Introduction
Uveal malignant melanoma (UMM) is a rare cancer of
the melanocytes of the iris, ciliary body, and choroid eye
structures (1). While UMM accounts
for only 5% of all melanoma cases and has a yearly incidence of
just 5 cases per one million in the United States (2,3), it
is the most common intraocular malignancy affecting adults and the
most commonly occurring non-cutaneous form of melanoma (4,5).
Despite effective primary tumor management, one half of UMM cases
progress to metastatic disease, after which survival is usually
<1 year (6,7).
The successful use of targeted gene therapies, such
as BRAF and MEK inhibitors in cutaneous malignant melanoma (CMM)
has not translated to UMM. This is due in part to the genetically
distinct mutational landscape of UMM, which lacks the driver
mutations common to CMM, and to the rarity of the disease, which
limits patient availability for clinical trials (8). Trials with traditional
chemotherapeutic agents, such as dacarbazine and with novel
immunotherapies, such as checkpoint inhibitors have both failed to
lead to a reduction in the mortality rate (9-11),
and there is currently no FDA-approved therapy available for UMM
(12).
In this study, we discuss our use of a
discovery-based approach to identify already-existing therapies
that have potential activity against uveal melanoma. Using
high-throughput library screening with a large panel of
FDA-approved drugs on a UMM cell line, we ranked the activity of
each drug against UMM, and then further characterized the
discovered top-ranking calcium channel blocker class of medications
using in vitro studies. Our findings suggest that the
calcium signaling pathway may mediate the downstream signaling of
mutant GNAQ, a common uveal melanoma driver mutation.
Calcium channel blockade may thus be a potential strategy for the
treatment of uveal melanoma.
Materials and methods
Melanoma cell lines
We have adopted stringent criteria for establishing
the identity of the cell lines used in this study. New lines are
purchased directly from trusted repositories (e.g., ATCC), while
legacy cell lines (e.g., those cell lines which are gifts from
collaborators) are stratified into 3 levels of confidence after
sequencing. ‘CONFIRMED' cell lines are either new lines from ATCC
or those which have demonstrated an unequivocal match between our
designation and the STR database after STR genotyping. For those
with ‘no hits' in the STR database, we performed a manual search of
key melanoma mutations [e.g., BRAF(V600E), NRAS(Q61), etc.]
reported for each line in either the Catalogue Of Somatic Mutations
In Cancer (COSMIC), the Cancer Cell Lines Encyclopedia (CCLE), or
in individual publications in the literature. We subsequently
compared Sanger or whole exome sequence information generated in
our laboratory for these lines, and if there is a direct match
between our sequence information and the public domain data, these
cell lines are designated as ‘CONSISTENT'. For cell lines without
public domain information, we analyzed the levels of MITF
(M-isoform) to determine whether the cells are compatible with
melanocytic cells. Those that express significant M-isoform MITF
are designated as ‘COMPATIBLE'.
When selecting cell lines for experimentation, the
order of preferential selection is ‘CONFIRMED' > ‘CONSISTENT'
> ‘COMPATIBLE'. Furthermore, all cell lines have been tested for
common pathogens, such as mycoplasma, and are pathogen-free. The
following are accession numbers for cell lines from Cellosaurus
(web.expasy.org/cgi-bin/cellosaurus/search): MP41
(CVCL_4D12), Mel-270 (CVCL_C302), Mel-202 (CVCL_C301), OMM2.3
(CVCL_C306), CHL-1 (CVCL_1122), MeWo (CVCL_0445), SK-MEL-2
(CVCL_0069), SK-MEL-119 (CVCL_6077), Mel JuSo (CVCL_1403), IPC-298
(CVCL_1307), UACC-903 (CVCL_4052), SK-MEL-28 (CVCL_0526) and A375
(CVCL_0132).
High-throughput candidate drug
screening
To initially identify UMM-active compounds, the UMM
cell line, OMM2.3, was subjected to viability screening using an
FDA-approved Drug Library (Selleckchem) that included 1,018 drugs
(Table SI). Cells were seeded at
2,000 cells/well using an automated plate filler in a 384-well
plate and incubated for 24 h at 37°C. Subsequently, a 3 µM
concentration of each drug was added to the plate via pin-transfer
and the plates were incubated for an additional 72 h at 37°C. Drug
efficacy was measured via a proliferation ratio using
CellTiter-Glo® Luminescence assay (Promega) and an
EnVision plate reader (PerkinElmer).
