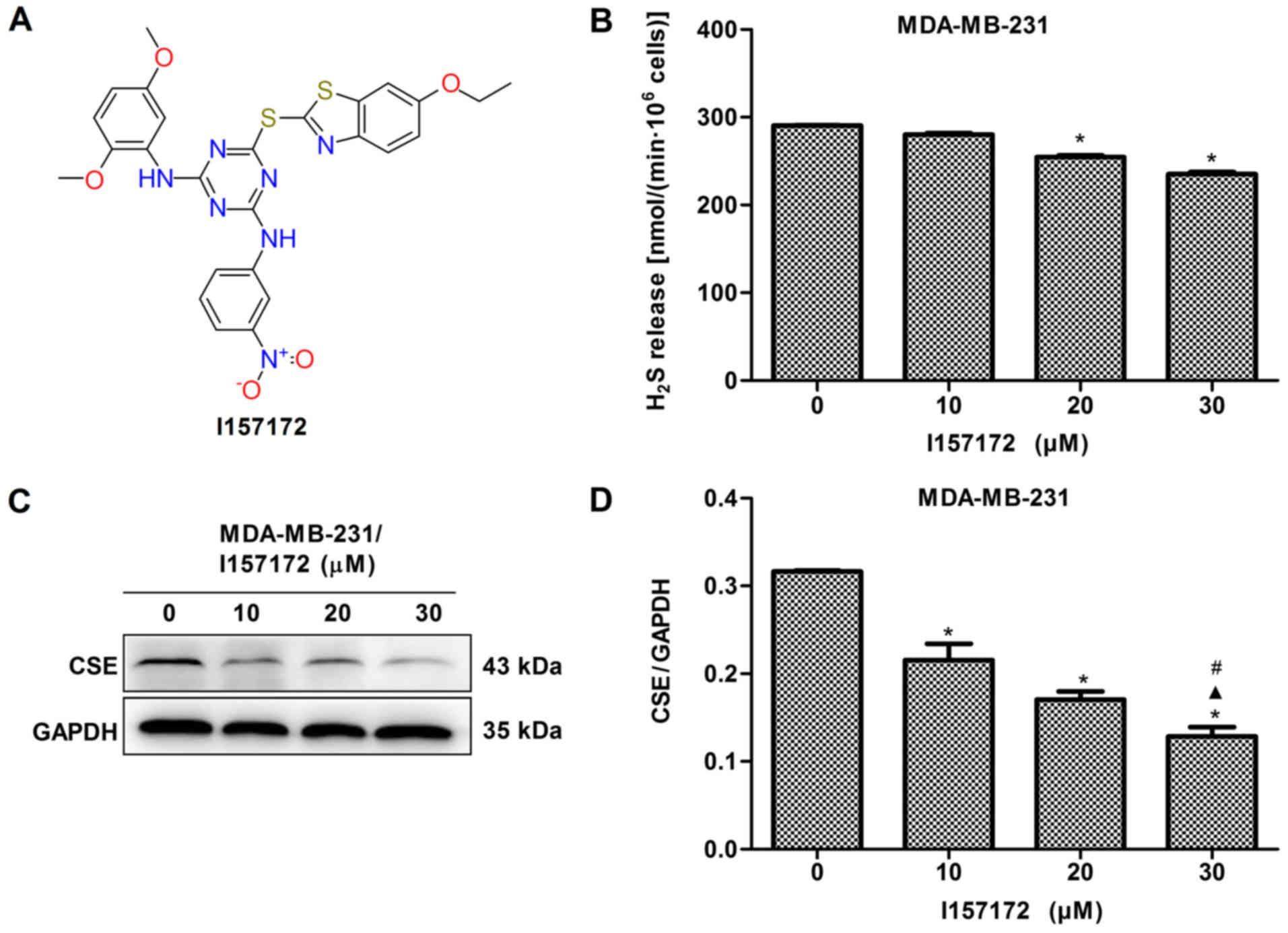
Novel CSE inhibitor I157172 inhibits the
proliferation, migration and invasion of MDA-MB-231 cells
A novel CSE inhibitor I157172 was obtained via
virtual screening and its chemical structure is presented in
Fig. 8A. The inhibitory activity
of I157172 on CSE was confirmed by analysis of the production of
H2S and expression of CSE (Fig. 8B-D). MTS, EdU and Transwell assays
were performed to evaluate the effects of I157172 on the
proliferation, migration and invasion of MDA-MB-231 cells. The data
revealed that I157172 significantly inhibited growth and
proliferation in a dose-dependent manner (Fig. 8E-F) and possessed a 20.36 μM
IC50 value in MDA-MB-231 cells. Decreased migration and
invasion rates were observed in MDA-MB-231 cells treated with
different concentrations of I157172 (Fig. 8G-J). Taken together, the CSE
inhibitor I157172 possessed antiproliferative and anti-metastatic
activities in the early metastatic MDA-MB-231 breast cancer cells,
which further confirmed the role of CSE in breast cancer
metastasis.
