Introduction
Lung cancer is widely acknowledged to be among the
most severe and most common malignancies worldwide, with ~228,150
newly diagnosed cases and 142,670 estimated deaths in the United
States in 2019 (1). Non-small cell
lung cancer (NSCLC) accounts for ~85% of all lung cancer cases and
is the predominant form of the disease largely attributed to
cancer-related deaths. Fortuitously, more effective treatment
protocols have been developed in recent years that have aided in
the ultimate goal of cancer remission, including improved screening
methods, surgical procedures, chemoradiotherapy and diagnostic
evaluation (2). However, despite
the substantial progress in the last few years, the majority of
NSCLC prognoses remain poor, with a 10% 5-year survival rate
(3). Additionally, patients with
NSCLC have been widely documented to experience poor sleep quality
and unexpected sleep disturbances (4).
Obstructive sleep apnea-hypopnea (OSAH) syndrome is
a condition characterized by various respiratory events during a
period of sleep accompanied by loud snoring, excessive daytime
sleepiness or breathing pauses (5). The morbidity of this syndrome is 3-7%
among males and 2-5% among females, and is as high as 33% in
patients from Sao Paulo with low socio-economic status, with a
apnea-hypopnea index between 5 to 15; however, research into this
condition has been inadequate in recent years (6). Thus, the current study sought to
identify a novel therapeutic approach in regard to the involvement
of OSAH model in NSCLC.
Connexin (CX) 26 is a transmembrane protein that is
part of gap junctions between adjacent cells and its connection
with the PI3K/Akt signaling pathway leads to the acquired gefitinib
resistance in NSCLC cells through the activation of
epithelial-mesenchymal transition (EMT) (7). More specifically, CX26 has been
reported to contribute to the proliferation of epithelial cells
involved in basal airway repair (8). The CX-CX interaction mechanism has
been revealed to be important in intercellular communication
regulation; for instance, CX26 can be co-expressed with CX43 in
human skin (9). Researchers have
highlighted the role of CX26 and CX43 signaling in the suppression
of tumor progression (10). In
recent years, the interaction between the tumor suppressor gene P53
and its negative regulator, murine double minute-2 (MDM2), has been
reported as a potential therapeutic target for patients with cancer
(11). P53 has been demonstrated
to induce apoptosis through blockade of cellular DNA damage repair,
while the transactivation ability and stability of P53 can be
repressed by MDM2 through ubiquitination (12). Based on the aforementioned
literature, the present study subsequently asserted the hypothesis
that an OSAH model may induce the internalization of CX26 and CX43
in pulmonary epithelial cells (PECs) with the involvement of the
P53/MDM2 signaling pathway.
Materials and methods
Ethics statements
The study was approved by the Ethics Committee of
The First Affiliated Hospital of Nanchang University. Written
informed consent was obtained from each participant. All animal
experimentation in this study was approved by the Institutional
Animal Care and Use Committee of The First Affiliated Hospital of
Nanchang University.
Study subjects
In total, 120 specimens (cancer pulmonary epithelial
tissues and paracancerous tissues) were obtained from 60 patients
with pathologically confirmed NSCLC that had previously undergone
surgical resection in The First Affiliated Hospital of Nanchang
University between January 2010 and December 2015. There were 38
males and 22 females with a mean age of 62.4±5.9 years. None of the
patients had OSAH. Each patient was free of any
immunologically-mediated disease, connective tissue disease or
comorbidity of significant organs, and did not receive anticancer
therapy, including chemoradiotherapy or immunotherapy prior to
surgery. The paracancerous tissues were confirmed to lack any
pathologic features of cancerization according to hematoxylineosin
(HE) staining.
Cell model establishment with
hypoxia
Following isolation from one patient and culture
(13), PECs derived from human
NSCLC were washed twice with PBS and cultured in DMEM/F12 (cat. no.
GNM-12500-S; Shanghai Jingke Chemical Technology Co., Ltd.)
containing 20% FBS (Sigma-Aldrich; Merck KGaA) in an incubator with
5% CO2 at 37°C. The medium was changed once every 3 days
as soon as the majority of the epithelial cells were detached. In
the event that cell confluence reached 80-90%, the medium was
subsequently discarded. The cells were then washed twice with PBS
and detached following treatment with 0.25% trypsin, after which
the density was adjusted to 1×106 cells/ml using DMEM
containing 10% calf serum (Sigma-Aldrich; Merck KGaA). The cells
were then further cultured in the plate and dish.
An in vitro model was established by exposing
the cells to hypoxic conditions over an extended period. The cells
were assigned to two groups: Hypoxia and normal group. Intermittent
hypoxia under normal pressure and temperature was applied in in
vitro model, which was established to mimic the conditions of
chronic intermittent hypoxia that occurs in the body due to snoring
during the night. A transparent plexiglass box was used (5 mm in
thickness; 75×50×50 cm) in which nitrogen (>95.5%) was input
circularly (~8 min/cycle) by the experimenter. The oxygen
concentration in the intermittent hypoxia box was monitored using
an oxygen monitor to control the amount of air-input and
air-removal. The minimum oxygen concentration required was 8.5%
during each cycle. With the removal of hypoxic gas and the input of
air (oxygen concentration, 20.9%), the oxygen concentration was
gradually restored to 21%. The volume fraction of oxygen in the gas
mixture was adjusted on a timely basis to 10±1.5% under the dynamic
monitoring of the gas analyzer (OX-100A; Jiande Meicheng Analysis
Instrument Factory). The water and CO2 content in the
intermittent hypoxia box were absorbed using anhydrous calcium
chloride and CO2 absorbent, both of which, along with
water and padding in the hypoxia box, were changed every 2 days.
Cells in the normal group were cultured under normal pressure and
temperature with O2 (volume fraction, 21%) between 10:00
p.m. and 06:00 a.m. each day in the dark. Cells in the hypoxia
group were cultured under intermittent hypoxia with O2
(volume fraction, 8.5%) between 10:00 p.m. and 06:00 a.m. each day
in the dark (14-16).
Cell treatment
All plasmids used for transfection were purchased
from Sangon Biotech Co., Ltd. The siRNA was expressed from plasmid
vectors. Hsp70 siRNA expression vector was constructed by
psuperpuro expression system (a primitive siRNA design) and
overexpression vector was constructed using pcDNA3.1-GALT-GFP.
Cells in the exponential growth phase were seeded into a 24-well
plate at 500 µl/well with a final cell density adjusted to
1.2×105 cells/ml. The transfection was performed in
accordance with the instructions of the Lipofectamine®
2000 kit (cat. no. 11668-019; Invitrogen; Thermo Fisher Scientific,
Inc.). Briefly, 100 pmol si-CX26, si-CX43, CX26, CX43 and negative
control (NC), or target plasmid were diluted in 250 µl
serum-free Opti-MEM (Gibco; Thermo Fisher Scientific, Inc.; final
concentration of 50 nM in the cells), mixed and incubated at room
temperature for 5 min; 4 µg was used for plasmids. An
additional 250 µl serum-free Opti-MEM was used to dilute 5
µl Lipofectamine 2000 followed by a 5-min incubation at room
temperature. The two solutions were then mixed, incubated at room
temperature for 20 min and added to the wells. Following incubation
at 37°C with CO2 for 6-8 h, the cells were further
cultured in complete culture medium for 48 h for the following
experiments. The sequences of the siRNAs were as follows: si-CX26,
5′-CCC AGU UGU UAG AUU AAG ATT-3′; si-CX26-NC, 5′-UUC UCC GAA CGU
GUC ACG UTT-3′; si-CX43, 5′-AGA GAU ACG AAC AAG AGA G-3′;
si-CX43-NC, 5′-AUA CAA CAG AGA GGG AAA G-3′.
The cells were assigned to 14 groups, as follows:
Normal (PECs exposed to normal oxygen); hypoxia (PECs exposed to
hypoxia); hypoxia + lactacystin (6 µM proteasome inhibitors)
(17); normal + lactacystin (6
µM proteasome inhibitors); si-NC-CX26 + si-NC-CX43 (PECs
transfected with NC), si-CX26 + si-NC-CX43 (PECs transfected with
si-CX26 and NC of si-CX43); si-NC-CX26 + si-CX43 (PECs transfected
with si-CX43 and NC of si-CX26); si-CX26 + si-CX43 (PECs
transfected with si-CX26 and si-CX43); NC-CX26 + NC-CX43 + hypoxia;
CX26 + NC-CX43 + hypoxia; NC-CX26 + CX43 + hypoxia; CX26 + CX43 +
hypoxia; hypoxia + monodansylcadaverine (MDC; 200 µmol/l
protein internalization inhibitors) (18); and normal + MDC. A flow-chart
(Fig. S1) was created to
demonstrate the experimental design.
Immunohistochemistry for CX26 and CX43
expression
The specimens were fixed in 10% formaldehyde at room
temperature for 5 min, embedded in paraffin and continuously sliced
at a thickness of 4 µm. The tissue sections were heated in
an oven at 60°C for l h, followed by conventional xylene
deparaffinization and dehydration in gradient alcohol (100, 95 and
80%) at room temperature for 10-15 min each. The slices were then
incubated in 3% H2O2 (Sigma-Aldrich; Merck
KGaA) at 37°C for 30 min, rinsed with PBS and boiled in 0.01 M
citrate buffer at 95°C for 20 min. After they had cooled to room
temperature, the slices were washed with PBS and then blocked with
10% normal goat serum (Sigma-Aldrich; Merck KGaA) at 37°C for 10
min. Subsequently, the slices were incubated with the following
primary antibodies overnight at 4°C: 50 µl rabbit anti-mouse
CX26 (1:100; cat. no. ab65969; Abcam) and CX43 (1:1,000; cat. no.
ab11370; Abcam). After they were rinsed in PBS, the sections were
incubated with the goat anti-rabbit IgG secondary antibody (cat.
no. ab150077; 1:1,000; Abcam) for 30 min at 37°C, which was
followed by three washes in PBS (3 min each time). Next, the slices
were developed using 3,3′-diaminobenzidine (Sigma-Aldrich; Merck
KGaA) at room temperature for 5 min, counterstained in 0.5%
hematoxylin (Shanghai Bogoo Biotech Co., Ltd.) at room temperature
for 3 min and sealed. Slides for which PBS was used instead of the
primary antibody served as negative controls, while normal tissue
served as the positive control. The samples were observed under an
optical microscope (XSP-36, Shenzhen Boshida Optical Instrument
Co., Ltd.) and images were obtained. Five high-power fields (×200)
were randomly selected from each slice, and 100 PECs were counted
in each field. Fields with <10% positive cells were considered
negative, while those with ≥10% positive cells were deemed positive
(19).