Two-dimensional cell viability
Based on the results of this initial drug screen, 4
UMM (MP41, Mel-270, Mel-202 and OMM2.3) and 10 CMM (CHL-1, MeWo,
SK-MEL-2, SK-MEL-119, Mel JuSo, IPC-298, UACC-903, SK-MEL-28,
MGH-CH-1 and A375) cell lines were selected for additional drug
screening analysis with the dihydropyridine calcium channel blocker
(CCB), amlodipine (Selleck Chemicals), and the non-dihydropyridine
CCBs, verapamil and diltiazem (Sigma-Aldrich). The cells were
seeded at 2,000 cells/well in 96-well plates, incubated at 37°C for
24 h, and subjected to 9 doses of each drug in triplicate. Cell
viability was measured after 72 h of incubation at 37°C in the
presence of drug using CellTiter-Glo® Luminescence assay
(Promega). An equivalent volume of DMSO was used as a control, and
the results were normalized to this control value. IC50
values were calculated using GraphPad Prism software (Version 7)
and compared between the UMM and CMM lines using two-sided t-tests.
Intracellular calcium depletion after 48 h of treatment with 0, 2,
4 and 8 µM amlodipine was additionally visualized using 1
µM calcium-sensitive Rhod 2 fluorescent dye (Thermo Fisher
Scientific) on one UMM line (OMM2.3) and one CMM line (A375).
All CMM lines were cultured in Dulbecco's
modification of Eagle's medium (DMEM; Thermo Fisher Scientific)
with 10% fetal bovine serum (FBS; Atlanta Biologicals) and 1%
penicillin/streptomycin (Thermo Fisher Scientific). UMM lines were
cultured in RPMI-1640 with 10% FBS, 1% each of HEPES, L-glutamine,
penicillin/streptomycin, and 0.1% of β-mercaptoethanol (all from
Thermo Fisher Scientific).
Three-dimensional (3D) spheroid
model
We then performed drug screening analyses on a
smaller cohort of melanoma cell lines in a 3D culture system to
evaluate the ability of the drug to penetrate a tumor-like
spheroid. For spheroid generation in 3D culture, 100 µl of
Matrigel® were added to 96-well plates followed by
incubation at 37°C for 1 h to allow for matrix solidification. Cell
lines suspended in 70 µl of their respective growth media
were then seeded on top of the matrix base at an
empirically-determined density to allow for proper growth over the
experimental duration (A375 cells, 800 cells/well; CHL-1 cells,
1,000 cells/well; IPC-298 cells, 2,000 cells/well; MP41 cells,
2,500 cells/well; OMM2.3 cells, 2,500 cells/well). Following an
additional incubation at 37°C for 30 min, a 10 µl mixture of
Matrigel® and DMEM (at a ratio of 1:10, respectively)
was carefully transferred to each well and allowed to settle to the
bottom of the cell culture medium. Thus, the cell layer was secured
in place between 2 layers of Matrigel® extracellular
matrix.
The cells were cultured for 11 days to allow for
spheroid formation, after which time a 20 µM concentration
of amlo-dipine was added on day 0. Following a 48-h incubation at
37°C in the presence of drug or the DMSO control, the diameter of
randomly-sampled spheroids in each colony was measured on day 2
using light microscopy (Zeiss Axiovert 100 TV) and ImageJ software.
Mean spheroid diameters for each colony were calculated from these
measurements and normalized to the mean diameter of the
corresponding cell line's colonies on day 0. The normalized mean
diameters on day 2 were compared between conditions using two-sided
t-tests.
Cell cycle, apoptosis and senescence
To characterize the mechanism responsible for the
observed inhibitory effects of calcium channel blockade, we
performed several functional assays on UMM and CMM cell lines. Cell
cycle analysis was first performed using propidium iodide (PI)
staining. The cell lines were plated on 6-well plates at 50%
confluence and cultured for 24 h. They were then treated with 5, 10
or 20 µM of amlodipine and incubated at 37°C for an
additional 24 h. The cells were then trypsinized, centrifuged at
room temperature (1,500 rpm, 5 min), and washed with PBS. Following
re-suspension in 100 µl of 1X PBS, 250 µl of 70% cold
ethanol was added drop-wise while vortexing. The samples were
frozen at -20°C overnight for fixation. The following day, the
cells were thawed, and the ethanol was removed. The cells were then
washed with 1X PBS and resuspended in 0.5 ml PI staining solution
[1 ml of PI (100 µg/ml), 80 µl of RNAse (100
µg/ml), and 20 µl of NP-40 (0.1%); Molecular Probes]
prior to incubation at room temperature in the dark for 30 min. All
analyses were performed on FACSCalibur Flow Cytometer (BD
Biosciences).
After observing significant growth inhibition in our
cell viability analyses, we hypothesized that amlodipine may be
initiating its effects on cell growth through the induction of
apoptosis. We used Annexin V expression as an early apoptotic
marker. An Alexa Fluor 488 Annexin V/PI staining kit was used
following the manufacturer's protocol (Life Technologies; Thermo
Fisher Scientific). Briefly, the cells were collected after 24 h of
5, 10 or 20 µM amlodipine treatment, washed twice with PBS,
and stained with Alexa Fluor Annexin V for 20 min followed by PI
for 1 min both at room temperature. Flow cytometry was performed on
a BD FACSAria (BD Biosciences), and all data were analyzed using
FlowJo 10.0.8 software.