Novel CSE inhibitor I157172 inhibits the
VEGF signaling pathway in MDA-MB-231 cells
To further confirm the underlying molecular
mechanism of CSE in promoting breast cancer metastasis, the effect
of novel CSE inhibitor I157172 on the VEGF signaling pathway was
investigated in MDA-MB-231 cells. The data revealed that I157172
significantly inhibited the expression levels of VEGF (Fig. 9A and B) and decreased the levels of
key proteins, including PI3K, Akt, pAkt, FAK, paxillin, Raf and
pERK1/2, of the VEGF downstream pathway in a dose-dependent manner
(Fig. 9C-H). These results
suggested that I157172 inhibited the metastasis of MDA-MB-231 cells
via downregulating the VEGF signaling pathway, which may be one of
the underlying molecular mechanisms by which CSE promotes breast
cancer metastasis.
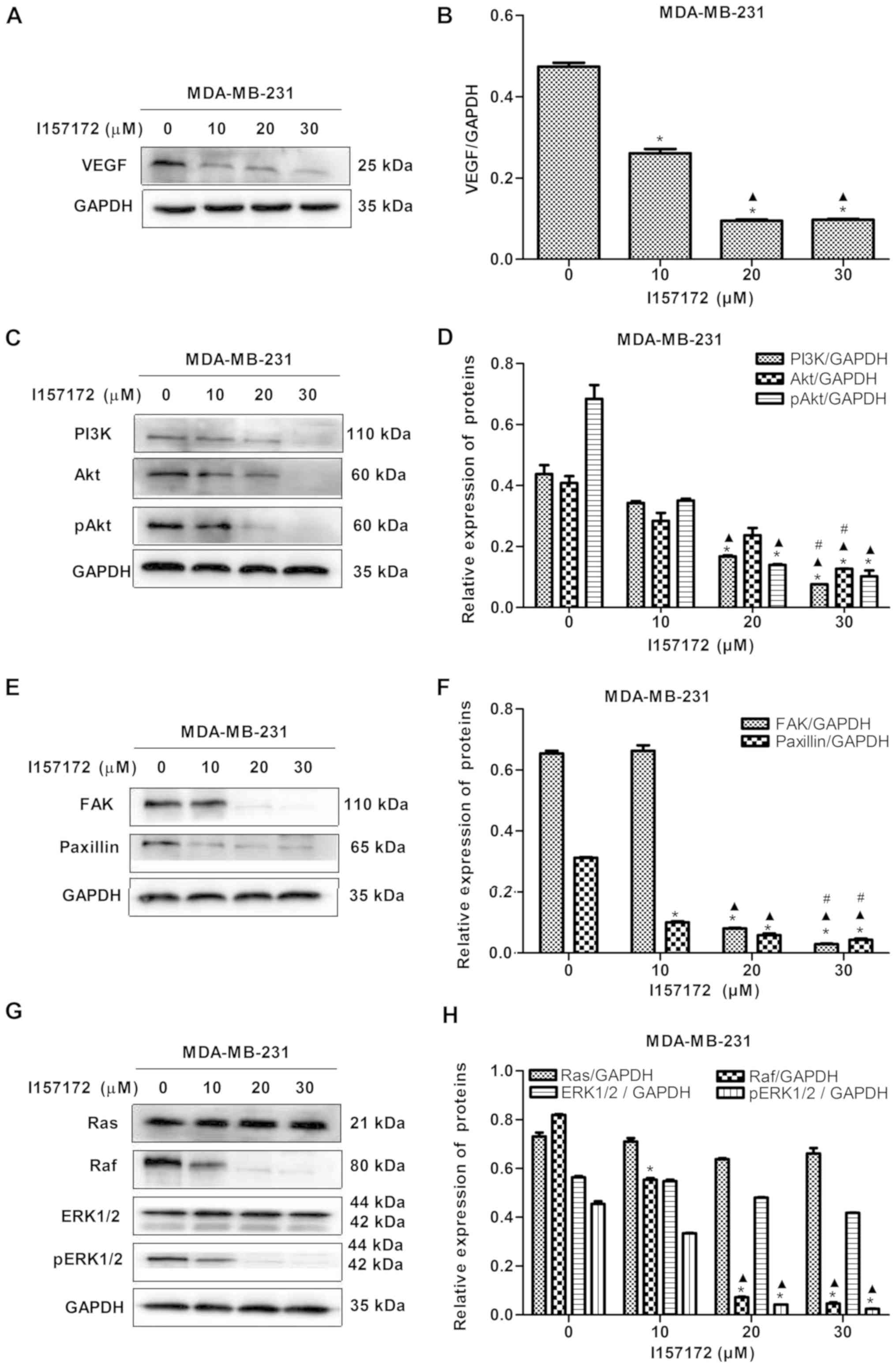 | Figure 9Novel CSE inhibitor I157172 inhibits
the VEGF signaling pathway in MDA-MB-231 cells. (A) Blots and (B)
graph showing the effect of different concentrations of I157172 on
the expression of VEGF. *P<0.05 vs. 0 μM
I157172 group; ▲P<0.05 vs. 10 μM I157172
group. (C) Blots and (D) graph showing the effect of different
concentrations of I157172 on the PI3K/Akt pathway.