Immunofluorescence for cellular
localization of CX26 and CX43
Following conventional detachment and transfection,
the cells were counted and cultured in immunofluorescence chambers
at a density of 2×105 cells/well. When cell confluence
reached ~90%, the cells were washed three times with PBS on ice and
fixed in 1 ml 4% paraformaldehyde at room temperature for 15 min,
followed by an additional three washes in PBS. The cells were then
treated with 0.3% Triton for 10 min at 37°C, followed by another
three washes in PBS and blocking in goat serum (Sigma-Aldrich;
Merck KGaA) for 30 min at 37°C. Next, the primary antibodies
(prepared in PBS) against CX26 (cat. no. 33-5800; 1:400; Thermo
Fisher Scientific, Inc.) and CX43 (cat. no. ab11370; 1:1,000;
Abcam) were added and incubated with the cells overnight at 4°C.
After three washes in PBS, the secondary antibody to Alexa Fluor
488 (cat. no. A11029; 1:500; Invitrogen; Thermo Fisher Scientific,
Inc.) was added and incubated with the sample at room temperature
for 1 h in the dark. The cells were then washed three times with
PBS, stained with DAPI staining solution for 15 min in the dark and
washed an additional three times with PBS. A fluorescence-quenching
agent was added for sealing. The samples were then finally observed
and images under a fluorescence microscope.
Co-immunoprecipitation (Co-IP) for the
binding of CX26 and CX43 to ubiquitin protein
The cells were washed twice with pre-cooled PBS
followed by the removal of PBS. After the addition of pre-cooled
RIPA buffer, the cells were scraped with a cell scraper. The
suspension was subsequently transferred to a 1.5 ml Eppendorf tube
and continuously shaken for 15 min in a gentle manner at 4°C, which
was followed by a centrifugation at 14,000 × g at 4°C for 15 min.
The supernatant was then transferred to a new centrifuge tube.
Ubiquitin agarose beads (Thermo Fisher Scientific, Inc.) were
prepared and washed twice with PBS. The concentration was then
adjusted to 50% using PBS. For every 1 ml total protein, 100
µl 50% ubiquitin agarose beads were added, which were then
shaken for 10 min at 4°C to remove non-specific hybrid protein and
to reduce background signal. Following centrifugation at 14,000 × g
at 4°C for 15 min, the supernatant was then transferred to a new
centrifuge tube, and the ubiquitin agarose beads were removed.
Total protein was diluted 10-fold. A protein standard curve was
plotted using the Bradford method followed by determination of the
protein concentration. A total of 200 µl diluted total
protein (10 µg/µl) was added to 0.2 µl primary
antibody against ubiquitin (cat. no. ab19247; Abcam) and shaken
gently for 1 h at 4°C. Subsequently, 20 µl ubiquitin agarose
beads were added for incubation at 4°C overnight. Centrifugation
was then performed at 1,500 × g for 5 min followed by the
collection of the agarose beads-antigen-antibody complex and the
removal of the supernatant. The complex was then washed three times
with 800 µl pre-cooled PBS, suspended in 60 µl 2X
loading buffer, gently mixed together and boiled for 5 min. Then,
the expression of CX26 and CX43 was analyzed by western blot
analysis.
Reverse transcription-quantitative PCR
(RT-qPCR) for determination of the expression of CX26, CX43 and
downstream target genes of the P53/MDM2 signaling pathway
RT-qPCR was applied to determine the expression of
CX26, CX43 and downstream target genes of the P53/MDM2 signaling
pathway. The primers of the downstream target genes were designed
using Primer 5.0 software (Premier Biosoft International) and were
analyzed for homology using BLAST software (blast.ncbi.nlm.nih.gov/Blast.cgi) with the related
reference sequences provided by GenBank (ncbi.nlm.nih.gov/genbank/), as depicted in Table I. Following transfection, the cells
were collected, from which total RNA was extracted using a TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). Next, cDNA was
obtained through RT in accordance with the instructions of
RevertAid First Strand cDNA Synthesis Kit (cat. no. K1622; Thermo
Fisher Scientific, Inc.), which was followed by PCR amplification
using SYBR® Premix Ex TaqTM II (Perfect Real Time) kit
(Takara Bio, Inc.). The reaction conditions were as follows:
Pre-denaturation at 95°C for 5 min, 45 cycles of denaturation at
95°C for 20 sec, annealing at 60°C for 1 min and extension at 72°C
for 30 sec. The qPCR results were subsequently analyzed using the
2−ΔΔCq method (20) to
determine the relative expression of the target genes at the
transcriptional level. β-actin was regarded as the internal
reference for CX26, CX43, β-catenin, E-cadherin, Vimentin, P53, P21
and MDM2. The experiment was repeated three times.
 | Table IPrimers for reverse
transcription-quantitative PCR. |
Table I
Primers for reverse
transcription-quantitative PCR.
| Name | Primer sequence
(5′-3′) |
|---|
| β-actin | Forward:
CATGTACGTTGCTATCCAGGC |
| Reserve:
CTCCTTAATGTCACGCACGAT |
| E-cadherin | Forward:
CGAGAGCTACACGTTCACGG |
| Reserve:
GGGTGTCGAGGGAAAAATAGG |
| N-cadherin | Forward:
TCAGGCGTCTGTAGAGGCTT |
| Reserve:
ATGCACATCCTTCGATAAGACTG |
| CX26 | Forward:
TCGCATTATGATCCTCGTTGTG |
| Reserve:
GGGGAAGTAGTGATCGTAGCAC |
| CX43 | Forward:
GGTGACTGGAGCGCCTTAG |
| Reserve:
GCGCACATGAGAGATTGGGA |
| P53 | Forward:
CAGCACATGACGGAGGTTGT |
| Reserve:
TCATCCAAATACTCCACACGC |
| P21 | Forward:
TGTCCGTCAGAACCCATGC |
| Reserve:
AAAGTCGAAGTTCCATCGCTC |
| MDM2 | Forward:
GAATCATCGGACTCAGGTACATC |
| Reserve:
TCTGTCTCACTAATTGCTCTCCT |
Western blot analysis for protein
expression measurement
The cells in each group were collected, and the
total protein was extracted. The concentration was determined using
a BCA kit (cat. no. 20201ES76; Yeasen Biotechnology Co., Ltd.). The
amount of protein was adjusted to 20 µg per well. Following
SDS-PAGE on 8% gels for 1 h, the separated protein band was
transferred to a PVDF membrane. The membrane was then blocked with
5% skim milk powder at room temperature for 1 h and incubated with
rabbit polyclonal antibodies, including ubiquitin (cat. no.
ab140601; 1:500), β-actin (cat. no. ab8227; 1:1,000), CX26 (cat.
no. ab65969; 1:1,000), CX43 (cat. no. ab11370; 1:8,000), P53
(ab131442; 1:500), P21 (ab109199; 1:1,000), MDM2 (cat. no. ab38618;
1:1,000), E-cadherin (cat. no. ab76055; 1:1,000), Vimentin (cat.
no. ab8978; 1:1,000), Oct4 (cat. no. ab181557; 1:1,000) and Nanog
(cat. no. ab218524; 1:1,000) overnight at 4°C. After three washes
in PBS with Tween-20 (PBST), the membrane was incubated with the
secondary antibody, horseradish peroxidase-labeled goat anti-rabbit
IgG (cat. no. ab150077; 1:1,000) at room temperature for 1 h,
followed by an additional three washes in PBST. All were purchased
from Abcam. The membrane was then immersed in the enhanced
chemiluminescence detection solution (cat. no. ECL808-25; Biomiga,
Inc.) in the dark. The images were visualized using a SmartView Pro
2000 (UVCI-2100; Major Science). The relative protein levels were
expressed by the gray value of the target protein band to that of
the β-actin protein band using the Quantity One v4.6.2 software
(Bio-Rad Laboratories, Inc.).
Transwell assay
The cells in the logarithmic growth phase were
starved for 24 h, detached, centrifuged, and resuspended to adjust
the final concentration into 2×105 cells/ml. The
extracellular matrix (ECM) gel was allowed to stand at 4°C
overnight and diluted with serum-free medium at a ratio of 1:9 to
reach a final concentration of 1 mg/ml. The diluted ECM gel (40
µl) was added to the polycarbonate membrane of each 24-well
Transwell apical chamber and incubated in a 5% CO2
incubator at 37°C for 5 h. After the ECM gel was polymerized to
form a gel, the DMEM was added (70 µl each chamber). After
another incubation in a 37°C incubator for 0.5 h, the ECM gel was
rehydrated. The cells in each group were starved for 24 h,
detached, centrifuged and resuspended with FBS-free DMEM to adjust
the final concentration into 2.5×105 cells/ml. A total
of 0.2 ml suspension was added into the apical chamber in which the
basement membrane has been hydrated, while 700 µl
pre-chilled DMEM containing 10% FBS was supplemented into the
basolateral chamber. The Transwell chamber was then cultured in a
5% CO2 incubator at 37°C for 24 h. The cells in the
chamber and basement membrane were removed by wet cotton swabs,
fixed with methanol for 30 min, stained with 0.1% crystal violet
for 20 min, dried, observed under an inverted microscope and
photographed. Five visual fields were randomly selected to count
the number of cells crossing the membrane to take the average.
Colony formation assay
Agarose (1.2%) was dissolved, and water-bathed in
46°C. Counted single cells were suspended in 40°C pre-warmed
RPMI-1640 medium containing 40% fetal calf serum. Afterwards, 325
µl cell suspension (1×105 cells) was added into
each well of the 24-well plate. Following the addition of 50
µl sample to be tested, 125 µl pre-warmed 1.2%
agarose was supplemented. After natural coagulation, the sample was
cultured in a 5% CO2 incubator at 37°C for 8-10 day and
observed under an inverted microscope.