To detect for the association of a
senescent-associated effect of amlodipine treatment, 2 UMM (MP41
and OMM2.3) and 2 CMM (A375, CHL-1 and IPC-298) cell lines were
plated on a 12-well plate and cultured for 24 h. The cells were
then treated with 3 different concentrations of (2, 4 and 8
µM) of amlodipine and incubated at 37°C for an additional 48
h. The medium was then removed, and the cells were washed with 1X
PBS followed by the addition of 0.5 ml of fixing solution for 15
min. The plate was again washed with 1X PBS prior to the addition
of 0.5 ml of staining solution mix detection kit (470 µl
staining solution, 5 µl staining supplement, 25 µl of
20 mg/ml SA-β-gal in DMF; BioVision). The cells were incubated
overnight at 37°C and senescence was visualized at 24 h using light
microscopy (Zeiss Axiovert 100 TV).
Western blot analysis
Western blot analysis was also preformed to validate
the apoptotic effects. Uveal lines (MP41 and OMM2.3) were grown on
60 mm plates, treated with either 10 µM of amlodipine or an
equivalent volume of DMSO for the controls, and incubated at 37°C
for 24 h. Protein extraction was then performed using RIPA buffer
(50 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate,
and 0.1% SDS; Boston Bioproducts). A Lowry assay was performed to
determine protein quantification, and sample concentrations were
adjusted accordingly based on the standard curve produced. A total
of 10 µg of each sample were loaded per well and run through
a 4-12% SDS-PAGE gel prior to transfer to a nitrocellulose membrane
(Bio-Rad Laboratories). Following incubation with 5% non-fat milk
in TBST for 1 h at room temperature, the membrane was washed with
TBST and probed with rabbit antibodies against cleaved PARP (cat.
no. 9541s; Cell Signaling Technology) and mouse antibodies against