*P<0.05 vs. 0 μM I157172 group;
▲P<0.05 vs. 10 μM I157172 group;
#P<0.05 vs. 20 μM I157172 group. (E) Blots and
(F) graph showing the effect of different concentrations of I157172
on the FAK/paxillin pathway. *P<0.05 vs. 0 μM
I157172 group; ▲P<0.05 vs. 10 μM I157172
group; #P<0.05 vs. 20 μM I157172 group. (G)
Blots and (H) graph showing the effect of different concentrations
of I157172 on the Ras/Raf/ERK1/2 pathway. *P<0.05 vs.
0 μM I157172 group; ▲P<0.05 vs. 10 μM
I157172 group. CSE, cystathionine-γ-lyase; VEGF, vascular
endothelial growth factor; PI3K, phosphatidylinositol 3-kinase;
Akt, protein kinase B; FAK, focal adhesion kinase; Ras, rat
sarcoma; Raf, rapidly accelerated fibrosarcoma; ERK, extracellular
signal-regulated protein kinase; pERK, phosphorylated ERK; pAkt,
phosphorylated Akt. |
Discussion
Breast cancer is associated with a high incidence of
metastasis, and breast cancer cells preferentially metastasize to
the bone and lung via the lymphatic system (19,20),
which leads to a poorer patient prognosis. Lymph node metastasis is
the most common site for secondary colonization of breast cancer
cells, and the likelihood of metastatic spread increases with
increasing tumor grade and in hormone receptor-negative cancer.
Metastasis is a complex process and metastatic
dissemination represents the main physiopathology of cancer.
Approximately 90% of breast cancer-associated mortality results
from the development of metastatic disease (21), therefore, numerous studies have
investigated the underlying molecular mechanisms and agents of
breast cancer metastasis. For example, Meehan and Welch (22) showed that the anti-metastatic
protein breast cancer metastasis suppressor-1 provides important
clues regarding the molecular mechanism underlying the cellular
processes of the initiation of metastasis (23). Davison et al (24) investigated the antimigration
potential of signal transduction inhibitors and co-administered
fish oil (24). Although a number
of studies have been performed with the aim of identifying
underlying molecular mechanisms and agents, metastasis remains the
leading cause of mortality in patients with breast cancer.
Previous studies have demonstrated that endogenous
H2S produced by CSE, a main enzyme catalyzing the
endogenous production of H2S, can promote the
proliferation of human cancer cells (14,15)
and contribute to the angiogenic process (12). Angiogenesis is an important concern
in the process of tumor metastasis. Therefore, in the present
study, the role of the CSE/H2S system in breast cancer
metastasis was investigated which, to the best of our knowledge,
revealed for the first time that CSE may promote the metastasis of
breast cancer.
The present study demonstrated that the expression
of CSE was higher in samples from patients with breast cancer
exhibiting lymph node metastasis than in those with no lymph node
metastasis. In addition, higher mRNA and protein levels of CSE were
observed in early metastatic MDA-MB-231 breast cancer cells
compared with those in non-metastatic MCF7 breast cancer cells.
These findings indicate that the metastasis of human breast cancer
may be associated with increased expression levels of CSE.
Triple negative breast cancer (TNBC), characterized
by invasive clinical behavior, has a propensity to metastasize and
establish secondary tumors (25).