5-Ethynyl-2′-deoxyuridine (EdU) staining
for cell proliferation
The cells in each group were cultured in 50
µM EdU (EdU labeling/detection kit; Guangzhou RiboBio Co.,
Ltd.) for 12 h, fixed in 4% paraformaldehyde for 30 min at room
temperature and further incubated in 5% glycine for 5 min at room
temperature. The cells were washed in PBS followed by treatment
with 0.5% Triton X-100 at room temperature for 30 min. The samples
in each well were incubated with 100 µl Apollo®
mixture (cat. no. C10338-2; Guangzhou RiboBio Co., Ltd.) for 30 min
at room temperature. The cells were observed and imaged under a
fluorescence microscope.
Flow cytometry for cell cycle and cell
apoptosis
At 48 h after transfection, the cells were collected
for cell cycle analysis. The cells were resuspended in PBS
(~1×106 cells), fixed in 75% ethanol at 4°C for 1 h and
incubated with 150 µl RNase (cat. no. GE101-01; TransGen
Biotech Co., Ltd.) for 40 min at 37°C. Afterwards, the cells were
stained with 200 µl propidium iodide (PI; Sigma-Aldrich;
Merck KGaA) for 2 h at 4°C. Finally, the cell cycle distribution
was measured through the detection of red fluorescence at an
excitation wavelength of 488 nm using a flow cytometer (CytoFLEX;
Beckman Coulter) and analyzed using ModfitLT for Mac 3.0 software
(Verity Software House, Inc.).
After transfection for 48 h, the cells were treated
with EDTA-free trypsin (cat. no. YB15050057; Shanghai Yu Bo
Biological Technology Co., Ltd.) and collected into a flow tube.
The supernatant was discarded after centrifugation at room
temperature at 1,200 × g for 5 min, which was followed by three
washes in pre-cooled PBS. Centrifugation was then performed again
after removal of the supernatant. Based on the instructions of the
Annexin-V-FITC cell apoptosis detection kit (cat. no. APOAF-20TST;
Sigma-Aldrich; Merck KGaA), Annexin-V-FITC/PI staining solution was
prepared using Annexin-V-FITC, PI and HEPES buffer at a ratio of
1:2:50. Then, 100 µl staining solution was used to resuspend
1×106 cells, followed by oscillation and mixing. The
specimens were then incubated at room temperature for 15 min, after
which 1 ml HEPES buffer solution (cat. no. PB180325; Procell Life
Science Co., Ltd.) was added and the suspension was oscillated and
mixed accordingly. Cell apoptosis was analyzed through the
detection of FITC and PI fluorescence by activating the band pass
at 525 and 620 nm at a wavelength of 488 nm using a flow
cytometer.
Xenograft tumors in nude mice
Male nude mice (n=20; 18-22 g; 4-6 weeks) were
housed in a temperature- (26-28°C) and humidity-controlled (40-60%)
environment with a 10/14 h light/dark cycle, and were provided free
access to aseptic water and chow. The cage was ventilated 10-15
times per h under positive air pressure of 0.65 cm H2O.
The animals were assigned to four groups, as follows: si-NC-CX26 +
si-NC-CX43; si-CX26 + si-NC-CX43; si-NC-CX26 + si-CX43; and si-CX26
+ si-CX43. After the preparation of lentivirus (2×107
TU; Shanghai GenePharma Pharmaceutical Technology Co., Ltd.)
carrying siRNAs, the lentivirus-packed si-NC-CX26 + si-NC-CX43,
si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43, and si-CX26 + si-CX43
were transduced into isolated human NSCLC PECs. The
2×106 cells (dissolved in 200 µl normal saline)
were subcutaneously injected into the right axilla of each nude
mouse. From the 7th day after injection, observations were
conducted once every 7 days. The length and width of the tumors
that appeared in the mice were recorded in detail. The approximate
tumor volume was calculated according to the formula, volume
(mm3)=length x width2/2. The mice were
subsequently euthanized on the 35th day to collect their
tumors.
HE staining
The lung tissues were obtained under a dissecting
microscope (Olympus Corporation) (21,22).
The tissues were fixed in 3% neutral formalin at room temperature
for 5 min and used to generate paraffin slices at a thickness of
5-8 µm. The prepared sections were deparaffinized twice in
xylene (5 min each time) and dehydrated in gradient ethanol
solutions (100, 95, 80 and 75%) for 1 min each. After they were
washed in running water for 2 min, the sections were stained with
hematoxylin for 2 min and washed under running water for 10 sec.
Next, the excess staining solution was removed with 1% hydrochloric
acid-75% ethanol. The sections were then washed with distilled
water for 1 min, stained with eosin for 1 min, washed again with
distilled water for 10 sec, dehydrated twice in gradient ethanol
solutions (95 and 100%; 1 min each) and cleared with xylene. All
above mentioned procedures were conducted at room temperature.
Finally, the sections were sealed with neutral gum for further
observation.
Statistical analysis
All experimental data were processed using SPSS 19.0
statistical software (IBM Corp.). Continuous data with normal
distribution and equal variance were expressed as the mean ±
standard deviation. Paired t-test was used to compare data between
cancer tissues and paracancerous tissues and an independent sample
t-test was used to analyze data between two groups. One-way ANOVA
or two-way ANOVA (tumor volume) with Tukey's post hoc test was used
for comparisons among multiple groups. Enumeration data were
presented as percentages and analyzed using Fisher's exact test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
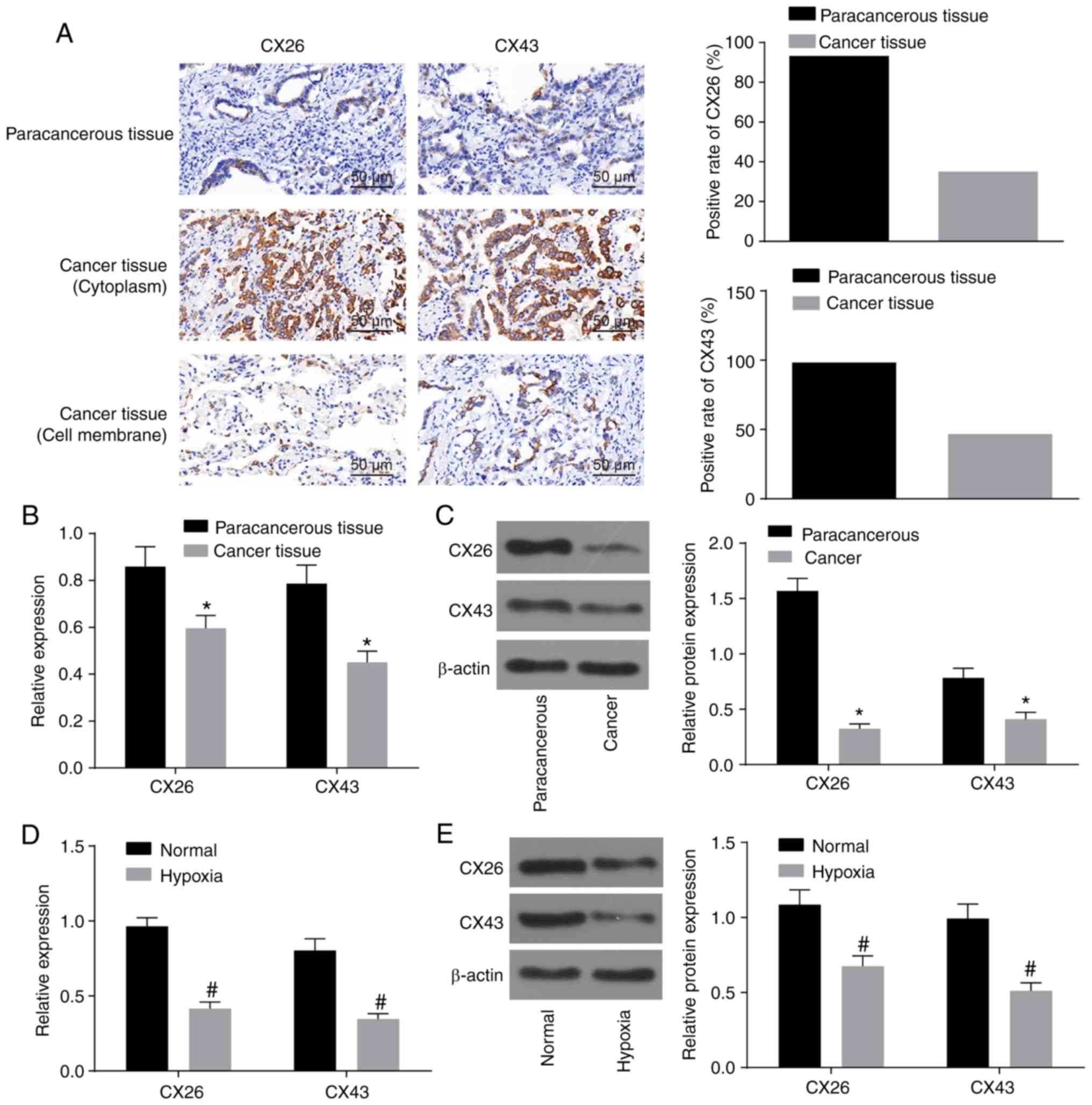
Hypoxia downregulates CX26 and CX43
expression in NSCLC PECs
Initially, to observe the effect of hypoxia on the
expression of CX26 and CX43 in PECs derived from NSCLC, the
expression of CX26 and CX43 was assessed by immunohistochemistry
and RT-qPCR. As illustrated in Table
II and Fig. 1A, the percentage
of CX26-positive samples was 93.33% (56/60) in paracancerous
tissues and 35.00% (21/60) in cancer tissues (P<0.05).