GAPDH as a loading control (cat. no. ab8245; Abcam) at a 1:3,000
dilution for 16 h at room temperature. The blots were washed again
with TBST and then incubated with a 1:5,000 dilution of horseradish
peroxidase-conjugated anti-rabbit (cat. no. 7074s; Cell Signaling
Technology) or anti-mouse antibodies (cat. no. 7076s; Cell
Signaling Technology) for 16 h at 4°C. The blots were washed again
with TBST and developed with Clarity western ECL substrate (Bio-Rad
Laboratories).
Statistical analysis
All viability and functional assays were performed
in triplicate and with biological replicates of each class of
melanoma cell lines (UMM and CMM) where appropriate. The Student's
t-test was used to perform both comparisons of mean IC50
values following two-dimensional CCB treatment in CMM vs. UMM and
comparisons of mean spheroid growth after three-dimensional CCB
treatment in treated vs. control groups of each individual cell
line. The comparison of apoptosis induction between treated and
control groups was performed using one-way ANOVA and Dunnett's
multiple comparisons test (with control group as reference).
Significant differences for all tests were assumed at P-values
<0.05. All statistical analyses were performed using GraphPad
Prism 8 software.
Results
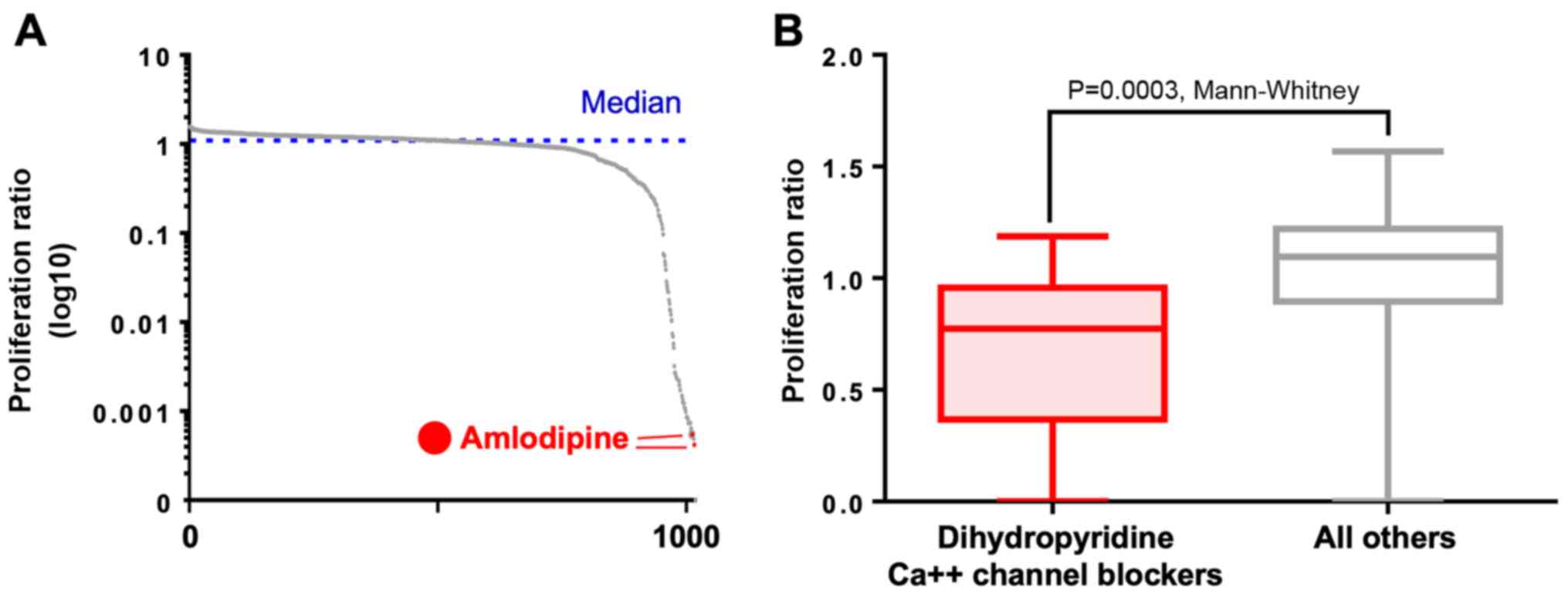
The initial library screening identified amlodipine
as the no. 2 and no. 8 most active compound in UMM out of the 1,018
screened compounds (Fig. 1A and
Table SI). The dihydropyridine
CCB class as a whole had a significantly lower proliferation ratio
than all other screened compounds (P=0.0003, Mann-Whitney U test;
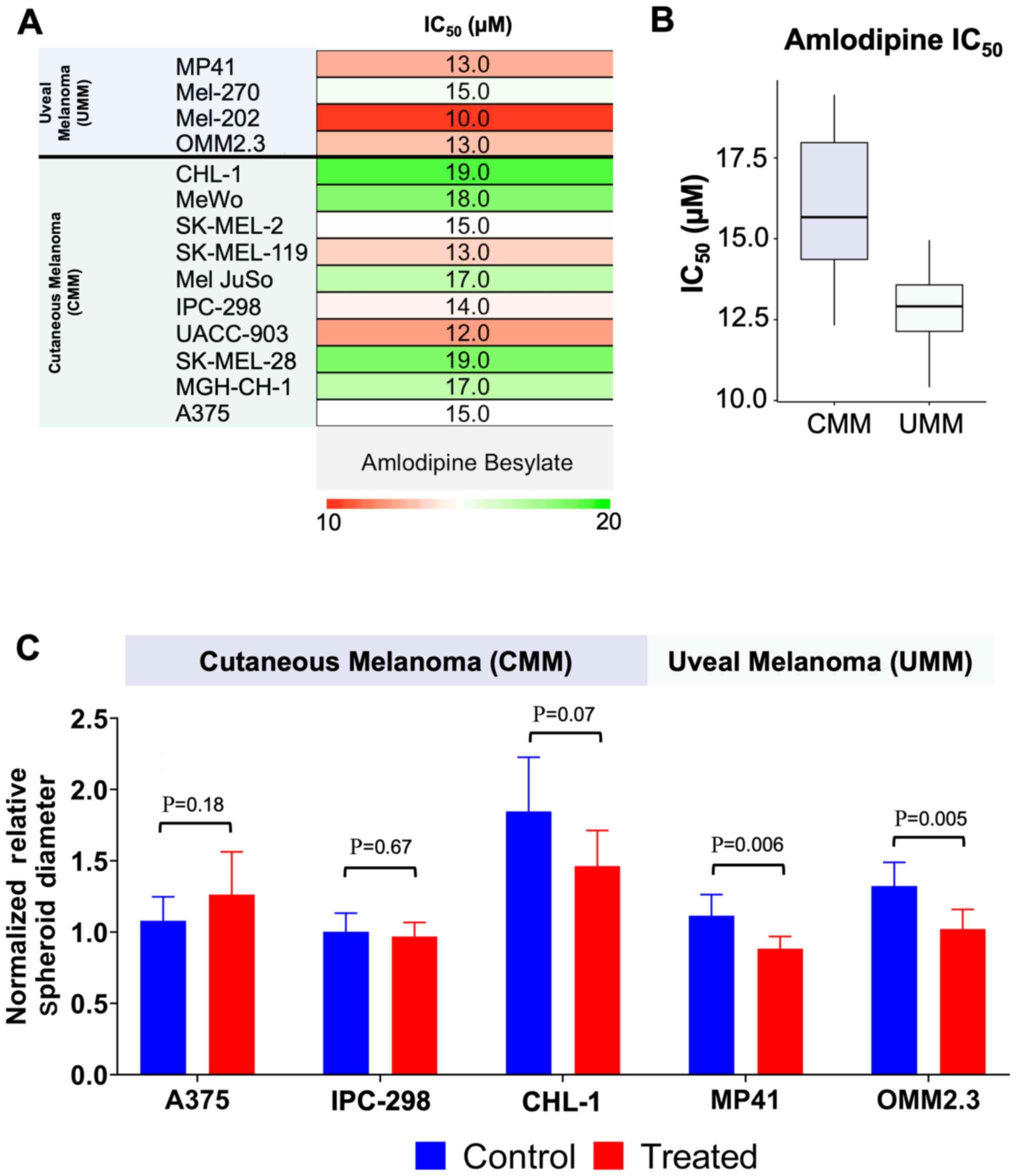
Fig. 1B). During 2D screening in
our cohort, amlodipine yielded an IC50 value of 12.8
µM in the UMM lines compared to 15.9 µM in the CMM
lines (P=0.035; Figs. 2A and B).