The targeted treatment of patients with TNBC has been limited due
the fact that patients with TNBC do not express any of the three
receptors (ER, PR and HER2). The recognition and validation of
novel targets is important for the inhibition of metastasis in
TNBC. Therefore, in the present study, the roles of the expression
of CSE in MDA-MB-231 TNBC cells was investigated. The function of
CSE protein in promoting breast cancer metastasis was confirmed
in vitro and in vivo. The expression of CSE promoted
the growth, migration and invasion of breast cancer cells in
vitro. In further examination of in vivo function, it
was revealed that CSE knockdown inhibited lung metastasis of
MDA-MB-231 in nude mice. It follows that the CSE/H2S
system possesses a function in promoting breast cancer
metastasis.
The present study further assessed the effects of
the expression of CSE on MMP-2 and MMP-9 and on the VEGF signaling
pathway in order to investigate the molecular mechanism underlying
the effect of CSE in promoting breast cancer metastasis. MMP-2 and
MMP-9, secreted by cancer cells, can degrade the basement membrane
and consequently promote tumor cells to enter the blood
circulation, facilitating diffusion and metastasis to the distal
end (26). The results of the
present study suggested that the expression of CSE increased the
levels of MMP-2 and MMP-9 in early metastatic breast cancer cells.
VEGFs constitute a sub-family of growth factors that stimulate the
growth of new blood vessels. Vascular VEGFs are important signaling
proteins involved in vasculogenesis and angiogenesis. VEGF is key
mediator of angiogenesis that is crucial for the development and
metastasis of tumors (27), which
is essential for tumor metastasis. In the present study, it was
revealed that the expression of CSE distinctly decreased the levels
of VEGF and inhibited its downstream pathway.
Based on the function of CSE protein in breast
cancer metastasis, a novel CSE inhibitor I157172 was obtained via
virtual screening, and it was revealed that I157172 possessed
antiproliferative and anti-metastatic activities in early
metastatic MDA-MB-231 breast cancer cells, which further confirmed
the role of CSE in breast cancer metastasis. In addition, I157172
downregulated the VEGF signaling pathway which further confirmed
the mechanism of CSE in promoting breast cancer metastasis.
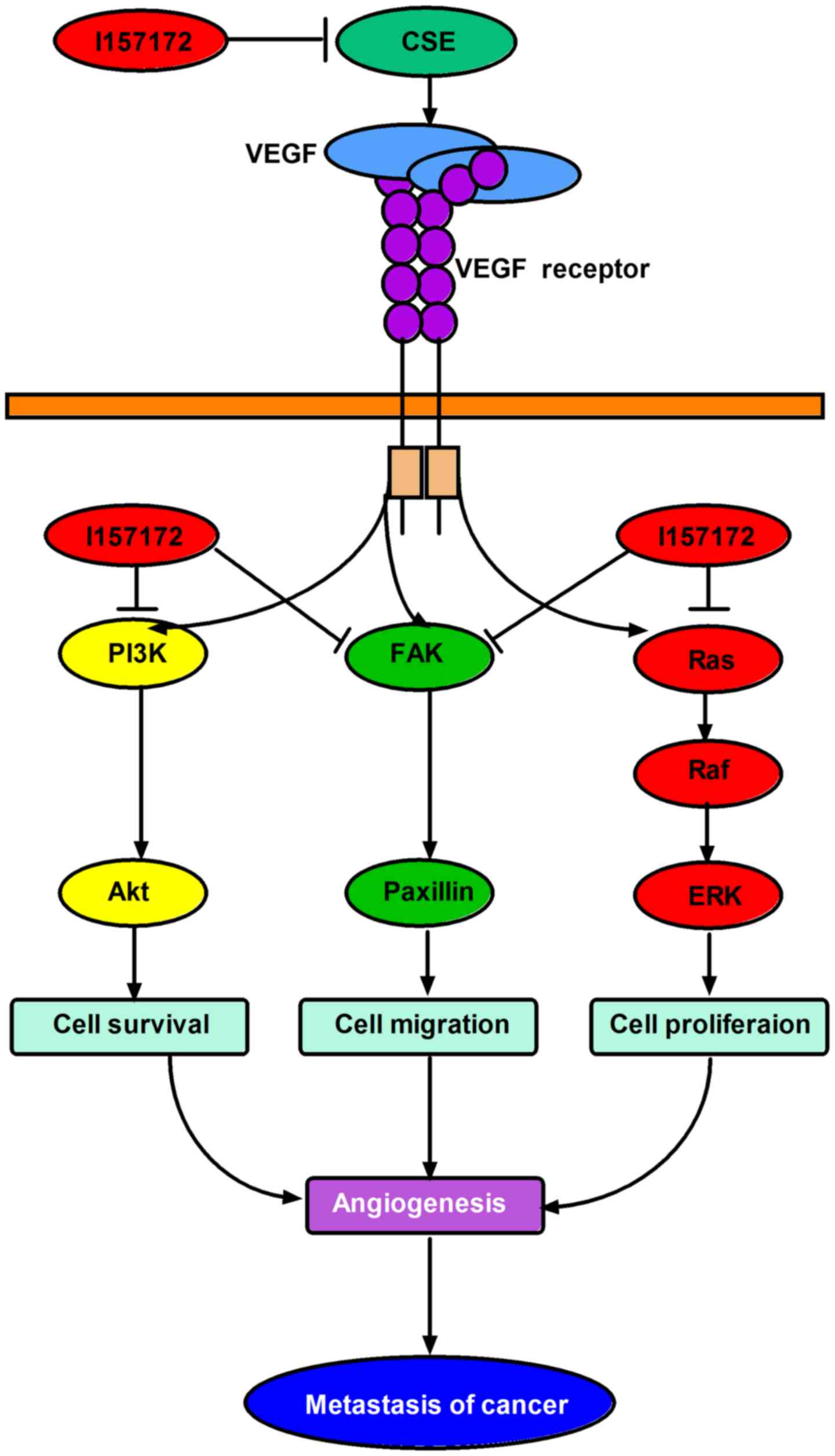
In conclusion, the increased expression levels of
CSE appeared to promote the metastasis of human breast cancer via
the VEGF signaling pathway, and the novel CSE inhibitor I157172
inhibited the metastasis of early metastatic breast cancer cells
via downregulating the VEGF signaling pathway (Fig. 10). This provides novel targets and