Additionally, there was a significant difference in the percentage
of CX43-positive samples, which was 98.34% (59/60) in paracancerous
tissues and 46.67% (28/60) in cancer tissues. The non-specific
response of CX26 in the tissue background was weak, while the
localization was clear with positive CX26 in a hairline or particle
shape. Among paracancerous tissues, CX26-positive particles were
detected on the clear-expressed membrane in a hairline shape with
unstained cytoplasm, while in cancer tissues, positively stained
particles were mainly distributed in the cytoplasm, and only two
cases with simultaneous partly stained membranes were observed.
Similar results were observed for CX43, but three cases with partly
stained membranes were observed (Fig.
1A). RT-qPCR and western blot analysis revealed that CX26 and
CX43 expression was decreased in cancer tissues compared with
paracancerous tissues (P<0.05; Fig.
1B and C). Following isolation and culture, human NSCLC PECs
were exposed to hypoxia, and the results revealed that CX26 and
CX43 expression was significantly reduced in the hypoxia group
compared with the normal group (P<0.05; Fig. 1D and E). The aforementioned data
demonstrated that the expression of CX26 and CX43 was downregulated
by hypoxia in PECs.
| Table IIAssociation analysis between
CX26/CX43 and carcinogenesis in pulmonary epithelial tissue. |
Table II
Association analysis between
CX26/CX43 and carcinogenesis in pulmonary epithelial tissue.
| Group | CX26
| CX43
|
|---|
| n | Positive rate
(%) | P-value | n | Positive rate
(%) | P-value |
|---|
| Paracancerous | 60 | 56/60 (93.33) | <0.001 | 60 | 59/60 (98.34) | <0.001 |
| Cancer | 60 | 21/60 (35.00) | | 60 | 28/60 (46.67) | |
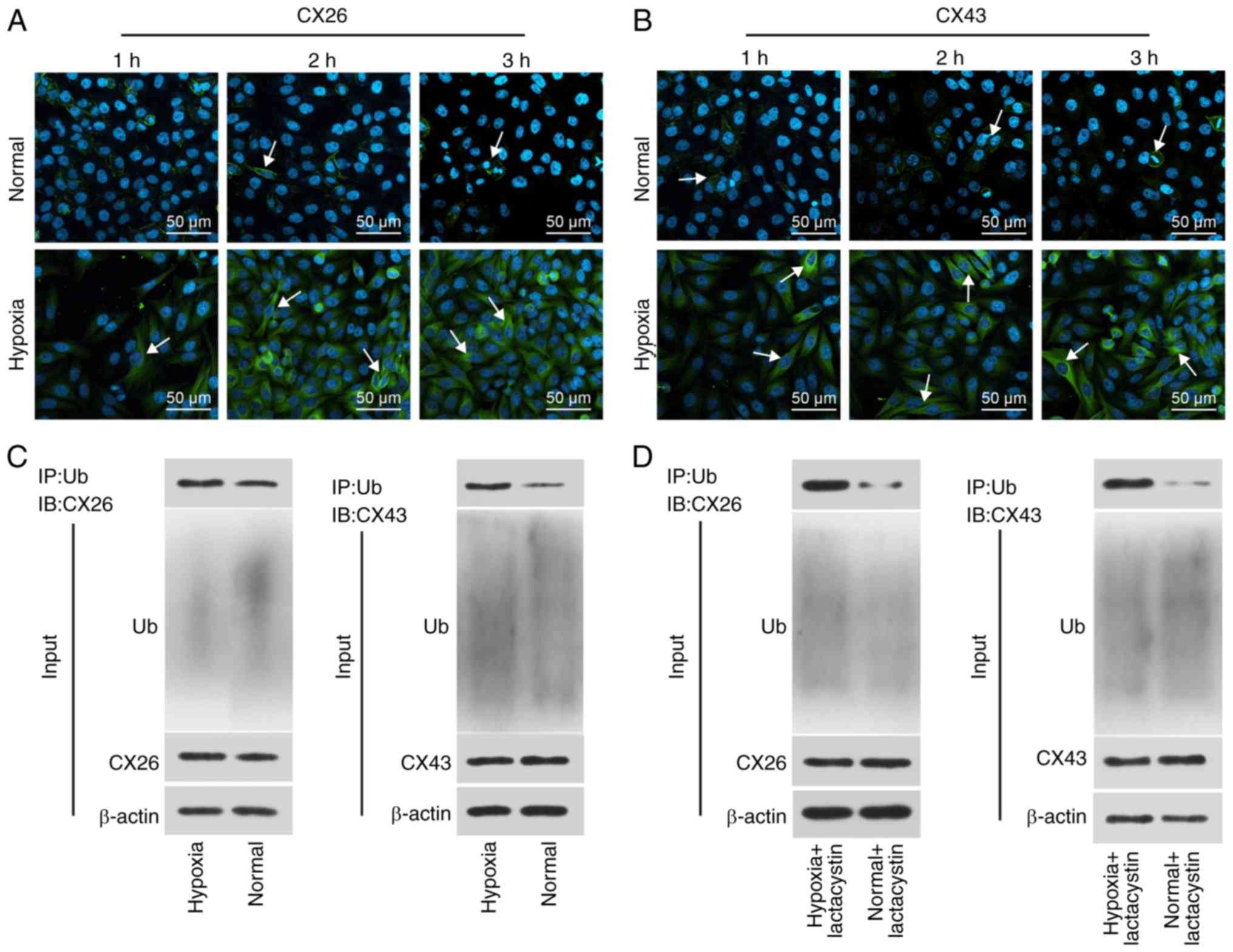
Hypoxia induces internalization and
degradation of CX26 and CX43
To determine the effect of hypoxia on the
internalization and degradation of CX26 and CX43, human NSCLC PECs
were isolated, cultured and exposed to hypoxic conditions for
extended periods followed by immunofluorescence staining. The
results revealed that CX26 and CX43 were both expressed on the
membrane in the normal group, while the expression and localization
of CX26 were not significantly different over time under normal
conditions. Compared with the normal group, the expression of CX26
and CX43 was increased in the cytoplasm, but reduced in the
membranes, in the hypoxia group, with progression over time
(Fig. 2A and B). These findings
suggested that expression of CX26 and CX43 in human NSCLC PECs was
gradually translocated from the membrane to the cytoplasm following
extended periods of hypoxia.
Subsequently, the binding of CX26 and CX43 to
ubiquitin was analyzed through a Co-IP, the results of which
demonstrated positive signals of CX26 and CX43 in the hypoxia
group; however, a weaker signal was detected in the normal group
(Fig. 2C). Following long-term
hypoxia exposure, PECs were further treated with the proteasome
inhibitor lactacystin, after which the binding of CX26 and CX43 to
ubiquitin protein was again detected by Co-IP. As illustrated in
Fig. 2D, the positive signal of
CX26 and CX43 was detected in the hypoxia + lactacystin group, but
weaker levels were observed in the normal + lactacystin group. The
aforementioned results highlighted that hypoxia promotes the
internalization of CX26 and CX43, and ubiquitination and
degradation are induced.
Downregulation of CX26 and CX43 induces
EMT in NSCLC PECs
To investigate the effects of CX26 and CX43 on EMT
in PECs, isolated human NSCLC PECs were exposed to hypoxia. RT-qPCR
and western blot analysis were performed to determine the
expression of E-cadherin and N-cadherin, while a Transwell assay
was performed to assess cell migration ability. The results
indicated lower E-cadherin expression and higher N-cadherin
expression in the hypoxia group (Fig.
3A-C), while cell migration ability was enhanced (Fig. 3D) compared with the normal group
(P<0.05). Next, human NSCLC PECs were transfected with
si-NC-CX26 + si-NC-CX43, si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43
and si-CX26 + si-CX43. To confirm the interference efficiency of
si-CX26 and si-CX43, the expression levels of CX26 and CX43 were
determined by RT-qPCR and western blot analysis. It was found that
the expression of CX26 in the si-CX26 + si-NC-CX43 and si-CX26 +
si-CX43 groups was decreased relative to the si-NC-CX26 +
si-NC-CX43 group (P<0.05), with no significant difference
observed in CX26 expression between the si-NC-CX26 + si-CX43 and
si-NC-CX26 + si-NC-CX43 groups (P>0.05). No significant
difference was found in CX43 expression between the si-CX26 +
si-NC-CX43 and si-NC-CX26 + si-NC-CX43 groups (P>0.05), but the
expression of CX43 was significantly diminished in the si-CX26 +
si-CX43 group compared with the si-NC-CX26 + si-NC-CX43 group
(P<0.05; Fig. 3E-G). RT-qPCR,
western blot analysis and Transwell assay were performed again
under hypoxic conditions with E-cadherin and N-cadherin analyzed.
The results revealed that, compared with the si-NC-CX26 +
si-NC-CX43 group, the si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43
and si-CX26 + si-CX43 groups all exhibited decreased E-cadherin
expression and increased N-cadherin expression (Fig. 3H-J) along with increased cell
migration (Fig. 3K; P<0.05).
Next, NC-CX26 + NC-CX43, CX26 + NC-CX43, NC-CX26 + CX43 and CX26 +
CX43 were used to transfect human NSCLC PECs and overexpress CX26
and CX43, which were then exposed to hypoxic conditions. RT-qPCR,
western blot analysis and a Transwell assay were performed for
investigative purposes. Compared with the NC-CX26 + NC-CX43 +
hypoxia group, elevated E-cadherin expression, reduced N-cadherin
expression (Fig. 3L-N) as well as
suppressed cell migration ability (Fig. 3O) were exhibited in the CX26 +
NC-CX43 + hypoxia, NC-CX26 + CX43 + hypoxia and CX26 + CX43 +
hypoxia groups (P<0.05). However, no significant difference was
observed between the CX26 + NC-CX43 + hypoxia and NC-CX26 + CX43 +
hypoxia groups. Based on these results, it was concluded that
hypoxia induced EMT in PECs, potentially through the downregulation
of CX26 and CX43.