Treatment with both the non-dihydropyridine CCBs, verapamil and
diltiazem, failed to reach 50% growth inhibition in either CMM or
UMM (maximum concentration: 30 µM; data not shown). Rhod-2
staining demonstrated effective intracellular calcium depletion
following 48 h of treatment with 1, 3 and 8 µM amlodipine in
UMM. Contrarily, intracellular calcium levels were unaffected by
amlodipine treatment in CMM (Fig.
S1). Based on these preliminary results, we focused primarily
on amlodipine rather than verapamil or diltiazem in all subsequent
assays.
Spheroids subjected to 48 h of treatment with 20
µM amlo-dipine displayed, on average, a significantly
decreased spheroid volume growth in UMM lines compared to the DMSO
control. In the UMM line, MP41, control spheroid diameter increased
by 11.5%, while the amlodipine-treated MP41 spheroids exhibited a
decay of 11.6% (P=0.006). The OMM2.3 spheroids grew by 32.4%, while
the amlodipine-treated spheroids grew an average of only 2.2%
(P=0.005). In 3 separate colonies of CMM spheroids (A375, CHL-1 and
IPC-298), there was no evidence of significant growth inhibition
with amlodipine treatment compared to the DMSO control, and, in
fact, the treated A375 spheroids appeared to grow more rapidly than
their control counterparts (Figs.
2C and S2).
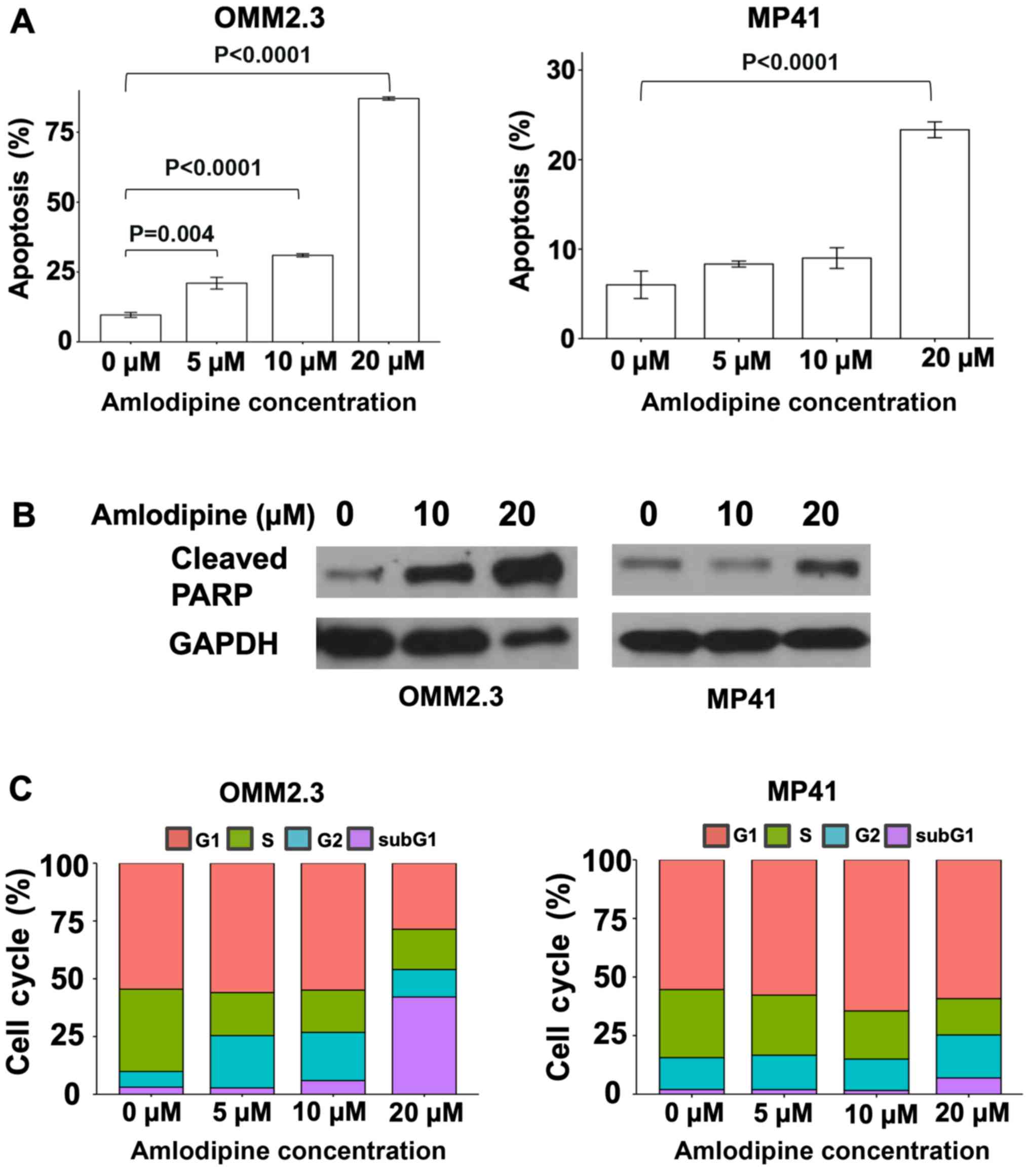
The UMM line, OMM2.3, exhibited an 87% induction of
the early apoptotic marker, Annexin V, with 20 µM amlodipine
treatment compared to 10% Annexin V induction with DMSO
(P<0.0001). The induction was less pronounced at 21 and 31% by
treatment with amlodipine at 5 and 10 µM (P=0.004 and
P<0.0001, respectively). The MP41 cells exhibited a 23% Annexin
V induction with 20 µM amlodipine and 6% with DMSO
(P<0.0001) (Fig. 3A). The
amount of induction with lower concentrations of amlodipine did not
differ significantly from the DMSO control. Western blot analysis
of the MP41 and OMM2.3 cells for cleaved PARP, a known indicator of
apoptosis, revealed that amlodipine treatment increased the levels
of cleaved PARP in both lines in a dose-dependent manner (Fig. 3B). The observed effect was more
prominent in the OMM2.3 cells. Cell cycle analysis demonstrated G1
and G2 phase arrest, S phase reduction and an increase in the
number of cells in the sub-G1 phase in both the MP41 and OMM2.3
lines treated with amlodipine compared to the DMSO control
(Fig. 3C). SA-β-gal did not reveal
any marked changes in the presence or absence of amlodipine in CMM
and UMM lines (Fig. S3).
Discussion