agents for the treatment and prognosis of breast cancer. Future
experiments aim to continue to examine the mechanism of CSE in
breast cancer and the anti-breast cancer effect and mechanism of
I157172 in vivo, in order to promote the development of
novel anti-breast cancer drugs.
Acknowledgments
Not applicable.
Funding
The authors would like to acknowledge the financial
assistance provided by the Key Science and Technology Fund of Henan
Province in China (grant no. 162300410035) and the Henan Province
University Science and Technology Innovation Team (grant no.
16IRTSTHN019).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TW and XS conceived and designed the experiments. LW
and HS performed the experiments. LW, HS, YL, WZ, XD and ML
analyzed the data and produced the figures. LW wrote and proofread
the paper. TW and XS revised the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee at the Medical School, Henan University (Kaifeng,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Davidson B, Konstantinovsky S, Nielsen S,
Dong HP, Berner A, Vyberg M and Reich R: Altered expression of
metastasis-associated and regulatory molecules in effusions from
breast cancer patients: A novel model for tumor progression. Clin
Cancer Res. 10:7335–7346. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hagemann T, Robinson SC, Schulz M, Trümper
L, Balkwill FR and Binder C: Enhanced invasiveness of breast cancer
cell lines upon co-cultivation with macrophages is due to TNF-α
dependent up-regulation of matrix metalloproteases. Carcinogenesis.
25:1543–1549. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hiscox S, Jordan NJ, Jiang W, Harper M,
McClelland R, Smith C and Nicholson RI: Chronic exposure to
fulvestrant promotes overexpression of the c-Met receptor in breast
cancer cells: Implications for tumour-stroma interactions. Endocr
Relat Cancer. 13:1085–1099. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pecorino L: Molecular Biology of Cancer.
Oxford University Press; Oxford: pp. p4002005
|
|
5
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Papapetropoulos A, Pyriochou A, Altaany Z,
Yang G, Marazioti A, Zhou Z, Jeschke MG, Branski LK, Herndon DN,
Wang R, et al: Hydrogen sulfide is an endogenous stimulator of
angiogenesis. Proc Natl Acad Sci USA. 106:21972–21977. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen DB, Feng L, Hodges JK, Lechuga TJ and
Zhang H: Human trophoblast-derived hydrogen sulfide stimulates
placental artery endothelial cell angiogenesis. Biol Reprod.
97:478–489. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang R: Hydrogen sulfide: The third
gasotransmitter in biology and medicine. Antioxid Redox Signal.