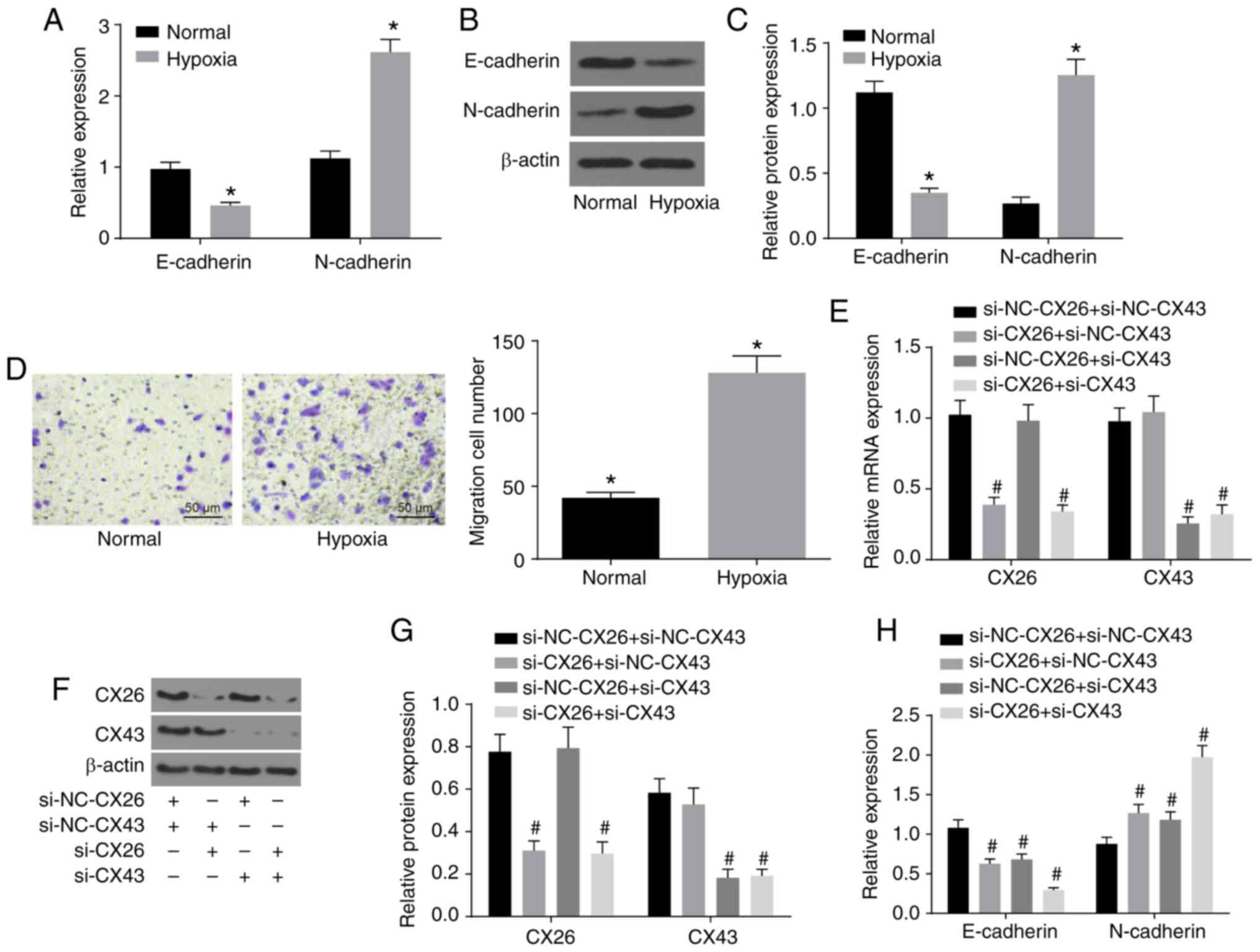 | Figure 3Downregulation of CX26 and CX43
induces epithelial-mesenchymal transition in PECs. (A) mRNA
expression of E-cadherin and N-cadherin in human NSCLC PECs
following exposure to hypoxia, as evaluated by RT-qPCR. (B) Protein
expression of E-cadherin and N-cadherin in human NSCLC PECs
following exposure to hypoxia, as assessed by western blot
analysis. (C) Semi-quantification of results from panel B. (D)
Migration ability of human NSCLC PECs following exposure to
hypoxia, as seen by microscopy (×200). Human NSCLC PECs were
transfected with si-NC-CX26 + si-NC-CX43, si-CX26 + si-NC-CX43,
si-NC-CX26 + si-CX43 or si-CX26 + si-CX43 and (E) mRNA expression
of CX26 and CX43 was evaluated by RT-qPCR, (F) protein expression
of CX26 and CX43 was assessed by western blot analysis and (G)
semi-quantified, (H) mRNA expression of E-cadherin and N-cadherin
was evaluated by RT-qPCR, (I) protein expression of E-cadherin and
N-cadherin was assessed by western blot analysis and (J)
semi-quantified, (K) migration ability was determined (×200).
Downregulation of CX26 and CX43 induces epithelial-mesenchymal
transition in PECs. Human NSCLC PECs were transfected with NC-CX26
+ NC-CX43, CX26 + NC-CX43, NC-CX26 + CX43 or CX26 + CX43
overexpression vectors, and exposed to hypoxia, then (L) mRNA
expression of E-cadherin and N-cadherin was evaluated by RT-qPCR,
(M) protein expression of E-cadherin and N-cadherin was assessed by
western blot analysis and (N) semi-quantified, and (O) migration
ability was determined (×200). *P<0.05 vs. normal
group; #P<0.05 vs. si-NC + si-NC group;
&P<0.05 vs. the NC-CX26 + NC-CX43 + hypoxia group
(n=3). NSCLC, non-small cell lung cancer; PECs, pulmonary
epithelial cells; RT-qPCR, reverse transcription-quantitative PCR;
si, small interfering RNA; NC, negative control; CX, connexin. |
Downregulation of CX26 and CX43 promotes
PEC proliferation and growth in NSCLC
Subsequently, to determine the effects of CX26 and
CX43 on the proliferation and growth of PECs, human NSCLC PECs were
transfected with si-CX26 and/or si-CX43, and an EdU assay was
applied to determine cell proliferation. Compared with the
si-NC-CX26 + si-NC-CX43 group, cell proliferation ability was
promoted in the si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43 and
si-CX26 + si-CX43 groups (Fig.
4A). Western blot analysis was performed to determine the
relative protein expression of the characteristic stem cell markers
Oct4 and Nanog, both of which were elevated in the si-CX26 +
si-NC-CX43, si-NC-CX26 + si-CX43 and si-CX26 + si-CX43 groups
compared with the si-NC-CX26 + si-NC-CX43 group (P<0.05;
Fig. 4B). Flow cytometry revealed
that the cell apoptosis was decreased in the si-CX26 + si-NC-CX43,
si-NC-CX26 + si-CX43 and si-CX26 + si-CX43 groups relative to the
si-NC-CX26 + si-NC-CX43 group (Fig.
4C) and that cells had been arrested in S phase (Fig. 4D). As illustrated in Fig. 4E, the colony formation assay
revealed that the number of colonies formed was higher in the
si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43 and si-CX26 + si-CX43
groups compared with the si-NC-CX26 + si-NC-CX43 group. These
findings suggested that CX26 and CX43 were involved in the PEC cell
cycle in NSCLC.
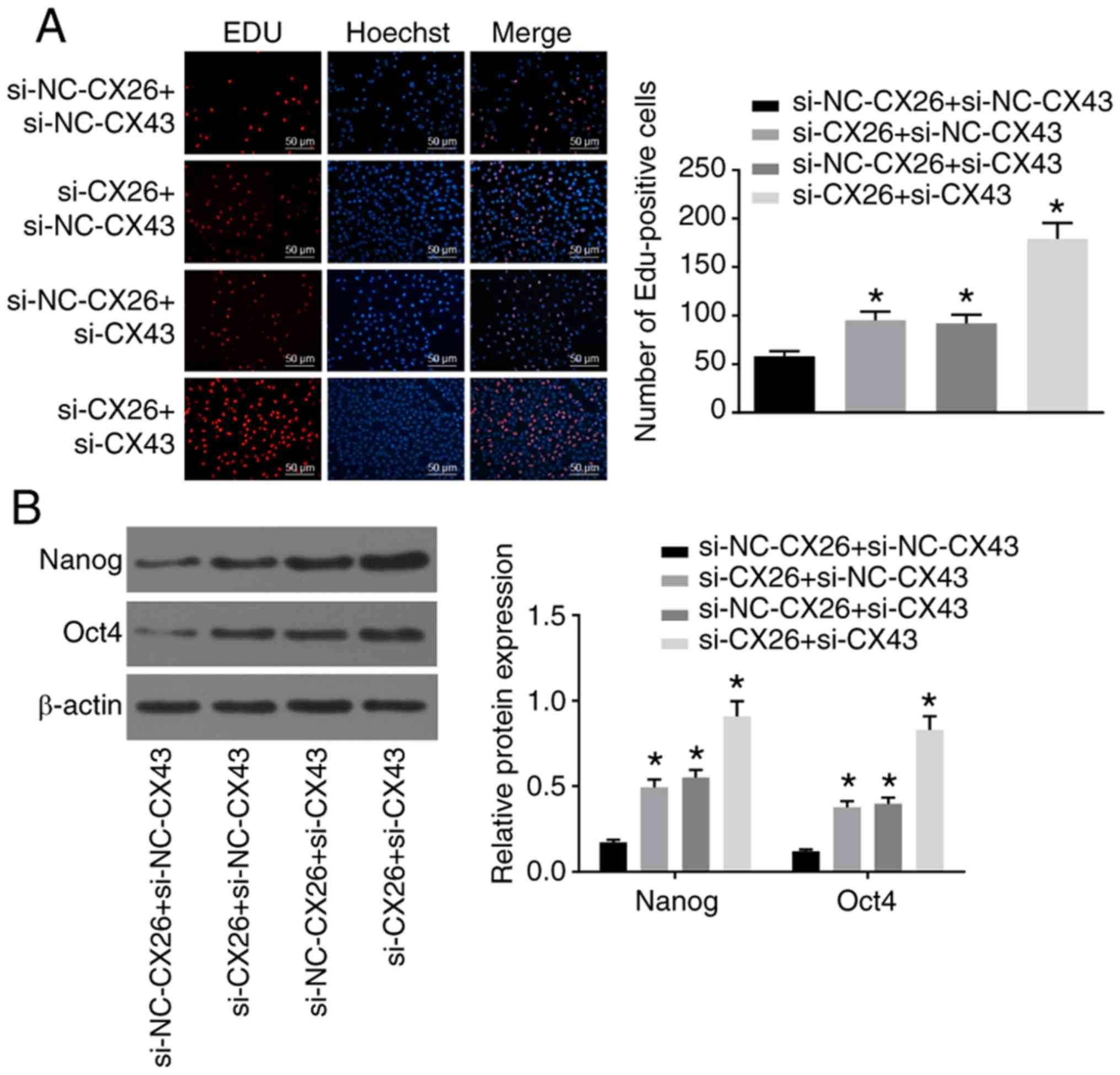 | Figure 4Downregulation of CX26 and CX43
promotes PEC proliferation and growth. PECs were transfected with
si-NC-CX26 + si-NC-CX43, si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43
or si-CX26 + si-CX43. (A) Cell proliferation was assessed by EdU
assay (scale bar, 50 µm). (B) Protein expression of Nanog
and Oct4 following transfection, as determined by western blot
analysis. (C) Cell apoptosis following transfection, as detected by
Annexin V-FITC/PI double staining. (D) Cell cycle analysis
following transfection, as detected by PI staining. (E) Colony
formation in vitro following transfection. PECs were
transfected with NC-CX26 + NC-CX43, CX26 + NC-CX43, NC-CX26 + CX43
or CX26 + CX43 and exposed to hypoxia. Downregulation of CX26 and
CX43 promotes PEC proliferation and growth. (F) Cell proliferation
following transfection was assessed by EdU assay (×200). (G)