Considering the results of our unbiased library
screening on the UMM line, OMM2.3, which was further confirmed in
2D and 3D experiments on larger cohorts, we hypothesized that the
calcium signaling pathway may be a potentially useful therapeutic
target in UMM. We demonstrated that amlodipine, which was
represented twice amongst the top 10 active compounds of our
candidate screening, significantly and selectively inhibited UMM
growth when compared to that of CMM in both 2D and 3D culture
systems. Furthermore, the spheroid growth inhibition observed in
this study suggests that amlodipine can act biologically as a
UMM-selective agent in a 3D tumor. We consider that amlodipine
likely causes growth inhibition by mediating an increase in
apoptosis and cell cycle arrest in UMM. We observed a consistent
increase in apoptosis independently through both Annexin V and PARP
assays, which further strengthens this finding. In addition, given
our preliminary results, we consider that senescence is unlikely to
markedly contribute to growth inhibition in UMM.
While our results support the role of calcium
signaling in the proliferation and survival of UMM, the mechanism
behind this association has not yet been defined. The calcium
signaling pathway, and its dysregulation, has a well-documented
association with cancer survival, proliferation, migration and
metastatic potential (13-16). For example, calcium signaling has
been reported to be involved in the proliferation of
RAS-driven cancers through the interaction of calmodulin
(CaM) and PI3K (17) and the
promotion of invasion and metastasis via ERK activation in both
BRAF-driven and non-BRAF melanoma cells (18,19).
One such example that could explain the selectivity we observed in
UMM is RasGRP3, which was identified by Chen et al (2017) as
a link between MAPK activation and the GNAQ/11 mutations
that characteristically drive UMM (20). RasGRP3, which is
overex-pressed in response to GNAQ/11 mutations, reportedly
drives the MAPK pathway through the activation of HRAS (20). RasGRP3 itself is activated both
through phosphorylation by protein kinase C (PKC), which is
calcium-activated, or through membrane recruitment by
diacylglycerol kinase (DAG) (21),
which similarly to RasGRP3 contains a calcium-binding EF-hand
motif. The ability of calcium to potentially alter this unique UMM
pathway, although speculative, is evidence of the multitude of
possible avenues for the involvement of calcium signaling, and
thereby calcium channel blockade, in the oncogenic landscape of
UMM.