12:1061–1064. 2010. View Article : Google Scholar
|
|
9
|
Kimura Y, Goto Y and Kimura H: Hydrogen
sulfide increases glutathione production and suppresses oxidative
stress in mitochondria. Antioxid Redox Signal. 12:1–13. 2010.
View Article : Google Scholar
|
|
10
|
Sheng J, Shim W, Wei H, Lim SY, Liew R,
Lim TS, Ong BH, Chua YL and Wong P: Hydrogen sulphide suppresses
human atrial fibroblast proliferation and transformation to
myofibro-blasts. J Cell Mol Med. 17:1345–1354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Popov D: An outlook on vascular hydrogen
sulphide effects, signalling, and therapeutic potential. Arch
Physiol Biochem. 119:189–194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Katsouda A, Bibli SI, Pyriochou A, Szabo C
and Papapetropoulos A: Regulation and role of endogenously produced
hydrogen sulfide in angiogenesis. Pharmacol Res. 113:175–185. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sbodio JI, Snyder SH and Paul BD:
Regulators of the transsulfu-ration pathway. Br J Pharmacol.
176:583–593. 2019. View Article : Google Scholar
|
|
14
|
Yin P, Zhao C, Li Z, Mei C, Yao W, Liu Y,
Li N, Qi J, Wang L, Shi Y, et al: Sp1 is involved in regulation of
cystathionine γ-lyase gene expression and biological function by
PI3K/Akt pathway in human hepatocellular carcinoma cell lines. Cell
Signal. 24:1229–1240. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cai WJ, Wang MJ, Ju LH, Wang C and Zhu YC:
Hydrogen sulfide induces human colon cancer cell proliferation:
Role of Akt, ERK and p21. Cell Biol Int. 34:565–572. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
You J, Shi X, Liang H, Ye J, Wang L, Han
H, Fang H, Kang W, Wang T, Han H, et al: Cystathionine-γ-lyase
promotes process of breast cancer in association with STAT3
signaling pathway. Oncotarget. 8:65677–65686. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fendri A, Kontos CK, Khabir A,
Mokdad-Gargouri R, Ardavanis A and Scorilas A: Quantitative
analysis of BCL2 mRNA expression in nasopharyngeal carcinoma: An
unfavorable and independent prognostic factor. Tumour Biol.
31:391–399. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nitti M, Piras S, Marinari UM, Moretta L,
Pronzato MA and Furfaro AL: HO-1 Induction in Cancer Progression: A
Matter of Cell Adaptation. Antioxidants. 6:292017. View Article : Google Scholar :
|
|
19
|
Cunnick GH, Jiang WG, Douglas-Jones T,
Watkins G, Gomez KF, Morgan MJ, Subramanian A, Mokbel K and Mansel
RE: Lymphangiogenesis and lymph node metastasis in breast cancer.
Mol Cancer. 7:232008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fidler IJ: The pathogenesis of cancer
metastasis: The 'seed and soil' hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lambert AW, Pattabiraman DR and Weinberg
RA: Emerging biological principles of metastasis. Cell.
168:670–691. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Meehan WJ and Welch DR: Breast cancer
metastasis suppressor 1: Update. Clin Exp Metastasis. 20:45–50.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Ye L, Tan Y, Sun P, Ji K and
Jiang WG: Expression of breast cancer metastasis suppressor-1,
BRMS-1, in human breast cancer and the biological impact of BRMS-1
on the migration of breast cancer cells. Anticancer Res.
34:1417–1426. 2014.PubMed/NCBI
|
|
24
|
Davison Z, Nicholson RI, Hiscox S and
Heard CM: Co-administration of fish oil with signal transduction
inhibitors has anti-migration effects in breast cancer cell lines,
in vitro. Open Biochem J. 12:130–148. 2018. View Article : Google Scholar :
|
|
25
|
Jitariu AA, Cîmpean AM, Ribatti D and
Raica M: Triple negative breast cancer: The kiss of death.
Oncotarget. 8:46652–46662. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu H, Zeng Z, Wang S, Li T, Mastriani E,
Li QH, Bao HX, Zhou YJ, Wang X, Liu Y, et al: Main components of
pomegranate, ellagic acid and luteolin, inhibit metastasis of
ovarian cancer by down-regulating MMP2 and MMP9. Cancer Biol Ther.
18:990–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carmeliet P: VEGF as a key mediator of
angiogenesis in cancer. Oncology. 69(Suppl 3): 4–10. 2005.
View Article : Google Scholar : PubMed/NCBI
|