Protein expression of Nanog and Oct4 following transfection and
hypoxia exposure, as determined by western blot analysis. (H) Cell
apoptosis following transfection and hypoxia exposure, as detected
by Annexin V-FITC/PI double staining. (I) Cell cycle analysis
following transfection and hypoxia exposure, as detected by PI
single staining. *P<0.05 vs. si-NC-CX26 + si-NC-CX43
group; #P<0.05 vs. NC-CX26 + NC-CX43 + hypoxia group
(n=3). PEC, pulmonary epithelial cell; EdU,
5-ethynyl-2′-deoxyuridine; si, small interfering RNA; NC, negative
control; CX, connexin; NC, negative control; PI, propidium
iodide. |
Following transfection with CX26 and/or CX43
overexpression vectors, human NSCLC PECs were exposed to hypoxia
after which an EdU assay, western blot analysis and flow cytometry
were performed to explore the effects on cell function. The results
showed that the CX26 + NC-CX43 + hypoxia, NC-CX26 + CX43 + hypoxia
and CX26 + CX43 + hypoxia groups had suppressed cell proliferation
ability, lower expression of Nanog and Oct4, enhanced cell
apoptosis ability and fewer cells in phase S compared with the
NC-CX26 + NC-CX43 + hypoxia group (P<0.05; Fig. 4F-I). However, there were no
significant differences detected in relation to the aforementioned
changes between the CX26 + NC-CX43 + hypoxia and NC-CX26 + CX43 +
hypoxia groups. Taken together, it was concluded that
downregulation of CX26 and CX43 promotes PEC proliferation and
growth, while inhibiting cell apoptosis in NSCLC cells.
CX26 and CX43 mediate activation of the
P53/MDM2 signaling pathway
The association between CX26 and CX43 and their
effects on the P53/MDM2 signaling pathway were also investigated
based on the quantification of related genes (P53, MDM2 and P21) in
human NSCLC PECs using RT-qPCR and western blot analysis. As shown
in Fig. 5A-C, the expression of
P53 and P21 decreased, while MDM2 expression increased in the
hypoxia group cells compared with the normal group (P<0.05),
which demonstrated the activation of the P53/MDM2 signaling pathway
under hypoxia.
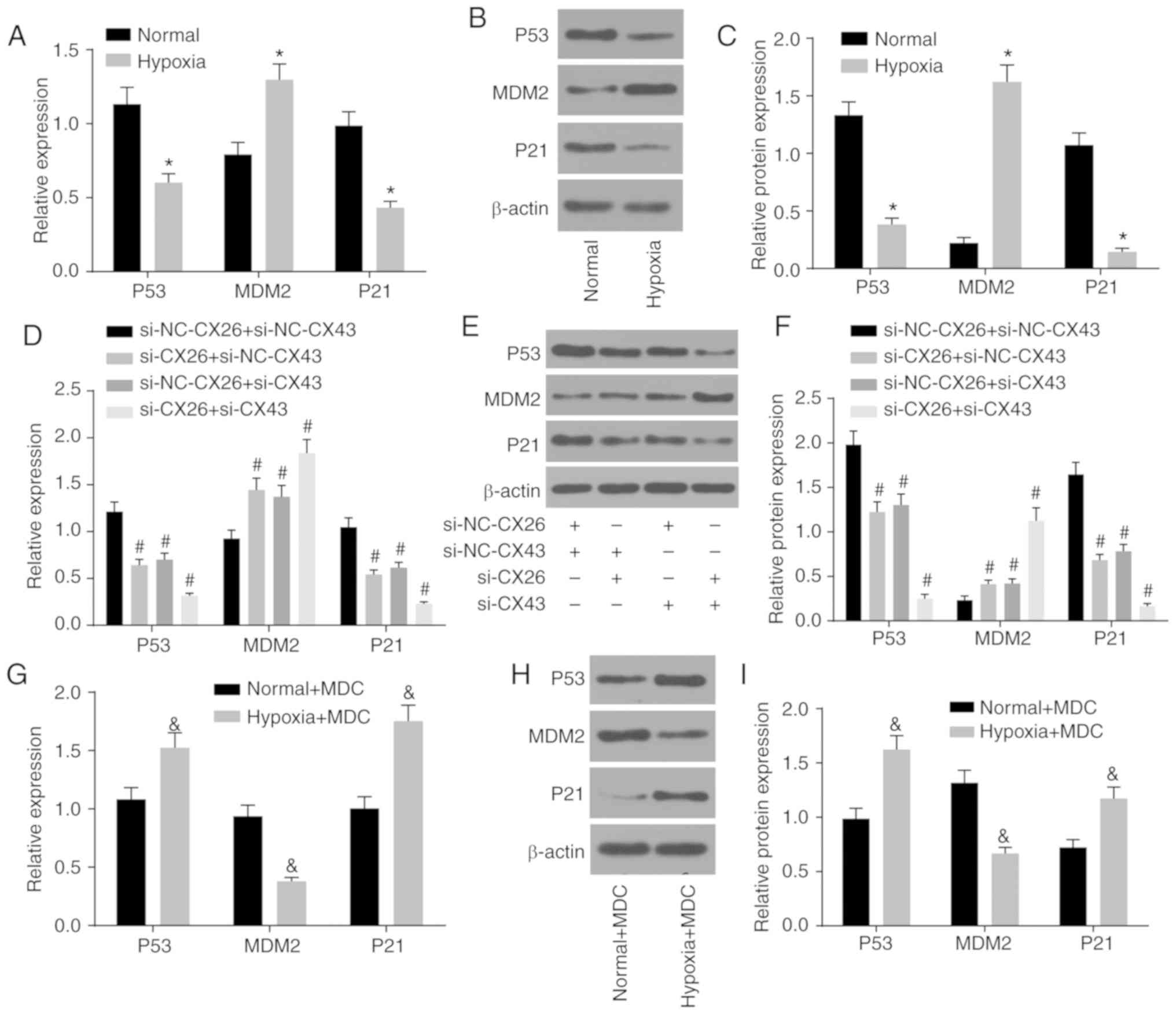 | Figure 5CX26 and CX43 mediate activation of
the P53/MDM2 signaling pathway. (A) mRNA and (B) protein expression
of P53, MDM2 and P21 in human NSCLC PECs following exposure to
hypoxia, as determined by RT-qPCR and western blot analysis,
respectively. (C) Semi-quantification of panel B. (D) mRNA and (E)
protein expression of P53, MDM2 and P21 in human NSCLC PECs
following transfection with si-NC-CX26 + si-NC-CX43, si-CX26 +
si-NC-CX43, si-NC-CX26 + si-CX43 or si-CX26 + si-CX43, as tested by
RT-qPCR and western blot analysis, respectively. (F)
Semi-quantification of panel E. (G) mRNA expression and (H) protein
expression of P53, MDM2 and P21 in human NSCLC PECs following MDC
treatment and hypoxia exposure, as evaluated by RT-qPCR and western
blot analysis, respectively. (I) Semi-quantification of panel H.
*P<0.05 vs. normal group; #P<0.05 vs.
si-NC-CX26 + si-NC-CX43 group; &P<0.05 vs. the
normal + MDC group (n=3). NSCLC, non-small cell cancer; PECs,
pulmonary epithelial cells; RT-qPCR, reverse
transcription-quantitative PCR; MDM2, murine double minute-2; si,
small interfering RNA; NC, negative control; CX, connexin; MDC,
monodansylcadaverine. |
Additionally, si-CX26 and si-CX43 were used to
transfect human NSCLC PECs for further exploration. Compared with
the si-NC-CX26 + si-NC-CX43 group, the si-CX26 + si-NC-CX43,
si-NC-CX26 + si-CX43 and si-CX26 + si-CX43 groups displayed lower
expression of P53 and P21, along with higher MDM2 expression
(P<0.05; Fig. 5D-F), which
suggested that the P53/MDM2 signaling pathway was activated through
downregulation of CX26 and CX43.
Moreover, protein internalization inhibitors were
used to treat human NSCLC PECs that had previously been exposed to
hypoxic conditions to assess activation of the P53/MDM2 signaling
pathway. The results showed that compared with the normal + MDC
group, expression of P53 and P21 was enhanced in the hypoxia + MDC
group, whereas MDM2 expression was diminished (P<0.05; Fig. 5G-I). These findings collectively
led to the conclusion that hypoxia could activate the P53/MDM2
signaling pathway through induction of CX26 and CX43
internalization.