It is of particular interest that
non-dihydropyridine CCBs demonstrated minimal to no inhibition in
our UMM or CMM cell lines, which further suggests the involvement
of a distinct and targetable alteration in the calcium signaling
pathway in melanoma. Both families of CCBs are known to bind L-type
calcium channels, but at different binding sites within the
α1-subunit (22). It has been
suggested that specific isoforms of the L-type channel are
particularly upregulated in melanoma, while others are widely
expressed in both melanoma and normal melanocytes (23), and each individual isoform has a
particular pattern of tissue expression (24). Therefore, it is possible that the
selectivity reported here is driven by a unique disparity of L-type
calcium channel expression in UMM and CMM.
We recognize several limitations in our study. Our
results stem from in vitro experiments and would likely
benefit from further confirmatory work using animal models or human
studies. Future research may also benefit from large-scale
population studies of patients on long-term therapy with amlodipine
and their risk for UMM, although this will remain challenging given
the considerably low incidence of UMM. We also acknowledge that the
variability in growth kinetics between UMM and CMM cell lines may
partially explain the observed differences in cell viability and
growth inhibition. We have partially addressed this limitation by
controlling our drug treatments with DMSO solvent. It is possible
that calcium plays a role in metabolic regulation over oncogenic
activation, and the significance of other pathways cannot be
excluded at this time. We recognize that the concentration of
amlodipine used in our experiments is higher than what is typically
achieved with standard dose for cardiovascular disease management,
but we found that it is comparable to the micromolar range
concentrations used in the literature to study amlodipine's
antiproliferative effects on various cancers and other cell types
(25-27). Finally, any mechanistic explanation
discussed herein is only speculative, and studies that aim to
further delineate the mechanism of action that validates our
demonstrated inhibitory effects of amlodipine on UMM are currently
underway.
In conclusion, UMM is an aggressive primary ocular
tumor with a high risk of metastasis and no known effective
treatments. We propose that calcium inhibition is one potential
strategy for targeting UMM, as it has already demonstrated
selective and significant in vitro growth inhibition in UMM
compared to CMM. Based on the information gathered in this study,
further preclinical trials in animal models and genetic studies
utilizing gene silencing techniques of candidate genes are required
to firmly establish calcium channel blockade as an effective
therapeutic strategy as well as uncovering a possible mechanism of
action.
Supplementary Data
Funding
This study was supported in part by the US NIH (K24
CA149202 to HT) and by generous donors to Massachusetts General
Hospital on behalf of melanoma research.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
MS, GLM, NK, ZJ, AR, RK, KF and HT were involved in
the experimental design and conception of the study. MS, GLM, NK,
ZH, AR and RK were involved in data acquisition. MS, GLM, NK, ZH,
AR, RK, KF and HT were involved in data analysis and
interpretation. MS, GLM, NK, ZH, AR, RK, KF and HT were involved in
the preparation of the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Shields CL, Furuta M, Thangappan A, Nagori
S, Mashayekhi A, Lally DR, Kelly CC, Rudich DS, Nagori AV, Wakade
OA, et al: Metastasis of uveal melanoma millimeter-by-millimeter in
8033 consecutive eyes. Arch Ophthalmol. 127:989–998. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chattopadhyay C, Kim DW, Gombos DS, Oba J,
Qin Y, Williams MD, Esmaeli B, Grimm EA, Wargo JA, Woodman SE, et
al: Uveal melanoma: From diagnosis to treatment and the science in
between. Cancer. 122:2299–2312. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shaughnessy M, Klebanov N and Tsao H:
Clinical and therapeutic implications of melanoma genomics. J
Transl Genet Genom. 2:142018.
|
|
4
|
Singh AD and Topham A: Incidence of uveal
melanoma in the United States: 1973-1997. Ophthalmology.
110:956–961. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kaliki S and Shields CL: Uveal melanoma:
Relatively rare but deadly cancer. Eye (Lond). 31:241–257. 2017.
View Article : Google Scholar
|
|
6
|
Grisanti S and Tura A: Uveal melanoma.
Noncutaneous melanoma. Scott JF and Gerstenblith MR: Codon
Publications; Brisbane: 2018, View Article : Google Scholar
|
|
7
|
Eskelin S, Pyrhönen S, Hahka-Kemppinen M,
Tuomaala S and Kivelä T: A prognostic model and staging for
metastatic uveal melanoma. Cancer. 97:465–475. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Field MG, Durante MA, Anbunathan H, Cai
LZ, Decatur CL, Bowcock AM, Kurtenbach S and Harbour JW: Punctuated
evolution of canonical genomic aberrations in uveal melanoma. Nat
Commun. 9:1162018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Augsburger JJ, Corrêa ZM and Shaikh AH:
Effectiveness of treatments for metastatic uveal melanoma. Am J
Ophthalmol. 148:119–127. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Desjardins L, Levy C, Lumbroso Le Rouic L,
Cassoux N, Piperno-Neumann S, Mariani P, Servois V, Dendale R,
Plancher C and Asselain B: Adjuvant intravenous therapy by
fotemustine in uveal melanoma: A randomised study. Acta
Ophthalmologica. Sep 15–2011.Epub ahead of print. View Article : Google Scholar
|
|
11
|
Komatsubara KM and Carvajal RD:
Immunotherapy for the treatment of uveal melanoma: Current status
and emerging therapies. Curr Oncol Rep. 19:452017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jovanovic P, Mihajlovic M,
Djordjevic-Jocic J, Vlajkovic S, Cekic S and Stefanovic V: Ocular
melanoma: An overview of the current status. Int J Clin Exp Pathol.
6:1230–1244. 2013.PubMed/NCBI
|
|
13
|
Stewart TA, Yapa KT and Monteith GR:
Altered calcium signaling in cancer cells. Biochim Biophys Acta.
1848:2502–2511. 2015. View Article : Google Scholar
|
|
14
|
Brzozowski JS and Skelding KA: The
multi-functional calcium/calmodulin stimulated protein kinase
(CaMK) family: Emerging targets for anti-cancer therapeutic
intervention. Pharmaceuticals (Basel). 12. pp. 122019, View Article : Google Scholar
|
|
15
|
Cui C, Merritt R, Fu L and Pan Z:
Targeting calcium signaling in cancer therapy. Acta Pharm Sin B.
7:3–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang X, Liu H, Xu Y, Xie J, Zhu D, Amos
CI, Fang S, Lee JE, Li X, Nan H, et al: Genetic variants in the
calcium signaling pathway genes are associated with cutaneous
melanoma-specific survival. Carcinogenesis. 40:279–288. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nussinov R, Wang G, Tsai CJ, Jang H, Lu S,
Banerjee A, Zhang J and Gaponenko V: Calmodulin and PI3K signaling
in KRAS cancers. Trends Cancer. 3:214–224. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maiques O, Barceló C, Panosa A, Pijuan J,
Orgaz JL, Rodriguez-Hernandez I, Matas-Nadal C, Tell G, Vilella R,
Fabra A, et al: T-type calcium channels drive migration/invasion in
BRAFV600E melanoma cells through Snail1. Pigment Cell Melanoma Res.
31:484–495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Umemura M, Baljinnyam E, Feske S, De
Lorenzo MS, Xie LH, Feng X, Oda K, Makino A, Fujita T, Yokoyama U,
et al: Store-operated Ca2+ entry (SOCE) regulates
melanoma proliferation and cell migration. PLoS One. 9:e892922014.
View Article : Google Scholar
|
|
20
|
Chen X, Wu Q, Depeille P, Chen P, Thornton
S, Kalirai H, Coupland SE, Roose JP and Bastian BC: RasGRP3
mediates MAPK pathway activation in GNAQ mutant uveal melanoma.
Cancer Cell. 31:685–696.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie S, Naslavsky N and Caplan S:
Diacylglycerol kinases in membrane trafficking. Cell Logist.
5:e1078431. 2015. View Article : Google Scholar
|
|
22
|
Opie LH: Pharmacological differences
between calcium antagonists. Eur Heart J. 18(Suppl A): A71–A79.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Das A, Pushparaj C, Bahí N, Sorolla A,
Herreros J, Pamplona R, Vilella R, Matias-Guiu X, Martí RM and
Cantí C: Functional expression of voltage-gated calcium channels in
human melanoma. Pigment Cell Melanoma Res. 25:200–212. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Striessnig J, Ortner NJ and Pinggera A:
Pharmacology of L-type calcium channels: Novel drugs for old
targets? Curr Mol Pharmacol. 8:110–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zou J, Li Y, Fan HQ and Wang JG: Effects
of dihydropyridine calcium channel blockers on oxidized low-density
lipoprotein induced proliferation and oxidative stress of vascular
smooth muscle cells. BMC Res Notes. 5:1682012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee H, Kang S and Kim W: Drug
repositioning for cancer therapy based on large-scale drug-induced
transcriptional signatures. PLoS One. 11:e01504602016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ohba T, Watanabe H, Murakami M,
Radovanovic M, Iino K, Ishida M, Tosa S, Ono K and Ito H:
Amlodipine inhibits cell proliferation via PKD1-related pathway.
Biochem Biophys Res Commun. 369:376–381. 2008. View Article : Google Scholar : PubMed/NCBI
|