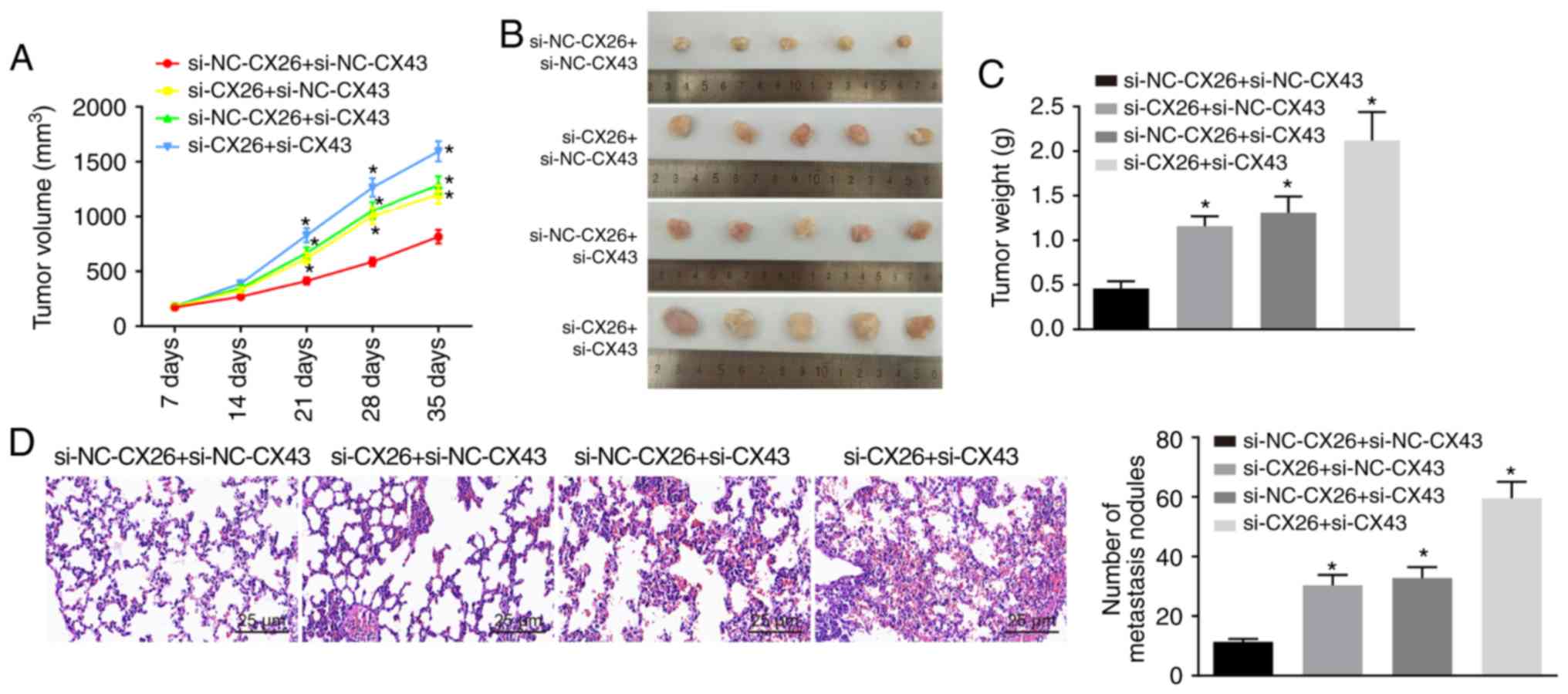
CX26 and CX43 promote tumorigenesis in
vivo
To test the effect of CX26 and CX43 on tumorigenesis
in vivo, nude mice were injected with PECs with CX26 and
CX43 knockdown. As shown, from the 21st day, tumor growth in the
si-CX26 + si-NC-CX43, si-NC-CX26 + si-CX43 and si-CX26 + si-CX43
groups exhibited accelerated growth (P<0.05; Fig. 6A and B) and the nude mice presented
with larger tumor weights than those in the si-NC-CX26 + si-NC-CX43
group (P<0.05; Fig. 6C). HE
staining was performed to observe tumor metastasis in the lung;
compared with normal lung tissues, metastatic tumor cells were
closely arranged, with deeply stained nuclear, clear nuclear
membrane and obvious nucleolus. Moreover, there were a higher
number of tumor metastatic nodes in the si-CX26 + si-NC-CX43,
si-NC-CX26 + si-CX43 and si-CX26 + si-CX43 groups compared with the
si-NC-CX26 + si-NC-CX43 group (P<0.05; Fig. 6D). The data indicated that
downregulation of CX26 and CX43 could promote tumorigenesis in
vivo.
Discussion
Lung cancer is widely considered to be among the
most prevalent malignancies worldwide, as studies have presented
cases of lung cancer in China that account for >1/3 of all
global cases, which places a severe burden on both individuals and
society as a whole (23). NSCLC is
one of the predominant histological types of lung cancer and
includes adenocarcinoma, squamous cell carcinoma and large cell
carcinoma, all of which usually arise from the alveoli, small
bronchi or bronchioles (24). The
alveoli, the structures where gas transfer occurs, play a central
role in lung function. Hypoxia causes lung dysfunction as a result
of impaired tissue function (25).
Mounting evidence has continued to highlight the significance of CX
isotypes in human cells partially due to their ubiquity and large
density in solid tissues (26). In
the current study, we investigated the roles of CX26 and CX43 under
hypoxic conditions in the progression of NSCLC through a series of
well-designed experiments and subsequently concluded that hypoxia
could induce the internalization of CX26 and CX43 via the P53/MDM2
signaling pathway, which ultimately further promotes the incidence
of NSCLC.
Initially, a key finding of our study indicated
aberrant expression of CX26 and CX43 in PECs induced by hypoxia,
which suggests that CX26 and CX43 were both downregulated in cancer
tissues, as well as cancer cells, after hypoxia exposure. Likewise,
researchers have reported that CX43 expression is reduced in NSCLC,
while demonstrating that the downregulation of CX26/CX43 is also
found in the alveolar epithelium in human lung cancer (27). Such changes have been widely
discussed in previous studies, which have indicated that CX
overexpression is observed at the early stage of cancer, while
reductions in function and expression have been noted in malignant
tumors, and at times, expression is even absent at more advanced
stages (26). Similar results have
been presented in another study in which subsequent changes,
including lung injury, induced the downregulation of CX43 and
reduced gap junctional intercellular communication (28). Furthermore, the role of CX-involved
channels has been shown to contribute to lung repair and
inflammatory response (27). It
has been confirmed by previous studies that hypoxia reduces the
expression of CX26 and CX43. Specifically, chronic prenatal hypoxia
causes promoter region hypermethylation and decreases the mRNA and
protein expression of CX26 (29).
Additionally, hypoxia is observed to decrease the CX43 protein
level by 30-50% (30), which was
consistent with the results of the present study.
Furthermore, additional evidence has been presented
in the current study that demonstrates that hypoxic conditions
promote the binding of CX26 and CX43 to ubiquitin protein in favor
of CX26 and CX43 internalization and degradation. In addition, the
expression CX26/CX43 was predominantly located within the cytoplasm
as opposed to the membrane. A prior study revealed that following
extended exposure to hypoxia, CX43 may translocate to the cytoplasm
from the membrane, which is accompanied by a decrease in
communication properties (31);
this was largely consistent with the results of the current study.
Ubiquitination is a crucial posttranslational modification
occurring during nonlysosomal protein degradation that is capable
of influencing the progression and development of cancer;
ubiquitination also shares a relationship with both tumor
suppressive and stimulatory pathways (32). Additionally, other studies have
previously indicated that degradation of CX co-exists with a
variety of physiological and pathological variations, including
cell migration (33), which was
further verified by the results of the present study.
In a subsequent experiment, the effects involved the
regulation of the biological functions of NSCLC PECs EMT and growth
involving CX26 and CX43 were determined to be enhanced when CX26
and CX43 were downregulated, which was further evidenced by the
increased expression of N-cadherin, Nanog and Oct4, decreased
E-cadherin expression and activation of the P53/MDM2 signaling
pathway. A conclusion drawn from the experiment performed by Zhao
et al (34) suggested that,
among the Chinese population, expression of CX43 and E-cadherin
decreases during NSCLC development, and thus the differentiation
and lymph node metastasis of cancer cells may be predicted. Oct4
and its downstream target Nanog have been investigated in depth as
essential homeobox transcription factors that participate in stem
cell self-renewal fate determination, while the ectopic
overexpression of both has been observed to operate in tandem to
promote EMT and tumor-initiating ability of lung adenocarcinoma,
the most prevalent histologic type of lung cancer (35). Evidence has been previously
presented that indicates that, if activated under hypoxic
conditions, P53 is able to function as an extraordinary tumor
suppressor regarding progression and prognosis through its negative
interaction with MDM2 (36). The
function of P53 is largely linked to cell proliferation and
apoptosis, along with cell cycle distribution, as P53 can promote
cell apoptosis (37). The
follow-up in vivo experiments further verified that
downregulated CX26 and CX43 promoted tumorigenicity. The
aforementioned literature and the observations of this study
support the promotive role of CX26 and CX43 in NSCLC.
Taken together, the data obtained in the current
study confirmed that hypoxia reduced the expression of CX26 and
CX43, and abnormal activation of the P53/MDM2 signaling pathway
through a combination of effects. These effects regulated
proliferation, migration and apoptosis of PECs, which ultimately
promoted NSCLC progression. Thus, in accordance with the evidence
presented, it is proposed that, under hypoxic conditions, CX26 and
CX43 may have therapeutic implications. Nevertheless, this is a
relatively new field of research in pulmonary diseases and provides
a platform for a better understanding of the mechanisms of NSCLC
progression. Therefore, more studies and additional research areas
are required to establish enhanced treatment methods for patients
with NSCLC. In the follow-up experiments, in addition to RT-qPCR
and western blot analysis performed to detect the efficiency of
RNAi, gene overexpression experiments for gene interference will be
conducted to ensure the specificity of RNAi.
Supplementary Data
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ and SZ performed and analyzed the studies. SZ and
XL initiated the work and analyzed the data. XL and JCL obtained
the results and validated them, and provided the figures and tables
and wrote the manuscript. All authors contributed to the revised
manuscript and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The First Affiliated Hospital of Nanchang University. Written
informed consent was obtained from each participant. All animal
experimentation in this study was approved by the Institutional
Animal Care and Use Committee of The First Affiliated Hospital of
Nanchang University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wakelee H, Kelly K and Edelman MJ: 50
Years of progress in the systemic therapy of non-small cell lung
cancer. Am Soc Clin Oncol Educ Book. pp. 177–189. 2014, View Article : Google Scholar
|
|
3
|
Lei Z, Xu G, Wang L, Yang H, Liu X, Zhao J
and Zhang HT: MiR-142-3p represses TGF-β-induced growth inhibition
through repression of TGFbetaR1 in non-small cell lung cancer.
FASEB J. 28:2696–2704. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grutsch JF, Wood PA, Du-Quiton J, Reynolds
JL, Lis CG, Levin RD, Ann Daehler M, Gupta D, Quiton DF and
Hrushesky WJ: Validation of actigraphy to assess circadian
organization and sleep quality in patients with advanced lung
cancer. J Circadian Rhythms. 9:42011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kleisiaris CF, Kritsotakis EI, Daniil Z,
Tzanakis N, Papaioannou A and Gourgoulianis KI: The prevalence of
obstructive sleep apnea-hypopnea syndrome-related symptoms and
their relation to airflow limitation in an elderly population
receiving home care. Int J Chron Obstruct Pulmon Dis. 9:1111–1117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bouloukaki I, Papadimitriou V, Sofras F,
Mermigkis C, Moniaki V, Siafakas NM and Schiza SE: Abnormal
cytokine profile in patients with obstructive sleep apnea-hypopnea
syndrome and erectile dysfunction. Mediators Inflamm.
2014:5689512014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang J, Qin G, Luo M, Chen J, Zhang Q, Li
L, Pan L and Qin S: Reciprocal positive regulation between Cx26 and
PI3K/Akt pathway confers acquired gefitinib resistance in NSCLC
cells via GJIC-independent induction of EMT. Cell Death Dis.
6:e18292015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Crespin S, Bacchetta M, Bou Saab J,
Tantilipikorn P, Bellec J, Dudez T, Nguyen TH, Kwak BR, Lacroix JS,
Huang S, et al: Cx26 regulates proliferation of repairing basal
airway epithelial cells. Int J Biochem Cell Biol. 52:152–160. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Garcia IE, Maripillán J, Jara O, Ceriani
R, Palacios-Munoz A, Ramachandran J, Olivero P, Perez-Acle T,
González C, Sáez JC, et al: Keratitis-ichthyosis-deafness
syndrome-associated Cx26 mutants produce nonfunctional gap
junctions but hyperactive hemichannels when co-expressed with wild
type Cx43. J Invest Dermatol. 135:1338–1347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sulkowska U, Febp AW and Sulkowski S:
Association of STAT3 with Cx26 and Cx43 in human uterine
endometrioid adenocarcinoma. Oncol Lett. 11:4134–4138. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deben C, Wouters A, Op de Beeck K, van Den
Bossche J, Jacobs J, Zwaenepoel K, Peeters M, Van Meerbeeck J,
Lardon F, Rolfo C, et al: The MDM2-inhibitor Nutlin-3 synergizes
with cisplatin to induce p53 dependent tumor cell apoptosis in
non-small cell lung cancer. Oncotarget. 6:22666–22679. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chang H, Li C, Huo K, Wang Q, Lu L, Zhang
Q, Wang Y and Wang W: Luteolin prevents H2O2-induced apoptosis in
H9C2 cells through modulating Akt-P53/Mdm2 signaling pathway.
Biomed Res Int. 2016:51258362016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reddy R, Buckley S, Doerken M, Barsky L,
Weinberg K, Anderson KD, Warburton D and Driscoll B: Isolation of a
putative progenitor subpopulation of alveolar epithelial type 2
cells. Am J Physiol Lung Cell Mol Physiol. 286:L658–L667. 2004.
View Article : Google Scholar
|
|
14
|
Bao B, Groves K, Zhang J, Handy E, Kennedy
P, Cuneo G, Supuran CT, Yared W, Rajopadhye M and Peterson JD: In
vivo imaging and quantification of carbonic anhydrase IX expression
as an endogenous biomarker of tumor hypoxia. PLoS One.
7:e508602012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tang D, Yan T, Zhang J, Jiang X, Zhang D
and Huang Y: Notch1 signaling contributes to hypoxia-induced high
expression of Integrin β1 in keratinocyte migration. Sci Rep.
7:439262017. View Article : Google Scholar
|
|
16
|
Hau S, Reich DM, Scholz M, Naumann W,
Emmrich F, Kamprad M and Boltze J: Evidence for neuroprotective
properties of human umbilical cord blood cells after neuronal
hypoxia in vitro. BMC Neurosci. 9:302008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bouley R, Lin HY, Raychowdhury MK,
Marshansky V, Brown D and Ausiello DA: Downregulation of the
vasopressin type 2 receptor after vasopressin-induced
internalization: Involvement of a lysosomal degradation pathway. Am
J Physiol Cell Physiol. 288:C1390–C1401. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim SJ, Kim MY, Lee EJ, Ahn YS and Baik
JH: Distinct regulation of internalization and mitogen-activated
protein kinase activation by two isoforms of the dopamine D2
receptor. Mol Endocrinol. 18:640–652. 2004. View Article : Google Scholar
|
|
19
|
Atkins D, Reiffen KA, Tegtmeier CL,
Winther H, Bonato MS and Storkel S: Immunohistochemical detection
of EGFR in paraffin-embedded tumor tissues: Variation in staining
intensity due to choice of fixative and storage time of tissue
sections. J Histochem Cytochem. 52:893–901. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Wilmanns C, Fan D, O'Brian CA, Bucana CD
and Fidler IJ: Orthotopic and ectopic organ environments
differentially influence the sensitivity of murine colon carcinoma
cells to doxorubicin and 5-fluorouracil. Int J Cancer. 52:98–104.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schlagbauer-Wadl H, Griffioen M, van Elsas
A, Schrier PI, Pustelnik T, Eichler HG, Wolff K, Pehamberger H and
Jansen B: Influence of increased c-Myc expression on the growth
characteristics of human melanoma. J Invest Dermatol. 112:332–336.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong QY, Wu GM, Qian GS, Hu CP, Zhou JY,
Chen LA, Li WM, Li SY, Wang K, Wang Q, et al: Prevention and
management of lung cancer in China. Cancer. 121(Suppl 17):
S3080–S3088. 2015. View Article : Google Scholar
|
|
24
|
Long F, Su JH, Liang B, Su LL and Jiang
SJ: Identification of gene biomarkers for distinguishing small-cell
lung cancer from non-small-cell lung cancer using a network-based
approach. Biomed Res Int. 2015:6853032015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olmeda B, Umstead TM, Silveyra P, Pascual
A, Lopez-Barneo J, Phelps DS, Floros J and Perez-Gil J: Effect of
hypoxia on lung gene expression and proteomic profile: Insights
into the pulmonary surfactant response. J Proteomics. 101:179–191.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Teleki I, Szasz AM, Maros ME, Gyorffy B,
Kulka J, Meggyeshazi N, Kiszner G, Balla P, Samu A and Krenacs T:
Correlations of differentially expressed gap junction connexins
Cx26, Cx30, Cx32, Cx43 and Cx46 with breast cancer progression and
prognosis. PLoS One. 9:e1125412014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Losa D, Chanson M and Crespin S: Connexins
as therapeutic targets in lung disease. Expert Opin Ther Targets.
15:989–1002. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu T, Li Y, Zhang B, Ma L, Liu W, Li Z
and Jin F: The role of phosphorylated Cx43 on PKC mediated Ser368
in lung injury induced by seawater inhalation. Inflammation.
38:1847–1854. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin J, Huang H, Lv G, Xu X, Lin W, Xu X,
Cheng J and Zheng M: Chronic prenatal hypoxia impairs cochlear
development, a mechanism involving connexin26 expression and
promoter methylation. Int J Mol Med. 41:852–858. 2018.
|
|
30
|
Wu X, Huang W, Luo G and Alain LA: Hypoxia
induces connexin 43 dysregulation by modulating matrix
metalloproteinases via MAPK signaling. Mol Cell Biochem.
384:155–162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Otto T, Gellhaus A, Lüschen N, Scheidler
J, Bendix I, Dunk C, Wolf N, Lennartz K, Koninger A, Schmidt M, et
al: Oxygen sensitivity of placental trophoblast Connexins 43 and
46: A role in preeclampsia? J Cell Biochem. 116:2924–2937. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Morrow JK, Lin HK, Sun SC and Zhang S:
Targeting ubiquitination for cancer therapies. Future Med Chem.
7:2333–2350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Falk MM, Kells RM and Berthoud VM:
Degradation of connexins and gap junctions. FEBS Lett.
588:1221–1229. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao JQ, Sun FJ, Liu SS, Yang J, Wu YQ, Li
GS, Chen QY and Wang JX: Expression of connexin 43 and E-cadherin
protein and mRNA in non-small cell lung cancers in Chinese
patients. Asian Pac J Cancer Prev. 14:639–643. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chiou SH, Wang ML, Chou YT, Chen CJ, Hong
CF, Hsieh WJ, Chang HT, Chen YS, Lin TW, Hsu HS and Wu CW:
Coexpression of Oct4 and Nanog enhances malignancy in lung
adenocarcinoma by inducing cancer stem cell-like properties and
epithelial-mesenchymal transdifferentiation. Cancer Res.
70:10433–10444. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang W and Hu Y: Small molecule agents
targeting the p53-MDM2 pathway for cancer therapy. Med Res Rev.
32:1159–1196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang P, Kratz AS, Salama M, Elabd S,
Heinrich T, Wittbrodt J, Blattner C and Davidson G: Expression
screening using a Medaka cDNA library identifies evolutionarily
conserved regulators of the p53/Mdm2 pathway. BMC Biotechnol.
15:922015. View Article : Google Scholar : PubMed/NCBI
|